
Синаптическая щель – Синапс — Википедия
Синапс нервной клетки. Анатомия мозга. Псеунок А.А..
Синапс – (от греческого synapsis — соприкосновение, связь) — место контакта двух нейронов или нейрона и мышцы.
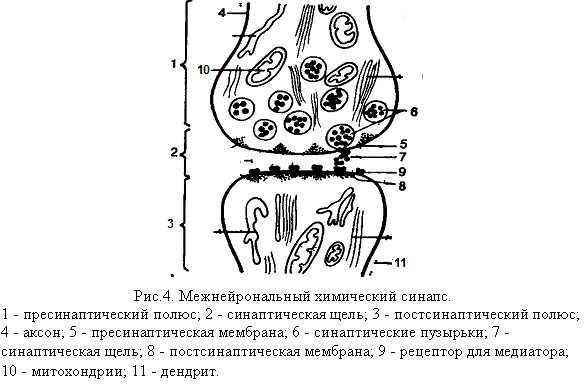
Межнейрональный химический синапс
Синапс состоит из 2-х мембран, соприкасающихся друг с другом: одна из них принадлежит разветвлению аксона одного нейрона, а другая – дендриту другого нейрона (рис. 4.). При исследовании синапса под электронным микроскопом ясно видна граница контактирующих друг с другом нейронов. На этой границе чётко вырисовываются две мембраны – пресинаптическая и постсинаптическая, отделённые друг от друга синаптической щелью. В центральной нервной системе синаптическая щель является непосредственным продолжением межклеточного пространства, их содержимое сообщается друг с другом. Ширина синаптической щели – от 2 до 30 нм, диаметр синаптического контакта – от 0,1 до 10 мкм.
Синаптическая щель – промежуток, разделяющий пресинаптическую мембрану аксона одной клетки и постсинаптическую мембрану тела или дендрита нейрона другой клетки или мышцы.
Пресинаптическая мембрана является продолжением поверхностной мембраны аксонального окончания, глиальные элементы не участвуют в образовании синапсов. Эта мембрана не сплошная, она имеет отверстия, через которые цитоплазма аксональных окончаний сообщается с синаптическим пространством. Постсинаптическая мембрана менее плотная, чем пресинаптическая, она не имеет отверстий. Толщина каждой из мембран синапса не превышает 5-6 нм. Несколько иначе построены органные синапсы, например, в области нервно-мышечного соединения. На поверхности мышечного волокна имеется углубление со множеством ветвящихся и взаимодействующих между собой складок, в которых размещаются разветвления аксона. Здесь также различаются пресинаптическая (аксональная) и постсинаптическая (мышечная) мембраны. Обе мембраны состоят из нескольких слоев, толщина каждой – около 10 нм; пространство между мембранами заполнено сильно гидратированным гелем.
Синапсы бывают двух видов – возбудительные и тормозные, с их помощью происходит соответственно передача или блокада нервного импульса.
Основной функцией синапса является передача возбуждения с одной нервной клетки на другую, либо с нейрона на эффекторный орган. По современным данным, в большинстве синапсов передача возбуждения осуществляется посредством
www.braintools.ru
подразделение, электрические и химические, работоспособность детей
 Что такое синапс? Синапс – это особая структура, предоставляющая передачу сигнала от волокон нервной клетки на прочую клетку или волокно от контактной клетки. Для чего необходимо наличие 2 нервных клеток. При этом синапс представлен в 3 функциональных участках (предсинаптический фрагмент, синаптическая щель и постсинаптический фрагмент) нервных клеток и располагается в области, где клетка контактирует с мышцами и железами человеческого организма.
Что такое синапс? Синапс – это особая структура, предоставляющая передачу сигнала от волокон нервной клетки на прочую клетку или волокно от контактной клетки. Для чего необходимо наличие 2 нервных клеток. При этом синапс представлен в 3 функциональных участках (предсинаптический фрагмент, синаптическая щель и постсинаптический фрагмент) нервных клеток и располагается в области, где клетка контактирует с мышцами и железами человеческого организма.
Подразделение синапсов
Система нейронных синапсов осуществляется по их локализации, типу деятельности и методу транзита имеющихся сигнальных данных. Относительно локализации синапсы различают: нейронейрональные, нервно-мышечные. Нейронейрональные на аксосоматические, дендросоматические, аксодендритические, аксоаксональные.
По типу деятельности на восприятие синапсы принято выделять: возбуждающие и не менее важные тормозящие. Относительно метода транзита информационного сигнала классифицируют их на:
- Электрический тип.
- Химический тип.
- Смешанный тип.
Этиология контактирования нейронов сводится к типу этого стыкования, которое может быть дистантным, контактным, а также пограничным. Соединение дистантного свойства выполняется посредством 2 нейронов, размещённых во многих частях организма.
Так, в тканях человеческого мозга генерируются нейрогормоны и вещества-нейропептиды, влияющие на присутствующие нейроны организма другого местоположения. Контактное соединение сводится к особым стыковкам плёнок-мембран типичных нейронов, составляющих синапсы химического направления, а также составляющих электрического свойства.
Смежная (пограничная) работа нейронов производится во время, в течение которого плёнки-мембраны нейронов перегорожены лишь синаптической щелью. Как правило, такое слияние наблюдается, если между 2 специальными плёнками-мембранами
Существует мнение, что смежный контакт провоцирует работу рядом расположенных нейронов в произведении общей функции. Это наблюдается по причине того, что метаболиты, плоды действия человеческого нейрона, проникая внутрь полости, расположенной, между клетками оказывают влияние на близлокализующиеся активные нейроны. Причём пограничное соединение часто может передавать данные электрического характера от 1 рабочего нейрона к 2 участнику процесса.
Синапсы электрического и химического направления
 Действие слияния плёнок-мембран принято считать электрическими синапсами. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 — 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Действие слияния плёнок-мембран принято считать электрическими синапсами. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 — 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Помимо отдельчатых типов синапсов существуют необходимые электрические типичные синапсы в виде единой синаптической щели, общий периметр которой простирается на 1000 мкм. Так, подобное синаптическое явление представлено в нейронах ресничного ганглия.
Электрические синапсы способны проводить качественное возбуждение в одностороннем порядке. Этот факт отмечается при фиксации электрического резерва синаптической составляющей. Например, в момент при касании афферентных канальцев синаптическая плёнка-мембрана деполяризуется, когда с касанием эфферентных частиц волокон напортив — гиперполяризуется. Считается, что синапсы действующих нейронов с общими обязанностями могут осуществлять требуемое возбуждение (между 2 пропускающими участками) в обе стороны.
Напротив, синапсы присутствующих нейронов с разным перечнем действий (моторные и сенсорные) проводят акт возбуждения односторонне. Основная работа синаптических составляющих обуславливается продуцированием безотлагательных реакций организма. Электрический синапс подлежит незначительной доли утомляемости, обладает значительным процентом устойчивости к внутренне-наружным факторам.
Химические синапсы обладают видом предсинаптического сегмента, функциональной синаптической щели с фрагментом постсинаптической составляющей. Предсинаптический фрагмент формируется увеличением размера аксона внутри собственного канальца или к его завершению. В этом фрагменте присутствуют гранулярные, а также агранулярные специальные мешочки, содержащие медиатор.
Предсинаптическое увеличение наблюдает локализацию активных митохондрий, генерирующую частицы вещества-гликогена, а также требуемую выработку медиатора и другое. В условиях частого соприкосновения с предсинаптическим полем резерв медиатора в имеющихся мешочках утрачивается.
Существует мнение, что малые гранулярные пузырьки имеют такое вещество, как норадреналин, а большие – катехоламины. Причём в агранулярных полостях (пузырьках) располагается ацетилхонин. Помимо этого, медиаторами усиленного возбуждения считаются вещества, образованные по типу вырабатываемой аспарагиновой или не менее значимой кислоты глутамина.
Действующие контакты синапса часто располагаются между:
- Дендритом и аксоном.
- Сомой и аксоном.
- Дендритами.
- Аксонами.
- Сомой клетки и дендритами.
Влияние выработанного медиатора относительно присутствующей постсинаптической плёнки-мембраны происходит из-за чрезмерного проникновения её частиц натрия. Генерация мощных изливаний частиц натрия из рабочей синаптической щели сквозь постсинаптическую плёнку-мембрану формирует её деполяризацию, образуя возбуждение постсинаптического резерва. Транзиту химического направления данных синапса свойственно синаптическое приостановление возбуждения по времени равному 0,5 мс с выработкой постсинаптического резерва, как реакция на предсинаптический поток.
Данная возможность в момент возбуждения представляется в деполяризации постсинаптической плёнки-мембраны, а в момент приостановления в её гиперполяризации. Из-за чего наблюдается приостановленный постсинаптический резерв. Как правило, во время сильного возбуждения повышается уровень проницаемости постсинаптической плёнки-мембраны.
Требуемое возбуждающее свойство фиксируется внутри нейронов, если в типичных синапсах работает норадреналин, вещество-дофамин, ацетил холин, важный серотонин, вещество Р и кислота глутамина.
Сдерживающий потенциал формируется во время влияния на синапсы из гамма-аминомаслянной кислоты и глицина.
Умственная работоспособность детей
 Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Точность и скорость умственных действий с возрастом осуществляется неравномерно в зависимости от прочих факторов, фиксирующих развитие и физический рост организма. Учащимся любого возраста, у которых присутствуют отклонения здоровья, характерна работоспособность низкого значения относительно окружающих крепких детей.
У здоровых первоклашек со сниженной готовностью организма к постоянному процессу обучения по некоторым показателям способность к действию является низкой, что усложняет борьбу с возникающими проблемами в процессе учёбы.
Скорость наступления ослабленности обуславливается исходным состоянием детской системы чувствительного нервного генеза, рабочим темпом и объёмом нагрузки. При этом дети склонны к переутомлению во время продолжительной неподвижности и когда выполняемые действия ребёнку неинтересны. После перерыва работоспособность становится прежней или становится выше прежней, причём лучше отдых делать не пассивным, но активным, переключившись на отличное от этого занятие.
Изменение работоспособности у детей
 Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
- Они глядят в окно.
- Невнимательно слушают слова учителя.
- Изменяют положения своего тела.
- Начинают разговаривать.
- Встают со своего места.
Специфически велики значения работоспособности у старшеклассников, обучающихся во 2 смену. Особенно важно обратить внимание на то, что достаточно коротко время для подготовки к занятиям до времени начала учебного действия в классе и не гарантирует полноценного избавления от пагубных изменений в центральной нервной системе. Умственная активность быстро истощается в первые часы уроков, что явно отмечается в отрицательном поведении.
Посему качественные сдвиги работоспособности наблюдаются у учеников младшего блока на уроках с 1 — 3, а блоках среднего-старшего звена на 4 — 5 занятии. В свою очередь, 6 урок проходит в условиях особенно сниженной способности к действию. При этом продолжительность занятия у 2 — 11 классников – 45 минут, что ослабляет состояние детей. Поэтому рекомендуется периодически сменять вид работы, а в середине урока провести активную паузу.
psiho.guru
синаптическая щель — это… Что такое синаптическая щель?
- синаптическая щель
- (fissura synaptica, LNH) пространство, разделяющее пресинаптическую и постсинаптическую мембраны синапса.
Большой медицинский словарь. 2000.
- синаптическая передача
- синаптический пузырек
Смотреть что такое «синаптическая щель» в других словарях:
синаптическая щель — Полость между двумя соседними нейронами [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN synaptic cleftsynaptic gap … Справочник технического переводчика
Синаптическая щель — – малое (приблизительно 20 30 нм) пространство между терминальной бляшкой пресинаптического нейрона и оболочкой постсинаптического нейрона, через которое проходят нейротрансмиттеры … Энциклопедический словарь по психологии и педагогике
СИНАПТИЧЕСКАЯ ЩЕЛЬ — Малое (приблизительно 20 30 нм) пространство между терминальной бляшкой пресинаптического нейрона и оболочкой постсинаптического нейрона, через которую проходят нейромедиаторы … Толковый словарь по психологии
Синаптическая щель — – пространство между пре и постсинаптической мембранами; синапс, см … Словарь терминов по физиологии сельскохозяйственных животных
Синаптическая пластичность — Для улучшения этой статьи желательно?: Добавить иллюстрации. Проставить для статьи более точные категории. Синаптическая пластичност … Википедия
Синапс — I Синапс (греч. synapsis соприкосновение, соединение) специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически С. образован… … Медицинская энциклопедия
Синапс — Основные элементы синапса Синапс[1] (греч … Википедия
СИНАПСЫ — (от греч. synapsis соединение, связь), специализир. функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. Термин «С.» ввёл Ч. Шеррингтон в 1897. Синаптич … Биологический энциклопедический словарь
синапс — а; м. [греч. synapsis соединение, связь] Физиол. Область соприкосновения нервных клеток друг с другом или тканями, содержащими нервные клетки. ◁ Синапсический; синаптический, ая, ое. С ая зона. * * * синапс (от греч. sýnapsis соединение),… … Энциклопедический словарь
Химический синапс — Химический синапс особый тип межклеточного контакта между нейроном и клеткой мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество. Состоит из трёх основных частей: нервного окончания с… … Википедия
dic.academic.ru
Синаптическая щель — Справочник химика 21
| Рис. 8.2. Схема никотинового холинэргического синапса. Пресинаптическое нервное окончание содержит компоненты для синтеза нейромедиатора (здесь ацетилхолина). После синтеза (I) нейромедиатор упаковывается в пузырьки (везикулы) (II). Эти синаптические везикулы сливаются (возможно, вре.мен-но) с пресинаптической мембраной (1П), и нейромедиатор высвобождается таким путем в синаптическую щель. Он диффундирует к постсинаптической мембране и связывается там со специфическим рецептором (IV). В результате образования нейромедиатор-рецепторного комплекса постсинаптическая мембрана становится проницаемой для катионов (V), т. е. деполяризуется. (Если деполяризация достаточно высока, то появляется потенциал действия, т. е. химический сигнал снова превращается в электрический нервный импульс.) Наконец, медиатор инактивируется , т. е. либо расщепляется ферментом (VI), либо удаляется из синаптической щели посредством особого механизма поглощения . В приведенной схеме только один продукт расщепления медиатора— холин — поглощается нервным окончанием (VII) и используется вновь. Базальная мембрана — диффузная структура, идентифицируемая методом электронной микроскопии в синаптической щели (рис. 8.3,а), здесь не показана. |
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30—80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит 40000 молекул ацетилхолина. При возбуждении высвобождение медиатора происходит квантами , т.е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100—200 квантов медиатора—количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Происходит это, по-видимому, следующим образом. Деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов Са в клетку. Временное увеличение внутриклеточной концентрации ионов Са стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуется примерно 4 иона Са . Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны —резко увеличивается ее пропускная способность для ионов Ка. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации, или удаления, чтобы подготовить синапс к восприятию нового импульса. [c.638]
Объемы выделяемых нейромедиаторов (называемых также и нейротрансмиттерами) невелики. Ацетилхолин, например, выделяется дозами, каждая из которых содержит около 10 ООО молекул. Эти молекулы распределяются по синаптической щели так, что сигнал достигает рецептора. Воздействие ацетилхолина на рецептор вызывает соответствующий физиологический ответ, после чего трансмиттер подвергается разрушению. Ацетилхолин гидролизуется под действием фермента ацетилхолинэстераза (АХЭ). [c.406]
Передача сигналов от клетки к клетке. может осуществляться либо путем прямого прохождения потенциалов действия (электрические синапсы), либо с помощью специальных молекул — нейромедиаторов (химические синапсы). В зависимости от своих специфических функций синапсы имеют очень разные структуры. В химических синапсах расстояние между клетками составляет — 20—40 нм синаптическая щель между клетками— это часть межклеточного пространства она содержит жидкость с низким электрическим сопротивлением, так что электрический сигнал рассеивается прежде, чем он достигнет следующей клетки. Электрическая передача, напротив, осуществляется только в специализированных структурах — щелевых контактах, где клетки находятся на расстоянии 2 нм и соединяются проводящими канала.ми. В действительности здесь имеется нечто сходное с постулированным ранее синцитием, или многоклеточным цитоплазматическим континуумом. По иронии истории нау- [c.188]
Как известно, клетки нервной системы (нейроны) не имеют непосредственного контакта друг с другом. Они разделены синаптическими щелями, через которые сигнал (передаваемый в виде бегущей по нейронной мембране волны поляризации-деполяризации) пройти не может без определенного посредника, называемого нейромедиатором (или нейротрансмиттером). Передача нервного импульса от одного нейрона к другому происходит следующим образом (рис. 3, схема А). По достижении нервным сигналом конца возбужденной клетки (нейрон 1) в ее пресинаптической области синтезируется нейротрансмиттер (АХ), который затем выбрасывается в синаптическую щель и быстро диффундирует к своему рецептору (R), расположенному в постсинаптической мембране покоящейся клетки (нейроне 2). [c.31]
Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (—КН,) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя указанные медиаторы. В последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из синаптической щели в результате вторичного поглощения симпатическими нервами вновь оказавшись в нервном волокне, медиатор, естественно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен. [c.640]
Рис 3 Передача нервного импульса ацетилхолином (АХ) через синаптическую щель Расширение ионофорного канма под действием АХ [c.31]
Как же ацетилхолин попадает в синаптическую щель [c.199]
Каждый пузырек, выбрасывая свое содержимое в синаптическую щель, вызывает изменение мембранного потенциала постсинаптической клетки, и это можно регистрировать с помощью внутриклеточного электрода. Стимуляция [c.97]
Медиатор диффундирует через синаптическую щель и воздействует на постсинаптическую метку, присоединяясь к рецепторным белкам постсинаптической мембраны. [c.101]
Он образует цилиндрический канал, который с одной стороны выступает на 65 А в синаптическую щель, а с другой — пронизывает липидный бцслой мембраны, входя на 15 А внутрь клетки. Этот узкий канал (или пора) расширяется до 20 А при «посадке» на рецептор нейромедиатора (комплекс RAX) за счет резкого уменьшения вращательного (конформационного) движения субъединиц. Увеличение размера канала облегчает прохождение ионов К+ и Na+ через мембрану против электрохимического фадиента. При этом изменяется мембранный потенциал покоящегося нейрона 2, и в нем генерируется нервный импульс. После этого нейромедиатор гидролизуется ацетилхолинэстера-зой до неактивного холина, и ионофорныи канал закрывается. [c.31]
Концы тонких нервных волокон утолщаются в синаптические пуговки, которые образуют контакты с дендритами других нейронов. Как правило, появление нервного сигнала на пресинаптическом конце нейрона стимулирует высвобождение химического нейромедиатора (или нейрогормона). Медиатор проходит через синаптическую щель между двумя клетками (ширина щели 10—50 нм обычно 20 нм) и вызывает деполяризацию постсинаптической мембраны следующего нейрона [c.325]
Во время передачи электрического импульса из пресинаптического нейрона в синаптическую щель выделяется медиатор, который диффундирует в щели к мембране следующего, постсинаптического нейрона и здесь связывается со своим рецептором. Затем медиатор оказывает действие на активность различных ферментов. Одновременно в постсинаптической мембране активируются специальные белки, образующие в мембране каналы, по которым натрий входит в постсннаптический нейрон, а калий выходит наружу. Система возвращается в исходное, невозбужденное, состояние в течение миллисекунды. [c.113]
Многие антвдепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т.е. количество медиаторов для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехол-аминовая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге. [c.641]
Расстояние между пресинаптической и посГсинаптической мембранами—синаптическая щель — может достигать 15—20 нм. В мионевральном соединении разрыв еще больше— до 50—100 нм. В то же время существуют синапсы с сильно сближенными и даже сливающимися пресинаптической и постсинаптической мембранами. Соответственно реализуются два типа передачи. При больших щелях передача является химической, при тесном контакте возможно прямое электрическое взаимодействие. Здесь мы рассмотрим химическую передачу. [c.382]
На рис. 8.2 представлена схема химического синапса. Он состоит из нервного окончания на пресинаптической стороне и специализированной области на поверхности принимающей сигнал клетки на постсинаптической стороне. Пре- и постсинаптические мембраны находятся на расстоянии 20—40 нм. Синаптическая щель, видимо, заполнена олигосахаридсодержащей соединитель—ной тканью — базальной мембраной, представляющей собой поддерживающую структуру для обеих объединенных клеток. [c.189]
В синапсе мембрана мышечной клетки ведет себя как преобразователь, который превращает химический сигнал, т.е. определенную концентрацию нейромедиатора, в сигнал электрический. Это осуществляется с помощью ли-ганд-зависимых ионных каналов, находящихся в постсинаптической мембране. Связывание нейромедиатора с этими каналами с наружной стороны мембраны вызывает изменение их конформации-каналы открываются, пропуская через мембрану ионы и тем самым изменяя мембранный потенциал. В отличие от потенциал-зависимых каналов, ответственных за возникновение потенциалов действия и выделение медиатора, лигаяд-зависимые каналы относительно нечувствительны к изменениям мембранного потенциала (рис. 18-29) и потому не способны к самоуснливающемуся возбуждению типа все или ничего . Вместо этого они генерируют электрический сигнал, сила которого зависит от интенсивности я продолжительности внешнего химического сигнала, т.е. от того, сколько медиатора выводится в синаптическую щель и как долго он там остается. Это свойство лиганд-зависимых каналов важно для обработки информации в синапсах, и мы рассмотрим его позднее. [c.99]
Когда потенциал действия достигает нервного окончания, он вызывает путем деполяризации высвобождение медиатора. Последний диффундирует через синаптическую щель к постсинаптической ме.мбране, вызывая изменения ее ионной проницаемости и, следовательно, ме.мбралного потенциала (гл. 5). Это в свою очередь может приводить к генерации потенциала действия. [c.194]
Ацетилхолин удаляется п синаптической щели путем хщффузии или в результате гидролиза [18] [c.100]
Де Робертис и Беннет в 1955 г. открыли в нервном окончании сферические структуры — так называемые синаптические везикулы (рис. 8.3). Они предположили, что эти структуры действуют как органеллы, содержащие запасенный медиатор, который, хак установили Кастильо и К Ц на основании своих работ по миниатюрным потенциалам концевых пластинок, высвобождается дпскретными квантами при нервном возбуждении, а также спонтанно в состоянии покоя. Постсинаптические потенциалы всегда кратны этому кванту (гл. 5). Синаптические везикулы были выделены, и наличие в них ацетилхолина определено одновременно лабораториями Уиттейкера и Де Робертиса в 1963 г. Остался лишь вопрос, высвобождался ли медиатор непосредственно в синаптическую щель или попадал туда через цитоплазму. Мы еще вернемся к этой проблеме при обсуждении механизма высвобождения медиатора, а здесь опишем только, как ацетилхолин попадает в запасающие его везикулы. [c.198]
Как молекула нейромедиатора, высвобождающаяся из пресинаптической мембраны, достигает постсинаптической мембраны Напрашивается простой ответ — посредством диффузии. Но здесь необходимо объяснить, как медиатор диффундирует мимо многочисленных молекул ацетилхолинэстеразы, которые присутствуют в синаптической щели и теоретически могли бы гидролизовать во много раз большие количества высвобожденного медиатора, сделав, следовательно, невозможным его взаимодействие с постсинаптической мембраной. Предполагается, что этому препятствуют либо структурные особенности вещества синаптической щели — базальной мембраны, которое, возможно, образует каналы, либо временное ингибирование ферментативной активности эстеразы, вероятно, из-за ее взаимодействия с иостспнантической мембраной или из-за насыщения субстратом. Высказано также предположение, что эстераза не присутствует в щели, т. е. на пути диффузии ацетилхолина, а находится в постсинаптической мембране, но такая модель не доказана [8]. [c.201]
Постсинаптический потенциал длится всего несколько миллисекунд, если он не усиливается дополнительным высвобождением молекул медиатора, а концентрация ацетилхолина в синаптической щели уменьшается в результате диффузии и гидролиза. Медиатор инактивируется ферментом ацетилхолинэстеразой (КФ 3.1.1.7), который был выделен в кристаллическом состоянии Нахманзоном [8] и является одним из наиболее часто обновляющихся ферментов. [c.205]
Имеются также данные о наличии Ыа+-зависимого высокоаффинного поглощения, которое отвечает за инактивацию этого медиатора путем его удаления из синаптической щели. Фармакология отдельных стадий серотонинового цикла исследована слабо. Мы уже упоминали здесь механизм действия LSD. п-Хлорофенилаланин представляет собой сильный ингибитор триптофангидроксилазы и благодаря такой специфичности используется для определения участия серотонина в том или ином типе поведения. [c.228]
Цикл медиатора 1) синтез, 2) поглошение везикулами 3) если первое и второе происходят в перикарионе, то экзо-плазматический транспорт к нервным окончаниям, 4) пресинаптическое высвобождение при деполяризации в синаптическую-щель (экзоцитоз), 5) диффузия к постсинаптической мембране, 6) узнавание и связывание специфическим рецептором, например мембранным белком (чтобы включился воротной механизм постсинаптической мембраны), 7) инактивация. [c.238]
Рис 45 Схематичное строение нервно мышечного синапса по С Куфлеру и Дж Николсу (1979) 1 — частички 2 — ямки 3 — синаптические визикулы 4 — пресинаптическая мембрана 5 — постсинаптическая мембрана 6 — складки постсинаптической мембраны 7 — синаптическая щель [c.131]
Сигналы, проводимые нейронами, передаются от одной клетки к другой в особых местах контакта, называемых синапсами (рис. 18-3). Обычно эта передача осуществляется, как это ни странно на первый взгляд, непрямым путем. Клетки электрически изолированы друг от друга пресииаптическая клетка отделена от постсинаптической промежутком-синаптической щелью. Изменение электрического потенциала в пресинаптической клетке приводит к высвобождению вещества, называемого ненромедиатором (или нейротрансмиттером), которое диффундирует через синаптическую щель и вызывает изменение электрофизиологического состояния постсинаптической клетки. Та- [c.73]
| Рис. 18-3. Схема типичного синапса. Элеггричесжий сигнал, приходящий в окончание аксона клетки А, приводит к высвобождению в синаптическую щель химическое го посредника (иейромеднатораХ который вызывает электрическое изменение в мембране деидрита клетки В. Широкая стрелка указывает направление передачи сигнала, Аксон одного нейрона, такого как изображенный на рис. 18-2, образует иногда тысячи выходных синаптических соединений с другими клетками. И наоборот, нейрон может принимать сигналы через тысячи входных синаптических соединений, находящихся на его дендритах и теле. |
chem21.info
Синаптическая щель — это… Что такое Синаптическая щель?
- Синаптическая щель
пространство, разделяющее пресинаптическую и постсинаптическую мембраны синапса.
1. Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982—1984 гг.
- Синапти́ческая переда́ча
- Синапти́ческий пузырёк
Смотреть что такое «Синаптическая щель» в других словарях:
синаптическая щель — Полость между двумя соседними нейронами [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN synaptic cleftsynaptic gap … Справочник технического переводчика
синаптическая щель — (fissura synaptica, LNH) пространство, разделяющее пресинаптическую и постсинаптическую мембраны синапса … Большой медицинский словарь
Синаптическая щель — – малое (приблизительно 20 30 нм) пространство между терминальной бляшкой пресинаптического нейрона и оболочкой постсинаптического нейрона, через которое проходят нейротрансмиттеры … Энциклопедический словарь по психологии и педагогике
СИНАПТИЧЕСКАЯ ЩЕЛЬ — Малое (приблизительно 20 30 нм) пространство между терминальной бляшкой пресинаптического нейрона и оболочкой постсинаптического нейрона, через которую проходят нейромедиаторы … Толковый словарь по психологии
Синаптическая щель — – пространство между пре и постсинаптической мембранами; синапс, см … Словарь терминов по физиологии сельскохозяйственных животных
Синаптическая пластичность — Для улучшения этой статьи желательно?: Добавить иллюстрации. Проставить для статьи более точные категории. Синаптическая пластичност … Википедия
Синапс — I Синапс (греч. synapsis соприкосновение, соединение) специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически С. образован… … Медицинская энциклопедия
Синапс — Основные элементы синапса Синапс[1] (греч … Википедия
СИНАПСЫ — (от греч. synapsis соединение, связь), специализир. функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. Термин «С.» ввёл Ч. Шеррингтон в 1897. Синаптич … Биологический энциклопедический словарь
синапс — а; м. [греч. synapsis соединение, связь] Физиол. Область соприкосновения нервных клеток друг с другом или тканями, содержащими нервные клетки. ◁ Синапсический; синаптический, ая, ое. С ая зона. * * * синапс (от греч. sýnapsis соединение),… … Энциклопедический словарь
Химический синапс — Химический синапс особый тип межклеточного контакта между нейроном и клеткой мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество. Состоит из трёх основных частей: нервного окончания с… … Википедия
dic.academic.ru
СИНАПС — Большая Медицинская Энциклопедия
СИНАПС (греч. sinapsis соединение, связь) — специализированная зона контакта между нервными клетками или нервными клетками и другими возбудимыми образованиями, обеспечивающая передачу приходящей информации с сохранением ее информационной значимости.
Изучение С. как структурного и функционального образования началось в конце прошлого века после того, как испанский гистолог С. Рамон-и-Кахаль установил, что все компоненты нервной системы построены из отдельных нервных клеток разнообразной формы и величины (см. Нервная клетка, Нервная система). В 1897 г. англ. физиолог Ч. Шеррингтон для.обозначения точки контакта окончания аксона с другой нервной клеткой ввел понятие «синапс». Изучение С. было тесно связано со становлением представления о хим. способе передачи возбуждений от клетки к клетке; его придерживались Эллиотт (Th. R. Elliott), О. Леви и Г. Дейл. Дальнейший прогресс в области исследования С. и способов передачи процесса возбуждения (см.) связан с исследованиями 3. Бака, Дж. Ленгли, Фелд-берга (W. Feldberg), Геддама (J. H. Gaddum). Большой вклад в изучение функций С. внесли советские исследователи А. Ф. Самойлов, А. В. Кибяков, А. Г. Гинецинский, X. С. Коштоянц. Физиол. анализ функционирования синаптического аппарата ц. н. с. был проведен Дж. Экклсом и П. Г. Костю ком. Структуре межнейронных связей посвящен ряд фундаментальных исследований С. А. Саркисова (1948), А. Д. Зурабашвили (1951), Г. И. Полякова (1973) и др.
Морфология
С. между двумя нервными клетками состоит из пресинаптического отростка, принадлежащего афферентному нейрону, и постсинаптического компонента, к-рым может быть дендрит, тело или аксон постсинаптического нейрона, мышечная или секреторная ткань. Синаптический контакт между нервными клетками иногда называют также синаптической бляшкой. Пресинаптическое окончание аксона было названо концевой пластинкой, С. между двумя нервными клетками — межнейронными (нейрональными) связями, или нервными С. Окончания аксонов на мышечных волокнах, имеющие черты специализации С., называют мионейрональными связями, пресинаптическое окончание на мышечном волокне — моторной бляшкой, их комплекс — нервно-мышечным соединением, а синаптический контакт аксона с железистой клеткой — нейросекреторным С. Новый этап в развитии учения о С. связан с электронной микроскопией (см.), позволившей изучить ультраструктуру С. и показать, что он представляет собой сложноорганизованный комплекс взаимосвязанных структур.
Пресинаптическое окончание (пресинаптический отросток) и участок постсинаптического нейрона разделены синаптической щелью. В пре-синаптическом окончании содержатся синаптические пузырьки, образующие, как правило, скопления у электронно-плотной пресинаптической мембраны. Сочетание синаптических пузырьков и участков повышенной электронной плотности синаптических мембран получило название активных участков контакта.
Синаптические пузырьки могут быть светлыми, электронно-прозрачными или иметь в центре осмиофиль-ный материал (гранулярные синаптические пузырьки). Изредка в пре-синаптическом отростке встречаются синаптические пузырьки, покрытые дополнительной оболочкой. Размеры синаптических пузырьков колеблются от 20 до 60 нм.
По строению синаптических мембран С. можно разделить на симметричные (с равномерным повышением электронной плотности синаптических мембран) и асимметричные (с преимущественным повышением электронной плотности постсинапти-ческой мембраны).
В большинстве случаев синаптическая щель асимметричных С. заметно шире, чем расстояние между мембранами двух соседних отростков клеток вне С. В синаптической щели С. этого типа имеется нек-рое количество электронно-плотного материала, к-рый иногда может выглядеть как прерывистая линия, идущая между синаптическими мембранами параллельно им. Асимметричные С. чаще располагаются на ветвях и шипиках дендритов, реже на крупных стволах дендритов. Симметричные С., как правило, локализованы на телах нервных клеток и стволах дендритов, реже на тонких разветвлениях дендритов.
Характерным для межнейронных связей является их полиморфизм, что послужило основанием для многочисленных классификаций С. Наиболее полная классификация С., основанная на светооптическом изучении межнейронных связей (по характеру разветвлений аксонов вокруг тел клеток и их взаиморасположения с ветвями дендритов), была предложена С. Рамон-и-Кахалем в 1954 г. Согласно этой классификации межнейронные связи формируются сетью разветвлений одного аксона или веточкой аксона с незначительным числом коллатералей, к-рые идут прямо к нейрону и образуют с ним С., сетью контактирующих между собой аксонов нескольких нейронов вокруг тела нейрона или какого-нибудь участка дендрита.
Электронограммы аксодендритического (а) и аксо-аксонального синапсов (б) в коре большого мезга крысы: 1 — цитоплазматическая мембрана пресинаптического окончания; 2 — синаптические пузырьки: 3 — митохондрии; 4 — цитоплазматическая мембрана дендрита; 5— активная зона синапсов; 6— цитоплазма дендрита; 7 —пресинаптические окончания аксонов; х 30 000.По локализации С. на нейронах межнейронные связи делят на 3 основные группы: аксодендритические (рис., а), аксосоматические и аксо-аксональные (рис., б). В свою очередь, аксодендритические С. разделяют еще на 3 подгруппы: С. на стволах дендритов, на шипиках дендритов и на тонких концевых веточках дендритов; реже встречаются денд-ро-дендритические, дендросомати-ческие и соматодендритические синапсы.
Описаны более сложные, комплексные формы межнейронных контактов. К ним относятся реципрок-ные С., расположенные между аксоном и дендритом (аксодендритические и дендроаксональные С., располагающиеся рядом), и сериальные С., т. е. идущие один за другим аксо-аксодендритические или аксо-аксосоматические контакты. Своеобразными формами межнейронных контактов являются гломерулы, в к-рых комплекс межнейронных связей как бы отделен от окружающей ткани мозга отростками глиальных клеток, а также синаптические поля, характеризующиеся сложной взаимосвязанностью окончаний аксонов различных систем волокон.
В ц. н. с. аксоны образуют межнейронные связи либо своими концевыми веточками, либо по ходу своего продвижения. С., образованные по ходу продвижения, служат, по-видимому, для передачи малоспецифической информации.
Существенную роль в организации межнейронного контакта играет площадь активной зоны контактирующих мембран С., хотя нет жесткой зависимости между площадью активной зоны контакта и площадью соприкосновения пресинаптического и постсинаптического участков. На основе исследования ультраструктуры синапсов H. Н. Боголеповым был предложен термин «информативность» межнейронного контакта, к-рая определяется степенью воздействия аксона на дендрит в области С. и (с морфологической точки зрения) зависит от площади активных зон С. в области контакта между пре-синаптическим отростком и постси-наптическим нейроном. С увеличением площади активной зоны С. увеличивается его информативность.
С.— это одни из наиболее пластичных и ранимых компонентов нейронов. Однако не все С. обладают равными пластическими возможностями; по степени пластичности они могут быть разделены на стабильные и динамичные. Стабильные С. в процессе онтогенеза созревают раньше, чем динамичные.
Исследование онтогенетических особенностей созревания межнейронных контактов показывает, что в ультраструктуре каждого из них отражается определенный путь развития. Он начинается с незрелого десмосовидного контакта, имеющего небольшое по площади увеличение электронной плотности контактирующих мембран и всего 2—3 синаптических пузырька в пресинапти-ческом отростке, и постепенно превращается в С., типичный для взрослого организма. В процессе онтогенеза также наблюдается неравномерность развития межнейронных связей различного типа. Наир., в зрительной области коры большого мозга С. образуются прежде всего на крупных стволах дендритов, затем на их основных разветвлениях и на телах нервных клеток. Позднее формируются С. на шипиках и тонких ветвях дендритов, образованных концевыми веточками аксонов, еще позднее происходит формирование аксо-аксональных С.
Неравномерность созревания С. создает основу для пластических изменений межнейронных связей у того или иного индивида в результате особенностей индивидуального развития, обучения, тренировки и ДР-
Особенности структурно-функциональной организации С. находят свое отражение и в закономерностях их деструктивных изменений при повреждении. При перерезке аксонов различных систем нейронов возникают разные формы деструктивных изменений С. Основными среди них являются так наз. темная, светлая и филаментарная дегенерации. Кроме того, описана очаговая дегенерация С. Наиболее распространена темная дегенерация, характеризующаяся повышением осмиофилии цитоплазмы пресинаптического отростка. При этом дегенерирующие волокна и их окончания сжимаются, приобретают неправильную, уродливую форму. Повышение электронной плотности пресинаптического отростка происходит вначале за счет накопления в нем мелкогранулярного материала. Затем цитоплазма гомогенизируется и становится столь осмиофильной, что на ее фоне лишь с трудом можно различить какие-либо дискретные структуры.
С темной дегенерацией по своему развитию сходна филаментарная дегенерация. Она характеризуется появлением в пресинаптическом отростке множества филаментов диаметром 6—10 нм, нередко образующих кольцеобразную структуру. При электронно-микроскопическом исследовании филаментарная реакция выявляется на 3—7-й день после перерезки соответствующего нервного волокна. В дальнейшем увеличение количества филаментов дополняется появлением мелкогранулярного материала в пресинаптическом отростке и последующее течение процесса мало отличается от темной дегенерации С.
Для светлой дегенерации С. характерны отек пресинаптического окончания, увеличение его размеров, просветление цитоплазмы и уменьшение числа синаптических пузырьков. Светлая дегенерация возникает в те же сроки, что и темная, в нек-рых случаях несколько раньше. Светлая дегенерация является также типичной формой патологии С. при гипоксии (см.).
При очаговой дегенерации С. наряду с просветлением пресинаптического отростка наблюдается частичный распад синаптических мембран и грубое нарушение контакта в этом участке.
Физиология
Успехи в исследовании строения клеточных мембран дали возхможность глубже изучить не только структуру, но и функцию С. Предполагают, что появление межклеточных контактов в филогенезе было тем ключевым моментом, к-рый привел к возможности взаимодействия клеток между собой, т. е. к образованию из одноклеточных организмов многоклеточных, появлению тканевых систем, в т. ч. нервной системы. С. как контакт нервной клетки с другими возбудимыми образованиями представляет собой специализированный вид межклеточного контакта. Поскольку именно С. представляют собой элементы, объединяющие отдельные нейроны в нейронные системы и обеспечивающие возможность регулирующего влияния ц. н. с. на другие возбудимые образования (мышцы, железы и др.), становится понятной их большая роль в деятельности всей нервной системы в целом.
По способу передачи возбуждения С. подразделяют на три группы. К первой группе относятся С., в к-рых передача возбуждения осуществляется с помощью хим. веществ-передатчиков (см. Медиаторы), ко второй — синапсы с передачей возбуждения без участия медиаторов, только за счет непосредственного перехода электрического сигнала с пре- на постсинаптические структуры (см. Эфапс), к третьей — «смешанные синапсы», в к-рых передача осуществляется и химическим, и электрическим способами.
По изменению потенциала постси-наптической мембраны различают тормозные и возбуждающие С. Кроме того, все С. подразделяют на центральные (находящиеся в головном и спинном мозге) и периферические (нервно-мышечные, нейросекреторные, а также С. вегетативных ганглиев).
Аксо-аксональные С. найдены в самых различных частях центральной и периферической нервной системы. Большинство исследователей считают, что именно с помощью аксо-аксональных С. осуществляется пресинаптическое торможение (см.).
В С. с химической передачей медиатор, выделяясь из пресинаптического окончания, попадает в синаптическую щель, к-рая является непосредственным продолжением межклеточного пространства. Содержимое синаптической щели обладает свойствами полисахаридного геля, в его состав входят гликозаминогликаны. В пресинаптической области локализованы митохондрии, гранулы гликогена, гранулы с мелкозернистым содержимым, различного рода филаментозные структуры. Предполагают, что спиралевидные филаментозные нити при отсутствии возбуждения пресинаптического окончания расположены параллельно субсинаптической мембране и служат своеобразным барьером для взаимодействия синаптических пузырьков с активными зонами пресинаптической мембраны. В том случае, когда нервный импульс достигает пресинаптического окончания, филаментозные нити распадаются на отдельные субъединицы, способствуя тем самым взаимодействию содержимого синаптического пузырька с активными зонами синапса. Т. о., выделение медиатора носит прерывистый характер в виде отдельных порций, или квантов. Большинство исследований, посвященных изучению квантового характера выброса медиаторов, было выполнено на синаптических пузырьках, содержащих ацетилхолин. Так, для С. млекопитающих число молекул медиатора, содержащихся в одном синаптическом пузырьке и одновременно изливающихся в синаптическую щель (т. е. квант ацетилхолина), колеблется в широких пределах и составляет от 4 * 102 до 4-104 молекул.
Между синаптическими пузырьками и выделением квантов медиатора существует функциональная связь. Так, при длительнохм раздражении в пресинаптическом окончании значительно уменьшается количество синаптических пузырьков, что сопровождается снижением эффективности передачи через С. распространяющегося возбуждения.
В процессе синаптической передачи медиатор из пресинаптического окончания попадает в синаптическую щель двумя путями: при контакте синаптического пузырька с внутренней поверхностью пресинаптической мембраны или при выделении свободного невезикулярного передатчика. В состоянии покоя (т. е. при отсутствии деполяризации пресинаптического окончания) происходит случайное столкновение синаптических пузырьков с пресинаптической мембраной. В результате экзоцитоза в синаптическую щель попадает небольшое количество медиатора, к-рый, взаимодействуя с клеточными рецепторами постсинаптической мембраны, приводит к возникновению миниатюрного постсинапти-ческого потенциала (МПП). МПП возникают апериодически, беспорядочно, их величина значительно меньше порога возбуждения постсинаптической мембраны. При деполяризации пресинаптической мембраны количество квантов медиатора, выделяющегося в синаптическую щель, увеличивается в несколько раз, в результате чего происходит возбуждение постсинаптической мембраны и появление распространяющегося возбуждения.
Эффект деполяризации пресинап-тического окончания и соответственно увеличение выделения квантов медиатора сказывается на состоянии возбудимости постсинаптической мембраны не сразу. Отмечается заметная задержка во времени от момента деполяризации пресинаптической мембраны до возникновения импульса в постсинаптической мембране — так наз. синаптическая задержка (СЗ). Она обусловлена рядом факторов: временем освобождения медиатора из синаптического пузырька при экзоцитозе, диффузией медиатора через синаптическую щель, взаимодействием медиатора с клеточными рецепторами постсинаптической мембраны и формированием распространяющегося процесса возбуждения в постсинаптических структурах. В частности, у мотонейронов спинного мозга продолжительность СЗ составляет 0,5 мсек, а в нек-рых случаях может достигать 2— 2,5 мсек. Продолжительность СЗ обусловлена многими факторами: температурой, содержанием различных ионов, pH среды и др.
Взаимосвязь между деполяризацией пресинаптической мехмбраны и высвобождением медиатора обеспечивают ионы кальция (см. Мембраны биологические). Именно кальций выполняет роль фактора, сопрягающего процесс изменения величины мембранного потенциала пресинаптической мембраны с усилением выделения медиатора в синаптическую щель. В связи с тем, что входящий ток в пресинаптической мембране связан с увеличением проницаемости натриевых каналов, а процесс секреции медиатора — с увеличением проницаемости кальциевых каналов, появилась возможность разобщить процессы деполяризации пресинаптической мембраны и секреции медиатора. Так, при действии ингибиторов синаптической передачи, па пр. тетродотоксина — блокатора проводимости натриевых каналов мембраны, может отсутствовать изменение потенциала пресинаптической мембраны, но продолжается секреция медиатора. Блокаду секреции медиатора можно вызвать с помощью факторов, связывающих ионы кальция (напр., этилендиаминтетрауксусной к-той — ЭДТА), или же путем удаления ионов кальция из внешней среды (см. Проницаемость). Секрецию медиатора можно усилить действием специфических кальциевых ионофоров (см.). Механизм секреции медиатора регулируется рядом биологически активных веществ, в т. ч. и самими медиаторами, а также циклическими нуклеотидами (см. Нуклеиновые кислоты), простагландинами (см.) и рядом нейропептидов.
В процессах изменения состояния постсинаптической мембраны большое значение имеет система адени-латциклаза — циклический АМФ. Предполагают, что взаимодействие медиатора с рецепторным участком аденилатциклазы приводит к активации ее каталитического центра, к образованию циклического АМФ с последующим повышением активности протеинкиназ цитоплазмы и ядра клетки. Это в свою очередь интенсифицирует фосфорилирование белков, непосредственно участвующих в изменении проницаемости постсинаптической мембраны, и ведет к появлению в постсинаптических структурах распространяющегося процесса возбуждения. При спонтанном выделении квантов медиатора (напр., в нервно-мышечном С.) в постсинаптической мембране регистрируется л8кальное изменение уровня мембранного потенциала, названного миниатюрным потенциалом концевой пластинки (МПКП).
В том случае, когда выделение квантов медиатора недостаточно для возникновения потенциала действия, в постсинаптической мембране регистрируется локальная деполяризация или гиперполяризация. В нейронных С. эта реакция названа возбуждающим постсинаптическим потенциалом (ВПСП) и тормозным постсинаптическим потенциалом (ТПСП), а в нервно-мышечных С.— потенциалом концевой пластинки (ПКП).
Возникновение ВПСП связано с функционированием в синаптических мембранах специальных каналов для таких катионов, как Na+ и К+. Анионы в подавляющем числе случаев не принимают участия в генерации ВПСП. Возникновение ВПСП в мотонейронах происходит в результате одновременного повышения натриевой и калиевой проводимости постсинаптической мембраны, однако возможно и избирательное увеличение ее проницаемости к ионам натрия. Обнаружена зависимость между величиной мембранного потенциала и значением ВПСП. Эта зависимость свидетельствует о том, что движение ионов по каналам постсинаптической мембраны осуществляется путем пассивной диффузии, а не с помощью активного транспорта (см. Биоэлектрические потенциалы).
Передача возбуждения в электрических С. осуществляется только с помощью электрических токов. Необходимо отметить, что электрические С. встречаются значительно реже, чем С. с химической передачей. Электрические синапсы (см. Эфапс) по сравнению с химическими отличаются большей быстротой передачи сигнала, высокой надежностью передачи, возможностью двусторонней передачи возбуждения. В то же время химические С., использующие сложный полифункциональ-ный нейрохимический аппарат, способны в неизмеримо большей степени обеспечить сохранение информационной значимости сигналов, кодируемой биологически активными веществами и метаболическими процессами, протекающими в постси-наптнческих структурах.
Т. о., основные этапы синаптической передачи схематично могут быть представлены следующими процессами: приход возбуждения в пре-синагхтическую мембрану и ее деполяризация, проникновение внутрь пресинаптической мембраны ионов кальция, взаимодействие синаптических пузырьков с активными участками пресинаптической мембраны, экзоцитоз и выделение квантов медиатора в синаптическую щель, диффузия медиатора к постсинапти-чевкой мембране, взаимодействие медиатора с клеточными рецепторами постсинаптической мембраны и как следствие этого изменение проницаемости последней к определенным ионам, образование постсинаптических потенциалов и возникновение распространяющегося процесса возбуждения. Возникновение ТПСП полностью предотвращает возможность появленрш возбуждения в постсинаптической мембране и связано с изменением проницаемости мембраны к ионам хлора или калия.
Основными методами исследования синаптических процессов являются микроэлектродная техника (см. Микроэлектродный метод исследования), гистоавторадиографические методы (см. Авторадиография), электронная микроскопия (см.), биохимические методы исследования (см.) и нейрофармакологический анализ.
Библиография: Бак 3. М. Химическая передача нервного импульса, пер. с франц., М., 1977; Боголепов H. Н. Ультраструктура синапсов в норме и патологии, М., 1975; он же, Ультраструктура мозга при гипоксии, М., 1978; Глебов Р. Н. и Крыжанов-с к и й Г. Н. Функциональная биохимия синапсов, М., 1978, библиогр.; К и-б я к о в А. В. Химическая передача нервного возбуждения, М.— JI., 1964; Саркисов С. А. Некоторые особенности строения нейрональных связей коры большого мозга, М., 1948.
С. А. Осиповский; H. Н. Боголепов (морфология).
xn--90aw5c.xn--c1avg
Организация и функция синапса
В синапсе различают пресинаптическую часть, постсинаптическую часть и расположенную между клетками синаптическую щель (см. рис 6–1Б и 6–6, а также рис. 6–5 в книге).
Пресинаптическая часть
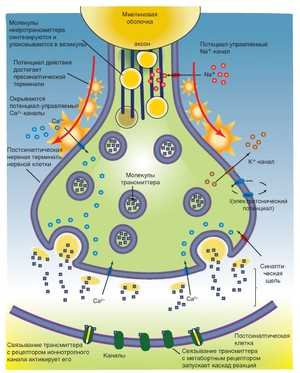
Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и митохондрии. В пресинаптическую мембрану встроены потенциалозависимые Ca2+‑каналы. При поступлении ПД к терминальному расширению мембрана деполяризуется, Ca2+‑каналы открываются, ионы Ca2+ входят в терминаль, запуская в активных зонах процесс слияния мембраны синаптического пузырька и пресинаптической мембраны, т.е. секрецию (экзоцитоз) нейромедиатора (рис. 6–6, позиции 2–4).
РольСа2+. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+и тем самым принимает участие в регуляции экзоцитоза (в том числе путём реорганизации примембранного цитоскелета).
Синаптическиепузырьки. Молекулы нейромедиатора накапливаются в нервной терминали, находясь внутри синаптических пузырьков вместе с АТФ и некоторыми катионами. В каждом пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант нейромедиатора.
Синтезнейромедиатора. Ферменты, необходимые для образования нейромедиаторов, синтезируются в перикарионе и транспортируются к синаптической терминали по аксонам (рис. 6–4).
Типыпузырьков— мелкие (диаметр порядка 50 нм) и крупные (диаметр 100–200 нм). Мелкие синаптические пузырьки содержат «классические» медиаторы (см. ниже). Крупные везикулы содержат нейропептиды.
Секреция. Когда ПД достигает нервной терминали, синаптические пузырьки сливаются с пресинаптической мембраной, что приводит к выделению квантов нейромедиатора в синаптическую щель. Незначительное количество квантов нейромедиатора постоянно (спонтанно) секретируется в синаптическую щель.
Узнавание. Предшествующий слиянию синаптических пузырьков и плазмолеммы процесс узнавания синаптическим пузырьком пресинаптической мембраны происходит при взаимодействии мембранных белков (синаптобревин, SNAP-25, синтаксин и другие).
Влияниетоксинов. Синтаксин, SNAP-25 и синаптобревин — мишени ботулинического токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. Мишень столбнячного токсина — синаптобревин.
Активныезоны(рис. 6–3). Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания — активных зонах — участках утолщения пресинаптической мембраны. Активная зона состоит из «плотной полоски» на пресинаптической мембране и сгруппированных около неё синаптических пузырьков, потенциалозависимых кальциевых каналов, специальных белков экзоцитоза и элементов цитоскелета. Количество активных зон в нервно-мышечном синапсе достигает 30–40, в межнейронных синапсах — около десятка. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.

Рис.6–3.Активныезонынервно-мышечногосинапсарасположены напротив постсинаптических складок — участков скоплений холинорецепторов. Пресинаптическая мембрана слева расщеплена на два листка.
Жизненныйциклсинаптическихпузырьков(рис. 6–4). Синаптические везикулы образуются в теле нейрона в эндоплазматическом ретикулуме и комплексеГольджи(1) и с аксонным транспортом поступают в нервные окончания (2). В нервном окончаниимелкиесинаптическиепузырькипосредством активного транспорта заполняются медиатором (3) и передвигаются к пресинаптической мембране (4). Освобождение медиатора (5) может осуществляться посредством экзоцитоза с полным («классический» механизм) либо неполным (механизм «kiss and run») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы (6), которые затем проходят стадию эндосомы (7) и снова заполняются медиатором (3). Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё (8) и повторно заполняется медиатором (3).Крупныесинаптическиевезикулызаполняются медиатором в теле клетки (9), их экзоцитоз происходит в других участках пресинаптической мембраны, а эндоцитоз опорожнённых пузырьков отсутствует (10).

Рис.6–4.Образование,транспортиэкзоцитозсинаптическихпузырьков.
Синаптическая щель— промежуток между пре- и постсинаптическими мембранами шириной 20–35 нм. В синаптическую щель из синаптических пузырьков выделяются молекулы нейромедиатора и путём диффузии достигают постсинаптической мембраны. В синаптической щели находятся ферменты, расщепляющие молекулы нейромедиатора (например, ацетилхолинэстераза, гидролизующая ацетилхолин), а в пресинаптическую мембрану вмонтированы переносчики, осуществляющие перенос нейромедиаторов–аминокислот и биогенных аминов (например, глутамата, аспартата, норадреналина) в пресинаптическую терминаль.
Таким образом, удаление нейромедиатора из синаптической щели происходит двояко: инактивация ферментом или захват пресинаптической терминалью.
Инактивациянейромедиатора. Кратковременность взаимодействия нейромедиатора с рецептором достигается разрушением нейромедиатора специальными ферментами (например, ацетилхолина — ацетилхолинэстеразой).
Захватнейромедиатора. В большинстве синапсов передача сигналов прекращается вследствие быстрого захвата нейромедиатора пресинаптической терминалью.
Транспортёры. Захватнорадреналина осуществляют специфические Na+— и Cl–-транспортирующие белки (например, норадреналин–транспортирующий белок 1) — мишени трициклических антидепрессантов (например, дезипрамин и имипрамин). Система захвата биогенных аминов — точка приложения антидепрессантов и таких препаратов, как кокаин и амфетамины. Дефекты транспортёров норадреналина исеротонина— кандидаты на роль первопричины при психиатрических расстройствах, таких как маниакально-депрессивные состояния.
studfiles.net