
Моторные нейроны – Моторный нейрон это что такое Моторный нейрон: определение — Психология.НЭС
Мотонейроны (нейроны эфферентные, двигательные)
Мотонейроны (нейроны эфферентные, двигательные)
Нейроны после дифференцировки утрачивают способность к пролиферации и становятся высокоспециализированными неделящимися клетками. Основная функция нейрона — принимать, хранить, перерабатывать и передавать информацию на другие нервные клетки, органы или мышцы. По функциям нейроны подразделяются на:
— афферентные (рецепторные, чувствительные) , передающие информацию от органов чувств в центральные отделы нервной системы;
— эфферентные (двигательные, моторные; лат. efferens — выносящий), посылающие импульсы к различным органам и тканям. Они находятся главным образом в передних рогах спинного мозга и в специализированных центрах головного мозга ;
— вставочные (замыкательные, кондукторные, промежуточные) , служащие для переработки и переключения импульсов. ЦНС на 90% состоит из вставочных нейронов.
Тела эфферентных нейронов находятся в ЦНС (или в симпатических и парасимпатических узлах). Их аксоны идут к рабочим органам (мышцам или железам). Различают два вида рабочих, или исполнительных, органов: анимальные — поперечнополосатые (скелетные) мышцы и вегетативные — гладкие мышцы и железы. Соответственно этому имеются нервные окончания аксонов эфферентных нейронов двух типов: двигательные и секреторные. Первые ( мотонейроны ) оканчиваются на мышечных волокнах, образуя бляшки, которые в поперечнополосатых мышцах представляют аксомышечные синапсы. Нервные окончания неисчерченной (гладкой) мышечной ткани образуют вздутия, в которых также содержатся синаптические пузырьки. Секреторные окончания контактируют с железистыми клетками.
Скелетные мышцы иннервируются мотонейронами.
Мотонейроны периферические (альфа- и гамма-мотонейроны)
Мотонейроны корковые
Ссылки:
Все ссылкиmedbiol.ru
Моторный нейрон это что такое Моторный нейрон: определение — Психология.НЭС
Читать PDFСовременные нейропсихологические представления о феномене левшества
Постоева В. А., Пахомов В. П.
В статье, носящей обзорный характер, рассматриваются современные концепции о феномене левшества в контексте нейропсихологического подхода, развиваемого в когнитивной психологии, анализируются научные
Читать PDFЭкспериментальные подходы к нейроэндокринотерапии постстрессорных депрессий
Шаляпина В.Г., Ракицкая В.В., Семенова О.Г.
Нейробиологические представления о самоорганизации психогенно индуцированной астмы
Языков К.Г., Немеров Е.В., Данилец А.В.
Читать PDFПоиск новых методов и приборов для оценки деятельности нейронов мозга в процессах обучения и памяти
Журавлев Борис Васильевич
Обсуждается гипотеза о переходе процессов запоминания и извлечения из памяти от биоэлектрических к нейрохимическим и далее к физическим конформационным изменениям молекул на основе жидкокристаллически
Читать PDFНейрохимические аспекты холинергической модуляции полового поведения при иммобилизационном стрессе
Байрамов А.А., Кудрявцева Т.А., Торкунова О.В.
Данная статья посвящена оценке роли холинергической медиаторной системы в нейрохимических механизмах реализации полового поведения при иммобилизационном стрессе.
Нейрофармакологический анализ межполушарной асимметрии мозга в регуляции поведения, болевой чувствит
Михеев В.В., Шабанов П.Д.
У мышей животных разных генетических линий (CC57W, BALB/c, DBA/2J, C57BL/6J, беспородные) функциональная межполушарная асимметрия головного мозга проявляется в регуляции видотипичного поведения, болев
Читать PDFНейронные константы в психологии
Лебедев Артур Николаевич
В статье ставится проблема определения содержания психологической службы в образовании.
Читать PDFВлияние центральных холинолитиков на половой диморфизм в нейромедиаторной активности мозга 20-дневны
Пренатальное воздействие многих нейротропных соединений вызывает нарушение половой дифференцировки мозга.
Читать PDFВзаимосвязь показателей «Нейротизма» и «Экстра-версии» с познотоническими реакциями человека в проце
Звоников В.М., Биркин А.А.
Читать PDFНейрохимические механизмы когнитивных нарушений при болезни Паркинсона
Шатова А.В.
В статье представлен нейрохимический анализ когнитивных нарушений при болезни Паркинсона. В исследовании приняло участие 117 пациентов с болезнью Паркинсона различной степени тяжести.
Читать PDFМолекулярно-генетический анализ ассоциаций полиморфных маркеров генов нейромедиаторных систем с уров
Гумерова О.В., Столбова О.В., Зарипова Т.Ю., Леконцев Е.В., Воробьева Е.В., Горбунова В.Ю.
Проведен молекулярно-генетический анализ генов нейромедиаторных систем и фермента МАОА в группе близнецов методом полимеразной цепной реакции синтеза ДНК.
Психолого-педагогический и нейропсихологический подход к решению проблемы неуспеваемости младших шко
Осипова В.В.
В статье анализируются причины неуспеваемости младших школьников с точки зрения психологии, педагогики и нейропсихических процессов детского организма, подчеркивается важная роль базовых факторов в пс
vocabulary.ru
Мотонейроны | Doctor-V.ru
Центральный мотонейрон
text_fields
text_fields
arrow_upward
Произвольные движения мышц происходят за счет импульсов, идущих по длинным нервным волокнам из коры большого мозга к клеткам передних рогов спинного мозга. Эти волокна формируют двигательный (корково-спинномозговой ), или пирамидный , путь . Они являются аксонами нейронов, расположенных в прецентральной извилине, в цитоархитектоническом поле 4. Эта зона представляет собой узкое поле, которое тянется вдоль центральной щели от латеральной (или сильвиевой) борозды к передней части парацентральной дольки на медиальной поверхности полушария, параллельно чувствительной области коры постцентральной извилины.
Нейроны, иннервирующие глотку и гортань, расположены в нижней части прецентральной извилины. Далее в восходящем порядке идут нейроны, иннервирующие лицо, руку, туловище, ногу. Таким образом, все участки тела человека спроецированы в прецентральной извилине как бы вверх ногами. Мотонейроны расположены не только в поле 4, они встречаются и в соседних кортикальных полях. В то же время, подавляющее большинство их занимает 5-й кортикальный слой 4-го поля. Они «ответственны» за точные, нацеленные единичные движения. Эти нейроны включают и гигантские пирамидные клетки Беца, имеющие аксоны с толстой миелиновой оболочкой. Эти быстропроводящие волокна составляют лишь 3,4–4 % от всех волокон пирамидного пути. Большинство волокон пирамидного пути исходит из малых пирамидных, или фузиформных (веретенообразных), клеток в двигательных полях 4 и 6. Клетки поля 4 дают около 40 % волокон пирамидного пути, остальные исходят из клеток других полей сенсомоторной зоны.
Мотонейроны поля 4 контролируют тонкие произвольные движения скелетных мышц противоположной половины тела, так как большинство пирамидных волокон переходит на противоположную сторону в нижней части продолговатого мозга.
Импульсы пирамидных клеток двигательной области коры идут по двум путям. Один – корково-ядерный путь – оканчивается в ядрах черепных нервов, второй, более мощный, корково-спинномозговой – переключается в переднем роге спинного мозга на вставочных нейронах, которые в свою очередь оканчиваются на больших мотонейронах передних рогов. Эти клетки передают импульсы через передние корешки и периферические нервы к двигательным концевым пластинкам скелетной мускулатуры.
Когда волокна пирамидного пути покидают двигательную область коры, они, проходя через лучистый венец белого вещества мозга, сходятся по направлению к задней ножке внутренней капсулы. В соматотопическом порядке они проходят внутреннюю капсулу (ее колено и передние две трети заднего бедра) и идут в средней части ножек мозга, нисходят через каждую половину основания моста, будучи окруженными многочисленными нервными клетками ядер моста и волокнами различных систем. На уровне понтомедуллярного сочленения пирамидный путь становится видимым извне, его волокна образуют удлиненные пирамиды по обе стороны от средней линии продолговатого мозга (отсюда его название). В нижней части продолговатого мозга 80–85 % волокон каждого пирамидного пути переходят на противоположную сторону в перекресте пирамид и образуют латеральный пирамидный путь . Остальные волокна продолжают спускаться неперекрещенными в передних канатиках в качестве переднего пирамидного пути . Эти волокна перекрещиваются на сегментарном уровне через переднюю комиссуру спинного мозга. В шейной и грудной частях спинного мозга некоторые волокна соединяются с клетками переднего рога своей стороны, так что мышцы шеи и туловища получают кортикальную иннервацию с обеих сторон.
Перекрещенные волокна спускаются в составе латерального пирамидного пути в латеральных канатиках. Около 90 % волокон образуют синапсы со вставочными нейронами, которые в свою очередь соединяются с большими альфа– и гамма-нейронами переднего рога спинного мозга.
Волокна, формирующие корково-ядерный путь , направляются к двигательным ядрам (V, VII, IX, X, XI, XII) черепных нервов и обеспечивают произвольную иннервацию лицевой и оральной мускулатуры.
Заслуживает внимания и другой пучок волокон, начинающийся в «глазном» поле 8, а не в прецентральной извилине. Импульсы, идущие по этому пучку, обеспечивают содружественные движения глазных яблок в противоположную сторону. Волокна этого пучка на уровне лучистого венца присоединяются к пирамидному пути. Затем они проходят более вентрально в задней ножке внутренней капсулы, поворачивают каудально и идут к ядрам III, IV, VI черепных нервов.
Периферический мотонейрон
text_fields
text_fields
arrow_upward
Волокна пирамидного пути и различных экстрапирамидных путей (ретикулярно-, покрышечно-, преддверно-, красноядерно-спинномозгового и др.) и афферентные волокна, входящие в спинной мозг через задние корешки, оканчиваются на телах или дендритах больших и малых альфа– и гамма-клеток (непосредственно либо через вставочные, ассоциативные или комиссуральные нейроны внутреннего нейронального аппарата спинного мозга) В противоположность псевдоуниполярным нейронам спинномозговых узлов нейроны передних рогов мультиполярны. Их дендриты имеют множественные синаптические связи с различными афферентными и эфферентными системами. Некоторые из них – облегчающие, другие – тормозящие по своему действию. В передних рогах мотонейроны образуют группы, организованные в колонки и не разделенные сегментарно. В этих колонках имеется определенный соматотопический порядок. В шейной части латеральные мотонейроны переднего рога иннервируют кисть и руку, а мотонейроны медиальных колонок – мышцы шеи и груди. В поясничной части нейроны, иннервирующие стопу и ногу, также расположены латерально в переднем роге, а иннервирующие туловище – медиальное. Аксоны клеток передних рогов выходят из спинного мозга вентрально как корешковые волокна, которые собираются по сегментам и образуют передние корешки. Каждый передний корешок соединяется с задним дистальнее спинномозговых узлов и вместе они образуют спинномозговой нерв. Таким образом, каждый сегмент спинного мозга имеет свою пару спинномозговых нервов.
В состав нервов входят также эфферентные и афферентные волокна, исходящие из боковых рогов спинномозгового серого вещества.
Хорошо миелинизированные, быстропроводящие аксоны больших альфа-клеток идут непосредственно к поперечно-полосатой мускулатуре.
Помимо больших и малых альфа-мотонейронов, передние рога содержат многочисленные гамма-мотонейроны. Среди вставочных нейронов передних рогов следует отметить клетки Реншо, тормозящие действие больших мотонейронов. Большие альфа-клетки с толстым и быстропроводящим аксоном осуществляют быстрые сокращения мышц. Малые альфа-клетки с более тонким аксоном выполняют тоническую функцию. Гамма-клетки с тонким и медленнопроводящим аксоном иннервируют проприорецепторы мышечного веретена. Большие альфа-клетки связаны с гигантскими клетками коры полушарий большого мозга. Малые альфа-клетки имеют связь с экстрапирамидной системой. Через гамма-клетки происходит регуляция состояния мышечных проприорецепторов. Среди различных мышечных рецепторов наиболее важными являются нервно-мышечные веретена.
Афферентные волокна, называемые кольцеспиральными , или первичными, окончаниями, имеют довольно толстое миелиновое покрытие и относятся к быстропроводящим волокнам.
Многие мышечные веретена имеют не только первичные, но и вторичные окончания. Эти окончания также отвечают на стимулы растяжения. Потенциал их действия распространяется в центральном направлении по тонким волокнам, сообщающимся со вставочными нейронами, ответственными за реципрокные действия соответствующих мышц-антагонистов. Только небольшое число проприоцептивных импульсов достигает коры больших полушарий, большинство передается по кольцам обратной связи и не достигает коркового уровня. Это элементы рефлексов, которые служат основой произвольных и других движений, а также статических рефлексов, противостоящих силе тяжести.
Экстрафузальные волокна в расслабленном состоянии имеют постоянную длину. При растяжении мышцы растягивается веретено. Кольцеспиральные окончания реагируют на растяжение генерацией потенциала действия, который передается в большой мотонейрон по быстропроводящим афферентным волокнам, а затем опять по быстропроводящим толстым эфферентным волокнам – экстрафузальной мускулатуре. Мышца сокращается, ее исходная длина восстанавливается. Любое растяжение мышцы приводит в действие этот механизм. Перкуссия по сухожилию мышцы вызывает растяжение этой мышцы. Немедленно реагируют веретена. Когда импульс достигает мотонейронов переднего рога спинного мозга, они реагируют, вызывая короткое сокращение. Эта моносинаптическая передача является базовой для всех проприоцептивных рефлексов. Рефлекторная дуга охватывает не более 1–2 сегментов спинного мозга, что имеет большое значение при определении локализации поражения.
Гамма-нейроны находятся под влиянием волокон, нисходящих от мотонейронов ЦНС в составе таких путей, как пирамидные, ретикулярно-спинномозговые, преддверно-спинномозговые. Эфферентные влияния гамма-волокон делают возможным тонкую регуляцию произвольных движений и обеспечивают возможность регулировать силу ответа рецепторов на растяжение. Это называется системой гамма-нейрон–веретено.
Методика исследования. Проводят осмотр, пальпацию и измерение объема мышц, определяют объем активных и пассивных движений, мышечную силу, мышечный тонус, ритмику активных движений и рефлексы. Для выявления характера и локализации двигательных нарушений, а также при клинически незначительно выраженных симптомах используются электрофизиологические методы.
Исследование двигательной функции начинают с осмотра мышц. Обращается внимание на наличие атрофии или гипертрофии. Измеряя сантиметром объем мышц конечности, можно выявить степень выраженности трофических расстройств. При осмотре некоторых больных отмечаются фибриллярные и фасцикулярные подергивания. При помощи ощупывания можно определить конфигурацию мышц, их напряжение.
Активные движения проверяются последовательно во всех суставах и выполняются обследуемым. Они могут отсутствовать или быть ограниченными в объеме и ослабленными по силе. Полное отсутствие активных движений называют параличом, ограничение движений или ослабление их силы – парезом. Паралич или парез одной конечности носит название моноплегии или монопареза. Паралич или парез обеих рук называют верхней параплегией или парапарезом, паралич или парапарез ног – нижней параплегией или парапарезом. Паралич или парез двух одноименных конечностей называют гемиплегией или гемипарезом, паралич трех конечностей – триплегией, четырех конечностей – квадриплегией или тетраплегией.
Пассивные движения определяются при полном расслаблении мышц обследуемого, что позволяет исключить местный процесс (например, изменения в суставах), ограничивающий активные движения. Наряду с этим определение пассивных движений – основной метод исследования тонуса мышц.
Исследуют объем пассивных движений в суставах верхней конечности: плечевом, локтевом, лучезапястном (сгибание и разгибание, пронация и супинация), движения пальцев (сгибание, разгибание, отведение, приведение, противопоставление I пальца мизинцу), пассивные движения в суставах нижних конечностей: тазобедренном, коленном, голеностопном (сгибание и разгибание, вращение кнаружи и внутрь), сгибание и разгибание пальцев.
Сила мышц определяется последовательно во всех группах при активном сопротивлении больного. Например, при исследовании силы мышц плечевого пояса больному предлагают поднять руку до горизонтального уровня, оказывая сопротивление попытке исследующего опустить руку; затем предлагают поднять обе руки выше горизонтальной линии и удерживать их, оказывая сопротивление. Для определения силы мышц плеча больному предлагают согнуть руку в локтевом суставе, а исследующий пробует ее разогнуть; исследуется также сила абдукторов и аддукторов плеча. Для исследования силы мышц предплечья пациенту дают задание выполнить пронацию, а затем супинацию, сгибание и разгибание кисти при сопротивлении во время выполнения движения. Для определения силы мышц пальцев больному предлагают сделать «колечко» из I пальца и каждого из остальных, а исследующий пробует его разорвать. Проверяют силу при отведении V пальца от IV и сведении других пальцев, при сжатии кистей в кулак. Силу мышц тазового пояса и бедра исследуют при задании поднять, опустить, привести и отвести бедро, оказывая при этом сопротивление. Исследуют силу мышц бедра, предлагая больному согнуть и разогнуть ногу в коленном суставе. Силу мышц голени проверяют следующим образом: больному предлагают согнуть стопу, а исследующий удерживает ее разогнутой; затем дается задание разогнуть согнутую в голеностопном суставе стопу, преодолев сопротивление исследующего. Исследуют также силу мышц пальцев стопы при попытке исследующего согнуть и разогнуть пальцы и отдельно согнуть и разогнуть I палец.
Для выявления пареза конечностей проводят пробу Барре: паретичная рука, вытянутая вперед или поднятая кверху, постепенно опускается, приподнятая над постелью нога также постепенно опускается, в то время как здоровая удерживается в приданном положении. При легком парезе приходится прибегать к пробе на ритмику активных движений; пронировать и супинировать руки, сжимать руки в кулаки и разжимать их, передвигать ноги, как на велосипеде; недостаточность силы конечности проявляется в том, что она скорее устает, движения выполняются не так быстро и менее ловко, чем здоровой конечностью. Сила кистей измеряется динамометром.
Тонус мышц – рефлекторное мышечное напряжение, которое обеспечивает подготовку к движению, сохранение равновесия и позы, способность мышцы сопротивляться растяжению. Выделяют два компонента мышечного тонуса: собственный тонус мышцы, который зависит от особенностей происходящих в ней метаболических процессов, и нервно-мышечный тонус (рефлекторный), рефлекторный тонус вызывается чаще растяжением мышцы, т.е. раздражением проприорецепторов, определяемым характером нервной импульсации, которая достигает этой мышцы. Именно этот тонус лежит в основе различных тонических реакций, в том числе антигравитационных, осуществляемых в условиях сохранения связи мышц с ЦНС.
В основе тонических реакций лежит рефлекс на растяжение, замыкание которого совершается в спинном мозге.
На тонус мышц оказывают влияние спинномозговой (сегментарный) рефлекторный аппарат, афферентная иннервация, ретикулярная формация, а также шейные тонические, в том числе вестибулярные центры, мозжечок, система красного ядра, базальные ядра и др.
Состояние мышечного тонуса оценивается при осмотре и ощупываниимышц: при снижении мышечного тонуса мышца дряблая, мягкая, тестообразная. при повышенном тонусе она имеет более плотную консистенцию. Однако определяющим является исследование тонуса мышц путем пассивных движений (сгибатели и разгибатели, приводящие и отводящие мышцы, пронаторы и супинаторы). Гипотония – снижение тонуса мышц, атония – его отсутствие. Снижение мышечного тонуса можно выявить при исследовании симптома Оршанского: при поднятии вверх (у лежащего на спине больного) разогнутой в коленном суставе ноги выявляется переразгибание ее в этом суставе. Гипотония и атония мышц возникают при периферическом параличе или парезе (нарушение эфферентного отдела рефлекторной дуги при поражении нерва, корешка, клеток переднего рога спинного мозга), поражении мозжечка, ствола мозга, полосатого тела и задних канатиков спинного мозга. Гипертония мышц – напряжение, ощущаемое исследующим при пассивных движениях. Различают спастическую и пластическую гипертонию. Спастическая гипертония – повышение тонуса сгибателей и пронаторов руки и разгибателей и аддукторов ноги (при поражении пирамидного пути). При спастической гипертонии наблюдается симптом «перочинного ножа» (препятствие пассивному движению в начальной фазе исследования), при пластической гипертонии – симптом «зубчатого колеса» (ощущение толчков во время исследования тонуса мышц в конечностях). Пластическая гипертония – равномерное повышение тонуса мышц, сгибателей, разгибателей, пронаторов и супинаторов, что встречается при поражении паллидонигральной системы.
doctor-v.ru
Что такое моторный нейрон и каковы его функции?
18 марта 2018Функция моторного нейрона заключается в переносе электрического сигнала в мышцу, что приводит к её сокращению или расслаблению. У позвоночных животных, включая людей, движение сочлененной внутренней скелетной структуры обеспечивается путем координации сокращений мышц. Только мозг способен к этой сложной координации, а электрическая сигнализация, возможно, является единственным быстрым средством отправки приказов мышцам. Средством доставки является электрически возбуждаемая клетка — нейрон.
Что из себя представляет моторный нейрон?
Моторный нейрон, иногда объединенный в сингулярный термин мотонейрон, представляет собой нервную клетку. Его основная структура включает в себя рецептор на одном конце и передатчик на другом, соединенные удлиненным телом, называемым аксоном. Некоторые из нейронов могут составлять 1 м в длину. Цепи нервных клеток соединяются в нервные волокна, которые проходят от мозга до мышц.
columbiamnc.org
Человеческая нервная система состоит из ветвящейся сети нервных волокон, пронизывающей всё тело, и центральной нервной системы, а именно, головного и спинного мозга. Все компоненты состоят из различных специализированных нейронов. Моторный нейрон определяется его эфферентной функцией: он передает сигналы от центральной нервной системы. Нейроны, несущие сигналы в направлении спинного и головного мозга, называются сенсорными нейронами. Не все двигательные движения управляются и контролируются мозгом; например, автоматический рефлекс рывков колена происходит от спинного мозга к мышцам бедра.
Стоит также отметить, что существуют и другие типы мышц, кроме длинных продольных связок, прикрепленных к скелету:
- Сердечная мышца специализируется на ритмическом сокращении.
- Гладкие мышцы, такие как те, которые продвигают пищу через пищеварительный тракт, специализируются на равномерном сжатии в соответствии с их различными формами, такими как сфинктеры и трубки. Хотя это в значительной степени непроизвольные мышечные действия, они, тем не менее, находятся под контролем мозга, который посылает сигналы через двигательные нейроны.
Нейроны, которые контролируют скелетные мышцы, называются соматическими; сердечные и гладкие мышцы контролируются моторными нейронами, называемыми висцеральными.
Люди не могут заряжаться от электрической розетки, поэтому задача моторного нейрона заключается в создании электричества и передаче заряда на следующий нейрон, а затем, пока терминальный нейрон не разрядится, в мышечную ткань. Это достигается посредством химической сигнализации. На конце рецептора нервная клетка расширяет полотно нитей, называемых дендритами, которые контактируют с соседними нейронами. Их клеточные мембраны имеют молекулярные каналы, через которые осуществляется сравнение внутриклеточных и внеклеточных концентраций ионных элементов. Когда разность достигает критической точки, ячейка генерирует электрический импульс, называемый потенциалом действия, который ускоряет аксон и активирует его концевые дендриты.
Электрическая стимуляция дендритов высвобождает химический нейротрансмиттер, называемый ацетилхолин, который соединяет микроскопический промежуток между двумя связанными нейронами, а также промежуток между нервной клеткой и мышечной клеткой. По сути, эти соединения открывают ионные каналы, которые позволяют клетке измерять разность зарядов и решить, проводить ли собственный электрический импульс дальше вниз по нервной системе.
Функция моторного нейрона отлично подходит для работы мышц. Электрический сигнал, который он передает, является положительным или отрицательным. Мышцы также имеют двоичное состояние — напряжение или расслабление.
Источник: wisegeek.com
Метки: мышцы, нейрон
Далее по теме:
www.medkurs.ru
Мотонейрон. Нервный импульс. Синапс. | steelbros.ru
Мотонейрон.Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов – нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления – аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну, подобно электрическим проводам присоединенным к домам.. Таким образом, один мотонейрон управляет целой группой волокон (так называемая нейромоторная единица), которая работает как единое целое.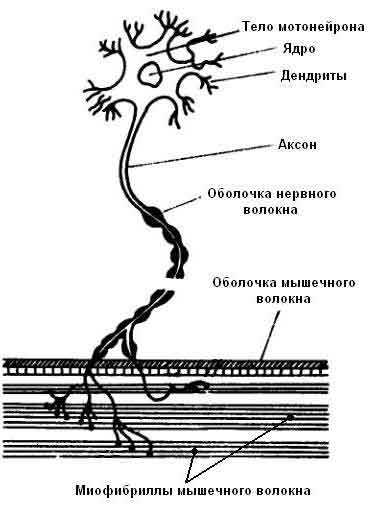
Мышца состоит из множества нейромоторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения.
Рассмотрим более детальное строение клетки нейрона.
Структурной и функциональной единицей нервной системы является нервная клетка – нейрон.
Нейроны – специализированные клетки, способные принимать, обрабатывать, передавать и хранить информацию, организовывать реакцию на раздражения, устанавливать контакты с другими нейронами, клетками органов.
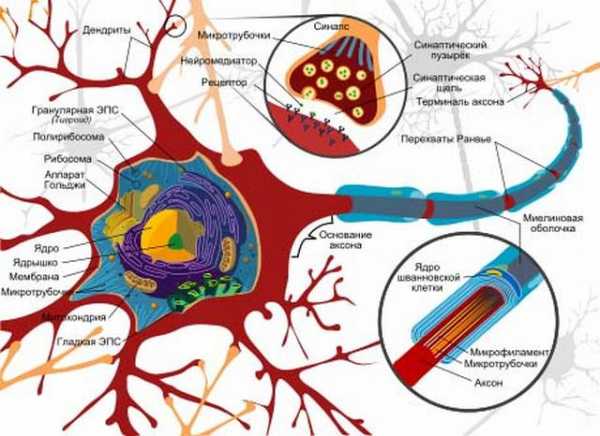
Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый эндоплазматический ретикулум с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксоны. Нейрон имеет развитый и сложный цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов).
Дендриты — ветвящиеся короткие отростки, воспринимающие сигналы от других нейронов, рецепторных клеток или непосредственно от внешних раздражителей. Дендрит проводит нервные импульсы к телу нейрона.
Аксоны – длинный отросток, для проведения возбуждения от тела нейрона.
Уникальными способностями нейрона являются:
— способность генерировать электрические заряды
— передавать информацию с помощью специализированных окончаний – синапсов.
Нервный импульс.
Итак, как же происходит передача нервного импульса?
Если раздражение нейрона превышает определенную пороговую величину, то в точке стимуляции возникает серия химических и электрических изменений, которые распространяются по всему нейрону. Передающиеся электрические изменения называются нервным импульсом.
В отличие от простого электрического разряда, который из-за сопротивления нейрона будет постепенно ослабевать и сумеет преодолеть лишь короткое расстояние, гораздо медленнее «бегущий» нервный импульс в процессе распространения постоянно восстанавливается (регенерирует).
Концентрации ионов (электрически заряженных атомов) – главным образом натрия и калия, а также органических веществ – вне нейрона и внутри него неодинаковы, поэтому нервная клетка в состоянии покоя заряжена изнутри отрицательно, а снаружи положительно; в результате на мембране клетки возникает разность потенциалов (т.н. «потенциал покоя» равен примерно –70 милливольтам). Любые изменения, которые уменьшают отрицательный заряд внутри клетки и тем самым разность потенциалов на мембране, называются деполяризацией.
Плазматическая мембрана, окружающая нейрон, – сложное образование, состоящее из липидов (жиров), белков и углеводов. Она практически непроницаема для ионов. Но часть белковых молекул мембраны формирует каналы, через которые определенные ионы могут проходить. Однако эти каналы, называемые ионными, открыты не постоянно, а, подобно воротам, могут открываться и закрываться.
При раздражении нейрона некоторые из натриевых (Na+) каналов открываются в точке стимуляции, благодаря чему ионы натрия входят внутрь клетки. Приток этих положительно заряженных ионов снижает отрицательный заряд внутренней поверхности мембраны в области канала, что приводит к деполяризации, которая сопровождается резким изменением вольтажа и разрядом – возникает т.н. «потенциал действия», т.е. нервный импульс. Затем натриевые каналы закрываются.
Во многих нейронах деполяризация вызывает также открытие калиевых (K+) каналов, вследствие чего ионы калия выходят из клетки. Потеря этих положительно заряженных ионов вновь увеличивает отрицательный заряд на внутренней поверхности мембраны. Затем калиевые каналы закрываются. Начинают работать и другие мембранные белки – т.н. калий-натриевые насосы, обеспечивающие перемещение Na+ из клетки, а K+ внутрь клетки, что, наряду с деятельностью калиевых каналов, восстанавливает исходное электрохимическое состояние (потенциал покоя) в точке стимуляции.
Электрохимические изменения в точке стимуляции вызывают деполяризацию в прилегающей точке мембраны, запуская в ней такой же цикл изменений. Этот процесс постоянно повторяется, причем в каждой новой точке, где происходит деполяризация, рождается импульс той же величины, что и в предыдущей точке. Таким образом, вместе с возобновляющимся электрохимическим циклом нервный импульс распространяется по нейрону от точки к точке.
Мы выяснили как нервный импульс проходит по нейрону, теперь разберемся с тем как же передается импульс от аксона к мышечному волокну.
Синапс.
Аксон размещается в мышечном волокне в своеобразных карманах, образующийся из выпячиваний аксона и цитоплазмы клеточного волокна.
Между ними образовывается нервно-мышечный синапс.
Нервно-мышечный синапс – нервное окончание между аксоном мотонейрона и мышечным волокном.
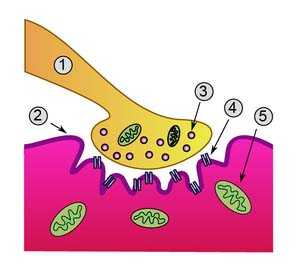
- Аксон.
- Клеточная мембрана.
- Синаптические везикулы аксона.
- Белок-рецептор.
- Митохондрия.
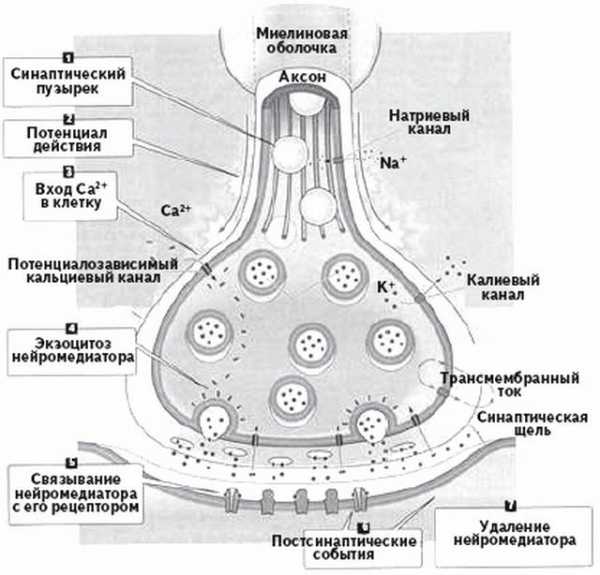
Синапс состоит из трех частей:
1) пресинаптического(отдающий) элемента, содержащего синаптические пузырьки (везикулы) с медиатором
2) синаптической щели ( щель передачи )
3) постсинаптического(воспринимающий) элемента с белками-рецепторами, обеспечивающими взаимодействие медиатора с постсинаптической мембраной и белками-ферментами, разрушающими или инактивирующими медиатор.
Пресинаптический элемент – элемент который отдает нервный импульс.
Постсинаптический элемент – элемент принимающий нервный импульс.
Синаптическая щель – промежуток в котором происходит передача нервного импульса.
Когда нервный импульс в виде потенциала действия (трансмембранный ток, обусловленный ионами натрия и калия) «приходит» к синапсу, в пресинаптический элемент поступают ионы кальция.
Медиатор – биологически активное вещество, выделяемое нервными окончаниями и передающее нервный импульс в синапсе. В передаче импульса к мышечному волокну используется медиатор – ацетилхолин.
Ионы кальция обеспечивают разрыв пузырьков и выход медиатора в синаптическую щель. Пройдя через синаптическую щель, медиатор связывается с белками-рецепторами на постсинаптической мембране. В результате этого взаимодействия на постсинаптической мембране возникает новый нервный импульс, который передается другим клеткам. После взаимодействия с рецепторами медиатор разрушается и удаляется белками-ферментами. Информация передается другим нервным клеткам в закодированном виде (частотные характеристики потенциалов, возникающих на постсинаптической мембране; упрощенным аналогом такого кода является штрих-код на упаковках товаров). «Расшифровка» происходит в соответствующих нервных центрах.
Не связавшийся с рецептором медиатор либо разрушается специальными ферментами, либо захватывается обратно в пузырьки пресинаптического окончания.
Завораживающее видео о том как проходит нервный импульс:
Еще более красивое видео
Синапс
Как проводится нервный импульс ( слайд шоу )
steelbros.ru
Мотонейрон — Медицинская википедия
Мотонейрон (лат. motor — приводящий в движение + нейрон; син. нейрон двигательный) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса.
Мотонейроны называют по той мышце, которую они иннервируют (четырёхглавые, икроножные, полусухожильные и т. п.).
Различают альфа-мотонейроны и гамма-мотонейроны:
| Вид | Функция |
|---|---|
| альфа-мотонейроны | иннервируют волокна скелетной мускулатуры (экстрафузальные волокна)и обеспечивают мышечное сокращение |
| гамма-мотонейроны | иннервируют рецепторы растяжения (интрафузальные волокна) |
См. также
| Гистология: Нервная ткань | |
|---|---|
| Нейроны (Серое вещество) | Сома · Аксон (Аксонный холмик[en], Терминаль аксона[en], Аксоплазма[en], Аксолемма[en], Нейрофиламенты[en]) · Конус роста · Аксонный транспорт · Валлерова дегенерация Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит[en], Базальный дендрит[en]) · Дендритная пластичность · Дендритный потенциал действия типы: Биполярные нейроны · Униполярные нейроны[en] · Псевдоуниполярные нейроны[en] · Мультиполярные нейроны[en] · Пирамидальный нейрон · Звёздчатый нейрон · Клетка Пуркинье · Гранулярная клетка · Интернейрон · Клетка Реншоу |
| Афферентный нерв[en]/ Сенсорный нейрон[en] | GSA · GVA · SSA[en] · SVA[en] · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено (Ib), II или Aβ-волокна[en], III или Aδ-волокна[en], IV или C-волокна[en]) |
| Эфферентный нерв[en]/ Моторный нейрон | GSE[en] · GVE[en] · SVE[en] · Верхний мотонейрон · Нижний мотонейрон[en] (α мотонейроны[en], γ мотонейроны[en]) |
| Синапс | Химический синапс · Нервно-мышечный синапс · Эфапс (Электрический синапс) · Нейропиль · Синаптический пузырёк |
| Сенсорный рецептор | Тельце Мейснера · Тельце Меркеля · Тельце Пачини · Тельце Руффини · Нервно-мышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица |
| Нейроглия | Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия |
| Миелин (Белое вещество?!) | ЦНС: Олигодендроцит |
| Соединительная ткань | Эпиневрий · Периневрий · Эндоневрий · Пучки нервных волокон · Мозговые оболочки: твёрдая, паутинная, мягкая |
Шаблон:Neuroscience-stub
[[Категория:Википедия:Статьи к доработке Шаблон:Rq/topics/getcategory]]medviki.com
Мотонейроны периферические (альфа- и гамма-мотонейроны)
Периферические мотонейроны подразделяются на альфа-мотонейроны и гамма-мотонейроны (рис. 21.2 ).
Меньшие по размеру гамма-мотонейроны иннервируют интрафузальные мышечные волокна . Активация гамма-мотонейронов увеличивает растяжение мышечных веретен, тем самым облегчаясухожильные и другие рефлексы, замыкающиеся через альфа-мотонейроны.
Каждую мышцу иннервирует несколько сотен альфа-мотонейронов. В свою очередь, каждый альфа-мотонейрон иннервирует множество мышечных волокон — около двадцати в наружных мышцах глаза и сотни в мышцах конечностей и туловища .
В нервно-мышечных синапсах выделяется ацетилхолин .
Аксоны периферических мотонейронов идут в составе черепных нервов и передних корешков спинного мозга . На уровне межпозвоночных отверстий передние корешки и задние корешкисливаются, образуя спинномозговые нервы . Несколько соседних спинномозговых нервов образуют сплетение, а затем разветвляются на периферические нервы . Последние тоже неоднократно разветвляются и иннервируют несколько мышц. Наконец, аксон каждого альфа-мотонейрона образует многочисленные разветвления, иннервируя многие мышечные волокна.
Каждый альфа-мотонейрон получает прямые возбуждающие глутаматергические входы откорковых мотонейронов и от чувствительных нейронов , иннервирующих мышечные веретена . Возбуждающие влияния поступают также к альфа- и гамма-мотонейронам от двигательных ядер ствола мозга и вставочных нейронов спинного мозга — как по прямым путям, так и с переключениями.
Прямое постсинаптическое торможение альфа-мотонейронов осуществляют клетки Реншоу — вставочные глицинергические нейроны . Непрямое пресинаптическое торможение альфа-мотонейронов и непрямое пресинаптическое торможение гамма-мотонейронов обеспечивают другие нейроны , образующие ГАМКергические синапсы на нейронах задних рогов .
Тормозное действие на альфа- и гамма-мотонейроны оказывают и другие вставочные нейроны спинного мозга, а также двигательные ядра ствола мозга .
Если преобладают возбуждающие входы, группа периферических мотонейронов активируется. Вначале возбуждаются мелкие мотонейроны. По мере того как сила сокращения мышцы нарастает, частота их разрядов нарастает и вовлекаются крупные мотонейроны. При максимальном сокращении мышцы возбуждена вся соответствующая группа мотонейронов.
Нейронные структуры и их свойства
Тела чувствительных клеток вынесены за пределы спинного мозга (рис.9.1.). Часть из них располагается в спинальных ганглиях. Это тела соматических афферентов, иннервирующих в основном скелетные мышцы. Другие находятся в экстра- и интрамуральных ганглиях автономной нервной системы и обеспечивают чувствительность только внутренних органов.
Чувствительные клетки имеют один отросток, который вскоре после отхождения от тела клетки делится на две ветви.
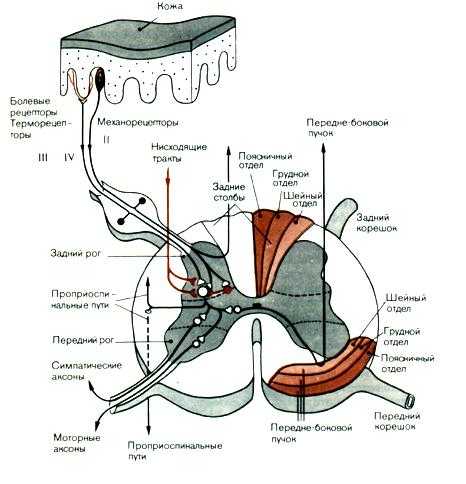
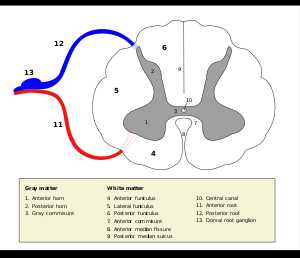
Рис.9.1. Поперечное сечение спинного мозга и связи кожных афферентов в спинном мозгу.
Одна из них проводит возбуждение от рецепторов к телу клетки, другая — от тела нервной клетки к нейронам спинного или головного мозга. Распространение возбуждения из одной ветви в другую может происходить без участия тела клетки.
Нервные волокна чувствительных клеток по скорости проведения возбуждения и диаметру классифицируют на А-, В- и С-группы. Толстые миелинизированные А-волокна с диаметром от 3 до 22 мкм и скоростью проведения возбуждения от 12 до 120 м/с разбиваются еще на подгруппы: альфа— волокна от мышечных рецепторов, бета — от тактильных рецепторов и барорецепторов,дельта — от терморецепторов, механорецепторов, болевых рецепторов. К волокнам группы В относят миелиновые отростки средней толщины со скоростью проведения возбуждения 3-14 м/с. По ним в основном передается ощущение боли. К афферентным волокнам типа С относится большинство безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения до 2 м/с. Это волокна от болевых, хемо- и некоторых механорецепторов.
Сам спинной мозг в целом содержит, например, у человека примерно 13 млн. нейронов. Из их общего числа только около 3% составляют эфферентные, двигательные или моторные нейроны, а остальные 97 % приходятся на долю вставочных, или интернейронов. Двигательные нейроны являются выходными клетками спинного мозга. Среди них различают альфа- и гамма-мотонейроны, а также преганглионарные нейроны автономной нервной системы.
Альфа-мотонейроны осуществляют передачу скелетным мышечным волокнам сигналов, выработанных в спинном мозгу. Аксоны каждого мотонейрона многократно делятся, и, таким образом, каждый из них охватывает своими терминалями до сотни мышечных волокон, образуя в совокупности с ними двигательную единицу. В свою очередь, несколько мотонейронов, иннервирующих одну и ту же мышцу, образуют мотонейронный пул, В его состав могут входить мотонейроны нескольких соседних сегментов. В связи с тем что возбудимость мотонейронов пула неодинакова, при слабых раздражениях возбуждается только часть из них. Это влечет за собой сокращение лишь части мышечных волокон. Другие моторные единицы, для которых это раздражение является подпороговым, тоже реагируют, хотя их реакция выражается лишь в деполяризации мембраны и повышении возбудимости. С усилением раздражения они еще больше вовлекаются в реакцию, и, таким образом, все двигательные единицы пула участвуют в рефлекторном ответе.
Максимальная частота воспроизведения ПД в альфа-мотонейроне не превышает 200-300 имп/с. Вслед за ПД, амплитуда которого составляет 80-100 мВ, возникает следовая гиперполяризациядлительностью от 50 до 150 мс. По частоте импульсации и выраженности следовой гиперполяризации мотонейроны разделяют на две группы: фазические и тонические. Особенности их возбуждения коррелируют с функциональными свойствами иннервируемых мышц. Фазически-ми мотонейронами иннервируются более быстрые, «белые» мышцы, тоническими — более медленные, «красные» .
В организации функции альфа-мотонейронов важным звеном является наличие системы отрицательной обратной связи, образованной аксонными коллатералями и специальными тормозными вставочными нейронами — клетками Реншоу. Своими возвратными тормозными влияниями они могут охватывать большие группы мотонейронов, обеспечивая, таким образом, интеграцию процессов возбуждения и торможения.
Гамма-мотонейроны иннервируют интрафузальные (внутриверетенные) мышечные волокна. Они разряжаются с более низкой частотой, и следовая гиперполяризация у них выражена слабее, чем у альфа-мотонейронов. Их функциональное значение сводится к сокращению интрафузальных мышечных волокон, не приводящему, однако, к появлению двигательного ответа. Возбуждение этих волокон сопровождается изменением чувствительности их рецепторов к сокращению или расслаблению экстрафузальных мышечных волокон.
Нейроны автономной нервной системы составляют особую группу клеток. Тела симпатических нейронов, аксоны которых являются преганглионарными волокнами, располагаются в интермедиолатеральном ядре спинного мозга. По своим свойствам они относятся к группе B-волокон. Характерной особенностью их функционирования является низкая частота свойственной им постоянной тонической импульсной активности. Одни из этих волокон участвуют в поддержании сосудистого тонуса, другие — обеспечивают регуляцию висцеральных эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых клеток).
Тела парасимпатических нейронов образуют крестцовые парасимпатические ядра. Они располагаются в сером веществе крестцовых сегментов спинного мозга. Для многих из них характерна фоновая импульсная активность, частота которой возрастает по мере повышения давления в мочевом пузыре. При раздражении висцеральных тазовых афферентных волокон в этих эфферентных клетках регистрируется вызванный разряд, характеризующийся чрезвычайно большим латентным периодом.
К вставочным, или интернейронам, спинного мозга относятся нервные клетки, аксоны которых не выходят за его пределы. В зависимости от хода отростков различают собственно спинальные и проекционные. Спинальные интернейроны ветвятся в пределах нескольких смежных сегментов, образуя внутрисегментарные и межсегментарные связи. Наряду с ними существуют интернейроны, аксоны которых проходят через несколько сегментов или даже из одного отдела спинного мозга в другой. Их аксоны образуют собственные пучки спинного мозга.
К проекционным интернейронам относятся клетки, длинные аксоны которых формируют восходящие пути спинного мозга. На каждом интернейроне расположено в среднем около 500 синапсов. Синаптические влияния в них опосредуются через ВПСП и ТПСП, суммация которых и достижение критического уровня приводят к возникновению распространяющегося ПД.
studfiles.net