
Как отреагирует фосфатная буферная система на понижение ph – Буферные системы внутренней среды организма. Тканевые гомеостатические обменные процессы
Фосфатная буферная система — МегаЛекции
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи.
Она образована дигидрофосфатом (Nah3PO4) и гидрофосфатом (Na2HPO4) натрия.
Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами.
В норме отношение HРO42– к h3РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (h3PO4–). В результате, благодаря связыванию введенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион > h3PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+, а расход ионов Н+ восполняется повышением диссоциации Nah3PO4.
h3PO4– + Катион-ОН > Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи.
В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого Nah3PO4:
Na2HPO4 + Н2СО3 > Nah3PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам.
В кислой среде подавляется диссоциация СООН-групп, а группы Nh3 связывают избыток Н+, при этом белок заряжается положительно.
В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови.
В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве.
Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера.
Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb — более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина выглядит так:
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (к вопросу о значении пранаямы! — ALG)
В легких после удаления СО2 (угольной кислоты) происходит защелачивание крови.
При этом присоединение О2 к дезоксигемоглобину H-Hb образует кислоту ННbО2 более сильную, чем угольная. Она отдает свои ионы Н+ в среду, предотвращая повышение рН:
Н-Hb + O2 > [H-HbO2] > НbO2 + Н+
В капиллярах тканей постоянное поступление кислот (в том числе и угольной) из клеток приводит к диссоциации оксигемоглобина НbO2 (Эффект Бора) и связыванию ионов Н+ в виде Н-Hb:
НbO2+ Н+ > [H-HbO2] > Н-Hb + O2
Длительная стабилизация сдвигов рН
Это так называемая физиологическая компенсация нарушений кислотно-основного состояния, которая происходит прежде всего за счет работы дыхательной системы и почек, и в меньшей степени — за счет печени и костной системы.
Дыхательная система
Легочная вентиляция обеспечивает удаление угольной кислоты, образованной при функционировании бикарбонатной буферной системы. По скорости реакции на изменение рН – это вторая система после буферных систем.
Дополнительная вентиляция легких приводит к удалению СО2, а значит и Н2СО3, и повышает рН крови, что компенсирует закисление межклеточной жидкости и плазмы крови продуктами метаболизма, в первую очередь, органическими кислотами.
Сдвиги значений рО2 не являются сильно значимыми для изменения легочной вентиляции. Только снижение рО2 до 8 кПа в артериальной крови (норма 11,04-14,36 кПа или 83-108 мм рт.ст.) приводит к увеличению активности дыхательного центра.
Более существенным фактором для активации дыхательной системы является концентрация ионов Н+.
Накопление ионов Н+ в крови уже через 1-2 минуты вызывает максимальную (для данной их концентрации) стимуляцию дыхательного центра, повышая его активность до 4-5 раз, что приводит к снижению рСО2 до 10-15 мм рт.ст.
И, наоборот, снижение кислотности крови понижает активность дыхательного центра на 50-75%, рСО2 при этом способен возрастать до 60 мм рт.ст и выше.
Костная ткань
Это наиболее медленно реагирующая система. Механизм ее участия в регуляции рН крови состоит в возможности обмениваться с плазмой крови ионами Са2+ и Na+ в обмен на протоны Н+. Происходит растворение гидроксиапатитных кальциевых солей костного матрикса, освобождение ионов Са2+ и связывание ионов НРО42– с Н+ с образованием дигидрофосфата, который уходит с мочой.
Параллельно при снижении рН (закисление) происходит поступление ионов H+внутрь остеоцитов, а ионов калия – наружу.
Печень
Существенную, но пассивную роль в регуляции кислотно-основного состояния крови берет на себя печень, в которой происходит метаболизм низкомолекулярных органических кислот (молочная кислота и др). Кроме этого, кислые и щелочные эквиваленты выделяются с желчью.
Почки
Развитие почечной реакции на смещение кислотно-основного состояния происходит в течение нескольких часов.
Регуляция концентрации ионов H+ осуществляется опосредованно, через потоки ионов Na+, движущихся по градиенту концентрации, и через перераспределение потоков ионов К+ и Н+, которые выходят из эпителиоцитов (секретируются) в обмен на ионы Na+.
Также для обеспечения электронейтральности внутри- и внеклеточной жидкости при реабсорбции ионов Na+ усиливается реабсорбция ионов Cl–, однако их не хватает, поэтому возникает необходимость в усилении реабсорбции и дополнительном синтезе ионов HCO3– (и вот тут-то как раз и играет свою роль сода — бикарбонат натрия NaHCO3. Если мы поставляем организму дополнительное количество ионов HCO3 посредством соды, мы существенно снижаем нагрузку с почек и помогаем им в работе — ALG)
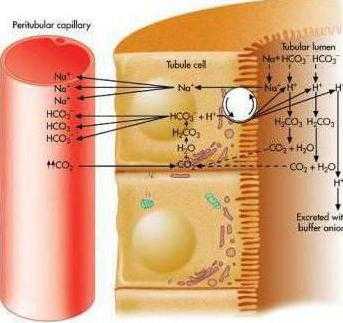
В почках активно протекают три процесса, связанных с уборкой кислых эквивалентов. Благодаря этим процессам рН мочи в состоянии снижаться до 4,5-5,2 :
1. Реабсорбция бикарбонатных ионов HCO3–.
2. Ацидогенез – удаление ионов Н+ с титруемыми кислотами (в основном в составе дигидрофосфатов Nah3PO4).
3. Аммониегенез – удаление ионов Н+ в составе ионов аммония Nh5+.
Реабсорбция бикарбонат-ионов
В проксимальных канальцах ионы Na+ мигрируют в цитозоль эпителиальных клеток в силу концентрационного градиента, который создается на базолатеральной мембране при работе фермента Na+,К+ АТФазы.
В обмен на ионы Na+ эпителиоциты канальцев активно секретируют в канальцевую жидкость ионы водорода.
Ионы HCO3– первичной мочи и секретируемые ионы Н+ образуют угольную кислоту Н2СО3.
В гликокаликсе эпителиоцитов фермент карбоангидраза катализирует распад угольной кислоты на СО2 и воду.
В результате возникает градиент концентрации углекислого газа между просветом канальцев и цитозолем и СО2 диффундирует в клетки.
Внутриклеточная карбоангидраза использует пришедший СО2 и образует угольную кислоту, которая диссоциирует.
Ионы НСО3– транспортируются в кровь, ионы Н+ – секретируются в мочу в обмен на ионы Na+. Таким образом, объем реабсорбции НСО3– полностью соответствует секреции ионов Н+.

Процесс реабсорбции бикарбонат-ионов
В проксимальных канальцах происходит реабсорбция 90% профильтрованного НСО3–.
В петле Генле и дистальных канальцах реабсорбируется оставшееся количество карбонат-иона. Всего в почечных канальцах реабсорбируется более 99% от фильтруемых бикарбонатов.
(Из всего вышесказанного становится очевидным, что дополнительное поступление в систему бикарбонат-ионов благодаря приему бикарбоната натрия — соды — снижает нагрузку на эту сторону работы почек. Чем больше свободных бикарбонат-ионов имеется в системе, тем меньше зависимость организма от этого процесса их реабсорбции. Соответственно, почки не сильно напрягаются в этом плане, в результате чего мы на выходе имеем мочу с более щелочной реакцией! Известно же, что у новорожденных показатель мочи близок к 8…ALG)
Ацидогенез
В процессе ацидогенеза в сутки с мочой выделяется 10-30 ммоль кислот, называемых титруемыми кислотами.
Фосфаты, являясь одной из этих кислот, играют роль буферной системы в моче.
Роль этой системы состоит экскреции кислых эквивалентов без потерь бикарбонат-ионов за счет дополнительного иона водорода в составе выводимого Nah3PO4 (по сравнению с NaHCO3):
Na2HPO4 + Н2СО3 > Nah3PO4 + NaНСО3
После того как бикарбонат натрия в почечных канальцах реабсорбируется, кислотность мочи зависит только от связывания ионов Н+с HPO42– и содержания дигидрофосфата.
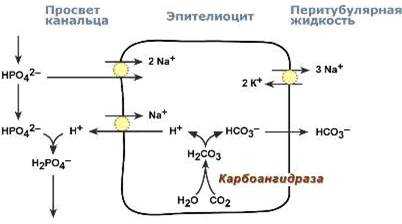
Процесс ацидогенеза в почечных канальцах
Хотя в крови соотношение HРO42– : h3РO4– равно 4 : 1, в клубочковом фильтрате оно меняется на 1 : 9.
Происходит это из-за того, что менее заряженный h3РO4– лучше фильтруется в клубочках.
Связывание ионов Н+ ионами HРO42– на протяжении всего канальца приводит к увеличению количества h3РO4–.
В дистальных канальцах соотношение может достигать 1 : 50.
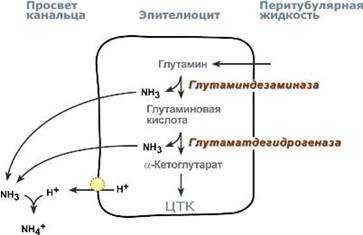
Аммониегенез
Аммониегенез происходит на протяжении всего почечного канальца, но более активно идет в дистальных отделах – дистальных канальцах и собирательных трубочках коркового и мозгового слоев. Глутамин и глутаминовая кислота, попадая в клетки канальцев, быстро дезаминируются ферментами глутаминаза и глутаматдегидрогеназа с образованием аммиака.
Являясь гидрофобным соединением, аммиак диффундирует в просвет канальца и акцептирует ионы Н+ с образованием аммонийного иона.
Источником ионов H+ первичной мочи в проксимальных отделах канальца является Na+, H+-антипорт. В дистальных отделах, в отличие от проксимальных, секреция ионов Н+ происходит с участием Н+-АТФазы, локализованной на апикальной мембране вставочных клеток.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Фосфатная буферная система
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи. Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия. Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (H2PO4–). В результате, благодаря связываниювведенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион → H2PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+ , а расход ионов Н+восполняется повышением диссоциации NaH2PO4.
H2PO4– + Катион-ОН → Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи. В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 → NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам. Вкислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно. В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Изменение заряда буферных групп белка при различных рН Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови. В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве. Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Гемоглобиновые подсистемы | |||
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера. Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb — более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (Газобмен в легких и тканях).
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина |
Н-Hb + O2 → [H-HbO2] → НbO2 + Н+
В капиллярах тканей постоянное поступление кислот (в том числе и угольной) из клеток приводит к диссоциации оксигемоглобина НbO2 (Эффект Бора) и связыванию ионов Н+ в виде Н-Hb:
НbO2+ Н+ → [H-HbO2] → Н-Hb + O2
studfiles.net
Фосфатная буферная система
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи. Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия. Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (H2PO4–). В результате, благодаря связываниювведенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион → H2PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+ , а расход ионов Н+восполняется повышением диссоциации NaH2PO4.
H2PO4– + Катион-ОН → Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи. В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 → NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам. Вкислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно. В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Изменение заряда буферных групп белка при различных рН Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови. В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве. Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Гемоглобиновые подсистемы | |||
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера. Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb — более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (Газобмен в легких и тканях).
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина |
Н-Hb + O2 → [H-HbO2] → НbO2 + Н+
В капиллярах тканей постоянное поступление кислот (в том числе и угольной) из клеток приводит к диссоциации оксигемоглобина НbO2 (Эффект Бора) и связыванию ионов Н+ в виде Н-Hb:
НbO2+ Н+ → [H-HbO2] → Н-Hb + O2
studfiles.net
классификация, пример и механизм действия
Огромную роль для нормального функционирования человеческого организма играет кислотно-основное равновесие. Циркулирующая в организме кровь представляет собой смесь живых клеток, которые находятся в жидкой среде обитания. Первой чертой охраны, что контролирует уровень pH в крови, выступает буферная система. Это физиологический механизм, который обеспечивает сохранение параметров кислотно-основного баланса, препятствуя перепадам pH. Что он собой представляет и какие имеет разновидности, узнаем ниже.
Описание
Буферная система — это уникальный механизм. В организме человека существует их несколько, и все они состоят из плазмы и клеток крови. Буферы представляют собою основания (белки и неорганические соединения), которые связывают или отдают Н+ и ОН-, уничтожая сдвиг pH в течение тридцати секунд. Способность буфера поддерживать кислотно-основный баланс зависит от количества элементов, из которых он слагается.
Виды буферов крови
Кровь, которая постоянно движется, представляет собой живые клетки, которые существуют в жидкой среде. pH в норме составляет 7,37-7,44. Связка же ионов происходит определенным буфером, классификация буферных систем приведена ниже. Сам же он состоит из плазмы и кровяных клеток и может быть фосфатным, белковым, бикарбонатным или гемоглобиновым. Все эти системы имеют достаточно простой механизм действия. Их деятельность нацелена на урегулирование уровня ионов в крови.
Особенности буфера гемоглобинового
Гемоглобиновая буферная система является самой мощной из всех, она представляет собой щелочь в капиллярах тканей и кислоту в таком внутреннем органе, как легкие. На ее долю приходится около семидесяти пяти процентов всей буферной емкости. Этот механизм участвует во множестве процессах, что происходят в крови человека, и имеет в своем составе глобин. При переходе гемоглобинового буфера в другую форму (оксигемоглобин), наблюдается изменение этой формы, изменяются и кислотные свойства действующего вещества.
Качество восстановленного гемоглобина меньшее, чем у угольной кислоты, но становится намного лучше, когда он окисляется. Когда приобретается кислотность pH, гемоглобин соединяет ионы водорода, получается так, что он уже восстановленный. Когда происходит очищение углекислого газа в легких, pH получается щелочным. В это время гемоглобин, который окислился, выступает донором протонов, при помощи чего происходит уравновешивание кислотно-основного баланса. Так, буфер, что состоит из оксигемоглобина и его калиевой соли, способствует выделению из организма углекислоты.
Эта буферная система выполняет немаловажную роль в дыхательном процессе, так как совершает транспортную функцию по переносу к тканям и внутренним органам кислорода и удалению из них углекислоты. Кислотно-основное равновесие внутри эритроцитов при этом придерживается на постоянном уровне, следовательно, в крови также.
Таким образом, когда кровь насыщается кислородом, гемоглобин превращается в сильную кислоту, а когда кислород он отдает, то превращается в достаточно слабую органическую кислоту. Системы оксигемоглобина и гемоглобина — взаимопревращающиеся, они существуют как одно целое.
Особенности бикарбонатного буфера
Бикарбонатная буферная система выступает также мощной, но и самой управляемой в организме. На ее долю приходится около десяти процентов всей буферной емкости. Она имеет универсальные свойства, которые обеспечивают ее двухстороннюю эффективность. В состав этого буфера входит сопряженная кислотно-щелочная пара, что состоит из таких молекул, как угольная кислота (источник протона) и бикарбонат аниона (акцептор протона).
Так, бикарбонатная буферная система способствует протеканию систематического процесса, где в кровь попадает мощная кислота. Этот механизм связывает кислоту с бикарбонатом анионов, образуя кислоту угольную и ее соль. При попадании щелочи в кровь буфер связывается с угольной кислотой, образуя бикарбонатную соль. Так как гидрокарбоната натрия в крови человека больше, чем угольной кислоты, данная буферная емкость будет иметь высокую кислотность. Другими словами, гидрокарбонатная буферная система (бикарбонатная) очень хорошо проводит компенсацию веществ, которые повышают кислотность крови. К ним относится и молочная кислота, концентрация которой увеличивается при интенсивных физических нагрузках, а данный буфер очень быстро реагирует на изменения кислотно-основного баланса в крови.
Особенности фосфатного буфера
Фосфатная буферная система человека занимает близко двух процентов всей буферной емкости, что связано с содержанием в крови фосфатов. Этот механизм поддерживает показатель pH в моче и жидкости, что находится внутри клеток. Буфер состоит из неорганических фосфатов: одноосновного (выполняет роль кислоты) и двухосновного (выполняет роль щелочи). При нормальном показателе pH соотношение кислоты и основания равняется 1:4. При увеличении количества ионов водорода фосфатная буферная система связывается с ними, образуя кислоту. Этот механизм больше кислотный, чем щелочной, поэтому он отлично нейтрализует поступающие в кровь человека кислые метаболиты, например, молочную кислоту.
Особенности белкового буфера
Белковый буфер играет не такую особую роль в стабилизации кислотно-щелочного баланса, по сравнению с другими системами. На его долю приходится около семи процентов всей буферной емкости. Белки состоят из молекул, которые объединяются в кислотно-щелочные соединения. В кислой среде они выступают щелочами, которые связывают кислоты, в щелочной среде все происходит наоборот.
Это приводит к тому, что образуется белковая буферная система, которая достаточно эффективна при значении pH от 7,2 до 7,4. Большая доля белков представлена альбуминами и глобулинами. Так как белковый заряд нулевой, то при нормальном показателе pH он находится в виде щелочи и соли. Эта буферная емкость зависит от количества белков, их структуры и свободных протонов. Данный буфер может нейтрализовать и кислые, и щелочные продукты. Но емкость она имеет больше кислотную, чем щелочную.
Особенности эритроцитов
В норме эритроциты имеют постоянный показатель pH – 7,25. Здесь оказывают действие гидрокарбонатный и фосфатный буферы. Но по мощности они отличаются от тех, что находятся в крови. В эритроцитах белковый буфер играет особую роль в обеспечении органов и тканей кислородом, а также удалению из них углекислоты. Кроме этого, он поддерживает постоянное значение внутри эритроцитов pH. Белковый буфер в эритроцитах тесно связан с гидрокарбонатной системой, так как соотношение кислоты и соли здесь меньшее, чем в крови.
Пример буферной системы
Растворы сильных кислот и щелочей, которые обладаю слабыми реакциями, имеют непостоянный показатель pH. Но смесь кислоты уксусной с ее солью сохраняет имеет стабильное значение. Даже если к ним добавить кислоту или щелочь, кислотно-основное равновесие не изменится. В качестве примера можно рассмотреть ацетатный буфер, который состоит из кислоты СН3СООН и ее соли СН3СОО. Если добавить сильную кислоту, то основание соли свяжет ионы Н+ и превратится в кислоту уксусную. Снижение уровня анионов соли уравновешивается увеличением молекул кислоты. В результате этого наблюдается незначительное изменение в соотношении кислоты и ее соли, поэтому pH изменяется совсем незаметно.

Механизм действия буферных систем
При поступлении в кровь кислых или щелочных продуктов буфер обеспечивает постоянное значение pH до тех пор, пока поступившие продукты не выведутся или не используются в процессах метаболизма. В крови человека представлены четыре буфера, каждый из которых состоит из двух частей: кислоты и ее соли, а также сильной щелочи.
Эффект буфера обуславливается тем, что он связывает и нейтрализует ионы, которые поступают соответствующим ему составом. Поскольку в природе организм больше всего сталкивается с недоокисленными продуктами обмена, свойства буфер имеет антикислотные больше, чем антищелочные.
Каждая буферная система имеет свой принцип работы. При снижении уровня pH ниже отметки 7,0 начинается их активная деятельность. Они начинают связывать излишки свободных ионов водорода, образуя комплексы, которые перемещают кислород. Он, в свою очередь, перемещается к системе пищеварения, легким, коже, почкам и так далее. Такая транспортировка кислых и щелочных продуктов способствует их разгрузке и выведению.
В организме человека только четыре буферные системы играют важные роли в сохранении кислотно-основного равновесия, но существуют и другие буферы, например, ацетатная буферная система, которая имеет слабую кислоту (донор) и ее соль (акцептор). Способность этих механизмов противостоять изменениям pH при попадании кислоты или соли в кровь является ограниченной. Они поддерживают кислотно-щелочное равновесие только в том случае, когда сильная кислота или щелочь поступают в определенном количестве. Если оно будет превышено, pH резко изменится, буферная система прекратит свое действие.
Эффективность буферов
Буферы крови и эритроцитов имеют различную эффективность. У последних она выше, так как здесь присутствует гемоглобиновый буфер. Уменьшение количества ионов происходит по направлению от клетки до межклеточной среды, а затем до крови. Это говорит о том, что самая большая буферная емкость у крови, а меньшую имеет внутриклеточная среда.
При метаболизме в клетках появляются кислоты, которые проходят в межклеточную жидкость. Это происходит тем легче, чем их больше появляется в клетках, поскольку переизбыток ионов водорода увеличивает проницаемость мембраны клетки. Нам уже известна классификация буферных систем. В эритроцитах они имеют более эффективные свойства, так как здесь еще играют роль коллагеновые волокна, которые реагируют набуханием на накопление кислоты, они ее поглощают и освобождают от ионов водорода эритроциты. Такая его способность обуславливается свойством абсорбции.

Взаимодействие буферов в организме
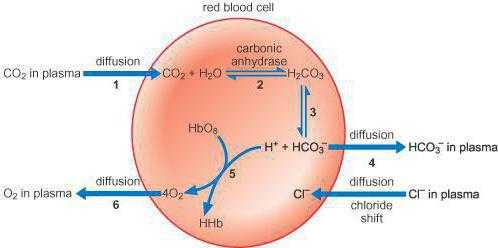
Все механизмы, которые находятся в организме, взаимосвязаны между собой. Буферы крови состоят из нескольких систем, вклад которых в поддержание кислотно-щелочного баланса различный. При попадании крови в легкие она получает кислород путем его связывания в эритроцитах гемоглобином, образуя оксигемоглобин (кислоту), что поддерживает уровень pH. При содействии карбоангидразы происходит параллельное очищение крови легких от углекислоты, которая в эритроцитах представлена в виде слабой двухосновной угольной кислотой и карбаминогемоглобином, а в крови – углекислотой и водой.
При уменьшении в эритроцитах количества слабой двухосновной угольной кислоты происходит проникновение ее из крови в эритроцит, и очищение крови от углекислоты. Таким образом, из клеток в кровь постоянно проходит слабая двухосновная угольная кислота, а из крови в эритроциты для соблюдения нейтральности поступают неактивные анионы хлорида. В результате этого в красных клетках крови среда более кислотная, чем в плазме. Все системы буферов обосновываются отношением донор-акцептор протона (4:20), что связано с особенностями метаболизма организма человека, который образует большее число кислотных продуктов, чем щелочных. Очень важным здесь является показатель кислотных буферных емкостей.

Обменные процессы в тканях
Кислотно-основной баланс поддерживается буферами и метаболическими превращениями в тканях организма. Этому помогают биохимические и физико-химические процессы. Они способствуют потере кислотно-щелочных свойств продуктов обмена веществ, их связыванию, образованию новых соединений, которые быстро выводятся из организма. Например, большое количество молочной кислоты выводится в гликоген, органические кислоты нейтрализуются солями натрия. Сильные кислоты и щелочи растворяются в липидах, а органические кислоты подвергаются окислению, образуя угольную кислоту.
Таким образом, буферная система — это первый помощник при нормализации кислотно-щелочного баланса в организме человека. Стабильность pH нужна для нормальной работы биологических молекул и структур, органов и тканей. При нормальных условиях буферные процессы поддерживают равновесие между появлением и удалением ионов водорода и углекислого газа, что способствует обеспечению в крови постоянного уровня pH.
Если происходит сбой в работе буферных систем, то у человека появляются такие патологии, как алкалоз или ацидоз. Все буферные системы взаимосвязаны и направлены на поддержание стабильного кислотно-основного равновесия. В организме человека постоянно образуется большое число кислых продуктов, которое эквивалентно тридцати литрам сильной кислоты.
Постоянство реакций внутри организма обеспечивают мощные буферы: фосфатный, белковый, гемоглобиновый и бикарбонатный. Существуют и другие буферные системы, но эти являются основными и самыми нужными для живого организма. Без их помощи у человека начнут развиваться различные патологии, которые могут привести к коме или летальному исходу.
fb.ru
Фосфатная буферная система — Мегаобучалка
Кислотно-основное равновесие.
Кислотно-основное равновесие – это соотношение концентрации водородных (Н+) и гидроксильных (ОН—) ионов в жидкостях организма.
Постоянство рН внутренней среды организма обусловлено совместным действием буферных систем и ряда физиологических механизмов.
1. Буферные системы крови и тканей:
Бикарбонатная: NaHCO3 + Н2СО3
Фосфатная: NaHРO4с + NaHРO4к
Белковая: протеин-Na+ + протеин-Н+
Гемоглобиновая: HbK+HbH+
2. Физиологический контроль:
Дыхательная функция легких
Выделительная функция почек
КЩР отражает клеточный метаболизм, газотранспортную функцию крови, внешнее дыхание и водно-солевой обмен.
В норме рН крови колеблется от 7,37 до 7,44, среднее значение рН=7,4.
Буферные системы поддерживают постоянство рН при поступлении кислых и основных (ОН—) продуктов. Буферное действие объясняется связыванием свободных Н+ и ОН— ионов компонентами буфера и переводом их в недиссоциированную форму слабой кислоты или воды.
Буферные системы организма состоят из слабых кислот и их солей с сильными основаниями.
Для устранения сдвига рН необходимо различное время:
Буферные системы – 30 сек
Дыхательный контроль – 1 – 3 мин
Выделительная функция почек – 10 – 20 час.
Буферные системы устраняют только сдвиги рН. Физиологические механизмы восстанавливают и буферную емкость.
Бикарбонатная буферная система.
На долю бикарбонатного буфера приходится около 10% всей буферной емкости крови.
Бикарбонатный буфер состоит из угольной кислоты, выполняющей роль донора протона, и бикарбонат-иона , выполняющего роль акцептора протона.
Н2СО3 – слабая кислота, трудно диссоциирует
Н2СО3 Н+ +
NaНСО3 – соль слабой кислоты и сильного основания диссоциирует полностью:
NaНСО3 Na+ +
Механизм действия буфера
1. При поступлении в кровь кислых продуктов водородные ионы взаимодействуют с ионами бикарбоната , образуется слабо диссоциирующая угольная кислота:
Н+ + NaНСО3 Na+ + Н2СО3
Восстанавливается соотношение Н2СО3/ NaНСО3, рН не изменяется (концентрация NaНСО3 незначительно понижается).
Легкие обеспечивают выведение углекислого газа.
2. При поступлении в кровь оснований из тканей, ионы ОН— взаимодействуют со слабой угольной кислотой (ионы ОН— взаимодействуют с Н+ из буфера, образуя Н2О)
Н2СО3 + ОН— Н2О +
рН сохраняется, [ ] увеличивается. Избыток усиливает диссоциацию Н2СО3, расход Н+ восполняется усилением диссоциации Н2СО3.
При нормальном значении рН крови концентрация ионов бикарбоната в плазме крови превышает концентрацию углекислого газа примерно в 20 раз:
Фосфатная буферная система
Компоненты буфера:
Na2НРО4с – соль – двузамещенный фосфат
NaН2РО4к – слабая кислота – однозамещенный фосфат
Соотношение
На долю фосфатной буферной системы приходится 1% буферной емкости крови.
Механизм действия буфера.
1. При поступлении в кровь кислых продуктов обмена веществ происходит связывание ионов Н+ с двузамещенным фосфатным ионом , образуется кислый однозамещенный ион , избыток которого удаляется почками с мочой:
Н+ + Na2НРО4 Na+ + Na2НРО4 выделяется почками.
2. При поступлении в кровь ионов ОН— происходит связывание избытка ОН— при взаимодействии с однозамещенным фосфатом (кислота) Na2НРО4. Избыток ОН— связывается с водородом (Н+) из с образованием воды:
ОН— + Na2НРО4 NaНРО + Н2О
Понижение концентрации [NaН2РО4] сдвигает диссоциацию NaН2РО4.
NaН2РО4 NaНРО + Н+ и восстанавливается недостаток ионов водорода.
Действие фосфатного буфера контролируется выделительной функцией почек. При ее нарушении происходит задержка кислых фосфатов.
Фосфатный буфер действует при изменении рН в интервале от 6,1 до 7,7. В крови максимальная емкость фосфатного буфера проявляется при 7,2.
megaobuchalka.ru
— быстрая компенсация — Биохимия
Буферные системы
Буферные системы – это соединения, противодействующие резким изменениям концентрации ионов Н+. Любая буферная система — это кислотно-основная пара: слабое основание (анион, А–) и слабая кислота (Н-Анион, H-А). Они минимизируют сдвиги количества ионов Н+ за счет их связывания с анионом и включения в плохо диссоциирующее соединение – в слабую кислоту. Поэтому общее количество ионов Н+ изменяется не так заметно, как это могло бы быть.
Существует три буферные системы жидкостей организма – бикарбонатная, фосфатная, белковая, гемоглобиновая.Они вступают в действие моментально и через несколько минут их эффект достигает максимума возможного.
Фосфатная буферная система
Фосфатная буферная система составляет около 2% от всей буферной емкости крови и до 50% буферной емкости мочи. Она образована гидрофосфатом (HPO42–) и дигидрофосфатом (H2PO4–). Дигидрофосфат слабо диссоциирует и ведет себя как слабая кислота, гидрофосфат обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов Н+) с двузамещенным фосфатом (HPO42‑) образуется дигидрофосфат (H2PO4–):
Удаление ионов H+ фосфатным буфером
В результате концентрация ионов Н+ понижается.
При поступлении в кровь оснований (избыток ОН–‑групп) они нейтрализуются поступающими в плазму от H2PO4– ионами Н+:
Удаление щелочных эквивалентов фосфатным буфером
Роль фосфатного буфера особенно высока во внутриклеточном пространстве и в просвете почечных канальцев. Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата (h3PO4–), т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
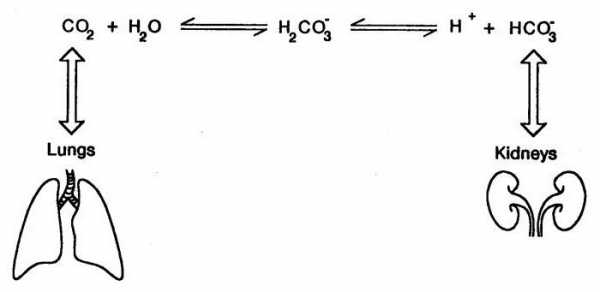
Бикарбонатная буферная система
Эта система самая мощная, на ее долю приходится 65% всей буферной мощности крови. Она состоит из бикарбонат-иона (НСО3–) и угольной кислоты (Н2СО3). В норме отношение HCO3– к H2CO3 равно 20 : 1.
При поступлении в кровь ионов H+ (т.е. кислоты) ионы бикарбоната натрия взаимодействуют с ней и образуется угольная кислота:
При работе бикарбонатной системы концентрация водородных ионов понижается, т.к. угольная кислота является очень слабой кислотой и плохо диссоциирует. При этом в крови не происходит параллельного значимого увеличения концентрации НСО3–.
Если в кровь поступают вещества с щелочными свойствами, то они реагируют с угольной кислотой и образуют ионы бикарбоната:
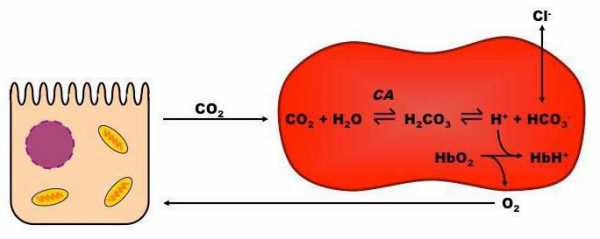
Работа бикарбонатного буфера неразрывно связана с дыхательной системой (с вентиляцией легких). В легочных артериолах при снижении плазменной концентрации СО2 и благодаря присутствию в эритроцитах фермента карбоангидразы угольная кислота быстро расщепляется с образованием CO2, удаляемого с выдыхаемым воздухом:
Н2СО3→ Н2О + СО2↑
Кроме эритроцитов, значительная активность карбоангидразы отмечена в эпителии почечных канальцев, клетках слизистой оболочки желудка, коре надпочечников и клетках печени, в незначительных количествах – в центральной нервной системе, поджелудочной железе и других органах.
Белковая буферная система
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам. Их вклад в буферизацию плазмы крови около 5%.
В кислой среде подавляется диссоциация СООН‑групп аминокислотных радикалов (в аспарагиновой и глутаминовой кислотах), а группы NH2 (в аргинине и лизине) связывают избыток Н+. При этом белок заряжается положительно.
В щелочной среде усиливается диссоциация COOH‑групп, поступающие в плазму ионы Н+ связывают избыток ОН–‑ионов и pH сохраняется. Белки в данном случае выступают как кислоты и заряжаются отрицательно.
Изменение заряда буферных групп белка при различных рН
Гемоглобиновая буферная система
Высокой мощностью в крови обладает гемоглобиновый буфер, на него приходится до 28% всей буферной емкости крови. В качестве кислой части буфера выступает оксигенированный гемоглобин H‑HbO2. Он имеет выраженные кислотные свойства и в 80 раз легче отдает ионы водорода, чем восстановленный Н‑Нb, выступающий как основание. Гемоглобиновый буфер можно рассматривать как часть белкового, но его особенностью является работа в теснейшем контакте с бикарбонатной системой.
Изменение кислотности гемоглобина происходит в тканях и в легких, и вызывается связыванием соответственно H+ или О2. Непосредственный механизм действия буфера заключается в присоединении или отдаче иона H+ остатком гистидина в глобиновой части молекулы (эффект Бора).
В тканях более кислый pH в норме является результатом накопления минеральных (угольной, серной, соляной) и органических кислот (молочной). При компенсации pH данным буфером ионы H+ присоединяются к пришедшему оксигемоглобину (HbО2) и превращают его в H‑HbО2. Это моментально вызывает отдачу оксигемоглобином кислорода (эффект Бора) и он превращается в восстановленный H‑Hb.
НbO2+ Н+ → [H-HbO2] → Н-Hb + O2
В результате снижается количество кислот, в первую очередь Н2СО3, продуцируются ионы НСО3‑ и тканевое пространство подщелачивается.
В легких после удаления СО2 (угольной кислоты) происходит защелачивание крови. При этом присоединение О2 к дезоксигемоглобину H-Hb образует кислоту более сильную, чем угольная. Она отдает свои ионы Н+ в среду, предотвращая повышение рН:
Н-Hb + O2 → [H-HbO2] → НbO2 + Н+
Работу гемоглобинового буфера рассматривают неотрывно от бикарбонатного буфера:
Вы можете спросить или оставить свое мнение.
biokhimija.ru
Фосфатная буферная система
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи. Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия. Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (H2PO4–). В результате, благодаря связываниювведенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион → H2PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+ , а расход ионов Н+восполняется повышением диссоциации NaH2PO4.
H2PO4– + Катион-ОН → Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи. В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 → NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам. Вкислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно. В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Изменение заряда буферных групп белка при различных рН Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови. В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве. Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Гемоглобиновые подсистемы | |||
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера. Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb — более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (Газобмен в легких и тканях).
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина |
Н-Hb + O2 → [H-HbO2] → НbO2 + Н+
В капиллярах тканей постоянное поступление кислот (в том числе и угольной) из клеток приводит к диссоциации оксигемоглобина НbO2 (Эффект Бора) и связыванию ионов Н+ в виде Н-Hb:
НbO2+ Н+ → [H-HbO2] → Н-Hb + O2
studfiles.net