
Проводимость это в физиологии – «Лекция 1. Физиология возбудимых тканей Кафедра физиологии им. А.Т. Пшоника Проф. Ю.И. Савченков.». Скачать бесплатно и без регистрации.
Проводимость и сократимость. Нормальная физиология
Проводимость и сократимость
Между клетками проводящей системы и рабочим миокардом имеются тесные контакты в виде нексусов, поэтому возбуждение, возникшее в одном участке сердца, проводится без затухания (без декремента) в другой. Скорость распространения возбуждения от предсердий к желудочкам составляет 0,8–1,0 м/с. Проходя атриовентрикулярный узел, возбуждение задерживается на 0,04 с. Далее, распространившись по пучку Гиса и волокнам Пуркинье, возбуждение охватывает мускулатуру желудочков со скоростью 0,75–4,0 м/с.
Таким образом, мышечная ткань сердца ведет себя как функциональный синцитий. Благодаря этой особенности сердце, в отличие от скелетной мышцы, подчиняется закону «все или ничего». Это означает, что на раздражение возрастающей силы, начиная от порогового, мышца сердца отвечает сразу возбуждением всех волокон, т. е. амплитуда сокращений одинакова. Если раздражитель подпороговый, то она совсем не реагирует. Однако если раздражать сердечную мышцу током возрастающей частоты, оставив его силу постоянной, то каждое увеличение частоты раздражителя вызовет возрастающее сокращение сердечной мышцы – феномен «treppe» – лестницы. Это явление можно объяснить попаданием каждого последующего импульса в фазу повышенной возбудимости и накоплением ионов Са2+ в области миофибрилл, что и дает усиление ответной реакции.
Сокращение сердца, как и у скелетных мышц, запускается ПД. Однако если у скелетной мышцы ПД составляет всего несколько миллисекунд и предшествует сокращению, то у сердечной ПД и фазы сокращения перекрывают друг друга. ПД заканчивается только после начала фазы расслабления. Это одна из особенностей электромеханического сопряжения сердечной мышцы. Другая особенность состоит в том, что существует взаимосвязь между внутриклеточным депо Са
med.wikireading.ru
29.Физиологические свойства нервной ткани. Понятие о возбудимости, проводимости и лабильности.
Основными свойствами нервной ткани являются возбудимость, проводимость и лабильность, которые в свою очередь связаны с одним из самых общих свойств всего живого — раздражимостью.
Изменения в окружающей среде или организме называют раздражителями, процесс действия раздражителя — раздражением, а ответные изменения в деятельности клеток и целого организма — биологическими реакциями.
Основные физиологические свойства нервной ткани, ее проводимость, возбудимость и лабильность характеризуют функциональное состояние нервной системы человека, определяют его психические процессы. Нарушение проводимости и возбудимости нервной ткани, например при общем наркозе, прекращает все психические процессы человека и приводит к полной потере сознания.
Возбудимость и возбуждение. Клетки нервной ткани в процессе эволюции приспособились к быстрой ответной реакции на действие раздражителя, поэтому нервную ткань называют возбудимой, а ее способность быстро реагировать на раздражение — возбудимостью.
Возбуждение нервной клетки связано с изменением обмена веществ и сопровождается появлением электрических потенциалов — электрических, или нервных, импульсов.
Проводимость . Проводимость — способность живой ткани проводить возбуждение. Проводимость нервной ткани связана с распространением по ней процессов возбуждения. Возникнув в одной клетке, электрический (нервный) импульс легко переходит на соседние клетки и может передаваться в любой участок нервной системы.
Проводимость нервной ткани связана с тем, что возникший в месте возбуждения потенциал действия в свою очередь вызывает изменения ионных концентраций в соседнем участке. Возникнув на новом участке, потенциал действия вновь вызывает изменение концентрации ионов в соседнем участке и, соответственно, новый потенциал действия и т. д. Таким способом волна возбуждения распространяется вдоль всей ткани или отдельной нервной клетки.
Лабильность . Способность возбудимой ткани отвечать максимальным числом потенциалов действия в ответ на определенную частоту раздражений Н. Е. Введенский назвал лабильностью или функциональной подвижностью. Иначе говоря, лабильность — свойство, характеризующее способность возбудимой ткани воспроизводить максимальное количество потенциалов действия в единицу времени. Оказалось, что нервная ткань обладает наибольшей лабильностью, у мышечной она значительно ниже, самая низкая лабильность у синапсов.
Лабильность ткани в значительной степени зависит от функционального состояния этой ткани. Патологические процессы и утомление приводят к снижению лабильности нервной ткани, а систематические специальные тренировки — к ее повышению.
30. Основные процессы в центральной нервной системе, их координация и возрастные особенности.
В основе деятельности нервной системы лежат два процесса: возбуждение и торможение нейронов.
Возбуждение в ЦНС. Основное свойство нервной системы имеет ряд особенностей в ЦНС по сравнению с возбуждением в нервном волокне. В связи с особенностями строения синапсов в ЦНС возможно только одностороннее проведение возбуждения — от окончания аксона, где освобождается медиатор, к постсинаптической мембране. В синапсах ЦНС отмечается замедленное проведение возбуждения. Известно, что возбуждение по нервным волокнам проводится быстро. В синапсах скорость проведения возбуждения примерно в 200 раз ниже скорости проведения возбуждения в нервном волокне.
В деятельности всех отделов нервной системы играет важную роль и процесс торможения, результатом которого является ослабление или подавление возбуждения.
Явление торможения в ЦНС было открыто И. М. Сеченовым.
Торможение участвует в осуществлении любого рефлекторного акта.
Взаимодействие процессов возбуждения и торможения обеспечивает всю сложную деятельность нервной системы и согласованную деятельность всех органов человеческого тела. На воздействия из внешней и внутренней среды организм реагирует как единое целое. Объединение деятельности различных систем организма в единое целое (интеграция) и согласование, взаимодействие, ведущее к приспособлению организма к различным условиям среды (координация), связаны с деятельностью ЦНС.
Координация нервных процессов. (Иррадиация, индукция, доминанта нервных процессов)
Любая реакция организма представляет собой результат деятельности нервной системы и зависит от функционального состояния многих нервных центров и составляющих их нейронов. Такое согласованное взаимодействие нейронов и нервных процессов называют координацией рефлекторных процессов.
Координация нервных процессов, без которой были бы невозможны согласованная деятельность всех органов детского организма и его адекватные реакции на воздействия внешней среды, основывается на следующих особенностях, или принципах.
Конвергенция нервных процессов. В связи с широкой межнейронной связью нервные импульсы к одному нейрону могут приходить из различных участков нервной системы. Например, на один и тот же нейрон могут конвергировать импульсы от слуховых, зрительных и кожных рецепторов.
Иррадиация нервных процессов. Возбуждение или торможение, возникнув в одном нервном центре, могут распространяться на другие нервные центры. Это явление называют иррадиацией.
Индукция нервных процессов. В каждом нейроне или их скоплениях (нервные центры) один нервный процесс легко переходит в свою противоположность. Это явление называют индукцией. Если возбуждение сменяется на торможение, говорят об отрицательной индукции. Если вслед за торможением наступает возбуждение, говорят о положительной индукции.
Концентрация нервных процессов. Явление концентрации противоположно иррадиации. При этом процессы возбуждения или торможения концентрируются в каком-либо участке нервной системы.
Возрастные особенности координации нервных процессов
Деятельность целостного организма всегда связана со сложной координацией безусловно-рефлекторной и условно-рефлекторной реакций и их двигательных и вегетативных компонентов. Особое значение имеет координация вегетативных функций, выражающаяся в согласованных изменениях дыхания, работы сердца и всей сердечно-сосудистой системы, деятельности желез внутренней секреции и т. д. Вся совокупность этих изменений связана с энергетическим обеспечением рефлекторных реакций ребенка и необходима для достижения полезного организму результата в кратчайший срок и с наименьшей энергетической издержкой.
Ребенок рождается с далеко несовершенной координацией рефлекторных реакций. Ответная реакция у новорожденного всегда связана с обилием ненужных движений и широкими неэкономичными вегетативными сдвигами.
Дети в сравнении со взрослыми имеют более высокую возбудимость нервной ткани, меньшую специализацию нервных центров, более распространенные явления конвергенции и более выраженные явления индукции нервных процессов.
Доминантный очаг у ребенка возникает быстрее и легче, чем у взрослого, с чем в значительной степени связана неустойчивость внимания детей. Новые раздражители легко вызывают и новую доминанту в мозге ребенка.
В процессе развития все недостатки координации рефлекторных процессов у детей и подростков сглаживаются. Своего совершенства координационные процессы достигают только к 18—20 годам
31. Нервные центры, их физиологические особенности.
Нервный центр— это совокупность нейронов, расположенных на различных уровнях центральной нервной системы и регулирующих сложный рефлекторный процесс.
Нервные центры состоят из множества нейронов, связанных между собой еще большим множеством синаптических связей. Это обилие синапсов определяют основные свойства нервных центров: односторонность проведения возбуждения, замедление проведения возбуждения, суммацию возбуждений, усвоение и трансформацию ритма возбуждений, следовые процессы и легкую утомляемость.
Односторонность проведения возбуждения в нервных центрах связана с тем, что в синапсах нервные импульсы проходят только в одном направлении — от синаптического окончания аксона одного нейрона через синаптическую щель на клеточное тело и дендриты других нейронов.
Замедление движения нервных импульсов связано с тем, что «электрический, способ передачи нервных импульсов в синапсах сменяется химическим, или медиаторным, скорость которого в тысячу раз меньше.
Чем больше синапсов на пути движения нервных импульсов, тем больше проходит времени от начала раздражения до начала ответной реакции. Это время называют временем реакции или латентным временем рефлекса.
У детей время центральной задержки больше, оно увеличивается также при различных воздействиях на организм человека.
Суммация возбуждений была открыта И. М. Сеченовым в 1863 г. В настоящее время различают пространственную и временную суммацию нервных импульсов.
Первая наблюдается при одновременном поступлении к одному нейрону нескольких импульсов, каждый из которых в отдельности является подпороговым раздражителем и не вызывает возбуждение нейрона. В сумме же нервные импульсы достигают необходимой силы и вызывают появление потенциала действия.
Временная суммация возникает при поступлении к постсинаптической мембране нейрона серии импульсов, в отдельности не вызывающих возбуждение нейрона. Сумма этих импульсов достигает пороговой величины раздражения и вызывает возникновение потенциала действия.
Усвоение и трансформация ритма возбуждений в нервных центрах были изучены А. А. Ухтомским. Сущность усвоения ритма возбуждений заключается в способности нейронов «настраиваться» на ритм поступающих раздражений, что имеет большое значение для оптимизации взаимодействия различных нервных центров при организации поведенческих актов человека. С другой стороны, нейроны способны трансформировать (изменять) поступающие к ним ритмические раздражения в свой собственный ритм.
После прекращения действия раздражителя активность нейронов, составляющих нервные центры, не прекращается. Время этого последействия, или следовых процессов, сильно варьирует у различных нейронов и в зависимости от характера раздражителей. Предполагают, что явление последействия имеет важное значение в понимании механизмов памяти.
Быстрая утомляемость — также связана в значительной степени с деятельностью синапсов. Существуют данные, что длительные раздражения приводят к постепенному истощению в синапсах запасов медиаторов, к снижению чувствительности к ним постсинаптической мембраны. В результате рефлекторные ответные реакции начинают ослабевать и в конечном итоге полностью прекращаются.
studfiles.net
Общая физиология возбудимых тканей

1. Мембраны в основном состоят из липидов и белков, количество которых неодинаково у разных типов клеток. Сложность изучения молекулярных механизмов функционирования клеточных мембран обусловлена тем, что при выделении и очистке клеточных мембран нарушается их нормальное функционирование. В настоящее время можно говорить о нескольких видах моделей клеточной мембраны, среди которых наибольшее распространение получила жидкостно-мозаичная модель.Согласно этой модели, мембрана представлена бислоем фосфолипидных молекул, ориентированных таким образом, что гидрофобные концы молекул находятся внутри бислоя, а гидрофильные направлены в водную фазу (рис. 2.1). Такая структура идеально подходит для образования раздела двух фаз: вне- и внутриклеточной. В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения. Здесь описана только общая схема строения клеточной мембраны и для других типов клеточных мембран возможны значительные различия. Электрические характеристики мембран. Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость. Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов. Кроме того, емкостные свойства клеточных мембран являются одной из причин, определяющих временные характеристики электрических процессов, протекающихщих на клеточных мембранах. Проводимость (g) — величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов. Через фосфолипидный бислой могут диффундировать различные вещества, причем степень проницаемости (Р), т. е. способность клеточной мембраны пропускать эти вещества, зависит от разности концентраций диффундирующего вещества по обе стороны мембраны, его растворимости в липидах и свойств клеточной мембраны. Скорость диффузии для заряженных ионов в условиях постоянного поля в мембране определяется подвижностью ионов, толщиной мембраны, распределением ионов в мембране. Для неэлектролитов проницаемость мембраны не влияет на ее проводимость, поскольку неэлектролиты не несут зарядов, т. е. не могут переносить электрический ток. Проводимость мембраны является мерой ее ионной проницаемости. Увеличение проводимости свидетельствует об увеличении количества ионов, проходящих через мембрану.
ФУНКЦИИ:
1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
5.На ее поверхности протекает большинство биохимических реакций.
6.Мембрана координирует и регулируют физические процессы в клетке.
7.мембрана играет важную роль в образовании межклеточных контактов.
По способу активации выделяют:
потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно осуществляется конформацией белковой молекулы при изменении потенциала мембраны). Примером может служить потенциал-зависимый натриевый канал, определяющий деполяризацию клетки при генерации потенциала действия.
механочувствительные ионные каналы (открываются при воздействии на мембрану клетки механического стимула, например, при активации механорецепторов кожи).
лиганд-активируемые ионные каналы. По способу активации они подразделены на две группы (экстраклеточные и внутриклеточные) в зависимости от того, с какой стороны мембраны воздействует лиганд. Если стимул (например, ацетилхолин) при осуществлении синаптической передачи возбуждения в нервно-мышечном синапсе действует на рецептор (в данном примере холинорецептор, представляющий собой одну из нескольких белковых субъединиц ионного канала), расположенный на внешней поверхности мембраны мышечной клетки, откроется ионный канал, проницаемый для катионов. Если лиганд-активируемые каналы зависят от вторичных посредников в клетке, их переход в открытое состояние осуществляется при изменении концентрации определенных ионов в цитоплазме. Примером может служить кальций-активируемый калиевый канал, активирующийся при увеличении концентрации ионов кальция в клетке. Такие каналы принимают участие в реполяризации мембраны при завершении потенциала действия.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. В состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация) (рис. 2.4). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула.
Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+, причем существуют разновидности каналов для этих ионов (см. табл. 2.1).
Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя.
Особый интерес представляют кальциевые каналы.
Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли «мессенджера», или вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током.
Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца.
Электровозбудиые: раздражитель (электрический ток) -> сдвиг мембранного потенциала(до критического потенциала) -> активация потенциалуправляемых ионных каналов -> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
Хемовозбудимые: раздражитель( химическое вещество) -> химическое связывание раздражителя и рецептора хемоуправляемого ионного канала -> изменение конформации лигандрецепторного комплекса и открытие рецептруправляемых( хемоуправляемых) ионных каналов-> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
Механовозбудимые: раздражитель (механический стресс)-> изменение натяжения мембраны -> открытие механоуправляемых ионных каналов -> -> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
2. Все клетки имеют свой электрический заряд, который формируется в результате неодинаковой проницаемости мембраны для различных ионов. Клетки возбудимых тканей (нервная, мышечная, железистая) отличаются тем, что они под действием раздражителя меняют проницаемость своей мембраны для ионов, в результате чего ионы очень быстро транспортируются согласно электрохимическому градиенту. Это и есть процесс возбуждения. Его основой является потенциал покоя. Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах от -30 до -90 мВ. Внутренняя сторона мембраны в покое заряжена отрицательно, а наружная – положительно из-за неодинаковых концентраций катионов и анионов внутри и вне клетки.
основную роль в создании отрицательного заряда внутри клетки играют ионы K+ и высокомолекулярные внутриклеточные анионы, главным образом они представлены белковыми молекулами с отрицательно заряженными аминокислотами (глутамат, аспартат) и органическими фосфатами. Эти анионы, как правило, не могут транспортироваться через мембрану, создавая постоянный отрицательный внутриклеточный заряд. Во всех точках клетки отрицательный заряд практически одинаков. Заряд внутри клетки является отрицательным как абсолютно (в цитоплазме анионов больше, чем катионов), так и относительно наружной поверхности клеточной мембраны. Абсолютная разность невелика, однако этого достаточно для создания электрического градиента. Обеспечивающим формирование потенциала покоя (ПП), является K+. В покоящейся клетке устанавливается динамическое равновесие между числом входящих и выходящих ионов K+. Это равновесие устанавливается тогда, когда электрический градиент уравновесит концентрационный. Согласно концентрационному градиенту, создаваемому ионными насосами, K+ стремится выйти из клетки, однако отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому (электрический градиент). В случае равновесия на клеточной мембране устанавливается равновесный калиевый потенциал. В покое вход Na+ в клетку низкий (намного ниже, чем K+), но он уменьшает мембранный потенциал. Влияние Cl- противоположно, так как это анион. Отрицательный внутриклеточный заряд не позволяет большому количеству Cl- проникнуть в клетку, поэтому Cl- это в основном внеклеточный анион. Как внутри клетки, так и вне ее Na+ и Cl- нейтрализуют друг друга, вследствие чего их совместное поступление в клетку не оказывает существенного влияния на величину потенциала покоя.
Наружная и внутренняя стороны мембраны несут на себе собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные составляющие мембранных молекул – гликолипидов, фосфолипидов, гликопротеинов. Ca2+, как внеклеточный катион, взаимодействует с наружными фиксированными отрицательными зарядами, а также с отрицательными карбоксильными группами интерстиция, нейтрализуя их, что приводит к увеличению и стабилизации потенциала покоя.
Для создания и поддержания электрохимических градиентов необходима постоянная работа ионных насосов. Ионный насос – это транспортная система, обеспечивающая перенос иона вопреки электрохимическому градиенту, с непосредственными затратами энергии. Градиенты Na+ и K+ поддерживаются с помощью Na/K – насоса. Сопряженность транспорта Na+ и K+ примерно в 2 раза уменьшает энергозатраты. В целом же траты энергии на активный транспорт огромны: лишь Na/K – насос потребляет около 1/3 всей энергии, расходуемой организмом в покое. 1АТФ обеспечивает один цикл работы – перенос 3 Na+ из клетки, и 2 K+ в клетку. Асимметричный перенос ионов способствует заодно формированию и электрического градиента (примерно 5 – 10мВ).
Активный транспорт- с затратой энергии АТФ, против концентрационного и электрического градиента(насосы)
-первичный(включает перенос отдельных ионов вопреки концентрационному и электрическому градиентам с помощью специальных ионных насосов, микровезикулярный транспорт( эндоцитоз, экзоцитоз, трансцитоз)и фильтрацию.
-вторичный(он осуществляется за счет энергии, запасенной ранее, которая создается в виде электрического и концентрационного градиентов, т. е. тоже в результате расхода энергии, поэтому называть его пассивным необоснованно, хотя частицы движутся согласно законам диффузии.)
Пассивный транспорт- без затраты энергии по концентрационному и электрическому градиенту.
-простая диффузия(ионные каналы)
-облегченная диффузия(белки-переносчики)
-осмос
В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно должна выравниваться. Имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Им являются ионные насосы, в частности Na/K- насос.
Ионный насос — обладающая АТФ-азной активностью молекула интегрального белка, обеспечивающая перенос ионов через мембрану с непосредственной затратой энергии вопреки концентрационному и электрическому градиентам.выведение Na+ сопряжено с транспортом К+, что мжно продемонстрировать при адалении К+ из наружного раствора. Если К+ вне клетки нет, работа насоса блокируется, перенос Na+ из клетки в этом случае падает,составляя примерно 30% от нормально уровня. Накопление Na+ в клетке стимулирует работу насоса, уменьшение Na+ в клетке снижает его активность, поскольку снижается вероятность контакта ионов с соответствующим переносчиком. В результате сопряженного транспорта Na+ K+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ- обеспечивает один цикл работы Na/K насоса: перенос трех ионов Na+ за пределы клетки и двух ионов К+ внутрь клетки. Такой ассиметричный перенос ионов поддерживает избыток положительно заряженных частиц вне клетки и отрицательных зарядов внутри клетки,что позволяет считать Na/K насос структурной электрогенной,дополнительно увеличивающей потенциал покоя примерно на несколько милливольт.
Блокаторы — это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином — адреноблокаторы; гистамина — гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
Блокаторы — это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином — адреноблокаторы; гистамина — гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
3.
Два основных класса рецепторов — это метаботропные рецепторыиионотропные рецепторы.
Ионотропные рецепторы представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом. Возникающие при этом ионные токи вызывают изменения трансмембранной разности потенциалов и, вследствие этого, возбудимости клетки, а так же меняют внутриклеточные концентрации ионов, что может вторично приводитъ к активации систем внутриклеточных посредников. Одним из наиболее полно изученных ионотропных рецепторов являетсян-холинорецептор.
Метаботропные рецепторы связаны с системами внутриклеточных посредников. Изменения их конформации при связывании с лигандом приводит к запуску каскада биохимических реакций, и, в конечном счете, измемению функционального состояния клетки.
Рецепторы, связанные с гетеротримерными G-белками(например, рецепторвазопрессина).
Рецепторы, обладающие внутренней тирозинкиназной активностью (например, рецепторинсулина).
Внутриклеточные рецепторы-факторы транскрипции (например, рецепторы глюкокортикоидов).
Рецепторы, связанные с G-белками, представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Сайт связывания с лигандом находится на внеклеточных петлях, домен связывания с G-белком — вблизи C-конца в цитоплазме.
Активация рецептора приводит к тому, что его α-субъединицадиссоциирует от βγ-субъединичного комплекса и таким образом активируется. После этого она либо активирует, либо наоборот инактивируетфермент, продуцирующий вторичные посредники.
Рецепторы с тирозинкиназной активностью фосфорилируютпоследующие внутриклеточные белки, часто тоже являющиеся протеинкиназами, и таким образом передают сигнал внутрь клетки. По структуре это — трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, субъединицы которых связаныдисульфидными мостиками. Внутриклеточные рецепторы после связывания с гормоном переходят в активное состояние, транспортируются в ядро клетки, там связываются с ДНК и либо индуцируют, либо супрессируют экспрессию некоторого гена или группы генов.
Имеется несколько видов G-белков (GS, Gi ,Gq ,GO), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (GS), так и ингибировать (Gi) образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.
4 вопрос Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны; 2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил: 1) силы диффузии; 2) силы электростатического взаимодействия. Значение электрохимического равновесия: 1) поддержание ионной асимметрии; 2) поддержание величины мембранного потенциала на постоянном уровне. В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим. Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Любая живая клетка поддерживает на мембране определенной величины потенциал. Его величина колеблется в значительных пределах, у нервных и мышечных клеток его величина составляет обычно 60 — 90 мВ, у других тканей не превышает 10 мВ. Различные внешние воздействия, способные изменяющие ионную проницаемость мембраны (то есть повлиять на воротный механизм канала), вызывают изменения величины ПП. Его уменьшение называется деполяризацией, а увеличение — гиперполяризацией мембраны
5 вопрос Возбужде́ние в физиологии — ответ ткани на раздражение, проявляющийся помимо неспецифических реакций (генерация потенциала действия, метаболические изменения) в выполнении специфической для этой ткани функции; возбудимыми являются нервная (проведение возбуждения), мышечная (сокращение) и железистая (секреция) ткани. Возбудимость — свойство клеток отвечать на раздражение возбуждением. Реакция клеток, тканей на раздражитель определяется законами раздражения 1. Закон «все или ничего»: При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца. 2. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. 3. Закон силы-длительности. Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы длительности. По этой кривой можно определить ряд параметров возбудимости. а) Порог раздражения — это минимальная сила раздражителя, при которой возникает возбуждение. б) Реобаза — это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. в) Полезное время — минимальное время действия раздражителя силой в одну реобазу за которое возникает возбуждение. г) Хронаксия — это минимальное время действия раздражителя силой в две реобазы, необходимое для возникнов4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя , возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией. Физиологическая лабильность (подвижность) — это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем Выше ее лабильность. Определение лабильности предложено Н.Е.Введенским. Наибольшая, лабильность у нервов, наименьшая у сердечной мышцы. ения возбуждения. Действие постоянного тока на возбудимые ткани. •возбудимость повышается, а под положительным — анодом снижается. Это называется законом действия постоянного Тока. Изменение возбудимости ткани (например: нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода — катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном, Под положительным — анодом, он возрастает. Возникает физический катэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризации, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами. Критический уровень деполяризации (Ек)- это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия. Ек — пороговая величин
6 вопрос Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки. Этому способствуют две причины: открываются потенциалзависимые Nа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс). Освобождение натриевых каналов от Са2+. Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут). Для характеристики фазы деполяризации вводится понятие реверсии — это та разность потенциалов, на которую потенциал действия превышает потенциал покоя. Р (реверсия)- это то количество мВ на которое произошла перезарядка мембраны. Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости. Фаза реполяризации характеризуется: снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация). Натрий накапливается на наружной поверхности клеточной мембраны; возрастает проницаемость мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране; изменение активности Nа+-К+ насоса. Реполяризация — это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы. Гиперполяризация – при частом возбуждении канал не успевает захлопнуться, калий выходит из клетки, мембрана становится более электроотрицателной. Так как в формировании потенциала покоя мембраны участвуют многие ионы, равновесие клетки может нарушаться посредством изменений проводимости различных ионов. Так, например, при дополнительном выходящем токе ионов К+ или при входящем токе ионов Сl может увеличиваться потенциал покоя мембраны, это означает что она гиперполяризуется. Гиперполяризация мембраны — противоположность возбуждения, т.е. определенные химические процессы на постсинаптической мембране могут вызывать торможение нейрона.
7. Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь клетки. Нормальное поляризованное состояние немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией,
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Изменения мембранного потенциала,следующие за пиком потэнциала действия,называют СЛЕДОВЫМИ ПОТЕНЦИАЛАМИ.
Различают 2 вида следовых потенциалов-следовую деполяризацию и следовую гиперполяризацию.Амплитуда следовых потенциалов обычно не превышает нескольких милливольт,а длительность их у различных волокон составляет от нескольких миллисекунд до десятков и сотен секунд.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Возникновению потенциала действия предшествует в точке раздражения мышцы или нерва активные под пороговые изменения мембранного потенциала. Они проявляются в форме локального (местного) ответа.
8. Для локального ответа характерны:
1) зависимость от силы раздражения
2) нарастание постепенно величины ответа.
3) нераспространение по нервному волокну.
Первые признаки локального ответа обнаруживаются при действии стимулов составляющих 50-70% пороговой величины. Локальный ответ как и потенциал действия обусловлен повышением натриевой проницаемости. Однако это повышение было недостаточно, чтобы вызвать потенциал действия.
Потенциал действия возникает когда деполяризация мембраны достигнет критического уровня. Но локальный ответ важен. Он подготавливает ткани к последующим воздействиям.
Сравнительная характеристика локального потенциала и ПД
Свойство | Локальный потенциал | Потенциал действия |
Распространение | На 1 – 2 мм с затуханием (декрементом) | Без декремента на большие расстояния по всей длине нервного волокна |
Зависимость от величины стимула | Возрастает с увеличением силы раздражителя, т. е. подчиняется закону «силы» | Не зависит (подчиняется закону «все или ничего») |
Явление суммации | Суммируется – возрастает при частых повторных подпороговых раздражениях | Не суммируется |
Амплитуда | 10 – 40 мВ | 80 – 130 мВ |
Возбудимость ткани при возникновении потенцала | Увеличивается | Уменьшается вплоть до абсолютной рефрактерности |
Повышение возбудимости клетки во время локального потенциала объясняется тем, что мембрана оказывается частично деполяризованной. Если КУД остается на постоянном уровне, то для его достижения требуется гораздо меньший раздражитель. Амплитуда ПД не зависит от силы раздражителя, потому что он возникает вследствие регенеративных процессов.
При стимуляции рецептора внешним воздействием, к которому чувствителен рецептор , происходит деполяризация сомы , которая исчезает с прекращением стимуляции. Деполяризация называется рецепторным потенциалом (или генераторным потенциалом ). Длительность его соответствует длительности стимула, а его амплитуда возрастает с увеличением интенсивности стимуляции, таким образом, он является отражением стимула, а не ответом по типу «все или ничего», как потенциал действия . Рецепторный потенциал обусловлен повышением Na+ — проводимости мембраны дендритов , в результате чего вход ионов натрия создает деполяризующий рецепторный потенциал, который электротонически распространяется к соме . Эта первичная трансформация стимула в рецепторный потенциал называется преобразованием, а рецептор, таким образом, является преобразователем, датчиком.
Пороговая Сила тока увеличивается при уменьшении крутизны его нарастания,а при некоторой минимальной крутизне ответы на раздражение исчезают.Это я вление принято обозначать термином « аккомодация »
В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей,развивающиеся во время медленно нарастающей деполяризации мембраны.Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных неравных волокон скорость аккомодации,как правило,значительно выше,чем у чувствительных волокон.
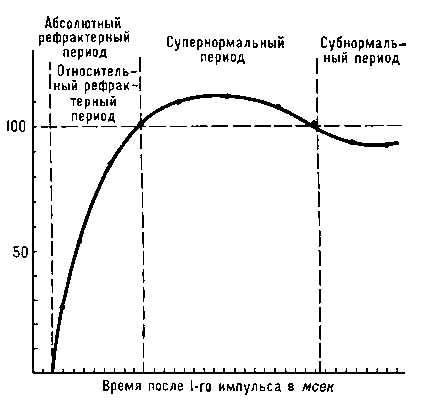
Кратковременное снижение возбудимости (См. Возбудимость) нервной и мышечной тканей непосредственно вслед за потенциалом действия (См. Потенциал действия). Р. обнаруживается при стимуляции нервов и мышц парными электрическими импульсами. Если сила 1-го импульса достаточна для возникновения потенциала действия, ответ на 2-й будет зависеть от длительности паузы между импульсами. При очень коротком интервале ответ на 2-й импульс отсутствует, как бы ни увеличивалась интенсивность стимуляции (абсолютный Рефрактерный период). Удлинение интервала приводит к тому, что 2-й импульс начинает вызывать ответ, но меньший по амплитуде, чем 1-й импульс (в опытах на нервных стволах, состоящих из большого числа параллельных нервных проводников), либо для возникновения ответа на 2-й импульс необходимо увеличить силу раздражающего тока (в опытах на одиночных нервных волокнах). Период сниженной возбудимости нервной или мышечной клетки называется относительным рефракторным периодом. За ним следует супернормальный период, или фаза экзальтации (См. Экзальтация), т. е. фаза повышенной возбудимости, сменяющаяся периодом несколько сниженной возбудимости — субнормальным периодом. В основе наблюдаемых колебаний возбудимости лежит изменение проницаемости биологических мембран (См. Проницаемость биологических мембран), сопровождающее возникновение потенциала действия (см. Биоэлектрические потенциалы). Длительность каждого периода определяется кинетикой этих процессов в данной ткани. В быстропроводящих нервных волокнах Р. длится не более 3—5 мсек, в мышце сердца период изменений возбудимости занимает до 500 мсек. Р. — один из факторов, ограничивающих частоту воспроизведения биологических сигналов, их суммацию и скорость проведения. При изменении температуры или действии некоторых лекарственных веществ длительность рефракторных периодов может меняться, чем пользуются для управления возбудимостью ткани, например сердечной мышцы: удлинение относительного рефрактерного периода приводит к снижению частоты сердечных сокращений и устранению нарушений ритма работы сердца.
studfiles.net
Фундаментальные процессы в цнс
Горобец
Нейрон рождается и умирает молодым. При ушибах мозга погибают нервные клетки.
Существует 2 фундаментальных базовых процесса нервной клетки:
— процессы возбуждения
— процессы торможения
Закономерность распространения возбуждения нервной клетки
Процессы возбуждения
Возбуждение – это форма ответной реакции на действие раздражителей. Раздражители могут быть как и с внешней так и с внутренней стороны. Из внешней – специфические раздражители(световая волна, химический раздражитель). Возбуждения сопровождаются интеграцией волнового, распространяющегося потенциала действия. Внутренним содержанием действия является изменение интенсивность процессов жизнедеятельности в нервных клетках.
Общие свойства возбудимых тканей
Возбудимость – способность возбуждаться.
Проводимость – способность проводить возбуждение, т.е формировать потенциал действия.
Сократимость – способность развивать силу или напряжение при возбуждении.
Лабильность – (функциональная подвижность) – это способность к ритмической активности.
Способность выделять секрет.
Для нервной ткани процесс возбуждения это основная форма проявления жизнедеятельности. Возбуждение передаётся либо через вставочные нейроны либо прямую передачу. Проведение импульсов по меилинизированному нерву происходит скачкообразно. Сальтоторная передача отличается тем что у неё очень большая скорость. В мякотных волокнах скорость возбужения от 3 до 18 метров в сек. А в безмякотных от 1 до 3х метров в сек.
Закономерности распространения возбуждения по нервному волокну:
Непрерывность
Двусторонняя проводимость
Изолированное проведение возбуждения
Существует 3 закона функционирования нервной клетки:
1)Закон силы – чтобы возникло возбуждение, раздражитель должен быть достаточно сильным.(Пороговый или выше порогового). Пороговая сила 20 мили вольт.
2) Закон времени – это зависимость пороговой силы раздражителя от времени его действия.
3) Закон градиента – что бы раздражитель вызвал возбуждение, он должен нарастать достаточно быстро.
Передача возбуждение с одного волокна на другое происходит благодаря химич веществам – медиаторам, они синтезируются внутриклеточно и выпрыскиваются в синаптическую щель и формируют возбуждающий постсинаптический потанциал(В П С П). Он формируется:
1)Ацетилхолин – медиатор
Он расширяет проходимость для ионов натрия.
Генерируется ВПСП
2) В перерывах между импульсами ацетилхолин разрушается под действием ацетилхалинной эстеразы разрушается до халина и уксусной кислоты.
3) Гаммааминомаслянная кислота является медиатором торможения. Взаимодействует с постсинаптической мембраной. Повышается диффузия кальция, блокируется поток натрия, при этом калий выходит из клетки в кальций поступает в клетку.
Торможение – это самостоятельная форма электрической активности клеточной мембраны(один из актов жизнедеятельности).
Ковалёв
Смирнов,Яковлев «Физиология ЦНС»
Шульговский «Нейрофизиология»
Шульговский «Физиология ЦНС»
Недоспасов «Физиология ЦНС»
Физиология возбудимых тканей
К возбудимым тканям относят :
— нервную
— мышечную
— железистую ткани
В эту группу их объединяет в зависимости от следующих свойств:
1)Возбудимость – способность возбудимых тканей реагировать на действие раздражителя развитием специфического процесса возбуждения. Возбуждение – изменение физико–химических свойств возбудимой ткани под действием раздражителя. В роле раздражителя выступает любой агент, способный вызвать видимо ответную реакцию со стороны возбудимых тканей.
2)Проводимость – это способность возбудимых тканей проводить раздражение.
3)Рефрактэрность – это временное снижение возбудимости, возникающая в данном образовании сразу после прохождения волны возбуждения( В первые моменты её наступления). Для каждого вида возбудимой ткани характерны так же специфические свойства. Для нервной ткани – это генерация или воспроизведение нервного импульса для мышечной ткани – сократимость или способность менять длину при возникновении раздражителя. Для железистой ткани характерно выделение особых веществ – секретов. Благодаря усилиям итальянским учёным в 18 веке Луиджи Гальдбани и Александра Вольта было открыто явление биологического электричества. Гольдбани изучая электричесво подвешивал препарат лапки лягушки к балконной решётке. При соприкосновении нерва препарата происходило сокращение мышц лапки, в результате чего Гольдбани предположил наличие биологического электричества. В качестве возражения Вольта выдвинул предположение, что сокращение конечности происходит благодаря разности потенциалов между металлами, составляющими решётку ,в результате Гольдбани ставит второй опыт: на повреждённый участок мышцы, в область ахилого сухожилия стеклянным крючком набрасывается нерв препарата, в результате чего происходит сокращение лапки лягушки. Однако объяснить природу биоэлектрических явлений стало возможно только после использования микроэлектродной техники, когда внутрь клетки вводили электроды, не повреждающие структуры мембраны. Второй электрод помещали на поверхность мембраны, в результате регистрировали разность потенциалов, между наружной и внутренней поверхностями мембраны. Выяснили, что эта разность потенциалов связана с физиологическими функциями ,свойствами и строением мембраны нервных клеток. Оказалось, что мембрана нервных клеток, согласно современным представлениям, состоит из двойного слоя липидов, жироподобных веществ, который состоит из гидрофильных головок, обращённых наружу и внутрь мембраны ,они притягивают к себе воду и гидрофобных хвостов, образующих промежуточный слой между головками. Гидрофобный слой не пропускает водорастворимые вещества. В билипидный слой вмонтированы белковые молекулы в форме глобул, которые могут пронизывать билипидный слой насквозь или прободать его частично снаружи или изнутри. С билипидным слоем ассоциирован слой гликокаликса, который обеспечивает узнавание веществ, поступающих на мембрану. Рисунок 1 .Мембрана обладает определёнными физиологическими свойствами:
1) Полупроницаемость – способность биомембраны проводить только определённые вещества, обладающие определённым зарядом
2) /Избирательная проницаемость мембраны означает что через мембрану вещества может проходить только в определённых концентрациях в определённом направлении и по специальным каналам, который предназначены только для данного вещества
Функции биологической мембраны :
Рецепторная функция – заключается в узнавании веществ, поступающих на мембрану нервных клеток. Связь мембраны с определённым веществом осуществляется по принципу ключ к замку, это означает, что данное вещество вступает в химическое взаимодействие со специальным рецептором мембраны клетки. Химическая структура мембранного рецептора специфична по отношению к определённому веществу.
Транспортная функция мембраны – способность мембраны проводить определённые вещества. Различают активные и пассивные виды транспорта
Активный транспорт осуществляется против градиента концентрации и против электрохимического градиента, с участием специальных молекул переносчиков и затратами энергии.
Пассивный транспорт осуществляется по градиенту концентрации ,т.е из области большей концентрации в область меньшей концентрации с минимальными затратами энергии и по электрохимическому градиенту ,т.е от большего заряда, в сторону меньшего заряда.
Разграничительная функция — мембрана нервной клетки отделяет нервную клетку от внеклеточной среды, создавая тем самым внутриклеточную среду.
Барьерная функция – мембрана клетки защищает клетку от механических повреждений
studfiles.net
1 Возбуждение — Стр 2
Одиночный цикл возбуждения.
К возбудимым тканям относятся только те, клетки которых генерируют потенциал действия (ПД). Это мышечные и нервные клетки. Нередко к возбудимым тканям необоснованно относят и «железистую ткань», хотя железистой ткани нет, а имеются различные железы и железистый эпителий как вид тканей. В процессе активной деятельности железы в ней действительно регистрируются биоэлектрические явления, поскольку железа как орган состоит из различных клеток: соединительной ткани, эпителиальной, мышечной. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма.
Невозбудимыми тканями являются эпителиальная и соединительная (собственно соединительная, ретикулярная, жировая, хрящевая, костная и гемотопоэтические ткани в совокупности с кровью), клетки этих тканей хотя и способны изменять свой мембранный потенциал, но не генерируют ПД при действии наних раздражителя.
Основными физиологическими свойствами возбудимых тканей являются: возбудимость, проводимость, рефрактерность, лабильность.Специфическим свойством мышечной ткани являетсясократимость.
Возбудимость — это свойство некоторых тканей генерировать потенциал действия (ПД) в ответ на раздражение. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. Раздражители вызывающие гиперполяризацию мембран будут приводить к процессу обратному возбуждению – торможению.
Возбудимость может быть охарактеризована кривой потенциала действия, в которой выделяют несколько фаз (рис.1 А). Отметим, что единой терминологии в классификации этих фаз нет, поэтому будем использовать наиболее часто употребляемые названия.
Рис. 1. Изменение мембранного потенциала (А) и возбудимости клетки (Б) в разные фазы потенциала действия.
МВ – фаза местного возбуждения;
Д – фаза деполяризации;
РБ– фаза быстрой реполяризации;
РМ – фаза медленной реполяризации;
Г – фаза следовой гиперполяризации;
Н – период нормальной возбудимости;
РА – период абсолютной рефрактерности;
РО– период относительной рефрактерности;
Н+– период первичной экзальтации;
Н++– период экзальтации;
Н–– период субнормальной возбудимости.
Вначале, под действием раздражителя, развивается местное возбуждение(фаза начальной деполяризации) — процесс медленной деполяризации мембраны от мембранного потенциала до критического уровня деполяризации (КУД). Если этот уровень не будет достигнут – ПД не формируется, а развивается только локальный ответ.
Разность между мембранным потенциалом покоя и критическим уровнем деполяризации называют пороговым потенциалом, его величина определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
Время фазы начальной деполяризации очень короткое, на кривой ПД она регистрируется только при большой развертке, и чаще всего является составной частью общей фазы деполяризации. Эта фаза развивается при достижении КУД, за счет открытия всех потенциалчувствительных Na+ — каналов и лавинообразного входа ионов Na+ в клетку по градиенту концентрации (входящий натриевый ток). В результате, мембранный потенциал очень быстро уменьшается до 0, и даже приобретает положительное значение. Графически – это восходящая часть кривой потенциала действия. В результате инактивации Na+ — каналов и прекращения поступления Na+ в клетку, рост кривой ПД прекращается и начинается ее снижение. Явление изменения знака мембранного потенциала называютреверсиейзаряда мембраны.
По мнению некоторых исследователей, фаза деполяризации заканчивается уже тогда, когда мембранный потенциал становится равным нулю, и весь период, когда величина мембранного потенциала превышает величину 0 мВ, следует считать отдельной фазой реверсии, т.к. ионные токи, определяющие развитие этой части ПД, имеют характерные особенности.
Период времени, в течение которого мембранный потенциал имеет положительное значение, называется овершут.
Нисходящая част кривой ПД – фаза реполяризации. Она определяется выходящим калиевым током. Калий выходит через постоянно открытые каналы утечки, ток через которые резко возрастает из-за изменения электрического градиента вызванного нехваткой снаружи ионов Na+ и через потенциалочувствительные, управляемыеК+- каналы, которые активируются на пике ПД.
Различают быструю и медленную реполяризацию. В начале фазы, когда активны оба типа каналов, реполяризация происходит быстро, к концу фазы, ворота потенциалочувствительных К+- каналов закрываются, интенсивность калиевого тока снижается и реполяризация замедляется. Она прекращается тогда, когда положительный заряд снаружи мембраны вырастет настолько, что окончательно затруднит выход калия из клетки.
Фазу медленной реполяризации называют иногда отрицательным следовым потенциалом, что не совсем верно, так как эта фаза не является потенциалом по определению и не является следовым процессом по механизму.
Фаза следовой гиперполяризации(следовой положительный потенциал) – увеличение мембранного потенциала выше величины потенциала покоя, которое наблюдается у нейронов. Развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+АТФ-азы.
Механизм наблюдаемой иногда следовой деполяризации(следовой отрицательный потенциал) до конца не ясен.
Изменение возбудимости клетки при развитии возбуждения. Рефрактерность.
Возбудимость в различные фазы развития одного цикла возбуждения, вообще является переменной величиной. В ходе развития одного цикла возбуждения возбудимость изменяется в сторону, как повышения, так и понижения. Повышение возбудимости называется экзальтацией, понижение –рефрактерностью.
В изменении возбудимости от момента нанесения раздражения до завершения одиночного цикла возбуждения отмечается несколько периодов (фаз). (Рис.1. Б)
В период развития местного возбуждения наблюдается некоторое повышение возбудимости, которое получило название первичной экзальтации. Каждое нанесенное в это время дополнительное раздражение, по силе даже ниже порогового, ускоряет развитие местного потенциала. Это связано с тем, что пороговый потенциал уменьшается, и открытие воротного механизма Na+ -каналов облегчается.
Как только местное возбуждение достигает критической величины и переходит в потенциал действия (фаза деполяризации), возбудимость начинает быстро снижаться и в точке пика потенциала практически становится равной нулю. Это связано с полной инактивацией Na+ -каналов на пике ПД.
Время, в течение которого происходит это снижение возбудимости называется абсолютной рефрактерной фазой(периодом), а само снижение возбудимости — абсолютной рефрактерностью. Раздражение любой сверхпороговой силы, нанесенное в этот период, практически не может повлиять на развитие текущего возбуждения (потенциала действия).
В фазе реполяризации возбудимость мембраны последовательно восстанавливается, до исходного уровня, за счет постепенного восстановления активности инактивированных Na+ -каналов. Пока активны не все каналы – то этот период называетсяотносительной рефрактерной фазой, а состояние, в котором находится живой объект — относительной рефрактерностью. Эта фаза продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Раздражение, нанесенное в этот период, может вызвать усиление возбуждения только в том случае, если по силе оно будет больше величины порогового потенциала Длительность относительной рефрактерной фазы может быть значительно больше, чем абсолютной.
Вслед за периодом относительной рефрактерности наступает фаза экзальтации (повышенной возбудимости). Это связано с тем что мембранный потенциал снижается до величины КУД, при которой восстанавливается активность большей части Na+ каналов, а разница между величиной мембранного потенциала и КУД – пороговый потенциал – минимальна. В этой фазе может возникнуть повторная волна возбуждения даже на раздражения, которые значительно ниже порогового потенциала. Фаза экзальтации длится до тех пор, пока не восстановится исходная величина мембранного потенциала – потенциал покоя, при этом восстанавливается исходная величина возбудимости.
В фазы следовой гипер- и деполяризации возбудимость меняется незначительно и связана с колебаниями порогового потенциала.
Биологический смысл фазового изменения возбудимости в ходе развития одиночной волны возбуждения заключается в следующем.
Начальная фаза повышения возбудимостиобеспечивает условие, при котором каждый дополнительный раздражитель ускоряет процесс подготовки (местное возбуждение) к специфической (для данной ткани) приспособительной реакции.
Состояние абсолютной рефрактерностипозволяет данной ткани «без помех» осуществлять текущую приспособительную реакцию. Если бы в этих условиях возбудимость была нормальной, то дополнительное раздражение, вызвав дополнительное возбуждение, могло бы исказить эту реакцию, превратив ее в избыточную или недостаточную для данных условий.
Абсолютная рефрактерность защищает ткань от чрезмерных энергетических трат в процессе осуществления текущей приспособительной реакции. Сходную роль играет и относительная рефрактерность, с той разницей, что в данном случае живое образование в состоянии реагировать на раздражения, требующие срочного ответа. Именно поэтому для большинства тканей и органов, работающих непрерывно и не имеющих длительных периодов физиологического покоя (например, сердце), характерна более длительная по сравнению со скелетной мускулатурой рефрактерность.
Кроме того, рефрактерность — один из факторов, определяющих максимальный (предельный) ритм импульсации клетки, что лежит в основе например кодирования и декодирования сигнала структурами нервной системы, регуляции восприятия, сокращения, обеспечении одностороннего проведения возбуждения по нервам и др.
Состояние зкзальтациисоздает условия готовности ткани к ответу на повторное раздражение не только прежней силы, но и более слабой.
Лабильность,или функциональная подвижность, одно из физиологических свойств живых тканей. Это свойство описано в 1892 г. Н. Е. Введенским, который установил, что скорость протекания процесса возбуждения в тканях различна. Каждая возбудимая ткань способна на раздражение отвечать только определенным количеством волн возбуждения. Так, нервное волокно способно воспроизводить до 1000 импульсов в секунду, поперечно-полосатая мышца только 200 250 имп/с.
Мерой лабильности, по Н. Е. Введенскому, является то наибольшее количество волн возбуждения, которое возбудимая ткань может воспроизводить в 1 с в точном соответствии с ритмом наносимых раздражений без явлений трансформации (переделки) ритма, т.е. не уменьшая и не увеличивая его.
Лабильность величина подвижная и может изменяться в достаточно широких пределах. В частности, лабильность широко варьирует в процессе ритмического раздражения. В одних случаях вследствие взаимодействия волн возбуждения лабильность может повыситься, в других понизиться. Повышение лабильности может привести к тому, что недоступные ранее ритмы деятельности станут доступными. На основании этого А. А. Ухтомский сформировал представление об «усвоении ритма», как способности ткани отвечать на раздражение более высоким или более низким ритмом возбуждения по сравнению с его исходным уровнем. Усвоение ритма зависит от текущих изменений обмена веществ в ткани во время ее деятельности
Явление усвоения ритма играет важную роль в процессах врабатывания и тренировки. Снижение лабильности, происходящее в процессе деятельности, приводит к иному результату, способность ткани к ритмической работе уменьшается. Лабильность может быть измерена косвенным путем по величине хронаксии(см. ниже) возбудимых тканей. Чем короче хронаксия, тем выше лабильность. Определение лабильности весьма важно в физиологии труда и спорта.
Проводимость — способность живой ткани проводить возбуждение, которое, возникая в рецепторе, распространяется по нервной системе и является для организма информацией, закодированной в нейроне в виде электрических или химических сигналов. Способностью к проведению возбуждения обладают практически все возбудимые ткани, но наиболее ярко она выражена в нервной ткани, для которой проводимость является одной из функций.
Подробно механизм и закономерности распространения возбуждения по мембранам возбудимых клеток рассмотрен в отдельном занятии.
Законы раздражения.
Процесс возбуждения начинается с действия на возбудимую клетку какого либо раздражителя.
Раздражитель-любое изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающееответную реакцию. По своей природе раздражители делятфизические (электрические, механические, температурные, световые)и химические.
В зависимости от степени чувствительности клеток к тому или иному раздражителю их подразделяют на адекватные и неадекватные. Адекватный раздражитель — это такой раздражитель, к которому клетка обладает наибольшей чувствительностью вследствие наличия специальных структур, воспринимающих этот раздражитель. Так, адекватным раздражителем для фоторецепторов сетчатки глаза, например, являются световые волны, адекватным раздражителем нейронов являются медиаторы и электрические импульсы.
Неадекватныераздражители в естественных условиях существования организма не воздействуют на возбудимые структуры. Однако, при достаточной силе и продолжительности действия, могут вызвать ответную реакцию со стороны возбудимых тканей, например, удар в глаз при достаточной силе может вызвать ощущение вспышки света.
В условиях физиологического эксперимента в качестве раздражителя чаше всего используют электрический ток. Электрический ток легко дозировать, и он является адекватным раздражителем для возбудимых тканей, так как их функциональная активность всегда сопровождается электрическими явлениями.
Определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани отражают законы раздражения. К законам раздражения относятся:
Закон силы.
Для возникновения возбуждения решающее значение имеет сила раздражителя. Возбуждение возникают только в том случае, если сила действующего раздражителя достигает минимальной, критической величины, которая характеризуется порогом возбуждения. По отношению к этой величине, по своей силе раздражители могут быть подпороговыми, пороговыми и надпороговыми.
Подпороговый раздражитель— это раздражитель такой силы, который не вызывает видимых изменений, но обусловливает возникновение физико-химических сдвигов в возбудимых тканях, например локального ответа. Однако степень этих сдвигов недостаточна для возникновения распространяющегося возбуждения.
Пороговый раздражитель— это раздражитель минимальной силы, который впервые вызывает минимальную измеримую ответную реакцию со стороны возбудимой ткани. Именно эту пороговую силу раздражителя называютпорогомраздражения или возбуждения. Порог раздражения и является мерой возбудимости ткани. Между порогом раздражения и возбудимостью существует обратная зависимость:чем выше порог раздражения, тем ниже возбудимость, чем ниже порог раздражения, тем возбудимость выше.При достижении раздражителем величины порога, возникновение потенциала действия становится неизбежным.
Следует отметить, что порог раздражения показатель достаточно изменчивый и значительно зависит от исходного функционального состояния возбудимой ткани и практически никак не зависит от характеристик самого раздражителя
Надпороговый раздражитель— это раздражитель, сила которого выше, чем сила порогового раздражителя.
Закон силы — характеризует взаимосвязь между силой раздражителя и электрическим ответом, он может быть применен для простых и сложных систем.
Простая возбудимая система– это одна возбудимая клетка, которая реагирует на раздражитель как единое целое. Исключением является сердечная мышца, которая вся реагирует как одна клетка.Закон силы для простых возбудимых систем — подпороговые раздражители не вызывают возбуждения, а пороговые и сверхпороговые раздражители вызывают сразу максимальное возбуждение (Рис. 2).
При подпороговых значениях раздражающего тока возбуждение (электротонический потенциал, локальный ответ) носит местный (не распространяется), градуальный (сила реакции пропорциональная силе действующего стимула) характер. При достижении порога возбуждения возникает ответ максимальной силы (ПД). Амплитуда ответа (амплитуда ПД) не изменяется при дальнейшем увеличении силы раздражителя.
Закон силы для простых возбудимых систем известен как закон «все ли ничего».
Сложная возбудимая система– система, состоящая из множества возбудимых элементов (мышца включает множество двигательных единиц, нерв – множество аксонов). Отдельные элементы (клетки) системы имеют неодинаковые пороги возбуждения.
Закон силы для сложных возбудимых систем — амплитуда ответа пропорциональна силе действующего раздражителя (при значениях силы раздражителя от порога возбуждения самого легковозбудимого элемента до порога возбуждения самого трудновозбудимого элемента) (рис. 3). Амплитуда ответа системы пропорциональна количеству вовлеченных в ответ возбудимых элементов. При возрастании силы раздражителя в реакцию вовлекается все большее число возбудимых элементов.
В случае сложных систем, от силы раздражителя будет зависеть не только электрический, но и физиологический (функциональный) ответ ткани, например сила сокращения. В этом случае закон силы будет звучать следующим образом: чем больше сила раздражителя, тем выше, до определенного предела, ответная реакция со стороны возбудимой ткани. Этот предел будет определяться функциональными возможностями ткани.
Ответ минимальной силы – едва заметное сокращение — возникнет при достижении раздражителем пороговой величины. При этом сократятся мышечные волокна имеющие наименьший порог возбуждения.
Ответная реакция на надпороговый раздражитель будет выше и по мере его увеличения некоторое время также возрастает за счет вовлечения в сокращение все новых мышечных волокон, которые имеют более высокие пороги возбуждения. По достижении определенной величины раздражителя, рост силы сокращения прекратится, значит, в сокращение вовлечены все мышечные волокна. Такую ответную реакцию называют максимальной, астепени силы раздражителя, находящиеся между пороговой и максимальной –субмаксимольными.
Амплитуда ответа а лЛ И Т У Д а ответа | | Амплитуда ответа | |


Рис. 2. Зависимость силы реакции простой Рис. 3. Зависимость силы реакции сложной
возбудимой системы от силы раздражителя. возбудимой системы от силы раздражителя. ПВ – порог возбуждения. ПВMIN– порог возбуждения самого
легковозбудимого элемента,
ПВMАХ– порог возбуждения самого
трудновозбудимого элемента.
Увеличение силы раздражителя выше максимального какое-то время не сказывается на величине ответной реакции. Такую силу раздражителя называют супермаксимальной. Но при достаточно большом увеличении силы раздражителя, сила ответной реакции начинает снижаться. Такую величину силы раздражителя называютпессимальной.
Пессимальный ответ и есть тот определенный предел, до которого может расти ответная реакция. Превышение этого предела при спортивных, интеллектуальных, эмоциональных и любых других нагрузках не имеет никакого физиологического смысла для получения результата.
Закон силы-времени (силы-длительности)
Эффективность раздражителя зависит не только от силы, но и от времени его действия. Длительность действия раздражителя, способна компенсировать недостаток силы раздражителя и при его недостатке привести, тем не менее, к возникновению распространяющегося потенциала действия, поэтому важно определять не только пороговую силу, но пороговую длительность раздражителя. Учение о хронаксии как пороговом времени необходимом для возникновения возбуждения было создано французским ученым Лапиком.
Связь между силой и временем действия раздражителя характеризует закон силы длительности — сила раздражителя, вызывающего процесс распространяющегося возбуждения, находится в обратной зависимости от длительности его действия, т.е., чем больше сила раздражителя, тем меньше времени он должен действовать для возникновения возбуждения. Зависимость между силой раздражителя и продолжительностью его воздействия, необходимого для возникновения минимальной ответной реакции живой структуры, очень хорошо можно проследить на так называемой кривой силы — времени (кривая Гоорвега — Вейса — Лапика) (Рис 4).
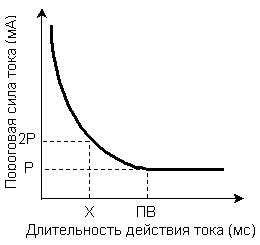
Рис.4.Зависимость пороговой силы раздражителя от времени его действия. Р – реобаза, ПВ – полезное время, Х – хронаксия.
Из кривой следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и как бы ни велика была сила раздражителя, при недостаточной длительности его воздействия ответной реакции не будет.
Минимальная сила раздражителя, способная, при неограниченном времени действия вызвать возбуждение, была названа Лапиком реобазой.Наименьшая длительность действия раздражителя силой в одну реобазу, достаточная для возникновения ответной реакции называется –полезным временем.
Но определение точки соответствующей на кривой величине полезного времени по техническим причинам затруднено, поэтому Лапиком было предложено измерять не полезное время, а условную величину – хронаксию.
Хронаксия – это наименьший промежуток времени, в течение которого ток, равный по силе удвоенной реобазе, вызывает в ткани возбуждение. Она и есть показатель пороговой длительности раздражения. Хронаксия измеряется в (тысячные доли секунды). По величине хронаксии можно судить о скорости возникновения возбуждения в ткани: чем меньше хронаксия, тем быстрее возникает возбуждение.
Определение хронаксии — хронаксиметрия —получило широкое распространение в физиологии спорта, в клинике, для определения функциональных возможностей и сохранности возбудимых тканей.
Закон крутизны нарастания силы раздражителя (закон аккомодации, закон Дюбуа-Реймона)
Для возникновения возбуждения имеет значение не только сила и время действия тока, но и скорость нарастания силы раздражителя в единицу времени от нулевой величины до величины порога. Степень крутизны, т.е. величина прироста силы раздражения в единицу времени, представляет собой градиент раздражения. Зависимость ответной реакции от градиента раздражения характеризует закон градиента (крутизны нарастания) раздражения — чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого образования (рис 5).
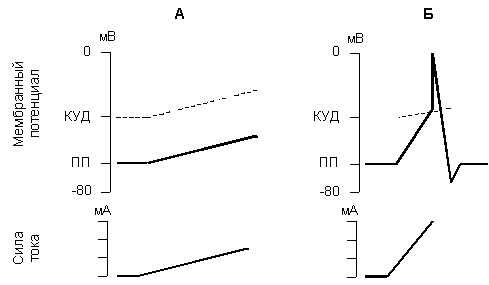
Рис. 5. Изменение мембранного потенциала и критического уровня деполяризации при медленном (А) и быстром (Б) нарастании силы раздражающего тока.
При действии медленно нарастающего раздражителя возбуждение возникает при его гораздо большей силе, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации.Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает.
Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого — окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще.
Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
studfiles.net
Лекции биопотенциалы восстановленный
5
Лекции № 1
Понятие о физиологии.
Физиология – наука о функциях и процессах, протекающих в организме или его составляющих системах, органах, клетках и механизмах их регуляции, обеспечивающих жизнедеятельность во взаимодействии с окружающей средой.
Функция – специфическая деятельность систем или органа.
Процесс – последовательная смена явлений или состояний в развитии какого-либо действия или совокупность последовательных действий, направленных на достижение определенного результата.
Система в физиологии – совокупность органов или тканей, связанных общей функцией.
Нормальная физиология – изучает жизнедеятельность организма в норме.
Норма – это пределы оптимального функционирования живой системы.
Физиология разделяется на несколько относительно самостоятельных, но связанных между собой областей. Выделяют:1)общую физиологию и частную; 2)сравнительную; 3)эволюционную; 4)прикладную, которая изучает закономерности изменения функций организма в связи с его специфической деятельностью. В этом плане различают: — авиационную, — возрастную, — клиническую, -космическую, -психофизиологию, -физиологию труда и спорта, -экологическую физиологию.
Методы физиологии.
1)экспериментальный
2)наблюдение
3)моделирование
Место физиологии в системе медицинского образования.
Физиология формирует понимание и дает знания об условиях, необходимых для обеспечения здоровья, методах оценки уровня здоровья, технологиях поддержания здоровья.
Тема: Возбудимые ткани. Кодирование информации в организме.
Понятие возбудимые ткани. Возбудимые ткани способны реагировать на раздражение генерацией потенциала действия. К возбудимым тканям относятся нервная ткань, мышечная и железистая.
Свойства возбудимых тканей:
Общие свойства: возбудимость, проводимость. Для мышечной — дополнительно сократимость, а для сердечной мышцы, некоторых гладкомышечных волокон и нервных клеток – дополнительно автоматия.
Возбудимость – способность отвечать на раздражение генерацией потенциала действия. Критерии возбудимости: 1.Порог раздражения – минимальная сила раздражения, способная вызвать ответную реакцию. 2.Полезное время — минимальная длительность действия раздражителя, необходимая для возникновения возбуждения. 3. Хронаксия – время, в течение которого раздражитель, силой в 2 порога, способен вызвать возбуждение. 4.Лабильность — способность ткани генерировать ответ синхронно с раздражением.
Проводимость – способность ткани проводить возбуждение от места возникновения к другим участкам или структурам.
Критерии проводимости – скорость распространения возбуждения .
Сократимость – способность ткани менять геометрию.
Критерии сократимости — изменении длины или тонуса мышцы.
Автоматия – способность ткани самопроизвольно возбуждаться без внешних раздражителей. Критерии автоматии – частота спонтанных возбуждений.
Кодирование информации в организме – это перевод информации в другую форму, понятную организму. Кодирование информации, поступающей в организм, начинается на уровне воспринимающих образований – рецепторов в виде возникающих рецепторных потенциалов. Частота и пространственно-временной рисунок рецепторных потенциалов зависит от параметров раздражителя: силы и длительности его действия. По мере прохождения сигнала через различные структуры ЦНС происходит его перекодирование. Расшифровка, т.е. декодирование информации, осуществляется в коре.
Принцип кодирования информации.
Кодирование осуществляется путем изменения биоэлектрической активности клеток возбудимых тканей.
Виды кодирования: — среднечастотное, — интервальное, — путем изменения количества импульсов в пачке.
Пачка импульсов – группа электрически ответов нервной клетки, возникающая в ответ на раздражение.
БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ВОЗБУДИМЫХ ТКАНЯХ
Биоэлектрические процессы развиваются на мембране клеток и представлены потенциалом покоя (ПП) и потенциалом действия (ПД).
Последовательность развития событий при кодировании информации можно представить в виде следующей схемы:
Информация → клетка → изменение ПП → локальный ответ →разновидности импульсной активности.
Теория, объясняющая существование биоэлектрических явлений в организме называется мембранно-ионной.
Роль мембраны.
1.Барьерная функция связана с наличием бислоя липидов, обеспечивающего избирательную проницаемость для жирорастворимых молекул и почти полную непроницаемость для ионов.
2.Транспртная функция связана с работой белковых каналов, по которым ионы перемещаются через мембрану.
Виды ионных каналов.
А.По способу открытия различают хемочувствительные и потенциалзависимые;
Б.По скорости открытия и закрытия – быстрые и медленные;
Каналы могут быть в открытом и закрытом состоянии. Открытие каналов приводит к пассивному транспорту ионов по градиенту концентрации и, как следствие, изменению заряда мембраны.
3.Рецепторная функция. Мембрана содержит рецепторы к химическим веществам. Взаимодействие данного вещества с рецептором открывает хемочувствительные ионные каналы и возникает ответ клетки на информацию, переданную гуморальным путем.
ПОТЕНЦИАЛ ПОКОЯ
И ЕГО ХАРАКТЕРИСТИКА ( величина, происхождение, колебания, механизм поддержания).
ПП — это разность потенциалов между наружной и внутренней сторонами мембраны. Внутри заряд отрицательный, снаружи – положительный.
Величина ПП: скелетные мышцы — -60 -90 мВ, нейрон — -50 — 80 мВ, кардиомиоцит — -85 -95 мВ железистая клетка — -30 мВ. Гладкомышечные клетки — -30 -20 мВ или -50 мВ (у клеток, не обладающих автоматией).
ПРОИСХОЖДЕНИЕ ПП.
1.Роль мембраны. В покое мембрана полупроницаема. Открыты каналы для К и практически все каналы для Na закрыты.
2.Роль ионов. В клетке существуют ионные градиенты. Внутри клетки преобладают ионы К (в 30 – 50 раз больше, чем снаружи). Na внутри находится в основном в связанном состоянии или в компартментах, т.е. иммобилизован. Снаружи клетки больше свободного Na ( в 10 раз больше, чем внутри) и в 30 раз больше Cl.
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПП.
Ионы К+ пассивно по градиенту концентраций выходят из клетки. Внутри остаются крупные анионы, которые не могут пройти через каналы для мелкого иона К и создают отрицательный заряд внутренней стороны мембраны. Снаружи заряд положительный. Но мембрана несколько проницаема для Na (идет внутрь и снижает разность потенциалов, создаваемую К).
Nа – К насос. Механизм поддержания мембранного потенциала и ионных градиентов.
Концентрационный градиент К и Na поддерживается ионным насосом. Так называют систему энергозависимого переноса ионов через мембрану против концентрационного градиента: с помощью переносчиков ионы транспортируются из растворов с меньшей в растворы с большей конценцентра -цией . Переносчиком для Na и К является Nа – К зависимая АТФ – аза. На 3 удаленных иона Na из клетки, в клетку возвращается 2 иона К. Такой перенос ионов сопровождается увеличением внутренней отрицательности, а насос называется электрогенным. Если же насос переносит эквивалентное количество ионов, то заряд мембраны при этом не меняется. Такой перенос ионов является электронейтральным.
Изменения ПП. Снижение внутренней отрицательности называется деполяризацией, повышение – гиперполяризацией.
Критический уровень мембранного потенциала (КУМП) – такое значение потенциала покоя, при достижении которого открываются потенциалзависимые каналы для натрия и возникает ПД.
Локальный ответ. Возникает в локальном участке в ответ на действие химических или электрических стимулов, составляющих 30 – 90% от порогововой силы раздражения. В итоге происходит открытие каналов для Na ( в некоторых образованиях для Ca).Возникает деполяризация мембраны, которая быстро сменяется реполяризацией, не достигнув КУМП.
Свойства локального ответа.
не распространяется
не зависит от силы раздражения
способен к суммации.
Возбудимость во время локального ответа повышена.
Локальный ответ играет важную роль в деполяризации мембраны до критического уровня, что является необходимым условием возникновения ПД.
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД), ЕГО ФАЗЫ, ИОННЫЙ МЕХАНИЗМ.
ПД – это быстрые колебания ПП под влиянием порогового или сверхпорогового раздражения.
Условия возникновения ПД. – пороговая сила раздражения
— достаточная длительность раздражения
— достаточная скорость нарастания раздражения.
Ионный механизм ПД .
Схема ПД. Ео – Ек = порогу раздражения. Восходящая фаза называется деполяризацией, называется деполяризацией, нисходящая – реполяризацией.
При достижении ПП значения Ек открываются быстрые потенциалзависимые каналы для Na, который лавиной устремляется внутрь клетки и снижает внутреннюю отрицательность сначала до 0, а затем возникает перезарядка мембраны – овершут. При перезарядке мембраны до +20 -30 мВ происходит инактивация натриевых каналов и начинается процесс реполяризации
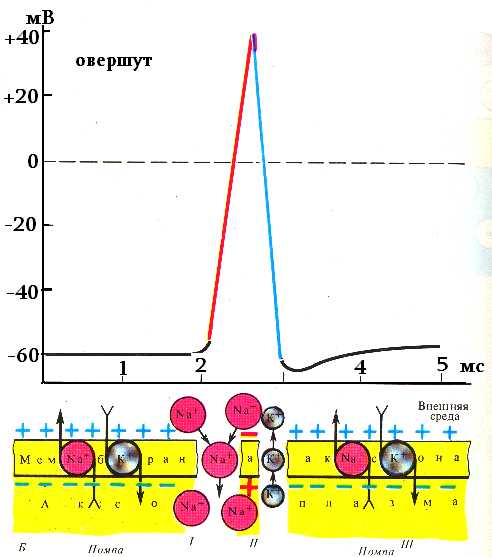
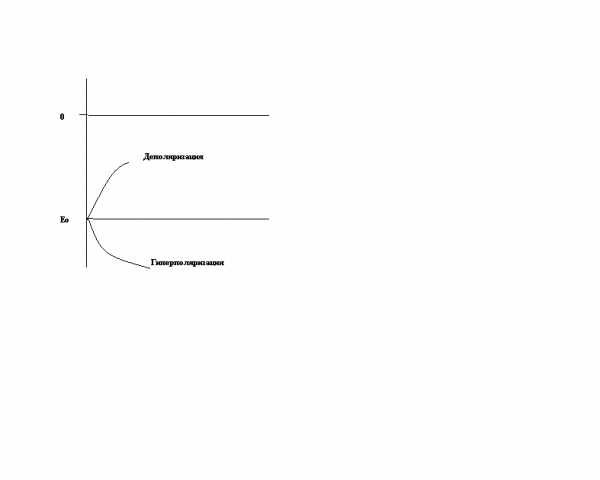
5
studfiles.net
Общая физиология возбудимых тканей
Биологические реакции. Живые организмы и все их клетки обладают раздражимостью, т.е. способностью отвечать на воздействия внешней среды или нарушения их состояния изменением своей структуры или функции, что неразрывно связано с количественными и качественными изменениями обмена веществ и энергии. Изменения структуры и функций организма и его клеток в ответ на различные воздействия называют биологическими реакциями, а сами воздействия, их вызывающие — раздражителями, или стимулами.
Понятие биологической реакции включает все виды ответной деятельности организма, его клеток и органов на различные воздействия. Реакции клеток проявляются в изменении их формы, структуры, их роста и процесса деления, в образовании в них различных химических соединений, преобразовании потенциальной энергии в кинетическую (электрическую, механическую, тепловую, световую), совершении той или иной работы (перемещении в пространстве, выделении тех или иных веществ, работе по концентрированию в клетке определенных электролитов и т.п). Еще более разнообразны реакции целостного организма, в особенности — сложные формы поведения. В процессе их осуществления меняется деятельность многих органов и бесчисленного множества клеток, ибо организм всегда реагирует на различные воздействия как единое целое, как единая сложная система.
Раздражители. Раздражителем живой клетки или организма как целого может оказаться любое изменение внешней среды или внутреннего состояния организма, если оно достаточно велико, возникло достаточно быстро, и продолжается достаточно долго.
Все бесконечное многообразие возможных раздражителей можно разделить на 3 группы: физические, физико-химические и химические. К числу физических раздражителей относятся температурные, механические (удар, укол, давление, перемещение, ускорение и т.п.), электрические, световые. Физико-химические раздражители представлены изменениями осмотического давления, активной реакции среды, электролитного состава, коллоидального состояния. К числу химических раздражителей относится множество веществ, имеющих различный состав и свойства, и способных изменить обмен веществ клеток (вещества пищи, лекарства, яды, гормоны, ферменты, метаболиты и т.п.).
Раздражителями клеток, вызывающими их деятельность, имеющими особо важное значение в жизненных процессах, являются нервные импульсы. Будучи естественными, т.е. возникающими в самом организме, электрохимическими раздражителями клеток, нервные импульсы, поступая по нервным волокнам от нервных окончаний в ЦНС или приходя от нее к периферическим органам, вызывают направленные изменения их состояния и деятельности.
Все раздражители по месту возникновения делят на внешние (экстеро-) и внутренние (интеро-) раздражители, а по физиологическому значению — на адекватные и неадекватные. Адекватными называют те раздражители, которые действуют на данную биологическую структуру в естественных условиях, к восприятию которых она специально приспособлена эволюцией и чувствительность к которым у нее обычно чрезвычайно велика (глаз — свет, ухо — звук и т.д.). Неадекватными называются те раздражители, для восприятия которых данная клетка или орган специально не приспособлен, но которые в определенных условиях могут вызвать изменения структуры или функции (мышца — может сократиться при ударе, быстром согревании, воздействии электротока, внезапном растяжении, действии кислоты и т.п.).
Возбудимость. Клетки нервной, мышечной и железистой тканей специально приспособлены к осуществлению быстрых реакций на раздражение (возбуждаться). Клетки этих тканей называют возбудимыми, а их способность отвечать на различные раздражения возбуждением — возбудимостью. Возбудимость — это свойство клеточной мембраны отвечать на действие раздражающего (возбуждающего) фактора изменением проницаемости и своего электрического состояния. Это явление и носит название возбуждение. Возбуждение представляет собою сложную биологическую реакцию, проявляющуюся в совокупности физических, физико химических и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны, (изменение ее мембранного потенциала, МП, и генерация распространяющегося потенциала действия, ПД). Возникнув в одной клетке или в одном ее участке, возбуждение распространяется на другие участки той же клетки или на другие клетки.
Ответ живой клетки на раздражение в форме ли возбуждения и связанной с ним электрической реакции, или в форме сокращения или секреции, происходит всегда после некоторого скрытого, или латентного периода. Так называют период времени между началом действия раздражителя и реакцией ткани на его действие. В течение латентного периода должны пройти изменения состояния ткани, необходимые для того, чтобы проявилась реакция. Латентный период возбудимых тканей короче, чем у невозбудимых, а латентный период электрической реакции ткани короче, чем мышечного сокращения и тем более секреторной реакции.
История открытия электрических явления в тканях.
В 1786 г. итальянский врач и физиолог Гальвани, развесив для просушки лягушачьи лапки на балконе заметил, что когда раскачиваемая ветром лапка соприкасается с металлической решеткой балкона, то возникает ее сокращение. Гальвани сделал вывод, что если между нервом и мышцей устанавливается замыкание посредством металлического проводника, и при этом мышца сокращается, то это есть доказательство проявления «животного электричества». Он считал, что нерв и мышца заряжены противоположно.
Однако, физик Вольта показал ошибочность вывода Гальвани путем проведения такого опыта: он подметил, что перила балкона были медные, а крючки, на которых висели лапки — железные. Попробовав приложить к лапке пинцет, одна ножка которого была сделана из меди, а другая из цинка или из железа, Вольта получил сокращение мышцы. Следовательно, заключил он, мышцы сокращаются не потому, что выделяется «животное электричество», а потому, что между двумя металлами, соприкасающимися с электролитом, течет ток, который и раздражает нервы лапки лягушки.
Не соглашаясь с Вольта, Гальвани поставил второй опыт. Он заключался в без металлическом сокращении мышцы. Сокращение достигалось путем накидывания нерва на отпрепарированную мышцу с помощью стеклянных инструментов. Однако оказалось, что сокращение удавалось получить только тогда, когда мышца была повреждена, а если мышца была отпрепарирована тщательно, без повреждения ее поверхности, то при таком опыте сокращения не возникало. Позднее немецкий физиолог Герман показал, что если к неповрежденной мышце приложить электроды гальванометра, то никакой разности потенциалов увидеть не удается. Но если на мышцу или нерв нанести повреждение, надрез, и погрузить в этот надрез один из электродов, то стрелка гальванометра отклоняется, что показывает, что между поврежденными и неповрежденными участками живой мышцы возникает электрический ток, причем поврежденный участок несет отрицательный заряд. Этот ток был назван током повреждения, или током покоя.
В 1837 г. Маттеучи показал, что ток покоя скелетной мышцы при ее сокращении уменьшается. Маттеучи проделал и еще один опыт. Он брал два нервно-мышечных препарата и нерв 2-го набрасывал на мышцу 1-го. При этом он раздражал нерв 1-го препарата, заставляя мышцу сокращаться. Оказалось, что и 2-я мышца при этом начинала сокращаться. Объяснить это влиянием на нерв тока покоя нельзя, так как сокращение второй мышцы происходило лишь при возбуждении первой. Еще демонстративнее этот опыт, если вместо первой мышцы взять работающее сердце лягушки. При набрасывании стеклянным крючком нерва нервно-мышечного препарата на сердце лягушки мышца лапки начинает сокращаться в ритме работающего сердца. Причина этого явления была обнаружена позже.
В 1850 г. знаменитый французский исследователь Дюбуа-Реймон, раздражая седалищный нерв лягушки, обнаружил, что вслед за раздражением по нерву пробегает волна электрического тока. В 1868 г. Герман показал, что причина этого в том, что возникающий при раздражении электрический ток достигает соседнего участка, возбуждает его, затем достигает следующего участка и путем таких контактов волна возбуждения бежит по нерву, как огонь по бикфордову шнуру.
Если раздражать одиночными ударами постоянного тока участок нерва, а от следующего участка отводить двумя электродами ток на гальванометр или на трубку катодного осциллографа, то вначале, в момент нанесения раздражения, никаких отклонений не регистрируется. так как под обоими отводящими электродами одинаковый потенциал. Через некоторое время, распространяясь. возбуждение достигает первого отводящего электрода и тогда гальванометр регистрирует разность потенциалов в виде отрицательного колебания — стрелка отклоняется влево (на осциллографе — вниз). Когда волна возбуждения — оказывается между электродами, стрелка возвращается в исходное положение. Затем волна возбуждения достигает второго электрода — стрелка отклоняется вправо (луч вверх). Когда волна возбуждения уходит дальше, и луч осциллографа и стрелка гальванометра возвращаются в исходное положение.
Из этих фактов можно сделать следующие выводы:
1. В покое разность потенциалов существует только между неповрежденным и поврежденным участками ткани (ток повреждения, или ток покоя).
2. При прохождении возбуждения по нерву в нем возникает ток действия.
3. Этот ток действия не остается на месте, а распространяется.
4. Ток действия представляет собой отрицательное колебание потенциала.
Более точное изучение механизмов электрических изменений в тканях в покое и при возбуждении стало возможным с прогрессом электроизмерительной и микроэлектродной техники. Переходим теперь к рассмотрению современных данных об электрических процессах в тканях.
Потенциал покоя. Оказалось, что между наружной поверхностью клетки и ее протоплазмой в состоянии покоя существует разность потенциалов порядка 60-90 мв., причем поверхность клетки заряжена электроположительно по отношению к протоплазме. Эта разность потенциала называется мембранным потенциалом, или потенциалом покоя. Точное его измерение возможно только с помощью внутриклеточных микроэлектродов.
Согласно мембранно-ионной теории Ходжкина-Хаксли, биоэлектрические потенциалы обусловлены неодинаковой концентрацией ионов K+,Na+,Cl- внутри и вне клетки, и различной проницаемостью для них поверхностной мембраны.
На основании данных электронной микроскопии, химического анализа и электрофизиологических исследований предполагают, что мембрана состоит из двойного слоя молекул фосфолипидов, покрытого изнутри слоем белковых молекул, а снаружи — слоем молекул белка и мукополисахаридов. Допускают, что в клеточной мембране имеются тончайшие каналы (поры) диаметром в несколько ангстрем. Через эти каналы молекулы воды и других веществ, а также ионы, имеющие соответствующий размеру пор диаметр, входят в клетку и покидают ее. На структурных элементах мембраны фиксируются различные заряженные группы, что придает стенкам каналов тот или иной заряд. Так, наличие в мембране нервных волокон диссоциированных фосфатных и карбоксильных групп является причиной того, что она (мембрана) значительно менее проницаема для анионов, чем для катионов.
Проницаемость мембраны для различных катионов также неодинакова и закономерно изменяется при различных функциональных состояниях ткани. В покое мембрана нервных волокон примерно в 25 раз более проницаема для ионов К, чем для ионов Na, а при возбуждении натриевая проницаемость примерно в 20 раз превышает калиевую.
Кроме проницаемости, большое значение для возникновения мембранного потенциала имеет градиент концентрации ионов по обе стороны мембраны. Показано, что цитоплазма нервных и мышечных клеток содержит в 30-59 раз больше ионов К+ (500 мэкв/л против 10 мэкв/л), но в 8-10 раз меньше ионов Na+ (35 мэкв/л против 350 мэкв/л) и в 50 раз меньше ионов Cl-, чем внеклеточная жидкость (см. табл.). Величина потенциала покоя нервных волокон и клеток определяется соотношением положительно заряженных ионов К+, диффундирующих в единицу времени из клетки наружу по градиенту концентрации, и положительно заряженных ионов Na+, диффундирующих по градиенту концентрации в обратном направлении. Так, в модельных опытах на аксоне кальмара при том градиенте концентрации К+, который имеет место в нервном волокне, величина тока К+ составляет -120 мв. Если же смоделировать в таком опыте только натриевый градиент, то величина тока Na+ составляет +30 мв. Реально измеряемый мембранный потенциал нерва равен сумме этих двух противоположно направленных токов, т.е. -90мв.
Несмотря на то, что скорость диффузии ионов Na+ и К+ через мембрану в покое мала, разность их концентрации вне клетки и внутри нее должна была бы в конечном итоге полностью выровняться, если бы в клетке не существовало специального механизма, который обеспечивает активное выделение («выкачивание») из протоплазмы проникающих в нее ионов Na+ и введение («нагнетание») ионов К+. Этот механизм получил образное название натрий калиевого насоса.
Для того, чтобы сохранялась ионная асимметрия, Na-К-насос должен совершать определенную работу против градиента концентрации ионов. Непосредственным источником энергии для работы насоса является расщепление АТФ, которое происходит под влиянием АТФ-азы, локализованной в мембране и активируемой ионами Na+ и К+ (т.н. Na-К-зависимая АТФ-аза). Торможение активности этого фермента приводит к нарушению работы насоса. В результате протоплазма обогащается Na+ и теряет К+. Прямым следствием этого является снижение или даже полное исчезновение МП (потенциала покоя, или мембранного потенциала).
Деполяризация мембраны возникает потому, что в силу градиента концентрации К+ выходит наружу, но из-за того, что ионы CL-, которые не в состоянии пройти через мембрану, электростатически удерживают положительные ионы, в пограничном слое создается избыток К+, и между наружной и внутренней поверхностями мембраны, заряженными соответственно положительно и отрицательно, возникает разность потенциалов величиной около -90 мв. Мембрана в покое постоянно деполяризована, так как в результате работы Na-K-насоса поддерживается нужный для этого градиент концентрации ионов.
Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя (например, толчка электрического тока), в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электро-отрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. «перезарядка мембраны». Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
В ПД принято различать его пик (т.н. спайк — spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
Кроме пика, в ПД различают два следовых потенциала — следовую деполяризацию (следовой отрицательный потенциал) и следовую гиперполяризацию (следовой положительный потенциал. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения.
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+ превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+ Поэтому поток ионов Na+ в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+ достигает величины +150 мв. Одновременно несколько уменьшается выход К+ из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
Повышение проницаемости мембраны для ионов Na+ продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
Наличие специальных Na- и К- каналов и сложного механизма запирания и открытия ворот изучено биофизиками достаточно хорошо. Показано, что существуют избирательные механизмы, регулирующие те или иные каналы. Например, яд тетродотоксин блокирует только Na-поры, а тетраэтиламмоний — только К-поры. Показано, что у некоторых клеток возникновение возбуждения связано в изменением проницаемости мембраны для Са++, в других — для Mg+. Исследования механизмов изменения проницаемости мембран продолжаются.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса.
Таким образом, в живой клетке существуют два различных типа движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным транспортом. Он ответственен за возникновение МП и ПД и ведет в конечном итоге к выравниванию концентраций ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в «выкачивании» ионов Na+ из протоплазмы и «нагнетании» ионов К+ внутрь клетки. Этот тип ионного транспорта возможет лишь при условии затраты энергии — это активный транспорт. Он является результатом работы специальных ферментных систем (т.н. насосов), и благодаря ему восстанавливается исходная разность концентраций, необходимая для поддержания МП.
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше — надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 — 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк)
Причины этого явления нам станут ясны позже, после выяснения некоторых механизмов действия постоянного электротока на возбудимые ткани.
В лабораторных условиях и при проведении некоторых клинических исследований для раздражения нервов и мышц применяют электрические стимулы, которые легко дозировать как по амплитуде и длительности, так и по форме, имитируя естественные нервные импульсы. Механизм раздражающего действия тока на ткань в принципе одинаков при всех видах стимулов, максимально приближается к механизму действия самих нервных импульсов, однако в наиболее отчетливой форме эти механизмы выявляются при использовании постоянного тока.
dendrit.ru