
Потенциал действия и мембранный потенциал покоя – CGI script error
Мембранный потенциал покоя. Мембранный потенциал действия. Проведение возбуждения
Мембранный потенциал покоя
text_fields
text_fields
arrow_upward
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной. Принимая потенциал наружного раствора за нуль, МПП записывают со знаком «минус». Величина МПП зависит от вида ткани и варьирует от -9 до -100 мв. Следовательно, в состоянии покоя клеточная мембрана поляризована. Уменьшение величины МПП называют деполяризацией, увеличение — гиперполяризацией, восстановление исходного значения МПП — реполяризацией мембраны.
Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К
Возникающая разность потенциалов препятствует выходу К+ из клетки и при некотором ее значении наступает равновесие между выходом К+ по концентрационному градиенту и входом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется
где Ек— равновесный потенциал для К+; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; п — валентность К+ (+1), [Кн+] — [К+вн] — наружная и внутренняя концентрации К+—
Если перейти от натуральных логарифмов к десятичным и подставить в уравнение числовые значения констант, то уравнение примет вид:
В спинальных нейронах (табл. 1.1) Ек = -90 мв. Величина МПП, измеренная с помощью микроэлектродов заметно ниже — 70 мв.
Таблица 1.1. Концентрация некоторых ионов внутри и снаружи спинальных мотонейронов млекопитающих
| Ион | Концентрация | (ммоль/л Н2О) | Разновесный потенциал (мв) |
внутри клетки | снаружи клетки | ||
| Na+ | 15,0 | 150,0 | +60 |
| К+ | 150,0 | 5,5 | -90 |
| Сl— | 9,0 | 125,0 | -70 |
Мембранный потенциал покоя = -70 мв | |||
Если мембранный потенциал клетки имеет калиевую природу, то, в соответствии с уравнением Нернста, его величина должна линейно снижаться с уменьшением концентрационного градиента этих ионов, например, при повышении концентрации К+ во внеклеточной жидкости. Однако линейная зависимость величины МПП (Мембранный потенциал покоя) от градиента концентрации К+ существует только при концентрации К
Величины натриевого и хлорного равновесных потенциалов для спинальных нейронов (табл. 1.1) равны соответственно +60 и -70 мв. Значение Е
Вклад каждого из равновесных потенциалов в величину МПП определяется соотношением между проницаемостью клеточной мембраны для каждого из этих ионов. Расчет величины мембранного потенциала производится с помощью уравнения Гольдмана:
Еm— мембранный потенциал; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; РK , PNaи РCl — константы проницаемости мембраны для К
Подставляя в это уравнение полученные в экспериментальных исследованиях концентрации ионов и величину МПП, можно показать, что для гигантского аксона кальмара должно быть следующее соотношение констант проницаемости Рк : PNa : РС1 = I : 0,04 : 0,45. Очевидно, что, поскольку мембрана проницаема для ионов натрия (Р
При повышении концентрации ионов калия в наружном растворе выше 15 мМ МПП увеличивается и соотношение констант проницаемости меняется в сторону более значительного превышения» Рк над PNa и РС1. Рк: PNa : РС1 = 1 : 0.025 : 0,4. В таких условиях МПП определяется почти исключительно градиентом ионов калия, поэтому экспериментальная и теоретическая зависимости величины МПП от логарифма отношения концентраций калия снаружи и внутри клетки начинают совпадать.
Таким образом, наличие стационарной разности потенциалов между цитоплазмой и наружной средой в покоящейся клетке обусловлено существующими концентрационными градиентами для К+, Na+ и Сl и различной проницаемостью мембраны для этих ионов. Основную роль в генерации МПП играет диффузия ионов калия из клетки в наружный наствор. Наряду с этим, МПП определяется также натриевым и хлорным равновесными потенциалами и вклад каждого из них определяется отношениями между проницаемостями плазматической мембраны клетки для данных ионов.
Все факторы, перечисленные выше, составляют так называемую ионную компоненту МПП (Мембранный потенциал покоя). Поскольку, ни калиевый, ни натриевый равновесные потенциалы не равны МПП. клетка должна поглощать Na+ и терять К+. Постоянство концентраций этих ионов в клетке поддерживается за счет работы Na+ К+-АТФазы.
Однако роль этого ионного насоса не ограничивается поддержанием градиентов натрия и калия. Известно, что натриевый насос электрогенен и при его функционировании возникает чистый поток положительных зарядов из клетки во внеклеточную жидкость, обуславливающий увеличение электроотрицательности цитоплазмы по отношению к среде. Электрогенность натриевого насоса была выявлена в опытах на гигантских нейронах моллюска. Электрофорети-ческая инъекция ионов Na
Из сказанного следует, что МПП может быть разделен на две компоненты — «ионную» и «метаболическую». Первая компонента зависит от концентрационных градиентов ионов и мембранных проницаемостей для них. Вторая, «метаболическая», обусловлена активным транспортом натрия и калия и оказывает двоякое влияние на
Мембранный потенциал действия
text_fields
text_fields
arrow_upward
Если на нерв или мышцу нанести раздражение выше порога возбуждения, то МПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной. Это кратковременное изменение МПП, происходящее при возбуждении клетки, которое на экране осциллографа имеет форму одиночного пика, называется
МПД в нервной и мышечной тканях возникает при снижении абсолютной величины МПП (деполяризации мембраны) до некоторого критического значения, называемого порогом генерации МПД. В гигантских нервных волокнах кальмара МПД равен — 60 мВ. При деполяризации мембраны до -45 мВ (порог генерации МПД) возникает МПД (рис. 1.15).
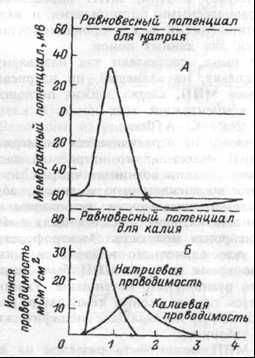 Рис. 1.15 Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
Рис. 1.15 Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).Во время возникновения МПД в аксоне кальмара сопротивление мембраны уменьшается в 25 раз, с 1000 до 40 Ом.см2, тогда как электрическая емкость не изменяется. Указанное снижение сопротивления мембраны обусловлено увеличением ионной проницаемости мембраны при возбуждении.
По своей амплитуде (100-120 мВ) МПД (Мембранный потенциал действия) на 20-50 мВ превышает величину МПП (Мембранный потенциал покоя). Другими словами, внутренняя сторона мембраны на короткое время становится заряженной положительно по отношению к наружной, — «овершут» или реверсия заряда.
Из уравнения Гольдмана следует, что лишь увеличение проницаемости мембраны для ионов натрия может привести к таким изменениям мебранного потенциала. Значение Ек всегда меньше, чем величина МПП, поэтому повышение проницаемости мембраны для К+ будет увеличивать абсолютное значение МПП. Натриевый равновесный потенциал имеет знак «плюс», поэтому резкое увеличение проницаемости мембраны для этих катионов приводит к перезарядке мембраны.
Во время МПД увеличивается проницаемость мембраны для ионов натрия. Расчеты показали, что если в состоянии покоя соотношение констант проницаемости мембраны для К+, Na+ и СГ равно 1 :0,04:0,45, то при МПД — Рк : PNa : Р = 1 : 20 : 0,45. Следовательно, в состоянии возбуждения мембрана нервного волокна не просто утрачивает свою избирательную ионную проницаемость, а, напротив, из избирательно проницаемой в покое для ионов калия она становится избирательно проницаемой для ионов натрия. Увеличение натриевой проницаемости мембраны связано с открыванием потенциал-зависимых натриевых каналов.
Механизм, который обеспечивает открывание и закрывание ионных каналов, получил название ворот канала. Принято различать активационные (m) и инактивационные (h) ворота. Ионный канал может находиться в трех основных состояниях: закрытом (m-ворота закрыты; h-открыты), открытом (m- и h-ворота открыты) и инактивированном (m-ворота открыты, h- ворота закрыты) (рис 1.16).
Рис. 1.16 Схема положения активационных (m) и инактивационных (h) ворот натриевых каналов, соответствующие закрытому (покой, А), открытому (активация, Б) и инактивированному (В) состояниям.
Деполяризация мембраны, вызываемая раздражающим стимулом, например, электрическим током, открывает m-ворота натриевых каналов (переход из состояния А в Б) и обеспечивает появление направленного внутрь потока положительных зарядов — ионов натрия. Это ведет к дальнейшей деполяризации мембраны, что, в свою очередь, увеличивает число открытых натриевых каналов и, следовательно, повышает натриевую проницаемость мембраны. Возникает «регенеративная» деполяризация мембраны, в результате которой потенциал внутренней стороны мембраны стремится достичь величины натриевого равновесного потенциала.
Причиной прекращения роста МПД (Мембранный потенциал действия) и реполяризации мембраны клетки является:
а) Увеличение деполяризации мембраны, т.е. когда Ем -» ENa, в результате чего снижается электрохимический градиент для ионов натрия, равный Ем —> ENa. Другими словами, уменьшается сила, «толкающая» натрий внутрь клетки;
б) Деполяризация мембраны порождает процесс инактивации натриевых каналов (закрывание h-ворот; состояние В канала), который тормозит рост натриевой проницаемости мембраны и ведет к ее снижению;
в) Деполяризация мембраны увеличивает ее проницаемость для ионов калия. Выходящий калиевый ток стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала.
Снижение электрохимического потенциала для ионов натрия и инактивация натриевых каналов уменьшает величину входящего натриевого тока. В определенный момент времени величина входящего тока натрия сравнивается с возросшим выходящим током — рост МПД прекращается. Когда суммарный выходящий ток превышает входящий, начинается реполяризация мембраны, которая также имеет регенеративный характер. Начавшаяся реполяризация ведет к закрыванию активационных ворот (m), что уменьшает натриевую проницаемость мембраны, ускоряет реполяризацию, а последняя увеличивает число закрытых каналов и т.д.
Фаза реполяризации МПД в некоторых клетках (например, в кардиомиоцитах и ряде гладкомышечных клеток) может замедляться, формируя плато ПД, обусловленное сложными изменениями во времени входящих и выходящих токов через мембрану. В последействии МПД может возникнуть гиперполяризация или/и деполяризация мембраны. Это так называемые следовые потенциалы. Следовая гиперполяризация имеет двоякую природу: ионную и метаболическую. Первая связана с тем, что калиевая проницаемость в нервном волокне мембраны остается некоторое время (десятки и даже сотни миллисекунд) повышенной после генерации МПД и смещает мембранный потенциал в сторону калиевого равновесного потенциала. Следовая гиперполяризация после ритмической стимуляции клеток связана преимущественно с активацией электрогенного натриевого насоса, вследствие накопления ионов натрия в клетке.
Причиной деполяризации, развивающейся после генерации МПД (Мембранный потенциал действия), является накопление ионов калия у наружной поверхности мембраны. Последнее, как это следует из уравнения Гольдмана, ведет к увеличению МПП (Мембранный потенциал покоя).
С инактивацией натриевых каналов связано важное свойство нервного волокна, называемое рефрактерностью.
Во время абсолютного рефрактерного периода нервное волокно полностью утрачивает способность возбуждаться при действии раздражителя любой силы.
Относительная рефрактерность, следующая за абсолютной, характеризуется более высоким порогом возникновения МПД (Мембранный потенциал действия).
Представление о мембранных процессах, происходящих во время возбуждения нервного волокна, служит базой для понимания и явления аккомодации. В основе аккомодации ткани при малой крутизне нарастания раздражающего тока лежит повышение порога возбуждения, опережающее медленную деполяризацию мембраны. Повышение порога возбуждения почти целиком определяется инактивацией натриевых каналов. Роль повышения калиевой проницаемости мембраны в развитии аккомодации состоит в том, что оно приводит к падению сопротивления мембраны. Вследствие снижения сопротивления скорость деполяризации мембраны становится еще медленнее. Скорость аккомодации тем выше, чем большее число натриевых каналов при потенциале покоя находится в инактивированном состоянии, чем выше скорость развития инактивации и чем выше калиевая проницаемость мембраны.
Проведение возбуждения
text_fields
text_fields
arrow_upward
Проведение возбуждения по нервному волокну осуществляется за счет локальных токов между возбужденным и покоящимися участками мембраны. Последовательность событий в этом случае представляется в следующем виде.
При нанесении точечного раздражения на нервное волокно в соответствующем участке мембраны возникает потенциал действия. Внутренняя сторона мембраны в данной точке оказывается заряженной положительно по отношению к соседней, покоящейся. Между точками волокна, имеющими различный потенциал, возникает ток (локальный ток), направленный от возбужденного (знак (+) на внутренней стороне мембраны) к невозбужденному (знак (-) на внутренней стороне мембраны) к участку волокна. Этот ток оказывает деполяризующее влияние на мембрану волокна в покоящемся участке и при достижении критического уровня деполяризации мембраны в данном участке возникает МПД (Мембранный потенциал действия). Этот процесс последовательно распространяется по всем участкам нервного волокна.
В некоторых клетках (нейронах, гладких мышцах) МПД имеет не натриевую природу, а обусловлен входом ионов Ca2+ по потенциал-зависимым кальциевым каналам. В кардиомиоцитах генерация МПД связана с входящими натриевым и натрий-кальциевым токами.
doctor-v.ru
1.4. Механизмы потенциала покоя и потенциала действия
1.4.1. Потенциал покоя.
Теория, объясняющая происхождение мембранного потенциала, базируется на двух основных положениях:
1. Мембрана избирательно проницаема для различных ионов. В состоянии покоя мембрана проницаема для катионов и практически непроницаема для анионов. В покое проницаемость для ионов калия гораздо выше, чем для ионов натрия.
2. В мембране имеется биохимический механизм (натрий–калиевая–АТФ-аза), обеспечивающий активный транспорт ионов калия внутрь клетки, а ионов натрия наружу.
В состоянии покоя в цитоплазме клетки ионов натрия в 10 раз меньше, чем снаружи, а ионов калия в 50 раз больше, чем снаружи.
Это состояние поддерживается работой насоса. Работает насос против градиента концентрации с затратой энергии.
В покое мембрана имеет более высокую проницаемость для ионов калия, чем для ионов натрия и тем более анионов, внутри клетки. По закону осмоса, несмотря на работу насоса, калий стремится выйти из клетки, а анионы не могут последовать за ним, что приводит к разделению зарядов и появлению на мембране потенциала, отрицательного внутри и положительного снаружи.
Величина мембранного потенциала зависит от концентрации ионов калия внутри клетки и снаружи и может быть вычислена на основе законов физической химии (уравнение Нернста и уравнение Гольдмана – Ходжкина – Катца).
Работа натриевого насоса при гидролизе одной молекулы АТФ сопровождается выведением из клетки трех ионов натрия и поглощением двух ионов калия. Поскольку при этом перенос зарядов не скомпенсирован, то в результате функционирования АТФ – азы на мембране клетки разность потенциалов суммируется.
Кроме того, в создании отрицательного заряда принимают участие и белковые молекулы протоплазмы клетки.
Потенциал действия.
Возникновение ПД связано с изменением проницаемости клеточной мембраны при ее возбуждении. При деполяризации мембраны до некоторого критического уровня (КУД) открываются натриевые каналы и ионы натрия по градиенту концентрации, без затрат энергии устремляются внутрь клетки, обусловливая фазу деполяризации потенциала действия.
Этот лавинообразный поток ионов натрия внутрь клетки продолжается до момента перезарядки мембраны. Во время овершута (перескока) наступает резкое снижение проницаемости для натрия (натриевая инактивация), но резко увеличивается проницаемость мембраны для ионов калия, которые по градиенту концентрации без затрат энергии, выходят из клетки, компенсируя вошедшие положительно заряженные ионы натрия и обусловливая возвращение мембранного потенциала на исходный уровень (фаза реполяризации).
Таким образом, по заряду (потенциалу) клетка вернулась на исходный уровень, а ионный состав ее нарушен. Внутри увеличилось количество ионов натрия, а снаружи увеличилось количество ионов калия. Это именно та ситуация, когда натриевый насос работает наиболее активно, восстанавливая ионное равновесие (точнее ионное неравновесие) клетки.
Возникновение потенциала действия связано в основном с движением ионов натрия внутрь. Поэтому ПД считают “натриевым потенциалом”, в отличие от потенциала покоя, который считается в основном “калиевым”.
studfiles.net
2_1 Мембранный потенциал покоя кратко (доцент Сазонов В.Ф.)
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
— На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
— На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка — нейрон.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом перемещаются электрические заряды на нервных клетках к как за счёт этого на мембране появляется электрический потенцила. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов — формирование потенциала покоя.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя — это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
«Потенциал» — это возможность, он сродни понятию «потенция». Электрический потенциал мембраны — это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы — ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные — положительно (+).
Таким образом, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках — это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать «плюсики», т.е. положительно заряженные ионы, а внутри — «минусики», т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки — отрицательные «персонажи». ((
Сущность
Сущность потенциала покоя — это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки — «отрицательность», а снаружи — «положительность».
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны , (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях — и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
1) Через ионные насосы-обменники натрий насильно выводится из клетки, а калий затаскивается в клетку.
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне — 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Попробую рассказать коротко, откуда берётся мембранный потенциал покоя в нервных клетках — нейронах. Ведь, как всем теперь известно, наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц — анионов и недостаток положительных частиц — катионов.
И вот тут исследователя и студента поджидает одна из логических ловушек: внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот — из-за потери некоторого количества положительных частиц (катионов).
И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов — катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомню, что это ионы натрия — Na+ и калия — K+.
Натрий-калиевый насос
А всё дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют «собственный» натрий клетки на наружный «чужой» калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Чтобы легче было запомнить, образно можно сказать так: «Клетка любит калий!» (Хотя об истинной любви здесь не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его и так полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от -10 до -70 mV, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки . А в экспериментах на мультипотентных мезенхимальных стромальных клетках (ММСК) костного мозга человека искусственная деполяризация ингибировала дифференцировку клеток (Fischer-Lougheed J., Liu J.H., Espinos E. et al. Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology 2001; 153: 677-85; Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology 1998; 510: 467-76; Sundelacruz S., Levin M., Kaplan D.L. Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. Plos One 2008; 3).
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка «заряжается любовью».
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Как ни странно, но результат этих двух видов любви — пустота!
Именно она, пустота, создаёт в клетке отрицательный электрический заряд — потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал — возможность затягивать внутрь клетки положительно заряженные частицы (ионы).
2. Ионный натриевый потенциал — возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какие-нибудь другие).
3. Ионный калиевый потенциал — возможновть выталкивать из клетки ионы калия (и именно калия, а не какие-нибудь другие).
1. Дефицит натрия (Na+) в клетке.
2. Избыток калия (K+) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь «полезет» этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выравнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно — концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
От химического — к электрическому
А теперь — самое главное, следите за излагаемой мыслью! Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и «плюсики» (положительные заряды). На их месте в клетке остаются «минусы» (отрицательные заряды). Это и есть мембранный потенциал покоя!
Мембранный потенциал покоя — это дефицит положительных зарядов внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Заключение
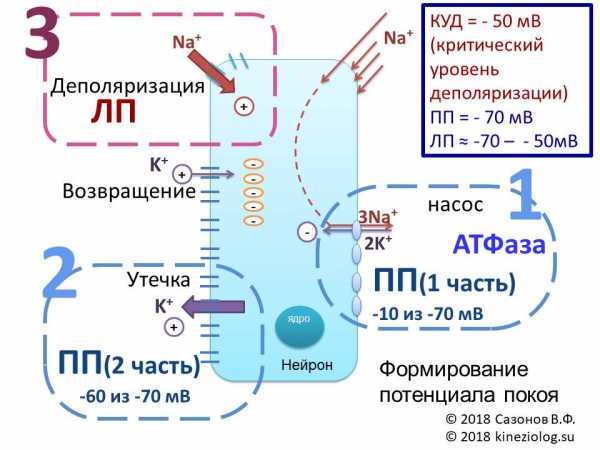
Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Потенциал покоя — отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть — это примерно -10 милливольт, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает «плюсиков» с натрием, чем закачивает обратно с калием).
2. Вторая часть — это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до -70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в -90 милливольт. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне -70 милливольт.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки «лишнего» калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть «мембранный потенциал покоя». Он измеряется внутри клетки и составляет обычно -70 мВ.
Выводы
Говоря образно, «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками».
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше «плюсиков» (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки — неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Итак:
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это — разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Видео: Мембранный потенциал покоя (Resting membrane potential)
© 2009-2018 Сазонов В.Ф. © 2009-2016 kineziolog.bodhy.ru, © 2016-2018 kineziolog.su,
kineziolog.su
Потенциал покоя, действия
Потенциал покоя
Мембраны, в том чикле плазматические, в принципе непроницаемы для заряженных частиц. Правда, в мембране имеется Na+/K+-АТФ-аза (Nа+/К+-АТР-аза), осуществляющая активный перенос ионов Na+ из клетки в обмен на ионы К+. Этот транспорт энергозависим и сопряжен с гидролизом АТФ (АТР). За счет работы «Nа+,К+-насоса» поддерживается неравновесное распределение ионов Na+ и К+ между клеткой и окружающей средой. Поскольку расщепление одной молекулы АТФ обеспечивает перенос трех ионов Na+ (из клетки) и двух ионов К+ (в клетку), этот транспорт электрогенен, т. е. цитоплазма клетки заряжена отрицательно по отношению к внеклеточному пространству.
Электрохимический потенциал. Содержимое клетки заряжено отрицательно по отношению к внеклеточному пространству. Основная причина возникновения на мембране электрического потенциала (мембранного потенциала Δψ,— существование специфических ионных каналов. Транспорт ионов через каналы происходит по градиенту концентрации или под действием мембранного потенциала. В невозбужденной клетке часть К+-каналов находится в открытом состоянии и ионы К+ постоянно диффундируют из нейрона в окружающую среду (по градиенту концентрации). Покидая клетку, ионы К+ уносят положительный заряд, что создает потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости различных ионов видно, что каналы, проницаемые для Na+ и Cl— , преимущественно закрыты. Ионы фосфата и органические анионы, например белки, практически не могут проходить через мембраны. С помощью уравнения Нернста можно показать, что мембранный потенциал нервной клетки в первую очередь определяется ионами К+, которые вносят основной вклад в проводимость мембраны.
Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na+, К+, Са2+ и Cl—. Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (или электровозбудимые), например быстрые Na+-каналы, и лиганд-управляемые (или хемовозбудимые), например никотиновые холинэргические рецепторы. Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами , белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — «закрыт» — и т.д.).
Потенциал действия
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия. Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа+-канала. Ионы Na+ устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала. При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время. В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциал принимает первоначальное значение, и даже превышает на короткое время потенциал покоя. После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
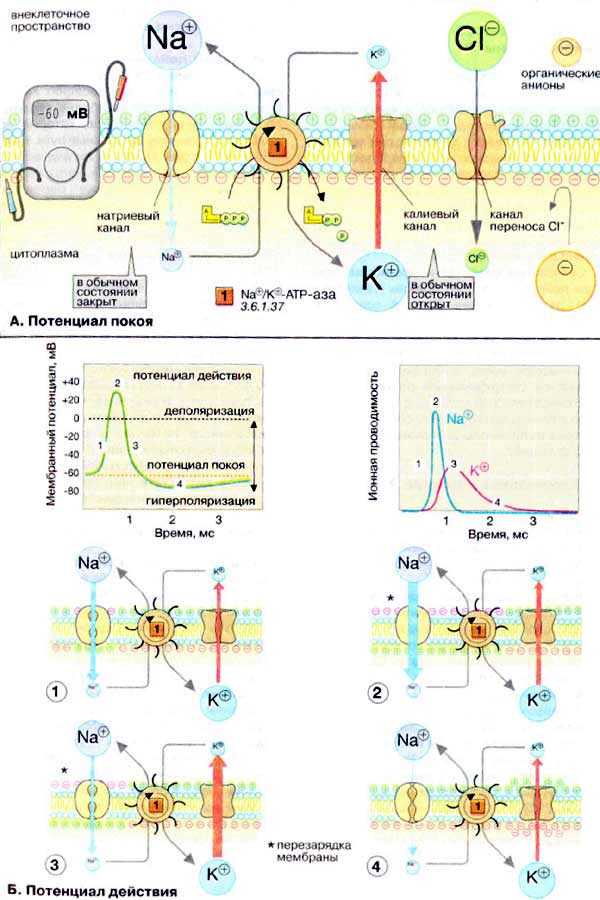
Потенциал покоя, потенциал действия
www.braintools.ru
Мембранный потенциал и его происхождение
МП, или потенциал покоя, — это разность потенциалов между наружной и внутренней поверхностями мембраны в условиях покоя. В среднем у клеток возбудимых тканей он достигает 50—80 мВ, со знаком «—» внутри клетки. Обусловлен преимущественно ионами калия. Как известно, в клетках возбудимых тканей концентрация ионов калия достигает 150 ммоль/л, в среде —4—5 ммоль (ионов калия намного больше в клетке, чем в среде). Поэтому по градиенту концентрации калий может выходить из клетки, и это происходит с участием калиевых каналов, часть которых открыта в условиях покоя. В результате из-за того, что мембрана непроницаема для анионов клетки (глутамат, аспартат, органические фосфаты), на внутренней поверхности клетки образуется избыток отрицательно заряженных частиц, а на наружной — избыток положительно заряженных частиц. Возникает разность потенциалов. Эта ситуация описывается уравнением Нернста:
RT Ke
Ем = ——- * lg ——-
F Ki
Чем выше концентрация калия в среде — тем меньше это отношение, тем меньше величина мембранного потенциала. Однако расчетная величина, как правило, ниже реальной. Например, по расчетам МП должен быть -90 мВ, а реально -70 мВ. Это расхождение обусловлено тем, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В частности, известно, что натрия больше в среде (140 ммоль/л против 14 ммоль/л внутриклеточной). Поэтому натрий может войти в клетку. Но большая часть натриевых каналов в условиях покоя закрыта. Поэтому в клетку входит лишь небольшая часть ионов натрия. Но и этого достаточно, чтобы хотя бы частично компенсировать избыток анионов. Ионы хлора, наоборот, входят в клетку (частично) и вносят отрицательные заряды. В итоге величина мембранного потенциала определяется в основном калием, а также натрием и хлором. Формула Гольдмана-Ходжкина (см. в учебнике) описывает это явление.
Для того чтобы МП поддерживался на постоянном уровне, необходимо поддержание ионного гетерогенитета — ионной асимметрии. Для этого, в частности, служит калий-натриевый насос (и хлорный), который восстанавливает ионную асимметрию, особенно после акта возбуждения. Доказательством калиевой природы МП является наличие зависимости: чем выше концентрация калия в среде, тем меньше величина МП. Для дальнейшего изложения важно понятие: деполяризация (уменьшение МП, например, от минус 90 мВ до минус 70 мВ) и гиперполяризация — противоположное явление.
Потенциал действия
Потенциал действия — это кратковременное изменение разности потенциала между наружной и внутренней поверхностями мембраны (или между двумя точками ткани), возникающее в момент возбуждения. При регистрации потенциала действия с помощью микроэлектродной техники наблюдается типичный пикообразный потенциал. В нем выделяют следующие фазы или компоненты:
10
1. Локальный ответ — начальный этап деполяризации. 2. Фазу деполяризации — быстрое снижение мембранного потенциала до нуля и перезарядка мембраны (реверсия, или овершут).
3. Фазу реполяризации — восстановление исходного уровня мембранного потенциала; в ней выделяют фазу быстрой реполяризации и фазу медленной реполяриэации; в свою очередь, фаза медленной реполяризацик представлена следовыми процессами (потенциалами):
следовая негативность (следовая деполяризация) и следовая позитивность (следовая гиперполяризация). Амплитудно-временные характеристики потенциала действия нерва, скелетной мышцы таковы: амплитуда потенциала действия 140—150 мВ; длительность пика потенциала действия (фаза деполяризации + фаза реполяризации) составляет 1—2 мс, длительность следовых потенциалов—10—50 мс.
Форма потенциала действия (при внутриклеточном отведении) зависит от вида возбудимой ткани: у аксона нейрона, скелетной мышцы — пикообразные потенциалы, у гладких мышц в одних случаях пикообразные, в других — платообразные (например, потенциал действия гладких мышц матки беременной женщины — платообразный, а длительность его составляет почти 1 минуту). У сердечной мышцы потенциал действия имеет платообразную форму.
При внеклеточной регистрации форма потенциала иная, и она зависит от того, какой вид отведения используется — монополярный или биполярный. При биполярном отведении в покое разность потенциалов между двумя точками ткани (например, между точкой 1 и 2) отсутствует. Когда появляется возбуждение, например, распространяющееся в направлении через точку 1 к точке 2 и далее, волна возбуждения достигает вначале электрода 1,— возникает разность потенциалов, при этом область у электрода 1 — электронегативна по отношению к области под электродом 2. Когда волна достигает области под электродом 2, то эта область становится электронегативной по отношению к области под электродом 1. В целом возникает двухфазное изменение разности потенциалов. При монополярном отведении в условиях, когда один из электродов заземлен, потенциал действия по форме однофазный, т. к. вторая область всегда имеет постоянный потенциал, равный 0.
Амплитудные характеристики: если при внутриклеточном отведении амплитуда ПД достигает 120—150 мВ, то при внеклеточном отведении, особенно при дистантном отведении, амплитуда ПД составляет всего 1—2 мВ (например, ЭКГ) или 100—200 мкВ (ЭЭГ). Иначе говоря, при внеклеточном отведении регистрируется только сотая часть потенциала. Важно подчеркнуть, что при внеклеточном отведении регистрируется активность множества одиночных образований, которые в определенной степени могут возбуждаться асинхронно. Это отражается на реальной форме внеклеточно отводимого потенциала. Такова, например, ситуация в отношении ЭКГ.
ПРИРОДА ПОТЕНЦИАЛА ДЕЙСТВИЯ
При исследовании ПД аксонов и сомы нервной клетки, ПД скелетной мышцы было установлено, что фаза деполяризации обусловлена значительным повышением проницаемости для ионов натрия, которые входят в клетку в начале процесса возбуждения и таким образом уменьшают существующую разность потенциала (деполяризация). Процесс этот имеет регенеративную природу — чем выше степень деполяризации, тем выше становится проницаемость натриевых каналов, тем больше входит ионов натрия в клетку и тем выше степень деполяризации. В этот период происходит не только снижение разности потенциалов до нуля, но и изменение поляризованности мембраны — на высоте пика ГЩ внутренняя поверхность мембраны заряжена положительно по отношению к наружной (явление реверсии, или овершута). Однако бесконечно этот процесс идти не может: в результате закрытия инактивационных ворот натриевые каналы закрываются, и приток натрия в клетку прекращается. Затем наступает фаза реполяризации. Она связана с увеличением выхода из клетки ионов калия. Это происходит за счет того, что в результате деполяризации большая часть калиевых каналов, которые в условиях покоя были закрыты, открываются и «+» заряды
11
уходят за пределы клетки. Вначале этот процесс идет очень быстро, потом — медленно, поэтому фаза реполяризации вначале протекает быстро (нисходящая часть пика ПД), а потом медленно (следовая негативность). Этот же процесс лежит в основе фазы следовой гиперполяризации. На фоне следовых потенциалов происходит активация калий-натриевого насоса. Если он работает в электронейтральном режиме (2 иона натрия выносятся из клетки в обмен на 2 вносимых в клетку иона калия), то на форме ПД этот процесс не отражается. Если же насос работает в электрогенном режиме, когда 3 иона натрия выносятся из клетки в обмен на 2 вносимых в клетку иона калия, то в результате на каждый такт работы насоса в клетку вносится на 1 катион меньше, чем выносится, поэтому в клетке постепенно возрастает избыток анионов, т. е. в таком режиме насос способствует появлению дополнительной разности потенциалов. Это явление может лежать в основе фазы следовой гиперполяризации.
Если насос заблокировать, например, с помощью специфического блокатора — стро-фантина G (уабаин), то постепенно клетка перестает отвечать на раздражители, т. к. ионный гетерогенитет как основа электрической активности резко снизится.
Одним из доказательств того, что в аксонах, нейронах, скелетных мышцах ПД по природе натриевый, являются опыты с гипонатриевыми растворами: если ткань помещена в такой раствор, то величина амплитуды ПД снижается и тем на большую величину, чем меньше в среде содержание натрия. Блокаторы натриевых каналов, например, тетродотоксин, также приводят к ингибированию процесса генерации ПД, к снижению амплитудных и временных характеристик ПД. Анестетики за счет их способности блокировать натриевые каналы тоже вызывают нарушение возбудимости, что используется в клинической практике для блокады проведения по нерву.
В сердечной мышце природа ПД иная: процесс деполяризации обусловлен ионами натрия и кальция — эти ионы входят внутрь клетки в начале фазы деполяризации. Поэтому потенциал действия сердечной мышцы может частично уменьшаться блокаторами натриевой проницаемости (тетродотоксином, новокаином), а частично — блокаторами кальциевой проницаемости (верапамилом, нифедипином). В настоящее время блокаторы кальциевой проницаемости используются с целью подавления активности клеток сердца в условиях клиники.
В гладких мышцах сосудов, желудка, кишечника, матки и других образований генерация ПД связана с тем, что в момент возбуждения в клетку входят главным образом не ионы натрия, а ионы кальция. Поэтому блокаторы кальциевой проницаемости, типа верапамила, тормозят активность гладкомышечных клеток. Это используется в клинике, в частности, при лечении угрозы прерывания беременности, т. е. для ингибирования сократительной активности матки.
studfiles.net
Потенциал покоя | Наука | FANDOM powered by Wikia
Потенциа́л поко́я (ПП) — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ[1]. У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки.
История открытия Править
В 1902 г. Ю. Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К+, и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазмой мышцы и окружающей средой, который составлял около — 70 мВ.
Согласно теории Ю. Бернштейна, при возбуждении клетки её мембрана повреждается, и ионы К+ вытекают из клетки по концентрационному градиенту до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя. Это утверждение, относящееся скорее к потенциалу действия, было опровергнуто Ходжкином и Хаксли в 1939 году.
Теорию Бернштейна касательно потенциала покоя подтвердил Кеннет Стюарт Коул (Kenneth Stewart Cole), иногда его инициалы ошибочно пишут как K.C. Cole, из-за его прозвища, Кейси («Kacy»). ПП и ПД изображены на известной иллюстрации Коула и Curtis, 1939. Этот рисунок стал эмблемой Membrane Biophysics Group of the Biophysical Society (см. иллюстрацию).
Общие положения Править
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Концентрации ионов в клетке скелетной мышцы и во внеклеточной среде
| Ионы | Концентрация в саркоплазме (ммоль) | Концентрация вне клетки (ммоль) |
|---|---|---|
| K+ | 140 | 2,5 |
| Na+ | 10 | 120 |
| Cl— | 3-4 | 120 |
| Ca2+ | <0,001 | 2 |
| A— (полипептиды) | 140 | 0 |
Различия в концентрации ионов на противоположных сторонах клеточной мембраны приводят к напряжению, названному мембранным потенциалом. Много ионов имеют градиент концентрации поперек мембраны, включая калий (K+), который является в высокой внутренней части и низкой концентрации вне мембраны. Натрий (Na+) и хлорид (Замкнутый–) ионы — при высоких концентрациях во внеклеточной области, и низких концентрациях во внутриклеточных областях. Эти градиенты концентрации обеспечивают потенциальную энергию вести формирование мембранного потенциала. Это напряжение установлено, когда мембрана имеет проходимость к одному или более ионам. В самом простом случае, иллюстрированном здесь, если мембрана выборочно водопроницаема к калию, они положительно обвиняли, что ионы могут распространить вниз градиент концентрации к внешней стороне ячейки, оставляя позади неданные компенсацию отрицательные обвинения. Это разделение обвинений-, каков вызывает мембранный потенциал. Отметьте, что оптовые решения любой стороны мембраны electo-нейтральны. Аналогично, система в целом нейтральна гальваностереотипом. «Неданные компенсацию» положительные обвинения вне ячейки, и неданных компенсацию отрицательных обвинений в ячейке, физически выстраиваются в линию на мембранной поверхности и привлекают друг друга поперек мембраны. Таким образом, мембранный потенциал физически расположен только в непосредственной близости мембраны. Это — разделение этих обвинений поперек них мембрана, которая является основанием мембранного напряжения. Отметьте также, что эта диаграмма — только приближение ионных вкладов в мембранный потенциал. Другие ионы, включая натрий, хлорид, кальций и другие играют более незначительную роль, даже при том, что они имеют сильные градиенты концентрации, потому что они более ограничили проходимость чем калий. Ключ: Синие пятиугольники — ионы натрия; Фиолетовые площади — ионы калия; Желтые круги — ионы Choloride; Оранжевые прямоугольники — Анионы (они являются результатом разнообразия источников, включая белки). Большая фиолетовая структура со стрелкой представляет трансмембранный канал калия и руководство чистого движения калия.
С помощью уравнения Нернста можно рассчитать равновесный трансмембранный потенциал для K+, который и определяет значение ПП. Но значение потенциала покоя полностью не совпадает с EK+, так как в создании его участвуют также ионы натрия и хлора, вернее, их равновесные потенциалы.
Впоследствии было доказано, что основной вклад в создание потенциала покоя вносит выходящий калиевый ток, который осуществляется через специфические белки-каналы — калиевые каналы постоянного тока. В покое калиевые каналы открыты, а натриевые каналы закрыты. Ионы калия выходят из клетки по градиенту концентрации, что создает на наружной стороне мембраны избыток положительных зарядов; при этом на внутренней стороне мембраны остаются отрицательные заряды. Некоторый (небольшой) вклад в создание потенциала покоя вносит также работа так называемого «натрий-калиевого насоса», который образован особым мембранным ферментом — натрий-калиевой АТФазой.
Потенциал покоя для большинства нейронов составляет величину порядка −60 мВ — −70 мВ. У клеток невозбудимых тканей на мембране также имеется разность потенциалов, разная для клеток разных тканей и организмов.
Формирование потенциала покоя Править
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.
Итак, мембранный потенциал покоя — это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
Дудель Й., Рюэгг Й., Шмидт Р. и др. Физиология человека: в 3-х томах. Пер. с англ / под ред Р. Шмидта и Г. Тевса. — 3. — М.: Мир, 2007. — Т. 1. — 323 с илл. с. — 1500 экз. — ISBN 5-03-000575-3.
ru.science.wikia.com
| Лекция 2. Общая физиология возбудимых тканей. Потенциал покоя. Потенциал действия. ۩ Сущность процесса возбуждения. Сущность процесса возбуждения можно сформулировать следующим образом. Все клетки организма имеют электрический заряд, который создается неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для различных ионов и работы ионных насосов. Процесс возбуждения начинается с действия на возбудимую клетку раздражителя. Сначала очень быстро повышается проницаемость её мембраны для ионов натрия и быстро возвращается к норме, затем – для ионов калия и также быстро, но с некоторым отставанием возвращается к норме. Вследствие этого ионы перемещаются в клетку и из клетки согласно электрохимическому градиенту – это и есть процесс возбуждения. Возбуждение возможно только в том случае, если клетка постоянно поддерживает потенциал покоя (мембранный потенциал) и при её раздражении быстро изменяется проницаемость клеточной мембраны. ۩ Потенциал покоя. Потенциал покоя (ПП) – это разность электрических потенциалов между внутренней и наружной средами клетки в состоянии её покоя. При этом внутри клетки регистрируется отрицательный заряд. Величина ПП в различных клетках различна. Так, в волокнах скелетной мышцы регистрируется ПП равный 60-90 мВ, в нейронах – 50-80 мВ, в гладких мышцах – 30-70 мВ, в сердечной мышце – 80-90 мВ. Органеллы клеток имеют свои вариабельные мембранные потенциалы. Непосредственной причиной существования потенциала покоя является неодинаковая концентрация анионов и катионов внутри и вне клетки (смотри таб.1!). Таблица 1. Внутри- и внеклеточные концентрации ионов в мышечных клетках.
Неравномерное расположение ионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для различных ионов и работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки электрохимическому градиенту. Проницаемость – это её способность пропускать воду, незаряженные и заряженные частицы согласно законам диффузии и фильтрации. Она определяется:
Проводимость – это способность заряженных частиц проходить через клеточную мембрану согласно электрохимическому градиенту. Различная проницаемость различных ионов играет важную роль в формировании ПП:
Кроме выше перечисленных ионов, в формировании ПП играют важную роль и поверхностные заряды мембраны (в основном отрицательные). Их формируют гликопротеиды, гликолипиды и фосфолипиды: фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП, а фиксированные внутренние отрицательные заряды мембраны, напротив, увеличивают ПП, суммируясь с анионами внутри клетки. Таким образом, потенциал покоя — это алгебраическая сумма всех положительных и отрицательных зарядов ионов вне и внутри клетки и поверхностных зарядов клеточной мембраны. Роль ионных насосов в формировании ПП. Ионный насос – это молекула белка, которая обеспечивает перенос иона с непосредственной затратой энергии вопреки электрическому и концентрационному градиентам. В результате сопряженного транспорта натрия и калия поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы Na/K-насоса – перенос трех ионов натрия за пределы клетки и двух ионов калия внутрь клетки. Таким образом, увеличивается ПП. Нормальная величина потенциала покоя является необходимым условием для формирования потенциала действия, то есть для формирования процесса возбуждения. ۩Потенциал действия. Потенциал действия – это электрофизиологический процесс, который выражается в быстром колебании мембранного потенциала вследствие изменения проницаемости мембраны и диффузии ионов в клетку и из клетки. Роль ПД заключается в обеспечении передачи сигналов между нервными клетками, нервными центрами и рабочими органами, в мышцах ПД обеспечивает процесс электромеханического сопряжения. ПД подчиняется закону «всё или ничего». Если сила раздражения мала, то возникает локальный потенциал, который не распространяется. Потенциал действия состоит из трех фаз: деполяризации, то есть исчезновения ПП; инверсии – изменения знака заряда клетки на обратный; реполяризации – восстановление исходного МП. Механизм возникновения потенциала действия. Фаза деполяризации. При действии раздражителя на клетку начальная частичная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Когда деполяризация достигает примерно 50% пороговой величины, возрастает проницаемость мембраны для Na+, причем в первый момент сравнительно медленно. В этот период движущей силой, обеспечивающей движение Na+ в клетку, являются концентрационный и электрический градиенты. Вспомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются), а концентрация Na+ вне клетки в 12 раз больше, чем внутри клетки. Условием, обеспечивающим дальнейший вход Na+ в клетку, является увеличение проницаемости клеточной мембраны, который определяется состоянием воротного механизма натриевых каналов. Воротный механизм натриевых каналов расположен на внешней и внутренней стороне клеточной мембраны, воротный механизм калиевых каналов – только на внутренней стороне мембраны. В каналах для натрия имеются активационные m-ворота, которые расположены с внешней стороны клеточной мембраны, и инактивационные h-ворота, расположенные с внутренней стороны мембраны. В условиях покоя активационные m-ворота закрыты, инактивационные h-ворота открыты. Калиевые активационные ворота закрыты, а инактивационных калиевых ворот нет. Когда деполяризация клетки достигает критической величины, которая обычно составляет 50 мВ, проницаемость мембраны для Na+ резко возрастает, так как открывается большое количество потенциалзависимых m-ворот натриевых каналов и ионы натрия лавиной устремляются в клетку. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, соответственно, проводимости натрия: открываются все новые и новые активационные m-ворота. В итоге ПП исчезает, то есть становится равным нулю. Фаза деполяризации на этом заканчивается. Ее длительность составляет примерно 0,2-0,5 мс. Фаза инверсии. Процесс перезарядки мембраны представляет собой вторую фазу ПД – фазу инверсии. Фаза инверсии делится на восходящую и нисходящую составляющие. Восходящая часть. После исчезновения ПП вход в клетку ионов натрия продолжается, так как натриевые активационные m-ворота еще открыты. В результате заряд внутри клетки становится положительным, а снаружи-отрицательным. В течение доли миллисекунды ионы натрия еще продолжают входить в клетку. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом Na+ в клетку. Нисходящая составляющая фазы инверсии. Примерно через 0,2-0,5 мс после начала деполяризации рост ПД прекращается в результате закрытия натриевых инактивационных h-ворот и открытия калиевых активационных ворот. Поскольку калий находится преимущественно внутри клетки, он, согласно концентрационному градиенту, начинает быстро выходить из нее, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. Во время нисходящей составляющей фазы инверсии выходу ионов калия из клетки способствует также и электрический градиент. К+ выталкивается положительным зарядом из клетки и притягивается отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки. Калий выходит из клетки не только по управляемым каналам, но и по неуправляемым каналам – каналам утечки. Амплитуда ПД складывается из величины ПП и величины фазы инверсии, составляющей у разных клеток 10-50 мВ. Фаза реполяризации. Пока активационные калиевые каналы открыты, K+ еще продолжает выходить из клетки, согласно химическому градиенту. Заряд внутри клетки становится отрицательным, а снаружи – положительным, следовательно, электрический градиент резко тормозит выход ионов калия из клетки. Но так как сила химического градиента больше силы электрического градиента, ионы калия продолжают очень медленно выходить из клетки. Затем активационные калиевые ворота закрываются, остается только выход ионов калия по каналам утечки, то есть по концентрационному градиенту через неуправляемые каналы. Таким образом, ПД вызывается циклическим процессом поступления ионов натрия в клетку и последующего выхода калия из нее. Роль Са2+ в возникновении ПД в нервных клетках незначительна. Однако Са2+ играет очень важную роль в возникновении ПД сердечной мышцы, в передаче импульсов от одного нейрона к другому, от нервного волокна к мышечному, в обеспечении мышечного сокращения. Вслед за ПД возникают следовые явления (характерные для нейронов) – сначала следовая гиперполяризация, а затем следовая деполяризация. Следовая гиперполяризация клеточной мембраны обычно является следствием еще сохраняющейся повышенной проницаемости мембраны для ионов калия. Следовая деполяризация связана с кратковременным повышением проницаемости мембраны для Na+ и входом его в клетку согласно химическому и электрическому градиентам. Кроме этого существуют: а) так называемая фаза абсолютной рефрактерности, или полная невозбудимость клетки. Она приходится на пик ПД и продолжатся 1-2 мс; и б) фаза относительной рефрактерности – период частичного восстановления клетки, когда сильное раздражение может вызвать новое возбуждение. Относительная рефрактерность соответствует конечной части фазы реполяризации и следовой гиперполяризации клеточной мембраны. В нейронах вслед за гиперполяризацией возможна частичная деполяризация клеточной мембраны. В этот период очередной потенциал действия можно вызвать более слабым раздражением, так как МП несколько меньше обычного. Этот период называется фазой экзальтации (период повышенной возбудимости). Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность. Лабильность, или функциональная подвижность, — это скорость протекания одного цикла возбуждения. Мерой лабильности возбудимого образования является максимальное число ПД, которое он может воспроизвести в 1 секунду. Обычно возбуждение продолжается менее 1 мс и подобно взрыву. Такой «взрыв» протекает мощно, но быстро завершается. Поделитесь с Вашими друзьями: | |||
zodorov.ru