
Концентрационный градиент физиология – Физиология биологических мембран. Структура биологической мембраны. Виды транспорта веществ через мембраны
4. Мембранные механизмы возникновения и проведения электрических сигналов
Медицинская реабилитация
4.1. Концентрационный и электрический градиенты
Для того, чтобы понять, как и почему возникает возбуждение в нервных или мышечных клетках, необходимо прежде всего, уяснить основные правила обмена веществами между клеткой и окружающей её средой, поскольку ионы и небольшие молекулы одновременно растворены в водной среде клетки и во внеклеточном пространстве, где их концентрация отличается от внутриклеточной. В среде биологов иногда говорят, что для исследования любой биологической проблемы Бог создал идеальный организм. Эксперименты, положенные в основу мембранной теории, были проведены в 40-х годах ХХ века на гигантских аксонах кальмара.
Диаметр этих аксонов достигает 1 мм, их можно увидеть даже невооружённым глазом, в них легко вводить электроды, чтобы исследовать возникновение электрических сигналов – потенциалов действия. Именно на таком объекте работали основоположники мембранной теории, британские физиологи Алан Ходжкин и Эндрю Хаксли (Hodgkin A., Huxley A.), лауреаты Нобелевской премии 1963 года. Цитоплазма гигантских аксонов кальмара отличается от окружающей внеклеточной жидкости по концентрации некоторых ионов (Таблица 4.1).
Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при котором ток ионов в клетку и из неё становится одинаковым, т.е. фактически ионы не перемещаются.
Как видно из таблицы, концентрация ионов калия внутри клетки намного больше, чем во внеклеточной жидкости, а концентрация ионов натрия и хлора, напротив, значительно больше во внеклеточной жидкости. Органические анионы представляют собой крупные молекулы, которые не проходят через клеточную мембрану.
Корректно или нет делать какие-либо выводы о мембранах клеток теплокровных животных, тем более человека, изучая нервные клетки кальмара? Сравним их гигантские аксоны, например, с мышечными клетками теплокровных (Таблица 4.2).
Результаты измерений концентраций ионов в разных клетках животных, относящихся к разным видам, дают, разумеется и разные значения этих концентраций, но общим для всех клеток, у всех видов животных является одно: концентрация ионов калия всегда больше в клетке, а концентрация ионов натрия и хлора – во внеклеточной жидкости.
Эта разница концентраций или концентрационный градиент является движущей силой для диффузии растворённых ионов в область меньшей концентрации или, в соответствии со вторым законом термодинамики, – к меньшему энергетическому уровню. Ещё раз взглянув на представленные в таблицах цифры, можно безошибочно предсказать, что катионы натрия должны диффундировать в клетку, а катионы калия – из неё.
Однако не всё так просто, поскольку надо учесть и проницаемость клеточной мембраны для различных ионов, а она изменяется в зависимости от состояния активности клетки. В покое у плазматической мембраны открыты лишь ионные каналы для калия, через которые не могут проходить другие ионы. Значит ли это, что через мембрану покоящейся клетки ионы калия могут выходить беспрепятственно?
Выходя из клетки, катионы калия уменьшают в ней количество положительных зарядов и одновременно увеличивают их количество на наружной поверхности мембраны. Остающиеся в клетке органические анионы начинают ограничивать дальнейший выход катионов калия, поскольку между анионами внутренней поверхности мембраны и катионами её наружной поверхности возникает электрическое поле и появляется электростатическое притяжение. Сама же клеточная мембрана оказывается поляризованной: на наружной её поверхности группируются положительные заряды, на внутренней – отрицательные.
Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного тока будут определять два обстоятельства: концентрационный градиент и действие электрического поля, причём концентрационный градиент может направлять ионы в одном направлении, а электрическое поле – в другом. Когда эти две силы уравновешиваются, ток ионов практически прекращается, поскольку количество входящих в клетку ионов становится равным количеству выходящих. Это состояние называется равновесным потенциалом (Е), а его значение можно вычислить с помощью уравнения Нернста (Nernst W., 1888):
где R – газовая постоянная, Т – абсолютная температура (310 при температуре тела), z – валентность иона (для калия = 1), F – константа Фарадея, [K ]a – концентрация ионов калия вне клетки, [K]i – концентрация ионов калия в клетке.
Если подставить в уравнение значение констант и концентрацию ионов, то равновесный потенциал мембраны аксона кальмара для ионов калия окажется равным – 75 мВ (для мышечной мембраны теплокровных – -97 мВ). Это означает, что при такой трансмембранной разности потенциалов и при таких значениях внутри- и внеклеточной концентрации ионов калия их ток из клетки становится равным току в клетку. Если трансмембранная разность потенциалов сделается меньше, то ионы калия станут выходить из клетки до тех пор, пока не восстановится значение равновесного потенциала.
У находящихся в состоянии покоя клеток глии мембрана пропускает только ионы калия, поэтому реальная трансмембранная разность потенциалов у них совпадает с расчётной, т.е. со значением равновесного потенциала для калия – 75 мВ. Но у большинства нейронов дело обстоит иначе, поскольку их мембрана в покое пропускает не только ионы калия, но в небольшом количестве и ионы натрия и хлора. В связи с этим трансмембранная разность потенциалов оказывается несколько меньше, чем равновесный калиевый потенциал, но незначительно, так как проницаемость для ионов калия в покое намного выше, чем для ионов натрия и хлора.
Пользуясь уравнением Нернста нетрудно найти значение равновесных потенциалов для любых ионов (для натрия и хлора они приводятся в таблице 1). Равновесный потенциал для натрия равен + 55 мВ, а его концентрация во внеклеточной среде намного больше, чем в клетке; и то, и другое побуждает ионы натрия входить в клетку. Но в состоянии покоя клеточная мембрана не даёт им такой возможности: её проницаемость для ионов натрия крайне мала.
4.2. Активный транспорт
Диффузия ионов должна уменьшать концентрационный градиент, но концентрационное равновесие означало бы для клетки гибель. Не случайно она более 1/3 своих энергетических ресурсов тратит на поддержание градиентов, на сохранение ионной асимметрии. Перенос ионов через клеточную мембрану против концентрационных градиентов является активным, т.е. энергозатратным видом транспорта, его обеспечивает натрий-калиевый насос.
Это большой интегральный белок клеточной мембраны, который непрерывно выносит из клетки ионы натрия и одновременно закачивает в неё ионы калия. Этот белок обладает свойствами АТФ-азы, фермента, расщепляющего АТФ на внутренней поверхности мембраны, там же белок присоединяет три иона натрия. Освободившаяся при расщеплении молекулы АТФ энергия используется для фосфорилирования определённых участков белка-насоса, после которого меняется конформация белка и он выносит три иона натрия из клетки, но одновременно забирает снаружи и вносит в клетку два иона калия (Рис. 4.1).
Таким образом, за один цикл работы насоса выносятся из клетки три иона натрия, вносятся в неё два иона калия, а на эту работу тратится энергия одной молекулы АТФ. Именно так поддерживается высокая концентрация калия в клетке, а натрия – во внеклеточном пространстве. Если учесть, что и натрий, и калий являются катионами, т.е. несут положительные заряды, то суммарным итогом одного цикла работы насоса для распределения электрических зарядов является удаление одного положительного заряда из клетки. В результате такой деятельности мембрана становится чуть более отрицательной изнутри и поэтому натрий-калиевый насос можно считать электрогенным.
За 1 секунду насос способен вынести из клетки около 200 ионов натрия и одновременно перенести в клетку приблизительно 130 ионов калия, а на одном квадратном микрометре мембранной поверхности может разместиться 100- 200 таких насосов. Кроме натрия и калия насос переносит в клетку против концентрационных градиентов глюкозу и аминокислоты; этот, как бы попутный транспорт, получил название: симпорт. Производительность натрий-калиевого насоса зависит от концентрации в клетке ионов натрия: чем больше она, тем быстрее работает насос. Если же концентрация ионов натрия в клетке понизится, то и насос уменьшит свою деятельность.
Наряду с натрий-калиевым насосом в клеточной мембране существуют специальные насосы для ионов кальция. Они тоже используют энергию АТФ для выноса ионов кальция из клетки, в результате создаётся значительный концентрационный градиент кальция: вне клетки его значительно больше, чем в клетке. Это заставляет ионы кальция постоянно стремиться войти в клетку, но в состоянии покоя клеточная мембрана эти ионы почти не пропускает. Однако порой мембрана раскрывает каналы для этих ионов и тогда они играют очень важную роль в освобождении медиаторов или в активации некоторых ферментов.
Таким образом, активный транспорт создаёт концентрационные и электрические градиенты, которые играют выдающуюся роль во всей жизни клетки.
4.3. Пассивный транспорт – диффузия
Создаваемые работой насосов градиенты дают ионам возможность перемещаться через мембрану от большего энергетического уровня к меньшему путём диффузии, если, конечно, есть открытые ионные каналы. Такой канал – это крупномолекулярный интегральный белок, молекула которого проходит через двойной слой мембранных липидов. В этой молекуле есть заполненная водой пора, диаметр которой не превышает 1 нм. Через такое отверстие способны пройти только ионы калия (Рис. 4.2).
Радиус иона калия – 0,133 нм, у иона натрия он даже меньше – 0, 098 нм, тем не менее, через постоянно открытые каналы может проходить лишь калий. Дело в том, что истинные размеры иона определяются толщиной его гидратной оболочки, которой в водном растворе покрываются все ионы. Молекулы воды ведут себя как диполи: электроны их атомов кислорода сильнее, чем у атомов водорода, а значит кислород несёт слабый отрицательный заряд. Именно поэтому молекулы воды притягиваются положительными зарядами катионов калия, натрия и кальция. Но, поскольку у атомов водорода в молекуле воды есть слабый положительный заряд, существует притяжение водных молекул и к анионам хлора.
При меньшем ионном радиусе электрическое поле у иона натрия сильнее, чем у калия, а потому гидратная оболочка у него толще. Она и не позволяет ионам натрия проходить через каналы, оказывающиеся доступными для прохождения одного лишь калия. Вот почему в состоянии покоя клеточной мембраны через неё происходит ток преимущественно одной разновидности ионов – калия, постоянно уходящего из клетки по концентрационному градиенту.
4.4. Управляемые каналы
Только что описанные каналы, через которые проходят ионы калия, открыты всегда: и в состоянии покоя, и при возбуждении клетки – они мало зависят от внешних условий и поэтому являются каналами пассивного типа. В противоположность этому существуют управляемые ионные каналы, большинство из которых в состоянии покоя клетки закрыты, а для того, чтобы их открыть, необходимо каким-то образом на них подействовать. Следовательно, такие каналы являются управляемыми, а в зависимости от способа управления их подразделяют на три типа:
1) потенциалзависимые;
2) хемозависимые;
3) управляемые механически.
Устройство, с помощью которого открываются или закрываются каналы, часто называют воротным механизмом или даже воротами, хотя такое сравнение не вполне корректно. Современные представления об ионных каналах сложились в связи с двумя методическими подходами к их изучению. Во-первых, это метод локальной фиксации (англ. patch clamp), позволяющий наблюдать ток ионов через одиночный канал. Эту технику изобрели в конце 70-х годов Эрвин Неер и Берт Закман (Neher E., Sakmann B.), лауреаты Нобелевской премии 1991 года. Во-вторых, пониманию свойств каналов способствовало построение их моделей на основе расшифрованного генетического кода многих канальных белков и установленной в связи с этим аминокислотной последовательностью молекул.
Каждый канал образован несколькими белковыми субъединицами (Рис. 4.3), которые представляют собой длинные цепи аминокислот, скрученных в a-спираль. Форма a-спирали может меняться, например, в связи с изменением трансмембранной разности потенциалов (что исключительно важно для потенциалзависимых каналов).
Изменение формы a-спирали приводит к перемещению аминокислот, в том числе несущих электрический заряд. В результате заряды таких аминокислот, как лизин или аргинин, могут оказаться во внутренней стенке ионного канала и сделать её гидрофильной: тогда покрытые гидратной оболочкой ионы смогут проходить через канал. Возвращение альфа-спирали к прежней форме приводит к тому, что во внутренней стенке канала снова оказываются гидрофобные участки и поэтому ток ионов прекращается.
В образовании разных типов каналов участвуют от двух до семи субъединиц, белковая цепь каждой субъединицы несколько раз пересекает клеточную мембрану, причём каждая область пересечения выполняет определённую задачу: одни образуют стенки канала, другие служат датчиками изменений электрического поля, третьи, выступающие за наружную сторону мембраны, являются рецепторами, четвёртые объединяют канал с цитоскелетом.
Потенциалзависимые каналы открываются или закрываются в связи с определёнными изменениями мембранного потенциала. Например, натриевые каналы в состоянии покоя закрыты, но, если мембранный потенциал уменьшится до критического значения, они открываются. Если деполяризация продолжится до положительного значения мембранного потенциала (т.е. на внутренней части мембраны окажется больше положительных зарядов, чем на наружной), то каналы закроются.
Хемозависимые каналы открываются вследствие присоединения нейромедиатора к выступающей наружу гликопротеиновой рецепторной области канального белка – такой тип каналов используется в синапсах (Рис. 4.4). Механически управляемые каналы характерны для чувствительных окончаний нейронов, реагирующих на растяжение и давление. Эти каналы особым способом связаны с цитоскелетом, что и приводит их к открытию при деформации клетки.
Сам момент открытия канала – всего лишь мгновение, длящееся миллионные доли секунды. Но и в открытом состоянии каналы находятся недолго – лишь несколько миллисекунд, после чего стремительно закрываются. Однако пропускная способность открытого канала поразительна: ток ионов происходит со скоростью до 100 000 000 ионов/с, что можно сравнить только с деятельностью самых «быстрых» ферментов, как, например, карбоангидраза, катализирующая образование и дегидратацию углекислоты в эритроцитах крови.
Кроме открытого и закрытого конформационных состояний каналы могут стать инактивированными: это значит, что они закрыты, но не подчиняются, как обычно, действию механизмов управления и не открываются. Состояние инактивации наблюдается немедленно после закрытия каналов, продолжается несколько мс и контролируется особыми субъединицами или особыми регионами белковой молекулы. Во время инактивации каналов клетка перестаёт отвечать на возбуждающие её стимулы, что определяется термином рефрактерность, т.е. временная невозбудимость.
Ионные каналы имеются в мембране любой клетки организма, но в мышечных и особенно в нервных клетках их плотность намного больше, чем в клетках других тканей. В нейронах, кроме высокой плотности каналов, обнаружено и их большое разнообразие. Это не случайно, поскольку именно каналы определяют условия возникновения электрических сигналов, характер самих сигналов, скорость их проведения и т.п., что собственно и позволяет нейронам выполнять свою главную задачу: принимать, перерабатывать и передавать информацию.
4.5. Блокаторы ионных каналов
Существует довольно много веществ, способных обратимо или необратимо связываться с молекулами канальных белков и, тем самым, блокировать их, т.е. выводить из подчинения управляющим механизмам. Блокированные каналы чаще всего оказываются закрытыми, хотя в некоторых случаях фиксируется открытое положение канала.
Блокировать каналы способны многие из давно известных ядов животного или растительного происхождения. Так, например, во внутренностях некоторых сростночелюстных рыб (Tetrodontiformes) содержится тетродотоксин, блокирующий натриевые каналы. К этому отряду относится печально знаменитая рыба фугу, унёсшая жизни немалого количества гурманов, а также плавающая в водах залива Петра Великого собака-рыба, способная раздуваться и издавать при этом довольно громкие звуки. Тетродотоксин достаточно давно применяется в экспериментальной практике, связанной с исследованием мембранной проницаемости.
Натриевые каналы может блокировать и другой животный яд – батрахотоксин, который содержится в слизи некоторых южноамериканских лягушек, например, пятнистого древолаза. Этим ядом индейцы отравляли свои стрелы, хотя и не догадывались, что батрахотоксин блокирует натриевые каналы, а такая блокада не позволяет нервным клеткам возбуждаться.
Другие южноамериканские индейцы готовили отравленные стрелы с помощью другого яда, растительного – это древесный сок кураре, добываемый из некоторых видов лиан. Яд кураре избирательно блокирует хемозависимые каналы нервно-мышечных синапсов. Эти же синапсы необратимо блокирует змеиный яд альфа-бунгаротоксин, который выделяют при укусе бунгары, они же крайты – близкие родственники кобр.
Вещество искусственного происхождения – тетраэтиламмоний специфически блокирует калиевые каналы; его часто использовали в экспериментальной практике. А в медицине применяют многие лекарственные вещества, точкой приложения которых являются ионные каналы: с помощью таких веществ можно управлять определёнными ионными каналами и тем самым влиять на активность нейронов.
4.6. Мембранный потенциал покоя
В покое на наружной стороне плазматической мембраны располагается тонкий слой положительных зарядов, а на внутренней стороне – отрицательных. Электрический заряд наружной поверхности принято считать нулевым, поэтому трансмембранная разность потенциалов или мембранный потенциал покоя имеет отрицательное значение. В типичном для большинства нейронов случае потенциал покоя равен приблизительно -60 – -70 мВ.
Техника прямого измерения потенциала покоя была создана в конце 40-х годов ХХ века. Был изготовлен специальный измерительный электрод: тонкий стеклянный капилляр с оттянутым кончиком диаметром не более 1 мкм и заполненный проводящим электрический ток солевым раствором (3М КСl). который не изменяет внутренний заряд мембраны. В этот раствор с широкого конца капилляра вставляли металлический проводник, а тонким концом протыкали клеточную мембрану. Второй электрод представлял собой хлорированную серебряную пластинку и помещался во внешнюю среду; использовались усилитель слабых электрических сигналов и гальванометр (Рис. 4.5). Объектом исследования был гигантский аксон кальмара, именно на нём удалось получить данные, послужившие основой для мембранной теории (Hodgkin Huxley).
Как же возникает мембранный потенциал покоя? Прежде, чем ответить на этот вопрос, следует ещё раз напомнить, что работой натрий-калиевого насоса в клетке создаётся высокая концентрация ионов калия, а в клеточной мембране для этих ионов есть открытые каналы. Выходящие из клетки по концентрационному градиенту ионы калия увеличивают количество положительных зарядов на наружной поверхности мембраны. В клетке много крупномолекулярных органических анионов и потому изнутри мембрана оказывается заряженной отрицательно. Все остальные ионы могут проходить через покоящуюся мембрану в очень небольшом количестве, их каналы, в основном, закрыты. Следовательно, потенциал покоя обязан своим происхождением, главным образом, току ионов калия из клетки.
Это заключение достаточно просто проверить экспериментально. Если, например, искусственно повысить концентрацию ионов калия вокруг клетки, то их ток из клетки уменьшится или даже вовсе прекратится, поскольку уменьшится концентрационный градиент – движущая сила для этого тока. И тогда начнёт уменьшаться потенциал покоя, он может сделаться равным нулю, если концентрация калия по обе стороны мембраны окажется одинаковой. Есть ещё одна возможность доказать калиевую природу потенциала покоя. Если блокировать калиевые каналы тетраэтиламмонием, то ток ионов калия прекратится, а вслед за этим начнёт уменьшаться потенциал покоя.
Мембрана находящейся в покое клетки пропускает в небольшом количестве ионы натрия и хлора. Две силы гонят ионы натрия в клетку: высокая наружная концентрация и электроотрицательная внутренняя среда клетки. Даже небольшое количество вошедшего в клетку натрия приводит к деполяризации мембраны – уменьшению потенциала покоя. Ионам хлора попасть в клетку труднее, поскольку их отталкивает электроотрицательный слой зарядов на внутренней поверхности мембраны, а значение равновесного потенциала хлора -60 мВ мало отличается от нормальной величины потенциала покоя. Связь между избирательной проницаемостью мембраны для каждого из трёх видов ионов и их концентрациями описывает уравнение Гольдмана:
где Е m – значение мембранного потенциала, Р – проницаемость мембраны, зависящая от её толщины и подвижности в ней иона, a – концентрация иона снаружи, i – концентрация его изнутри, R, T и F имеют то же значение, что и в уравнении Нернста.
Из этого уравнения следует, что реальное значение потенциала покоя (Еm = – 65 мВ) является компромиссным между равновесными потенциалами калия (- 75 мВ), натрия (+ 55 мВ) и хлора (- 60 мВ). Нетрудно предсказать, что повышение проницаемости мембраны для натрия приведёт к деполяризации, а увеличение её проницаемости для хлора – к гиперполяризации.
Если принять за 1 проницаемость мембраны в покое для ионов калия, то проницаемость её для ионов натрия составит 0,04, а для хлора – 0,45. Но при возбуждении мембраны это соотношение изменяется и на вершине пика потенциала действия составляет 1(К) : 20 (Na) : 0,45 (Cl).
4.7. Потенциал действия
Уравнение Гольдмана позволяет вычислить значение мембранного потенциала покоя, если известна концентрация ионов внутри клетки и снаружи, а также проницаемость для этих ионов. Реальная величина мембранного потенциала покоя ближе всего к значению равновесного потенциала для ионов калия, которые проходят через постоянно открытые для них каналы. Ситуация решительно изменяется при раздражении клетки, когда повышается натриевая проницаемость и появляется деполяризующий рецепторный потенциал или постсинаптический потенциал.
Потенциал действия возникает лишь при определённой величине деполяризующего сдвига, например, с -65 мВ до -55 мВ. Если деполяризация будет меньше, то потенциал действия не возникнет: такие деполяризующие сдвиги называются подпороговыми. Приведённые здесь цифры относительны, в разных клетках они могут быть меньше или больше, но всегда наименьший деполяризующий сдвиг, который вызовет появление потенциала действия, определяется, как пороговый.
Возникновение рецепторного или постсинаптического потенциалов связано со сравнительно небольшим местным повышением натриевой проницаемости мембраны. Вход ионов натрия в клетку и обусловленная этим местная деполяризация приводят к возникновению локального электрического тока. Его распространению вдоль мембраны препятствует электрическое сопротивление самой мембраны, поэтому начавшаяся в каком-нибудь месте пассивная деполяризация не может распространяться далеко – пассивные электрические ответы всегда локальны.
Но, если сумма локальных деполяризующих сдвигов всё-таки сможет деполяризовать мембрану триггерной зоны нейрона до критического уровня, до порогового значения, то произойдёт активный и максимальный ответ клетки по правилу «всё или ничего». Деполяризация до критического значения ведёт к конформационным изменениям внутренней стенки натриевых каналов и перемещению полярных аминокислот. Вследствие этого открывается пора диаметром 0,3 – 0,5 нм через которую могут пройти катионы натрия (См. рис. 4.3). Ток анионов через этот канал невозможен, поскольку его устье содержит отрицательные заряды карбоксильных групп глутаминовой кислоты, которые отталкивают отрицательные заряды анионов.
Равновесный потенциал натрия +55 мВ, а каналы для него открываются при значении мембранного потенциала -55 мВ, поэтому ионы натрия поступают в клетку с высокой скоростью: до 107 ионов/с через одиночный канал. Плотность натриевых каналов варьирует от 1 до 50 на одном квадратном микрометре. В результате этого за 0,2 – 0, 5 мс значение мембранного потенциала из отрицательного (-55 мВ) становится положительным (около +30 мВ), хотя и не достигает значения равновесного натриевого потенциала.
Такая стремительная деполяризация является саморегенерирующей: чем больше входит натрия в клетку и чем больше сдвиг мембранного потенциала, тем больше открывается натриевых каналов и тогда ещё больше натрия входит в клетку:
По мере приближения величины мембранного потенциала к значению равновесного натриевого потенциала движущая сила для ионов натрия слабеет, но одновременно растёт движущая сила, заставляющая выходить из клетки ионы калия, каналы для которых постоянно открыты. Когда мембранный потенциал становится положительным, натриевые потенциалзависимые каналы закрываются, а ток калия из клетки резко увеличивается. В связи с этим происходит реполяризация, т.е. восстановление первоначального значения мембранного потенциала (иногда выходной ток калия приводит даже к кратковременной следовой гиперполяризации). Две фазы потенциала действия – деполяризация и реполяризация образуют пик или спайк потенциала действия (Рис. 4.6).
Само открытие натриевых каналов происходит необычайно быстро, в течение не более, чем 10 микросекунд (т.е. миллионных долей секунды), открытыми они остаются несколько миллисекунд, затем быстро закрываются, причём на некоторое время конформация канального белка становится такой, что его невозможно активировать, а значит и открыть каналы. Это состояние называется рефрактерностью, около 1 мс она абсолютна, а затем относительна: при абсолютной рефрактерности нельзя открыть каналы никаким действием, при относительной их нельзя активировать пороговой деполяризацией, но можно надпороговой.
Общая продолжительность рефрактерного состояния определяет максимальную частоту возбуждения нейрона. Например, если рефрактерный период продолжается 2 мс, то за 1с нейрон может возбуждаться максимум 500 раз (1 с = 1000 мс: 2мс = 500). Некоторые нейроны могут возбуждаться чаще, чем 500/с, другие – реже: в соответствии с этим первые можно назвать более лабильными, чем вторые. Проблему лабильности или функциональной подвижности клеток в конце XIX – начале ХХ века исследовал российский физиолог Н.Е.Введенский, он же ввёл в употребление понятие о мере лабильности, как о наибольшем числе электрических осцилляций, которое может воспроизвести за секунду нерв или мышца. Так, например, нерв, по данным Введенского, способен возбуждаться до 500/с, а мышца – лишь до 200/с, т.е. нерв является более лабильным объектом, чем мышца.
4.8. Механизм проведения потенциалов действия
Чем более сложные задачи решает мозг, тем большее количество нейронов ему необходимо. Однако вся масса нейронов должна поместиться в ограниченном черепом и позвоночным каналом пространстве, а потому нервные клетки должны быть небольшими, а их отростки – достаточно тонкими. Но, как известно, чем тоньше и длиннее проводник, тем большее сопротивление он окажет распространяющемуся по нему току. Действующее напряжение в нейроне (V) не может быть больше, чем амплитуда потенциала действия, т.е приблизительно 100-120 мВ, а ток (I), согласно закону Ома, прямо пропорционален напряжению и обратно пропорционален сопротивлению: I = V/R
Из этого следует, что потенциал действия обычным для проведения электричества способом не может распространяться далеко. Очень тонкая мембрана аксона, окруженная электропроводной средой, имеет очень высокую ёмкость, что тормозит распространение электрического сигнала. Если сказать проще: тонкий цитоплазматический отросток – это очень плохой проводник. Но, несмотря на это, потенциалы действия распространяются по аксону с высокой скоростью, достигающей 100 м/с. Как это происходит?
Когда в возбуждённом участке мембраны повышается натриевая проницаемость и возникает потенциал действия, начинается электротоническое распространение положительных зарядов к невозбуждённому участку – этот процесс представляет собой круговой ток (Рис. 4.7). Такой ток деполяризует ещё не возбуждавшийся соседний участок, а когда эта деполяризация достигнет порога – возникнет потенциал действия. Теперь уже этот участок становится источником кругового тока, действующего на следующую область мембраны, теперь уже в этой области возникнет потенциал действия, все параметры которого будут стандартными для данного типа нейрона.
Вслед за повышением натриевой проницаемости в процессе формирования потенциала действия усиливается ток калия из клетки. Вместе с калием из клетки уходят положительные заряды и восстанавливается прежнее значение мембранного потенциала. При любой длине аксона амплитуда потенциалов действия повсюду оказывается неизменной, поскольку в каждом отдельном участке аксона они формируется фактически заново. В физиологическом смысле это важно потому, что постоянство сигнала означает передачу информации по аксону без искажений.
В миелинизированных аксонах круговой ток распространяется к соседнему перехвату, где и возникает потенциал действия. Плотность натриевых каналов в перехватах Ранвье гораздо выше, чем в обычной немиелинизированной мембране и пришедший сюда электротонически круговой ток легко деполяризует перехват до порогового значения. Появившийся вследствие этого потенциал действия служит источником кругового тока для следующего перехвата.
Проведение возбуждения в нерве или мышце можно зарегистрировать с помощью внеклеточных электродов, приложенных к двум различным точкам их поверхности и соединённым с регистрирующей аппаратурой. При распространении потенциала действия мембрана поочерёдно деполяризуется сначала под ближним к источнику возбуждения электродом, а затем – под дальним. И в том, и в другом случае между электродами регистрируется разность потенциалов, поскольку один из них будет находиться на деполяризованном, а значит электроотрицательном снаружи мембраны участке, а второй – в интактной электроположительной точке, где возбуждение ещё не началось, либо уже закончилось.
Регистрация проходящих по мембране потенциалов действия с помощью двух электродов называется биполярной. При таком способе регистрируются две фазы потенциала действия: положительная и отрицательная. Если расположенный под одним из электродов участок сделать невозбудимым (для этого можно подействовать на него каким-либо анестезирующим веществом, например, новокаином), то останется только одна фаза потенциала действия. Такое отведение называется униполярным (или монополярным).
При некоторых аутоиммунных и вирусных заболеваниях миелиновая оболочка разрушается, что приводит к многочисленным неврологическим нарушениям, вплоть до полной утраты некоторых функций; при этом может нарушаться и эмоциональная деятельность, и интеллект. Примером демиелинизирующих заболеваний может служить рассеянный склероз.
Резюме
Возникновение электрических сигналов связано со свойствами клеточной мембраны. Мембранные насосы создают концентрационные градиенты ионов. Открытые в покое ионные каналы для калия позволяют ему выходить из клетки и, тем самым, создавать мембранный потенциал покоя, близкий к равновесному потенциалу для калия. В случае его уменьшения до порогового значения открываются потенциалзависимые каналы для натрия и происходит саморегенерирующая деполяризация, значение мембранного потенциала становится положительным, Это вызывает закрытие натриевых каналов, которые на время инактивируются. Выходящий ток ионов калия восстанавливает прежнее значение мембранного потенциала. Возникновение потенциала действия вызывает появление кругового электрического тока, который деполяризует соседний участок мембраны до порогового значения. В связи с этим потенциал действия распространяется по аксону без уменьшения амплитуды.
Вопросы для самоконтроля
46. Концентрация каких ионов в клетке значительно больше, чем во внеклеточной жидкости?
А. Натрия; Б. Калия; В. Кальция; Г. Хлора; Д. Магния.
47. Какие ионные каналы при физиологическом покое клетки открыты?
А. Для всех катионов; Б. Для анионов; В. Для натрия; Г. Для калия; В. Для кальция.
48. Какова величина равновесного потенциала мембраны гигантского аксона кальмара для ионов калия?
А. +55 мВ; Б. +25-30 мВ; В. = 0; Г. -60 мВ; Д. -75 мВ.
49. Почему натрий-калиевый насос считается электрогенным?
А. Он расходует энергию АТФ; Б. Он создаёт концентрационный градиент калия; В. Он выносит из клетки натрий; Г. За один цикл он удаляет из клетки положительный заряд; Д. Он обеспечивает симпорт глюкозы и аминокислот.
50. Вхождению в клетку каких ионов препятствует электрическое поле между внутренней и наружной поверхностями мембраны?
А. Калия; Б. Натрия; В. Хлора; Г. Кальция; Д. Всех катионов.
51. Через каналы какого типа диффундируют ионы калия, когда клетка находится в состоянии физиологического покоя?
А. Потенциалзависимые; Б. Хемозависимые; В. Потенциалзависимые и хемозависимые; Г. Управляемые механически; Д. Пассивные.
52. Что из перечисленного ниже характерно для состояния рефрактерности?
А. Активированное состояние потенциалзависимых каналов; Б. Инактивированное состояние потенциалзависимых каналов; В. Открытое состояние потенциалзависимых каналов; Г. Закрытое состояние потенциалзависимых каналов; Д. Увеличение пропускной способности потенциалзависимых каналов.
53. Какое из перечисленных ниже веществ является блокатором ионных каналов для калия?
А. Тетраэтиламмоний; Б. Тетродотоксин; В. Батрахотоксин; Г. Кураре; Д. a-Бунгаротоксин.
54. Каким должен быть наименьший деполяризующий сдвиг, если мембранный потенциал равен -69 мВ, а критический уровень деполяризации – -56 мВ?
А. 6 мВ; Б. 9 мВ; В. 11 мВ; Г. 13 мВ; Д. 15 мВ.
55. Если рефрактерный период нейрона продолжается 3 мс, то с какой максимальной частотой он может возбуждаться?
А. 555 Гц; Б. 444 Гц; В. 333 Гц; Г. 222 Гц; Д. 111 Гц.
56. Для какого перемещения ионов через клеточную мембрану, находящейся в покое клетки, необходима энергия?
А. Кальция в клетку; Б. Натрия в клетку; В. Хлора в клетку; Г. Калия из клетки; Д. Кальция из клетки.
57. Какое перемещение ионов происходит только путём диффузии?
А. Натрия из клетки; Б. Калия из клетки; В. Кальция из клетки; Г. Калия в клетку; Д. Глюкозы в клетку.
58. Что заставляет закрыться открывающиеся при возбуждении потенциалзависимые каналы для натрия?
А. Процесс реполяризации; Б. Восстановление исходного значения мембранного потенциала; В. Установление положительного значения мембранного потенциала; Г. Достижение критического уровня деполяризации; Д. Возникновение гиперполяризации.
59. К каким последствиям должно привести повышение мембранной проницаемости для хлора при реальном значении мембранного потенциала -55 мВ?
А. Уменьшение мембранного потенциала; Б. Гиперполяризация; В. Деполяризация; Г. Значение мембранного потенциала не изменится; Д. Возникнет потенциал действия.
60. Каждый потенциал действия образуют две, последовательно сменяющие друг друга фазы – это:
А. Гиперполяризация-деполяризация; Б. Деполяризация-реполяризация; В. Гиперполяризация-реполяризация; Г. Реполяризация- деполяризация; Д. Реполяризация – восстановление первоначального значения мембранного потенциала.
aupam.ru
ГРАДИЕНТ — Большая Медицинская Энциклопедия
ГРАДИЕНТ (лат. gradiens, gradient[is] шагающий) — векторная величина, показывающая направление наиболее быстрого изменения какой-либо функции. Понятием Г. широко пользуются в физике, физ. химии, метеорологии и других науках для характеристики скорости изменения какой-либо величины на единицу длины в направлении ее максимального роста; Г. в биологии — это количественное изменение морфол, или функциональных (в т. ч. биохим.) свойств вдоль одной из осей тела, органа или клетки на любой стадии их развития. Г., отражающий изменение какого-либо физиол, показателя (напр., интенсивности обмена веществ), называют физиол, градиентом (см. Градиент физиологический). При рассмотрении различных биол, процессов чаще встречаются с Г. электрического поля, концентрационным Г., осмотическим Г., гидростатическим Г. и температурным Г.
Градиент электрического поля в биол, объектах возникает в результате перемещения ионов внутри клеток и тканей или вследствие приложения внешнего источника электрического поля, напр, при гальванизации (см. Гальванизация, Электрофорез). Особенно большие значения Г. электрического поля имеют место на биол, мембранах. Так, при толщине мембраны ок. 10 нм и при изменении потенциала на 10 же градиент электрического поля на ней составит 104 в/см. Такое значительное изменение внутреннего электрического поля мембраны может привести к изменению ее поляризации и степени упорядоченности ее структуры. Существует пороговое значение Г. потенциала, при к-ром клетки генерируют потенциал действия (см. Биоэлектрические потенциалы, Возбуждение).
Концентрационный градиент в живых тканях возникает при условии наличия значительной разницы в концентрации ионов во внутренней и внешней среде, напр, высокая внутренняя концентрация ионов калия и низкая концентрация ионов натрия и хлора. Так, внутри волокна сердечной мышцы крысы содержится 140 мкмолей ионов калия и 13 мкмолей ионов натрия на 1 г внутриклеточной воды. Во внешней среде содержится 2,7 мкмоля ионов калия и 150 мкмолей ионов натрия. Концентрационный Г. ионов калия может быть объяснен существованием так наз. доннановского равновесия (см. Мембранное равновесие) по обе стороны биол, мембраны. При этом недиффундирующие анионы (напр., анионы белковых макромолекул) вызывают неравномерное распределение концентрации как анионов (напр., C—), так и катионов (напр., K+) по обе стороны мембраны. Существование концентрационного Г. ионов натрия не может быть объяснено доннановским равновесием, и перенос ионов натрия против концентрационного Г. объясняют существованием активного транспорта ионов (см.). Концентрационный Г. ионов может возникать также в результате протекания метаболических процессов. В итоге все процессы перераспределения ионов по разные стороны биол, мембраны приводят к возникновению потенциалов покоя (см. Биоэлектрические потенциалы).
Поступление и выход различных веществ из клеток происходит вследствие наличия Г. их концентрации. Скорость диффузии веществ определяется соотношением: dn/dt =Dq grad C, где n — количество диффундирующих молекул через поверхность q, D — коэф. диффузии, grad С — градиент концентрации; коэффициент диффузии определяется вязкостью среды и размером молекул вещества. Различие в скорости диффузии катионов и анионов (их подвижности) приводит к появлению диффузионного потенциала φ, который возникает на границе двух соприкасающихся растворов и описывается уравнением Нернста:
где U — подвижность катиона, V — подвижность аниона, С1 и С2 — концентрация электролита в двух соприкасающихся р-рах; R — газовая константа, T — абсолютная t°, n — заряд иона, F — число Фарадея. Диффузионный потенциал минимален, когда подвижность катиона и аниона равны или близки, напр, в случае раствора KCl. Поэтому этот электролит используется в биологии и медицине в качестве жидкостного проводника при гальванизации, электрофорезе и т. д.
Осмотический градиент характеризует разницу в величине осмотического давления (см.) в системе растворитель — раствор, разделенных полупроницаемой мембраной, т. е. проницаемой для молекул растворителя, но непроницаемой для растворенного вещества. Осмотическое давление при этом определяется как величина силы, к-рую нужно приложить к р-ру, чтобы остановить движение растворителя в сторону р-ра. При изменении осмотического давления во внешней среде клетки (напр., при его увеличении) вода будет поступать в клетку; скорость поступления воды при этом будет пропорциональна осмотическому Г. (между внутренней и внешней средой клетки). Так, для эритроцитов скорость проникновения воды составляет величину 2,5 мкм3/мсм2-мин-атм. Величина осмотического давления крови высших животных ок. 40 мм вод. ст. и составляет малую часть от всего кровяного давления. При нарушении белкового или солевого обмена изменяется также и Г. осмотического давления, напр, при его увеличении вода будет поступать в ткань, вызывая отек (см.).
Гидростатический градиент характеризует перепад давления между внешней и внутренней средой клетки, целого организма или отдельных его частей. Так, работа сердца приводит к появлению гидростатического градиента. В артериальной части кровеносной системы возникает положительное гидростатическое давление, в венозной — отрицательное (см. Кровяное давление). Гидростатическое давление может компенсировать осмотическое, что имеет место в капиллярах кровеносной системы. При росте гидростатического Г. (напр., при гипертензии) усиливается выход воды из кровяного русла в ткани, что может привести к возникновению отеков.
Температурный градиент, возникающий вследствие разности температур внутри и вне клетки, существенно влияет практически на все процессы жизнедеятельности. Так, скорость диффузии электролитов увеличивается на 30— 40% при повышении температуры на 10°. Примерно на столько же увеличивается электропроводность клеток. Перенос тепла пропорционален Г. температуры по обе стороны поверхности; при этом Q = -λgrad T, где Q — количество тепла, переносимого через теплопроводящую поверхность, λ — коэф. теплопроводности, T — абсолютная температура. Основным источником тепла в организме человека и животных являются экзотермические процессы, протекающие при работе мышц и внутренних органов. Рассеивание тепла (напр., с поверхности тела человека) может происходить также путем конвекции, излучения и испарения. Все эти процессы ускоряются с ростом температурного Г.
Библиография: Байер В. Биофизика, пер. с нем., М., 1962; Биофизика, под ред. Б. Н. Тарусова и О. Р. Колье, М., 1968; Пасынский А. Г. Биофизическая химия, М., 1968.
Ю. М. Петрусевич.
xn--90aw5c.xn--c1avg
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
8
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Градиент концентрации (от лат. gradi, gradu, gradus — ход, движение, течение, приближение; con — с, вместе, совместно + centrum — центр) или концентрационный градиент — это векторная физическая величина, характеризующая величину и направление наибольшего изменения концентрации какого-либо вещества в среде. Например, если рассмотреть две области с различной концентрацией какого-либо вещества, разделенные полупроницаемой мембраной, то градиент концентрации будет направлен из области меньшей концентрации вещества в область с большей его концентрацией.
Активный транспорт — перенос вещества через клеточную или внутриклеточную мембрану (трансмембранный А.т.) или через слой клеток (трансцеллюлярный А.т.), протекающий против градиента концентрации из области низкой концентрации в область высокой, т. е. с затратой свободной энергии организма. В большинстве случаев, но не всегда, источником энергии служит энергия макроэргических связей АТФ.
Различные транспортные АТФазы, локализованные в клеточных мембранах и участвующие в механизмах переноса веществ, являются основным элементом молекулярных устройств — насосов, обеспечивающих избирательное поглощение и откачивание определенных веществ (например, электролитов) клеткой. Активный специфический транспорт неэлектролитов (молекулярный транспорт) реализуется с помощью нескольких типов молекулярных машин — насосов и переносчиков. Транспорт неэлектролитов (моносахаридов, аминокислот и других мономеров) может сопрягаться с симпортом — транспортом другого вещества, движение которого против градиента концентрации является источником энергии для первого процесса. Симпорт может обеспечиваться ионными градиентами (например, натрия) без непосредственного участия АТФ.
Пассивный транспорт — перенос веществ по градиенту концентрации из области высокой концентрации в область низкой, без затрат энергии (например, диффузия, осмос). Диффузия — пассивное перемещение вещества из участка большей концентрации к участку меньшей концентрации. Осмос — пассивное перемещение некоторых веществ через полупроницаемую мембрану (обычно мелкие молекулы проходят, крупные не проходят).
Существует три типа проникновения веществ в клетку через мембраны: простая диффузия, облегчённая диффузия, активный транспорт.
Простая диффузия
При простой диффузии частицы вещества перемещаются сквозь билипидный слой. Направление простой диффузии определяется только разностью концентраций вещества по обеим сторонам мембраны. Путём простой диффузии в клетку проникают гидрофобные вещества (O2,N2,бензол) и полярные маленькие молекулы (CO2, H2O, мочевина). Не проникают полярные относительно крупные молекулы (аминокислоты, моносахариды), заряженные частицы (ионы) и макромолекулы (ДНК, белки).
Облегченная диффузия
Большинство веществ переносится через мембрану с помощью погружённых в неё транспортных белков (белков-переносчиков). Все транспортные белки образуют непрерывный белковый проход через мембрану. С помощью белков-переносчиков осуществляется как пассивный, так и активный транспорт веществ. Полярные вещества (аминокислоты, моносахариды), заряженные частицы (ионы) проходят через мембраны с помощью облегченной диффузии, при участии белков-каналов или белков-переносчиков. Участие белков-переносчиков обеспечивает более высокую скорость облегченной диффузии по сравнению с простой пассивной диффузией. Скорость облегченной диффузии зависит от ряда причин: от трансмембранного концентрационного градиента переносимого вещества, от количества переносчика, который связывается с переносимым веществом, от скорости связывания вещества переносчиком на одной поверхности мембраны (например, на наружной), от скорости конформационных изменений в молекуле переносчика, в результате которых вещество переносится через мембрану и высвобождается на другой стороне мембраны. Облегченная диффузия не требует специальных энергетических затрат за счет гидролиза АТФ. Эта особенность отличает облегченную диффузию от активного трансмембранного транспорта.
Белки-переносчики
Белки-переносчики — это трансмембранные белки, которые специфически связывают молекулу транспортируемого вещества и, изменяя конформацию, осуществляют перенос молекулы через липидный слой мембраны. В белках-переносчиках всех типов имеются определенные участки связывания для транспортируемой молекулы. Они могут обеспечивать как пассивный, так и активный мембранный транспорт.
Все живые клетки отделены от окружающей среды поверхностью называемой клеточной мембраной. Кроме того, для эукариотов характерно образование внутри клеток нескольких компартментов. Они представлены рядом субклеточных органелл, ограниченных мембранами, например, ядро и митохондрии. Мембраны представляют собой не только статически организованные поверхности раздела, но и включают активные биохимические системы, отвечающие за такие процессы, как избирательный транспорт веществ внутрь и наружу клетки, связывание гормонов и других регуляторных молекул, протекание ферментативных реакций, передача импульсов нервной системы и т.д. Существуют различные типы мембран, отличающиеся по выполняемым функциям. Функции мембран обусловлены их строением.
Функции мембран
Химический состав
Мембраны состоят из липидных и белковых молекул, относительное количество которых варьирует (от 1/5 — белок + 4/5 — липиды до 3/4 — белок + 1/4 – липиды) у разных мембран. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5-10% вещества мембраны.
Липиды мембран
Основная часть липидов в мембранах представлена фосфолипидами, гликолипидами и холестерином. Строение этих липидов представлено на рисунке:

Строение липидов мембран
Липиды мембран имеют в структуре две различные части: неполярный гидрофобный «хвост» и полярную гидрофильную «голову». Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных липидов, расположенных таким образом, что неполярные гидрофобные «хвосты» молекул находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются «хвост к хвосту» так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. Белки мембран включены в липидный двойной слой двумя способами:
связаны с гидрофильной поверхностью липидного бислоя — поверхностные мембранные белки
погружены в гидрофобную область бислоя — интегральные мембранные белки.
Поверхностные белки своими гидрофильными радикалами аминокислот связаны нековалентными связями с гидрофильными группами липидного бислоя. Интегральные белки различаются по степени погруженности в гидрофобную часть бислоя. Они могут располагаться по обеим сторонам мембраны и либо частично погружаются в мембрану, либо прошивают мембрану насквозь. Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связаны с моносахаридными остатками или олигосахаридными цепями и представляют собой гликопротеины. Примеры расположения белков и липидов в мембране представлены на рисунке:
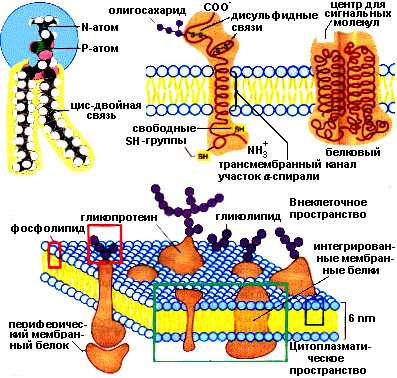
Структура плазматической мембраны
Асимметрия мембран
Хотя каждый монослой образован из липидов, ориентированных одинаковым образом, тем не менее, липидный состав монослоев различен. Например, в плазматической мембране эритроцитов фосфатидилхолины преобладают в наружном слое, а фосфатидилсерины во внутреннем слое мембраны. Углеводные части белков и липидов располагаются на наружной части мембраны. Кроме того, поверхности мембраны отличаются по составу белков. Степень такой асимметрии мембран различна у разных типов мембран и может меняться в процессе жизнедеятельности клетки и ее старения. Подвижность (жесткость) и текучесть мембран также зависят от ее состава. Повышенная жесткость обуславливается увеличением соотношения насыщенных и ненасыщенных жирных кислот, а также холестерина. Физические свойства мембран зависят от расположения белков в липидном слое. Липиды мембран способны к диффузии в пределах слоя параллельно поверхности мембраны (латеральная диффузия). Белки тоже способны к латеральной диффузии. Поперечная диффузия в мембранах сильно ограничена.
Мембранный транспорт
Транспорт веществ внутрь и наружу клетки, а также между цитоплазмой и различными субклеточными органеллами (митохондриями, ядром и т.д.) обеспечивается мембранами. Если бы мембраны были глухим барьером, то внутриклеточное пространство оказалось бы недоступным для питательных веществ, а продукты жизнедеятельности не могли бы быть удалены из клетки. В то же время при полной проницаемости было бы невозможно накопление определенных веществ в клетке. Транспортные свойства мембраны характеризуются полупроницаемостью: некоторые соединения могут проникать через нее, а другие — нет:

Проницаемость мембран для различных веществ
Одна из главных функций мембран — регуляция переноса веществ. Существуют два способа переноса веществ через мембрану: пассивный и активный транспорт:

Транспорт веществ через мембраны
Пассивный транспорт. Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т.е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую и облегченную.
Простая диффузия характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации.
Облегченная диффузия. Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков — переносчиков. Для облегченной диффузии, в отличие от простой, характерна высокая избирательность, так как белок переносчик имеет центр связывания комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка. Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Мало известно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.
Активный транспорт имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии клеткой. Активный транспорт служит для накопления веществ внутри клетки. Источником энергии часто является АТР. Для активного транспорта кроме источника энергии необходимо участие мембранных белков. Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ и K+ через клеточную мембрану. Эта система называется Na+ — K+ — насос. Она отвечает за поддержание состава внутриклеточной среды, в которой концентрация К+ выше, чем Na+ :
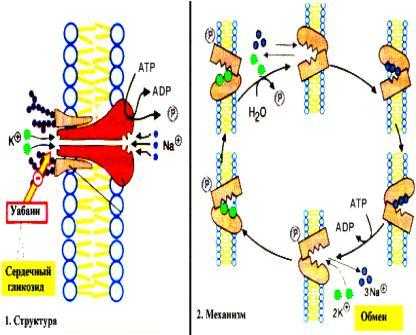
Механизм действия Na+, K+-АТР-азы
Градиент концентрации калия и натрия поддерживается путем переноса К+ внутрь клетки, а Na+ наружу. Оба транспорта происходят против градиента концентрации. Такое распределение ионов определяет содержание воды в клетках, возбудимость нервных клеток и клеток мышц и другие свойства нормальных клеток. Na+ ,K+ -насос представляет собой белок — транспортную АТР-азу. Молекула этого фермента является олигомером и пронизывает мембрану. За полный цикл работы насоса из клетки в межклеточное вещество переносится три иона Na+, а в обратном направлении — два иона К+. При этом используется энергия молекулы АТР. Существуют транспортные системы для переноса ионов кальция (Са2+ — АТР-азы), протонные насосы (Н+ — АТР-азы) и др. Симпорт это активный перенос вещества через мембрану, осуществляемый за счет энергии градиента концентрации другого вещества. Транспортная АТР-аза в этом случае имеет центры связывания для обоих веществ. Антипорт — это перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту своей концентрации. Симпорт и антипорт могут происходить при всасывании аминокислот из кишечника и реабсорбции глюкозы из первичной мочи. При этом используется энергия градиента концентрации ионов Na+, создаваемого Na+, K+-АТР-азой.
studfiles.net
понятие, формула. Транспорт веществ в биологических мембранах
Что такое концентрация? Если говорить в широком смысле, то это соотношение объема вещества и количества растворенных в нем частиц. Данное определение встречается в самых разнообразных отраслях науки, начиная от физики и математики, заканчивая философией. В данном случае, идет речь об употреблении понятия «концентрация» в биологии и химии.
Градиент
В переводе с латыни, это слово означает «растущий» или «шагающий», то есть это некий «указующий перст», который показывает направление, в котором возрастает любая величина. В качестве примера можно использовать, допустим, высоту над уровнем моря в разных точках Земли. Ее (высоты) градиент в каждой отдельной точке на карте будет показывать вектор увеличения значения до достижения самого крутого подъема.
В математике этот термин появился только в конце девятнадцатого века. Его ввел Максвелл и предложил свои обозначения этой величины. Физики используют данное понятие для того, чтобы описывать напряженность электрического или гравитационного поля, изменение потенциальной энергии.
Не только физика, но и другие науки используют термин «градиент». Это понятие может отражать как качественную, так и количественную характеристику вещества, например, концентрацию или температуру.
Градиент концентрации
Что такое градиент теперь известно, а что такое концентрация? Это относительная величина, которая показывает долю вещества, содержащегося в растворе. Она может высчитываться в виде процента от массы, количества молей или атомов в газе (растворе), доли от целого. Такой широкий выбор дает возможность выразить практически любое соотношение. И не только в физике или биологии, но и в метафизических науках.
А в общем, градиент концентрации является векторной величиной, которая одновременно дает характеристику количеству и направлению изменения вещества в среде.
Определение

Можно ли подсчитать градиент концентрации? Формула его представляет собой частность между элементарным изменением концентрации вещества и длинной пути, который придется преодолеть веществу для достижения равновесия между двумя растворами. Математически это выражается формулой С = dC/dl.
Наличие градиента концентрации между двумя веществами является причиной их смешивания. Если частицы движутся из области с большей концентрацией в меньшую, то это называется диффузией, а если между ними находится полупроницаемое препятствие — осмосом.
Активный транспорт

Активный и пассивный транспорт отражает движение веществ через мембраны или слои клеток живых существ: простейших, растений, животных и человека. Этот процесс проходит с использованием тепловой энергии, так как переход веществ осуществляется против градиента концентрации: от меньшего к большему. Наиболее часто для осуществления такого взаимодействия используется аденозинтрифосфат или АТФ — молекула, которая является универсальным источником энергии в 38 Джоулей.
Существуют разные формы АТФ, которые располагаются на мембранах клеток. Энергия, заключенная в них, высвобождается при переносе молекул веществ через так называемые насосы. Это поры в клеточной стенке, которые выборочно поглощают и откачивают ионы электролитов. Кроме того, существует такая модель транспорта как симпорт. В этом случае одновременно транспортируется два вещества: одно выходит из клетки, а другое в нее попадает. Это позволяет сэкономить энергию.
Везикулярный транспорт

Активный и пассивный транспорт включает в себя транспортировку веществ в виде пузырьков или везикул, поэтому процесс называется, соответственно, везикулярным транспортом. Выделяют два его вида:
- Эндоцитоз. В этом случае пузырьки образуются из мембраны клеток в процессе поглощения ею твердых или жидких веществ. Везикулы могут быть гладкими или иметь каемку. Такой способ питания имеют яйцеклетки, белые клетки крови, а также эпителий почек.
- Экзоцитоз. Исходя из названия, это процесс противоположный предыдущему. Внутри клетки есть органеллы (например, аппарат Гольджи), которые «упаковывают» вещества в пузырьки, а они, в последующем, выходят через мембрану.
Пассивный транспорт: диффузия

Движение по градиенту концентрации (от высокой к низкой) происходит без использования энергии. Выделяют два варианта пассивного транспорта – это осмос и диффузия. Последняя бывает простой и облегченной.
Основное отличие осмоса в том, что процесс перемещения молекул происходит через полупроницаемую мембрану. А диффузия по градиенту концентрации происходит в клетках, имеющих мембрану с двумя слоями липидных молекул. Направление транспорта зависит только от количества вещества с обеих сторон мембраны. Этим способом в клетки проникают гидрофобные вещества, полярные молекулы, мочевина, и не могут проникнуть белки, сахара, ионы и ДНК.
В процессе диффузии, молекулы стремятся заполнить весь доступный объем, а так же выровнять концентрацию по обе стороны мембраны. Бывает так, что мембрана непроницаема или плохо проницаема для вещества. В этом случае на нее воздействуют осмотические силы, которые могут как сделать преграду плотнее, так и растянуть ее, увеличив размеры насосных каналов.
Облегченная диффузия

Когда градиент концентрации не является достаточным основанием для транспорта вещества, на помощь приходят специфические белки. Они располагаются на мембране клеток точно так же, как и молекулы АТФ. Благодаря ним, может осуществляться как активный, так и пассивный транспорт.
Таким способом через мембрану проходят крупные молекулы (белки, ДНК), полярные вещества, к которым относятся аминокислоты и сахара, ионы. Благодаря участию белков скорость транспорта увеличивается в несколько раз, по сравнению с обычной диффузией. Но это ускорение зависит от некоторых причин:
- градиента вещества внутри и вне клетки;
- количества молекул-переносчиков;
- скорости связывания вещества и переносчика;
- скорости изменения внутренней поверхности мембраны клетки.
Несмотря на это, транспорт осуществляется благодаря работе белков-переносчиков, а энергия АТФ в данном случае не используется.
Основными чертами, которые характеризуют облегченную диффузию, являются:
- Быстрый перенос веществ.
- Избирательность транспорта.
- Насыщаемость (когда все белки заняты).
- Конкуренция между веществами (из-за сродства с белком).
- Чувствительность к специфическим химическим агентам – ингибиторам.
Осмос
Как уже упоминалось выше, осмос – это движение веществ по градиенту концентрации через полупроницаемую мембрану. Наиболее полно процесс осмоса описывает принцип Лешателье-Брауна. В нем говорится, что если на систему, находящуюся в равновесии, повлиять извне, то она будет стремиться вернуться в прежнее состояние. Первый раз с явлением осмоса столкнулись в середине XVIII столетия, но тогда ему не придали особого значения. Исследования феномена начались только сто лет спустя.
Самым важным элементом в феномене осмоса является полупроницаемая мембрана, которая пропускает через себя только молекулы определенного диаметра или свойств. Например, в двух растворах с разной концентрацией, через преграду будет проходить только растворитель. Это будет продолжаться до тех пор, пока концентрация с обеих сторон мембраны не станет одинаковой.
Осмос играет значительную роль в жизни клеток. Это явление позволяет проникать в них только тем веществам, которые необходимы для поддержания жизни. Красная клетка крови имеет мембрану, пропускающую только воду, кислород и питательные вещества, но белки, которые, образуются внутри эритроцита, не могут попасть наружу.
Явление осмоса нашло и практическое применение в быту. Даже не подозревая об этом, люди в процессе засаливания пищи использовали именно принцип движения молекул по градиенту концентрации. Насыщенный солевой раствор «вытягивал» на себя всю воду из продуктов, тем самым позволяя им дольше храниться.
fb.ru
Градиент концентрации Википедия
 При наличии градиента концентрации в растворе наблюдается диффузия — диффузионный поток растворённого вещества направлен против градиента концентрации.
При наличии градиента концентрации в растворе наблюдается диффузия — диффузионный поток растворённого вещества направлен против градиента концентрации. 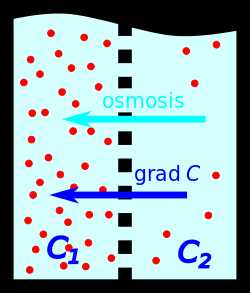 Если имеется полупроницаемая перегородка, пропускающая только молекулы растворителя (но не растворённого вещества), наблюдается осмос — поток растворителя, направлен по градиенту концентрации.
Если имеется полупроницаемая перегородка, пропускающая только молекулы растворителя (но не растворённого вещества), наблюдается осмос — поток растворителя, направлен по градиенту концентрации.Градиент концентрации или концентрационный градиент — это векторная физическая величина, характеризующая величину и направление наибольшего изменения концентрации какого-либо вещества в среде. Например, если рассмотреть две области с различной концентрацией какого-либо вещества, разделённые полупроницаемой мембраной, то градиент концентрации будет направлен из области меньшей концентрации вещества в область с большей его концентрацией. Вектор диффузионного потока направлен против вектора градиента концентрации, что, в соответствии с принципом Ле-Шателье, приводит со временем к уменьшению этого потока и градиента концентрации.
Определение
Градиент концентрации направлен по пути l{\displaystyle l}, соответствующему нормали к изоконцентрационной поверхности (полупроницаемой мембране). Значение градиента концентрации ∇C{\displaystyle \nabla C} равно отношению приращения концентрации dC{\displaystyle dC} к соответствующему приращению пути dl{\displaystyle dl}:
- ∇C=dCdl.{\displaystyle \nabla C={\frac {dC}{dl}}.}
При постоянном значении градиента концентрации C{\displaystyle C} на длине пути l{\displaystyle l}:
- ∇C=C1−C2l.{\displaystyle \nabla C={\frac {C_{1}-C_{2}}{l}}.}
Здесь C1{\displaystyle C_{1}} и C2{\displaystyle C_{2}} — начальное и конечное значение концентрации на длине пути l{\displaystyle l} (нормали к изоконцентрационной поверхности).
Градиент концентрации может быть причиной переноса веществ, например диффузии. Диффузия осуществляется против вектора градиента концентрации[источник не указан 762 дня].
Единицей измерения градиента концентрации в Международной системе единиц (СИ) является величина м−4 (моль/м4 или кг/м4), а также её дольные или кратные производные.
См. также
Литература
wikiredia.ru
Градиент концентрации
Химия — Градиент концентрации
01 марта 2011Градиент концентрации
Градиент концентрации или концентрационный градиент это векторная физическая величина, характеризующая величину и направление наибольшего изменения концентрации какого-либо вещества в среде. Например, если рассмотреть две области с различной концентрацией какого-либо вещества, разделенные полупроницаемой мембраной, то градиент концентрации будет направлен из области меньшей концентрации вещества в область с большей его концентрацией.
Определение
Градиент концентрации направлен по пути l , соответствующему нормали к изоконцентрационной поверхности. Значение концентрационного градиента gradC равно отношению элементарного изменения концентрации dC к элементарной длине пути dl :
При постоянном значении градиента концентрации C на длине пути l :
Здесь C1 и C2 начальное и конечное значение концентрации на длине пути l.
Градиент концентрации может быть причиной переноса веществ, например диффузии. Диффузия осуществляется против градиента концентрации.
Единицей измерения градиента концентрации является величина м, а также её дольные или кратные производные.
В научной литературе достаточно часто данный термин встречается в значении степени различия, то есть не векторной, а скалярной величины, показывающей разницу концентраций между двумя ограниченными областями, что является грубой ошибкой. В связи с этим, говоря, например, о пассивном транспорте, указывают, что он осуществляется по градиенту концентрации, имея ввиду по разности концентраций вещества, но это изменяет смысл термина, и потому такое его трактование неверно.
Просмотров: 3386
| Водородный электрод |
4108.ru
Концентрационный градиент — Справочник химика 21
Самопроизвольное разделение гомогенного раствора на две фазы в этом процессе представляется, на первый взгляд, неожиданным, поскольку в нем возникают концентрационные градиенты, а также фазовые границы, обладающие избыточной энергией. Статистическая трактовка, предложенная Онзагером [13, с. 456], вскрывает энтропийный характер коацервации. Вытянутые макромолекулы в растворе перекрываются сферами действия, в результате чего уменьшается свобода броуновского движения. Выделение части макромолекул в другую, более концентрированную фазу, значительно увеличивает свободу вращательного движения всех макромолекул, оставшихся в дисперсионной среде (мало изменяя ее для макромолекул коацервата), а следовательно и энтропию системы. [c.316]Обработка результатов. I. Рассчитывают значения мутности растворов т по формулам (124) и (125). Из величины т для каждого раствора вычитают мутность растворителя То. 2. Строят график зависимости показателя преломления растворов п от концентрации. Получают прямую линию. Определяют концентрационный градиент показателя преломления (1п/с1С как тангенс угла наклона этой прямой. 3. Используя полученное значение (1п/ 1С, рассчитывают постоянную Н по формуле (117). В качестве X берут значение эффективной длины волны света, пропускаемого светофильтром, который использовался при измерениях светорассеяния. 4. Рассчитывают значения функции НС/т—То для каждой концентрации. Результаты сводят в таблицу по следующей форме [c.163]
По мере утолщения образующихся при высокотемпературном окислении металлов пленок перемещение реагентов через них в преобладающем больщинстве случаев осуществляется диффузией (из-за наличия концентрационного градиента, созданного разностью химических потенциалов), которая часто и контролирует процесс окисления металлов, являющийся, таким образом, процессом реакционной диффузии (диффузии, при которой возникают или разлагаются химические соединения). Если исходить из преимущественной диффузии через окисную пленку кислорода (зона роста пленки при этом находится у поверхности раздела пленка—металл), то для скорости установившегося стационарного режима процесса можно написать уравнение [c.56]
Рост толщины слоев происходит благодаря диффузии металла или окислителя или их обоих, движущей силой которой является концентрационный градиент, созданный разностью химических потенциалов. [c.69]
Вагнера Диффузия под действием концентрационного градиента — — Окалина — — — [c.82]
Гомогенизация жидкостей. Число оборотов мешалки Пд а случае перемешивания в однофазной системе с целью снижения температурных и концентрационных градиентов может быть опреде/Тено из зависимости [c.253]
На рис. 90 приведены концентрационные градиенты (по Вагнеру) в окислах с избытком и недостатком металла при разных давлениях кислорода в предположении линейности распределения дефектов. [c.130]
Почти все существующие модели регенерации закоксованного слоя катализатора относятся к неподвижному слою [146, 147, 149, 150, 160-162]. В принципе полная математическая модель нестационарного процесса в слое катализатора учитывает продольный и радиальный перенос тепла и вещества в слое катализатора, а также наличие температурных и концентрационных градиентов внутри пористого зерна, т. е. включает в себя модель (4.15)-(4.16) [159]. Математическое описание такой модели представляется очень сложной системой дифференциальных уравнений в частных производных. Поэтому, чтобы математически моделировать такой сложный процесс, как регенерация катализатора, обычно прибегают к ряду упрощающих допущений. [c.83]
Возможность торможения в газовой пленке может быть грубо оценена также вычислением величины концентрационного градиента между свободным объемом реагента в реакционной среде и реагента на поверхности гранулы катализатора. Если он составляет величину, которая больше нескольких процентов от концентрации реагента в основной массе среды, то, вероятно, имеет место торможение в газовой пленке. Концентрационный градиент зависит от природы реагирующих веществ и степени турбулентности. Влияние турбулентности изменяется для различных систем, но для оценки его значения имеются достаточно точные экспериментальные данные. Эта оценка была предложена Уилером [8], который показал, что для реакции, которая полностью лимитируется диффузией, константа скорости составляет [c.51]
В уравнении (116) По и п—показатели преломления растворителя и раствора соответственно (1п/(1С — концентрационный градиент показателя преломления X—длина [c.157]
Флуктуации тем больше, чем меньше осмотическое давление, препятствующее им, поэтому т обратно пропорционально концентрационному градиенту осмотического давления. [c.158]
Обратная зависимость х от концентрационного градиента осмотического давления отвечает положению Эйнштейна, что флуктуации плотности в растворе тем больше, чем меньше осмотическое давление. [c.458]
В результате процесса термодиффузии концентрация примеси в одном из сосудов будет возрастать, что приводит к возникновению концентрационного градиента вдоль соединительной трубки. Последний вызывает обычную концентрационную диффузию, которая стремится выровнять состав смеси во всем объеме системы, вследствие чего примесь переносится по соединительной трубке в обратном направлении. Скорость этой концентрационной диффузии будет тем выше, чем больше разница в концентрациях примеси в сосудах / и. 2, и в соответствии с законом Фика может быть выражена соотношением [c.161]
При использовании неполяризующегося вспомогательного электрода и электролизера с малым омическим сопротивлением ом при потенциостатическом методе можно обеспечить постоянство потенциала исследуемого электрода. Все методы измерения перенапряжений, основанные на том или ином возмущении системы, можно еще подразделить на стационарные и переходные. В стационарных методах соблюдается не только стационарность переноса заряда, но и постоянство структуры поверхности электрода и примыкающих к нему областей в течение опыта. Должны оставаться постоянными концентрационные градиенты в электроде и в электролите. На твердых электродах в течение опыта должна быть постоянной и микроструктура поверхности. Последнее условие трудно соблюдать при электроосаждении или ионизации металлов. По этой причине весьма часто пользуются переходными методами, в которых измерения занимают достаточно короткое время и микроструктуру электрода можно считать постоянной. [c.39]
При протекании в жидкой фазе реакции между растворенным газообразным компонентом и поглотителем (или его активной составляющей) часть компонента переходит в связанное состояние и концентрация свободного компонента в жидкости понижается. Такое понижение приводит к увеличению концентрационного градиента и ускорению абсорбции в жидкой фазе по сравнению с физической абсорбцией. Это ускорение тем больше, чем выше скорость химической реакции, т. е. чем быстрее растворенные молекулы компонента переходят в связанное состояние. [c.129]
Если два необратимых процесса представляют теплопроводность и диффузию, коэффициент 12 соответствует термодиффузии. Это означает, что концентрационный градиент возникает в однородной смеси под влиянием температурного градиента. Заменяя потоки (3.3) их значениями в выражении (2.23) для источника энтропии, получим квадратичную форму [c.44]
Это соотношение является обобщением неравенства (11.26). Главная его особенность состоит в том, что неустойчивость, возникновение которой связано с обращением в нуль (11.98), зависит теперь от конкуренции не двух процессов, как в (11.26), а трех диссипации кинетической энергии, выделения энергии за счет выталкивающих сил, возникающих благодаря температурному и концентрационному градиентам. Каждый из последних двух эффектов может быть как стабилизирующим, так и дестабилизирующим. [c.171]
В настоящее время общепринят механизм окисления Вагнера, по которому основным фактором, определяющим скорость высокотемпературной коррозии, является интенсивность диффузии коррозионноактивных реагентов через оксидную пленку к металлу под воздействием концентрационного градиента. [c.11]
Распределение скоростей седиментации зависит от концентрационного градиента, который устанавливается в граничной области, и его изменения во времени. [c.114]
Для большинства газов DJ примерно равно средней длине свободного пробега и очень близко к 10 см при стандартных температуре и давлении (см. табл. VIII.3), так что в объеме 500 см гд 5 см) значение Р должно быть порядка 0,002/е мм рт. ст. или выше для того, чтобы диффузия имела значение для обрыва на стенках. Таким образом, если эффективность захвата радикала стенкой равна 1, то диффузия играет важную роль при давлениях выше 0,002 мм рт. ст. Однако если е = 10″, то это давление равно 20 мм рт. ст. или выше. Ниже этих давлений радикалы гибнут на стенках, но заметные концентрационные градиенты отсутствуют. [c.386]
Ионы Na» и 1 в реакции не участвуют. Распределение различных ионов в пленке показано на рис. V-8. Для каждого иона можно записать уравнение типа уравнения (1,31), выражающее скорость переноса этого иона как функцию от подвижностей и локальных концентраций и концентрационных градиентов всех присутствующих ионов. Для упрощения принято, что градиенты концентрации неизменны (например, для иона он равен р/б во всех точках), а значения концентрации каждого иона в уравнении (1,31) взяты усредненными в пленке, например р/2 — для Н +. Таким образом, можно записать четыре уравнения типа (1,31) для скоростей переноса всех четырех участвующих ионов, выраженных через концентрации т, п, р, q, S, толщины пленок б и б и подвижности ионов. Учитывая, что Ru+ = R — = —Roh- = (скорость абсорбции НС1) и i Na+ = о, можно избавиться от неизвестных т, s и б и получить выражение для Rb/p через подвижности ионов и qln и qlp. Скорость физической абсорбции хлористого водорода водой с той же толщиной пленки б была бы pDh i/6 отсюда коэффициент ускорения Е, показывающий, во сколько раз реакция ускоряет абсорбцию, выражается отношением R8Ip)IDh i- [c.143]
В частном случае, когда лимитирующей кинетической стадией является внешний перенос свободной влаги от материала к окружающей среде, температурный и концентрационный градиенты внутри материала обычно невелики. В этом случае температура материала может приниматься постоянной и равной температуре мокрого термометра, а процесс сушки рассматриваться как конвективный теплоперепос. В этих условиях постулируют, что количество удаленной влаги определяется количеством переданного тепла. Этот период сушки обычно называют периодом постоянной скорости сушки (или первым периодом). Продолжительность периода постоянной скорости обычно рассчитывается по уравнениям теплового баланса (для этого достаточно высоты слоя в 300—400 мм) или по уравнениям теплообмена. В последнем случае коэффициенты теплоотдачи могут быть определены по специальным расчетным формулам (см., например, гл. X этой книги или монографию Гельперина с соавт. ). [c.514]
Электродвижущая сила этого элемента Етв. возникает при уменьшении свободной энергии Абг реакции окисления металла, что приводит к появлению концентрационного градиента, вызывающего диффузию (градиент поля, приводящий к миграции заряженных частиц, по Вагнеру, не возникает из-за равномерного распределения положительных и отрицательных зарядов в объеме окисла). На поверхности раздела металл — пленка протекает анодная реакция по фор- Ме Пленпа Газ муле (44) [c.61]
Для определения кинетики необходимо использовать безгра-диентные или проточно-циркуляционные установки [39, 41, 121], позволяющие проводить реакцию в течение длительного времени, достаточного для того, чтобы образец катализатора достиг стационарных условий. Циркуляционный реактор похож на дифференциальный, но за счет внешней или внутренней циркуляции газа температурные и концентрационные градиенты по слою катализатора, обусловленные протеканием реакции, сводятся к минимуму. Как дифференциальный, так и циркуляционный реакторы применяют для того, чтобы добиться изотермического режима. Но на практике к нему приближается только циркуляционный реактор. Заметим, что для измерений истинной кинетики необходимо вместо таблеток использовать очень мелкие частицы катализатора с тем, чтобы свести к минимуму влияние массопереноса. [c.260]
Случай I. Отсутствуют температурные и концентрационные градиенты внутри и вне гранулы катализатора. Исиользуегся уравнение модели (21) — (25). [c.112]
Существуют температурные и концентрационные градиенты в пределах и в 1Ю1ран№шом слое гранулы катализатора. Предполагаем справедливой двухфазную модель. Они интегрируются последовательно с учетом соотаетствующих граничных условий. Первоначально интегрируются уравнения модели зерна (1)-(3) для задачи Неймана с использованием инвариантных соотношений (10)-(13). Реализуется следующий алгоритм [c.117]
Так как концентрационные градиенты в порах катализатора или адсорбента часто влияют на скорость реакции, следует привести данные, позволяющие оценить роль этого эффекта. Необходи.мо обсудить диффузионные ограничения. [c.342]
Некоторые результаты расчетов нестационарных режимов промышленного реактора окисления о-ксилола представлены на рис. 3.52. Они уточняют выводы по квазигомогенной модели. Из рис. 3.52 видно, что температурные градиенты в зерне незначительны, т.е. зерно изотермично. В противоположность этому имеются значительные концентрационные градиенты в зерне, указывающие на сильное влияние внутренней диффузии. Разности концентрации в зерне наиболее значительны в передних горячих участках слоя катализатора и уменьшаются с понижением температуры вследствие более сильного охлаждения по сравнению с тепловыделением в последних участках слоя. Разности концентрации и температуры между наружной поверхностью зерна и газовой фазой в общем были малы, только в горячей точке разность температуры превышала 5° по оси реактора. [c.173]
В точке С сектора III ситуация довольно неожиданная несмотря на то, что плотность уменьшается при переходе снизу вверх, система неустойчива. Этот пример показывает, что простая механическая интерпретация возникновения неустойчивости, данная нами в разд. 11.3, здесь неприменима. Теперь в (11,98) третий член дестабилизирующий (отрицательный), тогда как первые два—стабилизирующие (положительные). Неустойчивость является следствием того, что производство энергии выталкивающими силами, возникающими из-за концентрационных градиентов, превалирует над диссипацией энергии в двух первых эффектах. В результате появляется свободная конвекция. Однако возникающее при этом ячеечное движение совершенно отлично от того, которое наблюдается в однокомпонентной проблеме Бенара (подробно этот вопрос рассмотрен в работе [167]). [c.172]
Энергия, освобождающаяся при окислении субстратов и последующем переносе электронов в дыхательной цепи, используется не только на синтез АТФ, но и для осуществления других функций митохондрий, например для активного транспорта ионов a + через митохондриальную мембрану. Если к суспензии аэробно инкубируемых митохондрий в присутствии субстрата добавить некоторое количество ионов a + (в виде какой-либо его соли), то по истечении небольшого промежутка времени весь добавленный Са + оказывается во внутримитохондриальном пространстве. В процессе активного транспорта создается и поддерживается высокий концентрационный градиент ионов Са + по обе стороны митохондриальной мембраны. Когда функционирование дыхательной цепи полностью блокировано, транспорт может обеспечиваться за счет энергии гидролиза АТФ. [c.449]
К числу наиболее хорощо охарактеризованных эндергонических реакций митохондрий относятся наряду с окислительным фосфорилированием АДФ восстановление НАД+ сукцинатом, трансгидрогеназная реакция и перенос ионов Са + против концентрационного градиента. Изучение кинетических взаимоотнощений таких реакций при их одновременном протекании важно не только для понимания механизмов регуляции метаболизма митохондрий в целом, но и для выяснения механизмов, лежащих в основе любого из путей трансформации энергии в митохондриях. [c.469]
chem21.info