
Рецепторы медиаторов – 20.10 Определение медиаторов и синоптических рецепторов, их классификация и роль в проведении сигналов в возбуждающих и тормозных синапсах.
Медиаторы и рецепторы цнс
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
А. Ацетилхолин. Встречается в различных отделах ЦНС, известен в основном как возбуждающий медиатор: в частности, является медиатором α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина α -мотонейроны по коллатералям своих аксонов передают возбуждение на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и N-холинорецепторы. При взаимодействии ацетилхолина с рецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. Тормозное влияние ацетилхолин оказывает с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Б. Моноамины. Выделяют катехоламины, серотонин и гистамин. Большинство из них в значительных количествах содержится в нейронах ствола мозга, в меньших количествах они обнаруживаются в других отделах ЦНС.
Катехоламины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга — тормозящие влияния. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в лимбической системе. В нейронах названных структур выявлены ферменты, участвующие в синтезе серотонина. Полагают, что часть этих так называемых симпатических нейронов, а может быть и все, являются серотонинергическими нейронами вегетативной нервной системы.
Гистамин в довольно высокой концентрации обнаружен в гипофизе и гипоталамусе. В остальных отделах ЦНС уровень гистамина очень низкий. Выделяют Н1- и Н2-гистаминорецепторы. Н1-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, терморегуляции, секреции пролактина и антидиуретического гормона. Н2-рецепторы обнаружены на глиальных клетках.
В. Аминокислоты. Кислые аминокислоты (глицин, γ-аминомасляная кислота) являются тормозными медиаторами в синапсах ЦНС и действуют на тормозные рецепторы.
Нейтральные аминокислоты (α -глутамат, α -аспартат) передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Считается, что глутамат
Г. Полипептиды. В синапсах ЦНС они также выполняют медиаторную функцию. В частности, субстанция Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида в дорсальных корешках (радикс) спинного мозга. Субстанция Р в больших количествах содержится в гипоталамической области
Энкефалины и эндорфины — медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного и спинного мозга.
Ангиотензин участвует в передаче информации о потребности организма в воде, люлиберин — в половой активности и т.д.
Нейрофизиологические эффекты действия некоторых медиаторов головного мозга. 1.Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений;
2.дофамин -в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения.
4.ангиотензин — повышение АД, торможение синтеза катехоламинов, стимулирует секрецию гормонов; информирует ЦНС об осмотическом давлении крови.
5.Олигопептиды — медиаторы настроения, полового поведения; передачи ноцицептивного возбуждения от периферии в ЦНС, формирования болевых ощущений.
6.Эндорфины, энкефалины, пептид, вызывающий дельта-сон, обусловливают антиболевые реакции, повышение устойчивости к стрессу, сон.
7.Простагланди
Мозгоспецифичные белки различных отделов головного мозга влияют на процессы обучения.
Согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор или одни и те же медиаторы во всех разветвлениях своего аксона. Кроме основного медиатора, как недавно выяснилось, в окончаниях аксона могут выделяться и другие — сопутствующие медиаторы, играющие модулирующую роль.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — торможение, в синапсах гладкой мускулатуры пищеварительного тракта — возбуждение. Катехоламины вызывают возбуждение и торможение в стволе мозга.
studfiles.net
02. МЕДИАТОРЫ
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге к настоящему времени обнаружено около 30 биологически активных веществ). Вещество, из которого синтезируется медиатор (предшественник медиатора), попадает в нейрон или его окончание из крови или ликвора, в результате биохимических реакций под действием ферментов превращается в соответствующий медиатор, затем транспортируется в синапти-ческие везикулы. По химическому строению их можно разделить на несколько групп, главными из которых являются амины, аминокислоты, полипептиды. Достаточно широ-
ко распространенным медиатором является ацетилхолин.
А. Ацетилхолин встречается в коре большого мозга, в стволе, в спинном мозге, известен в основном как возбуждающий медиатор; в частности, является медиатором а-мо-тонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетил-холина а-мотонейроны по коллатералям своих аксонов передают возбуждающее влияние на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и Н-холинорецепторы. В ЦНС установлено 7 типов Н-холинорецеп-торов. В ЦНС основными М-холинорецепто-рами являются Мг и М2-рецепторы. М,-хо-линорецепторы локализуются на нейронах гиппокампа, полосатого тела, коры большого мозга.
Б. Амины (дофамин, норадреналин, серото-нин, гистамин). Большинство из них в значительных количествах содержится в нейронах ствола мозга, в меньших количествах выявляется в других отделах ЦНС.
Амины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, в черной субстанции, в лимбической системе, в полосатом теле. Норадренергические нейроны сконцентрированы в основном в области голубого пятна (средний мозг), где их насчитывается всего несколько сотен. Но ответвления их аксонов встречаются по всей ЦНС.
Норадреналин является тормозным медиатором клеток Пуркинье мозжечка и периферических ганглиев; возбуждающим — в гипоталамусе, в ядрах эпиталамуса. В ретикулярной формации ствола мозга и гипоталамусе обнаружены а- и р-адренорецепторы.
Дофаминорецепторы подразделяют на Дг и Д2-подтипы. Д,-рецепторы локализуются на клетках полосатого тела, действуют посредством дофаминчувствительной аденилатцикла-зы, как и Д2-рецепторы. Д2-рецепторы обнаружены в гипофизе. При действии на них дофамина угнетаются синтез и секреция про-лактина, окситоцина, меланоцитстимулиру-ющего гормона, эндорфина. Д2-рецепторы найдены на нейронах полосатого тела, где их функция пока не определена.
Серотонин. С его помощью в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга — тормозящие влияния. Имеется несколько типов серотонинорецепторов. Серотонин реализует свое влияние с помощью ионотропных и ме-таботропных рецепторов (цАМФ и ИФз/ДАГ). Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в лимбической системе, ядрах шва. В нейронах названных структур выявлены ферменты, участвующие в синтезе серотони-на. Аксоны этих нейронов проходят в буль-боспинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преган-глионарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих так называемых симпатических нейронов (а может быть, и все) являются серотонинергическими нейронами вегетативной нервной системы. Их аксоны, согласно последним данным, идут к органам желудочно-кишечного тракта и стимулируют их сокращения.
Гнетами н. Довольно высокая его концентрация обнаружена в гипофизе и срединном возвышении гипоталамуса — именно здесь сконцентрировано основное количество гистаминергических нейронов. В остальных отделах ЦНС уровень гистамина очень низок. Медиаторная роль его изучена мало. Выделяют Н,-, Н2— и Н3-гистаминорецепто-ры. Н,-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, терморегуляции, секреции пролактина и антидиуретического гормона. Н2-рецепторы обнаружены на глиальных клетках. Свое влияние гистамин реализует с помощью вторых посредников (цАМФ и ИФ3/ДАГ).
В. Аминокислоты. Кислые аминокислоты (глицин, гамма-аминомасляная кислота) являются тормозными медиаторами в синапсах ЦНС и действуют на соответствующие рецепторы (см. раздел 7.8), глицин — в спинном мозге, в стволе мозга, ГАМК — в коре большого мозга, мозжечке, стволе мозга, спинном мозге. Нейтральные аминокислоты (альфа-глутамат, альфа-аспартат) передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Предполагается, что глутамат может быть медиатором аффе-рентов в спинном мозге. Рецепторы глутами-новой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, та-ламуса, гиппокампа, коры большого мозга. Глутамат — основной возбуждающий медиатор ЦНС (75 % возбуждающих синапсов мозга). Рецепторы глутамата ионотропные (К+, Са2+, Na+) и метаботропные (цАМФ и ИФз/ДАГ).
Г. Полипептиды также выполняют медиа-торную функцию в синапсах ЦНС. В частности, вещество П является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга. Это послужило основанием к предположению, что вещество П может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Вещество П в больших количествах содержится в гипо-таламической области. Различают два вида рецепторов вещества П: рецепторы типа SP-Р, расположенные на нейронах мозговой перегородки, и рецепторы типа SP-E, расположенные на нейронах коры большого мозга.
Энкефалины и эндорфины — медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы, много их также на клетках черной субстанции, ядрах промежуточного мозга и солитар-ного тракта, имеются они на клетках голубого пятна, спинного мозга. Их лигандами являются р-эндорфин, динорфин, лей- и ме-тэнкефалины. Различные опиатные рецепторы обозначаются буквами греческого алфавита: ц, к, су, 1, е. К-рецепторы взаимодействуют с динорфином и лей-энкефалином, избирательность действия других лигандов на опиатные рецепторы не доказана.
Ангиотензин участвует в передаче информации о потребности организма в воде, люлиберин — в половой актив-
ности. Связывание ангиотензина с рецепторами вызывает увеличение проницаемости клеточных мембран для Са2+. Эта реакция обусловлена не конформационными изменениями рецепторного белка, а процессами фосфорилирования мембранных белков вследствие активации аденилатциклазной системы и изменением синтеза простаглан-динов. Рецепторы к ангиотензину обнаружены на нейронах головного мозга, на клетках среднего и промежуточного мозга, коры большого мозга.
На нейронах головного мозга выявлены ВИП-рецепторы и рецепторы к соматостати-ну. Рецепторы к холецистокинину обнаружены на клетках коры большого мозга, хвостатого ядра, обонятельных луковиц. Действие холе-цистокинина на рецепторы повышает проницаемость мембран для Са2+ посредством активации аденилатциклазной системы.
Д. АТФ также может выполнять роль классического медиатора, в частности в нейронах уздечки (возбуждающий эффект). В спинном мозге выделяется вместе с ГАМ К, но выполняет возбуждающую функцию. Весьма разнообразны рецепторы к АТФ, одни из них ионотропные, другие — метабо-тропные. АТФ и аденозин участвуют в формировании болевых ощущений, ограничивают перевозбуждение ЦНС.
Е. Химические вещества, циркулирующие в крови (некоторые гормоны, простагланди-ны), оказывающие модулирующее влияние на активность синапсов. Простагландины — ненасыщенные оксикарбоновые кислоты, высвобождаемые из клеток, влияют на многие звенья синаптического процесса, например на секрецию медиатора, работу адени-латциклаз. Они обладают высокой физиологической активностью, но быстро инактиви-руются и потому действуют локально.
Ж. Гипоталамические нейрогормоны. регулирующие функцию гипофиза, также выполняют медиаторную роль.
Физиологические эффекты действия некоторых медиаторов головного мозга. Н о р-адреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений; дофамин — в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. С е р о т о-н и н ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание; ангиотензин —
повышение АД, торможение синтеза катехол-аминов, стимулирует секрецию гормонов, информирует ЦНС об осмотическом давлении крови. Олигопептиды — медиаторы настроения, полового поведения; передачи ноцицептивного возбуждения от периферии к ЦНС, формирования болевых ощущений. Эндорфины, энкефалины, пептид, вызывающий дельт а-с о н, дают антиболевые реакции, повышение устойчивости к стрессу, сон. Простагландины вызывают повышение свертываемости крови; изменение тонуса гладких мышц, усиление физиологического эффекта медиаторов и гормонов. Мозгоспецифичные белки различных отделов головного мозга влияют на процессы обучения.
Согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор во всех разветвлениях своего аксона («один нейрон — один медиатор»). Кроме основного медиатора, как выяснилось, в окончаниях аксона могут выделяться и другие — сопутствующие медиаторы (комедиаторы), играющие модулирующую роль или более медленно действующие. Однако в спинном мозге установлено два быстродействующих типичных медиатора в одном тормозном нейроне — ГАМ К и глицин и даже один тормоз-ный (ГАМК.) и один возбуждающий (АТФ). Поэтому принцип Дейла в новой редакции сначала звучал так: «Один нейрон — один быстрый медиатор», а затем: «Один нейрон — один быстрый синаптический эффект».
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — торможение, в синапсах гладкой мускулатуры желудочно-кишечного тракта — возбуждение. Катехоламины стимулируют сердечную деятельность, но тормозят сокращения желудка и кишечника.
studfiles.net
Медиаторы и рецепторы ЦНС
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
А. Ацетилхолин. Встречается в различных отделах ЦНС, известен в основном как возбуждающий медиатор: в частности, является медиатором α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина α -мотонейроны по коллатералям своих аксонов передают возбуждение на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и N-холинорецепторы. При взаимодействии ацетилхолина с рецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. Тормозное влияние ацетилхолин оказывает с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Б. Моноамины. Выделяют катехоламины, серотонин и гистамин. Большинство из них в значительных количествах содержится в нейронах ствола мозга, в меньших количествах они обнаруживаются в других отделах ЦНС.
Катехоламины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга — тормозящие влияния. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в лимбической системе, ядрах шва. В нейронах названных структур выявлены ферменты, участвующие в синтезе серотонина. Аксоны этих нейронов проходят в бульбо-спинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преганглионарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих так называемых симпатических нейронов, а может быть и все, являются серотонинергическими нейронами вегетативной нервной системы. Их аксоны, согласно данным некоторых авторов, идут к органам пищеварительного тракта и стимулируют их сокращение.
Гистамин в довольно высокой концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса. В остальных отделах ЦНС уровень гистамина очень низкий. Медиаторная роль его изучена мало. Выделяют Н1- и Н2-гистаминорецепторы. Н1-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, терморегуляции, секреции пролактина и антидиуретического гормона. Н2-рецепторы обнаружены на глиальных клетках.
В. Аминокислоты. Кислые аминокислоты (глицин, γ-аминомасляная кислота) являются тормозными медиаторами в синапсах ЦНС и действуют на тормозные рецепторы (см. раздел 4.8). Нейтральные аминокислоты (α -глутамат, α -аспартат) передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Предполагают, что глутамат может быть медиатором афферентов в спинном мозге. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Полагают, что глутамат — самый распространенный медиатор ЦНС.
Г. Полипептиды. В синапсах ЦНС они также выполняют медиаторную функцию. В частности, субстанция Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида в дорсальных корешках спинного мозга. Это послужило основанием к предположению, что субстанция Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Субстанция Р в больших количествах содержится в гипоталамической области. Различают два вида рецепторов субстанции Р: рецепторы типа SР-Р, расположенные на нейронах мозговой перегородки, и рецепторы типа SР-Е, расположенные на нейронах коры большого мозга.
Энкефалины и эндорфины — медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного мозга и солитарного тракта, имеются они на клетках голубого пятна, спинного мозга. Их лигандами являются [3-эндорфин, динорфин, лей- и мет-энкефалины.
Ангиотензин участвует в передаче информации о потребности организма в воде, люлиберин — в половой активности.
Физиологические эффекты действия некоторых медиаторов головного мозга. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений; дофамин -в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. Серотонин ускоряет процессы обучения, формирования болевых ощущений, сенсорное восприятие, засыпание, ангиотензин — повышение АД, торможение синтеза катехоламинов, стимулирует секрецию гормонов; информирует ЦНС об осмотическом давлении крови. Олигопептиды — медиаторы настроения, полового поведения; передачи ноцицептивного возбуждения от периферии в ЦНС, формирования болевых ощущений. Эндорфины, энкефалины, пептид, вызывающий дельта-сон, обусловливают антиболевые реакции, повышение устойчивости к стрессу, сон. Простагландины вызывают повышение свертываемости крови, изменение тонуса гладких мышц, усиление физиологического эффекта медиаторов и гормонов. Мозгоспецифичные белки различных отделов головного мозга влияют на процессы обучения.
Согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор или одни и те же медиаторы во всех разветвлениях своего аксона. Кроме основного медиатора, как выяснилось, в окончаниях аксона могут выделяться и другие — сопутствующие медиаторы, играющие модулирующую роль.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — торможение, в синапсах гладкой мускулатуры пищеварительного тракта — возбуждение. Катехоламины вызывают возбуждение и торможение в стволе мозга.
www.braintools.ru
МЕДИАТОРЫ И РЕЦЕПТОРЫ ЦНС — Мегаобучалка
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды. Достаточно широко распространенным медиатором является ацетилхолин.
А. Ацетилхолин. Встречается в различных отделах ЦНС, известен в основном как возбуждающий медиатор: в частности, является медиатором а-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина а-мотонейроны по коллатералям своих аксонов передают возбуждение на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и К-холинорецепторы. При взаимодействии ацетилхолина с рецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. Тормозное влияние ацетилхолин оказывает с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Б. Моноамины. Выделяют катехоламины, серотонин и гистамин. Большинство из них в значительных количествах содержится в нейронах ствола мозга, в меньших количествах они обнаруживаются в других отделах ЦНС.
Катехоламины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга — тормозящие влияния. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в лимбической системе, ядрах шва. В нейронах названных структур выявлены ферменты, участвующие в синтезе серотонина. Аксоны этих нейронов проходят в бульбоспинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преганглионарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих так называемых симпатических нейронов, а может быть и все, являются серотонинергическими нейронами вегетативной нервной системы. Их аксоны, согласно данным некоторых авторов, идут к органам пищеварительного тракта и стимулируют их сокращение.
Гистамин в довольно высокой концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса. В остальных отделах ЦНС уровень гистамина очень низкий. Медиаторная роль его изучена мало. Выделяют Н1— и Н2-гистаминорецепторы. Н1-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, терморегуляции, секреции пролактина и антидиуретического гормона. Н2-рецепторы обнаружены на глиаль-ных клетках.
В. Аминокислоты. Кислые аминокислоты (глицин, у-амино-масляная кислота) являются тормозными медиаторами в синапсах ЦНС и действуют на тормозные рецепторы (см. раздел 4.8). Нейтральные аминокислоты (а-глутамат, а-аспартат) передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Предполагают, что глутамат может быть медиатором афферентов в спинном мозге. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса гиппокампа, коры большого мозга. Полагают, что глутамат — самый распространенный медиатор ЦНС.
Г. Полипептиды. В синапсах ЦНС они также выполняют ме-диаторную функцию. В частности, субстанция Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида в дорсальных корешках спинного мозга. Это послужило основанием к предположению, что субстанция Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Субстанция Р в больших количествах содержится в гипоталамической области. Различают два вида рецепторов субстанции Р: рецепторы типа SР-Р, расположенные на нейронах мозговой перегородки, и рецепторы типа SР-E, расположенные на нейронах коры большого мозга.
Энкефалины и эндорфины — медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного мозга и солитарного тракта, имеются они на клетках голубого пятна, спинного мозга. Их лигандами являются (3-эндорфин, динорфин, лей- и мет-энкефалины.
Ангиотснзин участвует в передаче информации о потребности организма в воде, люлиберин — в половой активности.
Физиологические эффекты действия некоторых медиаторов головного мозга. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений; дофамин — в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. Серотонин ускоряет процессы обучения, формирования болевых ощущений, сенсорное восприятие, засыпание, ангиотензин — повышение АД, торможение синтеза катехоламинов, стимулирует секрецию гормонов; информирует ЦНС об осмотическом давлении крови. Олигопептиды -медиаторы настроения, полового поведения; передачи ноцицептив-ного возбуждения от периферии в ЦНС, формирования болевых ощущений. Эндорфины, энкефалины, пептид, вызывающий дельта-сон, обусловливают антиболевые реакции, повышение устойчивости к стрессу, сон. Простагландины вызывают повышение свертываемости крови, изменение тонуса гладких мышц, усиление физиологического эффекта медиаторов и гормонов. Мозгоспецифичные белки различных отделов головного мозга влияют на процессы обучения.
Согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор или одни и те лее медиаторы во всех разветвлениях своего аксона. Кроме основного медиатора, как выяснилось, в окончаниях аксона могут выделяться и другие — сопутствующие медиаторы, играющие модулирующую роль.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — торможение, в синапсах гладкой мускулатуры пищеварительного тракта — возбуждение. Катехоламины вызывают возбуждение и торможение в стволе мозга.
megaobuchalka.ru
Медиаторы
Основные медиаторные системы головного мозга
Тема 4 Основные медиаторные системы головного мозга ^ Медиатор (нейротрансмиттер, нейропередатчик) — вещество, с помощью которого нервный сигнал передается через синапс. Медиатор образуется либо в теле нейрона (и попадает в синаптическую бляшку, пройдя через весь аксон), либо непосредственно в синаптической бляшке. В синаптической бляшке молекулы медиатора упаковываются в синаптические пузырьки, в которых они хранятся до момента высвобождения. Известно несколько медиаторных веществ, для большинства из них описаны системы синтеза, хранения, высвобождения, взаимодействия с постсинаптическими рецепторами (из которых наиболее хорошо изучен ацетилхолиновый рецептор), инактивации, возврата продуктов их расщепления в пресинаптические окончания. Имеются данные о том, что во всех пресинаптических окончаниях зрелого нейрона высвобождается один и тот же медиатор (принцип Дейла). Однако в процессе своего развития некоторые нейроны временно синтезируют и высвобождают более одного медиаторного вещества. Предполагают, что каждый нейрон можно отнести к категории либо возбуждающих, либо тормозных («концепция функциональной специфичности»). Однако возбуждающий или тормозной характер действия медиатора определяется свойствами постсинаптической мембраны, а не самого медиатора. Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора и каждый из этих рецепторов способен контролировать разный механизм ионной проницаемости («принцип множественности медиаторного сигнала»). Таким образом, в нервной системе могло бы быть достаточно только одного медиатора, который при связывании с соответствующими постсинаптическими рецепторами вызывал бы тормозные или возбуждающие эффекты. Поэтому разнообразие медиаторных веществ заставляет предполагать, что они выполняют и другие функции, возможно, служат хемотаксическими факторами или трофическими факторами. В настоящее время известны следующие вещества, выполняющие медиаторные функции: ацетилхолин, катехоламины (адреналин, норадреналин, дофамин), аминокислоты (гамма-аминомасляная кислота, глутаминовая кислота, глицин), гистамин, нейроактивные пептиды. Сейчас известно более 30 веществ, выполняющих медиаторные функции. Нейроны, высвобождающие ацетилхолин, называются холинэргическими нейронами, а катехоламины — адренергическими нейронами. Некоторые аминокислоты обнаружены в ЦНС в довольно высоких концентрациях, что и вызвало предположение об их медиаторной функции. Считается, что они используются в системах крупных афферентных путей и эфферентных путей (возбуждающих и тормозных), в отличие от ацетилхолина и катехоламинов, служащих медиаторами преимущественно в периферической и вегетативной нервной системе. Гамма-аминомасляная кислота (ГАМК) синтезируется только в нервной системе из глутаминовой кислоты при посредстве глутаматдекарбоксилазы. Она встречается в ЦНС повсеместно, в самых разных концентрациях. При электофоретическом нанесении гамма-аминомасляная кислота оказывает, как правило, тормозное действие. Существуют данные о том, что эта кислота участвует в пресинаптическом торможении в качестве медиатора в аксо-аксонных синапсах. Некоторые судорожные яды (алкалоид бикукулин, пикротоксин) оказались специфическими антагонистами этой кислоты. Широко распространенная аминокислота глицин также, видимо, служит медиатором в некоторых случаях постсинаптического торможения в спинном мозге. Специфическим антагонистом глицина является стрихнин. Глутаминовая кислота при электрофоретическом нанесении обладает возбуждающим действием. Так как глутаминовая кислота обнаружена в ЦНС повсюду, весьма вероятно, что она не только является предшественником гамма-аминомасляной кислоты, но, кроме того, сама действует как медиатор. Гистамин образуется путем декарбоксилирования аминокислоты гистидина. Довольно высокие концентрации гистамина обнаружены в гипофизе и в соседнем срединном возвышении гипоталамуса. В остальных отделах ЦНС уровень гистамина очень низок. Молекулы нейроактивных пептидов представляют собой более или мене длинные цепи аминокислот. Предполагается, что вещество, которое служит медиатором в первичных афферентных волокнах в спинном мозге, является нейроактивным пептидом. Некоторые из нейроактивных пептидов представляют собой нейрогормоны, т.е. вещества, которые высвобождаются из нервных клеток, а затем переносятся кровотоком к их мишеням (не являющимся нейронами). К таким пептидам относятся либерины (рилизинг — гормоны), которые действуют на аденогипофиз, антидиуретический гормон (вазопрессин) и окситоцин, которые синтезируются в гипоталамусе и хранятся в нейрогипофизе. Еще одна группа пептидов влияет на активность нейронов не через посредство синапсов, а присутствуя в качестве гормонов, например, эндорфины. Такие пептиды называются нейромодуляторами. Выяснение этапов синаптической передачи пролило свет на способ действия психотропных препаратов. Некоторые из них действуют, либо усиливая, либо ослабляя высвобождение данного медиатора из аксонных окончаний. Например, под действием сильного стимулятора амфетамина в мозге из нервного окончания выделяется дофамин – медиатор, связанный с системами бодрствования и удовольствия. Чрезмерное применение амфетамина приводит к расстройствам мышления, галлюцинациям и мании преследования, т.е. к симптомам, сходными с теми, какие наблюдаются при некоторых формах шизофрении. Следовательно, существует предположение о том, что в основе симптомов шизофрении, возможно, лежит повышенная активность дофаминовых систем мозга. Многие психотропные препараты действуют на уровне постсинаптических рецепторов, имитируя естественные медиаторы. Например, многие галлюциногены сходны по своей структуре с истинными медиаторами: мескалин похож на норадреналин и дофамин (бензольное кольцо), а ЛСД и псилоцибин сходны с серотонином (индольное кольцо). В последние годы в литературе находится все больше подтверждений, что межмедиаторное взаимодействие в ЦНС частично реализуется несинаптическим путём. Этот новый нейротрансмиттерный механизм получил название объёмной передачи. Он основан на медленной диффузии нейромедиаторов по межклеточному пространству и их действии на отдалённые от места выброса — несинаптические рецепторы. Например, в органах, имеющих в своём составе гладкую мышечную ткань мезенхимного типа, терминали нервных волокон заканчиваются не непосредственно на миоцитах, а между ними. И после поступления нервного импульса, медиатор распространяется диффузно, возбуждая сразу многие клетки. Такой механизм обнаружен в стенках кровеносных сосудов, трубчатых внутренних органов, а также в отдельных мелких мышцах (цилиарных).
Синтез
медиаторов и их постсинаптическое
действие ^
ДОФАМИН (DA). Локализуется
преимущественно в среднем мозге (черная
субстанция, вентральная покрышка),
обонятельной луковице, гипоталамусе и
перивентрикулярной области продолговатого
мозга.
DA производится в дофаминэргических
нейронах из предшественника тирозина,
который перемещается в нейрон активным
транспортным насосом, а затем
преобразовывается в DA двумя из трех
энзимов, которые также синтезируют
норадреналин. Разрушение DA также
осуществляется теми же самыми ферментами,
которые разрушают норадреналин, а именно
монаминоксидазой (МАО) и катехолметилтрансферазой
(COMT). DA нейрон имеет пресинаптический
транспортер, который является уникальным
для DA, но работает аналогично
нораденалиновому и серотониновому.
Рецепторы для DA регулируют дофаминэргичекую
нейропередачу. Существует множество
дофаминовых рецепторов, известно, по
крайней мере, пять фармакологических
подтипов и несколько их молекулярных
изоформ. Наиболее изучен D2-рецептор,
потому что он стимулируется
дофаминэргическими агонистами при
лечении болезни Паркинсона, и блокируется
дофаминовыми антагонистами при лечении
шизофрении.  ^
Рис.1 Дофаминэргические проводящие пути
в ЦНС (Stephen M., Stahl, 1999) Можно
выделить четыре главные дофаминергических
системы в ЦНС: (А)
Стриопалидарная система;
нигро-стриальный дофаминовый путь
проектируется от substantia nigra к basal ganglia и,
как полагают, управляет движениями
(Рис.1). (B)
Мезолимбическая система;
мезолимбический дофаминовый путь
проектируется от среднего мозга (ventral
tegmental area) к nucleus accumbens лимбической системы,
отвечающей за многие виды эмоционального
поведения типа эффектов радости, эйфории
при употреблении наркотиков, а также
возникновения галлюцинаций при психозах.
Антипсихотическое действие классических
нейролептиков обычно связано с их
дофаминблокирующей активностью в
мезолимбической системе. (C)
Мезокортикальная система;
мезокортикальный дофаминовый путь,
тесно связан с мезолимбическим дофаминовым
путем (В). Он также проектируется от
среднего мозга (ventral tegmental area), но аксоны
проходят в лимбическую кору мозга.
Полагают, что эта система опосредует
как позитивные, так и негативные
психотические симптомы, а также
когнитивные побочные эффекты
нейролептического средства при его
применении. С мезокортикальной системой
связывают действие атипичных
нейролептиков. (D)
Нейрогипофизарная система;
тубероинфундибулярный дофаминовый
путь управляет секрецией пролактина и
проектируется от гипоталамуса
(релизинг-факторы) до
гипофиза. СЕРОТОНИН (5-hydroxytryptamine,
5HT). Локализация: ядра шва в ростральной
части моста, нейроны дают проекции к
лимбической системе, базальным ганглиям,
коре больших полушарий. 5-НТ играет
важную роль в регуляции эмоционального
поведения, двигательной активности,
пищевого поведения, сна, терморегуляции,
участвует в контроле нейроэндокринных
систем.
Серотонин производится из
триптофана, после того как эта аминокислота
транспортируется в серотониновый
нейрон. Триптофановый насос сходен с
серотониновым. Ферменты триптофангидроксилаза
и декакорбаксилаза трансформируют
триптофан в конечный продукт — серотонин,
который скапливается в синаптических
пузырьках до тех пор, пока не поступит
нейронный импульс для его выброса. 5HT
разрушается ферментом монаминоксидазой
(МАО), превращаясь в неактивный метаболит.
Серотониновый нейрон имеет пресинаптический
транспортный насос аналогичный
дофаминовому (DA). Сейчас изветны по
крайней мере четыре главных типа
серотониновых рецепторов различных по
фармакологическим и молекулярным
свойствам. Имеется ключевой пресинаптический
серотониновый рецептор — 5HТ1D-рецептор,
а также множество других постсинаптических
серотониновых рецепторов (5HTn, где n —
буквено-цифровые обозначения
подтипа).
Известно, по крайней мере,
пять проводящих путей для разнообразных
регулирующих влияний серотонина (Рис.2).
Эффекты антидепрессантов связаны с
блокадой серотонинового насоса — это
путь от шва среднего мозга до префронтальной
коры — (А). Этот проводящий путь к
префронтальной коре опосредуют влияние
серотонина на когнитивные функции.
Действие
лекарств при обсессивно-компульсивном
расстройстве, вероятно опосредовано
путем от шва среднего мозга до базальных
узлов — (В). Этот путь также теоретически
может регулировать с помощью серотонина
и движения.
^
Рис.1 Дофаминэргические проводящие пути
в ЦНС (Stephen M., Stahl, 1999) Можно
выделить четыре главные дофаминергических
системы в ЦНС: (А)
Стриопалидарная система;
нигро-стриальный дофаминовый путь
проектируется от substantia nigra к basal ganglia и,
как полагают, управляет движениями
(Рис.1). (B)
Мезолимбическая система;
мезолимбический дофаминовый путь
проектируется от среднего мозга (ventral
tegmental area) к nucleus accumbens лимбической системы,
отвечающей за многие виды эмоционального
поведения типа эффектов радости, эйфории
при употреблении наркотиков, а также
возникновения галлюцинаций при психозах.
Антипсихотическое действие классических
нейролептиков обычно связано с их
дофаминблокирующей активностью в
мезолимбической системе. (C)
Мезокортикальная система;
мезокортикальный дофаминовый путь,
тесно связан с мезолимбическим дофаминовым
путем (В). Он также проектируется от
среднего мозга (ventral tegmental area), но аксоны
проходят в лимбическую кору мозга.
Полагают, что эта система опосредует
как позитивные, так и негативные
психотические симптомы, а также
когнитивные побочные эффекты
нейролептического средства при его
применении. С мезокортикальной системой
связывают действие атипичных
нейролептиков. (D)
Нейрогипофизарная система;
тубероинфундибулярный дофаминовый
путь управляет секрецией пролактина и
проектируется от гипоталамуса
(релизинг-факторы) до
гипофиза. СЕРОТОНИН (5-hydroxytryptamine,
5HT). Локализация: ядра шва в ростральной
части моста, нейроны дают проекции к
лимбической системе, базальным ганглиям,
коре больших полушарий. 5-НТ играет
важную роль в регуляции эмоционального
поведения, двигательной активности,
пищевого поведения, сна, терморегуляции,
участвует в контроле нейроэндокринных
систем.
Серотонин производится из
триптофана, после того как эта аминокислота
транспортируется в серотониновый
нейрон. Триптофановый насос сходен с
серотониновым. Ферменты триптофангидроксилаза
и декакорбаксилаза трансформируют
триптофан в конечный продукт — серотонин,
который скапливается в синаптических
пузырьках до тех пор, пока не поступит
нейронный импульс для его выброса. 5HT
разрушается ферментом монаминоксидазой
(МАО), превращаясь в неактивный метаболит.
Серотониновый нейрон имеет пресинаптический
транспортный насос аналогичный
дофаминовому (DA). Сейчас изветны по
крайней мере четыре главных типа
серотониновых рецепторов различных по
фармакологическим и молекулярным
свойствам. Имеется ключевой пресинаптический
серотониновый рецептор — 5HТ1D-рецептор,
а также множество других постсинаптических
серотониновых рецепторов (5HTn, где n —
буквено-цифровые обозначения
подтипа).
Известно, по крайней мере,
пять проводящих путей для разнообразных
регулирующих влияний серотонина (Рис.2).
Эффекты антидепрессантов связаны с
блокадой серотонинового насоса — это
путь от шва среднего мозга до префронтальной
коры — (А). Этот проводящий путь к
префронтальной коре опосредуют влияние
серотонина на когнитивные функции.
Действие
лекарств при обсессивно-компульсивном
расстройстве, вероятно опосредовано
путем от шва среднего мозга до базальных
узлов — (В). Этот путь также теоретически
может регулировать с помощью серотонина
и движения.  ^
Рис. 2. Главные серотониновые проводящие
пути в ЦНС (Stephen M., Stahl, 1999) Путь
от шва до лимбической коры оказывает
регулирующие функции серотонина на
эмоции, включая панику и тревожность
— (С). Влияние
серотонина на пищевое поведение и
аппетит связаны с путем от шва среднего
мозга до гипоталамуса — (D). Влияние
серотонина на половое функционирование
могут быть связаны с путем, который
проектируется от шва вниз в спинной
мозг —(E). Есть
еще другие серотониновые центры в
мозговом стволе, которые влияют на
регуляцию цикла сна.
Серотониновые
нейроны из серединного шва среднего
мозга и дофаминовые нейроны из черной
субстанции проецируются на базальные
ганглии, где могут взаимодействовать,
причем серотонин тормозит выброс
дофамина.
^
Рис. 2. Главные серотониновые проводящие
пути в ЦНС (Stephen M., Stahl, 1999) Путь
от шва до лимбической коры оказывает
регулирующие функции серотонина на
эмоции, включая панику и тревожность
— (С). Влияние
серотонина на пищевое поведение и
аппетит связаны с путем от шва среднего
мозга до гипоталамуса — (D). Влияние
серотонина на половое функционирование
могут быть связаны с путем, который
проектируется от шва вниз в спинной
мозг —(E). Есть
еще другие серотониновые центры в
мозговом стволе, которые влияют на
регуляцию цикла сна.
Серотониновые
нейроны из серединного шва среднего
мозга и дофаминовые нейроны из черной
субстанции проецируются на базальные
ганглии, где могут взаимодействовать,
причем серотонин тормозит выброс
дофамина.
^ АЦЕТИЛХОЛИН (АСh). Основная локализация: медиальное ядро перегородки, диагональная связка, базальное гигантоклеточное ядро. Аксоны этих нейронов проецируются на гиппокамп, проходя через кору больших полушарий. Холинергические системы участвуют в таких функциях как память, регуляция движения, уровень бодрствования (ретикулярная формация ствола мозга, базальные ганглии). В спинном мозге ацетилхолин является нейромедиатором в синапсах, образуемых a-мотонейронами на клетках Реншоу. В вегетативной нервной системе АСh – медиатор во всем парасимпатическом отделе и в преганглионарных нервных окончаниях симпатического отдела. ACh образуется в холинергических нейронах от двух предшественников: холина и ацетилкоэнзима (Ac-CoA). Холин поступает с пищей и внутринейронных источников, а Ac-CoA образуется из глюкозы в митохондриях нейрона. Эти два субстрата взаимодействуют с ферментом ацетилтрансферазой в процессе образования ацетилхолина. ACh разрушается ферментом ацетилхолинэстеразой (AchE), которая преобразовывает ACh в неактивные продукты. Один из них — холин, который может быть закачан назад (80%) в нейрон пресинаптическим транспортером холина, подобным транспортерам для других нейромедиаторов. Имеются многочисленные рецепторы для ACh. Главное их подразделение — никотиновые (N) и мускариновые (M) холиноэргические рецепторы. Имеются также многочисленные их подтипы, лучше всего описанные для мускриновых подтипов рецептора (M1,М2, Mx). Возможно M1 постсинаптический рецептор ключевой к опосредованию функций памяти и к периферийным побочными эффектами антихолинэргических препаратов в виде сухости во рту, нечеткости зрения и др. Дофамин и ацетилхолин имеют взаимные отношения в нигростриарном дофаминовом пути. Дофаминовые нейроны здесь имеют постсинаптические связи с холинэргическими нейронами. В отсутствии дофамина холинэргический нейрон — активный. Дофамин обычно подавляет ацетилхолиновую деятельность, в присутствии дофамина холинэргическая продукция уменьшается.
НОРАДРЕНАЛИН. Наряду с адреналином и дофамином относится к катехоламинам. Локализация: в мосте (голубое пятно, латеральная ретикулярная формация моста), в продолговатом мозге и ядре одиночного тракта. Многочисленные (несколько сотен) нейроны голубого пятна образуют диффузные проекции большой протяженности, достигая практически всех отделов ЦНС – коры больших полушарий, лимбической системы, таламуса, гипоталамуса, спинного мозга. В ЦНС, как правило, – тормозной медиатор (кора), реже – возбуждающий (гипоталамус). Является медиатором во всех постганглионарных симпатических окончаниях, за исключением потовых желез. АДРЕНАЛИН. У млекопитающих мало адреналиновых путей. Адреналин секретируется диффузно (в мозговом слое надпочечников) и выполняет, в первую очередь, роль гормона.
^
ГЛУТАМИНОВАЯ КИСЛОТА (L-глутамат,
Glu). Является главным возбуждающим
медиатором, локализуется во всех отделах
ЦНС. Его преобладающее использование
в организме — не как нейромедиатор, а
как аминокислота белкового синтеза.
Когда она используется как нейромедиатор,
то она синтезируется от глютамина
посредством фермента глютаминазы в
митохондриях. Глутамат накапливается
в синаптических пузырьках для последующего
выпуска при нейротрасмиссии. Глютамин
поступает в нейрон от смежных с ним
клеток глии. Влияния глутамата
остановливаются не ферментативным
разрушением, как в других системах
нейромедиатора, а удалением двумя
транспортными насосами. Первый из этих
насосов — пресинаптический
глютамат-транспортер, который работает
подобно всем другим транспортерам
нейромедиаторов. Второй транспортный
насос расположен на близлежащей глии,
которая и удаляет глютаминовую кислоту
от синапса.
Глутамат, взаимодействуя
с глутаматными рецепторами, увеличивает
проницаемость мембраны для инов натрия,
вызывает деполяризацию и возбуждающий
эффект. Существуют много разновидностей
рецепторов глутамата, связанных с
ионными каналами: NMDA, AMPA, каиновые
рецепторы. NMDA – TV-methyl-D-aspartate; AMPA —
ct-amino-3-hydroxy-5-methyl-4-isoxazole-propionic кислота;
каиновая кислота — аминокислота из
морских водорослей с нейротоксическим
действием, разрушает тела нейронов с
глутаматными рецепторами.
Из них
наибольшое внимание уделяется
NMDA-рецепторам (см. рис3.), которые связаны
с кальциевыми каналами. В эксперименте
было показано, что блокада этих рецепторов
(например, веществом МК-801) предупреждает
дегенерацию нейронов при при ишемии
мозга. Некоторые анастетики (кетамин)
являются антогонистами
NMDA-рецепторов.
NMDA-рецептор глутамат
— кальциевого канала реагирует не
только на глутамат. Он имеет дополнительные
рецепторные участки для нейромедиатора
глицина, полиаминов и цинка. Кроме того,
ион магния может блокировать канал
кальция на одном из таких модулирующих
участков. Еще один подавляющий модуляцию
участок, который расположен внутри
канала, иногда называют «PCP участком»,
так как препарат психодислептика PCP
(phencyclidine) связывается с этим участком.
В норме NMDA-рецептор при выбросе глутамата
заставляет канал кальция открываться,
и нейрон будет возбуждаться. 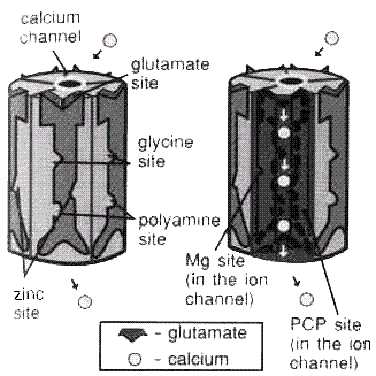 ^
Рис.3. NMDA-рецептор АСПАРАГИНОВАЯ
КИСЛОТА (L-аспартат).
Возбуждающий медиатор, который обнаружен
преимущественно в среднем мозге, а также
в переднем и заднем столбе спинного
мозга. Полагают, что его действие имеет
много общего с глутаматом.
Глутамат
и аспартат (ВАК) являются основными
медиаторами возбуждения в ЦНС. С
гиперактивностью соответствующих
систем связывают развитие многих
положительных реакций: усиление
когнитивных способностей, памяти,
обучения, контроля пищевого поведения
и др. Снижение функциональной активности
этих систем сопровождается развитием
психотических реакций, что хорошо
известно на примере использования
препаратов, обладающих антиВАКергическим
действием на уровне блокады NMDA-рецепторов
(кетамин, фенциклидин, дизоцилпин и
др.). При слабости ВАКергической передачи,
наряду с симптомами психоза, будет
присутствовать и негативная симптоматика.
Полагают, что формирование позитивной
и негативной симптоматики еще лучше,
чем с дофаминергической, согласуется
с другой концепцией патогенеза шизофрении,
а именно – антиглутамат/антиаспартатергической
(Kim J.S., Kornhuber H.H., 1982).
^
Рис.3. NMDA-рецептор АСПАРАГИНОВАЯ
КИСЛОТА (L-аспартат).
Возбуждающий медиатор, который обнаружен
преимущественно в среднем мозге, а также
в переднем и заднем столбе спинного
мозга. Полагают, что его действие имеет
много общего с глутаматом.
Глутамат
и аспартат (ВАК) являются основными
медиаторами возбуждения в ЦНС. С
гиперактивностью соответствующих
систем связывают развитие многих
положительных реакций: усиление
когнитивных способностей, памяти,
обучения, контроля пищевого поведения
и др. Снижение функциональной активности
этих систем сопровождается развитием
психотических реакций, что хорошо
известно на примере использования
препаратов, обладающих антиВАКергическим
действием на уровне блокады NMDA-рецепторов
(кетамин, фенциклидин, дизоцилпин и
др.). При слабости ВАКергической передачи,
наряду с симптомами психоза, будет
присутствовать и негативная симптоматика.
Полагают, что формирование позитивной
и негативной симптоматики еще лучше,
чем с дофаминергической, согласуется
с другой концепцией патогенеза шизофрении,
а именно – антиглутамат/антиаспартатергической
(Kim J.S., Kornhuber H.H., 1982).
ГАМК. К обычным тормозным медиаторам головного мозга относится гамма-аминомасляная кислота (ГАМК), не входящая в состав белков. ГАМК вырабатывается исключительно в головном и спинном мозгу. Не менее трети (до 50%) синапсов головного мозга используют в качестве медиатора ГАМК. Например, для хореи Гентингтона – наследственного неврологического заболевания характерен специфический дефицит ГАМК в мозгу. Возникающие при этом непроизвольные движения с наступающим в среднем возрасте постепенным разрушением полосатого тела. Посмертные исследования показали, что в основе болезни лежит дефицит ГАМК. Бенозодиазепиновые транкливизаторы являются непрямыми агонистами ГАМК. Они воздействуют на хлорные ионные каналы, соединенные с ГАМК-рецепторами. Обнаружено несколько подтипов бензодиазепиновых рецепторов: ?-1 (опосредуют седативные эффекты), ?-2 (опосредуют снижение тревоги, антиконвульсивное действие, ухудшение памяти и нарушение координации движений), ?-3 (расположены в спинном мозге, связаны с мышечно-релаксирующими эффектами). ГАМК удаляется из синаптической щели путем захвата пресинаптическим окончанием, а также клетками глии. Глия играет важную роль, как в захвате, так и в метаболизме ГАМК. Молекула медиатора связывается с мембранным белком, при этом имеется механизм очищения щели и для обратного захвата (50%). Однако последующая реакция в постсинаптическом окончании более сложна. Рецепторный белок аденилатциклаза активирует внутренний рецептор – протеинкиназу, что приводит к фосфорилированию белка. Завершается этот процесс изменением ионной проводимости мембраны. Этот механизм участвует в опосредовании реакций на такие разные вещества как, например, биогенные амины. ГЛИЦИН. В спинном мозге опосредует постсинаптическое торможения активности мотонейронов, высвобождаясь из окончаний клеток Реншоу. Также является нейромедиатором в тормозных интернейронах промежуточного мозга и ретикулярной формации. АТФ. Является медиатором во всех синапсах, образуемых метасимпатическим отделом вегетативной нервной системы на гладких мышцах. Действие АТФ при этом опосредуется пуриновыми рецепторами, сопряженными с кальциевыми ионными каналами. ^ Нейромодуляторы, комедиаторы, нейропептиды Нейромодуляторы — вещества не обладающие самостоятельным физиологическим действием, а модифицирующие эффект нейромедиаторов. Действие нейромодуляторов имеет тонический характер – медленное развитие и большую продолжительность действия. Происхождение – не обязательно нейронное, например, глия может синтезировать ряд нейромодуляторов. Действие не инициируется нервным импульсом и не всегда сопряжено с эффектом медиатора. Мишени: не только рецепторы на постсинаптической мембране, а разные участки нейрона, в том числе и внутриклеточные. Комедиаторы – это сопутствующие синаптические посредники, характеризующиеся прежде всего совместной локализацией, совместным высвобождением и общей клеткой-мишенью. Под совместной локализацией понимается синтез и депонирование медиаторов в одном и том же нейроне, их происхождение в одних и тех же пресинаптических окончаниях, но не обязательно в одних и тех же пузырьках. Под совместным высвобождением понимается экзоцитоз двух (и более) медиаторов, в результате одной и той же активации пресинаптического окончания, под которым в данном случае подразумевается не одиночный пресинаптический потенциал действия, а разряд потенциалов действия с одной и той же частотой. Нейропептиды — система химических посредников в головном мозге. Нейропептиды представляют цепочки из аминокислотных остатков (от 2 до 39). Многие из них локализованы в аксонных окончаниях. Нейропептиды отличаются от ранее идентифицированных медиаторов тем, что они организуют такие сложные явления как память, жажда, половое поведение и др. Например, соматостатин подавляет выделение гормона роста гипофизом, регулирует секрецию инсулина и глюкагона поджелудочной железой и функционирует в качестве медиатора в спинном и головном мозге.
Нейромедиатор (нейротрансмиттер, нейропередатчик) – это вещество, которое синтезируется в нейроне, содержится в пресинаптических окончаниях, высвобождается в синаптическую щель в ответ на нервный импульс, и действует на специальные участки постсинаптической клетки, вызывая изменения мембранного потенциала и метаболизма клетки.
Существует 4 типа медиаторов:
амины;
аминокислоты;
пуриновые нуклеотиды;
нейропептиды.
Классические медиаторы и их постсинаптическое действие. Ацетилхолин.
Основная локализация: медиальное ядро перегородки, диагональная связка, базальное гигантоклеточное ядро. Аксоны этих нейронов проецируются на гиппокамп, проходя через кору больших полушарий.
Холинергические системы участвуют в таких функциях как память, регуляция движения, уровень бодрствования (ретикулярная формация ствола мозга, базальные ганглии).
В спинном мозге ацетилхолин является нейромедиатором в синапсах, образуемых a-мотонейронами на клетках Реншоу. В вегетативной нервной системе АХ – медиатор во всем парасимпатическом отделе и в преганглионарных нервных окончаниях симпатического отдела.
Норадреналин.
Наряду с адреналином и дофамином относится к катехоламинам.
Локализация: в мосте (голубое пятно, латеральная ретикулярная формация моста), в продолговатом мозге и ядре одиночного тракта. Многочисленные (несколько сотен)нейроны голубого пятна образуют диффузные проекции большой протяженности, достигая практически всех отделов ЦНС – коры больших полушарий, лимбической системы, таламуса, гипоталамуса, спинного мозга.
В ЦНС, как правило, – тормозной медиатор (кора), реже – возбуждающий (гипоталамус). Является медиатором во всех постганглионарных симпатических окончаниях, за исключением потовых желез.
Адреналин.
У млекопитающих мало адреналиновых путей. Адреналин секретируется диффузно (в мозговом слое надпочечников) и выполняет, в первую очередь, роль гормона.
ДОФАМИН.
Локализуется в среднем мозге (черная субстанция, вентральная покрышка), обонятельной луковице, гипоталамусе и перивентрикулярной области продолговатого мозга.
СЕРОТОНИН.
Локализация: ядра шва в ростральной части моста, эти нейроны дают проекции к лимбической системе, базальным ганглиям, коре больших полушарий.
5-НТ (5-гидрокситриптамин) играет важную роль в регуляции эмоционального поведения, двигательной активности, пищевого поведения, сна, терморегуляции, участвует в контроле нейроэндокринных систем.
L-ГЛУТАМИНОВАЯ КИСЛОТА.
Является главным возбуждающим медиатором, локализуется во всех отделах ЦНС.
АСПАРАГИНОВАЯ КИСЛОТА.
Возбуждающий медиатор в среднем мозге, и в переднем и заднем столбе спинного мозга.
ГАМК.
К обычным тормозным медиаторам головного мозга относится g-аминомасляная кислота (ГАМК), не входящая в состав белков. ГАМК вырабатывается исключительно в головном и спинном мозгу. Не менее трети (до 50%) синапсов головного мозга используют в качестве медиатора ГАМК. Например, для хореи Гентингтона – наследственного неврологического заболевания характерен специфический дефицит ГАМК в мозгу. Возникающие при этом непроизвольные движения с наступающим в среднем возрасте постепенным разрушением полосатого тела. Посмертные исследования показали, что в основе болезни лежит дефицит ГАМК.
studfiles.net
Нейромедиаторы рецепторы — Справочник химика 21
| Рис. 8.2. Схема никотинового холинэргического синапса. Пресинаптическое нервное окончание содержит компоненты для синтеза нейромедиатора (здесь ацетилхолина). После синтеза (I) нейромедиатор упаковывается в пузырьки (везикулы) (II). Эти синаптические везикулы сливаются (возможно, вре.мен-но) с пресинаптической мембраной (1П), и нейромедиатор высвобождается таким путем в синаптическую щель. Он диффундирует к постсинаптической мембране и связывается там со специфическим рецептором (IV). В результате образования нейромедиатор-рецепторного комплекса постсинаптическая мембрана становится проницаемой для катионов (V), т. е. деполяризуется. (Если деполяризация достаточно высока, то появляется потенциал действия, т. е. химический сигнал снова превращается в электрический нервный импульс.) Наконец, медиатор инактивируется , т. е. либо расщепляется ферментом (VI), либо удаляется из синаптической щели посредством особого механизма поглощения . В приведенной схеме только один продукт расщепления медиатора— холин — поглощается нервным окончанием (VII) и используется вновь. Базальная мембрана — диффузная структура, идентифицируемая методом электронной микроскопии в синаптической щели (рис. 8.3,а), здесь не показана. |
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30—80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит 40000 молекул ацетилхолина. При возбуждении высвобождение медиатора происходит квантами , т.е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100—200 квантов медиатора—количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Происходит это, по-видимому, следующим образом. Деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов Са в клетку. Временное увеличение внутриклеточной концентрации ионов Са стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуется примерно 4 иона Са . Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны —резко увеличивается ее пропускная способность для ионов Ка. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации, или удаления, чтобы подготовить синапс к восприятию нового импульса. [c.638]
Особого внимания заслуживает ненасыщенная жирная кислота, называемая арахидоновой. Она высвобождается фосфолипазой Аг и выступает как в роли предшественника простаглан-динов, тромбоксанов и лейкотриенов, так и в роли важного клеточного регулятора (или сама по себе, или после окисления липоксидазой). Было показано, например, что арахидоновая кислота стимулирует гуанилатциклазу обусловленное нейромедиатором образование сОМР (гл. 9), видимо, регулируется высвобождением арахидоновой кислоты, опосредованным рецептором. С другой стороны, образование сАМР в ряде тканей регулируется, вероятно, некоторыми простагландинами. [c.44]
Как известно, клетки нервной системы (нейроны) не имеют непосредственного контакта друг с другом. Они разделены синаптическими щелями, через которые сигнал (передаваемый в виде бегущей по нейронной мембране волны поляризации-деполяризации) пройти не может без определенного посредника, называемого нейромедиатором (или нейротрансмиттером). Передача нервного импульса от одного нейрона к другому происходит следующим образом (рис. 3, схема А). По достижении нервным сигналом конца возбужденной клетки (нейрон 1) в ее пресинаптической области синтезируется нейротрансмиттер (АХ), который затем выбрасывается в синаптическую щель и быстро диффундирует к своему рецептору (R), расположенному в постсинаптической мембране покоящейся клетки (нейроне 2). [c.31]
Какова природная функция рецепторов алкалоидов опия Логично предположить, что они предназначены для связывания каких-то нейромедиаторов или модуляторов. В последние годы было показано, что следующие два пентапептида (называемые энкефалинами), а таюке [c.345]Скорость переноса метаболитов через мембраны, разделяющие различные отсеки клетки, ограничена и строго контролируется на поверхности мембран расположены рецепторы, являющиеся мишенями для большинства гормонов и нейромедиаторов эти рецепторы могут подвергаться химической модификации. [c.68]
На третьей — фармакодинамической — стадии изучаются проблемы распознавания лекарственного вещества (или его метаболитов) мишенями и их последующего взаимодействия. Мишенями могут служить органы, ткани, клетки, клеточные мембраны, ферменты, нуклеиновые кислоты, регуляторные молекулы (гормоны, витамины, нейромедиаторы и т.д.), а также биорецепторы. Рассматриваются вопросы структурной и стереоспе-цифичной комплементарности взаимодействующих структур, функционального и химического соответствия лекарственного вещества или метаболита (например, фармакофорной группировки) его рецептору. Взаимодействие между лекарственным веществом и рецептором или акцептором, приводящее к активации (стимулированию) или дезактивации (ингибированию) биомишени и сопровождающееся ответом организма в целом, в основном обеспечивается за счет слабых связей — водородных, электростатических, ван-дер-ваальсовых, гидрофобных. [c.13]
Наряду с АХ известны и другие нейромедиаторы. Таковыми являются адреналин, норадреналин, дофамин и некоторые другие вещества. После связывания АХ с рецептором открываются [c.385]
Рецепторы нейромедиаторов исключительно важны для молекулярной нейробиологии, так как они играют ключевую роль при переносе нервных импульсов и являются центрами важных регуляторных процессов и тех изменений, которые происходят при некоторых нервных заболеваниях. Поскольку такие рецепторы представляют собой мишень действия многих нейрофармакологических препаратов, они представляют как практический, так и теоретический интерес. Поэтому биохимия рецепторных молекул — одно из наиболее активно развивающихся сейчас направлений нейрохимических исследований. [c.241]
До сих пор только один рецептор, никотиновый ацетилхолиновый рецептор, был тщательно очищен и биохимически охарактеризован. Но этого оказалось мало для создания общей теории связи между структурой рецептора и его функцией. Очевидно, что рецепторы имеют двойную функцию они принимают и узнают специфические сигналы и одновременно инициируют первую стадию клеточного ответа на сигнал. Наряду с другими своими функциями рецепторы нейромедиатора регулируют ионную проницаемость постсинаптической мембраны (рис. 9.1) Связывание молекулы медиатора сопряжено, таким образом, с ответным открытием ионных каналов. Вопрос механизма функционирования рецепторов сводится по сути к представлению а механизме такого сопряжения. [c.243]
При изучении нейромедиаторов важное значение имеет подбор специфических агонистов, имитирующих действие медиатора, или антагонистов, блокирующих это действие. В зависимости от чувствительности к одной или другой группе соединений холинэргические нейроны делятся на мускариновые (активируемые мускарином, рис. 16-6) или никотиновые (активируемые никотином) [46]. Мускариновые рецепторы, имеющиеся во многих нейронах автономной нервной системы, специфически блокируются атропином и декаметонием (рис. 16-6). Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда D-тубо-курарин (рис. 16-6), а также белок из змеиного яда а-бунгаротоксин (рис. 16-7). Этот токсин был, в частности, использован для титрования рецепторов ацетилхолина в моторной концевой пластинке диафрагмы крысы. Было показано, что количество рецепторов в расчете на одну пластинку составляет примерно 4-10 (или 13000 рецепторов на [c.332]
Исходя из такой интерпретации, рецептор нейромедиатора (или гормона) должен существовать по крайней мере в двух состояниях неактивном или состоянии покоя (I) и в активном (А) состоянии (рис. 9.2). Равновесие между этими состояниями зависит от природы нейромедиатора (или гормона). Соединения, которые, подобно медиатору, благоприятствуют активному состоянию, называются агонистами, а те, которые смещают равновесие в сторону неактивного состояния,— антагонистами. Обсудим несколько моделей, используемых для количественного анализа результатов и установления взаимосвязи между концентрацией и ответом в случае медиаторов и других фармакологически активных соединений. [c.247]
Для любого специалиста в этой области — работает ли он в основном с ферментами или с рецепторами — трудно отличить одну модель от другой. В случае нейромедиаторов интерпретация экспериментальных данных даже более затруднена, так как антагонист всегда ингибирует связывание агониста. Он может также ингибировать одну из стадий процесса, протекающую после связывания, например транспорт ионов через открытый канал, закрыв его как пробка в трубке, или сопряжение между связывающим центром медиатора и ионным каналом, т. е. открывание канала. Первый механизм, по-видимому, лежит в основе действия многих местных анестетиков, тогда как второй относится к некоторым эффекторам адренэргических рецепторов (см. ниже). [c.248]
Физиологически рецепторы функционируют как регуляторные белки. Число, сродство и активность рецепторов находятся под контролем различных механизмов регуляции. Они также являются местом действия многочисленных экзогенных эффекторов, а именно лекарств и токсинов. Некоторые заболевания нервной системы имеют рецепторную природу (миастения и, возможно, шизофрения). Некоторые, так называемые рецепторы, особенно участки связывания лекарств, могут быть в действительности регуляторными связывающими центрами или субъединицами истинных комплексов нейромедиатор — рецептор. Таким образом, мягкие транквилизаторы, бенздиазепины и барбитураты, которые усиливают ингибиторное действие GABA-эргических нейронов, по-видимому, действуют путем стимуляции связывания GABA с ее рецептором. [c.300]
На многих тканях показано, что десенсибилизация рецепторов — это не разрушение, не инактивация гормонсвязывающих белко В, а восстановление чувствительности — это не синтез белка de novo. При длительном воздействии гормонов и нейромедиаторов рецепторы переходят в латентное состояние. Устранение гормона из организма или же применение антагониста приводит к демаскировке соответствующих рецепторов. [c.152]
Он образует цилиндрический канал, который с одной стороны выступает на 65 А в синаптическую щель, а с другой — пронизывает липидный бцслой мембраны, входя на 15 А внутрь клетки. Этот узкий канал (или пора) расширяется до 20 А при «посадке» на рецептор нейромедиатора (комплекс RAX) за счет резкого уменьшения вращательного (конформационного) движения субъединиц. Увеличение размера канала облегчает прохождение ионов К+ и Na+ через мембрану против электрохимического фадиента. При этом изменяется мембранный потенциал покоящегося нейрона 2, и в нем генерируется нервный импульс. После этого нейромедиатор гидролизуется ацетилхолинэстера-зой до неактивного холина, и ионофорныи канал закрывается. [c.31]
В 1999 г. был открыт необычный нейромедиатор — D-серин, HO h3 H(Nh3) OOH. Оказалось, что эта правосторонняя а-аминокислота вырабатывается в организме человека из левосторонних а-аминокислот (из их L-форм). Еще одна неожиданность заключалась в том, что биосинтез D-серина осуществляется не в самих нейронах, а в астроцитах — клетках, покрывающих нейроны. Из астроцита этот нейромедиатор затем диффундирует в нервную клетку и взаимодействует со специальными рецепторами. Начинается разработка лекарственных веществ, регулирующих активность фермента, контролирующего синтез D-серина. Эти лекарства, как ожидается, могут оказаться полезными при инсультах, гипертонических кризах и помогут защищать нейроны от необратимых повреждений. [c.38]
Механизм их биодействия связан, по-видимому, с активацией главного нейромедиатора торможения в ЦНС — у-аминомасляной кислоты (ГАМК) — посредством воздействия на специфические бензодиазепиновые рецепторы. [c.174]
По данным [1031, в отличие от [101], некоторые другие органы крыс также способны в незначительной степени связывать диазепам. Немеченый диазепам и другие бенздиазепины вытесняли его радиоактивный аналог, который был связан с митохондриями печени, почек и легких крыс. Вещество Ro 4884, которое очень слабо вытесняло диазепам, связанный с мембранами мозга, обладало чрезвычайно выраженной способностью вытеснять диазепам, связанный с митохондриями почек. В то же время клоназепам — сильный ингибитор взаимодействия диазепама с мембранами мозга — слабо влиял на сродство последнего к митохондриям почек. Специфическое связывание Н-диазепама с препаратами толстого и тонкого кишечника, а также скелетных мышц не выявлено. Трипсин химотрипсин полностью подавляли специфическое связывание диазепама с препаратами мозга и почек. Таким образом, рецепторы для бенздиазепинов отличаются от всех известных в мозгу рецепторов, с которыми взаимодействуют нейромедиаторы. [c.263]
Д-специфич нейромедиатор для дофаминовых рецепторов, в больших дозах стимулирует также а- и р-адре-норецепторы и т обр увеличивает сердечный выброс, вызывая небольшие изменения артериального давления, а также силу и частоту сердечных сокращений без увеличения общего периферич сопротивления В отличие от адреналина и норадреналина уменьшает сопротивление сосудов кишечника и почек, увеличивает почечный кровоток и диурез (см также Адреномиметические средства и Kamexoi-амины) [c.115]
Осн. ф-ция К.-активация мн. ферментов аденилатциклазы, фосфодиэстеразы циклич. нуклеотидов, киназы фосфо-рилаз и легких цепей миозина (киназы-ферменты, катализирующие перенос фосфорильной группы с АТФ на субстрат), Са -зависимой протеинкиназы цитоплазмы и мембран, фосфолипазы Aj и др. Благодаря этому он влияет на гликогенолиз и липолиз, секрецию нейромедиаторов, адренергич. передачу регуляторного сигнала, изменяет функциональные св-ва рецепторов, ускоряет активный транспорт Са в сердце и мозге, препятствует гуанозинтрифосфат-зависимой полимеризации тубулина (белок, из к-рого состоят жгутики и реснички клеток животных и растений), влияет на скорость деления клеток. [c.293]
Ингибирование ферментов лежит в основе действия антибиотиков и других химиотерапевтических препаратов (см., например, дополнение 6-А). Однако многие лекарственные препараты взаимодействуют с рецепторами, расположенными на клеточной поверхности, которые не являются ферментами в обычном смысле этого слова. Согласно теории рецепторов, разработанной примерно в 1937 г., близкие по структуре лекарственные препараты часто оказывают аналогичное действие, поскольку связываются с одним и тем же рецептором. В нормальных условиях рецептор может связывать гормон, нейромедиатор или какой-либо метаболит, структурно близкий лекарственному препарату. С»вязывание с соответствующим рецептором препаратов одного класса, называемых в фармакологической литературе агонистами, вызывает в клетке ту же реакцию, что и связывание гормона. В то же время соединения с родственной структурой могут. действовать и как антагонисты связывание их с рецептором не вызывает должного ответа. Вза имоотношения агониста и антагониста часто носят конкурентный характер, подобный конкурентному ингибированию ферментов. [c.32]
В книге четко сформулированы основные проблемы и достижения нейрохимии, выделены ключевые вопросы. Обсуждение построено с привлечением большого числа примеров разнообразных объектов, процессы функционирования нейроклеток рассмотрены во взаимосвязи с другими сторонами деятельности живых организмов. В книгу включен прекрасный иллюстративный материал, помогающий усваивать довольно непростые концепции и гипотезы, касающиеся наиболее сложных высших функций живой природы — работы нервной системы. Строение и функции нейрональных мембран, механизмы синаптической передачи и характеристика рецепторов нейромедиаторов, ионные каналы и активный транспорт — вот наиболее важные и существенные проблемы, которые подробно рассмотрены. В книге хорошо отражена связь нейрохимии с развитием других смежных направлений — нейрофизиологии, нейрофармакологии, нейроэндокринологии и т. д. [c.5]
Трансдуцин, также называемый G-белком, удивительно похож на N-белки — описанные в гл. 9 мембранные компоненты, передающие некоторые гормональные сигналы или сигналы нейромедиаторов от их рецепторов к ферменту аденилатциклазе. И трансдуцин, и N-белки состоят из трех полипептидных цепей действия, по-видимому, очень сходны. Подобие системы родопсин — трансдуцин — фосфодиэстераза и системы -адренэргический рецептор — N-белок — аденилатцик-лаза так велико, что возможна, например, перекрестная рекомбинация (замена) отдельных компонентов этих систем. В одном из таких экспериментов по реконструкции было показано, что трансдуцин способен передавать сигналы от -рецепторов к аденилатциклазе в клетках с недостаточным количеством N-белка. [c.18]
Неясно, почему А] (или Р2) вызывает заболевание более того, почему симптохмы проявляются на относительно малом участке пораженного организма Предполагают [20], что причина, вероятно, связана со структурной аналогией между триптофаном в положении 116 на А1 (см. выше) и нейромедиатором серотонином (5-гидрокситриптамином) ЭАЭ, таким образом, возникает как иммунный ответ на рецептор серотонина, препят-ствуюший нормальному действию серотонина в качестве нейро-медиатора в специфических областях центральной нервной системы. Если данное предположение удастся доказать, то, возможно, это будет первый шаг к хемотерапии заболевания. [c.106]
Как мы увидим в гл. 9, специфичность и разнообразие эффектов, вызываемых относительно небольшим количеством нейромедиаторов, присутствующих в нервной системе, значительно возрастает благодаря механизму множественных рецепторов. Так, например, а- и р-рецепторы подразделяются на а и ссг, Р1 и Рг. Имеются Вр и Ог-рецепторы в допаминэргических и Нг и Нг-рецепторы в гистаминэргических окончаниях. Мускариновый и никотиновый ацетилхолиновый рецепторы и г-, у.- и [c.221]
Опиаты — это лекарства с болеутоляющим и эйфорическим действием. В мозге были обнаружены специфические рецепторы этих лекарств, и фактически, как уже отмечалось, в настоящее время известно несколько типов опиатных рецепторов. Давно предполагалось, что должны существовать и эндогенные лиганды для этих рецепторов, но только в 1974 г. группами Костер-лица и Хьюза и Терениуса и Валстрёма были открыты в экстрактах мозга пептиды, которые обладали свойствами, ожидаемыми для эндогенных опиатов. Пока еще неясно, являются ли эти пептиды истинными нейромедиаторами или просто нейромодуляторами здесь приведен краткий обзор имеющихся по этому вопросу данных. [c.233]
В настоящее время термин рецептор применяется в двух различных значениях. Во-первых, этим термином обозначают первичные приемники сенсорных стимулов — света, осязания, температуры и боли. В этом смысле рецептор представляет собой орган, состоящий из одной или более клеток палочки и колбочки ретины (сетчатки) являются, например, фоторецепторами. Во-вторых, термин рецептор описывает на молекулярном уровне связывающий центр для низкомолекулярного активного соединения. Такое определение опять-таки не вполне точно многие исследователи считают рецептором любой центр, который специфично связывает лиганд независимо от их эндогенного или экзогенного происхождения. Нейрохимики же имеют в виду исключительно центры — мишени эндогенных эффекторов типа гормонов, простагландинов и нейромедиаторов. Согласно такому толкованию, термин рецептор не охватывает участки связывания нейротоксинов в аксональных ионных каналах или на ганглиозидах нервной мембраны он относится в основном к пре- и постсинаптическим рецепторам, которые всегда являются белками, связывающими пресинаптически высвобождающийся медиатор и тем самым обеспечивающими первую стадию химического возбуждения мембраны. Данное определение не исключает того факта, что такие рецепторы, как опиатный, обнаружены и охарактеризованы с помощью экзогенных лекарственных препаратов, и это особенно справедливо в тех случаях когда эндогенный медиатор еще неизвестен. [c.241]
Здесь могут быть использованы методы, известные из энзимологии и белковой химии равновесный диализ, ультрацентрифугирование, гель-хроматография и ультрафильтрация. Основной проблемой является здесь выяснение различий между специфическим и неспецифическим связыванием, в особенности при изучении связывания, проводимом на препаратах мембран и на частично очищенных белковых фракциях. В центральной нервной системе концентрации рецептора могут составлять несколько пикомолей на 1 г ткани, а нейромедиаторы благодаря своим полярным свойствам легко взаимодействуют, хотя и слабо, с полярными мембранными компонентами, расположенными вне рецепторной области. Если даже такое неспецифическое связывание имеет сродство на несколько порядков ниже, чем специфическое связывание с рецептором (т. е. более высокую Ко), оно все же оказывается важным фактором, который следует принимать во внимание из-за большого числа присутствующих неспецифических связывающих центров. Таким образом, обнаружение рецептора в центральной нервной системе посредством изучения связывания возможно, если имеется лиганд с очень высоким сродством и с очень высокой удельной радиоактивностью (>185-10 ° раоп./(с-ммоль). [c.249]
Для некоторых гормонов и нейромедиаторов вместо сАМР в качестве регулируемого циклического нуклеотида выступает сОМР (рис. 9.12). Так, если допамин, серотонин, адреналин (для рецепторов. 1, р2, аг), гистамин (для Нг-рецептора), октопамин и пептидные нейромедиаторы регулируют систему сАМР, то аце- [c.275]
Этот первый рецептор нейромедиатора, выделенный из центральной нервной системы, имеет следующие биохимические характеристики он является гликапротеином, состоящим из нескольких полипептидных цепей (по данным электрофореза в ДСН- полиакриламидН Ом геле М 48 000, 59 000 и 92 000) центр связывания стрихнина, по-видимому, локализован на самой короткой полипептидной цепи, и функции других цепей в настоящее время неизвестны рецепторный белок содержит единственный тип независимых связывающих участков для стрихнина. [c.295]
chem21.info
Медиаторы и рецепторы центральной нервной системы — МегаЛекции
Медиатор (лат. mediator — посредник) — химическое вещество, с помощью которого сигнал передается от одной клетки к другой. В головном мозге к настоящему времени обнаружено около 30 БАВ (табл. 5).
Таблица 5. Основные медиаторы и нейропептиды ЦНС: место синтеза и физиологические эффекты
| Вещество | Синтез и транспорт | Физиологическое действие |
| Норадреналин (возбуждающий медиатор) | Ствол мозга, гипоталамус, ретикулярная формация, лимбическая система, симпатический отдел ВНС | Регуляция настроения, эмоциональные реакции, поддержание бодрствования, формирование сна, сновидений |
| Дофамин (допамин) (возбуждающий, может оказывать тормозное действие) | Средний мозг, черная субстанция, лимбичесая система | Формирование чувства удовольствия, регуляция эмоциональных реакций, поддержание бодрствования |
| Влияние на полосатое тело (бледный шар, скорлупа) базальных ганглиев | Участе в регуляции сложных движений | |
| Серотонин (возбуждающий и тормозной медиатор) | Спинной мозг, ствол мозга (ядро шва), головной мозг, гипоталаму, таламус | Терморегуляция, формирование болевых ощущений, сенсорное восприятие, засыпание |
| Ацетилхолин (возбуждающий медиатор) | Спинной и голвной мозг, ВНС | Возбуждающие влияние на эффекторы |
| ГАМК (гамма-аминомасляная кислота) тормозной медиатор | Спинной и головной мозг | Сон, торможение в ЦНС |
| Глицин (тормозной медиатор) | Спинной и головной мозг | Торможение в ЦНС |
| Ангиотензин II | Ствол мозга, гипоталамус | Повышение давления, торможение синтеза катехоламинов, стимуляция синтеза гормонов, информирует ЦНС об осмотическом давлении крови |
| Олигопептиды: | Лимбическая система, гипофиз, гипоталамус | Эмоциональные реакции, настроение, половое поведение |
| 1.Веществ Р | Передача болевого возбуждения от периферии в ЦНС, формирование болевых ощущений | |
| 2.Энкефалины, эдорфины | Антиболевые (обезболивающие) рекции головного мозга | |
| 3.Пептид, вызывающий дельта-сон | Повышение устойчивости к стрессу, сон | |
| 4.Гастрин | Информирует мозг о пищевой потребности | |
| Простогландины | Кора больших полушарий, мозжечок | Формирование болевых ощущений, повышение свертываемости крови; регуляциятонуса гладких мышц; усиление физиологическог эффекта медиаторов и гормонов |
| Моноспецифические белки | Различные отделы головного мозга | Влияние на процессы обучения, память, биоэлектрическую активность и химическую чувствительность нервных клеток |
Вещество, из которого образуется медиатор (предшественник медиатора), попадает в сому или аксон из крови и ликвора, в результате биохимических реакций под действием ферментов превращается в соответствующий медиатор, затем транспортируется в синаптические везикулы. Медиатор может синтезироваться в теле нейрона или его окончании. При передаче сигнала с нервного окончания на другую клетку медиатор высвобождается в синаптическую щель и действует на рецептор постсинаптической мембраны. Как отмечалось выше, по механизму реагирования на медиатор все эффекторные рецепторы подразделяют на ионотропные и метаботропные. Большинство ионотропных и метаботропных рецепторов связано с G-белками (ГТФ-связывающие белки).
При действии медиатора на ионотропные рецепторы открываются ионные каналы непосредственно с помощью G-белка, и вследствие движения ионов в клетку или из клетки формируются ВПСП или ТПСП. Ионотропные рецепторы называют также рецепторами быстрого ответа (например, N-холинорецептор, ГАМК1-, глицино-, 5-НТ3(S3)- серотонинорецепторы).
При действии медиатора на метаботропные рецепторы ионные каналы активируются через G-белок с помощью вторых посредников. Далее формируются ВПСП, ПД, ТПСП (электрофизиологические явления), с помощью которых запускаются биохимические (метаболические) процессы; при этом возбудимость нейрона и амплитуда ВПСП могут быть повышенными в течение секунд, минут, часов и даже дней. Вторые посредники могут также изменять активность ионных каналов.
Амины (дофамин, норадреналин, серотонин, гистамин) встречаются в разных отделах ЦНС, в значительных количествах — в нейронах ствола мозга. Амины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, в черной субстанции, в лимбической системе, в полосатом теле.
Серотонин является возбуждающим и тормозным медиатором в нейронах ствола мозга, тормозным — в коре большого мозга. Известно семь типов серотонинорецепторов (5-НТ, Б-рецепторы), большинство из них метаботропные (вторые посредники — цАМ Ф и ИФ3/ДАГ). Ионотропным является S3-рецептор (имеется, в частности, в ганглиях ВНС). Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в ядрах шва (ЯШ), лимбической системе. Аксоны этих нейронов проходят в бульбоспинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преганглионарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих симпатических нейронов (а может быть, и все) являются серотонинергическими нейронами ВНС. Их аксоны, согласно последним данным, идут к органам ЖКТ и оказывают мощное стимулирующее влияние на его моторику. Повышение уровня серотонина и норадреналина в нейронах ЦНС типично для маниакальных состояний, снижение — для депрессивных.
Норадреналин является возбуждающим медиатором в гипоталамусе, в ядрах эпиталамуса, тормозным — в клетках Пуркинье мозжечка. В ретикулярной формации (РФ) ствола мозга и гипоталамусе обнаружены α- и β-адренорецепторы. Норадренергические нейроны сконцентрированы в области голубого пятна (средний мозг), где их насчитывается всего несколько сотен, но ответвления их аксонов встречаются по всей ЦНС.
Дофамин является медиатором нейронов среднего мозга, гипоталамуса. Дофаминорецепторы подразделяют на Д1— и Д2-подтипы. Д1-рецепторы локализуются на клетках полосатого тела, действуют посредством дофаминчувствительной аденилатциклазы, как и Д2-рецепторы. Последние обнаружены в гипофизе.
При действии на них дофамина угнетаются синтез и секреция пролактина, окситоцина, меланоцитстимулирующего гормона, эндорфина. Д2-рецепторы найдены на нейронах полосатого тела, где их функция пока не очень ясна. Содержание дофамина в нейронах ЦНС повышено при шизофрении и снижено при паркинсонизме.
Гистамин реализует свое влияние с помощью вторых посредников (цАМФ и ИФ3/ДАГ). В значительной концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса — здесь же локализовано основное количество гистаминергических нейронов. В остальных отделах ЦНС уровень гистамина очень низок. Медиаторная роль гистамина изучена мало. Выделяют Н1-, Н2— и Н3-гистаминорецепторы. Н1-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, в терморегуляции, секреции пролактина и антидиуретического гормона (АДГ). Н2-рецепторы обнаружены на глиальных клетках.
Ацетилхолин встречается в коре большого мозга, в спинном мозге. Известен в основном как возбуждающий медиатор; в частности, является медиатором α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина α-мотонейроны по коллатералям своих аксонов передают возбуждающее влияние на тормозные клетки Реншоу; ацетилхолин имеется в РФ ствола мозга, в гипоталамусе. Обнаружены М- и N-холинорецепторы. Установлено семь типов М-холинорецепторов; основными являются и М1— и М2-рецепторы. М1-холинорецепторы локализуются на нейронах гиппокампа, полосатого тела, коры большого мозга, М2—холинорецепторы — на клетках мозжечка, ствола мозга. N—холинорецепторы довольно плотно расположены в области гипоталамуса и покрышки. Эти рецепторы изучены достаточно хорошо, они выделены с помощью α-бунгаротоксина (основной компонент яда ленточного крайта) и α-нейротоксина, содержащегося в яде кобры. При взаимодействии ацетилхолина с N-холинорецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. При взаимодействии ацетилхолина с М-холинорецептором активация ионных каналов (К+, Са2+) осуществляется с помощью вторых внутриклеточных посредников (цАМФ — циклический аденозинмонофосфат — для М2-рецептора; ИФ3/ДАГ — для М1рецептора).
Ацетилхолин активирует и тормозные нейроны с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Аминокислоты. Глицин и γ-аминомасляная кислота (ГАМК) являются тормозными медиаторами в синапсах ЦНС и действуют на соответствующие рецепторы, глицин — в основном, в спинном мозге, ГАМК — в коре большого мозга, мозжечке, стволе мозга, спинном мозге. Передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы α-глутамат и α-аспартат . Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Глутамат — это основной возбуждающий медиатор ЦНС (75% возбуждающих синапсов мозга). Глутамат реализует свое влияние посредством метаботропных (связанных с активацией цАМФ и ИФ3/ДАГ) и ионотропных (связанных с К+-, Са2+-, Na+-ионным и каналами рецепторов).
Полипептиды встречаются в синапсах различных отделов ЦНС.
Энкефалины и эндорфины — опиоидные медиаторы нейронов, блокирующих, например, болевую импульсацию. Реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного мозга и солитарного тракта, имеются и на клетках голубого пятна, спинного мозга. Их лигандами являются (β-эндорфин, динорфин, лей- и метэнкефалины. Различные опиатные рецепторы обозначаются буквами греческого алфавита: α, ε, κ, μ, χ.
Вещество Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга. Это позволило предположить, что вещество Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Большое количество вещества Р содержится в гипоталамической области. Различают два вида рецепторов вещества Р: рецепторы типа 8Р-Е (Р1, расположенные на нейронах коры большого мозга, и рецепторы типа 8Р-Р (Р2), расположенные на нейронах мозговой перегородки.
Вазоинтестинальный пептид (ВИП), соматостатин, холецистокинин (ХЦК) также выполняют медиаторную функцию. ВИП-рецепторы и рецепторы к соматостатину выявлены на нейронах головного мозга. Рецепторы к ХЦК обнаружены на клетках коры большого мозга, хвостатого ядра, обонятельных луковиц. Действие ХЦК на рецепторы повышает проницаемость мембран для Са2+ посредством активации аденилатциклазной системы.
Ангиотензин участвует в передаче информации о потребности организма в воде. Рецепторы к ангиотензину обнаружены на нейронах коры большого мозга, среднего и промежуточного мозга. Связывание ангиотензина с рецепторами вызывает увеличение проницаемости клеточных мембран для Са2+. Эта реакция обусловлена процессами фосфорилирования мембранных белков вследствие активации аденилатциклазной системы и изменением синтеза простагландинов.
Люлиберин участвует в формировании половой потребности.
Пурины (АТФ, аденозин, АДФ) выполняют в основном моделирующую функцию. В частности, АТФ в спинном мозге выделяется вместе с ГАМК. Рецепторы к АТФ весьма разнообразны: Одни из них ионотропные, другие — метаботропные. АТФ и аденозин ограничивают перевозбуждение ЦНС и участвуют в формировании болевых ощущений.
Гипоталамические нейрогормоны, регулирующие функцию гипофиза, также выполняют медиаторную роль.
Физиологические эффекты действия некоторых медиаторов головного мозга. Дофамин участвует в формировании чувства удовольствия, в регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений. Серотонин ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание. Эндорфины, энкефалины, пептид, дают антиболевые эффекты, повышают устойчивость к стрессу, способствуют сну. Простагландины вызывают повышение свертываемости крови, изменение тонуса гладких мышц, усиливают физиологические эффекты медиаторов и гормонов. Олигопептиды — медиаторы настроения, полового поведения, передачи ноцицептивного возбуждения от периферии к ЦНС, формирования болевых ощущений.
В последние годы получены факты, вызвавшие необходимость внесения коррективов в известный принцип Дейла. Так, согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор во всех разветвлениях своего аксона («один нейрон — один медиатор»). Однако выяснилось, что, кроме основного медиатора, в окончаниях аксона могут выделяться и другие, сопутствующие медиаторы (комедиаторы), играющие модулирующую роль или более медленно действующие. Кроме того, в тормозных нейронах в спинном мозге в большинстве случаев имеется два быстродействующих типичных медиатора в одном тормозном нейроне — ГАМК и глицин.
Таким образом, нейроны ЦНС возбуждаются или тормозятся, в основном, под влиянием специфических медиаторов.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны и вторых посредников. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — только торможение, в синапсах гладкой мускулатуры ЖКТ — только возбуждение. Катехоламины тормозят сокращения желудка и кишечника, но стимулируют сердечную деятельность. Глутамат является только возбуждающим медиатором ЦНС.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru