
Клетки нейроглии – Нейроглия — Википедия
Глиальная клетка. Функции и особенности глиальных клеток
Нервная система состоит не только из нейронов и их отростков. На 40 % она представлена глиальными клетками, которые играют важную роль в ее жизнедеятельности. Они буквально ограничивают головной мозг и нервную систему от остальных сред организма и обеспечивают его автономную работу, что действительно важно для человека и других животных, имеющих центральную нервную систему. Причем клетки нейроглии способны делиться, что отличает их от нейронов.
Общее понятие о нейроглии
Совокупность глиальных клеток называется нейроглией. Это особые клеточные популяции, которые находятся в центральной нервной системе и на периферии. Они поддерживают форму головного и спинного мозга, а также снабжают его питательными веществами. Известно, что в центральной нервной системе из-за наличия гематоэнцефалического барьера нет иммунных реакций. Однако при попадании чужеродного антигена в головной или спинной мозг, а также в ликвор глиальная клетка, редуцированный аналог макрофага периферических тканей, фагоцитирует его. Более того, именно отделение мозга от периферических тканей обеспечивает нейроглия.
Иммунная защита мозга
Мозг, где протекает множество биохимических реакций, а значит, образуется масса иммуногенных веществ, должен быть защищен от гуморального иммунитета. Важно понимать, что нейрональная ткань мозга очень чувствительна к повреждениям, после которых нейроны восстанавливаются лишь частично. Значит, появление места в центральной нервной системе, где будет проходить местная иммунная реакция, повлечет и гибель некоторых окружающих клеток либо демиелинизацию отростков нейронов.
На периферии тела это повреждение соматических клеток вскоре заполнится вновь образованными. А в мозге восстановить функцию потерянного нейрона невозможно. И именно нейроглия ограничивает головной мозг от контакта с иммунной системой, для которой центральная нервная система – это огромнейшее количество чужеродных антигенов.
Классификация глиальных клеток
Глиальные клетки делятся на два вида в зависимости от морфологии и происхождения. Выделяют клетки микроглии и макроглии. Первый вид клеток берет свое начало от мезодермального листка. Это мелкие клетки с многочисленными отростками, способные фагоцитировать твердые вещества. Макроглия — это производное эктодермы. Глиальная клетка макроглии делится на несколько видов в зависимости от морфологии. Выделяют эпендимальные и астроцитарные клетки, а также олигодендроциты. Такие виды клеточных популяций также делятся на несколько типов.
Эпендимальная глиальная клетка
Эпендимальные глиальные клетки встречаются в специфических участках центральной нервной системы. Они образуют эндотелиальную выстилку мозговых желудочков и центрального спинномозгового канала. Свое начало в эмбриогенезе они берут из эктодермы, а потому представляют собой особый вид нейроэпителия. Он многослойный и выполняет ряд функций:
- опорная: составляет механический каркас желудочков, который также поддерживается за счет гидростатического давления ликвора;
- секреторная: выделяет в ликвор некоторые химические вещества;
- разграничительная: отделяет мозговое вещество от ликвора.
Виды эпендимоцитов
Среди эпендимоцитов есть и свои виды. Это эпендимоциты 1-го и 2-го порядка, а также танициты. Первые образуют начальный (базальный) слой эпендимальной оболочки, а эпендимоциты лежат вторым слоем над ними. Важно, что эпендимальная глиальная клетка 1-го порядка участвует в образовании гематоглифического барьера (между кровью и внутренней средой желудочков). Эпендимоциты 2-го порядка имеют ворсинки, ориентированные в сторону тока ликвора. Также существуют танициты, которые представляют собой рецепторные клетки.

Они находятся в латеральных участках дна 3-го мозгового желудочка. Имея микроворсинки на апикальной стороне и один отросток на базальной, они могут передавать информацию нейронам о составе ликворной жидкости. При этом сам ликвор через небольшие многочисленные щелевидные отверстия между эпендимоцитами 1-го и 2-го порядка может попадать непосредственно к нейронам. Это позволяет говорить, что эпендима представляет собой особый вид эпителия. Его функциональный, но не морфологический аналог на периферии тела – эндотелий кровеносных сосудов.
Олигодендроциты
Олигодендроциты – это типы глиальных клеток, которые окружают нейрон и его отростки. Они встречаются как в центральной нервной системе, так и рядом с периферическими смешанными и вегетативными нервами. Сами олигодендроциты представляют собой полигональные клетки, оснащенные 1-5 отростками. Ими они сцепляются между собой, изолируя нейрон от внутренней среды организма и обеспечивая условия для нервного проведения и генерации импульсов. Существует три вида олигодендроцитов, которые различаются по морфологии:
- центральная клетка, расположенная около тела мозгового нейрона;
- сателитная клетка, окружающая тело нейрона в периферическом ганглии;
- шванновская клетка, охватывающая нейрональный отросток и образующая его миелиновую оболочку.

Олигодендроцитарные глиальные клетки встречаются как в головном и спинном мозге, так и в периферических нервах. Причем пока неизвестно, чем отличается сателитная клетка от центральной. Учитывая, что генетический материал у всех клеток организма, кроме половых, одинаков, то, вероятно, эти олигодендроциты могут взаимно заменять друг друга. Функции олигодендроцитов следующие:
- опорная;
- изолирующая;
- разделительная;
- трофическая.

Астроциты
Астроциты – это глиальные клетки мозга, которые составляют мозговое вещество. Они имеют звездчатую форму и отличаются небольшими размерами, хотя они больше, чем клетки микроглии. При этом существует всего два типа астроцитов: волокнистый и протоплазматический. Первый вид клеток расположен в белом и сером веществе головного мозга, хотя их значительно больше в белом.

Это значит, что они наиболее распространены в тех участках, где есть значительное число нейрональных миелинизированных отростков. Протоплазматические астроциты – это также глиальные клетки: встречаются в белом и сером веществе мозга, но их значительно больше в сером. Значит, их функцией является создание опоры для тел нейронов и структурная организация гематоэнцефалического барьера.

Микроглия
Микроглиальные клетки – это последний вид нейроглии. Однако в отличие от всех других клеток центральной нервной системы они имеют мезодермальное происхождение и представляют собой особые типы моноцитов. Их предшественниками являются стволовые кровяные клетки. Из-за особенностей строения нейронов и их отростков за иммунные реакции в центральной нервной системе отвечают как раз глиальные клетки. И их функции практически аналогичны таковым у тканевых макрофагов. Они ответственны за фагоцитоз и распознавание и презентацию антигена.

Микроглия содержит особые виды глиальных клеток, которые имеют рецепторы кластеров дифференцировки, что подтверждает их костномозговое происхождение и реализацию иммунных функций в ЦНС. Также они ответственны за развитие демиелинизирующих заболеваний, болезни Альцгеймера и синдрома Паркинсона. Однако сама клетка – это лишь способ реализации патологического процесса. Потому, вероятно, когда удастся найти механизм активации микроглии, будут пресечены процессы развития данных болезней.
fb.ru
Нейроглия (глия): общие сведения
Нейроглия (глия): общие сведения
Помимо нейронов нервная ткань содержит клетки еще одного типа — клетки глии, глиальные клетки, или глия (от греч. «глия» — клей). Они выполняют опорную и защитную функции , а также участвуют в нейронофагии. По численности их в 10 раз больше, чем нейронов (10 в 13-ой и 10 в 12-ой степени, соответственно) и они занимают половину объема центральной нервной системы (ЦНС). Глиальные клетки окружают нервные клетки и играют вспомогательную роль Глиальные клетки более многочисленные, чем нейроны: составляют по крайней мере половину объема ЦНС ( рис. 1-18 ).
Существует несколько типов глии. Так одни глиальные клетки участвуют в поддержании состава межклеточной среды вокруг нейронов , другие образуют миелиновую оболочку вокруг аксонов , благодаря которой увеличивается скорость проведения потенциалов действия. Следовательно, не принимая прямого участия в краткосрочных коммуникативных процессах в нервной системе, клетки нейроглии способствуют осуществлению этой функции нейронами.
Таким образом, глия не только выполняет опорные функции, но и обеспечивает многообразные метаболические процессы в нервной ткани, а также способствует восстановлению нервной ткани после травм и инфекций.
Между нейронами и глиальными клетками существуют сообщающиеся между собой щели размером 15-20 нм, так называемое интерстициальное пространство , занимающее 12-14% общего объема мозга.
Глиальные клетки невозбудимы: во время деполяризации глиальных клеток проводимость их мембран не повышается.
Клетки нейроглии делятся на несколько типов. Клетки эпендимы выстилают желудочки головного мозга и спинномозговой канал и образуют эпителиальный слой в сосудистом сплетении . Они соединяют желудочки с нижележащими тканями.
Клетки макроглии делятся на две категории - астроциты и олигодендроциты .
Протоплазматические астроциты локализованы в сером веществе; от тела клетки, содержащей овальное ядро и большое количество гликогена , отходят сильно разветвленные короткие и толстые отростки.
Фибриллярные астроциты локализованы в белом веществе . Ядро у них также овальное, и тело клетки содержит много гликогена , но отростки длинные и менее разветвленные, некоторые ветви буквально упираются в стенки кровеносных сосудов. Эти клетки переносят питательные вещества из крови в нейроны.
Астроциты двух типов взаимосвязаны и образуют обширное трехмерное пространство, в которое погружены нейроны. Они часто делятся, образуя в случае повреждений центральной нервной ситемы рубцовую ткань.
Олигодендроциты локализованы в сером и белом веществе. Они мельче астроцитов и содержат одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек, а само оно содержит цитоплазму с большим количеством рибосом. Шванновские клетки — это специализированные олигодендроциты, синтезирующие миелиновую оболочку миелинизированных волокон .
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. От каждого конца маленького продолговатого тела клетки, содержащей лизосомы и хорошо развитый аппарат Гольджи , отходит по толстому отростку. От всех его ветвей отходят более мелкие боковые веточки. При повреждении мозга эти клетки превращаются в фагоциты и, перемещаясь при помощи амебоидного движения, противостоят вторжению чужеродных частиц.
Глия является системой трофического обеспечения нервной системы , а также принимает активное участие в специфическом функционировании нервной ткани: в норме тормозит гиперактивность нейронов , способствует активному поглощению из синаптической щели и утилизации медиаторов и других агентов, участвующих в повреждении нейронов. В условиях ишемии микроглиальные клетки индуцируют синтез не только нейротоксичных веществ, но и сигнальных молекул, клеточных регуляторов, трофических факторов, способствующих выживаемости нейронов и уменьшающих процессы постишемического рубцевания
Микроглия — единственный иммунокомпетентный компартмент в центральной нервной системе
В ЦНС к нейроглии относятся астроциты и олигодендроциты, а в периферической нервной системе — шванновские клетки и клетки-сателлиты .
Клетки микроглии и эпендимы считаются центральными глиальными клетками ( рис. 32.7 , рис. 32.10 ).
Ссылки:
Все ссылкиГлиальные клетки (нейроглия) // Дуглас Филдз ≪ Scisne?
В недавно опубликованной книге «Облава на м-ра Альберта» (Driving Mr. Albert) рассказана подлинная история патологоанатома Томаса Харви, который в 1955 г. произвёл вскрытие Альберта Эйнштейна. Выполнив работу, Харви самым непочтительным образом забрал мозг учёного домой, где в течение 40 лет хранил его в пластиковой банке с дезинфицирующей жидкостью. Время от времени патологоанатом отдавал маленькие срезы мозговой ткани исследователям из разных частей света, пытавшимся выяснить причины гениальности Эйнштейна. Когда Харви перевалило за 80, он погрузил остатки мозга в багажник своего «Бьюика» и повёз возвращать их внучке гения.
Одной из тех, кто изучал срезы мозговой ткани Эйнштейна, была Мэриан Даймонд (Marian C. Diamond) — авторитетный гистолог из Калифорнийского университета в Беркли. Она установила, что числом и размерами нервных клеток (нейронов) головной мозг великого физика ничем не отличается от мозга обычного человека. Но в ассоциативной области коры, ответственной за высшие формы мыслительной деятельности, Даймонд обнаружила необычайно большое количество вспомогательных элементов нервной ткани — клеток нейроглии (глии). В мозге Эйнштейна их концентрация была намного больше, чем в голове среднестатистического Альберта.
Обзор: ГЛИЯ
|
Любопытное совпадение? Возможно. Но сегодня учёные получают всё больше данных, указывающих на то, что глиальные клетки играют гораздо более важную роль в деятельности мозга, чем предполагалось ранее. Долгие десятилетия всё внимание физиологов было сосредоточено на нейронах — главных, по их мнению, приёмопередатчиках мозга. Хотя глиальных клеток в 9 раз больше, чем нейронов, учёные отводили им скромную роль элементов, поддерживающих жизнедеятельность мозга (транспорт питательных веществ из кровеносных сосудов в нейроны, поддержание нормального баланса ионов в мозге, обезвреживание болезнетворных микробов, ускользнувших от преследования иммунной системы, и т. д.). А тем временем нейроны, поддерживаемые глией, были вольны общаться друг с другом через крошечные контактные точки (синапсы) и формировать сложнейшие сети соединений, благодаря которым мы думаем, вспоминаем прошлое или испытываем радость.
Неизвестно, как долго просуществовала бы ещё такая модель устройства мозга, если бы не недавно обнаруженные факты, свидетельствующие о том, что на протяжении всей жизни человека (от периода эмбрионального развития и до глубокой старости) нейроны и глия ведут весьма оживлённый диалог. Глия влияет на образование синапсов и помогает мозгу определять, какие нервные связи усиливаются или ослабевают с течением времени (эти изменения напрямую связаны с процессами общения и долгосрочной памяти). Последние исследования показали, что глиальные клетки общаются и друг с другом, влияя на деятельность мозга в целом. Нейробиологи с большой осторожностью наделяют глию новыми полномочиями. Однако можно вообразить, какое волнение они испытывают при мысли о том, что большая часть нашего мозга почти не изучена и, следовательно, может ещё раскрыть множество тайн.
Глиальные клетки общаются с нейронами
Мы представляем себе нервную систему в виде переплетения проводов, соединяющих нейроны. Каждый нейрон снабжён одним длинным отростком — аксоном, переносящим электрические сигналы от тела нейрона к расширенным участкам на его конце — аксонным терминалям. Каждая терминаль высвобождает в синаптическую щель молекулы химического посредника — нейротрансмиттера, которые достигают соответствующих рецепторов на коротких ветвящихся отростках (дендритах) соседнего нейрона. Пространства между нейронами и аксонами заполнены массой разнообразных клеток глии. К тому времени, как скончался Эйнштейн, нейробиологи уже подозревали, что глиальные клетки принимают участие в переработке информации, но доказательств у них не было. В конце концов они оставили глию в покое.
Причина того, что учёные не смогли обнаружить обмен сигналами между глиальными клетками, отчасти была связана с несовершенством методик. Но главными виновниками неудач были сами исследователи, ошибочно считавшие, что если клетки глии наделены способностью к общению, то обмениваться информацией они должны точно так же, как и нейроны, — с помощью электрических сигналов. Предполагалось, что клетки глии тоже должны генерировать электрические импульсы (потенциалы действия), стимулирующие выброс в синаптическую щель нейротрансмиттеров, которые, в свою очередь, вызывают импульсы в других клетках. Исследователи обнаружили, что глиальные клетки обладают несколькими типами ионных каналов, ответственных за генерирование электрических сигналов в аксонах, но они предположили, что эти каналы нужны глии просто для того, чтобы чувствовать уровень активности соседних нейронов. Было установлено, что мембрана глиальных клеток не обладает свойствами, необходимыми для проведения потенциалов действия. Нейробиологи, однако, упустили из виду одно обстоятельство, которое удалось обнаружить только благодаря современным методам исследования: глиальные клетки передают друг другу сообщения с помощью химических, а не электрических сигналов.
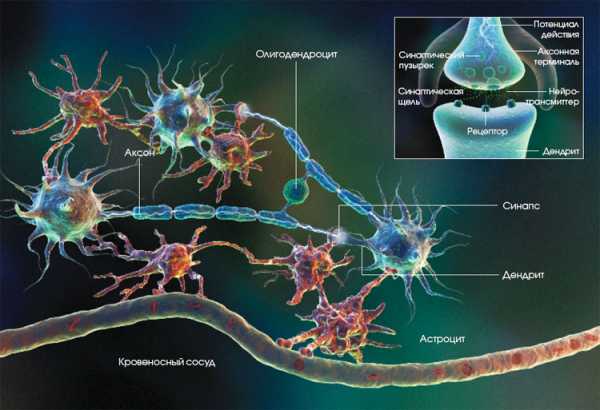 Глия и нейроны работают в головном и спинном мозге согласованно. Нейрон посылает по аксону сигнал, который через синаптическую щель достигает дендрита другой нервной клетки. Астроциты поставляют нейронам питательные вещества, а также окружают синапсы и регулируют их деятельность. Олигодендроциты вырабатывают миелин и образуют вокруг аксонов изолирующие миелиновые оболочки… |
Важный вклад в понимание механизмов, позволяющих глии распознавать нейронную активность, был сделан в середине 1990-х гг., когда учёные обнаружили в мембранах глиальных клеток рецепторы, реагирующие на разнообразные химические вещества, включая и нейротрансмиттеры. Это открытие навело их на мысль, что клетки глии способны общаться друг с другом с помощью сигналов, которые не распознаются нервными клетками.
Экспериментально было установлено, что показателем активации глиальных клеток служит поглощение ими кальция. На основании этого наблюдения учёные разработали метод, позволяющий визуально определять, обладают ли терминальные шванновские клетки (один из типов глиальных клеток, окружающих синапсы в области контакта нервов с мышечными клетками) чувствительностью к нервным сигналам, приходящим к этим синапсам. Было показано, что шванновские клетки действительно реагируют на синаптические импульсы и что такая реакция сопровождается проникновением в них ионов кальция.
Но ограничивается ли участие глии в нервных процессах только «подслушиванием» нейронных переговоров? Ведь шванновские клетки окружают аксоны как в области синапсов, так и по ходу нервов в разных частях тела, а глиальные клетки другого типа (олигодендроциты) образуют оболочки вокруг аксонов в центральной нервной системе (т. е. в головном и спинном мозге). Сотрудники лаборатории Национального института здравоохранения решили выяснить, способна ли глия отслеживать и нервные сигналы, распространяющиеся по аксонам в нервных цепях. И если такое общение между глией и нейронами существует, какие механизмы лежат в его основе и, что ещё важнее, как влияют на работу глиальных клеток «подслушанные» ими нервные сообщения?
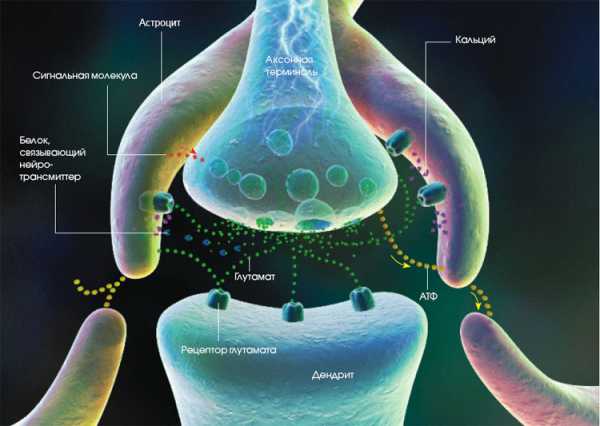 Астроциты регулируют синаптическую передачу сигнала несколькими способами. Аксон передаёт нервный сигнал дендриту за счёт выброса нейротрансмиттера (обозначен зелёным цветом) — в данном случае глутамата. Кроме того, аксон высвобождает АТФ (жёлтый). Эти соединения вызывают перемещение кальция (фиолетовый) внутрь астроцитов, что побуждает их вступить в общение друг с другом за счёт высвобождения собственного АТФ… |
Чтобы ответить на эти вопросы, мы культивировали сенсорные нейроны (клетки дорсально-корешкового ганглия, ДКГ) мыши в специальных лабораторных чашках с электродами, с помощью которых можно было вызывать потенциалы действия в аксонах. В одни чашки с нейронами мы добавили шванновские клетки, в другие — олигодендроциты. Необходимо было одновременно контролировать активность и аксонов, и глии. За активностью нервных и глиальных клеток мы следили визуально, вводя в них краситель, который при связывании с ионами кальция должен был флуоресцировать. Когда по аксону пробегает нервный импульс, потенциалозависимые ионные каналы в нейронной мембране открываются, и ионы кальция проникают в клетку. Следовательно, распространение импульсов по аксонам должно сопровождаться зелёными вспышками внутри нейронов. По мере роста концентрации кальция в клетке флуоресценция должна становиться ярче. Её интенсивность можно измерить с помощью фотоэлектронного умножителя, а искусственно окрашенные изображения светящейся клетки воспроизвести в реальном времени на экране монитора. Если глиальные клетки реагируют на нервные сигналы и поглощают в это время ионы кальция из окружающей среды, они тоже должны засветиться — только немного позднее, чем нейроны.
Сидя в затенённой комнате и напряжённо вглядываясь в экран монитора, мы с биологом Бетом Стивенсом (Beth Stevens) собирались приступить к эксперименту, на подготовку которого у нас ушло несколько месяцев. На включение стимулятора нейроны ДКГ тут же отреагировали изменением цвета: по мере увеличения концентрации кальция в их аксонах они превратились из синих в зелёные, затем — в красные и, наконец, побелели. Поначалу ни в шванновских клетках, ни в олигодендроцитах никаких изменений не обнаружилось, но спустя 15 долгих секунд они, подобно ёлочным лампочкам, начали загораться. Каким-то неведомым образом клетки глии почувствовали, что по аксонам пробегают импульсы, и отреагировали на это событие увеличением концентрации кальция в цитоплазме.
Глиальные клетки общаются друг с другом
Нам удалось показать, что глия способна распознавать импульсную активность в аксонах, реагируя на неё поглощением кальция. В нейронах он активирует ферменты, ответственные за выработку нейротрансмиттеров. Вполне вероятно, что поступление кальция в глиальные клетки также вызывает активацию ферментов, связанных с развитием какой-то реакции. Но какой?
Кадры фильма (окраска искусственная), отснятого с помощью лазерной конфокальной микроскопии, показывают, что глиальные клетки способны реагировать на обмен сигналами между нейронами. Сенсорные нейроны (две крупные клетки диаметром 20 микрон) (a) и более мелкие шванновские клетки были помещены в культуральную среду, содержащую ионы кальция. В клетки вводился краситель, который при связывании ионов кальция начинал флуоресцировать. Стимуляция нейронов электрическим током небольшого напряжения заставляла их генерировать распространяющиеся по аксонам (длинные линии) импульсы (потенциалы действия)… |
Исследование ещё одного типа глиальных клеток — астроцитов, транспортирующих питательные вещества из капилляров в нервные клетки и поддерживающих оптимальный уровень ионов, необходимый для генерирования нервных импульсов в окружающей нейроны среде (включая и удаление избытка нейротрансмиттеров и ионов, высвобождаемых нейронами во время импульсации), поможет ответить на этот вопрос. В 1990 г. Стивен Смит из Йельского университета показал, что если в культуру астроцитов добавить нейротрансмиттер глутамат, концентрация кальция в клетках резко возрастает. Клетки ведут себя так, словно только что произошёл выброс нейротрансмиттера из нейрона и они горячо обсуждают друг с другом вызвавшую его импульсацию нейронов.
Некоторые нейробиологи пытались выяснить, не явилось ли общение глиальных клеток следствием простого перемещения ионов кальция или связанных с ним сигнальных молекул из одного астроцита в соседний через соединяющие их открытые ворота. В 1996 г. Бен Кейтер (Ben Kater) из Университета штата Юта опроверг это предположение. С помощью острого микроэлектрода он разрезал слой астроцитов в культуре на две части, оставив между ними зазор, не содержавший клеток и разделявший популяцию астроцитов. Когда концентрация кальция в клетках по одну сторону разреза возрастала, то же происходило и по другую сторону. Таким образом выяснилось, что астроциты посылали друг другу сигналы через внеклеточную среду.
АТФ как химический посредник
Выявленные закономерности привели исследователей в замешательство. Коммуникация глиальных клеток, как и нейронов, контролируется токами кальция. Однако если изменения его уровня в нейронах вызывают электрические импульсы, то в глии — нет. Возникает вопрос: не было ли инициировано перемещение ионов кальция в глию каким-то другим электрическим феноменом? А если нет, то какова природа механизма?
Когда учёные экспериментировали с глией, в поле зрения им постоянно попадала знакомая всем молекула аденозинтрифосфата (АТФ). Будучи основным источником энергии в живых клетках, АТФ обладает многими признаками, благодаря которым прекрасно подходит на роль химического посредника между клетками. В окружающей среде он содержится в больших количествах, а во внеклеточном пространстве его мало. Благодаря небольшим размерам молекула способна к быстрой диффузии и легко разрушается ферментами. Более того, АТФ присутствует в аксонных терминалях, где обычно и хранятся молекулы нейротрансмиттеров, и может высвобождаться в синаптическую щель.
В 1999 г. Питер Гатри (Peter B. Guthrie) и его сотрудники из Университета штата Юта показали, что при возбуждении астроциты выбрасывают в окружающую среду АТФ. Затем он связывается рецепторами на соседних астроцитах, заставляя открываться ионные каналы и способствуя перемещению кальция внутрь клеток. В свою очередь, повышение уровня кальция в клетках заставляет их высвобождать во внеклеточную среду новые порции АТФ — так в популяции астроцитов инициируется цепная реакция, связанная с изменением внутриклеточного уровня кальция и опосредованная АТФ.
Как общаются глиальные клетки? В культуральную среду, содержавшую кальций, помещались астроциты (а) и сенсорные нейроны. После того как под влиянием электрической стимуляции нейроны принялись генерировать распространяющиеся по аксонам (зигзаги молний) (b) импульсы (потенциалы действия), глия начала флуоресцировать — признак того, что глиальные клетки отреагировали на это событие поглощением кальция. Спустя 10 и 12,5 секунд (с и d) по всей популяции астроцитов прокатились две огромные волны проникновения кальция внутрь клеток. О росте концентрации кальция в астроцитах свидетельствует изменение их цвета: вначале они были зелёными, затем стали синими и наконец красными. |
В результате наблюдений родилась модель, позволившая объяснить способность околоаксонной глии распознавать нейронную активность, и передавать затем сообщения другим глиальным клеткам, окружающим синапс. Импульсация нейронов побуждает глиальные клетки, окружающие аксон, высвобождать АТФ, который вызывает поглощение кальция соседними глиальными клетками. Это стимулирует выброс новых порций АТФ, что активирует передачу сообщения по длинной цепочке глиальных клеток иногда на значительное расстояние от нейрона, инициировавшего всю последовательность данных событий. Но каким образом глиальным клеткам, участвовавшим в нашем эксперименте, удавалось распознавать импульсацию нейронов — ведь аксоны не образуют с глией синаптических контактов и в области синапсов не было никаких глиальных клеток? Участием нейротрансмиттеров феномен объяснить нельзя: из аксонов они не диффундируют. Быть может, его причиной был АТФ, способный каким-то образом просачиваться из аксонов?
Для проверки гипотезы мы решили провести электрическую стимуляцию чистых культур аксонов ДКГ и последующий химический анализ культуральной среды. Воспользовавшись ферментом, ответственным за свечение брюшка у жуков-светляков (эта реакция требует участия АТФ), мы наблюдали свечение среды во время распространения импульса по аксонам, что свидетельствовало о высвобождении из них АТФ. Затем мы добавили в культуру шванновские клетки, также начинавшие светиться после того, как по аксонам пробегали потенциалы действия. Но когда мы добавили в среду фермент апиразу, быстро разрушающую АТФ и не дающую ему достигнуть шванновских клеток, глия во время импульсации аксонов оставалась тёмной. Таким образом, содержание кальция в шванновских клетках не менялось, т. к. они не получали АТФ-сигнала.
АТФ, высвободившийся из аксонов, на самом деле стимулировал транспорт кальция внутрь шванновских клеток. С помощью биохимического анализа и цифровой микроскопии нам удалось показать, что в результате этого события сигнальные молекулы перемещаются от клеточной мембраны к ядру и включают здесь различные гены. Таким образом, мы обнаружили поразительный факт: генерируя импульсы, призванные обеспечить общение с другими нейронами, нервная клетка и её аксон могут влиять на считывание генов в глиальной клетке и изменять тем самым её поведение.
Аксоны определяют судьбу глиальных клеток
Какие функции глии могут контролировать гены, включённые АТФ? Приказывают ли они глиальным клеткам действовать таким образом, чтобы повлиять на окружающие их нейроны? Стивенс попытался ответить на вопрос, обратив внимание на процесс, способствующий образованию миелиновой изолирующей оболочки вокруг аксонов. Благодаря ей аксоны способны проводить нервные импульсы с огромной скоростью на значительные расстояния. Её образование позволяет малышу всё крепче удерживать голову в вертикальном положении, а разрушение вследствие некоторых болезней (например, рассеянного склероза) превращает человека в инвалида.
Мы решили выяснить, как незрелая шванновская клетка, расположенная на аксоне в периферической нервной системе плода или младенца, узнаёт, нуждается ли отросток в миелинизации и когда нужно приступить к его «пеленанию» миелином. Или, напротив, следует ли ей превратиться в клетку, которая не будет сооружать миелиновую оболочку? Вообще говоря, в миелине нуждаются только аксоны большого диаметра. Могут ли аксональные нервные импульсы или высвобождение АТФ влиять на выбор шванновской клетки? Мы обнаружили, что шванновские клетки в культуре пролиферировали медленнее в том случае, когда окружали импульсирующие, а не молчащие аксоны. Более того, они приостанавливали своё развитие и прекращали выработку миелина. Добавление АТФ вызывало такие же эффекты.
А Витторио Галло (Vittorio Gallo) из соседней лаборатории НИЗ, изучая олигодендроциты, образующие миелиновые оболочки вокруг аксонов в головном мозге, обнаружил совершенно иную картину. АТФ не угнетал пролиферацию клеток, но аденозин (вещество, в которое превращается молекула АТФ после отщепления от неё остатков фосфорной кислоты) стимулировал созревание клеток и выработку миелина.
Понимание механизмов миелинизации имеет важнейшее значение. Болезни, сопровождающиеся разрушением миелиновой оболочки, каждый год уносят тысячи человеческих жизней и вызывают паралич и слепоту. Неизвестно, какой фактор инициирует миелинизацию, но аденозин стал первым веществом «аксонального происхождения», у которого была выявлена способность стимулировать этот процесс. Тот факт, что аденозин высвобождается из аксонов в ответ на распространение импульсов, означает, что электрическая активность мозга действительно влияет на процесс миелинизации. Подобные открытия помогут учёным вести поиск средств для лечения болезней демиелинизации. Возможно, эффективными окажутся лекарства, напоминающие своей химической структурой аденозин. И не исключено, что добавление аденозина в культуру стволовых клеток превратит их в миелинизирующие глиальные клетки, которые можно будет использовать в качестве трансплантатов.
Вырываясь из путнейронных сетей
Ограничивается ли участие глии в регуляции нейронных функций образованием вокруг аксонов миелиновой оболочки? По-видимому, нет. Ришар Робитайль (Richard Robitaille) из Монреальского университета обнаружил, что величина электрического потенциала, возникающего в мышце лягушки под влиянием стимуляции синапса, увеличивалась или уменьшалась в зависимости от того, какие химические вещества он вводил в шванновские клетки, окружающие этот синапс. Когда Эрик Ньюман (Eric A. Newman) из Миннесотского университета прикасался к сетчатке крысы, посылаемые глией «кальциевые сигналы» изменяли частоту импульсации зрительных нейронов. А Майкен Недергард (Maiken Nedergaard) из Нью-Йоркского медицинского колледжа, изучавший срезы гиппокампа крысы (эта область мозга принимает участие в процессах памяти), наблюдал усиление электрической активности синапсов в то время, когда окружающие астроциты увеличивали поглощение кальция. Подобные изменения эффективности синапсов учёные рассматривают в качестве главного фактора пластичности нервной системы, т. е. её способности изменять реакции на основании прошлого опыта, и глия, таким образом, может играть важную роль в клеточных процессах обучения и памяти.
Глия контролирует синапсы
Бен Баррес из Стэнфордского университета обнаружил, что если выращивать нейроны из сетчатки крысы в лабораторной культуре, не содержавшей астроцитов, синапсов на нейронах образуется очень мало. Когда же учёный добавил в культуру астроциты или просто среду, в которой прежде находились астроциты, синапсы появились в большом количестве. Затем он обнаружил присутствие в среде двух химических веществ, высвобождаемых астроцитами для стимуляции образования синапсов, — жировой комплекс под названием ароЕ/холестерин и белок тромбоспондин. Немного позднее Ле Тиан и Уэсли Томпсон из Техасского университета в Остине изучали мышей, которым вводились вещества, заставлявшие флуоресцировать шванновские клетки. Это позволило им воочию наблюдать за деятельностью глиальных клеток в области контактов нервов с мышечными волокнами. После того как учёные перерезали подходящий к мышце аксон, нервно-мышечный синапс исчезал, но на его «мышечной стороне» оставалась группа рецепторов нейротрансмиттера. Исследователям, конечно, было известно, что аксон вновь сможет прорасти к покинутым им рецепторам. Но как он найдёт к ним путь? Следя за флуоресценцией, Томпсон увидел, что шванновские клетки, окружавшие интактные синапсы, почувствовали, что синапс-сосед оказался в беде. Тогда они дружно выпустили в его сторону отростки, дотянулись ими до повреждённого синапса и образовали своего рода мостик, по которому аксон мог послать к своему синапсу новую проекцию (см. фото). Эти данные указывают на то, что глия помогает нейронам определить место, где нужно образовывать синаптические соединения. Сегодня учёные пытаются использовать эту способность глии для лечения повреждений спинного мозга: они трансплантируют шванновские клетки в повреждённые участки спинного мозга лабораторных животных. |
В связи с описанными выше наблюдениями остро встаёт одна проблема. Поглощение кальция распространяется по всей популяции астроцитов, подобно прокатывающимся по стадиону волнам взявшихся за руки болельщиков. Такая дружная реакция эффективна для управления работой всей группы клеток, но она слишком груба для передачи сложных сообщений. Принцип «все как один!» может оказаться полезным для координации общей активности мозга во время цикла сон-бодрствование, но чтобы входить во все тонкости переработки информации, глиальные клетки должны уметь «переговариваться» и со своими непосредственными соседями.
Стивен Смит предполагает, что нейроны и клетки глии способны вести друг с другом беседы и более «интимного свойства». Экспериментальные методы, которыми располагали в то время учёные, не позволяли им апплицировать нейротрансмиттеры в таких ничтожно малых дозах, которые могли бы воспроизвести истинные «переживания» астроцита, находящегося рядом с синапсом. Филипу Хейдону (Philip G. Haydon) из Пенсильванского университета удалось добиться этого только в 2003 г. с помощью современного лазерного метода аппликации нейротрансмиттеров. Учёный стимулировал в срезах гиппокампа выброс такого ничтожного количества глутамата, которое мог обнаружить только один-единственный астроцит. Хейдон наблюдал при этом, что астроцит посылает специфические кальциевые сигналы лишь небольшому числу окружающих его астроцитов. Исследователь предположил, что наряду с «кальциевыми волнами», оказывающими широкомасштабное воздействие, «между астроцитами существуют и близкодействующие связи». Иными словами, разрозненные цепочки астроцитов в головном мозге координируют свою активность в соответствии с активностью нейронных цепей.
Описанные выше открытия позволили Хейдону, автору этой статьи, сформулировать рабочую гипотезу, согласно которой обмен сигналами помогает астроцитам активировать нейроны, чьи аксоны оканчиваются на сравнительно большом от них расстоянии. А также утверждать, что эта активация способствует высвобождению нейротрансмиттеров из отдалённых синапсов. Это позволяет астроцитам регулировать готовность отдалённых синапсов к изменению своей силы (эффективности), что является клеточной основой процессов памяти и обучения.
Результаты исследований, представленные на ежегодном съезде Общества нейробиологии в ноябре 2003 г., подкрепляют эту гипотезу и даже свидетельствуют об участии глии в образовании новых синапсов. Следует упомянуть выполненную два года назад работу Бена Барреса (Ben A. Barres) и Фрэнка Фрайгера (Frank W. Pfrieger) из Стэнфордского университета, сообщивших, что выращенные в культуре нейроны крысы в присутствии астроцитов образуют большее количество синапсов. Впоследствии сотрудники из лаборатории Барреса обнаружили, что белок тромбоспондин, предположительно астроцитарного происхождения, выполняет функцию химического посредника и стимулирует образование синапсов. Чем большее количество этого белка учёные добавляли в культуру астроцитов, тем больше синапсов появлялось на нейронах. Возможно, тромбоспондин отвечает за связывание белков и других соединений, необходимых для образования синапсов во время роста молодых нервных сетей и, следовательно, может участвовать в модификации синапсов, когда эти сети подвергаются старению.
Будущие исследования расширят наши представления о влиянии глии на нейронную часть головного мозга. Возможно, учёным удастся доказать, что наша память (или её клеточный аналог — такой, как долговременная потенциация) зависит от функционирования синаптических астроцитов. Не исключено также, что будет установлено, каким образом сигналы, передаваемые по цепочкам астроцитов, оказывают влияние на отдалённые синапсы.
Сравнение головного мозга показывает, что чем более высокое положение занимают животные на «эволюционной лестнице», тем выше у них соотношение между числом глиальных клеток и нейронов. Хейдон предполагает, что увеличение связности астроцитов может повышать способности животных к обучению. Эта гипотеза проверяется сегодня экспериментально. Не исключено, что высокие концентрации глиальных клеток в мозге, а возможно, и наличие в нём более «действенной» глии, и превращает некоторых людей в гениев. Эйнштейн учил нас думать нетрадиционно. Его примеру последовали учёные, дерзнувшие «выпутаться» из нейронных сетей и решившие наконец выяснить, какое участие в переработке информации принимает нейроглия.
Об авторе:
Дуглас Филдз (R. Douglas Fields) —
заведующий отделением развития и пластичности нервной системы
Национального института охраны здоровья ребёнка и развития
человека, а также адъюнкт-профессор Мэрилендского университета (руководитель Программы развития нейробиологии и когнитивных наук). После защиты докторской диссертации работал в Йельском и Стэнфордском университетах.
«В мире науки»
scisne.net
Нейроглия: классификация клеток нейроглии
Нейроглия: классификация клеток нейроглии
Нейроглия состоит из макро (См. Клетки макроглии) — и микроглиальных клеток. К нейроглиальным элементам также относят эпендимные клетки , которые у некоторых животных сохраняют способность к делению.
Макроглия подразделяется на астроциты, или лучистые глиоциты , и олигодендроциты ( рис. 2 ). Астроциты — самые разнообразные глиальные клетки, имеющие звездчатую или паукообразную форму. Астроцитарная глия состоит из протоплазматических и фиброзных астроцитов ( рис. 3 ).
В сером веществе мозга встречаются преимущественно протоплазматические астроциты. Их тело имеет относительно крупные размеры (15-25 мкм) и многочисленные ветвистые отростки.
В белом веществе мозга располагаются волокнистые, или фиброзные, астроциты. У них небольшое тело (7-11 мкм) и длинные малоразветвленные отростки.
Астроциты — единственные клетки, располагающиеся между капиллярами ( рис. 3 ) и телами нейронов и участвующие в транспорте веществ из крови к нейронам и транспорте продуктов метаболизма нейронов обратно в кровь ( рис. 2 ). Астроциты формируют гематоэнцефалический барьер . Он обеспечивает избирательное прохождение из крови в ткань мозга различных веществ. Благодаря гематоэнцефалическому барьеру в экспериментах многие продукты обмена, токсины, вирусы, яды при введении в кровь почти не обнаруживаются в спинномозговой жидкости.
Олигодендроциты — мелкие (размеры тела около 5-6 мкм) клетки со слаборазветвленными, относительно короткими и немногочисленными отростками. Одна из основных функций олигодендроцитов — формирование оболочек аксонов в ЦНС . Олигодендроцит наматывает свою мембрану вокруг нескольких аксонов нервных клеток, образуя многослойную миелиновую оболочку ( рис. 2 ).
Олигодендроциты выполняют еще одну очень важную функцию — они участвуют в нейронофагии (от греч. фагос — пожирающий), т.е. удаляют омертвевшие нейроны путем активного поглощения продуктов распада.
В периферической нервной системе функции олигодендроглии выполняют шванновские клетки нейроэктодермального происхождения. Они отличаются от олигодендроглии тем, что охватывают обычно только один участок отдельного аксона . Длина такого охвата не превышает 1 мм. Между отдельными шванновскими клетками формируются своеобразные границы, которые носят название перехватов Ранвье ( рис. 2 ).
Тело микроглиальных клеток неправильной формы, с тонкими многочисленными отростками, усеянными шипиками ( рис. 3 ). Микроглиальные клетки способны к активной миграции, они распределены по всей ЦНС и выполняют фагоцитарные функции.
Эпендимные клетки (эпендимоциты) ( рис. 3 ) выстилают поверхность желудочков головного мозга и центральный канал спинного мозга . Эпендимоциты ближе всего к астроцитам . На ранних стадиях онтогенеза апикальные участки эпендимных клеток снабжены ресничками, которые способствуют перемещению спинномозговой жидкости . На более поздних стадиях онтогенеза реснички утрачиваются, сохраняясь только в водопроводе мозга .
Ссылки:
medbiol.ru
Особенности глиальных клеток
Нервная система состоит из двух главных типов клеток: нервных клеток (нейронов) и глиальных клеток. Глиальные клетки в совокупности называют нейроглией или глией. Они составляют по крайней мере половину объёма центральной нервной системы. Число гиальных клеток в 10-50 раз больше, чем нейронов. Нейроны центральной нервной системы окружены глиальными клетками. Глия является средой для нейронов. За пределами центральной нервной системы многие аксоны окружены оболочками (миелинизированные волокна), образованными глиальными клетками (шванновские клетки). Нейроны и глиальные клетки разделены межклеточными щелями. Их ширина ~15-20 нм. Щели сообщаются друг с другом, образуя внеклеточное (интерстициальное) пространство нейронов и глии, заполненное интерстициальной жидкостью. Интерстициальное пространство занимает 12-14% общего объёма мозга. Через интерстициальную жидкость путем диффузии к нервным, глиальным клеткам доставляются кислород, питательные вещества из плазмы крови кровеносных капилляров и удаляются конечные продукты метаболизма.
Глиальные клетки служат опорным и защитным аппаратом для нейронов. Метаболизм глиальных клеток тесно связан с метаболизмом нейронов, которые они окружают. Возможно, глиальные клетки участвуют в процессах памяти. Сателлиты нейроглии, называемые шванновскимя клетками, синтезируют оболочки миелинизированных нервных волокон периферических нервов. Некоторые глиальные клетки выполняют функции фагоцитов.
Клетки нейроглии делят на ряд типов. Эпендимоциты макроглии выстилают желудочки головного мозга и спинномозговой канал и образуют эпителиальный слой в сосудистом сплетении. Они соединяют желудочки с подлежащими тканями и выполняют разграничительную, опорную и секреторную функции. Клетки макроглии делятся на две категории -астроциты и олигодендроциты. Астроциты выполняют опорную функцию, осуществляют транспорт питательных веществ к нейронам, поглощение погибших клеток, участвуют в регулировании состава жидкости экстрацеллюлярного пространства (глюкоза, аминокислоты, ионы, в частности, являются буфером и депо ионов калия). Протоплазматические астроциты локализованы в сером веществе мозга. От тела астроцита, содержащего овальное ядро и большое количество гликогена, отходят сильно разветвленные короткие и толстые отростки. Фибриллярные астроциты локализованы в белом веществе мозга. Ядро у них также овальное, и тело клетки также содержит много гликогена, но отростки длинные и менее разветвленные. Некоторые ветви фибриллярных астроцитов буквально упираются в стенки кровеносных сосудов. Полагают, что эти клетки переносят питательные вещества из крови в нейроны. Астроциты этих двух типов взаимодействуют в обширной трехмерной сети, в которой расположены нейроны. Здесь астроциты часто делятся и в случае повреждений мозга образуют рубцовую ткань.
Олигодендроциты локализованы в сером и белом веществе мозга. Их главной функцией является изоляция миелином нейронов центральной нервной системы. Олигодендроциты мельче астроцитов и имеют одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек. Цитоплазма олигодендроцитов содержит большое количество рибосом. Шванновские клетки-это специализированные олигодендроциты, синтезирующие миелиновую оболочку миелинизированных волокон.
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. Эти клетки продолговатой формы и содержат лизосомы и хорошо развитый аппарат Гольджи. От каждого конца тела клетки отходит толстый отросток. Отростки ветвятся мелкими боковыми веточками. При повреждениях мозга клетки микроглии превращаются в фагоциты, способные перемещаться посредством амебоидного движения.
Основные отличия глиальных клеток от нейронов:
1) Глиальные клетки имеют только один тип отростков, в то время как нейроны
имеют два типа отростков — аксоны и дендриты.
2) Глиальные клетки, хотя и имеют потенциал покоя (~89 мв), не могут
генерировать потенциал действия как нейроны.
3) Глиальные клетки не имеют химических синапсов как нейроны.
4) Глиальные клетки, в отличие от зрелых нейронов, способны к делению.
5) Число глиальных клеток в центральной нервной системе в 10-50 раз больше, чем
нейронов.
Содержание ДНК в нейронах колеблется в пределах 6–8 нг на клетку, в астроцитах оно достигает 11 нг. Олигодендроциты характеризуются меньшим содержанием ДНК, что свидетельствует о практическом отсутствии полиплоидии и невысоком содержании митохондрий в олигодендроглиальных клетках.
Большое значение для сравнения метаболических превращений и роли ДНК в глии и нейронах имеет исследование путей ее синтеза и деградации. К сожалению, таких сравнительных работ еще недостаточно, чтобы сделать окончательные выводы. Однако установлены некоторые различия в каталитических свойствах ДНК-полимераз, выделенных из нейронов и нейроглии. Эти различия касаются предпочтительного использования матриц, субстратной специфичности и отношения к ингибиторам и отражают способность глиальных клеток к интенсивному размножению.
Качественный состав РНК в основном сходен в нейронах и нейроглии, хотя количество отдельных фракций РНК различается. Так, в глии обнаружено больше 4S РНК, Кроме того, РНК нейронов и нейроглии отличаются по общему нуклеотидному составу. Наибольшие различия касаются таких оснований, как аденин и цитозин. Исследование метаболизма РНК в нейронах и нейроглии проводится с использованием различных меченых предшественников. Установлено, что включение Н-аденина и Н-цитозина в нуклеотиды нейроглии происходит более интенсивно по сравнению с нейронами. Удалось установить цикличность биосинтеза РНК нейроглии, что дает основание предполагать существование в глиальных клетках двух пулов РНК, обладающих различной метаболической активностью.
Морфология фитоценоза
Биологические системы
Обмен газов в легких
Минералокортикоиды
Показатели легочной вентиляции
Регуляция дыхания
Гемолиз
Физиология анализаторов
Функциональное значение отделов ствола головного мозга
Основы зрительной рецепции
Собственные стволовые клетки
biofile.ru
Глиальные клетки: нейроглия или глия
Нейроглия в буквальном переводе означает – нервный клей (термин ввел немецкий цитолог Рудольф Вирхов). Она сопровождает нервные элементы и образует строму, в которой расположены нейроны. Кроме поддерживающей функции глии свойственны: трофическая, разграничительная, защитная и секреторная функции. У человека содержится приблизительно 10 глиоцитов, которые составляют почти половину объема мозга.
Глия подразделяется на глиоциты (макроглию) и микроглию – глиальные макрофаги. Различат следующие виды макроглии:
1) эпендимная глия, состоящая из эпендимоглиоцитов;
2) астроцитная глия, представленная астроцитами;
3) олигодендроглия, клетки которой называются олигодендроглиоциты.
Все виды макроглии способны к пролиферации.
Эпендимная глия развивается из внутренней эпителиоподобной выстилки нервной трубки. Эпендимоглиоциты имеют вид кубического или низкого, или кубического эпителия. От основания клеток отходят тонкие отростки, вплетающиеся в глиальный остов. На верхушке клеток имеются реснички и микроворсинки. Цитоплазма клеток богата митохондриями и аппаратом Гольджи, но мало эндоплазматической сети и свободных рибосом, содержит много пузырьков и гранул. Свободная поверхность клеток имеет сложную структуру: плазматическая мембрана многократно впячивается и возникают многочисленные микроворсинки. В эмбриогенезе эпендимная глия выполняет также пролиферативную функцию.
Во взрослом организме эпендимоглиоциты выстилают полости мозга: центральный канал спинного мозга и желудочки мозга. Эпендимоглиоцтиы выполняют разграничительную функцию и кроме того, предполагается их участие в выработке некоторых компонентов цереброспинальной жидкости. Эпендимный слой может быть источником развития очень злокачественной опухоли – медулобластомы, эпендимомы.
Астроцитная глияв эмбриональном периоде развивается из эпендимной глии. Астроцитная глия составляет остов органов центральной нервной системы. Клетки её имеют многочисленные отростки, отсюда и произошло их название – астроциты (астер – звезда).
Астроциты могут быть волокнистыми, или фибриллярными, и плазматическими (коротколучистые).
Плазматические астроциты лежат в сером веществе мозга, а фибриллярные астроциты составляют остов белого вещества.
В астроцитах содержится светлое, бедное хроматином овальное ядро. Цитоплазма выглядит светлой и в ней обнаруживается аппарат Гольджи, эндоплазматическая сеть и митохондрии, но органоидов мало, гранулы гликогена. В отростках фибриллярных астроцитов лежат тонкие нити, собранные в пучки. Это глиальные фибриллы, они всегда лежат интерцеллюлярно. Эти фибриллы представляют собой опорные элементы. Астроцитная глия выполняет различные функции: опорную, трофическую, разграничительную, т.к. отростки их клеточных элементов входят в состав глиальных мембран. Астроциты выполняют роль дворников, удаляющих из внеклеточного пространства избыток ионов калия и тем самым защищает нейроны от излишней деполяризации.
В метаболических процессах ЦНС астроциты в норме малоактивны, обладая слабой окислительной способностью. Однако, по данным ряда исследователей их уровень обмена очень высок.
Созревание астроцитов заканчивается к 5-ти годам. В первый год жизни в спинном мозге насчитывается 20-35 клеток в поле зрения. Уменьшение их числа происходит после 35 летнего возраста. Все виды астроглии у взрослых сохраняют способность к митотическому делению. Астроциты выделяют ряд веществ, способствующих росту аксонов: фактор роста нервов, фибронектин и ламинин.
Благодаря отросткам, астроциты обладают чрезвычайно большой поверхностью и непосредственно граничат с 50 % межклеточного пространства серого вещества. Астроциты способны к фагоцитозу, особенно при ряде патологических состояний: они превращаются в пролиферирующие, мигрирующие и фагоцитирующие клетки. Одни отростки астроцитов доходят до кровеносных сосудов, при этом они на концах расширяются и распластываются на поверхности капилляров образуя астроцитную ножку, которая контактирует с ножкой другой клетки. Это глиальная мембрана. Она принимает участие в образовании гемато-энцефалического барьера, благодаря которому некоторые вещества могут переходить из крови в мозг очень медленно или вообще не попадают туда.
В настоящее время в нервной ткани выявлены биологически – активные вещества (трефоны), которые вырабатываются нервными клетками и клетками глии. Эти вещества путем эндоцитоза затем попадают в другие нервные клети и регулируют внутриклеточный метаболизм, их рост, развитие, дифференцировку. Не исключено их воздействие на генетический аппарат клетки, что обусловливает долговременные пластические изменения как нейронов, так и иннервируемых тканей.
Олигодендроглия является самой распространённой. Она окружает тела нейронов и образует оболочки нервных волокон. Клетки олигодендроглии имеют меньшие размеры чем астроциты и называются олигодендроглиоцитами. Они имеют многочисленные короткие отростки.
Ядра их богаты хроматином, в цитоплазме много органелл, в том числе много свободных рибосом, гранул эндоплазматической сети и микротрубочек, благодаря чему возникает сходство с нейронами. Уровень окислительных процессов их очень велик.
Эндоплазматическая сеть представлена широкими цистернами и каналами. Много набухших митохондрий. Методом меченых атомов установлен высокий уровень обмена РНК и протеинов. Много включений холестерина. Обнаружены редкие митозы.
Олигодендроглиоциты имеют короткие отростки, которые окружают тела нервных клеток и нервные волокна. В отличие от других глиальных клеток олигодендроглиоциты часто располагаются группами. Олигодендроглия выполняет ряд функций: трофическую, так как клетки глии, благодаря своему положению между нейронами и капиллярами, обеспечивает поступление питательных веществ нервным клеткам, разграничительную, принимает участие в передаче нервных импульсов и в регенерации нервных волокон.
Глиальные клетки более многочисленные, чем нейроны. Нейроны и глиоциты разделены межклеточной щелью шириной 15-20 нм. При этом, щелт сообщаются друг с другом. Образуя заполненное жидкостью внеклеточное пространство нейронов и глии, что имеет значение в обеспечении нервных клеток кислородом и питательными веществами.
Микроглия – глиальные макрофаги появляются в органах нервной системы с момента прорастания в них кровеносных сосудов. Она представлена мелкими клетками с небольшими отростками. Клетки способны к активному перемещению. Функция микроглии фагоцитарная. Она развивается из мезенхимы (однако у них нет рецепторов, характерных для фагоцитов). Количество клеток микроглии увеличивается с возрастом и достигает в спинном мозге максимум к 15 годам. Нельзя исключить, что они имеют нейроэктодермальное происхождение, а возможно и то и другое.
Микроглия особенно сильно активируется при тех болезненных процессах когда происходит сильный распад нервной ткани: увеличение числа и размеров лизосом, появляются клетки больших размеров, фаголизосомы. Микроглия способна фагоцитировать целые некротические клетки (их можно встретить в составе фаголизосом), синаптические структуры.
Между структурными компонентами нервной ткани располагается внеклеточное пространство, на долю которого приходится около 20-25%. В его состав входит вода, обусловливающая высокую гидратацию нервной ткани. Вода является главнейшим веществом, поступающим в мозг путём диффузии. Полупериод обмена воды в нервной ткани мозга составляет около 12-30 минут, что зависит от степени капилляризации. Вода свободно переходит в мозг или из мозга в зависимости от изменения осмотических условий плазмы крови. Это свойство широко используется в клинике для осмотерапии отёка мозга.
В отличие от плазмы здесь очень много ионов натрия и значительно меньше белков. Здесь очень много содержится гликопротеинов, гликозаминогликанов (особенно гиалуроновой кислоты и хондроитинсульфатом), гликолипиды. Благодаря такому строению по внеклеточному пространству легко диффундируют кислород, углекислый газ, Na2O, летучие анестетики. У детей транспорт различных веществ, в том числе ионов, осуществляется значительно быстрее, чем у взрослых.
1.Швалев В.Н. и др. Нервная ткань и нейроглия //Руководство по гистологии. В 2 т. Т.1.-Спб.,2001.-С.388-434.
2.Хэм А, Кормак Д. Гистология.В 5 т. Т.3.-М.,1983.-С.163-240.
3.Физиология человека /Под ред. Покровского В.М. и Коротько Г.Ф. В 2 т.Т.1.-М.,1997.- С.27-70.
4.Быков В.Л.Цитология и общая гистология.Спб.,1999.- С.456-497.
5.Гистология/Под ред.Улумбекова Э.Г.,Челышева Ю.А.М.,2001.-С.193-224.
6.Гистология /Под ред.Ю.И.Афанасьева, Юриной Н.А.,2001.-С.268-284.
7.Мяделец О.Д.Основы цитологии, эмбриологии и общей гистологии,М.,2002.-С.318-340.
Материалы: http://studopedia.ru/14_81758_neyrogliya.html
vekoff.ru
students-library.com