
Мионевральный синапс это – 17) Нервно-мышечный синапс (мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне.
17) Нервно-мышечный синапс (мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне.
Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой мышечного волокна, образующийся из выпячиваний аксона и цитолеммы мышечного волокна, создавая глубокие «карманы». Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором (ацетилхолином).
Механизмы передачи возбуждения в синапсах на примере мионеврального синапса
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР—ХЭ—ХР—ХЭ—ХР—ХЭ.
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
В нервно-мышечном синапсе ( рис. 382.1 ) ацетилхолин синтезируется в окончаниях двигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат к N-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых — гидролизуется АХЭ .
Потенциал концевой пластинки аналогичен ВПСП в межнейронных синапсах.
Однако амплитуда одиночного ПКП существенно выше, чем ВПСП, потому что в нервно-мышечном соединении высвобождаемый нейромедиатор попадает на более обширную поверхность, где связывается с гораздо большим количеством рецепторов и где, следовательно, открывается намного больше ионных каналов. По этой причине амплитуда одиночного ПКП обычно бывает более чем достаточна для того, чтобы в смежной с концевой пластинкой области плазматической мышечной мембраны возник местный электрический ток, инициирующий потенциал действия. Затем потенциал действия распространяется по поверхности мышечного волокна посредством такого же механизма ( рис. 30.19 ), что и в мембране аксона. Большинство нервно- мышечных соединений расположены в срединной части мышечного волокна, откуда возникший потенциал действия распространяется к обоим его концам.
Мионевральный синапс — это… Что такое Мионевральный синапс?
- Мионевральный синапс
-

Нервно-мышечный синапс (мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне.
Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой мышечного волокна, образующийся из выпячиваний аксона и цитолеммы мышечного волокна, создавая глубокие «карманы». Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором (ацетилхолином).
Двигательные нервные окончания в гладкой мышечной ткани построены проще — безмиелиновые пучки аксонов прникают между глиоцитами к пласту гладких мышц и образуют булавовидные расширения, которые содержат холинергические и адренергические пузырьки.
Гистология: Нервная ткань Нейроны
(Серое вещество)Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты)
Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит, Базальный дендрит)
типы: Биполярные нейроны · Псевдополярные нейроны · Мультиполярные нейроны · Пирамидальные клетки · Клетки Пуркинье · Зернистые клеткиАфферентный нерв/
Сенсорный нерв/
Сенсорный нейрон· GVA · SSA · SVA · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено, II or Aβ, Aδ-волокна, C-волокна) Эфферентный нерв/
Моторный нерв/
Моторный нейронGSE · GVE · SVE · Верхний моторный нейрон · Нижний моторный нейрон (α мотонейроны, γ мотонейроны) Синапс Нейропиль · Синаптический пузырек · Сенсорный рецептор Чувствительное тельце Мейснера · Нервное окончание Меркеля · Тельца Пачини · Окончание Руффини · Нервномышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица Нейроглия Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия Миелин
(Белое вещество)CNS: Олигодендроцит
PNS: Клетки Шванна · Невролемма · Перехват Ранвье/Межузловой сегмент · Насечка миелинаСоединительная ткань Эпиневрий · Периневрий · Эндоневрий · Нервные пучки · Оболочки мозга
Wikimedia Foundation. 2010.
Смотреть что такое «Мионевральный синапс» в других словарях:
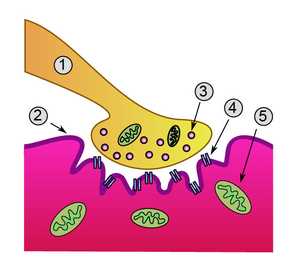
Нервно-мышечный синапс — 1. Пресинаптическое окончание 2. Сарколемма 3. Синаптический пузырек 4. Никотиновый ацетилхолиновый рецептор 5. Митохондрия Нервно мышечный синапс (также … Википедия
НЕРВНОЕ ОКОНЧАНИЕ — (terminatio nervi), специализированное образование в концевом разветвлении отростков нейрона, лишённых миелиновой оболочки; служит для приёма или передачи сигналов. Чувствительные, или сенсорные, Н. о,, осуществляющие приём сигналов (рецепцию),… … Биологический энциклопедический словарь
Нервно-мышечное окончание — Нервно мышечный синапс (мионевральный синапс) эффекторное нервное окончание на скелетном мышечном волокне. Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой… … Википедия
dic.academic.ru
Синапсы. Механизм проведения возбуждения в мионевральном синапсе. — КиберПедия
Синапс-контакт между 2-умя возбудимыми клетками,цитоплазма каждой из которых окружена отдельной электрогенной мембраной.
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану.
После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин. Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняетферментативную функцию.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу.
Мионевральный синапс обладает следующими основными свойствами.
1. Синапс проводит возбуждение только в одном направлении – в направлении от пресинаптической мембраны к постсинаптической.
2. В синапсе имеет место синаптическая задержка возбуждения, т. е. скорость проведения возбуждения по синапсу значительно меньше, чем по нервному волокну. Это связано с определенной продолжительностью времени, необходимого для выделения медиатора и взаимодействия его с рецепторами.
3. В синапсе отмечается облегчение проведения каждого последующего возбуждения, что, по всей вероятности, связано с накоплением медиатора в синаптической щели.
4. При длительном возбуждении синапса в нем может наблюдаться снижение чувствительности рецепторов к медиатору, обусловленное закрытием части натриевых каналов, за счет включения системы инактивации.
5. В синапсах быстро развивается процесс утомления, связанный с быстрым метаболическим истощением запасов медиатора в везикулах пресинаптических утолщений.
Нейтрофильные лейкоциты, их количество в периферической крови, морфологические особенности разных видов лейкоцитов, функции.
Методичка КРОВЬ стр 54
3. Систолический и минутный объёмы крови, факторы, на них влияющие. Методы определения этих показателей.
Методичка КРОВООБРАЩЕНИЕ стр 58
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ №23
1. Виды безусловных рефлексов в зависимости от категории раздражаемых рецепторов. Их особенности, значение и примеры.
Методичка ФИЗИОЛОГИЯ ЧЕЛОВЕКА 010034
Механизмы гемостаза, классификация, общая характеристика.
Методичка КРОВЬ
Секреторная деятельность желудочно-кишечного тракта и ее регуляция. Пищеварение в 12-перстной кишке.
Методичка ОБМЕН ВЕЩЕСТВ
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ №24
Понятие о гомеостазе (гомеокинезе). Значение постоянства состава и свойств внутренней среды организма для жизнедеятельности. Биологические константы организма (жесткие и пластичные), механизмы их поддерживающие.
Учение об относительном постоянстве внутренней среды организма было создано в 1878 году Клодом Бернаром. В 1929 году Кеннон показал, что способность к поддержанию гомеостаза организма является следствием работы его систем регулирования и предложил термин — гомеостаз. Гомеостаз — постоянство внутренней среды (крови, лимфы, тканевой жидкости). Это устойчивость физиологических функций организма. Это основное свойство, отличающее живые организмы от неживого. Чем выше организация живого существа, тем более оно независимо от внешней среды. Внешняя среда — это комплекс факторов, определяющий экологический и социальный микроклимат, действующий на человека. Гомеокинез — комплекс физиологических процессов, обеспечивающий поддержание гомеостаза. Он осуществляется всеми тканями, органами и системами организма, включая ФУС. Параметры гомеостаза являются динамическими и в нормальных пределах изменяются под влиянием факторов внешней среды. Пример: колебание содержания глюкозы в крови. Живые системы не просто уравновешивают внешние воздействия, а активно противодействуют им. Нарушения гомеостаза приводит к гибели организма.
Постоянство внутренней среды организма предстает перед врачом в виде нормативных показателей — констант — постоянных показателей. Константы отражают норму, нормальное значение.
Константы внутренней среды организма делятся на: жесткие и пластичные.
Жесткие константы — это такие константы, которые могут отклоняться от нормы, от своего исходного уровня в процессе жизнедеятельности на небольшую величину (т.е. колебания есть, так как человек живет, но лишь на небольшую величину). Существенное отклонение жестких констант от своей исходной величины не совместимо с жизнью.
(Пример: рН крови)/
Пластичные константы — это тоже постоянные константы, но которые в процессе жизнедеятельности колеблются в значительном диапазоне величин. Однако и при значительном колебании это совместимо с жизнью. Правда и у пластичных констант существуют пределы, выход за которые несовместим с жизнью.
Пример: артериальное давление.
2. Гормоны щитовидной железы. Физиологическое значение тиреоидных гормонов. Регуляция образования гормонов щитовидной железы.
010057 Методичка ФИЗИОЛОГИЯ ЧЕЛОВЕКА
cyberpedia.su
Нервно-мышечный синапс (мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне.
Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой мышечного волокна, образующийся из выпячиваний аксона и цитолеммы мышечного волокна, создавая глубокие «карманы». Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором (ацетилхолином).
Механизмы передачи возбуждения в синапсах на примере мионеврального синапса
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР—ХЭ—ХР—ХЭ—ХР—ХЭ.
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
В нервно-мышечном синапсе ( рис. 382.1 ) ацетилхолин синтезируется в окончаниях двигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат к N-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых — гидролизуется АХЭ .
Потенциал концевой пластинки аналогичен ВПСП в межнейронных синапсах.
Однако амплитуда одиночного ПКП существенно выше, чем ВПСП, потому что в нервно-мышечном соединении высвобождаемый нейромедиатор попадает на более обширную поверхность, где связывается с гораздо большим количеством рецепторов и где, следовательно, открывается намного больше ионных каналов. По этой причине амплитуда одиночного ПКП обычно бывает более чем достаточна для того, чтобы в смежной с концевой пластинкой области плазматической мышечной мембраны возник местный электрический ток, инициирующий потенциал действия. Затем потенциал действия распространяется по поверхности мышечного волокна посредством такого же механизма ( рис. 30.19 ), что и в мембране аксона. Большинство нервно- мышечных соединений расположены в срединной части мышечного волокна, откуда возникший потенциал действия распространяется к обоим его концам.
Похожие статьи:
poznayka.org
Механизмы передачи возбуждения в синапсах на примере мионеврального синапса и его структура
Механизмы передачи возбуждения в синапсах на примере мионеврального синапса и его структура
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану.
После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинап-тическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с преси-наптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холи-нэстеразой (ХЭ), которые находятся на постсинапти-ческой мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР—ХЭ—ХР—ХЭ—ХР—ХЭ.
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате сум-мации образуется ВПСП – возбуждающий постсинап-тический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинап-тической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
cribs.me
НЕРВНО-МЫШЕЧНЫЙ (МИОНЕВРАЛЬНЫЙ) СИНАПС
Изучен наиболее полно. Для него характерно наличие большого числа изгибов на пресинаптической и, особенно, на постсинаптической мембране. Благодаря этому возрастает
площадь контакта пресинапса с постсинапсом, что увеличивает вероятность взаимодействия. Пресинапс заполнен везикулами ацетилхолина (АХ). Примерно каждый из них содержит до 1000—10000 молекул АХ. В основном везикулы расположены в определенных местах пресинапса — около так называемых активных зон. В норме везикулы не подходят близко к пресинаптической мембране, вероятно, из-за того, что имеют такой же заряд, как и пресинапс, хотя возможно и наличие жесткой структуры, которая удерживает везикулы в подвешенном состоянии. Постсинаптическая мембрана представлена в виде гребешков. Они следуют с интервалом примерно в I мкм. На вершине гребешка концентрация холинорецепто-ров достигает максимальных значений (примерно 20000 рецепторов на 1 мкм2), а в устьях — т. е. в глубине — около 1000 рецепторов. Во внесинаптической зоне концентрация холи-норецепторов меньше — порядка 50 на 1 мкмг. В синаптической щели расположен гликока-ликс — волокна, которые выполняют опорную функцию (связь механическая). Здесь расположена ацетилхолинэстераза (АХЭ), способная расщеплять ацетилхолин со скоростью 1 молекула/мс.
Холинорецепторы нервно-мышечного синапса относятся к Н-ХР (никотинчувствитель-ный холинорецептор), т. е. к рецептору, который помимо ацетилхолина способен возбуждаться и под влиянием никотина. Его молекулярная масса, определенная с помощью методики связывания рецептора бунгаротоксином (яд полосатой крайоты — вид змеи), равна 250.000. Молекула рецептора состоит из 5 субъединиц — 2 альфа, бета, гамма и дельта. Узнающая субьединица — это альфа. Внутри рецептора (его внешний вид — гриб) проходит ионный канал, пропускающий натрий. При взаимодействии АХ с ХР вследствие кон-формационных изменений меняется состояние канала — он становится доступным для ионов натрия, что порождает вход их внутрь мышечного волокна в месте постсинаптической мембраны и, как результат, вызывает деполяризацию. В отличие от других синапсов, этот вид деполяризации получил название потенциала концевой пластинки (ПКП).
В условиях покоя, когда пресинаптическая мембрана не деполяризована, способность везикул выходить из пресинапса почти отсутствует. Однако с интервалом примерно в 1 с спонтанно одна из везикул открывается в синаптичёскую щель и выбрасывает квант медиатора. Этот вид активности получил название «миниатюрный потенциал концевой пластинки». Наличие МПКП свидетельствует о квантовой природе выделения медиатора.
Когда к пресинапсу приходит потенциал действия, он вызывает значительную деполяризацию мембраны. Это приводит к тому, что кальций извне поступает внутрь пресинапса и, вероятно, подобно ситуации в мышцах, вызывает сокращение структур синапса (актино-подобных волокон), в результате чего везикулы близко подходят к пресинаптической мембране и происходит экзоцитоз — выброс порции АХ. Если заблокировать кальциевые каналы, то процесс передачи возбуждения в синапсе прекращается. В целом, на 1 ПД выделяется 100 (синапсы лягушки) или 200—300 (синапсы млекопитающих) везикул (квантов) медиатора и в результате генерируется достаточно мощный постсинаптический потенциал (потенциал концевой пластинки), который достигает критического уровня деполяризации, вызывая генерацию полноценного потенциала действия, способного распространяться по обе стороны от постсинаптической области во внесинаптические ареалы. Вышедший в синаптичёскую щель ацетилхолин мгновенно разрушается ацетилхолинэстеразой (АХЭ), превращаясь в холин (он вновь захватывается для последующего синтеза) и ацетат.
Как и все синапсы, нервно-мышечный синалс подвергается фармакологической модификации: можно блокировать проведение ПД по пресинаптическому элементу (новокаином), блокировать высвобождение медиатора, например, за счет удаления из среды ионов кальция или добавления ионов марганца, или при помощи ботулинического токсина, можно заблокировать синтез АХ, угнетая захват холина. Наконец, что в практическом отношении чрезвычайно важно, можно блокировать сами рецепторы и, тем самым, полностью прекратить передачу возбуждения в синапсе. Это можно сделать, используя такие вещества, которые при высоких концентрациях способны вытеснять ацетилхолин с ХР. Этим свойством обладают кураре и курареподобные вещества (д-тубокурарин, диплацин и т. д.). Эта про-
цедура находит широкое применение в хирурпш. Существует также возможность управлять активностью ацетилхолинэстеразы (АХЭ). Если ее активность уменьшить до определенной степени, то это будет способствовать накоплению АХ в синаптической щели. Такая ситуация при определенной патологии (миастения гравис, когда количество выбрасываемого медиатора резко снижено) является благоприятной. Если инактивация АХЭ достигает значительной величины, то это сопровождается развитием стойкой деполяризации в области синапса и приводит к блокаде проведения возбуждения через мионевральный синапс. В конечном итоге это может привести к гибели организма. На этом явлении основано применение ФОС (фосфороорганических соединений) в качестве отравляющих веществ (ОВ) или в качестве инсектицидов (дихлофос, хлорофос и т.д.).
Рекомендуемые страницы:
lektsia.com
НЕРВНО-МЫШЕЧНЫЙ (МИОНЕВРАЛЬНЫЙ) СИНАПС — Мегаобучалка
Изучен наиболее полно. Для него характерно наличие большого числа изгибов на пресинаптической и, особенно, на постсинаптической мембране. Благодаря этому возрастает
площадь контакта пресинапса с постсинапсом, что увеличивает вероятность взаимодействия. Пресинапс заполнен везикулами ацетилхолина (АХ). Примерно каждый из них содержит до 1000—10000 молекул АХ. В основном везикулы расположены в определенных местах пресинапса — около так называемых активных зон. В норме везикулы не подходят близко к пресинаптической мембране, вероятно, из-за того, что имеют такой же заряд, как и пресинапс, хотя возможно и наличие жесткой структуры, которая удерживает везикулы в подвешенном состоянии. Постсинаптическая мембрана представлена в виде гребешков. Они следуют с интервалом примерно в I мкм. На вершине гребешка концентрация холинорецепто-ров достигает максимальных значений (примерно 20000 рецепторов на 1 мкм2), а в устьях — т. е. в глубине — около 1000 рецепторов. Во внесинаптической зоне концентрация холи-норецепторов меньше — порядка 50 на 1 мкмг. В синаптической щели расположен гликока-ликс — волокна, которые выполняют опорную функцию (связь механическая). Здесь расположена ацетилхолинэстераза (АХЭ), способная расщеплять ацетилхолин со скоростью 1 молекула/мс.
Холинорецепторы нервно-мышечного синапса относятся к Н-ХР (никотинчувствитель-ный холинорецептор), т. е. к рецептору, который помимо ацетилхолина способен возбуждаться и под влиянием никотина. Его молекулярная масса, определенная с помощью методики связывания рецептора бунгаротоксином (яд полосатой крайоты — вид змеи), равна 250.000. Молекула рецептора состоит из 5 субъединиц — 2 альфа, бета, гамма и дельта. Узнающая субьединица — это альфа. Внутри рецептора (его внешний вид — гриб) проходит ионный канал, пропускающий натрий. При взаимодействии АХ с ХР вследствие кон-формационных изменений меняется состояние канала — он становится доступным для ионов натрия, что порождает вход их внутрь мышечного волокна в месте постсинаптической мембраны и, как результат, вызывает деполяризацию. В отличие от других синапсов, этот вид деполяризации получил название потенциала концевой пластинки (ПКП).
В условиях покоя, когда пресинаптическая мембрана не деполяризована, способность везикул выходить из пресинапса почти отсутствует. Однако с интервалом примерно в 1 с спонтанно одна из везикул открывается в синаптичёскую щель и выбрасывает квант медиатора. Этот вид активности получил название «миниатюрный потенциал концевой пластинки». Наличие МПКП свидетельствует о квантовой природе выделения медиатора.
Когда к пресинапсу приходит потенциал действия, он вызывает значительную деполяризацию мембраны. Это приводит к тому, что кальций извне поступает внутрь пресинапса и, вероятно, подобно ситуации в мышцах, вызывает сокращение структур синапса (актино-подобных волокон), в результате чего везикулы близко подходят к пресинаптической мембране и происходит экзоцитоз — выброс порции АХ. Если заблокировать кальциевые каналы, то процесс передачи возбуждения в синапсе прекращается. В целом, на 1 ПД выделяется 100 (синапсы лягушки) или 200—300 (синапсы млекопитающих) везикул (квантов) медиатора и в результате генерируется достаточно мощный постсинаптический потенциал (потенциал концевой пластинки), который достигает критического уровня деполяризации, вызывая генерацию полноценного потенциала действия, способного распространяться по обе стороны от постсинаптической области во внесинаптические ареалы. Вышедший в синаптичёскую щель ацетилхолин мгновенно разрушается ацетилхолинэстеразой (АХЭ), превращаясь в холин (он вновь захватывается для последующего синтеза) и ацетат.
Как и все синапсы, нервно-мышечный синалс подвергается фармакологической модификации: можно блокировать проведение ПД по пресинаптическому элементу (новокаином), блокировать высвобождение медиатора, например, за счет удаления из среды ионов кальция или добавления ионов марганца, или при помощи ботулинического токсина, можно заблокировать синтез АХ, угнетая захват холина. Наконец, что в практическом отношении чрезвычайно важно, можно блокировать сами рецепторы и, тем самым, полностью прекратить передачу возбуждения в синапсе. Это можно сделать, используя такие вещества, которые при высоких концентрациях способны вытеснять ацетилхолин с ХР. Этим свойством обладают кураре и курареподобные вещества (д-тубокурарин, диплацин и т. д.). Эта про-
цедура находит широкое применение в хирурпш. Существует также возможность управлять активностью ацетилхолинэстеразы (АХЭ). Если ее активность уменьшить до определенной степени, то это будет способствовать накоплению АХ в синаптической щели. Такая ситуация при определенной патологии (миастения гравис, когда количество выбрасываемого медиатора резко снижено) является благоприятной. Если инактивация АХЭ достигает значительной величины, то это сопровождается развитием стойкой деполяризации в области синапса и приводит к блокаде проведения возбуждения через мионевральный синапс. В конечном итоге это может привести к гибели организма. На этом явлении основано применение ФОС (фосфороорганических соединений) в качестве отравляющих веществ (ОВ) или в качестве инсектицидов (дихлофос, хлорофос и т.д.).
megaobuchalka.ru