
Эволюция человека антропогенез – Происхождение и эволюция человека. Презентация. Основные этапы, стадии
Антропогенез человека — факторы, стадии, теория антропогенеза
Антропогенез – это становление человеческой особи в историко-эволюционном аспекте, его физическое формирование, начальное развитие речи, деятельности и общинный способ проживания. В ходе антропогенеза состоялось появление Homo sapiens, его отделение и обособление от обезьян и млекопитающих. Антропогенез человека изучает ряд наук, в основном антропология, палеоантропология, а также лингвистика, генетика, физиология, этнография. Важнейшими факторами процесса антропогенеза были: прямохождение человека, использование разных вещей для добывания еды, изготовление орудий для труда, стадное существование, возникновение языка. Существует много мыслей и соображений об антропогенезе человека, но сегодня самой научно обусловленной считается теория Чарльза Дарвина.
Что такое антропогенез?
Историю появления людей, развитие их рода изучать стали еще в XVIII веке. До сего времени постоянным и центральным было убеждение, что мир, все живое, как и неживое сотворил Бог таким, каковыми они являются на текущий момент. Но с прогрессом науки, проведений множеств исследований началось изменяться мнение о создании мира. На смену убеждению о неизменности всего существующего пришло понимание об эволюции, становление всего живого. Значимое место во всем этом занял антропогенез человека, его формирование, происхождение, отделение, развитие.
Изучением антропогенеза человеческой расы занимались многие ученые, начиная с Карла фон Линнея (шведского ученого, биолога, зоолога, врача), который классифицировал человека в одном ряду с человекоподобными обезьянами, отнеся его к животному миру. Огромный вклад в обоснование теории об антропогенезе имели исследования Буше де Перта, французского археолога, который открыл наличие каменного орудия, что применялись первобытным человеком в эпоху мамонтов. Долгое время такие открытия антропогенеза в науке не признавались и встречали бурю сопротивления, так как противоречили Библии.
Проблема антропогенеза существует в нескольких вопросах: датировка и место происхождения первых людей; дискуссии об основных стадиях антропогенеза, влияние факторов антропогенеза в разные периоды развития; соотношение физического влияния в антропогенезе с социальным; формирование первых общин.
Проблемы антропогенеза изучаются с помощью исследований нескольких наук. В значительной степени антропологии и палеоантропологии, а также психологии, физиологии, лингвистики, морфологии, археологии, этнографии и др.
Переворот в знаниях об антропогенезе человека сделали идеи Чарльза Дарвина. Согласно теории дарвинизма человеческая раса эволюционировала от обезьянообразного предка. Исследователи пришли к решению, что человекоподобные обезьяны – дальние предки нашей расы, так как они анатомически схожи с человеком. Центральным в антропогенезе человека, его адаптации к изменившимся условиям существования Дарвин обозначал естественный отбор. Теория трудовой деятельности Энгельса утверждает, что центральным фактором исторической эволюции человека, его антропогенеза есть трудоспособность, умение работать в условиях общины.
Антропогенез человека отличен от эволюции органического мира тем, что последняя регулируется только природными законами, а осознание своих возможностей в деятельности позволило человеку воздействовать на природу и уменьшить воздействие биологических факторов. Дарвиновская симидная теория антропогенеза учла в себе разные научные взгляды и аргументировала происхождение Homo sapiens от приматов. Подтверждение этому служит сходство живущих теперь человекоподобных обезьян из нынешним человеком в анатомическом строении, формою зародышей, физиологическими показателями. Дарвин доказал, что вся человеческая раса пошла от одного вида обезьян, и был уверен в становлении древних людей в Африке.
Проблема антропогенеза еще в том, что нет остаточного решения о древней родине человека. Одни ученые полагают, человек пошел из местностей Африки, другие – с южной Евразии, исключается только Австралия, Америка и северная Евразия.
Факторы антропогенеза
Основополагающими факторами антропогенеза человека считаются биологические и социальные моменты.
Антропогенез – это происхождение вида человеческой расы в физиологическом аспекте. Социальный фактор – это становление людского общества. Дарвин отмечал важность биологических обстоятельств в формировании человеческого существа. Такие факторы как естественный (природный) отбор, наследственность, склонность изменяться отыгрывали значимую роль на первоначальных этапах антропогенеза. Изменчивость предопределяет появление новых особенностей и функций в анатомическом строении человека. Наследственность укрепляет и передает в поколения эти преобразования. Как следствие естественного отбора сумели выжить сильнейшие и наиболее приспособленные. Значимость социальных факторов (мышление, умение разговаривать, стремление к общности, трудовая деятельность) в антропогенезе описывал Ф.Энгельс в собственных исследованиях о ценности труда в становлении из обезьяны человека.
Как уверяет наука, нашими пращурами были человекообразные обезьяны, что жили в лесу. В ходе изменения климата, уменьшения лесной территории они были принуждены приспосабливаться: становиться на ноги, чтоб мочь быстрее ориентироваться в новой местности. Постепенно прямохождение стало удобным и выгодным качеством нового вида человекоподобных, теперь передние конечности могли выполнять ряд новых функций трудовой деятельности.
Много стало изменяться в ходе антропогенеза человека: привлечение к труду, группирование в общины, коллективные виды работы, охота, защита общины от врагов. Все это побуждало искать способы общаться один с одним. Поначалу это было примитивное общение посредством жестов рук, звуков, дальнейшие коммуникации привели к неизбежности появления второй сигнальной системы — речи. Так, на телесном уровне прошли изменения ротового аппарата и строения гортани для формирования речи. Умение говорить, трудоспособность, пребывание в общине начало формировать мышление. Вследствие этого мозг увеличился в размерах, сформировалась кора головного мозга.
Одним из главенствующих биологических факторов антропогенеза считается естественный отбор, благодаря которому в разные периоды развития сохраняются выгодные для приспособления изменения и уничтожаются неблагоприятные, вредные качества. Следовательно, антропогенез человека привел его к самому оптимальному усовершенствованию в условиях его проживания, и эти сформировавшиеся особенности переходили по наследству.
Человек стал применять огонь с целью приготовления еды. Данный фактор способствовал изменению формы лица человека, его жевательного аппарата, пищеварительной системы для полноценного усвоения обработанной термически еды. Использование пламени для обогрева своих мест проживания дало способность человеку расселиться в местностях с холодным климатом.
На первых периодах антропогенеза центральное значение в развитии человека отыгрывали биологические факторы. Благодаря естественному отбору сформировались морфологические характеристики человека: s-образный позвоночник для ходьбы в вертикальном состоянии, широкая кость стопы, грудная клетка, строение мозга. Во время антропогенеза человек сумел так адаптироваться к окружающей природе, что его изменения стали менее независимы от воздействия естественного отбора. Позже человеческая особь уже овладела умением пользоваться и изготовлять орудие для работы, могла приготавливать пищу, оборудовать жилье, проживать в общинах, не зависеть от влияния природных процессов. Воздействие биологических факторов снижалось, а действие социальных увеличивалось.
Антропогенез человека вне общины становился невозможным. Хоть биологические факторы потеряли главную роль во влиянии на формирование вида человека, все же естественный отбор отыгрывает стабилизирующую функцию, а мутация сохраняет свое действие и в нынешнем мире. Иногда частота и сила мутаций даже увеличивается в некоторых регионах нашей планеты из-за разного рода загрязнений. В ряду с ослаблением воздействия естественного отбора, мутации могут приводить к ухудшению качественных характеристик выживания человека.
Подводя итоги, заметим, что главными факторами антропогенеза человека были биологические и социальные. Под их воздействием произошла эволюция человеческой расы. Физиологические качества переходили по наследству, а речь, умение думать, склонность к трудовой деятельности формируются в социуме во время образования и воспитания.
Стадии антропогенеза
Антропогенез, как центральный процесс становления социума и исторического становления человека, зависимо от смены трудовой деятельности, формирования сознания общины, изменений в физиологической структуре тела, был разбит на ряд стадий. Согласно одним источникам исследователи обозначают три стадии антропогенеза:
— антропоидные предки – это приматы, двигающиеся на двух нижних конечностях, которые умели использовать обычные предметы (камни, кости животных, палки) в роли орудия деятельности;
— архантропы и палеоантропы, так называемые, древнейшие и древние люди, — начали изготовлять орудие, охотились, создавали общины, были пещерными жителями, пользовались огнем. Внешний вид отдаленно схож на нынешнего человека. Отличаются они тем, что имели толстую дугу над бровями, низкий лоб, выступающий затылок. Строение мозга очень примитивное;
— неоантропы – имели физическое строение нынешнего человека, изменилась форма черепной кости, увеличился мозга объем, высокий рост. Они умели разговаривать на примитивном уровне, собирали овощи, фрукты, проводили охоту, строили жилье, создавали наскальные рисунки, шили одежду. Начало – эпоха позднего палеолита.
За другими данными стадии антропогенеза дифференцировали на пять этапов.
Погнидно-гоминидная – стадия формирующегося человека, 16-18 млн. лет назад. Это наиболее ранний период антропогенеза, который представляли: амфипитек, открыт около Бирмы, олигопитек – в Египте, египтопитек, группа дриопитеков, возраст близко 20 млн. лет назад. Все представители первых гоминид проживали стадным способом, имели незначительное подобие высшим человекообразным.
Прегоминидная стадия (австралопитеков, человекоподобных обезьян) около 5-2 млн. лет назад. Австралопитеки – двуногие гоминиды, что жили в широтах Африки. Австралопитеки существовали афарские, африканские, робустусы. Их рост был не более 1 м 30 см, вес до 40 кг, объем мозга – 700 куб.см. Самые поздние австралопитековые (президжантропы) умели уже изготовлять примитивное оружие, создали раннюю галечную культуру. Это первые образцы человека умелого, какого позже заменил человек прямоходящий. Развитие австралопитеков есть тупиковой ветвью антропогенеза человека.
Архантропная стадия – их назвали древнейшими людьми, сюда относится находка француза Дюбуа на острове Ява древних гоминид, названого питекантропом. Ростом питекантропы были 1 м 70 см, мозг был объемом до 1000 куб.см., выпирающий лоб, массивные дуги над бровями, весомая челюсть, двигались они на полусогнутых нижних конечностях. Представитель древнейших назывался человеком выпрямленным. Архантропы выявлены были в Южной Африке, Азии. Выявлены в Китае фрагменты синантропа, очень похожего по физиологическому строению на архантропов. В Европе самая давняя находка питекантропа – гейдельбергский человек. Архантропы уже владели начальной формой речи, пользовались огнем, проживали 2 – 0,5 млн. лет назад.
Четвертая стадия – стадия палеоантропов, назваными древними людьми, представителями какой будут неандертальцы. Проживали они приблизительно от 0,5 млн. — 30 тысяч лет назад. Древнейшие раскопки неандертальцев открыты в Германии, их ветвь развития была тупиковой. Другая часть ветви развития палеоантропов – близкий сородич неандертальского человека, которая вымерла, прожив около 70 тысяч лет. Имеют много похожего с нынешним человеком: строение мозга почти не различается, объем даже несколько больше – 1450 против 1350 кубичных сантиметров. Орудие для труда, охоты уже различно по виду и по назначению, особенность – это орудие, сложенное из пластин, отделяемых из каменного ядра. Показательно для этой стадии антропогенеза: групповая охота на очень крупного дикого животного, строительство примитивных жилищ, развитие речи с целью общения внутри племени.
Представителями палеоантропов есть ранние и поздние европейские неандертальцы, переднеазиатские неандертальцы. Данный вид человекоподобных отличается массивностью телосложения, выступающей лицевой половиной черепа, развитой надбровной частью, широкими носовыми отверстиями, увеличенной височной передней и средней долей мозга, усовершенствованным артикуляционным аппаратом. Анатомическое строение этого вида людей имело особенности к адаптации в суровом климате Европы. Хотя антропогенез человека этой стадии и двигался к развитию, но палеоантропы были слишком подвержены воздействию природных обстоятельств. Виды древних людей под действием неблагоприятных условий проживания, в силу свей низкой организованности и сознательной культуры приобрели морфологические качества, что задерживали их эволюционирование.
В поздний период стадии антропогенеза палеоантропов исследователями обнаружены первые захоронения. Этому свидетельствует найденные раскопки похороненного неандертальского мальчика в Узбекистане около 45 тысяч лет назад. Данный факт имел вид сознательного погребения с ритуалами. Известно около шестидесяти погребений неандертальцев.
На местностях Украины, Белоруссии, России, остальных странах СНГ также обнаружены жилища и стоянки неандертальцев.
Неоантропная стадия (Homo sapiens) – последняя стадия антропогенеза, прототипами которого были люди схожего физического образца с теперешним человеком – кроманьонцы, жившие около 200 — 50 тыс. лет тому назад. Начальные раскопки кроманьонцев открыты во Франции, их распределение по миру широко: арктические регионы, Америка, Австралия, Европа, регионы бывшего СССР.
Проблема антропогенеза данной стадии – это отсутствие определения четкого времени возникновения неоантропов. До недавнего определялась самой древней раскопка сформированного полностью неоантропа из Калимантана в пещере Ниа – 39 тысяч 600 лет назад. Археологи считают начало периода палеолита (40-35 тыс. лет назад) отсчетом времени рождения человека разумного. Также существуют взгляды антропологов о ранней организации Homo sapiens. Поиски ответа о дате возникновения неоантропа продолжаются. Так в Эфиопии в 1969 году найдены останки черепов подобных физиологическим строением на неоантропов, которые датированные 130 тыс. лет тому назад. В конце 20 ст. были раскрыты в раскопках африканских пещер следы проживания неоантропов на землях Африки многое раньше, чем в ландшафтах Европы — на 50 тыс. лет. Поэтому возможно рождение Homo sapiens намного ранее признанных дат.
Есть две гипотезы антропогенеза неоантропов. Первая – происхождение из Субсахары более 100 000 лет назад, потом неоантропы распространились по азиатских областях и 30 тыс. лет назад выдавили поздние виды неандертальцев. Вторая — ранние африканские гоминиды эволюционировали в человека разумного своим естественным путем.
За анатомическим строением кроманьонцы были подобные много в чем на нынешнего человека за формой черепа, формой нижней из челюстей с наличием большого подбородка, узким носом, прямым лбом, рост имели 180-190 сантиметров. Кроманьонцы умели создавать оружие из костей животных, камня, обнаружены на стенах пещер рисунки процесса охоты, животных. Центральное значение в антропогенезе неоантропов и их широкому расселению по миру было умение общаться. Посредством общения передавать ценную информацию о накопленном опыте, умениях, производств от племен к племенам, из поколения в следующее поколение. Выживали те племена, в каких социальная организованность, групповая деятельность была на значительном уровне.
Значимым моментом для выживания рода было приручение животных, их одомашнивание, а также занятие земледелием, окультуривание растений, что дало реальный шанс человеку пережить голод. Общение давало людям возможность сохранять и упорядочивать знания, конкретные технические навыки, наблюдения за законами природы, устанавливать правила внутри общины для большей продуктивности коллектива, его самовыживания и размножения. Постепенно влияние окружающей природы на антропогенез человека уменьшалось и утрачивало контроль. В последствии естественный отбор перестал быть значимым в антропогенезе Homo sapiens и, в следствие тому, эволюция человеческого вида прекратилась.
Подытоживая вышесказанное, следует отметить, что антропогенез не был процессом линейным и плавным. На каждой фазе антропогенеза формировались виды нескольких течений, и каждый шел своим путем развития. Под действием природных, а также социальных факторов развивалось несколько разновидностей гоминид.
psihomed.com
4.6. Антропогенез – биологическая эволюция человека
Особенность человека состоит в том, что он, будучи сложной и целостной системой, в то же время входит в качестве составного элемента в две другие системы – биологическую и социальную. Человек принадлежит одновременно природе и сложившемуся в ходе его развития социальному миру. Выступая субъектом научного познания, человек составляет одновременно и объект естественно-научного познания.
Проблема антропогенезасостоит в прослеживании превращения биологического организма, представляющего собой определенное звено в цепи эволюции, в существо социальное – носителя не только разума, но и культуры.
Научному осмыслению проблемы антропогенеза в значительной степени способствовало утверждение эволюционной теории, однако даже в рамках современной картины мира антропогенез выглядит задачей со многими неизвестными. Дело в том, что и дарвинизм, и синтетическая теория эволюции, впитавшая в себя новейшие достижения генетики, не затрагивают роли трудовой деятельности в процессе антропогенеза. Сторонники трудовой теории антропогенезаименно с трудовой деятельностью связывают (и не без оснований) развитие речи, мышления, сотрудничества людей, приведшее к их объединению в социальные коллективы.
Безусловно, трудовая деятельность стала одним из важнейших движущих факторов антропосоциогенеза. Примечательным моментом здесь является изготовление в процессе трудовой деятельностиорудий труда, в которых синтезированы способ мышления человека, его умения, навыки, знания, т. е. то, что составляет социальный опыт человечества на определенном этапе его развития. Являясь основным средством хранения и передачи социального опыта, орудия труда лежат в основесоциальной формы наследования, остающейся за рамками теории биологической эволюции, но связанной с передачей из поколения в поколение информации, важной с точки зрения выживания вида в условиях естественного отбора.
Таким образом, процесс антропосоциогенеза, т. е. становления человека и общества, представляет собой явление двуединое, в котором обе стороны взаимосвязанны и оказывают друг на друга постоянное влияние. Важная роль в этом процессе принадлежит трудовой деятельности, необходимость в которой, тем не менее, возникла в ходе жесткого естественного отбора.
В согласии с теорией самоорганизации появление человека разумного современная антропология рассматривает как скачок. Действительно, «первым человеком», по словам уже не раз упоминавшегося П. Тейяр де Шардена, может быть толькомножестволюдей. Следы первобытных орудий труда, датируемые одним и тем же периодом, встречаются повсеместно в Европе, Азии и Африке, свидетельствуя об одновременности и коллективном характере появления в истории вида Homo sapiens.
Чем дальше в глубь веков пытается человек проследить свою предысторию, тем в большей степени приходится ему ограничиваться предположениями, поскольку ископаемые материалы представлены весьма скудно. До недавнего времени развитие современного человека достаточно определенно прослеживалось как линейный процесс, начиная с питекантропа, жившего в интервале от 1,5 до 0,5 млн. лет назад. Следующее звено –синантроп, появившийся около 400 тыс. лет назад. Наконец, ранней формой Homo sapiens считалсянеандерталец, обитавший 200–35 тыс. лет назад. Что касаетсякроманьонца, костные остатки которого найдены во всех частях света, а самые ранние датируются временем около 40 тыс. лет назад, то его относят к человеку современного вида –неоантропу, или Homo sapiens sapiens.
В последнее время эволюция человека рассматривается уже как ветвящийся процесс. Предполагается, в частности, что самым древним известным предком и человека, и высших обезьян является рамапитек– человекообразная обезьяна, жившая на территории юга Азии и Африки 14–8 млн. лет назад. Также предполагается, что одна из ветвей рамапитеков (дриопитек) дала общего предка гориллы, шимпанзе и человека, обосновавшегося в Африке. В качестве непосредственного предка человека рассматриваетсяавстралопитек, обитавший в Юго-Восточной Африке от 4 до 2 млн. лет назад. Установлено, что австралопитек первым из приматов передвигался на двух конечностях.
Собственно род Homo (человек) начинает отсчет с Homo habilis (человека умелого), произошедшего, вероятно, от одной из четырех разновидностей австралопитека. Этот вид существовал от 2,5 до 1,5 млн. лет назад на юге и юго-востоке Африки. Именно здесь обнаружены древнейшие орудия труда, датируемые временем около 2,5 млн. лет назад, и остатки жилищ.
Примерно 1,6 млн. лет назад от Homo habilis наиболее вероятно произошел более крупный, обладающий мозгом большего объема Homo erectus (человек прямоходящий). Более развитый интеллект, новая техника изготовления орудий, использование огня помогли этому охотнику раннего каменного веказаселить всю Африку, Европу и Юго-Восточную Азию. Яванского питекантропа и пекинского синантропа большинство специалистов относят именно к Homo erectus. Вымер этот вид около 200 тыс. лет назад, а на смену ему пришел уже упоминавшийся Homo sapiens neandertalensis. Любопытно отметить, что объем мозга у неандертальца был больше, чем у современного человека. Вполне возможно, что судьба этого подвида свидетельствует о приходе линейного (количественного) роста объема мозга в критическую точку, успешный выход из которой оказался возможным только за счет качественно нового уровня совершенствования информационных возможностей материального носителя разума.
Надо сказать, что по поводу правомочности включения всех этих предполагаемых предков современного человека в его эволюционное дерево в среде ученых имеется немало версий и сомнений. Однако не следует забывать, что представлять антропогенез в виде линейного, чисто последовательного процесса было бы слишком серьезным упрощением этого сложного, ветвящегося, явления. Следуя теории самоорганизации, необходимо исходить из нелинейности процесса антропогенеза. Попытки доказать, что в палеонтологической летописи должна существовать линейная последовательность ископаемых форм, по всей видимости, не соответствуют реальной картине антропогенеза и эволюции вообще. На самом деле эволюция представляет собой процесс постоянного возникновения многочисленных ответвлений, большинство из которых отвергается естественным отбором. Поэтому ископаемые остатки человекоподобных приматов могут быть связаны между собой только общими предками, отнюдь не являясь последовательными переходными формами к человеку. Так, на основании результатов молекулярно-генетического анализа считается, что живущие в настоящее время человекообразные обезьяны: шимпанзе и горилла – имели общего предка с человеком около 5–7 млн. лет назад, причем за это время в каждой из этих двух ветвей эволюции исчезло, вымерло множество оказавшихся тупиковыми ответвлений. Такой анализ позволяет предположить, что современный человек произошел около 100 тыс. лет назад от архаической формы Homo sapiens, сохранившейся в Африке и повторно расселившейся по планете. Другие же ветви эволюционного дерева оказались тупиковыми.
Таким образом, в теоретической разработке проблемы антропогенеза существует много неясностей и «белых пятен», которые еще предстоит раскрыть. Очевиден, однако, тот факт, что феномен человека – это закономерный результат естественного развития биосферы Земли.
studfiles.net
Происхождение и эволюция человека. Антропогенез
Проблема происхождения человека всегда интересовала людей. У разных народов и племен существуют различные легенды, сказания, предания, объясняющие его происхождение. Религиозные учения также дают различные объяснения происхождению человека. Научное объяснение этой проблемы учёные смогли дать лишь на основе теории эволюции.
Изучением происхождения и эволюции человека занимается наука антропология. Именно она ищет ответы на вопросы, связанные с процессом формирования анатомического типа человека, развития его трудовой деятельности, речи, общественных отношений. Этот процесс называют антропогенезом.
В основе современных научных представлений о происхождении человека лежит концепция, в соответствии с которой человек вышел из мира животных. Широко распространено мнение, что человек произошёл от человекообразных обезьян. Это не совсем верно.
Развитие человека и человекообразных обезьян — это не последовательные ступени, а параллельные ветви эволюции, расхождение между которыми с эволюционной точки зрения очень глубокое.
Находки антропологов свидетельствуют, что приматы появились приблизительно 70-60 млн лет назад в Африке. Их предками были примитивные насекомоядные животные. Поэтому современных ежей и землероек мы можем считать нашими далёкими родственниками.
Первым приматам приходилось соперничать за пищу и места обитания с грызунами. Поэтому приматы перешли к жизни на деревьях. Это и послужило толчком к развитию их характерных черт: пятипалой конечности, острого стереоскопического зрения, хорошо развитой ключицы, большого и сложного головного мозга. Многие миллионы лет наши предки жили на деревьях. Климат в то время был жарким и влажным.
Затем, около 25 млн лет назад, от общей исходной формы приматов разошлись две ветви эволюции, которые в дальнейшем развивались независимо друг от друга. Первая привела к появлению современных высших обезьян. Развитие второй в конечном счёте завершилось появлением человека современного анатомического типа.
Выделяют четыре стадии антропогенеза: предшественники человека, древнейшие люди, древние люди и ископаемые люди современного анатомического типа.
 Основные стадии эволюции человека
Основные стадии эволюции человека
Предшественники человека, или протоантропы, жили 6 — 1 млн лет назад. Впервые их останки были обнаружены в Южной Африке. Во многом они были гораздо ближе к людям, чем любые человекообразные обезьяны. Наиболее существенные морфологические преобразования на данном этапе антропогенеза происходили в скелете и мускулатуре задних конечностей. Эти изменения были связаны с переходом к постоянному передвижению на двух ногах.
Предшественники человека добывали себе пищу собирательством и охотой. Вероятно, они достаточно часто употребляли мясную пищу, охотясь на различных животных с помощью камней и тяжелых костей крупных копытных. Именно протоантропы стали использовать камни как первое метательное оружие. Позднее отдельные группы предшественников человека научились самостоятельно изготавливать простейшие орудия труда и использовать огонь. Это дало им заметное преимущества перед окружающими животными, и их численность стала возрастать. От них произошли предки современного человека — древнейшие люди, или архантропы. Увеличение объёма головного мозга и усложнение его структуры способствовали, по-видимому, появлению у архантропов членораздельной речи. Их поздние представители овладели высокой технологией обработки камней и изготовления из них разнообразных орудий.
Следующая стадия антропогенеза — древние люди, или палеоантропы. Остатки их скелетов впервые были найдены в долине реки Неандерталь в Германии, откуда и пошло другое название — неандертальцы. Они обитали на Земле в то время, когда наблюдалось значительное похолодание — ледниковый период, поэтому целыми группами жили в пещерах, в которых постоянно поддерживали огонь. В целях защиты от холода древние люди стали изготавливать одежду из звериных шкур. Их орудия труда значительно превосходили по совершенству орудия труда архантропов.
 Остатки первобытной культуры человека
Остатки первобытной культуры человека
Люди современного анатомического типа, или неоантропы, появились 60-50 тыс. лет назад и начали стремительно вытеснять древних людей, которым они значительно уступали по физической силе, но обладали и обладают более развитым головным мозгом. Их останки впервые были найдены в гроте Кроманьон во Франции, так появилось другое название — кроманьонцы. В целом они уже не имели существенных отличий от современных людей, принадлежащих к виду человек разумный. Со времени формирования человека современного типа его анатомические параметры почти не изменились. Биологическая эволюция человека сильно затормозилась, и на смену ей пришла социальная эволюция.
Влияние биологических и социальных факторов на эволюцию человека
Современный человек является не только биологическим существом, но и существом социальным — общественным. Благодаря значительным изменениям в строении тела и его функционировании человек смог приспособиться к систематическому труду с использованием изготовленных им самим орудий. Их применение позволило ему более эффективно воздействовать на окружающую среду и тем самым её изменять.
Существование в окружении других людей способствовало формированию у человека членораздельной речи, а затем и письма. Всё это позволило людям общаться и передавать накопленный опыт не только друг другу, но и из поколения в поколение.
Таким образом, для современного человека ведущими и определяющими стали общественно-трудовые отношения. Но это не означает, что возникновение социальной сферы отменило действие биологических факторов, оно лишь изменило их проявление.
Социальные изменения и успехи здравоохранения, связанные с улучшением жизни людей, непосредственно влияют на состояние их здоровья и, следовательно, на уменьшение зависимости человека от негативного влияния условий внешней среды.
Все свои индивидуальные особенности, приобретаемые и формируемые в процессе исторического развития, человек как биологический вид передает по наследству.
Предки человека
Древнейшими общими предками человека и человекообразных обезьян принято считать парапитеков (от греч. пара — возле, около и питекос — обезьяна) — обезьян, которые вели как древесный, так и наземный образ жизни. От них произошла вымершая впоследствии сборная группа обезьян — дриопитеки (от греч. дрис — дерево). Ученые считают, что именно от какой-то группы дриопитеков около 8 млн лет назад начинается эволюционная линия гоминид (людей).
Наиболее ранними представителями семейства гоминид считаются австралопитеки (от лат. австралис — южный). Им было присуще прямохождение. Первыми представителями рода Человек, по мнению большинства специалистов, был прогрессивный вариант австралопитека — человек умелый. В морфологическом плане он незначительно отличается от австралопитеков, но способность к изготовлению примитивных орудий труда (чопперов) позволила ему преодолеть грань, отделявшую ископаемых человекообразных обезьян от древнейших людей (архантропов). Ученые считают, что все архантропы относились к одному виду – человек прямоходящий.
blgy.ru
Антропогенез — Википедия РУ
Антропогене́з[1] — часть биологической эволюции, которая привела к появлению человека разумного (лат. Homo sapiens), отделившегося от прочих гоминид, человекообразных обезьян и плацентарных млекопитающих, процесс историко-эволюционного формирования физического типа человека, первоначального развития его трудовой деятельности, речи. Изучением антропогенеза занимается множество наук, в частности антропология, палеоантропология, генетика, лингвистика.
В эволюционном контексте термин «человек» относится не только к ныне живущим людям, но и к представителям вымерших видов рода Homo. Кроме того, исследования антропогенеза распространяются на других гоминид, например, австралопитеков. Род Homo отделился от австралопитеков или подобных им гоминин около 2 млн лет назад в Африке. Существовало несколько видов людей, большинство из которых вымерло[2]. К ним, в частности, относятся эректусы и неандертальцы.
Важнейшими этапами антропогенеза, отделившими человека от других гоминид и выделившими его из мира животных, были начало изготовления орудий труда, освоение огня и появление языка.
Начиная с H. habilis, люди использовали каменные орудия, всё более искусно изготовленные (см. Палеолит). В последние 50 тыс. лет технология и культура изменяются быстрее, чем в предшествующие эпохи.
История вопроса об антропогенезе
| 1850 | |
| 1900 | |
| 1950 | |
| 2002 | |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. | |
Проблемы антропогенеза стали изучаться в XVIII веке. До этого времени господствовало представление, что человек и природа всегда были и являются такими, как их создал Бог. Однако постепенно в науке, культуре, общественном сознании утверждалась идея развития, эволюции, в том числе и применительно к человеку.
В XVIII и XIX веках
В середине XVIII века К. Линней дал начало научному представлению о происхождении человека. В своей «Системе природы» (1735) он отнёс человека к животному миру, помещая его в своей классификации рядом с человекообразными обезьянами. В XVIII веке зарождается и научная приматология; так, в 1766 году появилась научная работа Ж. Бюффона об орангутане. Голландский анатом П. Кампер показал глубокое сходство в строении основных органов человека и животных.
В XVIII — первой половине XIX века археологи, палеонтологи, этнографы накопили большой эмпирический материал, который лег в основу учения об антропогенезе. Большую роль сыграли исследования французского археолога Буше де Перта. В 1840—1850-х годах он искал каменные орудия и доказывал, что их использовал первобытный человек, живший одновременно с мамонтом и др. Эти открытия опровергали библейскую хронологию, встретили бурное сопротивление. Только в 1860-е годы идеи Буше де Перта признали в науке.
Однако даже Ламарк не решался довести до логического завершения идею эволюции животных и человека и отрицать роль Бога в происхождении человека (в своей «Философии зоологии» он писал об ином происхождении человека, чем только лишь от животных).
Революционную роль в учении об антропогенезе сыграли идеи Чарльза Дарвина. Он писал: «Тот, кто не смотрит, подобно дикарю, на явления природы как на нечто бессвязное, не может больше думать, чтобы человек был плодом отдельного акта творения».
После находки скелета неандертальца (1856) и целого ряда предшествовавших подобных находок в первой половине XIX века в науке появилось новое направление — палеоантропология. Оно дало фактический материал для постановки вопроса не только об анатомическом сходстве человека и некоторых обезьян, но и о биологической эволюции человека в прошедшие эпохи. Этот вопрос был поставлен Ч. Дарвином вскоре после опубликования «Происхождения видов», хотя уже в этой книге он писал: «Будет пролит свет на происхождение человека и его историю». Впрочем, этот аспект дарвинизма был его современникам вполне очевиден.
Эволюция человека была основным предметом дебатов между Томасом Хаксли и Ричардом Оуэном. Хаксли убедительно описал сходство и различия между человеком и обезьянами в своей книге «О положении человека в природе» (1863). Ч. Дарвин к этому времени также опубликовал следующую книгу «Происхождение человека», которая, несмотря на широкое распространение его идей, вновь спровоцировала всплеск бурных дискуссий. Даже сторонники самой идеи эволюции, такие как Альфред Уоллес и Чарлз Лайель не понимали, как могли умственные способности и нравственность появиться у людей в результате естественного отбора.
Со времен Линнея учёные считали, что человекообразные обезьяны — ближайшие родственники людей, поскольку анатомически они очень близки. В XIX веке предполагали, что самым близким к человеку из ныне существующих видов обезьян являются шимпанзе и что у людей и африканских обезьян был когда-то общий предок. Поэтому было бы логичнее всего искать останки этого существа в Африке. Тем не менее, первые находки, в которых заподозрили промежуточное звено между обезьяной и человеком, были сделаны Эженом Дюбуа на острове Ява. Это был питекантроп, то есть обезьяночеловек.
В XX—XXI веке
В 1920-х годах в Африке действительно были обнаружены останки существа, которое Раймонд Дарт назвал австралопитеком. Первой важной находкой был череп детёныша этого вида, обнаруженный в Южной Африке. Мозг ребёнка был слишком велик для детёнышей обезьян, 410 см³, и имел округлую форму, как у современных людей. Клыки были короткими, а основание черепа характерным для прямоходящих существ. Эти признаки убедили Дарта в том, что существо было переходной формой между обезьяной и человеком.
Понадобилось около 20 лет, чтобы подтвердить открытие Дарта новыми находками костей австралопитеков. В то время преобладало мнение о том, что развитие интеллекта предшествовало переходу к прямохождению, а черты австралопитека говорили об обратном. Со второй половины XX века австралопитек считается прямым предком рода Homo, к которому принадлежит и современный человек. Вместе с Homo sapiens австралопитек входит в трибу гоминини. Тем не менее, накапливаются новые данные, которые ставят под сомнение происхождение людей от австралопитека, и этот род может, в конце концов, оказаться боковой и тупиковой ветвью в антропогенезе. Судя по недавним находкам останков сахелантропа и оррорина, австралопитекам предшествовали другие человекообразные обезьяны, которые больше них похожи на сравнительно поздних предков человека. Однако последние находки ещё ждут подтверждений независимыми авторами.
Первоначально австралопитеков разделяли на грацильных и робустных. В 1930-х годах последних отнесли к особому роду парантропы, в 1960-х объединили в один род с австралопитеками, а сейчас прежняя классификация возвращается, хотя некоторые авторы по-прежнему считают, что это обособленная часть видов того же рода.
Сенсационное открытие принадлежит профессору Ли Бергеру (Professor Lee Berger) из Университета Витвотерсренд в Йоханнесбурге (Witwatersrand University in Johannesburg). Сделано оно во время раскопок в пещере Малапа (Malapa Cave) в районе Стеркфонтейн: был найден почти целый скелет австралопитека (скелет ребёнка), жившего около 2 миллионов лет назад (были и другие находки, например, 1994 года — 3,3 миллиона лет назад)[3][4][5].
Эволюция приматов до возникновения человека
Изменения средней температуры Земли за последние 500 млн летПриматы — одна из старейших групп современных плацентарных млекопитающих. Эволюционная история приматов может быть прослежена примерно на 90 млн лет назад, когда приматообразные разделились на приматов и шерстокрылов. Около 87 млн лет назад[6]сухоносые приматы отделились от мокроносых. Около 80 млн лет назад разошлись линии долгопятообразных и обезьянообразных, а лемурообразные отделились от лориобразных. Останки древнейших приматов пока известны лишь из палеоценовых и эоценовых отложений Северной Америки, Евразии и Африки (плезиадаписы, алжирипитек, нотарктус, дарвиниус и др). Только для пургаториуса предполагается верхнемеловое время.
После глобального похолодания, когда около 30 млн лет назад, в раннем олигоцене, Антарктида начала покрываться льдом, приматы вымерли повсеместно кроме Африки, Америки и юга Азии. Одним из выживших был грифопитек — ископаемая обезьяна, жившая на территории современной Германии и Турции около 16,5 млн лет назад, на 1,5 млн лет раньше, чем подобные виды появились в Африке. Возможно, первые человекообразные обезьяны тоже появились не в Африке, а в Евразии. С другой стороны, высказываются предположения, что предки гоминид мигрировали в Евразию из Африки около 17 млн лет назад, когда эти континенты некоторое время были соединены, прежде чем вновь разделились при расширении Средиземного моря. В начале миоцена (23,03 млн лет назад) климат снова стал тёплым, и они могли процветать в Евразии, после чего ареал одного из них, дриопитека, распространился из Европы или западной части Азии в Африку[7].
Возможно, что уцелевшая после климатических изменений тропическая популяция обезьян, хорошо представленная в слоях верхнего эоцена и нижнего олигоцена Файюмского оазиса в Египте (биретия, каранисия, сахарогалаго, протеопитек, египтопитек, катопитек и др.), дала начало всем ныне существующим приматам — лемурам Мадагаскара, лори Юго-Восточной Азии, африканским галаго, широконосым обезьянам Нового Света и узконосым обезьянам Старого Света (человекообразные обезьяны и мартышкообразные).
В раннем миоцене примитивные узконосые обезьяны Восточной Африки пережили длительный период видообразования. Среди выделяемых ныне видов и родов обезьян этой эпохи известны, в частности, камойяпитек, моротопитек, лимнопитек, проконсул, афропитек, кениапитек, чорорапитек, экваториус, отавипитек, ньянзапитек, викториапитек, дендропитек, все из Восточной Африки, а также ореопитек, который обитал в Италии около 9 млн лет назад и европейские — пиеролапитек, анойяпитек, дриопитек, уранопитек, удабнопитек, грекопитек, анкарапитек. При сравнении ДНК современных обезьян было показано, что гиббоны отделились от общего ствола гоминид около 18 млн лет назад, а орангутаны — около 14 млн лет назад. За исключением дендропитека останки ископаемых гиббонов науке неизвестны, и их происхождение остается неясным. Ископаемыми прото-орангутанами считаются сивапитек, обитавший в Азии около 12—10 млн лет назад, и хорапитек из Таиланда.
Предполагается, что видами, близкими к общему предку горилл, шимпанзе и людей, были накалипитек из Кении и грекопитек[8] с Балканского полуострова. По данным молекулярной биологии, около 7—8 млн лет назад сначала гориллы, а потом и шимпанзе отделились от предков людей. ДНК шимпанзе совпадает с человеческой примерно на 99 %[9] (ранее считалось, что на 98,7 %). Из-за влажного климата тропических лесов, в кислых почвах которого кости плохо сохраняются, а также отчасти из-за невнимания исследователей, сосредоточенных преимущественно на поиске предков человека, ископаемых горилл и шимпанзе до сих пор практически не найдено.
В середине миоцена климат снова стал холоднее и засушливее, что спровоцировало новое массовое вымирание фауны. Однако гоминины, как и многие другие виды (антилопы, гиены, собаки, свиньи, слоны, лошади), пережили климатические изменения и сумели к ним адаптироваться. Их дальнейшая эволюция привела к образованию множества новых родов, из которых самыми древними сейчас считаются Sahelanthropus tchadensis (7 млн лет назад) и Orrorin tugenensis (6 млн лет назад). За ними, в частности, последовали:
- Ardipithecus (5,8—4,4 млн лет назад), с видами Ar. kadabba и Ar. ramidus;
- Australopithecus (4—2 млн лет назад), с видами Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali и Au. garhi;
- Paranthropus (3—1,2 млн лет назад), с видами P. aethiopicus, P. boisei и P. robustus;
- Kenyanthropus (3 млн лет назад), вид Kenyanthropus platyops;
- Homo (от 2 млн лет назад), с видами Homo habilis (или Australopithecus habilis), Homo rudolfensis, Homo ergaster, Homo georgicus, Homo antecessor, Homo cepranensis, Homo erectus, Homo heidelbergensis, Homo rhodesiensis, Homo neanderthalensis, Homo sapiens idaltu, Homo sapiens sapiens, Homo floresiensis.
-10 —
–
-9 —
–
-8 —
–
-7 —
–
-6 —
–
-5 —
–
-4 —
–
-3 —
–
-2 —
–
-1 —
–
0 —
Род Homo
Слово homo в переводе с латыни означает «человек» и было выбрано для классификации ещё Линнеем. В латинском языке оно произошло от пра-и.е. *dʰĝʰem- «земля»[10].
В современной таксономии Homo sapiens — единственный ныне существующий вид рода Homo, и хотя продолжающиеся исследования происхождения Homo sapiens дают всё новую и новую информацию о других видах Homo, все эти виды давно уже вымерли. Некоторые из этих видов могли быть предками современных людей, но многие являются лишь «кузенами» и эволюционировали в сторону от нашего вида[11]. В то же время продолжаются и дискуссии о том, какие из них считать отдельными видами, а какие — лишь расами одного вида. В некоторых случаях причиной разногласий является ограниченность или полное отсутствие необходимой информации, в других — различия в подходах к классификации.
Считается, что есть две главные научные школы, по-разному объясняющие движущие силы эволюции человека. Раньше других появились представления об адаптации предков людей, обитавших на деревьях, к жизни в саванне, куда они вышли для охоты на травоядных. Теория саванны, впервые сформулированная Раймондом Дартом, не исключает, что к саванне смогли приспособиться лишь те обезьяны, которые ещё в лесах приобрели необходимую для этого анатомию и поведенческие навыки[12].
В частности, отсутствие волосяного покрова могло иметь несколько объяснений, включая половой отбор. Наличие шерсти — эволюционное преимущество даже в жарком климате: шерсть защищает от солнца и имеется у всех хищников. Но прямохождение уменьшает полученную телом суточную солнечную радиацию примерно на треть; а в полдень она меньше в целых четыре раза. При беге тело лучше охлаждается ветром, когда нет шерсти.[13] Убедительный эксперимент с беговой дорожкой под инфракрасным рефлектором показал: человек в шерстяном свитере может бежать 10-15 минут и перегревается, этот же человек без свитера способен бежать часами.
Некоторые антропологи, например, Бернард Вуд, Кевин Хант и Филипп Тобиас, считают теорию саванны устаревшей. Альтернативная гипотеза допускает, что человек эволюционировал под влиянием приспособления к земноводному существованию, то есть к собиранию моллюсков и прочей пищи на мелководье, что требовало, в частности, способности плавать и нырять, отличающей человека от прочих обезьян. Эта гипотеза объясняет многие анатомические особенности современного человека, такие, как прямохождение[14], отсутствие шерсти[15], развитый слой подкожного жира[16], низкое положение гортани относительно носоглотки, характерное для морских млекопитающих[17], vernix caseosa, или первородная смазка новорождённых детей, также характерная для морских млекопитающих, но не обезьян[17], крупный мозг[18], высокий нос с направленными вниз ноздрями (не вперёд, как у обезьян), предотвращающий попадание воды в носоглотку, и жирная кожа с обилием сальных желёз, которая может служить для защиты от воды[19]. Обсуждается несколько вариантов приспособления протолюдей к жизни в водной стихии, в том числе собирательство на мелководье и развитие новых способов передвижения в воде и доставки собранной пищи на берег[14], плавание[20] и ныряние[16][21][22]. Получить палеоантропологические доказательства земноводного обитания протолюдей крайне сложно, по крайней мере, из-за повышения уровня моря по окончании ледникового периода, из-за чего бывшее мелководье оказалось теперь на глубине 100—120 м[23]. Однако археология и палеонтология позволяют исследовать рацион питания различных видов Homo и его влияние на эволюцию анатомии и поведения[24][25][26][27][28].
Антропогенез до появления людей современного типа
В настоящее время признается, что эволюция гоминид была не линейной, а, скорее, кустообразной. Часто одновременно существовало по три, четыре и может быть даже больше видов гоминид, в том числе на одной и той же территории.
Вся ранняя эволюция гоминид происходила в Африке. 6—7 млн лет назад в Африке жил сахелантроп. Около 6 млн лет назад там же жил оррорин, а примерно 4,2 млн лет назад появились австралопитеки. Отличительной особенностью всех этих существ было передвижение на двух ногах (бипедализм). На сегодняшний день стало ясно, что бипедализм был свойствен гомининам изначально, то есть практически сразу после разделения линий человека и шимпанзе. Эта адаптация не была напрямую связана с жизнью на безлесных пространствах. Существует целый ряд теорий, объясняющих происхождение бипедализма. Таким образом, в период примерно от 6 до 1 млн лет назад в Африке жила довольно большая и разнообразная группа обезьян, передвигавшихся на двух ногах. Однако по размеру мозга эти обезьяны не отличались от современного шимпанзе, и нет оснований предполагать, что они превосходили его по своим интеллектуальным способностям.
Примерно 2,4 млн лет назад в одной из линий гоминид наметилась новая эволюционная тенденция — началось увеличение мозга. Первый представитель гоминин, у которого объём мозга превысил типичные для шимпанзе и австралопитеков 400—450 куб см, — Homo habilis. Он первым стал изготавливать простейшие каменные орудия. По некоторым данным, наиболее примитивная олдувайская культура обработки камня возникла около 3,3[29]—2,7 млн лет назад, а исчезла около 1 млн лет назад. Эти гоминины, по-видимому, начали питаться падалью крупных животных, а свои каменные орудия они, возможно, использовали для разделки туш или соскребания мяса с костей.
У Homo ergaster, который появился около 1,9 млн лет назад, объём мозга, а также размеры тела ещё увеличились. Предполагается, что это связано с увеличением доли мясной пищи в рационе. Возможно, Homo ergaster научился охотиться на крупную и среднюю дичь, или он просто научился более эффективно конкурировать с другими падальщиками.
В Дманиси (Грузия) были найдены кости возрастом около 1,85 млн лет[30]. Грузинские учёные относят их к отдельному виду Homo georgicus, а западные учёные рассматривают их как останки раннего представителя Homo ergaster или Homo erectus, или переходной формой между H. habilis и H. ergaster.
1,76 млн лет назад в Африке появилась более развитая ашельская культура.
Homo erectus заселили обширные территории Евразии. Это была первая волна расселения людей за пределами Африки. Около 1,1—1,2 млн лет их потомки появились и в Западной Европе (Испания). Они описаны как особый вид Homo antecessor. По-видимому, они близки к общему предку неандертальцев и современных людей. В то же время считается, что аббевильская культура в Европе возникла приблизительно 1,5 миллиона лет назад. Около 550—475 тыс. лет назад в Европе существовала клектонская культура.
Первые свидетельства использования огня людьми относятся к периоду примерно 1,5 миллиона лет назад. Приготовление пищи на огне привело к улучшению питания.
Первые люди с чертами прото-неандертальца появляются в Европе 600—350 тысяч лет назад[31]. С неандертальцами связывается мустьерская культура, возникшая 300 тыс. лет назад. В Африке ей соответствовала сангойская культура, возникшая 500 тыс. лет назад.
Денисовские люди стали вторым после неандертальцев видом вымерших гоминин, для которого стал известен полный митохондриальный и почти полный ядерный геномы. Впервые новый вид приматов был выделен исключительно на основании генетических исследований.
Команда учёных из лейпцигского Института эволюционной антропологии общества Макса Планка под руководством шведского биолога Сванте Паабо секвенировала ДНК, извлечённую из фрагмента кости фаланги детского пальца, найденного в 2008 году российскими археологами в Денисовой пещере на Алтае. Выяснилось, что митохондриальная ДНК этого образца отличается от мтДНК современного человека по 385 нуклеотидам, в то время как митохондриальная ДНК неандертальцев отличается от ДНК Homo sapiens на 202 нуклеотида. Статья, посвящённая этому открытию, была опубликована в журнале «Nature» 24 марта 2010 года[32].
Позднее, когда были обработаны последовательности, относящиеся к ядерному геному, оказалось, что денисовский человек всё же ближе к неандертальцу и их эволюционное расхождение произошло около 640 тыс. лет назад. На основе анализа ДНК исследователи полагают, что остатки кости датируются периодом 75—82 тысячи лет назад[33]. Возраст находок, найденных в пещере в тех же самых слоях, был определён при помощи радиоуглеродного анализа в 40 тысяч лет.
Исследовав останки 311 гоминин, живших от 4,4 млн л. н. до последнего ледникового периода, антропологи выяснили, что около 2,2—1,9 млн лет назад представители рода Homo прибавили в росте (около 20 см) и весе (15—20 кг). Между 1,4—1,6 млн лет назад, вскоре после появления Homo erectus, люди ещё подросли на 10 см (исключая виды Homo naledi и Homo floresiensis) при этом вес остался прежним. Около 0,5—0,4 млн лет назад в летописи окаменелостей появляются потяжелевшие на 10—15 кг представители рода Homo, что сигнализирует о адаптации к окружающей среде к северу от Средиземного моря[34].
Появление Homo sapiens
Древнейшие представители вида Homo sapiens появились в результате эволюции 400—250 тыс. лет назад. Господствующей в наши дни гипотезой происхождения людей является африканская, согласно которой наш вид появился в Африке и оттуда распространился по всему свету, замещая существовавшие популяции Homo erectus и неандертальцев. Альтернативная гипотеза называется мультирегиональной. Данные современной генетики поддерживают африканскую теорию.
Древнейшие люди современного типа в культурном отношении ничем не превосходили современных им ранних неандертальцев из Европы. У тех и других были примерно одинаковые среднепалеолитические каменные орудия.
Сравнение полиморфизмов митохондриальной ДНК и датирование окаменелостей позволяют заключить, что Homo sapiens происходит из Африки, где около 200 тыс. лет назад жил последний общий предок ныне живущих людей по женской линии («митохондриальная Ева»)[35].
В 2009 году группа учёных под руководством Сары Тишкофф из Университета Пенсильвании опубликовала в журнале «Сайенс» результаты комплексного исследования генетического разнообразия народов Африки. Они установили, что самой древней ветвью, испытавшей наименьшее количество смешиваний, как раньше и предполагалось, является генетический кластер, к которому принадлежат бушмены и другие народы, говорящие на койсанских языках. Скорее всего, они и являются той ветвью, которая ближе всего к общим предкам всего современного человечества[36].
Можно предположить, что 60 000 — 40 000 лет назад люди мигрировали в Азию и оттуда в Европу (40 000 лет назад), Австралию и Америку (35 000 — 15 000 лет назад)[35] (см. Ранние миграции человека).
Сравнительная таблица видов рода Homo
| Вид | Латинское название | Эпоха (млн лет назад) | Ареал | Средний рост (м) | Масса тела (кг) | Объём головного мозга (см³) | Ископаемые останки | Дата открытия/первой публикации |
|---|---|---|---|---|---|---|---|---|
| Человек умелый | Homo habilis | 2,6—2,5 | Африка | 1,0—1,5 | 30—50 | 650 | множество | 1960/1964 |
| Человек прямоходящий | H. erectus | 2—0,03 | Африка, Евразия (Ява, Китай, Кавказ) | 1,5—1,8 | 60 | 850 (ранние подвиды) — 1100 (поздние подвиды) | множество | 1891/1892 |
| Человек рудольфский | H. rudolfensis | 2,0—1,78 | Кения | 1,5—1,8 | 45—80 | 1 череп | 1972/1986 | |
| Человек грузинский | H. georgicus | 1,8 | Грузия | 1,5—1,7 | 40—50 | 600—680 | несколько | 1999/2002 |
| Человек работающий | H. ergaster | 1,8—1,4 | Южная и Восточная Африка | 1,3—1,7 | 750—1250 | множество | 1975 | |
| Человек-предшественник | H. antecessor | 1,2—0,8 | Испания | 1,6—1,8 | 90 | 1000 | 2 стоянки | 1997 |
| Человек из Чепрано | H. cepranensis | 0,9—0,8? | Италия | 1000 | 1 черепная крышка | 1994/2003 | ||
| Гейдельбергский человек | H. heidelbergensis | 0,8—0,345 | Европа, Африка, Китай | <1,5 | 60 | 1100—1400 | множество | 1908 |
| Неандерталец | H. neanderthalensis | 0,14—0,024 | Европа, Передняя Азия | 1,65 | 55—70 (коренастые) | 1400—1740 | множество | (1829)/1864 |
| Родезийский человек | H. rhodesiensis | 0,3—0,12 | Замбия | 1,8 | 1280 | очень мало | 1921 | |
| Человек разумный разумный | H. sapiens sapiens | 0,2—по н. в. | повсеместно | 1,4—1,9 | 50—100 | 1000—1850 | ныне живущий | —/1758 |
| Человек разумный старейший | H. sapiens idaltu | 0,16—0,15 | Эфиопия | 1450 | 3 черепа | 1997/2003 | ||
| Человек флоресский | H. floresiensis | 0,10—0,012 | Индонезия | 1 | 25 | 400 | 7 особей | 2003/2004 |
Примечание: численная информация взята преимущественно из соответствующих статей.
Эволюция человека в будущем
Считается, что в условиях современного общества (в первую очередь высокого уровня развития медицины) влияние на эволюцию человека таких факторов, как естественный отбор, волн численности и изоляции, значительно снизилось. Неизменным осталось лишь влияние мутационного процесса. Делается вывод, что в обозримом будущем ожидать существенного изменения биологического облика человека не приходится[37][неавторитетный источник?] И единственное направление эволюции, в котором человек будет продолжать эволюционировать, это путь по приобретению резистентности к болезням, которые до сих пор приводят к летальному исходу. По сей день производятся исследования по выяснению путей эволюции человека[38][39].
Исследования показывают, что в современном обществе идет отрицательный отбор по генам, которые способствуют высокому уровню образования и интеллекта, а также хорошим показателям физического здоровья. Этот вывод сделан, в частности, по результатам масштабного исследования 110 тыс. человек. Скорость деградации невелика, но достаточна, чтобы средний IQ снизился на 30 пунктов за тысячу лет, что означало бы «распад цивилизации».[40][41] Другое исследование, охватившее 730 тысяч тестов IQ норвежцев, родившихся в 1962—1991 годы, показало более высокую скорость деградации — 0,3 пункта ежегодно[42].
См. также
Примечания
- ↑ Антропогенез // БРЭ. Т.2. М., 2005.
- ↑ Mark Collard, Bernard Wood; How reliable are human phylogenetic hypotheses? quote: . existing phylogenetic hypotheses about human evolution are unlikely to be reliable http://www.pnas.org/content/97/9/5003.abstract
- ↑ В ЮАР наконец-то найдено то самое недостающее звено
- ↑ Scientists discover ‘missing link between man and apes’
- ↑ Scientists unearth missing link between ‘man and apes’, using ’two million-year-old’ remains of an African child
- ↑ A Molecular Phylogeny of Living Primates, March 17, 2011
- ↑ Kordos L, Begun DR (2001). «Primates from Rudabánya: allocation of specimens to individuals, sex and age categories». J. Hum. Evol. 40 (1): 17–39. DOI:10.1006/jhev.2000.0437. PMID 11139358.
- ↑ Jochen Fuss, Nikolai Spassov, David R. Begun, Madelaine Böhme. Potential hominin affinities of Graecopithecus from the Late Miocene of Europe. PLOS ONE (англ.)русск. (22 мая 2017). Проверено 25 мая 2017.
- ↑ Wong, Kate. Tiny Genetic Differences between Humans and Other Primates Pervade the Genome, Scientific American (September 1, 2014).
- ↑ «dhghem», The American Heritage Dictionary of the English Language (4th ed.), Houghton Mifflin Company, 2000, <http://www.bartelby.org/61/roots/IE104.html>
- ↑ Strait DS, Grine FE, Moniz MA (1997). «A reappraisal of early hominid phylogeny». J. Hum. Evol. 32 (1): 17–82. DOI:10.1006/jhev.1996.0097. PMID 9034954.
- ↑ Richard Leakey, Roger Lewin. Origins Reconsidered. — Little, Brown & Co, 1992. — ISBN 0 349 10345 3.
- ↑ Почему у человека редуцировался волосяной покров на теле и остался на голове и в других местах?
- ↑ 1 2 Niemitz, C (2002). «A Theory on the Evolution of the Habitual Orthograde Human Bipedalism — The «Amphibisce Generalistheorie»». Anthropologischer Anzeiger 60: 3–66.
- ↑ Morgan, Elaine. The Aquatic Ape. — Stein & Day Pub, 1982. — ISBN 0-285-62509-8.
- ↑ 1 2 Hardy, A. (1960). «Was man more aquatic in the past». New Scientist 7: 642–645.
- ↑ 1 2 Morgan, Elaine. The Aquatic Ape Hypothesis. — Souvenir Press, 1997. — ISBN 0-285-63518-2.
- ↑ Crawford, M et al (2000). «Evidence for the unique function of docosahexanoic acid (DHA) during the evolution of the modern hominid brain». Lipids 34: S39–S47.
- ↑ Kingdon, Jonathan. (2003) Lowly Origin Princeton University Press, 242
- ↑ Patrick, John. Human Respiratory Adaptations for Swimming and Diving. — Souvenir Press, 1991. — ISBN 0-285-63033 4.
- ↑ Scars of Evolution— Presented by Sir David Attenborough, a two part BBC radio 4 series examining the ‘aquatic ape hypothesis’
- ↑ Morris, Desmond. The Naked Ape. — April 1999 edition. — 1983. — P. 44. — ISBN 0385334303.
- ↑ Verhaegen M & Munro S (2002). «The continental shelf hypothesis». Nutrition & Health 16: 25–28.
- ↑ Ungar, Peter S. Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. — US : Oxford University Press, 2006. — P. 432. — ISBN 0195183460.
- ↑ Ungar, Peter S. & Teaford, Mark F. Human Diet: Its Origin and Evolution. — Westport, CT : Bergin & Garvey, 2002. — P. 206. — ISBN 0897897366.
- ↑ Bogin, Barry. The evolution of human nutrition // The Anthropology of Medicine: From Culture to Method / Romanucci-Ross, Lola; Moerman, Daniel E.; & Tancredi, Laurence R.. — 3. — South Hadley, Mass. : Bergen and Garvey, 1997. — P. 96–142. — ISBN 0897895169.
- ↑ Barnicot NA (2005, April/June). «Human nutrition: evolutionary perspectives». Integr Physiol Behav Sci 40 (2): 114–17. DOI:10.1007/BF02734246. PMID 17393680.
- ↑ Leonard WR, Snodgrass JJ, Robertson ML (2007). «Effects of brain evolution on human nutrition and metabolism» (PDF). Annu Rev Nutr. 27: 311–27. DOI:10.1146/annurev.nutr.27.061406.093659. PMID 17439362. Проверено 2008-12-29.
- ↑ World’s oldest stone tools discovered in Kenya, 2015
- ↑ Reid Ferring et al.: Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma. In: PNAS. Band 108, Nr. 26, 2011, S. 10432-10436, doi:10.1073/pnas.1106638108
- ↑ J. L. Bischoff et al. (2003). «The Sima de los Huesos Hominids Date to Beyond U/Th Equilibrium (>350 kyr) and Perhaps to 400–500 kyr: New Radiometric Dates». J. Archaeol. Sci. 30 (30). DOI:10.1006/jasc.2002.0834.
- ↑ Потерянное поколение, Лента.ру (26 марта 2010 года)..
- ↑ New DNA analysis shows ancient humans interbred with Denisovans. Nature (31 августа 2012). Архивировано 25 октября 2012 года.
- ↑ Height and weight evolved at different speeds in the bodies of our ancestors, 2017
- ↑ 1 2 Cavalli-Sforza L. L., Feldman M. W. The application of molecular genetic approaches to the study of human evolution. Nature genetics, 2003, v. 33, p. 266—275
- ↑ Африканское кроворазделение
- ↑ Яблоков А. В., Юсупов А. Г. Эволюционное учение: Учебное пособие для биол. спец. вузов. 6-е изд., испр. М.: Высшая школа, 2006. стр. 251
- ↑ Эволюция продолжается? В какую сторону?
- ↑ Финские биологи доказали: человек продолжает эволюционировать наряду с животными
- ↑ 10 главных открытий года о происхождении человека и эволюции («Александр Марков о деградации человека, природе сна и предках домашних кошек»), republic.ru, 15 декабря 2017 года.
- ↑ Selection against variants in the genome associated with educational attainment, Augustine Konga, Michael L. Friggea, Gudmar Thorleifssona, Hreinn Stefanssona, Alexander I. Youngc, Florian Zinka, Gudrun A. Jonsdottira, Aysu Okbayd, Patrick Sulema, Gisli Massona, Daniel F. Gudbjartssona, Agnar Helgasona, Gyda Bjornsdottira, Unnur Thorsteinsdottira, and Kari Stefanssona. Edited by Andrew G. Clark, Cornell University, Ithaca, NY, 2016.
- ↑ Исследование: в конце XX века люди стали глупее. В снижении IQ виноваты гаджеты, качество образования и питания. Данные норвежских ученых показывают, что эффект Флинна перестал действовать, republic.ru, 13 июня 2018 года.
Литература
- Групповой отбор, происхождение человека и происхождение семьи (А. И. Фет. Инстинкт и социальное поведение. Второе издание)
- Алексеев В. П. Становление человечества. — М.: Политиздат, 1984. — 462 с.: ил.
- Борисковский П. И. Древнейшее прошлое человечества. — М.; Л.: Изд-во АН СССР, 1957. — 224 с.: ил. — (Научно-популярная серия АН СССР).
- Гремяцкий М. А. Как произошел человек. — М.: МГУ, 1954. — 176 с.: ил.
- Дробышевский С. В. Антропогенез. — М.: Модерн, 2017. — 168 с.: ил. — ISBN 978-5-94193-074-6.
- Ефименко П. П. Первобытное общество. Очерки по истории палеолитического времени. — 3-е изд. — Киев: Изд-во АН УССР, 1953. — 664 с.: ил.
- История первобытного общества: Общие вопросы. Проблемы антропосоциогенеза / Ин-т этнографии им. Н. Н. Миклухо-Маклая АН СССР. — М.: Наука, 1983. — 432 с.
- Ламберт Давид. Доисторический человек. Кембриджский путеводитель / Пер. с англ. В. З. Махлина. — Л.: Недра, 1991. — 256 с.: ил. — ISBN 5-247-01726-9.
- Ларичев В. Е. Сад Эдема. — М.: Политиздат, 1980. — 400 с.: ил.
- Мерцалов В. Л. Логика антропогенеза. Происхождение человека. — СПб.: Алетейя, 2008. — 296 с.: ил. — (Мир культуры). — ISBN 978-5-91419-072-6.
- Нестурх М. Ф. Происхождение человека / Отв. ред. проф. Я. Я. Рогинский. — М.: Изд-во АН СССР, 1958. — 388 с.: ил.
- Робертс Эллис. Происхождение человека. Эволюция / Пер. с англ. И. В. Павловой, О. В. Сергеевой. — М.: ООО «АСТ», ОГИЗ, 2014. — 256 с.: ил. — ISBN 978-5-17-084157-8.
- Семенов Ю. И. На заре человеческой истории. — М.: Мысль, 1989. — 318 с. — ISBN 5-244-00092-6.
- Уэйд Николас. На заре человечества. Неизвестная история наших предков / Пер. с англ. В. А. Мезина. — М.: Альпина-нон-фикшен, ЛКИ, 2017. — 408 с.: ил. — (Научно-популярная литература). — ISBN 978-5-91671-607-8.
Ссылки
http-wikipediya.ru
Происхождение и эволюция человека. Презентация. Основные этапы, стадии
Профессор кафедры антропологии биологического факультета МГУ, д.б.н.От Редактора: перед Вами — текст доклада, прозвучавшего 24 февраля 2011 года на биологическом факультете МГУ.
Как правило, люди, далекие от антропологии, представляют себе теорию о происхождении человека консервативной и мало подверженной переменам. На самом деле современная эволюционная антропология – наиболее динамичная и изменчивая часть науки о человеке. Она во многом отличается от традиционного антропогенеза, с которым знакомили нас наши преподаватели еще, казалось бы, совсем недавно – в 60-е – 70-е годы прошлого века.
При этом концептуальный скелет эволюционной антропологии остается неизменным. Это представление о длительной биологической эволюции человека от древних приматов через целый ряд ископаемых гоминид к современному виду Homo sapiens. Однако частные концепции, формирующие тело, теоретическую плоть современной науки о происхождении человека, стремительно меняются. Так, на протяжении большей части 20-го века главной объяснительной теорией о причинах и факторах антропогенеза была так называемая трудовая теория Ф. Энгельса, согласно которой развитие прямохождения и головного мозга было напрямую связано с трудовой деятельностью и развитием руки. Ядро теории Ф. Энгельса составляла триада «рука – мозг – прямохождение», в которой обозначалась последовательность развития основных морфо-функциональных систем человеческой эволюции.
Далее, по мере накопления сведений о древнейших гоминидах трудовая теория антропогенеза Ф. Энгельса постепенно лишалась фактологической основы, а знаменитая триада «рука – мозг – прямохождение» утрачивала свою монолитность. Решающую роль в этом процессе сыграли находки австралопитеков и признание их эволюционной значимости. Согласно находкам, в то время как прямохождение уже стало привычным способом передвижения древнейших гоминид, их мозг по объему не превосходил мозг шимпанзе, а способность руки к трудовым операциям вызывала очень большие сомнения. В итоге к концу 20-го века только признак прямохождения остался главным и несомненным критерием гоминизации, то есть принадлежности ископаемых находок к семейству гоминид. Дискуссии в этой области велись лишь по поводу деталей – когда, почему и как возникло прямохождение, но сам способ передвижения сомнению не подвергался.
Но вот появляются новые находки, более подробно изучаются старые, и ситуация вновь меняется, неожиданно возникает новая проблема древней гоминидной локомоции вообще.
Вначале обнаружилось, что практически все австралопитековые, и даже ранние Homo обладали довольно странной особенностью, которую западные исследователи назвали «функциональной дихотомией», хотя более точно было бы обозначить её как «функциональная двойственность» [Johanson, Edey, 1981; Leakey et al., 1995; Alemseged, 2006]. Проявляется эта особенность в несколько разной степени выраженности комплекса прямохождения для верхних и нижних конечностей древнейших гоминид. Нижние конечности по толщине кортикального слоя длинных костей, форме и длине шейки бедра, расположению вертелов бедренной кости и другим особенностям полностью соответствуют прямохождению. В то же время строение верхних конечностей — пропорции, морфология костей плечевого пояса, изогнутость фаланг пальцев – говорит о способности к древесному передвижению. Как писала Е.Н. Хрисанфова, это можно рассматривать как свидетельство «широкого локомоторного репертуара» древнейших гоминид в условиях флуктуации климата и других факторов внешней среды [Хрисанфова, Перевозчиков, 1999].
Возможно и другое объяснение «функциональной дихотомии». Признаки древесного передвижения могли оказаться нейтральными и несколько «задержаться» на верхних конечностях, не мешая их владельцам практиковать полноценную бипедию.
Слайд 1. Ардипитек рамидус
Казалось бы, вопрос решен, объяснение найдено, и основные положения о значимости прямохождения как древней ключевой адаптации не пострадали. Однако новые находки показали, что не только верхние конечности древнейших гоминид были приспособлены к древолазанию, но и стопы некоторых из них имели явно обезьянье строение! Речь, прежде всего, идёт об ардипитеке с датировкой 4,4 млн. лет, находке которого был посвящен весь журнал «Science» в октябре 2009 г.
В этих публикациях ардипитек реконструируется как гоминид, который вел преимущественно древесный образ жизни, медленно перемещаясь по ветвям деревьев с опорой на стопы и ладони. Что касается наземного двуногого передвижения, то оно, по мнению исследователей, носило «эпизодический характер» [White et al., 2009]. Несмотря на приданное ардипитеку выпрямленное положение тела, нельзя не обратить внимание на довольно узкие тазовые кости, которые формируют таз явно понгидного типа. Но самое поразительное – это стопа ардипитека с резко противопоставленным большим пальцем! Согласно графику, приведенному в одной из статей, противопоставление большого пальца стопы у этого гоминида даже больше, чем у шимпанзе. Очень трудно представить себе развитую бипедию у примата с подобной конструкцией стопы! Авторы описания выстраивают гипотетическую эволюционную последовательность от ардипитека к австралопитеку афарскому, но в таком случае приходится допустить, что всего за 500 тыс. лет крайне специализированная стопа ардипитека могла эволюционировать в стопу афарского австралопитека с ее сводчатостью и приведенным первым пальцем. Слайд 1.
От редакции: рекомендуем ознакомиться с мнением нашего научного редактора на эту тему.
Слайд 2. Стопа Homo habilis OH 8
И здесь уместно вспомнить находку OH8, представляющую собой почти полную стопу, которую приписывают представителю самых ранних людей, Homo habilis. Наряду с чисто человеческими особенностями — укороченные пальцы, увеличенный большой палец, расположение пальцев в одной плоскости, полное развитие двойного свода стопы, — отмечаются следы хорошего развития задней большеберцовой мышцы, которая поворачивает стопу наружу, что может рассматриваться как маркер древолазания.
Возможно, в отношении древних гоминид вскоре можно будет говорить не только о широком локомоторном репертуаре, но о довольно широком спектре локомоторных адаптаций, представленных разными находками австралопитековых и даже ранних Homo. И в этом случае вновь возникает проблема критерия гоминизации – если это не рука, не крупный мозг и не устойчивое прямохождение, то что же? Получается, что могли успешно существовать частично древесные и четвероногие гоминиды? Могли ли такие существа быть предками рода Homo?
Очевидно, на все эти вопросы мы пока не можем дать ответ, но сама постановка подобных проблем, которые ещё недавно было просто невозможно себе представить, является знаковым признаком современной эволюционной антропологии.
В начале и середине 20-го века сформировалась и стадиальная теория антропогенеза, постулирующая последовательную смену эволюционных стадий, представленных конкретными находками. Согласно стадиальной теории, каждый эволюционный уровень представлен лишь одним вариантом гоминид, поэтому схемы эволюции неизбежно носят линейный характер, имеют вид ломаной линии, а каждая стадия отображается точкой на этой линии. Кроме того, предполагается, что гоминиды каждого эволюционного уровня являются носителями вполне определенной, доступной на их биологическом уровне культуры, отраженной в археологических орудиях. Согласно этим представлениям, развитие культуры идёт вслед за биологической эволюцией и зависит от неё.
Слайд 3.
Однако на сегодняшний день каждая эволюционная стадия оказывается представленной несколькими ископаемыми формами. Слайд 3. В отношении всех эволюционных этапов установлен факт высокого разнообразия одновременно существующих гоминидных вариантов. При этом в большинстве случаев мы ничего не можем сказать о том, как они соотносятся друг с другом. Исходя из данных о низком физиологическом барьере среди приматов в целом, а также из сложности критерия вида в отношении ископаемых форм, мы вправе предполагать некую возможность биологических смешений между этими формами. В этом случае наиболее приемлемой оказывается модель сетевидной, нелинейной эволюции, которая предполагает возможность генетических обменов между синхронными популяциями гоминид. Принимая концепцию сетевидной эволюции, мы вынуждены отказаться от традиционного представления об эволюционных линиях и в качестве представителей каждой эволюционной стадии рассматривать несколько вариантов гоминид. Идея сетевидной эволюции в современной антропологии наиболее ярко представлена гипотезой мультирегионального происхождения современного человека. Согласно мультирегиональной концепции, для эволюции человечества в планетарном масштабе оказалось вполне достаточно древнейшего исхода архантропов с территории африканского материка, после чего общность базового генофонда человечества поддерживалась благодаря сети локальных генных потоков.
Нелинейность эволюции можно признавать на словах, но принять это представление как основу наших взглядов на эволюцию человека оказывается весьма непросто. Проявлением линейного подхода в современной антропологии является модель «Out of Africa», в которой мы вновь имеем дело с эволюционными линиями. Африканская гипотеза предполагает последовательный исход с территории африканского континента нескольких миграционных волн, древнейшая из которых представлена архантропами, а самая поздняя – ранними сапиенсами.
Срабатывает инерция мышления, и одной из главных проблем для многих современных исследователей оказывается вопрос о том, какой же из одновременно существовавших вариантов гоминид дал начало дальнейшей прогрессивной линии эволюции (всё-таки линии!). С другой стороны, оперировать с понятием «эволюционная линия» намного более удобно, чем с понятием «пласт», и, выстраивая линейные схемы эволюции, многие современные исследователи подразумевают наличие в узловых точках неких обширных популяционных объединений.
Нетрудно видеть, что в модели мультирегиональной эволюции постоянные генные потоки между синхронными популяциями неизбежно формируют сменяющие друг друга пласты форм.
Слайд 4. Синхронные пласты гоминид и «эволюционный конус»
Автором в своё время был предложен термин «гетерогенный эволюционный пласт гоминид» для обозначения совокупности синхронных форм, репродуктивная изоляция которых не может быть ни доказана, ни опровергнута [Бахолдина, 1988]. Прогрессивные признаки могли быть рассеяны в пределах такого синхронного пласта, концентрируясь постепенно под действием отбора, по мере усиления процессов интеграции. Схема последовательно сменяющих друг друга эволюционных пластов может быть представлена в виде своеобразного конуса, внутри которого пунктиром обозначена магистральная линия прогресса, вокруг которой и происходит постепенная концентрация прогрессивных особенностей. За пределами конуса, когда видовой эволюционный полиморфизм гоминид сменяется видовым единообразием на уровне Homo sapiens sapiens, пунктир переходит в сплошную линию. Слайд 4.
Возникает вопрос – что дальше? Представляет ли эволюция современного человека линейный процесс? Скорее всего, нет, но эволюционные прогнозы — занятие неблагодарное. Необходимо добавить, что одна из наиболее плодотворных идей современной эволюционной антропологии, идея магистральной линии прогресса, подробно разработана в трудах А.А. Зубова [1985; 1996; 2004].
При этом нельзя сказать, что стадиальная теория эволюции полностью стала достоянием прошлого, ибо никто не отказывается от представления об эволюционных стадиях и их смене. Просто эти представления видоизменились и усложнились, вобрав в себя огромный массив новых данных.
Слайд 5.
Одной из особенностей нынешней ситуации в эволюционной антропологии является постоянное перемещение некоторых эволюционных пластов и отдельных находок «вверх» или «вниз» вдоль эволюционной вертикали. Относительно недавно, например, произошел дрейф единичной находки нижней челюсти, найденной в начале 20-го века недалеко от города Гейдельберг от уровня древнейших людей, архантропов, к более прогрессивному эволюционному пласту, в котором эта находка объединилась вместе с находками так называемых пренеандертальцев типа Сванскомб, Эрингсдорф и Штейнгейм. Образовался новый таксон, который стал называться, как и нижняя челюсть, Homo heidelbergensis, гейдельбергские люди. Сейчас этот таксон представлен обширным пластом находок из Африки, Азии и Европы. Другие находки попали в этот же таксон из более «высоких» пластов, как, например, знаменитый череп родезийского человека, который в учебниках антропологии 20-го века описывался в качестве африканского неандертальца. Слайд 5.
Представления о неандертальцах претерпели, пожалуй, наиболее радикальные изменения Главное из них состоит в перемещении неандертальцев от одного эволюционного уровня к другому – от уровня пусть спорной, но все же предковой формы, к уровню, занимаемому современным Homo sapiens. Слайд 5.
Сейчас неандертальцы и ископаемые сапиенсы рассматриваются как полиморфное человечество эпохи позднего плейстоцена (позже 200 тыс. лет).
Слайд 6. Находки, свидетельствующие о метисации между неандертальцами и сапиенсами
Антропологи пытаются решить вопрос: насколько это древнее человечество было единым? Иными словами, можно ли с уверенностью судить о наличии или отсутствии биологических смешений между сапиенсами и неандертальцами? Что касается ископаемых свидетельств, то существует довольно много находок, которые можно рассматривать в качестве морфологических подтверждений смешений между сапиенсами и неандертальцами. Это и известные находки с территории Передней Азии – Схул, Табун, Кафзех, Амуд, Кебара, и хорватское местонахождение Крапина, недавние находки с территории Испании и Португалии [Duarte et al., 1999; Schoenherr, 2008]. Слайд 6.
Однако до самого последнего времени генетики активно выступали против возможных смешений сапиенсов и неандертальцев, поскольку результаты изучения древней ДНК показывали очень большой масштаб различий между ними. Правда, постепенно накапливались и единичные генетические данные иного рода. Так, например, американские генетики показали, что самый распространенный вариант гена microcephalin, который определяет рост головного мозга, появился в популяции Homo sapiens 37 000 лет назад в результате скрещивания с каким-то другим архаичным видом. Этот вариант гена настолько сильно отличается от других, более редких его вариантов, которые встречаются у современных людей, что он не мог возникнуть в результате мутаций, а был получен извне. Появились и сведения о том, что связанный с речью ген FOXP2, с мутациями, характерными для сапиенсов, был обнаружен в ДНК неандертальцев.
Но самый сенсационный результат палеогенетических исследований был опубликован в мае прошлого года. Группа генетиков под руководством Сванте Паабо из Института эволюционной антропологии имени Макса Планка в Лейпциге расшифровала, наконец, 2/3 генома неандертальца. В качестве материала послужили кости скелета трех неандертальских женщин из пещеры в Хорватии. Оказалось, что генетические различия между неандертальцами и сапиенсами составляют всего 0,16 %! Гены, по которым наблюдаются различия, кодируют некоторые белки кожи, в том числе её пигментацию, связаны с энергетическим обменом, определяют степень активности мужских половых клеток.
Сравнение с отдельными группами современного человечества показало наибольшую близость неандертальцев к населению Евразии, и, прежде всего, к европейцам. Как пишет Елена Наймарк, излагая итоги этого исследования, «… среди древних вариантов аллелей имеются такие, которые существенно чаще встречаются у европейцев, чем у африканцев. Всего было известно 13 таких географически капризных вариантов. Так вот выяснилось, что 10 из этих 13 вариантов имеются и у неандертальцев!» http://elementy.ru/news/431316
Слайд 7. Происхождение, миграции и зоны смешений неандертальцев и сапиенсов Европы и Передней Азии
Результаты сравнения генома неандертальцев с современными обитателями разных регионов Земли представляются особенно интересными, потому что это тот редкий случай, когда данные генетики совпадают с данными палеоантропологии. Согласно нашим современным представлениям, неандертальцы – это древнейшее, коренное население Европы. Предполагается, что неандертальцы возникли в результате узконаправленной эволюции европейских популяций гейдельбергского человека. Из Европы отдельные группы неандертальцев могли мигрировать на юго-восток и расселиться на территории современного Кавказа и Передней Азии. Возможно, в этом регионе встретились миграционные потоки европейских неандертальцев и древнейших сапиенсов – выходцев из Африки. Слайд 7. Именно с территории Передней Азии и Европы известны находки смешанного типа, имеющие промежуточный между неандертальцами и сапиенсами морфологический облик.
Таким образом, мы теперь точно знаем, что смешения между двумя вариантами древнего человечества, действительно, имели место, и в наших жилах течет и неандертальская кровь! Но тогда неандертальцы и сапиенсы представляли собой не два разных вида, а две большие расы древнего человечества, и контакты между ними следует рассматривать не как межвидовые, а как межрасовые взаимоотношения. И в этом случае современные сапиенсы в некоторой степени оказываются потомками метисов между двумя верхнеплейстоценовыми расами человечества, что является еще одним ударом для сторонников «расовой чистоты», к которой, очевидно, наши предки были полностью равнодушны.
По-прежнему актуальной остается и проблема факторов антропогенеза. Многочисленные исследования в этой сфере обнаруживают высокую поливариантность и множественность причин и движущих сил человеческой эволюции. Большое значение, например, придается стратегиям питания, перемены в которых могли сыграть решающую роль в поворотные моменты биологической истории человечества.
Слайд 8.
Очень интересны исследования, касающиеся значимости полового отбора и фактора восприятия внешности, который сыграл важнейшую роль в исчезновении многих архаических и появлении современных признаков. Обычно роль этого фактора восстанавливается по результатам исследований о предпочтениях того или иного типа внешности современными мужчинами и женщинами. Однако последствия действия этого фактора можно обнаружить и на краниологическом материале. Автором было показано, что признаки лицевого скелета вносят наибольший вклад в общую морфологическую изменчивость и намного более эффективны при разграничении краниологических серий по сравнению с признаками мозгового черепа. Слайд 8. Возможно, эти различия носят эволюционный характер и сформировались в процессе становления вида Homo sapiens. На ранних этапах эволюции происходило ускоренное изменение размеров и формы мозгового черепа, а на поздних этапах началось интенсивное преобразование лицевого скелета под влиянием, в том числе, полового отбора, и антропологическая дифференциация человеческих популяций во многом происходила именно по этой системе признаков.
Меняются и основные тренды развития эволюционной антропологии. В начале и середине 20-го века главным вектором её движения был поиск новых находок, новых фактов и создание основных теоретических концепций, объясняющих эти факты. Сейчас одной из постоянных проблем является приведение в систему всего огромного разнообразия имеющихся данных. И в этом я вижу не просто сложную задачу, но некую ответственную миссию специалистов-антропологов, призванных не только выстраивать такую систему для самих себя, но и приобщать к ней широкий круг людей, заинтересованных в познании сложных процессов развития и становления современного человечества.
Литература:
- Бахолдина В.Ю. Гетерогенный пласт гоминид как объект общего эволюционного процесса // Вопр. Антропологии. 1988. Вып. 81. С. 18 – 28.
- Зубов А.А. Иерархия факторов антропогенеза // Вестник антропологии.1996. Вып. 2. С. 195 – 206.
- Зубов А.А. Магистрилизация и демагистрализация в ходе эволюционного процесса // Вопр. Антропологии. 1985. Вып. 75. С. 14 – 26.
- Зубов А.А. Палеоантропологическая родословная человека. М., 2004. 551 с.
- Хрисанфова Е.Н., Перевозчиков И.В. Антропология. М.: Изд-во Московского университета, 1999. 400 с.
- Alemseged Z., Spoor F., Kimbel W.H., Bobe R., Geraads D., Reed D., Wynn J.G. A juvenile early hominin skeleton from Dikika, Ethiopia // Nature. 2006. 443 (7109). P. 296–301.
- Duarte C, Maur?cio J, Pettitt PB, Souto P, Trinkaus E, van der Plicht H, Zilh?o J. The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia // Proceedings of the National Academy of Sciences 1999. 96 (13). P. 7604–7609.
- Johanson, Donald; Maitland Edey (1981). Lucy: The Beginnings of Humankind. New York: Simon and Schuster, 1981. 416 р.
- Leakey M. G., Feibel C. S., I. MacDougall & A. Walker. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya // Nature. 1995. 376 (6541). P. 565–571.
- Schoenherr N. Late Neandertals and modern human contact in southeastern Iberia // Washington University in St. Louis News & Information. 2008, 10 Dec. P. 1.
- White T., Asfaw B., Beyene Y., Haile-Selassie Y., Lovejoy C.O., Suwa G., WoldeGabriel G. Ardipithecus ramidus and the Paleobiology of Early Hominids // Science. 2009. 326. P. 75 — 86.
antropogenez.ru
Биологическая эволюция человека. Современные теории
Научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. ЛомоносоваЭтот доклад был прочитан в рамках семинара «Эволюция человека сегодня», прошедшего в Государственном Дарвиновском музее 3 марта 2011 г.
За последние десять-пятнадцать лет палеоантропология обогатилась, наверное, больше, чем за предыдущие сто лет. Невозможно даже вкратце упомянуть все находки и сделанные на их основе заключения. Рассмотрение одних только открытий в области палеоприматологии заняло бы несколько часов. Поэтому возможно лишь вкратце осветить наиважнейшие и самые яркие открытия в области антропогенеза.
Находка черепа саадании (Saadanius hijazensis) из среднего олигоцена (28-29 млн.л.н.) запада Саудовской Аравии достойна первого упоминания, поскольку по морфологии и датировке саадания идеально заполняет пробел между проплиопитековыми с одной стороны и мартышковых с гоминоидами – с другой. В настоящее время эта обезьяна – последнее общее звено мартышковых и гоминоидов, отчего была выделена в особое надсемейство сааданиоид (Saadanioidea) с единственным семейством сааданиид (Saadaniidae).
Необычайно расширились знания о проконсулидах – древнейших гоминоидах. В частности, в 1999 году описан целый скелет Nacholapithecus kerioi из среднего миоцена (15 млн.л.н.) Кении. В 2002 году ряд ранее известных нижне- и среднемиоценовых родов выделены в особое семейство дендропитецид (Dendropithecidae) и даже предложено повысить их ранг до надсемейства Dendropithecoidea. Кроме того, в последнее время описан целый ряд человекообразных обезьян неясного таксономического положения, например, Ugandapithecus major (2000) и Kogolepithecus morotoensis (2003) из миоцена Уганды, Samburupithecus kiptalami (1998) из миоцена Кении, Otavipithecus namibiensis (1992) из миоцена Намибии, Langsonia liquidness (1995) с севера Вьетнама с древностью 475 тыс.л.н., а также множество прочих приматов.
В Испанской провинции Каталония найдена и в 2004 году описана значительная часть скелета родственного горилле Pierolapithecus catalaunicus, жившего в миоцене (12,5-13 млн.л.н.). В первоописании он даже отнесён к cемейству Hominidae, хотя отстоит дальше от человека, чем горилла. Близкий род Ouranopithecus, известный ранее из Греции, в 2007 году пополнился турецким верхнемиоценовым видом Ouranopithecus turkae (7,4-8,7 млн.л.н.).
В самое последнее время найдены останки наиболее вероятных предков орангутанов. Таиландские находки включают два вида: среднемиоценового Khoratpithecus chiangmuanensis, описанного в 2003 году (10-13,5 млн.л.н.) и верхнемиоценового Khoratpithecus piriyai, описанного в 2004 году.
Совсем недавно, в 2007 году, был описан, вероятно, прямой предок гориллы – Chororapithecus abyssinicus, обитавший на территории современной Эфиопии, в Афаре, в верхнем миоцене (10-10,5 млн.л.н.). Наконец, в том же 2007 году описаны найденные в 2005 году челюсть и зубы Nakalipithecus nakayamai из Кении (верхний миоцен, 9,8-9,9 млн.л.н.) – последнего общего предка горилл, шимпанзе и гоминид, очередного «недостающего звена» эволюции. Таким образом, разрыв между Накалипитеком и Сахелянтропом, хотя и весьма значителен, но меньше, чем между Накалипитеком и предшествующими ему обезьянами.
Группа ранних австралопитеков вся в целом была открыта в последние пятнадцать лет. В 2002 г. был описан Sahelanthropus tchadensis, в основном по черепу, получившему имя Тумай. Это существо, жившее 6-7 миллионов лет назад на территории современной Республики Чад, имело небольшие клыки и, судя по расположению большого затылочного отверстия, было прямоходящим (или почти прямоходящим?), но по множеству особенностей напоминало современную гориллу. Как следствие, многие учёные склонны отводить Сахелянтропу почётную роль «двоюродного пра-пра-прадедушки», но не считают его одним из наших непосредственных предков.
В 2001 г. было опубликовано научное описание вида Orrorin tugenensis, громко прозванного в популярной печати «Человеком Тысячелетия». Оррорины жили 5,6-6,2 миллиона лет назад на территории современной Кении. Судя по бедренным костям, Оррорины были вполне прямоходящими существами, хотя особенности мышечного релефа плечевой кости и сильный изгиб фаланги пальца руки свидетельствуют о способностях к древолазанию лишь несколько меньших, нежели у шимпанзе.
В 2001 году был описан подвид Ardipithecus ramidus kadabba, сейчас принаваемый за самостоятельный вид Ardipithecus kadabba. Останки его представителей были найдены в местонахождениях Алайла и Гона и имеют древность от 4,3 до 5,5-5,8 миллиона лет назад.
Все вышеназванные находки интересны и замечательны, но меркнут на фоне Ardipithecus ramidus. Описание нового вида состоялось лишь в 1994 году, а рода – в 1995-ом, когда были описаны многочисленные находки из местности Арамис в долине реки Средний Аваш в Эфиопии, имеющие возраст 4,4 миллиона лет назад. История описания скелета Ардипитека – ярчайший пример научной добросовестности. Ведь между его обнаружением – в 1994 г. – и описанием – в конце 2009 г. – прошло 15 лет! См. подробней здесь.
В 1995 году на основе находок в кенийских местонахождениях Канапои и Аллия Бей был описан вид Australopithecus anamensis – более древний и примитивный, чем Australopithecus afarensis. Анамские австралопитеки жили 3,9-4,2 миллиона лета назад. Любопытно, что анамские австралопитеки, будучи преимущественно прямоходящими, могли иногда передвигаться на четвереньках, опираясь на согнутые фаланги пальцев, как это делают гориллы и шимпанзе, о чём свидетельствует форма их лучевой кости. По этой и другим причинам не все учёные видят этот вид в числе наших предков.
За последние годы было найдено множество новых останков афарских и африканских австралопитеков. Из них первейшего упоминания заслуживает скелет детёныша афарского австралопитека из Дикики в Эфиопии. Несмотря на многие очень примитивные черты, ростовые процессы головного мозга были скорее человеческими, нежели обезьяньими. Замечателен скелет Australopithecus afarensis из Ворансо-Милле в Эфиопии: он имеет большие размеры и более человеческие пропорции, чем это доныне предполагалось для австралопитеков. В строении же скелета Australopithecus africanus из Грота Сильберберг – древнейшего для этого вида – имеется слишком много примитивных признаков для зачисления его в наши предки, в том числе сильно отставленный большой палец стопы.
В 1996 г. последовало открытие первого австралопитека из Северной Африки – Australopithecus bahrelghazali. Обломок его челюсти, найденный в Республике Чад и датированный 3,0-3,5 миллиона лет назад, по большинству характеристик очень похож на челюсть Australopithecus afarensis. В 2001 году группа учёных описала новый кенийский род и вид Kenyanthropus platyops, живший 3,2-3,5 миллиона лет назад. Впрочем, его самостоятельность может быть поставлена под сомнение, ведь по большинству черт Кениантроп мало отличается от давно и хорошо известного вида Australopithecus afarensis.
На исходе века, в 1999 году, научный мир узнал о новом виде австралопитеков – Australopithecus garhi. Фрагменты его черепа и конечностей, найденные в Эфиопии, имеют возраст 2,5 миллиона лет назад. Таким образом, этот вид является самым поздним из группы так называемых грацильных австралопитеков. Но самое замечательное, что останки сопровождались примитивными каменными орудиями труда и расколотыми костями антилоп. Как всегда в таких случаях, возникло две версии такого сочетания: либо австралопитеки сами стали добычей более продвинутых гоминид, либо они умели изготавливать орудия и охотились на антилоп. На костях австралопитеков никаких следов орудий не найдено, так что вторая версия выглядит предпочтительной. А это значит, что способность изготавливать каменные орудия не является уникальной особенностью наших предков! Ведь челюсти и зубы Australopithecus garhi слишком специализированы, этот вид не входил в число наших предков; кроме того, к этому времени уже появился Homo habilis – «человек умелый» – наиболее вероятный прямой наш предок. Следовательно, в это время существовали как минимум две независимые, хотя и родственные, группы, находившиеся на пути очеловечивания.
Древнейшие представители нашего рода Homo тоже были обнаружены сравнительно недавно – в 1994 и 2002 годах, в Хадаре в Эфиопии и в Начукуи в Кении. Их датировка – 2,33 и 2,34 млн.л.н.
В 2010 году по двум фрагментарным скелетам из Малапы в Южной Африке с датировкой 1,8-2 млн.л.н. был описан вид Australopithecus sediba. По совокупности признаков он занимает строго промежуточное место между австралопитеками и «ранними Homo«. Особое значение этой находки в том, что Южная Африка доныне считалась окраиной эволюционной арены, а теперь может рассматриваться как её возможный центр.
В 2010 году по ранее известным находкам из Южной Африки был описан новый вид «ранних Homo» – Homo gautengensis. Авторами описания предполагается, что он занимает особое место в эволюции человека, хотя такая позиция уязвима по многим причинам.
Из прочих африканских находок последнего времени особо можно упомянуть черепа из Буйи в Эритрее и Боури в Эфиопии, имеющие возраст 1 миллион лет. Находки этого времени очень немногочисленны, фактически это – самые первые Homo erectus времени первых исходов из Африки.
Сам исход приобрёл зримый образ в лице людей из Дманиси в Грузии. Их многочисленные останки стали известны с 1991 года. В настоящее время найдены уже четыре черепа и почти полный скелет. Их наиболее вероятная датировка – 1,7 млн.л.н. Крайне архаичная морфология, промежуточная между Homo habilis и Homo ergaster, а также некоторые специфические черты позволяют говорить о тупиковости этих пионеров Евразии. Но факт достижения эволюционного уровня, достаточного для жизни за пределами африканской прародины, важен сам по себе.
Останки древнейшего европейца были найдены в 2008 году. Они представляют собой фрагмент нижней челюсти из пещеры Сима дель Элефанте в Испании, в Атапуэрке, с датировкой 1,1-1,2 млн.л.н. К несколько более поздним временам относятся найденные в 1994 году череп из Чепрано в Италии (800-900 тыс.л.н.) и кости нескольких человек из Гран Долина в Атапуэрке в Испании (780 тыс.л.н.), описанные в качестве особого вида Homo antecessor, предположительно предкового для Homo heidelbergensis.
Из множества новых находок в Азии особо можно упомянуть череп из Кокабас в Турции с датировкой 490-510 тыс.л.н., поскольку доныне Турция оставалась «белым пятном» на палеоантропологической карте, хотя неизбежно должна была лежать на пути расселения гоминид. Находка в Салхит в Монголии не имеет датировки, однако морфология черепной крышки позволяет отнести её к виду Homo heidelbergensis, что необычайно удревняет сроки заселения Центральной Азии и даёт новый ракурс представлениям об адаптационных возможностях этого вида.
Серия зубов из пещеры Кезем в Израиле была широко разрекламирована как принадлежащая древнейшим сапиенсам, хотя авторы открытия такого не заявляли. Зубы имеют датировку 200-400 тыс.л.н. и по морфологии могут быть отнесены к предкам людей «группы Схул-Кафзех», доныне плохо известным.
Из китайских находок важнейшей и позднейшей по описанию является фрагмент нижней челюсти из пещеры Жирендун, поскольку при датировке 100-115 тыс.л.н. она имеет зачаточный подбородочный выступ. Китайские антропологи склонны рассматривать человека из Жирендун как древнейшего сапиенса Азии.
Впрочем, древнейшие сапиенсы всё же найдены в Африке. В 2003 году были описаны три черепа из Херто в Эфиопии с датировкой 154-160 тыс.л.н. Смесь архаичных и прогрессивных признаков позволила выделить новый подвид Homo sapiens idaltu. В 2010 году в Темаре в Марокко найден скелет ребёнка 7-8 лет с датировкой 108 тыс.л.н. Его описание пока не опубликовано, но это может быть один из древнейших сапиенсов Северной Африки.
Много нового и на неандертальском фронте. В 2011 году опубликованы результаты исследования зубного камня неандертальцев из Спи и Шанидара, показавшие активное использование в пищу растений неандертальцами. Но самое интересное, что неандертальцы, похоже, умели эти растения варить! Этого от них, кажется, никто не ожидал.
Особого рода сенсация потрясла антропологический мир в 2004 году, когда был описан карликовый вид людей из пещеры Лянг Буа на индонезийском острове Флорес – Homo floresiensis, – живший 95-12 тыс. лет назад и обладавший крайне малым мозгом – около 340 см3, ростом около 1 м, сильно удлинёнными руками, но изготавливавший не самые простые орудия труда и охотившийся на местных карликовых слонов-стегодонов. Правда, ряд учёных склонны видеть в этих «хоббитах» лишь патологию – микроцефалов или людей с синдромом Ларона, но новейшие изыскания скорее подтверждают версию о эндемичном видовом статусе «хоббитов», приобрётших свой неповторимый облик в процессе сотен тысяч лет островной изоляции. Особенно важно, что ряд признаков «хоббитов» – в том числе пропорции конечностей и строение мозга – больше сближают их с австралопитеками, а не с архантропами.
В 2006 году на острове Палау в Микронезии были найдены почти современные скелеты карликовых людей с весьма архаичными чертами, во многом напоминающими «хоббитов» Флореса. Это открытие позволяет надеяться, что в будущем мы узнаем о существовании ещё не одного вида эндемичных островных людей.
Таранная кость из местонахождения Байгара в Тюменской области с датировкой 50 тыс.л.н. похожа на кость Схул IV, что позволяет предположить раннее заселение Сибири сапиенсами или их ближайшими родственниками. В Узбекистане же в пещерах Ангиляк и Оби-Рахмат в то же время жили неандертальцы, судя по найденным тут в 2003 году фрагментам.
Находка в 1998 году скелета ребёнка 3,5-5 лет из Абриго до Лагар Вельго I в Португалии замечательна тем, что при полностью сапиентном строении черепа пропорции конечностей выраженно «неандертальские». Такое сочетание было даже интерпретировано как свидетельство метисного неандертальско-сапиентного происхождения ребёнка, но датировка 24,5 тыс.л.н. делает такую возможность крайне маловероятной. В любом случае, Абриго до Лагар Вельго I выделяется своими «арктическими» пропорциями на фоне прочих «тропических» кроманьонцев.
В 1998 году на суд учёного мира была представлена детально обоснованная «гипотеза вулканической зимы». 73-74 тысячи лет назад на острове Суматра произошло грандиозное извержение вулкана Тоба – одно из сильнейших за всю историю планеты. В воздух было выброшено столько пепла, что в некоторых частях Малакки и Индии его отложения и сейчас достигают 6 м толщины. Кроме того, что выпавший пепел напрямую уничтожал всё живое, в первую очередь растительность, он ещё буквально затмил небо над всей планетой едва ли не на несколько лет. Как следствие, температура Земли упала, и 73 тысячи лет назад начался один из самых сильных пиков оледенения. Вероятно, практически всё население Южной Азии вымерло в результате этого катаклизма. В Европе неандертальцы именно тогда приобрели свой «классический ледниковый» облик и в борьбе с тяжелейшими условиями жизни отстали в своём развитии, а Африка, будучи менее прочих регионов задета катастрофой, стала родиной нового, наиболее прогрессивного вида – Homo sapiens.
В последние семь лет стали известны древнейшие представители нашего вида во всех частях света – из пещеры Тяньянь в Китае (39-42 тыс.л.н.), Пештера ку Оаз в Румынии (34-36 тыс.л.н.), Хофмейр в Южной Африке (36 тыс.л.н.) и Пайзли 5 Майл Пойн Кэйвз в Орегоне (12,3 тыс.л.н.). В Австралии в Манго открыты древнейшие следы австралийцев в буквальном смысле.
Наконец, палеоантропологические исследования на рубеже XXI века вышли на новый уровень с появлением методик выделения и анализа ДНК. Не вдаваясь в подробности, можно суммировать результаты изысканий в этой сложной области в следующих тезисах. Во-первых, неандертальцы ОЧЕНЬ отличаются от сапиенсов. Первоначально эти отличия даже были посчитаны достаточно большими для признания неандертальцев особым видом. Во-вторых, особый вариант ДНК был найден в останках людей из Денисовой пещеры на Алтае, так что «денисовцы» ОЧЕНЬ отличаются как от неандертальцев, так и от сапиенсов, хотя ближе всё же к неандертальцам. В-третьих, у современных НЕафриканцев есть 2,5% неандертальских генов.
Этот новейший вывод замечателен в том отношении, что опровергает более ранние построения генетиков, утверждавших, что ВСЕ современные НЕафриканцы имеют строго монофилетичное происхождение без какой-либо посторонней примеси. Теперь же кристалльная чистота сапиенсов несколько замутнилась неандертальской ДНК, а так называемая «гипотеза африканской Евы» или «концепция замещения» оказалась не столь уж стопроцентной. В-четвёртых, у современных меланезийцев, кроме помянутых неандертальских, обнаружились 4,8% «денисовских» генов, что даёт широкий простор для предположений об извилистых загогулинах древних миграций. Наконец, в-пятых, ДНК верхнепалеолитических людей принципиально не отличается от современной ДНК. Более детальное исследование верхнепалеолитических людей в будущем может дать ценнейшую информацию о путях сложения современных рас.
antropogenez.ru
Антропогенез — это… Что такое Антропогенез?
Антропогенез — часть биологической эволюции, которая привела к появлению человека разумного (лат. Homo sapiens), отделившегося от прочих гоминид, человекообразных обезьян и плацентарных млекопитающих, процесс историко-эволюционного формирования физического типа человека, первоначального развития его трудовой деятельности, речи. Изучением антропогенеза занимается множество наук, в частности антропология, палеоантропология, генетика, лингвистика.
В эволюционном контексте термин «человек» относится не только к ныне живущим людям, но и к представителям вымерших видов рода Homo. Кроме того, исследования антропогенеза распространяются на других гоминид, например, австралопитеков. Род Homo отделился от австралопитеков или подобных им гоминин около 2 млн лет назад в Африке. Существовало несколько видов людей, большинство из которых вымерло[1]. К ним, в частности, относятся эректусы и неандертальцы.
Важнейшими этапами антропогенеза, отделившими человека от других гоминид и выделившими его из мира животных, были начало изготовления орудий труда, освоение огня и появление языка.
Начиная с H. habilis, люди использовали каменные орудия, всё более искусно изготовленные (см. Палеолит). В последние 50 тыс. лет технология и культура изменяются быстрее, чем в предшествующие эпохи.
История вопроса об антропогенезе
| 1850 | |
| 1900 | |
| 1950 | |
| 2002 | |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. | |
Проблемы антропогенеза стали изучаться в XVIII веке. До этого времени господствовало представление, что человек и природа всегда были и являются такими, как их создал Бог. Однако постепенно в науке, культуре, общественном сознании утверждалась идея развития, эволюции, в том числе и применительно к человеку.
В XVIII и XIX веках
В середине XVIII века К. Линней дал начало научному представлению о происхождении человека. В своей «Системе природы» (1735 г.) он отнёс человека к животному миру, помещая его в своей классификации рядом с человекообразными обезьянами. В XVIII веке зарождается и научная приматология; так, в 1766 г. появилась научная работа Ж. Бюффона об орангутане. Голландский анатом П. Кампер показал глубокое сходство в строении основных органов человека и животных.
В XVIII — первой половине XIX века археологи, палеонтологи, этнографы накопили большой эмпирический материал, который лег в основу учения об антропогенезе. Большую роль сыграли исследования французского археолога Буше де Перта. В 40—50-х гг. XIX века он искал каменные орудия и доказывал, что их использовал первобытный человек, живший одновременно с мамонтом и др. Эти открытия опровергали библейскую хронологию, встретили бурное сопротивление. Только в 60-е гг. XIX века идеи Буше де Перта признали в науке.
Однако даже Ламарк не решался довести до логического завершения идею эволюции животных и человека и отрицать роль Бога в происхождении человека (в своей «Философии зоологии» он писал об ином происхождении человека, чем только лишь от животных).
Революционную роль в учении об антропогенезе сыграли идеи Чарльза Дарвина. Он писал: «Тот, кто не смотрит, подобно дикарю, на явления природы как на нечто бессвязное, не может больше думать, чтобы человек был плодом отдельного акта творения».
После находки скелета неандертальца (1856 г.) и целого ряда предшествовавших подобных находок в первой половине XIX в. в науке появилось новое направление — палеоантропология. Оно дало фактический материал для постановки вопроса не только об анатомическом сходстве человека и некоторых обезьян, но и о биологической эволюции человека в прошедшие эпохи. Этот вопрос был поставлен Ч.Дарвином вскоре после опубликования «Происхождения видов», хотя уже в этой книге он писал: «Будет пролит свет на происхождение человека и его историю». Впрочем, этот аспект дарвинизма был его современникам вполне очевиден.
Эволюция человека была основным предметом дебатов между Томасом Хаксли и Ричардом Оуэном. Хаксли убедительно описал сходство и различия между человеком и обезьянами в своей книге «О положении человека в природе» (1863 г.). Ч. Дарвин к этому времени также опубликовал следующую книгу «Происхождение человека», которая, несмотря на широкое распространение его идей, вновь спровоцировала всплеск бурных дискуссий. Даже сторонники самой идеи эволюции, такие как Альфред Уоллес и Чарлз Лайель не понимали, как могли умственные способности и нравственность появиться у людей в результате естественного отбора.
Со времен Линнея учёные считали, что человекообразные обезьяны — ближайшие родственники людей, поскольку анатомически они очень близки. В XIX веке предполагали, что самым близким к человеку из ныне существующих видов обезьян являются шимпанзе и что у людей и африканских обезьян был когда-то общий предок. Поэтому было бы логичнее всего искать останки этого существа в Африке. Тем не менее, первые находки, в которых заподозрили промежуточное звено между обезьяной и человеком, были сделаны Эженом Дюбуа на острове Ява. Это был питекантроп, т.е обезьяночеловек.
В XX—XXI веке
В 1920-х годах в Африке действительно были обнаружены останки существа, которое Раймонд Дарт назвал австралопитеком. Первой важной находкой был череп детёныша этого вида, обнаруженный в Южной Африке. Мозг ребёнка был слишком велик для детёнышей обезьян, 410 см³, и имел округлую форму, как у современных людей. Клыки были короткими, а основание черепа характерным для прямоходящих существ. Эти признаки убедили Дарта в том, что существо было переходной формой между обезьяной и человеком.
Понадобилось около 20 лет, чтобы подтвердить открытие Дарта новыми находками костей австралопитеков. В то время преобладало мнение о том, что развитие интеллекта предшествовало переходу к прямохождению, а черты австралопитека говорили об обратном. Со второй половины XX века австралопитек считается прямым предком рода Homo, к которому принадлежит и современный человек. Вместе с Homo sapiens австралопитек входит в трибу гоминини. Тем не менее, накапливаются новые данные, которые ставят под сомнение происхождение людей от австралопитека, и этот род может, в конце концов, оказаться боковой и тупиковой ветвью в антропогенезе. Судя по недавним находкам останков сахелантропа и оррорина, австралопитекам предшествовали другие человекообразные обезьяны, которые больше них похожи на сравнительно поздних предков человека. Однако последние находки ещё ждут подтверждений независимыми авторами.
Первоначально австралопитеков разделяли на грацильных и робустных. В 1930-х годах последних отнесли к особому роду парантропы, в 1960-х объединили в один род с австралопитеками, а сейчас прежняя классификация возвращается, хотя некоторые авторы по-прежнему считают, что это обособленная часть видов того же рода.
Сенсационное открытие принадлежит профессору Ли Бергеру (Professor Lee Berger) из Университета Витвотерсренд в Йоханнесбурге (Witwatersrand University in Johannesburg). Сделано оно во время раскопок в пещере Малапа (Malapa Cave) в районе Стеркфонтейн был найден почти целый скелет австралопитека (скелет ребёнка), жившего около 2 миллионов лет назад (были и другие находки, например, 1994 года — 3,3 миллиона лет назад)[2][3][4].
В СССР и постсоветской России
Человек — одновременно и биологическое существо, и социальное, поэтому антропогенез неотрывно связан с социогенезом, представляя собой по сути единый процесс антропосоциогенеза.
В соответствии с современными научными представлениями, современный человек является результатом развития материи на протяжении 13,7 миллиардов лет и представляет собой наиболее сложно организованную её разновидность из всего наблюдаемого — независимо от того, говорим мы о человеческом мозге, сознании или какой-либо форме социальной организации. Считается, что в конечном итоге инфляционный вакуум (состояние материи, предшествующее большому взрыву, где частицы обладали «виртуальным» существованием) проделал путь до современного человека (универсальный эволюционизм).
Вопросы антропогенеза, как вопросы возникновения существенных свойств человека, возникших относительно мировой эволюции сравнительно недавно, фокусируются на появлении вида Homo sapiens — современного человека, в связи с чем тесно связываются с вопросами биологической эволюции. Начиная с Дарвина она объясняется приспособлением живого к условиям изменяющейся среды при воздействии трёх определяющих факторов — изменчивости (мутации), отбора (оценка средой эффективности этих мутаций) и наследственности (передача этих мутаций). Таким образом полагается, что в соответствии с этим эволюционным законодательством, под воздействием изменений окружающей среды обезьяноподобные предки современного человека приобрели характерные для него свойства. Среди свойств человека, представляющих в нём специфически человеческое, прежде всего выделяются когнитивные способности (сознание) и язык.
Формирование составляющих сознание когнитивных особенностей современного человека — целеполагания, способности к абстрагированию, воображения, памяти — было определено изменениями организма предков людей. Так, это увеличение объёма головного мозга — от 500 кубических сантиметров у современных крупных обезьян до 1450 у современного человека, противопоставление большого пальца руки (сделало возможным манипуляцию предметами), формирование современного вида стопы (сделало возможным прямохождение) — «гоминидная триада».
В свою очередь эти изменения были вызваны изменением внешних, прежде всего — климатических, условий. Так, определяющую роль сыграли периоды значительных похолоданий («ледниковый период»), вынудив предков современного человека переместиться из гибнущих лесов на равнину. Здесь у них развивалось прямохождение, потребность в добывании пищи и защите от хищников провоцировала использование предметов окружающего мира.
Вопрос ключевого преобразования, непосредственно сделавшего из животного человека, рассматривается различными исследователями по разному — в зависимости и от определения ими этой границы. Так, в отечественной науке существует представление, что ключевым фактором в этом процессе стала деятельность по производству орудий труда как основание для выделения человеком объективных свойств предметов:
«Сознание возникает как отражение прежде всего тех объективных свойств природных предметов, которые выявляются в процессе производства орудий труда. В этом процессе необходимо взаимодействие между собой двух (по крайней мере) природных предметов (камней, палок, костей). Результат их взаимодействия (то есть орудие труда) определяется объективными свойствами таких природных предметов. Ставя во взаимодействие между собой два природных предмета, человек получает возможность выделять их объективные свойства (удар одним камнем по другому дает совсем иной результат, чем удар камнем по дереву: так проявляется объективное свойство твёрдости). Взаимодействие двух материальных предметов между собой позволяет выделять их объективные свойства, то есть свойства, не зависящие от того, кто ставит во взаимодействие эти предметы. … Без учета объективных свойств материальных предметов систематическое производство орудий труда просто невозможно. Другими словами, производя орудия труда, субъект получает возможность выявлять не только преимущественно ситуативные, относительные связи между организмом и средой (что свойственно психике животных), но и объективные связи между предметами, вещами самой природной среды.» (Найдыш В. М.)
Советско-российский философ Юрий Бородай связывает развитие предков людей с развитием их воображения. Согласно его оригинальной концепции, претендование и проявление желания относительно занятой самки в среде уже вооружённых примитивным оружием предков человека несло опасность для жизни. Поэтому удовлетворять эти потребности приходилось в воображении, активно тем самым его развивая.
Другую, ещё более эпатирующую концепцию антропогенеза выдвинул историк и социолог Борис Поршнев в книге «О начале человеческой истории». Поршнев утверждал, что речь, мышление и сознание появились только у вида Homo sapiens, сформировавшегося, с точки зрения автора, лишь 35-40 тысяч лет назад; предки человека, неспособные конкурировать с животными-хищниками, питались падалью, и прямохождение, навыки использования орудий и даже огня были выработаны ещё в животном состоянии.
Эволюция приматов до человека
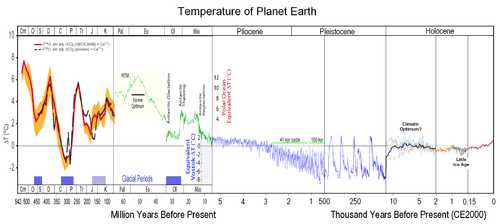 Изменения средней температуры Земли за последние 500 млн лет
Изменения средней температуры Земли за последние 500 млн летПриматы — одна из старейших групп современных плацентарных млекопитающих. Эволюционная история приматов может быть прослежена примерно на 90 млн лет назад, когда приматообразные разделились на приматов и шерстокрылов. 85 млн лет назад сухоносые приматы отделились от мокроносых. Около 80 млн л.н. разошлись линии долгопятообразных и обезьянообразных, а лемурообразные отделились от лориобразных. Останки древнейших приматов известны лишь из палеоценовых и эоценовых отложений Северной Америки, Евразии и Африки (плезиадаписы, антрасимия, нотарктус, дарвиниус (Ида) и др). Только для пургаториуса предполагается верхнемеловое время.
После глобального похолодания, когда около 30 млн лет назад, в раннем олигоцене, Антарктида начала покрываться льдом, приматы вымерли повсеместно кроме Африки, Америки и юга Азии. Одним из выживших был грифопитек — ископаемая обезьяна, жившая на территории современной Германии и Турции около 16,5 млн лет назад, на 1,5 млн лет раньше, чем подобные виды появились в Африке. Возможно, первые человекообразные обезьяны также появились не в Африке, а в Евразии. С другой стороны, высказываются предположения, что предки гоминид мигрировали в Евразию из Африки около 17 млн лет назад, когда эти континенты некоторое время были соединены, прежде чем вновь разделились при расширении Средиземного моря. В начале миоцена (23.03 мл лет назад) климат снова стал тёплым, и они могли процветать в Евразии, после чего ареал одного из них, дриопитека, распространился из Европы или западной части Азии в Африку[5].
Возможно, что уцелевшая после климатических изменений тропическая популяция обезьян, хорошо представленная в слоях верхнего эоцена и нижнего олигоцена Файюмского оазиса в Египте (биретия, каранисия, сахарогалаго, протеопитек, египтопитек, катопитек и др.), дала начало всем ныне существующим приматам — лемурам Мадагаскара, лори Юго-Восточной Азии, африканским галаго, широконосым обезьянам Нового Света и узконосым обезьянам Старого Света (человекообразные обезьяны и мартышкообразные).
В раннем миоцене примитивные узконосые обезьяны Восточной Африки пережили длительный период видообразования. Среди выделяемых ныне видов и родов обезьян этой эпохи известны, в частности, камойяпитек, моротопитек, лимнопитек, проконсул, афропитек, кениапитек, чорорапитек, экваториус, отавипитек, ньянцапитек, викториапитек, дендропитек, все из Восточной Африки, а также ореопитек, который обитал в Италии около 9 млн лет назад и европейские — пиеролапитек, анойяпитек, дриопитек. При сравнении ДНК современных обезьян было показано, что гиббоны отделились от общего ствола гоминид ~ 18 млн лет назад, а орангутаны — около 14 млн лет назад. За исключением дендропитека останки ископаемых гиббонов науке неизвестны, и их происхождение остается неясным. Ископаемым прото-орангутаном считается сивапитек, обитавший в Азии около 12—10 млн лет назад и хорапитек из Таиланда.
Предполагается, что видами, близкими к общему предку горилл, шимпанзе и людей, были накалипитек из Кении и уранопитек с Балканского полуострова. По данным молекулярной биологии, около 7 — 8 млн лет назад сначала гориллы, а потом и шимпанзе отделились от предков людей. ДНК шимпанзе на 98,4 % идентична человеческой. Из-за влажного климата тропических лесов, в кислых почвах которого кости плохо сохраняются, а также отчасти из-за невнимания исследователей, сосредоточенных преимущественно на поиске предков человека, ископаемых горилл и шимпанзе до сих пор практически не найдено.
В середине миоцена климат снова стал холоднее и засушливее, что спровоцировало новое массовое вымирание фауны. Однако гоминины, как и многие другие виды (антилопы, гиены, собаки, свиньи, слоны, лошади) пережили климатические изменения и сумели к ним адаптироваться. Их дальнейшая эволюция привела к образованию множества новых видов, из которых самыми древними сейчас считаются Sahelanthropus tchadensis (7 млн лет назад) и Orrorin tugenensis (6 млн лет назад). За ними, в частности, последовали:
- Ardipithecus (5.8-4.4 млн лет назад), с видами Ar. kadabba и Ar. ramidus;
- Australopithecus (4-2 млн лет назад), с видами Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali и Au. garhi;
- Paranthropus (3-1.2 млн лет назад), с видами P. aethiopicus, P. boisei и P. robustus;
- Kenyanthropus (3 млн лет назад), вид Kenyanthropus platyops
- Homo (от 2 млн лет назад), с видами Homo habilis (или Australopithecus habilis), Homo rudolfensis, Homo ergaster, Homo georgicus, Homo antecessor, Homo cepranensis, Homo erectus, Homo heidelbergensis, Homo rhodesiensis, Homo neanderthalensis, Homo sapiens idaltu, Homo sapiens sapiens, Homo floresiensis.
Род Homo
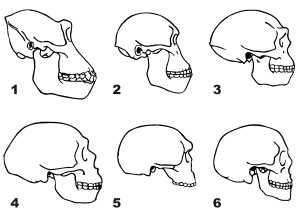
Слово homo в переводе с латыни означает «человек» и было выбрано для классификации ещё Линнеем. В латинском языке оно произошло от общеиндоевропейского корня «dhghem», земля[6].
В современной таксономии Homo sapiens — единственный ныне существующий вид рода Homo, хотя продолжающиеся исследования происхождения Homo sapiens дают всё новую и новую информацию о других видах Homo, давно уже вымерших. Некоторые из этих видов могли быть предками современных людей, но многие являются лишь «кузенами» и эволюционировали в сторону от нашего вида[7]. В то же время продолжаются и дискуссии о том, какие из них считать отдельными видами, а какие — лишь расами одного вида. В некоторых случаях причиной разногласий является ограниченность или полное отсутствие необходимой информации, в других — различия в подходах к классификации.
Считается, что есть две главные научные школы, по-разному объясняющие движущие силы эволюции человека. Раньше других появились представления об адаптации предков людей, обитавших на деревьях, к жизни в саванне, куда они вышли для охоты на травоядных. Теория саванны, впервые сформулированная Раймондом Дартом, не исключает, что к саванне смогли приспособиться лишь те обезьяны, которые ещё в лесах приобрели необходимую для этого анатомию и поведенческие навыки[8].
Некоторые антропологи, например, Бернард Вуд, Кевин Хант и Филипп Тобиас, считают теорию саванны устаревшей. Альтернативная гипотеза допускает, что человек эволюционировал под влиянием приспособления к земноводному существованию, то есть к собиранию моллюсков и прочей пищи на мелководье, что требовало, в частности, способности плавать и нырять, отличающей человека от прочих обезьян. Эта гипотеза объясняет многие анатомические особенности современного человека, такие как прямохождение[9], отсутствие шерсти[10], развитый слой подкожного жира[11], низкое положение гортани относительно носоглотки, характерное для морских млекопитающих,[12], vernix caseosa или первородная смазка новорождённых детей, также характерная для морских млекопитающих, а не обезьян[12], крупный мозг[13], высокий нос с направленными вниз ноздрями (не вперёд, как у обезьян), предотвращающий попадание воды в носоглотку и жирная кожа с обилием сальных желёз, которая может служить для защиты от воды[14]. Обсуждается несколько вариантов приспособления протолюдей к жизни в водной стихии, в том числе собирательство на мелководье и развитие новых способов передвижения в воде и доставки собранной пищи на берег[9], плавание[15] и ныряние[11][16][17]. Получить палеоантропологические доказательства земноводного обитания протолюдей крайне сложно, по крайней мере, из-за повышения уровня моря по окончании ледникового периода, из-за чего бывшее мелководье оказалось теперь на глубине 100—120 м[18]. Однако археология и палеонтология позволяют исследовать рацион питания различных видов Homo и его влияние на эволюцию анатомии и поведения[19][20][21][22][23].
Антропогенез до появления людей современного типа
В настоящее время признается, что эволюция гоминин была не линейной, а, скорее, кустообразной. Часто одновременно существовало по три, четыре и может быть даже больше вида гоминин, в том числе на одной и той же территории.
Вся ранняя эволюция гоминин происходила в Африке. 6-7 млн лет назад в Африке жил сахелантроп. Около 6 млн лет назад там же жил оррорин, а примерно 4,2 млн лет появились австралопитеки. Отличительной особенностью всех этих существ было передвижение на двух ногах (бипедализм). На сегодняшний день стало ясно, что бипедализм был свойствен гомининам изначально, то есть практически сразу после разделения линий человека и шимпанзе. Эта адаптация не была напрямую связана с жизнью на безлесных пространствах. Существует целый ряд теорий, объясняющих происхождение бипедализма. Таким образом, в период примерно от 6 до 1 млн назад в Африке жила довольно большая и разнообразная группа обезьян, передвигавшихся на двух ногах. Однако по размеру мозга эти обезьяны не отличались от современного шимпанзе и нет оснований предполагать, что они превосходили его по своим интеллектуальным способностям.
Примерно 2,4 миллиона лет назад в одной из линий гоминин наметилась новая эволюционная тенденция — началось увеличение мозга. Первый представитель гоминин, у которого объём мозга превысил типичные для шимпанзе и австралопитеков 400—450 куб см. — это Homo habilis. Он первым стал изготавливать простейшие каменные орудия. По некоторым данным, наиболее примитивная олдувайская культура обработки камня возникла около 2,7 миллионов лет назад, а исчезла около 1 миллиона лет назад. Эти гоминины, по-видимому, начали питаться падалью крупных животных, а свои каменные орудия они, возможно, использовали для разделки туш или соскребания мяса с костей.
У Homo ergaster, которые появились около 1,9 млн лет назад, объём мозга, а также размеры тела ещё увеличились. Предполагается, что это связано с увеличением доли мясной пищи в рационе. Возможно, Homo ergaster научился охотиться на крупную и среднюю дичь, или он просто научился более эффективно конкурировать с другими падальщиками.
1,76 млн лет назад в Африке появилась более развитая ашёльская культура. Это первая человеческая культура, покинувшая пределы Африки. В Грузии были найдены кости возрастом около 1,75 млн лет. Грузинские учёные относят их к отдельному виду Homo georgicus, а западные учёные рассматривают их как останки раннего представителя Homo ergaster или Homo erectus.
Homo erectus заселили обширные территории Евразии. Это была первая волна расселения людей за пределами Африки. Около 1,1-1,2 млн лет их потомки появились и в Западной Европе (Испания). Они описаны как особый вид Homo antecessor. По-видимому, они близки к общему предку неандертальцев и современных людей. В то же время считается, что аббевильская культура в Европе возникла приблизительно 1,5 миллиона лет назад. Около 550—475 тыс. лет назад в Европе существовала клектонская культура.
Первые свидетельства использования огня людьми относятся к периоду примерно 1,5 миллиона лет. Приготовление пищи на огне привело к улучшению питания.
140 тыс. лет назад в Европе появились неандертальцы. С неандертальцами связывается мустьерская культура, возникшая, однако, 300 тыс. лет назад. В Африке ей соответствовала сангойская культура, возникшая 500 тыс. лет назад.
Появление Homo sapiens
Древнейшие представители вида Homo sapiens появились в результате эволюции 400—250 тыс. лет назад. Господствующей в наши дни гипотезой происхождения людей является африканская, согласно которой наш вид появился в Африке и оттуда распространился по всему свету, замещая существовавшие популяции Homo erectus и неандертальцев. Альтернативная гипотеза называется мультирегиональной. Согласно последней, люди, начиная, по-видимому, от Homo erectus, эволюционировали как единый вид, в котором генные потоки могли свободно циркулировать. Имеющиеся в настоящее время данные палеоантропологии не позволяют сделать окончательный выбор между этими гипотезами[24], хотя данные генетики поддерживают африканскую теорию.
Древнейшие люди современного типа в культурном отношении ничем не превосходили современных им ранних неандертальцев из Европы. У тех и других были примерно одинаковые среднепалеолитические каменные орудия.
Сравнение полиморфизмов митохондриальной ДНК и датирование окаменелостей позволяют заключить, что Homo sapiens происходит из Африки, где около 200 тыс. лет назад жил последний общий предок (англ.)русск. ныне живущих людей по женской линии («митохондриальная Ева»)[25].
В 2009 году группа учёных под руководством Сары Тишкофф из Университета Пенсильвании опубликовала в журнале Science результаты комплексного исследования генетического разнообразия народов Африки. Они установили, что самой древней ветвью, испытавшей наименьшее количество смешиваний, как раньше и предполагалось, является генетический кластер, к которому принадлежат бушмены и другие народы, говорящие на койсанских языках. Скорее всего, они и являются той ветвью, которая ближе всего к общим предкам всего современного человечества[26].
Можно предположить, что 60 000 — 40 000 лет назад люди мигрировали в Азию, и оттуда в Европу (40 000 лет назад), Австралию и Америку (35 000 — 15 000 лет назад)[25] (см. Ранние миграции человека).
Эволюция человека в будущем
Гипотетический облик «человека будущего». Сконструирован Быстровым А. П., исходя из предположения о незавершенности морфологической эволюции человека[27]В условиях современного общества влияние на эволюцию человека таких факторов, как естественный отбор, волны численности и изоляция, значительно снизилось. Неизменным осталось лишь влияние мутационного процесса. Таким образом в обозримом будущем ожидать существенного изменения биологического облика человека не приходится[28]. Впрочем, исходя из предположения о незавершенной морфологической эволюции человека советский антрополог Быстров А. П. предположил в 1957 году гипотетический облик «человека будущего» (см. рисунок).
Сравнительная таблица видов рода Homo
| Виды | Эпоха (млн лет назад) | Ареал | Средний рост (м) | Масса тела (кг) | Объём головного мозга (см³) | Ископаемые останки | Дата открытия/первой публикации |
|---|---|---|---|---|---|---|---|
| H. habilis | 2,6 — 2,0 | Африка | 1,0 — 1,5 | 30 — 50 | 660 | множество | 1960/1964 |
| H. erectus | 1,89 — 0,143 | Африка, Евразия (Ява, Китай, Кавказ) | 1,45 — 1,85 | 40 — 68 | 900 — 1200 | множество | 1891/1892 |
| H. rudolfensis | 1,9 | Кения | 1 череп | 1972/1986 | |||
| H. georgicus | 1,8 | Грузия | 600 | несколько | 1999/2002 | ||
| H. ergaster | 1,9 — 1,4 | Южная и Восточная Африка | 1,9 | 700—850 | множество | 1975 | |
| H. antecessor | 1,2 — 0,8 | Испания | 1,75 | 90 | 1000 | 2 стоянки | 1997 |
| H. cepranensis | 0,9 — 0,8? | Италия | 1000 | 1 черепная крышка | 1994/2003 | ||
| H. heidelbergensis | 0,6 — 0,25 | Европа, Африка, Китай | 1,8 | 60 | 1100—1400 | множество | 1908 |
| H. neanderthalensis | 0,35 — 0,03 | Европа, Западная Азия | 1,6 | 55 — 70 (коренастые) | 1200—1700 | множество | (1829)/1864 |
| H. rhodesiensis | 0,3 — 0,12 | Замбия | 1300 | очень мало | 1921 | ||
| H. sapiens sapiens | 0,2 — до наст. вр. | повсеместно | 1,4 — 1,9 | 50 — 100 | 1000—1850 | ныне живущий | —/1758 |
| H. sapiens idaltu | 0,16 — 0,15 | Эфиопия | 1450 | 3 черепа | 1997/2003 | ||
| H. floresiensis | 0,10 — 0,012 | Индонезия | 1 | 25 | 400 | 7 особей | 2003/2004 |
В формате JPG. BY Sentteriys & Scriny
См. также
Примечания
- ↑ Mark Collard, Bernard Wood; How reliable are human phylogenetic hypotheses? quote: .. existing phylogenetic hypotheses about human evolution are unlikely to be reliable http://www.pnas.org/content/97/9/5003.abstract
- ↑ В ЮАР наконец-то найдено то самое недостающее звено
- ↑ Scientists discover ‘missing link between man and apes’
- ↑ Scientists unearth missing link between ‘man and apes’, using ’two million-year-old’ remains of an African child
- ↑ Kordos L, Begun DR (2001). «Primates from Rudabánya: allocation of specimens to individuals, sex and age categories». J. Hum. Evol. 40 (1): 17–39. DOI:10.1006/jhev.2000.0437. PMID 11139358.
- ↑ «dhghem», «The American Heritage Dictionary of the English Language» (4th ed.), Houghton Mifflin Company, 2000, <http://www.bartelby.org/61/roots/IE104.html>
- ↑ Strait DS, Grine FE, Moniz MA (1997). «A reappraisal of early hominid phylogeny». J. Hum. Evol. 32 (1): 17–82. DOI:10.1006/jhev.1996.0097. PMID 9034954.
- ↑ Richard Leakey Roger Lewin Origins Reconsidered. — Little, Brown & Co, 1992. — ISBN 0 349 10345 3
- ↑ 1 2 Niemitz, C (2002). «A Theory on the Evolution of the Habitual Orthograde Human Bipedalism — The «Amphibisce Generalistheorie»». Anthropologischer Anzeiger 60: 3–66.
- ↑ Morgan Elaine The Aquatic Ape. — Stein & Day Pub, 1982. — ISBN 0-285-62509-8
- ↑ 1 2 Hardy, A. (1960). «Was man more aquatic in the past». New Scientist 7: 642–645.
- ↑ 1 2 Morgan Elaine The Aquatic Ape Hypothesis. — Souvenir Press, 1997. — ISBN 0-285-63518-2
- ↑ Crawford, M et al (2000). «Evidence for the unique function of docosahexanoic acid (DHA) during the evolution of the modern hominid brain». Lipids 34: S39–S47.
- ↑ Kingdon, Jonathan. (2003) Lowly Origin Princeton University Press, 242
- ↑ Patrick John Human Respiratory Adaptations for Swimming and Diving. — Souvenir Press, 1991. — ISBN 0-285-63033 4
- ↑ Scars of Evolution— Presented by Sir David Attenborough, a two part BBC radio 4 series examining the ‘aquatic ape hypothesis’
- ↑ Morris Desmond The Naked Ape. — April 1999 edition. — 1983. — P. 44. — ISBN 0385334303
- ↑ Verhaegen M & Munro S (2002). «The continental shelf hypothesis». Nutrition & Health 16: 25–28.
- ↑ Ungar, Peter S. Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. — US: Oxford University Press, 2006. — P. 432. — ISBN 0195183460
- ↑ Ungar, Peter S. & Teaford, Mark F. Human Diet: Its Origin and Evolution. — Westport, CT: Bergin & Garvey, 2002. — P. 206. — ISBN 0897897366
- ↑ Bogin Barry The evolution of human nutrition // The Anthropology of Medicine: From Culture to Method / Romanucci-Ross, Lola; Moerman, Daniel E.; & Tancredi, Laurence R.. — 3. — South Hadley, Mass.: Bergen and Garvey, 1997. — P. 96–142. — ISBN 0897895169
- ↑ Barnicot NA (2005, April/June). «Human nutrition: evolutionary perspectives». Integr Physiol Behav Sci 40 (2): 114–17. DOI:10.1007/BF02734246. PMID 17393680.
- ↑ Leonard WR, Snodgrass JJ, Robertson ML (2007). «Effects of brain evolution on human nutrition and metabolism» (PDF). Annu Rev Nutr. 27: 311–27. DOI:10.1146/annurev.nutr.27.061406.093659. PMID 17439362. Проверено 2008-12-29.(недоступная ссылка)
- ↑ Leakey Richard The Origin of Humankind. — New York, NY: Basic Books, 1994. — P. 87–89. — ISBN 0465053130
- ↑ 1 2 Cavalli-Sforza L. L., Feldman M. W. The application of molecular genetic approaches to the study of human evolution. Nature genetics, 2003, v. 33, p. 266—275
- ↑ Африканское кроворазделение
- ↑ Быстров А. П. Прошлое, настоящее, будущее человека. Медгиз, Ленинградское отд., 1957, 314 c.
- ↑ Яблоков А. В., Юсупов А. Г. Эволюционное учение: Учебное пособие для биол. спец. вузов. 6-е изд., испр. М.: Высшая школа, 2006. стр. 251
Литература
Ссылки
dic.academic.ru