
Какие ядра таламуса относят к неспецифическим – .
Таламус, как коллектор чувствительной информации. Специфические ядра таламуса, их функциональная роль. Неспецифические ядра таламуса, характер их влияния на кору головного мозга.
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
cyberpedia.su
Неспецифическое ядро — Большая Энциклопедия Нефти и Газа, статья, страница 1
Неспецифическое ядро
Cтраница 1
Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает параллельно в разные отделы коры. Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса. [1]
Через неспецифические ядра таламуса в кору мозга поступают восходящие активирующие влияния от ретикулярной формации мозгового ствола. Система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы. [2]
Электрическое раздражение неспецифических ядер таламуса вызывает в коре больших полушарий периодические колебания потенциалов, синхронные с ритмом активности таламических структур. Реакция в коре возникает с большим скрытым периодом и значительно усиливается при повторении. Таким образом, нейроны коры больших полушарий вовлекаются в процесс активности как бы постепенно. Такая реакция вовлечения коры мозга отличается от специфических ее ответов своей генерализованностью, охватом обширных областей коры. Импульсы, идущие по путям болевой чувствительности, формируются при раздражении различных областей тела и внутренних органов. Скрытые периоды ответов в таламусе отличаются большой длительностью и вариабельностью. [3]
Другой тип окончаний таламокорти-кальных проекций образуется аксонами нейронов
При регистрации электрической активности различных отделов головного мозга кролика было обнаружено, что реакции в виде увеличения числа мыленных волн и веретен возникают одновременно во всех отведениях ( при скорости записи 15 мм / с), а наиболее интенсивную реакцию наблюдали в гипоталамусе, затем следовала кора сензомоторная, зрительная, специфические ядра таламу-са, неспецифические ядра таламуса. Можно заключить, что наиболее реактивными образованиями ЦНС при воздействии ПМП являются кора и гипоталамус. [6]
Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны
Через неспецифические ядра таламуса в кору мозга поступают восходящие активирующие влияния от ретикулярной формации мозгового ствола. Система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы. [8]
Для изучения механизма образования условных рефлексов существенное значение имеет не только точная регистрация самой ответной реакции ( слюноотделения, движения и т.п.), но также исследование электрической активности, возникающей в различных мозговых структурах во время действия условного и безусловного раздражителей. Для регистрации электрической активности применяют электроды, хронически вживляемые в различные области или слои коры большого мозга, а также в специфические и неспецифические ядра таламуса, ретикулярную формацию, гиппокамп и другие отделы головного мозга. В опытах с условными рефлексами широко используются микроэлектродные методы, позволяющие регистрировать электрическую активность отдельных нейронов, участвующих в осуществлении условнорефлекторной реакции. Для автоматического анализа электроэнцефалограмм, регистрируемых с различных областей коры, в опытах на животных непосредственно во время условнорефлекторных реакций используют электронно-вычислительные машины. [9]
Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает параллельно в разные отделы коры. Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса. [10]
Нейроны специфического комплекса ядер посылают по направлению к коре аксоны, почти не имеющие коллатералей. В отличие от него нейроны неспецифической системы посылают аксоны, дающие множество коллатералей. Вместе с тем и волокна, приходящие из коры к нейронам специфических ядер, характеризуются топографической локали-зованностью своих окончаний в противоположность широко разветвленной системе диф-фузно оканчивающихся волокон в неспецифических ядрах. [11]
Спиноталамический путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинальном ганглии, откуда они посылают в спинной мозг медленно проводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спиноталамического тракта направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса ( дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спиноталамического пути лишь частично дают проекции в соматосенсорную зону коры. [13]
Страницы: 1
www.ngpedia.ru
Таламус. Нормальная физиология
Зрительный бугор является местом переключения всех чувствительных проводников, идущих от экстеро-, проприои интерорецепторов, поднимающихся в кору головного мозга. В нем происходит обработка всей информации, поступающей в кору из спинного мозга и подкорковых структур. По мнению А. Уолкера, выдающегося исследователя зрительного бугра, «таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым и корковым центрам таким образом, чтобы организм смог адекватно приспособиться к постоянно меняющейся среде. Таламус, как видно, таит в себе тайну многого из того, что происходит в коре больших полушарий».
О полифункциональности таламуса говорит наличие в нем около 120 ядер, которые топографически можно разделить на три основные группы: переднюю, имеет проекции в поясную кору, медиальную – в лобную, латеральную – в теменную, височную, затылочную.
По функциональным признакам ядра зрительного бугра делят на три группы: специфические, образующие с соответствующими областями коры специфическую таламокортикальную систему, неспецифические, составляющие диффузную, неспецифическую таламокортикальную систему, и ассоциативные.
В состав группы специфических ядер входят переднее вентральное, медиальное, вентролатеральное, постмедиальное, постлатеральное, а также медиальные и латеральные коленчатые тела. Специфические ядра содержат так называемые «релейные» (передаточные) нейроны, имеющие мало дендритов и длинный аксон, заканчивающийся в III -IV слоях коры больших полушарий (соматосенсорная зона).
Если раздражать какое-либо из специфических ядер электрическими импульсами, то в соответствующих проекционных областях коры с коротким латентным периодом возникает реакция, названная первичным ответом.
Каждое из специфических ядер отвечает за свой вид чувствительности, так как они, так же как и кора больших полушарий, имеют соматотопическую локализацию, т. е. к ним поступают сигналы от тактильных, болевых, температурных, мышечных рецепторов, а также от интерорецепторов зон проекции блуждающего и чревного нервов.
Латеральное, или наружное, коленчатое тело – это подкорковый центр зрения, таламическое реле для зрительных импульсов. Оно имеет афферентные связи с сетчаткой глаза и буграми четверохолмия и эфферентные – с затылочной долей коры больших полушарий.
Медиальное коленчатое тело – подкорковый, таламический центр слуха, получает афферентные импульсы из латеральной петли и нижних бугров четверохолмия и посылает информацию в височную долю коры больших полушарий.
К неспецифическим ядрам таламуса относятся: срединный центр, парацентральное ядро, центральное медиальное и латеральное, субмедиальное, вентральное переднее, парафасцикулярпое, ретикулярное ядро, перивентрикулярное и центральная серая масса.
Нейроны неспецифических ядер являются клетками ретикулярной формации, аксоны которых контактируют со всеми слоями коры больших полушарий, образуя диффузные связи. В свою очередь, к неспецифическим ядрам поступает информация от ретикулярной формации ствола мозга, лимбической системы, базальных ганглиев и специфических ядер таламуса.
Раздражение неспецифических ядер электрическим током вызывает возникновение в коре больших полушарий не локально, а диффузно специфической электрической активности, имеющей длинный латентный период и вид веретена, названной сонными веретенами, или реакцией вовлечения.
Основная функция неспецифических ядер состоит в облегчении или торможении специфических ответов коры, т. е. в изменении их возбудимости.
Ассоциативные ядра таламуса включают в себя медиодорсальное, латеральное дорсальное ядро и подушку. Нейроны этих ядер имеют разную форму и количество отростков, что позволяет им выполнять разнообразные функции, связанные с переработкой информации различных модальностей, после чего она поступает в 1-е и 2-е слои ассоциативной зоны коры, частично – в проекционные зоны коры (4-е и 5-е слои).
При поражении таламических ядер, отвечающих за переработку всей сенсорной информации, в том числе и болевой, могут возникать сильнейшие боли. С наличием застойного очага возбуждения в таламусе и коре больших полушарий связаны «фантомные боли» (в ампутированной конечности).
Таламус обеспечивает двигательные и вегетативные реакции, связанные с сосанием, жеванием, глотанием и смехом.
Поделитесь на страничкеСледующая глава >
med.wikireading.ru
Функции таламуса
Краткая анатомия
Основную массу промежуточного мозга (20г) составляет таламус. Парный орган яйцевидной формы, передняя часть которого заострена (передний бугорок), а задняя расширенная (подушка) нависает над коленчатыми телами. Левый и правый таламусы соединены межталамической спайкой. Серое вещество таламуса разделено пластинками белого вещества на переднюю, медиальную и латеральную части. Говоря о таламусе, включают также метаталамус (коленчатые тела), принадлежащий к таламической области. Таламус наиболее развит у человека. Таламус, зрительный бугор, — ядерный комплекс, в котором происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация
Таламус (thalamus), зрительный бугор, — ядерный комплекс, в котором происходит
обработка и интеграция практически всех сигналов, идущих в кору большого мозга
от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга. В
ядрах таламуса происходит переключение информации, поступающей от экстеро-,
проприорецепторов и интерорецепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела являются подкорковыми центрами зрения и слуха, а
узел уздечки и переднее зрительное ядро участвует в анализе обонятельных
сигналов, можно утверждать, что зрительный бугор в целом является подкорковой
«станцией» для всех видов чувствительности. Здесь раздражения внешней и
внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений,
эмоций. Возможность получать информацию о состоянии множества систем организма
позволяет таламусу участвовать в регуляции и определении функционального
состояния организма. В целом (подтверждением тому служит наличие в таламусе
около 120 разнофункциональных ядер).
Функции ядер таламуса
Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы. Передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга. Медиальная — в лобную долю коры. Латеральная — в теменную, височную, затылочную доли коры. Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
Специфические сенсорные и несенсорные ядра
К специфическим ядрам относятся переднее вентральное, медиальное,
вентролатеральное, постлатеральное, постмедиальное, латеральное и медиальное
коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха
соответственно. Основной функциональной единицей специфических таламических ядер
являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их
функция заключается в переключении информации, идущей в кору большого мозга от
кожных, мышечных и других рецепторов.
В свою очередь специфические (релейные) ядра делятся на сенсорные и несенсорные.
От специфических сенсорных ядер информация о характере сенсорных стимулов
поступает в строго определенные участки III-IV слоев коры большого мозга.
Нарушение функции специфических ядер приводит к выпадению конкретных видов
чувствительности, так как ядра таламуса, как и кора большого мозга, имеют
соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса
возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут
сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют
сигналы от интерорецепторов зон проекции блуждающего и чревного нервов,
гипоталамуса. Латеральное коленчатое тело имеет прямые эфферентные связи с
затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с
передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному
реагируют на цветовые раздражения, включение, выключение света, т.е. могут
выполнять детекторную функцию. В медиальное коленчатое тело поступают
афферентные импульсы из латеральной петли и от нижних бугров четверохолмий.
Эфферентные пути от медиальных коленчатых тел идут в височную зону коры большого
мозга, достигая там первичной слуховой области коры.
Несенсорные ядра переключают в кору несенсорную импульсацию, поступающую в таламус из разных отделов головного мозга. В передние ядра импульсация поступает в основном из сосочковых тел гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору, откуда аксонные связи идут к гиппокампу и опять к гипоталамусу, в результате чего образуется нейронный круг, движение возбуждения по которому обеспечивает формирование эмоций («эмоциональное кольцо Пейпеца»). В связи с этим передние ядра таламуса рассматриваются как часть лимбической системы. Вентральные ядра участвуют в регуляции движения, выполняя таким образом моторную функцию. В этих ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая после этого проецируется в моторную и премоторную кору. Через эти ядра таламуса происходит передача в моторную кору сложных двигательных программ, образованных в мозжечке и базальных ганглиях.
Неспецифические ядра
Эволюционно более древняя часть таламуса, включающая парные ретикулярные ядра и интраламинарную (внутрипластинчатую) ядерную группу. Ретикулярные ядра содержат преимущественно мелкие, многоотростчатые нейроны и функционально рассматриваются как производное ретикулярной формации ствола мозга. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя диффузные связи. К неспецифическим ядрам поступают связи из ретикулярной формации ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Благодаря этим связям неспецифические ядра таламуса выступают в роли посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единый функциональный комплекс.
Ассоциативные ядра
Ассоциативные ядра принимают импульсацию от других ядер таламуса. Эфферентные
выходы от них направляются, главным образом, в ассоциативные поля коры.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные
трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные
функции. Ряд нейронов изменяет активность только при одновременном комплексном
раздражении. Подушка получает главную импульсацию от коленчатых тел и
неспецифических ядер таламуса. Эфферентные пути идут от нее в
височно-теменно-затылочные зоны коры, участвующие в гностических (узнавание
предметов, явлений), речевых и зрительных функциях (интеграция слова со
зрительным образом), а также в восприятии «схемы тела». Медиодорсальное ядро
получает импульсацию от гипоталамуса, миндалины, гиппокампа, таламических ядер,
центрального серого вещества ствола. Проекция этого ядра распространяется на
ассоциативную лобную и лимбическую кору. Оно участвует в формировании
эмоциональной и поведенческой двигательной активности. Латеральные ядра получают
зрительную и слуховую импульсацию от коленчатых тел и соматосенсорную
импульсацию от вентрального ядра.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических,
неспецифических и ассоциативных ядер, позволяет ему организовывать такие
двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции
интегрируются в таламусе с вегетативными процессами, обеспечивающими эти
движения.
biofile.ru
Зрительный бугор строение и принципы работы
Нервные клетки таламуса группируются в большое количество ядер (до 40), которые топографически разделяют на:
- Передний таламус: неспецифические ретикулярные ядра.
- Задний таламус: ЛКТ и МКТ.
- Срединный таламус (интраламинарная область): неспецифические ядра ретикулярной формации.
- Латеральный таламус: двигательные ядра.
По функции таламические ядра можно дифференцировать на специфические (сенсорно-релейные), неспецифические, ассоциативные и моторные (несенсорно-релейные).
В специфических, или проекционных, ядрах таламуса происходит синаптическое переключение сенсорной информации с аксонов восходящих афферентных путей на следующие, конечные нейроны, отростки которых идут в соответствующие сенсорные проекционные области коры больших полушарий. То есть, в специфическом таламусе проходят все сенсорные афференты, идущие на кору. Повреждение специфических ядер приводит к необратимому выпадению определенных видов чувствительности. Эти экспериментальные факты свидетельствуют о том, что специфические ядра являются передаточной станцией на пути афферентных импульсов от периферических рецепторов к коре больших полушарий.
Релейные (переключающие ядра): к ядру подходит сенсорный афферент, и идет переключение с аксона на следующий нейрон. Типичные релейные клетки расположены в). Релейные ядра, за счет механизма возвратно-коллатерального торможения, обладают способностью к генерации ритмической активности.
Среди основных проекционных ядер таламуса можно выделить вентробазальное ядро. Оно является главным реле для переключения соматосенсорной афферентации. Здесь переключаются тактильная, вкусовая, висцеральная, частично температурная и болевая чувствительность. Здесь также проецируются такие тонко организованные части тела, как лицо, язык.
Микроэлектродные исследования нейронов вентробазального комплекса показали, что данное ядро, как и прочие специфические ядра, организовано по топическому принципу. Суть этого принципа состоит в том, что каждый нейрон активируется раздражением рецепторов определенного участка кожи, причем смежные участки туловища проецируются на смежные части вентробазального комплекса.
Специфичность данных нейронов проявляется также в том, что каждый из них возбуждается одним типом рецепторов. Вентробазальные ядра связаны с соматосенсорной корой постцентральной извилины, в которой формируются соответственные ощущения.
Специфическим ядром зрительной сенсорной системы является латеральное коленчатое тело (ЛКТ), имеющее прямые связи с затылочными (зрительными) проекционными областями коры больших полушарий.
Латеральное коленчатое тело имеет слоистую структуру и состоит из 6 слоев. Аксоны, идущие в ЛКТ из зрительного тракта, распределяются в нем с поразительной четкостью: три слоя ЛКТ (5,3,2) связаны с ипсилатеральным глазом, а три остальные (6,4,1) — с контралатеральным. Клетки более глубоких слоев (1,2) крупнее, чем клетки менее глубоких. Нейроны ЛКТ обладают концентрическими рецептивными полями, центр и периферия которых антагонистичны и по-разному реагируют на изменение освещенности. Рецептивное поле нейрона ЛКТ представляет собой концентрически организованную совокупность рецепторов сетчатки, имеющую либо возбуждающий (при включении света) центр и периферическую тормозную часть, либо, напротив, тормозный центр и возбуждающую данный нейрон ЛКТ периферическую область.
Такая организация рецепторного поля позволяет нейронам ЛКТ хорошо реагировать на контраст при определении границы между темным и светлым и на суммарную яркость светового стимула.
У млекопитающих некоторые нейроны ЛКТ обладают цветоспецифичными рецепторными полями и могут возбуждаться или, наоборот, тормозиться в зависимости от длины волны светового стимула. Таким образом, нейроны ЛКТ, также как и нейроны сетчатки, принимают участие в анализе зрительной информации.
Восходящие пути слуховой системы, идущие из нижних бугров четверохолмия и по волокнам латеральной петли, проецируются в специфическое таламическое ядро — медиальное коленчатое тело (МКТ), от которого начинается тракт, достигающий первичной слуховой коры в верхней части височных долей. Медиальное коленчатое тело состоит из мелкоклеточной и крупноклеточной частей и обладает специализиацией своих нейронов. Так, например, нервные клетки мелкоклеточной части МКТ имеют довольно узкую настройку на восприятие звуков различной высоты и принимают участие в анализе и передаче акустической информации.
Итак, таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым и корковым центрам.
В специфические ядра таламуса проецируются афференты не только от экстерорецепторов и рецепторов двигательного аппарата. Электрофизиологические исследования показали, что в таламусе имеются области проекций блуждающего и чревного нервов, чувствительные волокна которых несут информацию от интероцепторов. В то же время таламус имеет связи с гипоталамусом, где сосредоточены главные вегетативные центры.
Ассоциативные ядра, в отличие от специфических ядер, не могут быть отнесены к какой-либо одной сенсорной системе и получают афферентные импульсы от специфических проекционных ядер. Три ядра этой группы имеют связи с главными ассоциативными областями коры: ядро подушки связано с ассоциативной зоной теменной и височной коры, заднее латеральное ядро — с теменной корой, медиальное дорсальное ядро — с лобной долей. Четвертое ядро — переднее — имеет связи с лимбической корой больших полушарий. По-видимому, ассоциативные ядра участвуют в высших интегративных процессах, однако их функция изучена еще недостаточно. Ассоциативные ядра расположены в подушке. Разрушение подушки приводит к нарушению речевых функций и «схемы тела».
К моторным ядрам таламуса относится вентролатеральное ядро, которое имеет вход от мозжечка и базальных ганглиев и одновременно дает проекции в моторную зону коры больших полушарий. Моторные ядра таламусы являются релейными несенсорными ядрами. Это ядро включено в систему регуляции движений, и, как показал материал клиники, разрушение некоторых его участков ослабляет симптомокомплекс болезни Паркинсона. Разрушение вентролатерального ядра приводит к возникновению хорей и патологического тремора.
Неспецифические ядра функционально связаны с ретикулярной формацией ствола. Неспецифические ядра имеют широкие связи со всеми основными структурами мозга, в том числе, с ретикулярной формацией и лимбической системой. К числу этих ядер относится срединная и интроламинарная группа ядер таламуса, которая получает афферентный вход от волокон, восходящих из ретикулярной формации, и, кроме того, имеет двусторонние связи со специфическими ядрами таламуса. Эти ядра обнаруживают не локальные, в отличие от специфических, а диффузные проекции во все области коры. Основная функция состоит в регуляции возбудимости и электрической активности корковых нейронов.
При сопоставлении функций специфических и неспецифических ядер таламуса возникает резонный вопрос о взаимодействии этих двух систем, которые могут влиять на одни и те же нейроны коры больших полушарий. Как показали электрофизиологические исследования, восходящие влияния неспецифических ядер таламуса проявляются не в вызове разряда коркового нейрона, а в изменении его возбудимости. Неспецифические влияния из таламуса, повышая возбудимость корковых нейронов, облегчают их деятельность, при этом ответы корковых нейронов на импульсы, приходящие из специфических проекционных ядер, усиливаются. Вместе с тем неспецифические влияния могут иметь и противоположный знак и обнаруживать угнетающее действие на разряды корковых нейронов. Существует точка зрения, что неспецифические ядра включены в восходящую активирующую систему и являются посредниками между корой и ретикулярной формацией ствола, которая получает информацию от всех органов чувств. Таким образом, неспецифические ядра передают активирующие влияния ретикулярной формации и участвуют в поддержании оптимального тонуса коры. Неспецифические ядра оказывают модулирующее влияние, обеспечивают плавную перестройку высшей нервной деятельности.
К неспецифическим ядрам таламуса относятся ретикулярные ядра. Они выполняют функцию затормаживания релейных специфичных нейронов, переключающих сенсорные пути. Торможение осуществляется по принципу возвратного торможения: релейный нейрон отсылает коллатераль аксона к тормозному нейрону, который и затормаживает релейный нейрон.
wiki.1vc0.ru
Функции Промежуточного мозга: таламусаа, гипоталамуса и гипофиза
Основные образования промежуточного мозга
text_fields
text_fields
arrow_upward
Промежуточный мозг интегрирует сенсорные, двигательные и вегетативные реакции, необходимые для целостной деятельности организма. Основными образованиями промежуточного мозга являются:
- таламус,
- гипоталамус,
- гипофиз.
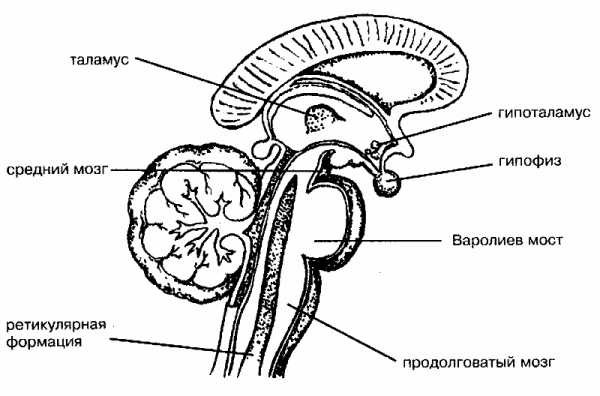
Функции таламуса
text_fields
text_fields
arrow_upward
Таламус — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору головного мозга от нейронов спинного мозга, среднего мозга, мозжечка, базальных ганглиев. Возможность получать информацию о состоянии множества систем организма позволяет ему участвовать в регуляции и определять функциональное состояние организма в целом. Это подтверждается уже тем, что в таламусе около 120 разнофункциональных ядер.
Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на три группы:
- передняя — проецирует аксоны своих нейронов в поясную кору;
- медиальная — в любую;
- латеральная — в теменную, височную, затылочную.
По проекциям определяется и функция ядер. Такое деление не абсолютно, так как часть волокон от ядер таламуса идет в корковые образования, часть — в разные зоны мозга.
Функциональная значимость ядер таламуса определяется не только их проекциями на другие структуры мозга, но и тем, какие структуры посылают к нему свою информацию. В таламус приходят сигналы от зрительной, слуховой, вкусовой, кожной, мышечной систем, от ядер черепно-мозговых нервов ствола, мозжечка, бледного шара, продолговатого и спинного мозга.
Функционально, по характеру нейронов входящих и выходящих из таламуса, его ядра делят на специфические, неспецифические и ассоциативные.
К специфическим ядрам относят:
- переднее вентральное, медиальное;
- вентролатеральное, постлатеральное, постмедиальное;
- латеральное и медиальное коленчатые тела.
Последние относятся, соответственно, к подкорковым центрам зрения и слуха.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, которые имеют мало дендритов, длинный аксон и выполняют переключательную функцию — здесь происходит переключение путей, идущих в кору от кожной, мышечной и других видов чувствительности.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки 3-4 слоев коры (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности. Это связано также с тем, что сами ядра таламуса имеют (так же, как и кора) соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются афферентациеи, поступающей только от своего типа рецепторов. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интероцепторов зон проекции блуждающего и чревного нервов, от гипоталамуса.
Ассоциативные ядра — медиодорсальные, латеральные, дорсальные и подушка таламуса. Основные клеточные структуры этих ядер: мультиполярные, биполярные, трехотростчатые нейроны, т.е. нейроны, способные выполнять полисенсорные функции. Наличие полисенсорных нейронов способствует взаимодействию на них возбуждений разных модальностей и созданию интегрированного сигнала для передачи в ассоциативную кору мозга. Аксоны от нейронов ассоциативных ядер таламуса идут 1 и 2 слоями ассоциативных и частично проекционных областей, по пути отдавая коллатерали в 4 и 5 слои коры, образуя аксосоматические контакты с пирамидными нейронами.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексом, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют связи по ретикулярному типу. Их аксоны поднимаются в кору и контактируют со всеми слоями коры, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из ретикулярной формации ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функций неспецифических ядер затрудняет появление веретенообразной активности, т.е. развитие сонного состояния.
Сложное строение таламуса, наличие здесь взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Читайте также:
Функции гипоталамуса
text_fields
text_fields
arrow_upward
Гипоталамус (подбугорье) — структура промежуточного мозга, организующая эмоциональные, поведенческие, гомеостатические реакции организма.
Функционально ядра гипоталамуса делят на переднюю, среднюю и заднюю группы ядер. Окончательно созревает гипоталамус к 13-14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. Мощные афферентные связи гипоталамуса с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, орбитальной, височной и теменной корой определяют его информативность о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые определяют специфику функций самого гипоталамуса. К этим особенностям относятся: чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреиии пептидов, нейромедиаторов и др.
Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Возбуждение ядер передней группы гипоталамуса приводит к реакции организма, его систем по парасимпатическому типу, т.е. реакциям, направленным на восстановление и сохранение резервов организма.
Возбуждение ядер задней группы вызывает симпатические эффекты в работе органов:
- происходит расширение зрачков,
- повышается кровяное давление,
- учащается ритм сердечных сокращений,
- тормозится перистальтика желудка и т.д.
Стимуляция ядер средней группы гипоталамуса приводит к снижению влияний симпатической системы. Указанное распределение функций гипоталамуса не абсолютно: все структуры гипоталамуса способны, но в разной степени, вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсируюшие отношения.
В целом, за счет большого количества входных и выходных связей, полифункциональности структур, гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций.
Так, в гипоталамусе располагаются центры:
- гомеостаза,
- теплорегуляции,
- голода и насыщения,
- жажды и ее удовлетворения,
- полового поведения,
- страха, ярости,
- регуляции цикла «бодрствование-сон».
Все эти центры реализуют свои функции путем активации или торможения вегетативной нервной системы, эндокринной системы, структур ствола и переднего мозга.
Нейроны передней группы ядер гипоталамуса продуцируют так называемые рилизинг-факторы (либерины) и ингибируюшие факторы (статины), которые регулируют активность передней доли гипофиза — аденогипофиз.
Нейроны срединной группы ядер гипоталамуса обладают детектирующей функцией, они реагируют на изменение температуры крови, электромагнитный состав и осмотическое давление плазмы, количество и состав гормонов крови.
Терморегуляция со стороны гипоталамуса проявляется в изменении теплопродукции или теплоотдачи организмом. Возбуждение задних ядер сопровождается усилением обменных процессов, увеличением частоты сердечных сокращений, дрожанием мышц туловища, что приводит к росту теплопродукции в организме.
Раздражение передних ядер гипоталамуса
- расширяет сосуды,
- усиливает дыхание, потоотделение — т.е. организм активно теряет тепло.
Пищевое поведение в форме поиска пищи, слюноотделения, усиления кровообращения и моторики кишечника наблюдается при стимуляции ядер заднего гипоталамуса. Повреждение других ядер вызывает голодание (афагия) или чрезмерное потребление пищи (гиперфагия), и, как следствие — ожирение.
В гипоталамусе расположен центр насыщения, чувствительный к составу крови — по мере поедания пищи и ее усвоения, нейроны этого центра тормозят активность нейронов центра голода.
Исследования во время хирургических операций показали, что у человека раздражение ядер гипоталамуса вызывает эйфорию, эротические переживания. В клинике отмечено также, что патологические процессы в области гипоталамуса сопровождаются ускорением полового созревания, нарушением менструального цикла, половой способности.
Гипоталамус является также, как уже отмечалось, одним из центров регуляции цикла «бодрствование-сон», при этом задний гипоталамус активизирует бодрствование, передний гипоталамус при стимуляции вызывает сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон.
Читайте также:
Функции Гипофиза
text_fields
text_fields
arrow_upward
Гипофиз тесно связан с гипоталамусом структурно и функционально. Задние отделы гипофиза (нейрогипофиз) накапливают гормоны, продуцируемые гипоталамусом и регулирующие водно-солевое равновесие, контролирующие функции матки и молочных желез.
Передние отделы гипофиза (аденогипофиз) вырабатывают:
- адренокортикотропный гормон — АКТГ, который стимулирует работу желез надпочечников;
- тиреотропный гормон — стимулирует рост и секрецию щитовидной железы;
- гонадотропный гормон — регулирует активность половых желез;
- соматотропный гормон — обеспечивает развитие костной системы; пролактин — стимулирует рост и активность молочных желез и др.
В гипоталамусе и гипофизе образуются также нейрорегуляторные энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса.
Читайте также:
Читайте также:
doctor-v.ru
|
Специфические ядра таламуса Специфические ядра имеют отчетливую топографическую и функциональную разграниченность проекций к определенным об-ластям коры больших полушарий. Специфические ядра называют также релейными, переключающими. Специфические ядра делят на сенсорные релейные и несенсорные релейные. Несенсорные релейные ядра, в свою очередь, делятся на моторные ядра и переднюю группу. Отдельные морфологи переднюю группу и ряд неспецифических ядер называют лимбическими ядрами таламуса, учитывая их проекции на лимбическую кору. Например, специфические несенсорные ядра – переднее дорсальное, переднее медиальное и переднее вентральное – проецируются на различные поля поясной извилины. Релейные ядра таламуса получают афференты от лемнисковых систем (спинальной, тригемиальной, слуховой и зрительной), от некоторых структур головного мозга (вентральное переднее ядро таламуса, мозжечок, ги-поталамус, полосатое тело) и имеют прямой выход в кору головного мозга (проекционные области, моторная и лимбическая кора).
Related news items: Newer news items: Older news items: |
flex4launch.ru