
Свойства нервных центров обусловленные их структурой и метаболизмом – Вопрос 30.2 Определение нервного центра (классическое и современное). Свойства нервных центров, обусловленные их структурными звеньями (иррадация, конвергенция, последействием возбуждения)
Основные свойства нервных центров | Doctor-V.ru
Различают девять Основные свойства нервных центров:
1. Пространственная и временная суммация
Пространственная и временная суммация основана на свойстве каждого нейрона в центре к суммации как возбуждения, так и торможения. Поскольку каждый нервный центр имеет много параллельно расположенных афферентных или входных волокон от рецептивного поля рефлекса, слабые раздражения нескольких участков рецептивного поля, в отдельности не способные реализовать рефлекс, вызывают в нейронах центра несколько ВПСП, которые суммируются, приводя к формированию на мембране нервной клетки потенциалов действия, распространяющихся по эфферентным проводникам, вызывая рефлекторную реакцию. Это явление называют пространственной суммацией. При увеличении частоты афферентных сигналов в единицу времени амплитуда ВПСП нарастает до критического уровня из-за повышения эффективности синаптического проведения, что также вызывает возбуждение нейронов и рефлекторный ответ на слабые частые раздражения. Это явление называют
2. Центральная задержка рефлекса
Центральная задержка рефлекса, характеризуется временем распространения информации в структурах нервного центра, главным образом в синапсах, где скорость проведения сигнала существенно меньше, чем в нервных проводниках. Поэтому, центральная задержка рефлекса зависит от количества синапсов между нейронами центра и представляет собой сумму синаптических задержек;
3. Посттетаническая потенциация
Посттетаническая потенциация — увеличение амплитуды ВПСП после серии частых (тетанизирующих) ритмических возбуждений, что связано с временной суммацией частых ВПСП и активацией синаптического проведения из-за увеличения числа квантов медиатора. Длительность состояния потенциации синапсов может достигать нескольких часов, что играет роль в процессах обучения и памяти:
4. Последействие и пролонгирование возбуждения
Последействие и пролонгирование возбуждения — связаны с длительными следовыми потенциалами в нейронах, улучшением синаптического проведения, наличием кольцевых нейронных цепей и реверберацией возбуждения. Все эти процессы также играют роль в процессах обучения и памяти;
5. Трансформация ритма возбуждений
Трансформация ритма возбуждений, т.е. увеличение или уменьшение частоты нервных импульсов и эфферентных проводниках (на выходе) по сравнению с частотой афферентной импульсации (на входе центра), что связано с механизмом синаптической передачи (трансформация ритма как свойство синапса) и интегративной деятельностью нейронов;
6. Спонтанная (фоновая) электрическая активность
Спонтанная (фоновая) электрическая активность — периодическое генерирование импульсов возбуждения (потенциалов действия) нервными клетками центра в состоянии покоя, т.е. без специфичес кого раздражения рецептивного поля рефлекса. Наличие спонтанной активности обусловлено тем, что организму не свойственно абсолютное отсутствие раздражителей или информационный покой, при этом за счет дивергенции и конвергенции возбуждений в нервных сетях нейроны центра всегда получают возбуждающие импульсы и от клеток других нервных центров. В происхождении спонтанной активности нейронов играют роль также метаболические сдвиги внутриклеточной среды и микроокружения клеток, а также суммация миниатюрных потенциалов постсинаптической мембраны, формирующихся из- за «утечки» единичных квантов медиатора;
7. Тонус нервного центра
Тонус нервного центра — состояние некоторого уровня активности нейронов, обеспечивающей их готовность к рефлекторной деятельности и проявляющейся в постоянной эфферентной импульсации низкой частоты к органам-эффекторам. Тонус нервных центров обусловлен небольшим уровнем афферентных сигналов от различных рецептивных полей (т.е. имеет рефлекторную природу), а также действием на нейроны метаболитов и других гуморальных раздражителей из клеточной микросреды. Проявлением тонуса нервных центров является спонтанная электрическая активность нейронов и некоторая фоновая активность эффекторов, например, тонус скелетной мускулатуры, гладких мышц сосудов и т.п.;
8. Пластичность нервных центров
Пластичность нервных центров — способность перестраивать функциональные свойства для более эффективной регуляции функций, осуществления новых, ранее несвойственных этому центру рефлексов или восстановления функций после повреждения части нейронов центра. Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками, является рабочим механизмом обучения. В основе пластичности лежат функциональные особенности синапсов и мембран нейронов («тренировка» синапсов, посттетаническая потенциация, периодичность функционирования синапсов и нейронов, пространственная и временная суммация постсинаптических потенциалов), а также наличие многочисленных дублирующих систем нейронов и нервных волокон;
9. Утомление нервных центров
Утомление нервных центров — снижение эффективности их деятельности в виде повышения порогов возбуждения, связанное с утомлением синапсов и метаболическими сдвигами типа энергетического истощения в нервных клетках. Утомление формируеся при чрезмерной продолжительности действия раздражителей или их интенсивности, напряженном умственном труде или физической работе.
Читайте также:
doctor-v.ru
Вопрос 30.2 Определение нервного центра (классическое и современное). Свойства нервных центров, обусловленные их структурными звеньями (иррадация, конвергенция, последействием возбуждения)
Нервный центр- это динамическая совокупность взаимосвязанных нейронов, расположенных в разных отделах ЦНС, которая формируется в процессе развития организма и обеспечивать какую-либо функцию.
Нервный центр-это совокупность взаимосвязанных нейронов, расположенных в какой-либо области ЦНС, которая обеспечивает рефлекторных ответ.
Свойства НЦ:
1.чувствительность к гипоксии
2.чувствительность к конц. Глюкозы в крови
4. Иррадиация возбуждения. Иррадация — распределение возбуждения от одного к множеству. В естественных условиях ее не бывает. Возбуждение неограниченно распространяется.
5. Суммация возбуждения.
6. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка).
7.Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением
8. Тонус. Тонус, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки
9. Пластичность.
10. Конвергенция. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны.
11. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
12. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
13. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы.
Вопрос 31.3 Физиологические свойства нервных центров: пространственная и временная суммация, трансформация ритма, посттетаническая потенциация, низкая лабильность, утомляемость, чувствительность к нейротропным веществам, автоматия.
1. Пространственная и временная суммация.
Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД.
2. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне (рис). Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
3. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы,. выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
4. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов.
6. Автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
studfiles.net
6.2. Свойства и принципы функционирования нервных центров
Свойства нервных центров. К ним относят:
замедленное распространение возбуждения, обусловленное наличием синапсов и многих нейронов на путях передачи возбуждения;
одностороннее распространение возбуждения, обусловленное тем, что основную роль в распространении возбуждения играют химические синапсы, в которых возбуждение передается только от пресинаптических структур на постсинап- тические структуры;
иррадиация возбуждения — распространение возбуждения от активированного нервного центра на соседние и функционально связанные нейронные структуры. Это свойство обусловлено наличием многочисленных ветвлений аксонов, заканчивающихся синаптическими контактами. В результате при активации группы нейронов от них идут возбуждающие влияния на синаптически связанные с ними другие нейронные группы. При прекращении притока возбуждающей импульсации активность нервного центра уменьшается и может наступать концентрация возбуждения — процесс, обратный иррадиации;
суммация возбуждения, проявляющаяся увеличением интенсивности рефлекторной реакции при возрастании длительности раздражения, его силы или площади раздражаемого рецепторного поля. Различают временную и пространственную суммацию. При временной суммации количество активируемых синаптических образований на эфферентном нейроне не изменяется, но увеличивается частота импульсаций, приходящих к каждому синапсу. При пространственной суммации из-за воздействия на новые рецепторы увеличивается количество одновременно активируемых синаптических входов и эфферентный нейрон быстрее и эффективнее возбуждается. В обоих случаях возрастает величина ответной реакции эффекторных структур. Например, увеличивается интенсивность сокращения мышцы, секреции слезной или слюнной железы.
трансформация ритма возбуждений, проявляющаяся изменением частоты импульсаций, генерируемых эфферентными нейронами, по отношению к частоте импульсаций, передаваемых афферентными нейронами. Такое изменение становится возможным благодаря суммации ВПСП, функционированию локальных нейронных цепей и т.д.;
явление последействия, заключающееся в способности нервных центров длительно поддерживать возбуждение после устранения раздражителя, вызвавшего его. Одним из механизмов этого служит возможность длительной циркуляции возбуждения в замкнутых нейронных цепях;
тонус нервных центров, проявляющийся длительным непрерывным возбуждением нервных центров. Такое состояние поддерживается за счет непрерывного притока афферентных импульсаций, влияния биологически активных веществ, доставляемых гуморальным путем, циркуляцией возбуждения в замкнутых нейронных цепях, спонтанной активностью отдельных нейронов;
пластичность нервных центров — способность к перестройке функциональных свойств и изменению выполняемых функций. Например, экспериментально показана способ
но ность диафрагмальных мотонейронов вместо обеспечения ритмических сокращений диафрагмы выполнять функцию регуляции шагательных движений ног;
повышенная утомляемость нервных центров по отношению к утомляемости периферических нервных структур и мышц- В системе нервный центр — нервно-мышечный препарат первое место по утомляемости занимает нервный центр, второе — нервно-мышечный синапс, третье — скелетная мышца и наименее утомляемым оказывается нервное волокно;
высокая чувствительность нервных центров к действию биологически активных веществ и ядов, нарушениям гомеостаза и недостатку кислорода. При прекращении доставки кислорода клетки коры мозга могут сохранять жизнедеятельность не более 5 мин, уменьшение содержания глюкозы в крови ниже 800 мг/л сопровождается гипогликемической комой.
Принципы функционирования, интегративной и координационной деятельности ЦНС. Интегративная деятельность ЦНС проявляется восприятием, объединением и переработкой комплекса поступающей к нервным центрам информации и принятием решений, обеспечивающих регуляцию гомеостаза, а также вегетативных и соматических реакций, направленных на достижение полезного для организма приспособительного результата. Координационной деятельностью называют регуляцию распределения возбуждения и торможения в нейронных структурах, а также взаимодействия нервных центров, обеспечивающие адекватные рефлекторные и произвольно вызываемые реакции.
Принцип рефлекторных регуляций — важнейший принцип функционирования нервной системы.
Рефлексом называют стереотипную ответную реакцию организма на действие раздражителя, осуществляющуюся благодаря регуляторной роли нервных клеток, составляющих рефлекторную дугу.
Абсолютное большинство рефлексов замыкается в головном и спинном мозге. Но есть также рефлексы, рефлекторная дуга которых замыкается вне центральной нервной системы в вегетативных внеорганных ганглиях или даже в пределах одного органа (например, сердца или кишечника).
Рефлекторная дуга включает 5 звеньев. Начальным звеном является рецептор, затем идет афферентный (чувствительный, Центростремительный) нейрон, ассоциативный (вставочный)
нейрон, эфферентный (двигательный, центробежный) нейрон и эффектор. Эффектором может быть любая структура (мышца, железа и т.д.), на которой заканчивается синапсом эфферентный нейрон. Вставочный нейрон может быть один или их может быть много. Они располагаются в нервных центрах.
Следовательно, в образовании рефлекторной дуги участвует как минимум 3 нейрона. Исключение составляет лишь один вид рефлексов так называемые сухожильные рефлексы, рефлекторная дуга которых включает только 2 нейрона: афферентный и эфферентный. Отросток афферентного нейрона входит в спинной мозг через задние корешки и, проникая в передние рога серого вещества, формирует синапс прямо на эфферентном нейроне. Примеры схем двух- и трехнейронной рефлекторной дуги соответственно сухожильного и оборонительного сгибательного рефлекса, вызываемого раздражением кожи, представлены на рис. 6.4.
Область сосредоточения рецепторов, с которых запускается определенный рефлекс, называют рецепторным полем этого рефлекса.
Разрез спинного мозга
Кожа
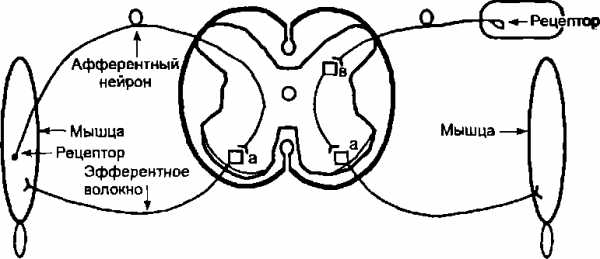
Рис. 6.4. Схема двухнейронной рефлекторной дуги сухожильного рефлекса и трехнейронной рефлекторной дуги сгибательного оборонительного
рефлекса: а — а-мотонейрон, в — вставочный нейрон
Двухнейронная Трехнейронная
рефлекторная рефлекторная
дуга дуга
Классификация рефлексов: все рефлекторные реакции подразделяют на безусловные и условные. Безусловные врожденные и проявляются при воздействии специфическогораздражителя на строго определенное рецепторное поле. Условные — приобретаются или вырабатываются в процессе жизни. Подробная их характеристика будет дана при изучении высшей нервной деятельности. Здесь же рассмотрим наиболее употребительные варианты классификации безусловных рефлексов.
По биологической значимости рефлекторной реакции выделяют пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы.
По виду рецепторов различают эстероцептивные, инте- роцептивные, проприоцептивные рефлексы. Среди последних выделяют сухожильные и миотатические рефлексы.
По участию в рефлексе соматических или вегетативных отделов ЦНС и органов эффекторов различают соматические и вегетативные рефлексы. Если эффектор и рецепторное поле рефлекса относятся к соматическим структурам, то такие рефлексы называют соматическими. Вегетативными называют рефлексы, эффектором в которых являются внутренние органы, а эфферентная часть рефлекторной дуги образована вегетативными нейронами. Примером вегетативного рефлекса является рефлекторное замедление сердечной деятельности, вызванное воздействием на рецепторы желудка. Примером соматического рефлекса является сгибание руки в ответ на болевое раздражение кожи.
По уровню замыкания рефлекторной дуги в центральной нервной системе выделяют спинальные, бульварные (замыкающиеся в продолговатом мозге), мезенцефальные и талами- ческие рефлексы.
По количеству нейронов, участвующих в замыкании рефлекторной дуги, и числу центральных синапсов выделяют двухнейронные, трехнейронные, мультинейронные; моносинап- тические, полисинаптические рефлексы.
Благодаря принципу обратной связи (обратной афферентации) формируется замкнутый регуляторный контур. Только замкнутая регуляторная система может обеспечить устойчивое, адекватное регулировние. Благодаря обратной связи осуществляется контроль за исполнением, результатом влияния управляющего сигнала на эффекторные структуры и величиной регулируемого параметра.
Участие обратной связи даже в простейших регуляциях можно рассмотреть на примере реализации сгибательного
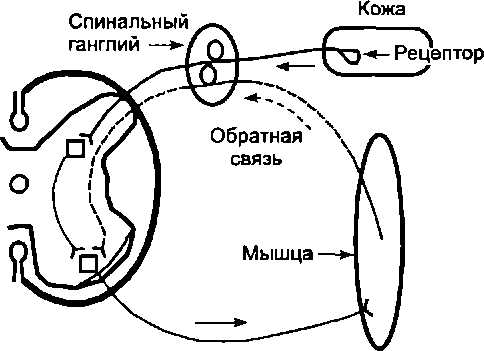
Рис. 6.5. Обратная связь в простейших рефлекторных реакциях
рефлекса, вызываемого прикосновением к коже (рис. 6.5). При рефлекторном сокращении мышцы изменяется активность находящихся в ней рецепторов и частота импульсаций в афферентных волокнах, идущих к мотонейронам. В результате формируется замкнутый контур регулирования, в котором роль канала обратной связи выполняют афферентные волокна, передающие импульсацию в нервные центры от рецепторов мышц, а роль канала прямой связи — эфферентные волокна, идущие к мышцам от мотонейронов. Таким образом, нервный центр (его мотонейроны) получает информацию об изменении состояния мышцы, вызванном передачей импульсов по двигательным волокнам. Благодаря обратной связи формируется своеобразное регуляторное кольцо. Поэтому некоторые авторы даже предлагали вместо термина «рефлекторная дуга» применять термин «рефлекторное кольцо»
Участие обратной связи в механизме регуляции кровообращения, дыхания, температуры тела и поведенческих реакций рассматривается ниже в соответствующих разделах.
Принцип реципрокной связи проявляется во взаимодействии центров-антагонистов по функциональному назначению, например группы мотонейронов, ответственных за сгибание руки, и группы, управляющей мышцами-разгибателями руки. При реципрокной связи возбуждение одного из антагонистических центров приводит к торможению другого. Это осуществляется за счет активации тормозных нейронов на конечном участке проводящих путей, идущих от возбужденного центра к антагонисту.
Принцип конвергенции заключается в схождении, поступлении к одному и тому же нейрону импульсаций от различных нервных центров или рецепторов различных модальностей (различных органов чувств). На основе конвергенции самые разные раздражители могут вызвать однотипную реакцию. Например, сторожевой рефлекс (поворот головы, насторажи- вание) может быть вызван и световым, и звуковым, и тактильным воздействием. Возможность конвергенции импульсаций с разных входов на одни и те же эфферентные нейроны называют принципом общего конечного пути.
Принцип дивергенции утверждает возможность расхождения импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.
Принцип доминанты характеризует особенности взаимодействия нервных центров. Доминантный очаг возбуждения обладает стойкой высокой активностью, он подавляет возбуждение в других нервных центрах, подчиняет их своему влиянию, притягивает к себе афферентные импульсации, адресуемые к другим центрам, и усиливает свою активность за счет этих импульсаций. Доминантный центр может длительно находиться в состоянии возбуждения без признаков утомления.
Примером рефлекторной реакции, вызванной доминантным очагом возбуждения, может служить обнимательный рефлекс самца лягушки, проявляющийся в период размножения. Если в этот период положить самца лягушки брюшком на палец, то у животного возникает обнимательный рефлекс. Самец обхватывает и сжимает передними лапками палец. Затем металлическим пинцетом проводят по спинке животного. Это воздействие обычно вызывает оборонительную реакцию (бегства). Но в данных условиях воздействие пинцетом лишь усиливает обнимательный рефлекс, самец сильнее сдавливает палец.
Контрольные вопросы и задания
Какова роль нервной системы в организме ?
Как подразделяют нервную систему ? Какие клетки ее образуют?
Какова роль глиальных клеток в ЦНС?
Каковы структура и функции нейронов и их частей?
Каковы особенности поляризации мембрены нейрона и возбудимости его отделов?
Каковы особенности передачи возбуждения в центральных синапсах? Что такое медиаторы?
7 Перечислите виды и механизмы торможения.
Что такое интеграция возбуждения на нейроне?
Какова классификация нейронов?
Что такое нейронные цепи?
Каковы свойства нервных центров?
Дайте характеристику рефлекса и рефлекторной дуги.
Какова классификация рефлексов?
Каковы принципы осуществления нервных регуляций?
Ситуационные задачи
Как соотносится длительность латентных периодов сухожильных и оборонительных рефлексов, вызываемых раздражением кожи? Почему имеются такие различия?
Зарисуйте схему нейронной сети, которая может увеличить частоту эфферентных импульсов по отношению к частоте афферентных.
Какие механизмы обусловливают способность спинальных мотонейронов генерировать частоту возбуждения до 50 имп/с, а вставочных — до 500 имп/с?
Почему при наличии обнимательного рефлекса у самца лягушки ряд раздражителей перестает вызывать оборонительную реакцию, а вместо нее лишь усиливает обнимательный рефлекс?
—»— РазделП
ЧАСТНАЯ ФИЗИОЛОГИЯ
studfiles.net
11.Понятие о нервном центре. Свойства нервных центров. Принципы координации работы нервных центров в цнс.
Нервный центр – совокупность нервных клеток, регулирующиж определеную функцию организма.
Свойства нервных центров
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2.Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное число межнейронных со единений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой яв ляется постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов.
4 Наличие синаптической задержки. Время рефлекторной ре акции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относи тельно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка).
5.Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомле нием.
6. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
В координационной деятельности центральных нервных образо ваний значительная роль взаимодействия рефлексов, ко торая проявляется в различных эффектах (в облегчении, или суммации, и в угнетении, или подавлении, возбуждения).
Координационной деятельностью называют регуляцию распределения возбуждения и торможения в нейронных структурах, а также взаимодействия нервных центров, обеспечивающие адекватные рефлекторные и произвольно вызываемые реакции.
12.Строение и функции спинного мозга. Основные рефлексы спинного мозга.
Спинной мозг – это орган, относящийся к центральной нервной системе. Спинной мозг расположен в спинномозговом канале, который образован телами и дужками позвонков. Однако, спинной мозг заполняет собой не весь спинномозговой канал, и продолжается он до 2 – 3 поясничного позвонка. Далее в качестве продолжения мозга идет так называемая спинномозговая нить.
Снаружи спинной мозг покрыт, как и головной, оболочками: мягкой и твердой. Между этими оболочками находится спинномозговая жидкость – это тот же ликвор.
Как и головной мозг. Спинной так же состоит из серого и белого вещества. однако, в отличие от головного мозга, в спинном мозге серое вещество находится внутри белого.
Для удобства спинной мозг разделен на сегменты (как и головной на доли и извилины), однако на глаз они неразличимы.
Спинной мозг выполняет две основные функции: рефлекторную и проводниковую. Рефлекторная функция спинного мозга обеспечивает движение. Через спинной мозг проходят рефлекторные дуги, с которыми связано сокращение мышц тела (кроме мышц головы). Пример простейшего двигательного рефлекса — коленный рефлекс. Спинной мозг вместе с головным мозгом регулирует работу внутренних органов: сердца, желудка, мочевого пузыря, половых органов.
Белое вещество спинного мозга обеспечивает связь и согласованную работу всех отделов центральной нервной системы, осуществляя проводниковую функцию. Нервные импульсы, поступающие в спинной мозг от рецепторов, передаются по восходящим проводящим путям в головной мозг. Из головного мозга импульсы по нисходящим проводящим путям поступают к нижележащим отделам спинного мозга и оттуда — к органам.
Примерами спинномозговых рефлексов могут служить:
-Сгибательный (флексорный) рефлекс — рефлекс защитного типа направленный на удаление повреждающего раздражителя (отдергивание руки от горячего).
-Рефлекс на растяжения (проприоцептивный) — предотвращающий чрезмерное растяжение мышцы. Особенностью этого рефлекса является, то что рефлекторная дуга содержит минимум элементов — мышечные веретена генерируют импульсы которые проходят в спинной мозг и вызывают моносинаптическое возбуждение в α-мотонейронах той же мышцы.
-Сухожильный, разнообразные тонические и ритмические рефлексы.
-У четвероногих животных можно наблюдать экстензорный толчок.
studfiles.net
2.1.2. Свойства нервных центров
Интеграция нервных процессов и рефлекторная деятельность ЦНС, лежащие в основе приспособительных реакций организма, во многом определяется общими свойствами нервных центров:
односторонним проведением возбуждения,
замедленным проведением возбуждения,
низкой лабильностью,
повышенной утомляемостью,
способностью к иррадиации,
способностью к суммации,
последействием (пролонгированием),
трансформацией ритма,
высокой пластичностью,
способность к тонической активности,
повышенной чувствительностью к недостатку питательных веществ и кислорода.
Одностороннее проведение возбуждения – это способность нервных центров проводить возбуждение только в одном направлении — от афферентов к эфферентам.
Если раздражать электрическим током афферент, то в эфферентных нервных волокнах возникает серия ПД. Однако, при раздражении эфферента, в афферентных волокнах возбуждение не возникает. Одностороннее проведение сигналов обусловлено возможностью передачи возбуждения в центральных химических синапсах только от пресинаптической мембраны к постсинаптической.
Схема эксперимента, доказывающего одностороннее проведение возбуждения в нервном центре
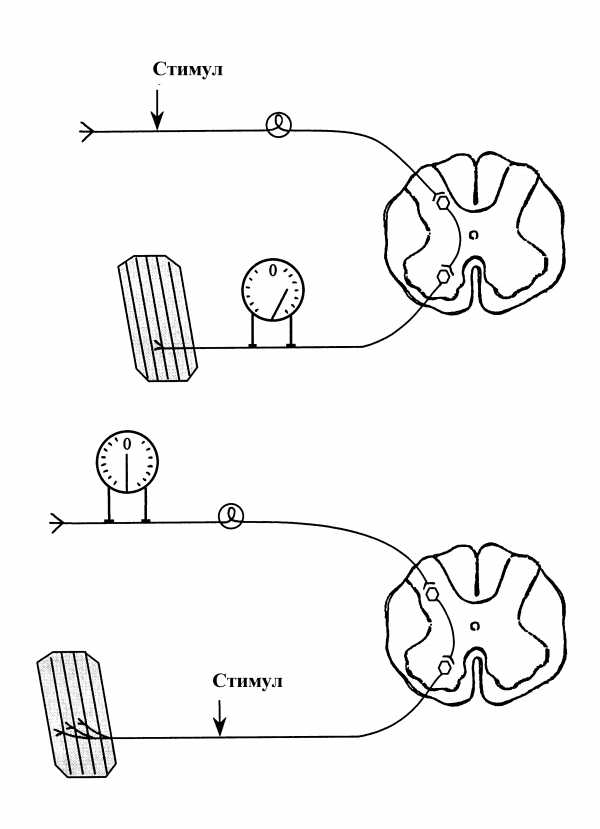
Замедленное проведение возбуждения — это свойство нервных центров, передавать возбуждение с низкой скоростью.
Между моментом поступления нервного импульса к окончанию аксона и формированием ВПСП на постсинаптической мембране проходит около 0,5 мс. Время от поступления нервного импульса к окончанию аксона до формирования ВПСП называется синаптической задержкой.
Низкая лабильность — это свойство нервных центров, характеризующееся относительно небольшой частотой импульсации, которую они способны воспроизвести без искажения ритма стимуляции афферента.
Лабильность нервных центров не превышает 50-70 ПД/с. Это обусловлено синаптической задержкой передачи возбуждения в центральных химических синапсах.
Повышенная утомляемость — это свойство нервных центров, которое проявляется в постепенном снижении или полном прекращении рефлекторного ответа при продолжительном раздражении афферентных волокон.
Снижение работоспособности нервных центров связано с нарушением передачи возбуждения в межнейронных химических синапсах и обусловлено повышенной утомляемостью центральных химических синапсов.
Иррадиация – это свойство нервного центра передавать возбуждение в ЦНС на другие нервные центры при достаточно сильном воздействии раздражителя на рефлексогенную зону.
Благодаря распространению возбуждения на несколько нервных центров рефлекторная реакция приобретает генерализованный характер. Иррадиации препятствуют процессы торможения, которые в нормальных условиях ограничивают распространение возбуждения в ЦНС.
При заболеваниях зубов афферентная импульсация от них существенно возрастает, что приводит к генерализованному распространению возбуждения. Иррадиация возбуждения при интенсивной зубной боли (например, при пульпите), приводит к тому, что пациент не в состоянии точно указать больной зуб.
В основе иррадиации возбуждения в ЦНС лежит морфологический принцип дивергенции (дихотомического ответвления аксонов нейронов), который заключается в способности клеток нервного центра устанавливать многочисленные синаптические контакты с нейронами других нервных центров.
Схема иррадиации возбуждения в ЦНС
1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентные нейроны.
Способность к суммации — это свойство нервных центров формировать рефлекторную реакцию при действии на рецепторы раздражителей, которые являются подпороговыми для нервного центра.
Различают два вида суммации: последовательную (временную) и одновременную (пространственную). Последовательная суммация наблюдается при взаимодействии возбуждений, которые поступают в нервный центр с коротким интервалом времени друг за другом по одним и тем же афферентам.
Если на рецептор действует один пороговый для рецептора и афферента, но подпороговый для возбуждения нервного центра раздражитель, то разрядная деятельность в области аксонного холмика центрального нейрона, а значит и рефлекторный ответ, не возникают. Однако, при последовательном действии серии таких раздражителей, в центральном синапсе увеличивается выделение медиатора. Вследствие этого на постсинаптической мембране происходит суммация ВПСП и начинается циркуляция местных ионных токов, сила которых достаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и для возникновения ответной рефлекторной реакции эффектора.
Примером проявления подобной суммации может служить чихание, которое возникает рефлекторно в результате длительной стимуляции рецепторов слизистой оболочки носа, накопившейся слизью, частицами пыли или другими раздражающими факторами.
Пространственная суммация возникает при взаимодействии возбуждений, одновременно приходящих в нервный центр по нескольким афферентам. При одновременном действии раздражителя на несколько рецепторов одного рецептивного поля, сигналы от которых конвергирует к одному нейрону, увеличивается выделение медиатора в центральных синапсах. Вследствие этого происходит суммация ВПСП на постсинаптической мембране, и начинают циркулировать местные ионные токи, сила которых достаточна для генерации серии эфферентных ПД в области аксонного холмика, что приводит к рефлекторному ответу исполнительного органа.
Последействие — это способность нервного центра продлевать рефлекторный ответ после прекращения раздражения рецепторов.
Известны два основных механизма последействия: кратковременный и долговременный. Кратковременное рефлекторное последействие связано со следовой деполяризацией постсинаптической мембраны нейрона после прекращения раздражения афферента. После прекращения раздражения рецепторов на постсинаптической мембране центрального синапса сохраняется следовая деполяризация в виде ВПСП, который поддерживает высокую возбудимость эфферентного нейрона. В это время за счет теплового движения молекул в синаптическую щель может самопроизвольно выделиться нескольких квантов медиатора, что приводит к образованию миниатюрного ВПСП, который суммируется со следовым потенциалом. В результате этого продолжается циркуляция местных ионных токов и поддерживается разрядная деятельность эфферентного нейрона, а следовательно и рефлекторная реакция.
За счет кратковременного механизма разрядная деятельность нейрона после прекращения раздражения афферента продлевается на несколько десятков мс. По мере уменьшения амплитуды ВПСП импульсная активность нейрона постепенно уменьшается, а затем прекращается совсем.
Долговременное рефлекторное последействие обусловлено реверберацией — циркуляцией возбуждения по замкнутым нейронным цепям в нервных центрах.
После прекращения раздражения возбуждение продолжает циркулировать по замкнутым нейронным цепям-«ловушкам», возвращаясь к одному и тому же эфферентному нейрону. Это обеспечивает поддержание на высоком уровне его возбудимости и продолжение разрядной деятельности. Благодаря нейронным «ловушкам» возбуждение может длительно циркулировать в нервном центре до тех пор, пока не наступит утомления одного из синапсов или же активность нейронов не будет приостановлена процессом торможения.
Схема организации нейронной ловушки
1) афферент, 2) эфферентный нейрон, 3) коллатераль, 4) вставочные нейроны.
Трансформация ритма — это свойство нервных центров изменять частоту проходящих через них импульсов.
Частота импульсации при прохождении через нервный центр может увеличиваться или уменьшаться. Способность нервных центров увеличивать частоту проходящих через них импульсов, называют мультипликацией ритма. Выделяют два основных механизма мультипликации: кратковременный и долговременный.
Кратковременная мультипликация, как и последействие, связана с суммацией после одиночного стимула на постсинаптической мембране миниатюрного и следового ВПСП.
Долговременная мультипликация обусловлена распространением возбуждения по дихотомически разветвляющимся коллатералям аксонов афферентов и вставочных нейронов, которые затем конвергируют (сходятся) к одному эфферентному нейрону. Вследствие схождения к эфференту нескольких вставочных нейронов, которые связаны с одним афферентом, нервный центр на одиночное раздражение афферента отвечает серией эфферентных ПД.
Схема нейронной организации долговременной мультипликации
1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентный нейрон.
Пластичность — это способность нервных центров изменять свое функциональное назначение в зависимости от особенностей условий деятельности.
Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками. Она является рабочим механизмом обучения. Наибольшей пластичностью обладает кора больших полушарий головного мозга. При повреждении какой-либо зоны коры через некоторое время другие ее участки могут брать на себя выполнение функции утраченного отдела.
Способность к тонической активности заключается в постоянной минимальной импульсной активности нервных центров, которая поддерживается за счет афферентных сигналов от рефлексогенных зон. Тоническая активность нервных центров обеспечивает тонус большинства исполнительных органов и их постоянную готовность к деятельности даже в состоянии относительного покоя.
Повышенная чувствительность к недостатку питательных веществ и кислорода обусловлена высоким уровнем метаболизма нервной ткани. Специфичность обмена веществ в нейронах состоит в исключительности использования в качестве питательного материала глюкозы, тогда как другие клетки используют также различные моносахариды, жирные кислоты и аминокислоты. Кроме того, в нейронах преобладает окислительное фосфорилирование, требующее непрерывной доставки О2.
При относительно небольшой массе, составляющей не более 2% от веса тела, мозг человека потребляет до 20% всего поступившего в организм кислорода и 17% глюкозы. Интенсивность потребления глюкозы настолько велика, что в нейронах не успевают образоваться запасы гликогена. Поэтому уменьшение доставки глюкозы к ЦНС быстро влечет за собой нарушение функции нервных центров.
Интенсивность потребления нервными клетками кислорода составляет около 50 мл в мин. Потребляя такое большое количество О2, мозг и особенно кора больших полушарий высокочувствительны к его недостатку. Поэтому уже через 5-7 секунд после прекращения кровообращения в мозге человек теряет сознание, а через 5-6 минут нервные клетки больших полушарий подвергаются необратимым биологическим изменениям.
Центры ствола мозга менее чувствительны к недостатку кислорода: их функция может восстановиться после 20 минут полного прекращения кровообращения. Центры спинного мозга еще более выносливы. Их функция восстанавливается даже через 30 минут после полного прекращения притока крови.
При гипотермии — понижении температуры тела, вследствие снижения уровня обмена веществ, ЦНС дольше переносит недостаток О2 и питательных веществ. Гипотермия при температуре тела 25-280С повышает устойчивость коры больших полушарий к кислородной недостаточности с 5 до 15-20 минут.
studfiles.net
Свойства нервных центров — Мегаобучалка
Нервные центры обладают большими возможностями, чем один нейрон. Это проявляется в их свойствах. Во многом эти свойства определяются наличием синапсов. Нервные центры обладают односторонностью проведения возбуждения.
Возбуждение в нервном центре проводитсяот афферентного нейрона к эфферентному. Нервный центр обладает трансформацией ритма; трансформация может быть повышающей (на один импульс нервный центр может ответить «автоматной очередью») или понижающей (в ответ на серию возбуждений нервный центр дает одиночный импульс или не отвечает вовсе). В нервном центре обнаружено последействие.Это свойство связано, вероятно, с наличием в нервных центрах нейронных цепей с положительной обратной связью и с таким свойством синапсов, как постсинаптическая потенциация. Последействие проявляется в продолжающемся рефлекторном процессе при отсутствии раздражителя, вызвавшего эту реакцию. Нервному центру характерна высокая чувствительность к недостатку кислорода. Это связано с интенсивными метаболическими процессами в нейронах. Нервный центр избирательно чувствителен к различным биологически активным веществам, ядам и другим химическим веществам. Это свойство называется хемотропностью. Хемотропность определяется специфическим набором медиаторов, ферментов, нейропептидов и других веществ, которые синтезируются системой нейронов, входящей в конкретный нервный центр. Нервные центры обладают низкой лабильностью и быстрой утомляемостью,а также замедлением проведения возбуждения через нервный центр.Эти свойства связаны с наличием в нервных центрах огромного числа синапсов. Нервные центры всегда находятся в тонусе.Тонус нервных центров (определенный уровень возбудимости) поддерживается притоком различной афферентации из периферии и поступлением возбуждения из нервных центров высших отделов ЦНС. Понижение или повышение тонуса нервных центров лежит в основе регуляции и организации функций.
5. Принципы координационной деятельности
центральной нервной системы
Принцип рефлекса
Принцип рефлекса – это универсальная и своеобразная форма взаимодействия организма со средой, происходящая при участии нервной системы. Понятие о рефлексе возникло в XVI веке в учении
Р. Декарта (1596-1650) о механической картине мира. Р. Декарт жил в эпоху расцвета механики, физики и математики. На его мировоззрение определяющее влияние оказало открытие У. Гарвеем механизма кровообращения и новаторские идеи А. Везалия о том, что носителями психики являются «животные духи», которые вырабатываются в желудочках мозга и передаются по нервам к соответствующим органам. Р. Декарт представлял нервные процессы по образцу системы кровообращения, используя существовавшие в то время принципы оптики и механики.
Под рефлексом Р. Декарт понимал движение «животных духов» от мозга к мышцам по типу отражения светового луча. Согласно его схеме внешние предметы действуют на периферические окончания расположенных внутри нервных «трубок» нервных «нитей», которые, натягиваясь, открывают клапаны отверстий, ведущих из мозга в нервы. По каналам этих нервов «животные духи» перемещаются в соответствующие мышцы, которые в результате раздуваются, и, таким образом, происходит движение. Однако, находясь под влиянием социально-исторических противоречий своей эпохи, Р. Декарт сделал серьезные уступки идеализму: сознание человека он рассматривал в виде субстанционного начала, способного воздействовать посредством мозговой шишковидной железы на подчиненные рефлекторным законам телесные процессы. Согласно Р. Декарту, тело и душа – самостоятельные субстанции. Поведение и сознание разводились, превращались в два независимых ряда явлений. В этом проявился дуализм Р. Декарта.
Биологическая концепция рефлекса была сформирована чешским анатомом и физиологом Йиржи Прохазкой(1749-1820). Свои представления о рефлексе Й. Прохазка выразил следующим образом: внешние впечатления, возникающие в чувствительных нервах, быстро распространяются по всей их длине до самого начала. Там они отражаются по определенному закону, переходят на соответствующие им двигательные нервы и по ним очень быстро направляются к мышцам, которые затем производят точные и строго ограниченные движения. Впервые термин «рефлекс» был введен в научный язык Й. Прохазкой. В отличие от Р. Декарта, Й. Прохазка утверждает монистическое представление о нервной системе, в целом относящейся к композиции «общего чувствилища», телесная часть которого локализуется в спинном мозге, а душевная – в головном.
Большой вклад в развитие рефлекторной теории сделали английский анатом и врач Чарльз Белл(1774-1842) и французский физиолог Ф. Мажанди(1783-1855). Они экспериментально показали наличие анатомической основы рефлекса. Так, Ч. Белл в 1811 году в трактате «О новой анатомии мозга» писал, что можно произвести перерезку заднего пучка нервов, исходящего из задней части спинного мозга, без конвульсивных сокращений мышц спины. Однако это становится невозможным даже при одном прикосновении кончиком ножа к переднему корешку. Переход нервного возбуждения по афферентным нервам через спинной мозг на эфферентные нервы получил название закона Белла-Мажанди. Ч. Белл создал теорию о «мышечной чувствительности» и сформулировал физиологическое обоснование циклической функции нервной системы. Между мозгом и мышцей имеется замкнутый нервный круг: один нерв передает влияние от мозга к мышце, а другой передает в мозг чувство состояния мышцы. Если круг будет разомкнут перерезкой моторного нерва, то исчезнет движение. Если же он размыкается перерезкой чувствительного нерва, пропадает ощущение самой мышцы, а вместе с тем исчезает и регуляция ее деятельности. Так, например, у женщины на одной руке была потеряна чувствительность, а на другой – способность к движению. Она могла держать ребенка на руке, потерявшей только чувствительность, до тех пор, пока на него смотрела. Как только женщина отводила взор от ребенка, сразу же возникала опасность его падения на пол.
Рефлексы спинного мозга широко использовались клиницистами, среди которых наиболее значительными фигурами были английский врач Маршал Холл и немецкий физиолог Иоганес Мюллер. Именно М. Холлу принадлежит термин «рефлекторная дуга». Согласно его учению, рефлекторная дуга состоит из афферентного нерва, спинного мозга и эфферентного нерва. М. Холл и И. Мюллер настаивали на принципиальном отличии работы спинного мозга от головного. По их мнению, рефлекторный механизм свойственен только спинному мозгу. Головной мозг оказывался все дальше от сферы влияния физиологии.
Следующий этап развития рефлекторной теории связан с работами великого русского физиолога И.М. Сеченова(1829-1905). Само понятие о рефлекторном характере нервной деятельности у И.М. Сеченова претерпело существенные изменения. Основные положения его теории заключаются в следующем.
1. Рефлекс им понимался как универсальная и своеобразная форма взаимодействия организма со средой, опирающаяся на эволюционную биологию. И.М. Сеченов поставил вопрос о существовании двух родов рефлексов. Он выделил, во-первых, рефлексы постоянные врожденные, осуществляемые низшими отделами нервной системы, которые назвал «чистыми» рефлексами; во-вторых, рефлексы головного мозга изменчивые, приобретенные в индивидуальной жизни. Последние И.М. Сеченов представлял себе одновременно и физиологическим, и психическим явлением.
Таким образом, была впервые показана неотделимость психических процессов от мозга и одновременно обусловленность психики внешним миром. Важнейшим для И.М. Сеченова было положение о единстве организма и условий внешней среды. Большое значение в становлении рефлексов он придавал факторам эволюции.
2. Физиологический субстрат рефлекторных актов характеризуется как нейродинамика, отличная от динамики других систем. Открытие центрального торможения И.М. Сеченовым в 1862 году было первым шагом к созданию им новой физиологии головного мозга. Деятельность нервных центров мыслится ученым как непрерывная динамика процессов возбуждения и торможения.
3. На первый план ставятся межцентральные координационные отношения. Физиологическому анализу начинают подвергаться высшие мозговые центры. Если до И.М. Сеченова усиление или угнетение рефлекторных реакций трактовалось усилием воли, сознания, разума, то И.М. Сеченов переводит все это на строгий физиологический язык и показывает, как центры головного мозга могут задерживать или усиливать спинальные рефлексы.
4. Функции мозговых центров трактуются в широком плане биологической адаптации. Центры влияют усиливающим или тормозным образом на движения не потому, что высвобождается заложенная в них «психическая сила», и не потому, что укорачивается или удлиняется путь прохождения нервного импульса. И.М. Сеченов вводит понятие «физиологического состояния центра», которое непосредственно связано с биологическими потребностями. Само же состояние центра, отражая характер отношений со средой, представляет собой нервный субстрат потребности. Таким образом, в учение о рефлексах вносится существенное дополнение. Реакция становится в непосредственную зависимость не только от наличных раздражений, но и от всей суммы прежних воздействий, оставивших длительно сохраняющиеся следы в нервных центрах.
5. И.М. Сеченов считает, что мышечное чувство при выполнении одного движения становится в порядке ассоциации рефлексов сигналом для другого движения. Принцип ассоциации рефлексов лежит в основе обучения человека сложным формам трудовой деятельности. Устанавливается общий характер для движений и для психической деятельности – это наличие мышечной чувствительности.
По вопросу соотношения физиологического и психического И.М. Сеченов занял совершенно определенную позицию, которая отражена в его словах: «Для нас, как физиологов, достаточно и того, что мозг есть орган души, то есть такой живой механизм, который, будучи приведен какими ни на есть причинами в движение, дает в окончательном результате тот же ряд внешних явлений, которыми характеризуется психическая деятельность».
Для большей убедительности своих взглядов И.М. Сеченову не хватало экспериментального подтверждения.
Блестящие догадки, предвидения и мысли И.М. Сеченова экспериментально подкрепил И.П. Павлов. Он создал научную концепцию условного рефлекса, которая благодаря его блестящим экспериментам была введена в строгие рамки лабораторного опыта. Вот основные элементы рефлекторной теории И.П. Павлова.
1. Прежде всего был создан лабораторный метод объективного изучения приспособительной деятельности человека и животных – метод условных рефлексов.
2. Изучая условные рефлексы на целостном организме, И.П. Павлов подчеркивал их приспособительно-эволюционный смысл для животного мира.
3. И.П. Павлов предпринял попытку локализовать сам нервный процесс замыкания нервных связей в коре головного мозга у высших животных и человека. При этом он не исключал специфического участия других отделов мозга в этом процессе.
4. И.П. Павлов констатировал наличие в коре головного мозга процесса торможения, чем подкрепил сеченовские идеи о тормозящем влиянии головного мозга.
5. Было четко сформулировано учение о физиологии анализаторов, под которыми И.П. Павлов, как и И.М. Сеченов, мыслил конструкции, состоящие из периферических рецепторов, проводящих путей и мозговых центров, вплоть до коры больших полушарий.
6. И.П. Павлов сформировал представление о коре головного мозга как мозаике из возбуждений и торможений.
7. В конце творческой жизни И.П. Павлов выдвинул принцип системности в работе коры головного мозга, способной формировать динамический стереотип деятельности, уже в какой-то мере независимый от качества внешних раздражений.
Дальнейшее развитие рефлекторного принципа происходит в работах А.А. Ухтомского и П.К. Анохина.
А.А. Ухтомский подчеркивал: «… рефлекс – это такая реакция, которая мотивируется вполне отчетливо текущей обстановкой или средой. Этим, однако, не уничтожается спонтанное действие субстрата, оно лишь ставится в определенные границы в его противоположении факторам среды, и от этого оно становится более определенным по поддержанию и значению. Рефлекс рисуется не чисто пассивным передвижением шарика под влиянием полученного им удара извне, так рефлекс мог изображаться, пока надо было подчеркнуть в особенности его мотивировку из среды. Но в полноте своей он представляется встречею во времени двух условий: с одной стороны, деятельности заготовленной, или сложившейся в самом субстрате (клетке) за предыдущую его историю, и, с другой стороны, внешних импульсов текущего момента». Принципы историзма и соотношения реактивности и активности в целостном поведении как способе разрешения противоречий, принесенные в физиологию А.А. Ухтомским, существенно обогатили рефлекторную теорию, которая окончательно избавилась от дуализма и механицизма картезианского толка, встав на твердые диалектические позиции.
П.К. Анохинрассматривал поведение как системный процесс. В основе организации поведения, согласно П.К. Анохину, лежат функциональные системы. Созданием теории функциональных систем П.К. Анохин определил дальнейшее развитие рефлекторного принципа.
Определение рефлекса
megaobuchalka.ru
Свойства нервных центров
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие, особенности:
1. однострочное проведение, оно идет от афферентного, через вставочный к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2.Центральная задержка проведения возбуждения т.е по НЦ возбуждения идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой т.к больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого, время рефлекса, это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Эго объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются 11 и мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения — изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне. Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения
нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие- это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров — состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессор метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов.
biofile.ru