
Длительность одиночного мышечного сокращения 200 мсек – 2. Физиология мышечного сокращения (Задачи №№ 2-1 – 2- 2 из сd- приложения к Учебнику «Физиология м основами морфологии» под ред. Судакова к.В., и в.Ф. Волкова [ 3 ])
зеленый практикуп по фозиологии — Стр 6
Изучение физиологических свойств и особенностей сердечной и гладкой мышечной ткани, будет проведено на занятиях по разделам
«Физиология сердца и кровообращение», «Пищеварение» и др.
Необходимо объяснить важность этого занятия для понимания других разделов физиологии и других дисциплин, связанных с изучением функции скелетных мышц.
Оставшуюся часть этого этапа занятия следует посвятить разбору,
контролю и коррекции исходного уровня знания студентов,
нарисовать и разобрать следующие схемы:
1.Кривую одиночного мышечного сокращения, с указанием его фаз в изометрическом и изотоническом режимах.
2.Кривые суммации одиночных мышечных сокращений (полная и неполная), с объяснением механизма.
3.Кривые зубчатого и гладкого тетанусов, посттетанической контрактуры.
4.Кривые, позволяющие одновременно оценить возбуждение (ПД),
изменение возбудимости и сокращение во время одиночного
мышечного сокращения и при суммации.
5.Кривые, позволяющие одновременно оценить сокращение и изменение концентрации свободного цитоплазматического кальция.
6.Кривые оптимума и пессимума сокращения мышц и условия их возникновения.
7.Кривые работоспособности различных видов ДЕ.
8.Кривые развития утомления в нервно-мышечномпрепарате.
9.Схемы строения саркомера и входящих в него структур.
10.Строение миозиновых и актиновых нитей, тропомиозина и тропонина.
51
11.Схемы, объясняющие процесс циклизации актомиозиновых мостиков.
12.Сопряжения процессов возбуждения-сокращенияи расслабления.
13.Схемы функционирования ионтранспортирующих систем
(каналов, | насосов | и | обменников), | участвующих | в |
электромеханическом сопряжении.
14.Схемы биохимических процессов энергетического обеспечения скелетных мышц при выполнении работы.
15.Физиологические свойства и особенности гладкой мышцы.
Следует обратить внимание студентов на то, что формирование мышечной ткани и её последующая дифференцировка — сложный многоэтапный процесс, в котором уровень конечной организации достигается только после завершения полового созревания. В
процессе онтогенеза развиваются не только отдельные мышечные волокна, но и их объединения — двигательные единицы, в которых состояние мышечных волокон определяется развитием соответствующих мотонейронов.
В эмбриональном периоде формирование мышечной ткани начинается на 4 — 6-йнеделе внутриутробного развития и характеризуется образованием первичных мышечных веретён
(миотрубки), к которым в последующем прирастают отростки мотонейронов спинного мозга. С этой стадии процесс формирования мышечных волокон связан в первую очередь с развитием мотонейронов спинного мозга. Это происходит на 6 — 7-оммесяце внутриутробного развития, и ребенок рождается с мышцами, уже частично прошедшими этап первичной дифференцировки, примерно
53 . В возрасте от 1 до 2 лет, дифференцировочные процессы резко усиливаются и уже можно выделить волокна с “быстрым” (тип DE II
52
B), с “медленным” (тип DE I) и “промежуточным” (тип DE II A)
миозином.
В возрасте от 5 до 10 лет в соотношении между волокнами различного типа устанавливается относительная стабильность, но затем в возрасте 11 — 12 лет наступает резкое увеличение числа волокон с
“быстрым” миозином. В возрасте 14 лет наблюдается увеличение количества волокон типа DE I. На этом этапе наблюдается резкое увеличение темпа роста мышечной ткани. К 17 — 18 годам устанавливается характерное для взрослых, соотношение мышечных волокон разного типа и их поперечные размеры.
В биохимическом отношении мышцы у детей, характеризуются низким содержанием миофибриллярных белков, сниженной АТФ —
азной и высокой холинэстеразной активностью. По мере роста ребёнка эти отличия сглаживаются с показателями детей старшего возраста и взрослыми. Растёт содержание миозина и актина,
тропомиозина и тропонина, белков саркоплазматического ретикулума.
С возрастом уменьшается количество гликогена, молочной кислоты,
нуклеиновых кислот и воды относительно массы мышечной ткани.
В функциональном отношении мышцы новорождённого характеризуются повышенной чувствительностью к некоторым гуморальным агентам, в том числе и к ацетилхолину. Отмечается низкая чувствительность к действию электрического тока при внутриутробном развитии, которая сохраняется и после рождения,
имея высокие показатели хронаксии. В этот период мышцы воспроизводят лишь 3 — 4 сокращения в секунду. С возрастом число сокращений доходит до 60 — 80, с последующей способностью к формированию тетанических сокращений. При этом, созревание
53
мионеврального синапса приводит к более лучшей передаче возбуждения (более чем в 4 раза) с нерва на мышцу.
2. Основной этап занятия.
На этом этапе занятия студенты выполняют практические работы,
обсуждают полученные результаты и делают выводы. Перед началом выполнения практических работ, преподаватель объясняет особенности выполнения работ по методическому пособию и в дальнейшем следит за правильностью выполнения. Необходимо акцентировать внимание студентов на значимости проводимых работ.
3. Заключительный этап занятия.
В заключение занятия преподаватель подводит итоги полученных знаний и умений студентов посредством либо тестового контроля,
либо решения ситуационных задач. В заключение занятия преподаватель дает задание на следующее занятие, проверяет и подписывает отчеты студентов.
Лабораторные работы
1.Приготовление нервно-мышечногопрепарата.
2.Знакомство с работой измерительно-регистрирующейустановки.
3.Анализ одиночного мышечного сокращения.
4.Зависимость сокращения мышц от силы раздражения.
6.Физиологические свойства и особенности гладких мышц.
7.Динамометрия.
8.Эргография.
9.Работы на компьютерных моделях мышечного сокращения.
10.Выполнение работ с использованием виртуального практикума.
11.Просмотр видеофильмов.
5. Задачи для определения конечного уровня знаний студентов.
54
Задача №1.
Какие процессы протекают в мышце во время латентного периода при непрямом раздражении?
Эталон ответа.
Раздражение называется непрямым, если оно производится через нерв, подходящий к мышце. От момента раздражения нерва до момента начала мышечного сокращения происходят следующие события:
Возбуждение нерва — движение возбуждения по нерву — возбуждение пресинаптической мембраны — электросекреторное сопряжение —
диффузия медиатора — возбуждение мембраны мышечного волокна —
движение возбуждения по мышечному волокну —
электромеханическое сопряжение — активация актомиозинового комплекса — сокращение.
Задача №2.
Длительность рефрактерного периода мышцы 10 мсек. Длительность одиночного сокращения 200 мсек. Назовите интервал частот раздражения, при которых данная мышца будет сокращаться в режиме гладкого тетануса?
Эталон ответа.
Для гладкого тетанического сокращения необходимо, чтобы интервал между раздражениями был длиннее рефрактерного периода, но короче продолжительности фазы укорочения. В данном случае этот интервал лежит в пределах от 10 до 70 мсек, значит при частоте от 15 до 100 Гц будет наблюдаться тетанус. При меньшей частоте будут одиночные сокращения, при большей — пессимум.
Задача №3.
55
Длительность потенциала действия мышцы 10 мсек. Какую частоту раздражения следует дать, чтобы каждый импульс попадал в фазу супернормальной возбудимости?
Эталон ответа.
Так как длительность потенциала действия совпадает с периодом рефрактерности, после которой следует фаза супернормальной возбудимости, интервал между раздражениями должен быть 10 мсек.
Это соответствует частоте раздражения 100 Гц. Чтобы импульс попал в субнормальный период, необходимо применение более частых раздражений.
Задача №4.
При раздражении нерва, нервно-мышечногопрепарата, мышца доведена до утомления. Что произойдет, если в это время подключить прямое раздражение мышцы?
Эталон ответа.
Мышца снова начнет сокращаться, так как при раздражении нервно-
мышечного препарата утомление раньше всего наступает в синапсе.
Задача №5.
При непрямом раздражении, мышца сокращается тетанически. Как изменится ритм сокращения, если в перфузируемый раствор ввести d-
тубокурарин?
Эталон ответа.
Мышца расслабится, так как d-тубокураринблокирует передачу импульсов в холинэргических синапсах.
Задача №6.
Площадь физиологического поперечного сечения мышцы 25 кв. см.
Рассчитайте удельную силу мышцы, если она в состоянии поднять максимально 200 кг.
56
Эталон ответа.
Удельная сила мышцы равна отношению максимального груза к площади физиологического поперечного сечения. В данном случае она равна 8 кг/см2.
Задача №7.
Мышцу нервно-мышечногопрепарата подвергают непрямому раздражению. Через некоторое время амплитуда сокращений начинает уменьшаться. Означает ли это, что в мышце наступило утомление? Как проверить это предположение?
Эталон ответа.
Нет. Утомление может возникнуть как в мышце, так и синапсе. Чтобы проверить это предположение нужно перейти к прямому раздражению мышцы. Если при этом амплитуда сокращений возрастает, утомление возникло не в мышце, а в синапсе.
Задача №8.
К покоящейся мышце подвесили груз. Как при этом изменится ширина Н-зонысаркомера?
Эталон ответа.
Н-зонаэто центральный участок толстой протофибриллы, который не перекрыт актиновыми нитями. При растяжении мышцы степень перекрытия миозиновых нитей уменьшается, так как актиновые нити частично выходят из промежутков между миозиновыми.
Соответственно, ширина Н-зоныувеличивается.
Задача №9.
Почему быстрые мышцы при сокращении потребляют в единицу времени больше энергии АТФ, чем медленные?
Эталон ответа.
57
При быстром сокращении мостики совершают больше гребковых движений в единицу времени, и на это затрачивается больше энергии АТФ.
Задача №10.
При перерезке двигательного нерва мышца, которую он иннервировал, атрофируется. Чем можно это объяснить?
Эталон ответа.
Иннервация обеспечивает не только сократительную деятельность, но и трофические влияния. При денервации нарушается синтез белков в мышце и уменьшается ее масса — происходит атрофия.
ПЕРЕЧЕНЬ ВОПРОСОВ К ИТОГОВОМУ ЗАНЯТИЮ ПО
РАЗДЕЛУ:
«ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ».
1.Плазматическая мембрана. Классификация, строение и молекулярный состав. Роль, значение и функции плазматических мембран.
2.Транспорт веществ через биологические мембраны. Активный и пассивный транспорт, его виды.
3.Общие свойства возбудимых тканей. Определение возбудимости.
Критерии оценки возбудимости. Соотношение между силой раздражителя и временем его действия на возбудимую ткань.
Хронаксия. Реобаза. Полезное время.
4. Мембранно-ионнаятеория происхождения потенциала покоя.
Ионный градиент и причины его возникновения.
5. Потенциал действия, его величина, способы регистрации, ионные механизмы происхождения. Следовые потенциалы.
58
6.Фазовые изменения возбудимости при возбуждении. Закон «Все или ничего».
7.Локальный ответ в сравнении с импульсным возбуждением.
8.Изменение физиологических свойств возбудимых тканей при прохождении постоянного тока. Законы действия постоянного тока.
Явление аккомодации.
9.Парабиоз нерва, его фазы и причины возникновения. Значение учения о парабиозе для практической медицины.
10.Функции нервов. Классификация нервных волокон. Законы проведения возбуждения по нервам.
11.Механизмы проведения возбуждения по нервным волокнам.
12.Физиологические свойства мышечной ткани. Двигательные единицы. Изометрическое, изотоническое и ауксотоническое сокращение.
13.Тетанус. Механизмы возникновения тетануса (Р. Гельмгольц, Е.Б.
Бабский, Н.Е. Введенский). Факторы, влияющие на величину тетануса. Контрактура и причины ее возникновения.
14. Оптимум и пессимум частоты и силы раздражения по методу Н.Е.
Введенского.
15.Ультрамикроскопическое строение скелетной мышцы. Теория скользящих нитей. Сопряжение возбуждения и сокращения. Роль АТФ, кальция, сократительных и регуляторных белков в мышечном сокращении.
16.Работа и сила мышц. Зависимость работы от величины нагрузки и ритма мышечных сокращений. Закон средних нагрузок.
17.Утомление, его характеристика. Теории утомления. Активный отдых по методу И.М. Сеченова. Адаптационно-трофическоевлияние симпатической нервной системы на скелетную мускулатуру.
59
18. Строение и механизм передачи возбуждения через нервно-
мышечный синапс. Особенности проведения возбуждения через синапс. Миорелаксанты. Классификация синапсов.
19.Строение и механизм передачи возбуждения через электрический синапс. Особенности проведения возбуждения через данный вид синапса.
20.Физиологические свойства и особенности гладких мышц.
60
studfiles.net
Суммация одиночных мышечных

План:
Суммация сокращений и тетанусы.
Оптимум и пессимум (Введенский).
Эргография. Закон средних нагрузок.
Основные отличия в строении и функционировании скелетной и гладкой мышц. Особенности сократительной деятельности у детей.
Одиночное сокращение и его фазы.
Различают 2 основных вида мышечных сокращений:
одиночные и
тетанические.
Механический ответ мышечного волокна или отдельной мышцы на однократные их раздражения нервным импульсом или кратким толчком тока, называется одиночным сокращением.
При этом в миоплазме мышцы происходит кратковременный подъем концентарации Ca++ вн., сопровождаемый кратковременной работой — тягой мостиков, сменяющейся снова покоем. В изометрическом режиме одиночное напряжение начинается через 2 мсек. после пика ПДм, причем напряжению предшествует краткое и незначительное, так называемое латентное расслабление, создаваемое выходом Ca++ из саркоплазматического ретикулума, что приводит к потере ретикулумом тургора.
В естественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон.
Механограммы мышечных сокращений
Анализ одиночного мышечного сокращения
Действующий раздражитель
Латентный период
Фаза расслабления
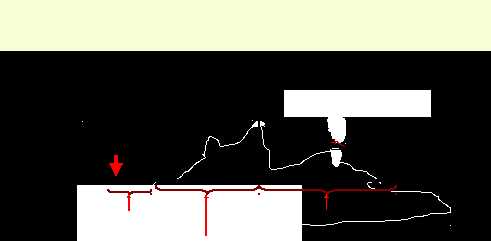
Фаза укорочения
Время, 0,01 с
Соотношение фаз возбудимости, ПД и сокращения.
При нанесении одиночного порогового и сверхпорогового раздражения на мышцу, мышца сокращается, а затем расслабляется, то есть наблюдается одиночное сокращение мышц.
Имеет место ряд последовательных явлений:
после нанесения раздражения сокращение мышцы наступает не сразу, а через некоторый интервал 2,5 мсек. (латентный период).
Реакция мышцы на раздражение начинается с генерации
ПД продолжительностью 3-5 мсек. и в это же время
(фаза сокращения),
затем следует фаза расслабления, по продолжительности либо равна или может быть несколько больше фазного сокращения. Длительность зависит от морфофункциональных свойств.
Так, у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения равна 7-10 мсек., а у медленных волокон камбаловидной мышцы равна 50-100мсек.
Возбудимость мышц в ходе одиночного
сокращения меняется.
Во время развития ПД, т.е. в начале сокращения мышцы наступает абсолютная рефрактерная фаза (состояние невозбудимости, длительностью 3-5 мсек.).
рефрактерности, а затем
Длительность этих периодов различна у животных разных видов и зависит от функционального состояния нервно- мышечного аппарата.
Тетанус и его виды
Суммация сокращений и тетанус возникают в том случае, если на мышцу наносится не одно, а несколько раздражений с определенным интервалом времени.
Рассмотрим пример:
Если интервал между раздражениями будет больше длительности одиночного сокращения (больше 100 мсек.), то зарегиструются 2 одиночных сокращения. Уменьшая интервал между раздражениями от
100 до 50 мс ек
( частота 10-20 Гц), т.е. второе раздражение будет наноситься в фазу расслабления и амплитуда сокращения мышцы на второе раздражение будет больше, т.к. второе сокращение возникает тогда, когда мышца еще не успела полностью расслабиться, т.е. происходит суммация сокращений. При этом на миограмме регистрируется две вершины.
Если интервал между раздражениями ме ньше 50 мс ек
(частота более 20 ГЦ), то второе раздражение воздействует на мышцу в конечный период фазы сокращения и произойдет полное слияние двух сокращений. Однако это будет наблюдаться до тех пор, пока интервал между раздражениями превышает длительность возникающего перед сокращением ПД, т.к. во время его развития резко падает возбудимость (фаза абсолютной рефрактерности.) и на второе раздражение, следующее с интервалом ме нее 5 мс ек .
(частота более 200 Гц) — не реагирует.
Зубчатый тетанус
Развивается на ряд последовательных раздражений, интервал между которыми больше, чем длительность фазы сокращения мышцы.
Гладкий тетанус
Возникает тогда, когда интервал между раздражениями меньше длительности фазы сокращения, но больше чем продолжительность потенциала действия.
У разных мышц длительность фаз одиночного сокращения неодинакова, а, следовательно, частота стимуляции или естественного возбуждения мышцы для получения тетанического сокращения различны.
Тетанус характеризуется слиянием одиночных мышечных сокращений в непрерывное укорочение мышцы, которое по амплитуде превышает уровень, достигаемый при одиночном сокращении.
В случае гладкого тетануса после нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое двигательными единицами напряжение в 2-4 раза больше, чем при одиночных сокращениях.
В режиме тетанического сокращения мышца способна работать лишь короткое время. Это объясняется тем, что из-за отсутствия периода расслабления она не может восстановить свой энергетический потенциал и работает как бы « в долг».
сокращений
 Механизм суммации одиночных
мышечных сокращений
Механизм суммации одиночных
мышечных сокращений
Одиночное сокращение 2
Суммация сокращений
Одиночное сокращение 1
Действующие раздражители
1 2
Время, 0,01 с
Зубчатый тетанус Гладкий тетанус

Сокращение мышцы в зависимости от
частоты раздражения
Свыше 100 раздражений / c
До 35 раздражений / c
До 25 раздражений / c
До 20 раздражений / c
Одиночное сокращение
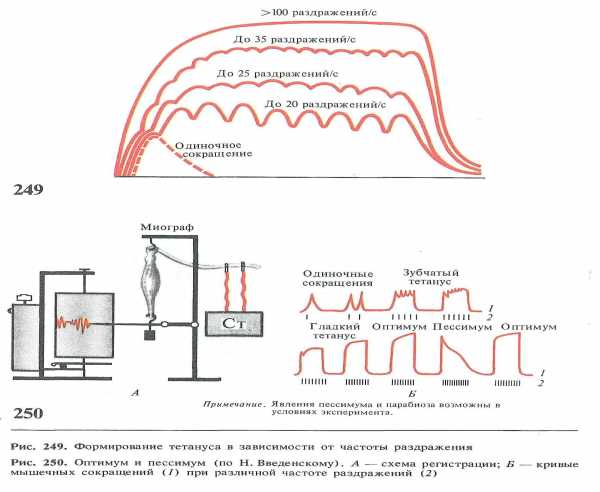
В естественных условиях мышечные волокна, двигательные единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон.
Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 12 импульсов в секунду, а быстрых волокон глазодвигательных мышц – при скорости импульсации мотонейрона менее 50 импульсов в секунду.
Гладкий тетанус для быстрых и медленных мышц достигается при разных частотах импульсации мотонейронов. Зависит это от времени одиночного сокращения. Так, гладкий тетанус для быстрой глазодвигательной мышцы проявляется при частоте свыше 150-200 импульсов в секунду, а у медленной
камбаловидной мышцы – при частоте около 30 импульсов в секунду.
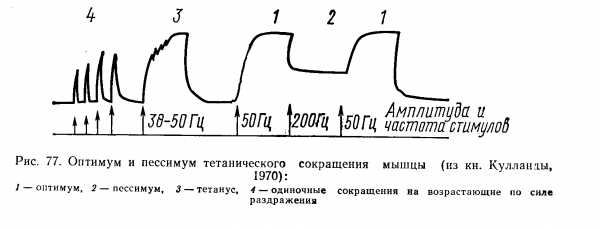
Оптимум и пессимум ( Введенский)
Высокие частоты раздражения, вызывающие уменьшение сокращения мышцы, Введенский назвал пессимальными частотами раздражения, а ответ мышцы на это раздражение — пессимальным сокращениям. Уменьшение частоты стимуляции тотчас приводит к восстановлению высокого уровня тетанического сокращения.
Чтобы разбираемая ситуация имела конкретные числовые выражения, что облегчит понимание, условно определим интервалы временной продолжительности
Рассмотрим фазы: аболютная рефрактерность=5 мсек.
относительная рефрактерность от 5-10 мсек.
супернормальная. возбудимость от 10 до 40 мсек
Отсюда можно рассчитать частоту ритма раздражения, при котором импульсы раздражающего тока будут совпадать с той или иной фазой возбудимости.
Расчет показывает, что
при раздражении с частотой ритма 200 импульсов/сек многие импульсы тока будут действовать на ткань, когда она находится в состоянии абсолютной рефрактерности и не способна отвечать на них и, следовательно, они будут неэффективными, а ритм раздражения подвергается трансформации в более медленный ритм возбуждения. Если подобное возникает при исследовании сократительной способности мышц, то в этих условиях степень сокращения мышц будет меньше, чем при раздражении тока меньшей частоты.
При увеличении интервала между раздражениями от 5 до 10 мсек.( от 200 до 100 импульсов в секунду), каждое следующее раздражение будет наноситься в фазу относительной рефрактерности. В этом частотном интервале выявляется наивысшая частота, которая способна воспроизводиться исследуемой структурой без искажения ритма, что соответствует лабильности данной структуры.
Последующее увеличение интервала между раздражениями от 10 до 40 мсек.( частоты от 100 до 25 импульсов в секунду) создает условия, когда каждое раздражение приходится на фазу супернормальной возбудимости, что благоприятствует возникновению возбуждения, и при этом можно ожидать максимального сокращения мышцы. Частота, которая вызывает максимальный сократительный эффект, была названа Введенским оптимальной частотой раздражений, а сокращение — оптимальным.
Изменяя силу раздражений при их фиксированной частоте, Введенский показал, что оптимум и пессимум сокращения зависят и от силы раздражения.
Сила и работа мышц. Динамометрия.
Эргография. Закон средних нагрузок
Сила, развиваемая мышцей или пучком мышечных волокон, является суммой сил отдельных волокон. Чем толще мышца и больше ее
«физиологическая S» поперечного сечения (физиологическая сумма S поперечного сечения отдельных волокон), тем сильнее мышца. Так,
например, при мышечной гипертрофии сила и толщина мышцы возрастают в одинаковой степени.
Мышечная сила зависит не только от активирующего влияния со стороны ЦНС, но также определяется внешними механическими условиями, при которых работает мышца.
Итак, сила мышцы оценивается весом груза, который она при максимальном возбуждении способна удерживать не меняя своей длины.
Сила мышцы зависит от:
суммы сил мышечных волокон ( их сократительной способности)
количества мышечных волокон в мышце и количества функциональных единиц, одновременно возбуждающихся при развитии напряжения
исходной длины мышцы (предварительно растянутая мышца
развивает
большую силу)
характера регулятивных явлений
условий взаимодействия мышцы с костями скелета
В организме человека скелетные мышцы передают силу костям скелета через посредство эластических, обладающих растяжимостью структур- сухожилий. Во время развития силы возникает тенденция к укорочению мышцы и, следовательно, к растяжению и развитию напряжения эластических структур, прикрепляющих мышцы к костям.
Мышечное сокращение, когда длина мышцы уменьшается с увеличением ее силы, наз. ауксотоническим сокращением. Максимальная ауксотоническая сила гораздо меньше, чем сила сокращения, развиваемого мышцей при постоянной длине, то есть при изометрическом сокращении.
Сила изометрического сокращения и длина
мышцы
В состоянии расслабления мышца, удерживаемая при «длине покоя» путем фиксации обоих ее концов, не развивает силу, которая передавалась бы на держатель. Однако, если один конец потянуть чтобы волокна растянулись в мышце развивается пассивное напряжение.
Таким образом, покоящаяся мышца эластична, в отличие от резиновой полоски ее напряжение не возрастает линейно при напряжении.
Отложив измеряемую силу в зависимости от длины в прямоугольной системе координат, получим график длина— напряжение для покоящейся мышцы, т.е. кривую напряжения покоя.
Кривая нарастает тем круче, чем больше степень растяжения мышцы.
Следовательно, модуль эластичности покоящейся мышцы возрастает с растяжением. Эластичность свойственна главным образом растяжимым структурам, которые располагаются параллельно по отношению к сократительным миофибриллам, к ним относятся
сарколемма, окружающая мышечное волокно,
продольная система саркоплазматического ретикулума,
элементы соединительной ткани между волокнами.
В отличие от этих структур миофибриллы в расслабленном состоянии практически не оказывают сопротивления растяжению, актиновые и миозиновые нити не связаны поперечными мостиками и могут легко передвигаться друг относительно друга.
Степень предварительного растяжения определяет не только величину пассивного эластического напряжения покоящейся мышцы, но и величину дополнительной силы, которую может развивать мышца в случае ее активации при данной исходной длине. Изометрический прирост силы во время сокращения суммируется с пассивным напряжением покоящейся мышцы. Пассивные эластические силы растянутых продольных трубочек и сакролеммы суммируются с активными сократительными силами миофибрилл.
Сократительная способность мышцы характеризуется ее абсолютной силой (сила, приходящаяся на 1 см 2 поперечного сечения мышечных волокон).
Показатель рассчитывается так:
Силу мыш цы
S ее физиологического поперечника (т.е. сумму S всех мышечных волокон мышцы).
У перистых мышц физиологический поперечник больше, чем у II
волокнистых, и поэтому их сила больше.
Абсолютная сила мышц (в кг на 1 см) у человека:
Икроножная- 6, 24;
Разгибательная шеи – 9.0;
Жевательная- 10,0
Трехглавая- 16,8
Одиночное мышечное волокно способно развить усилие в 20-30
тонн.
ЦНС изменяет силу сокращения мышцы путем изменения количества одновременно участвующих в сокращении функциональных единиц, а также частоты поступающих к ним импульсов. Симпатическая НС увеличивает силу сокращений.
Работа мышцы
Измеряется произведением поднятого груза на величину укорочения мышцы.
Между грузом, который поднимает мышца и выполняемой ею работой существует зависимость.
Внешняя работа мышцы равна 0, если мышца сокращается без
нагрузки.
По мере увеличения груза работа вначале увеличивается, а затем постепенно уменьшается.
При очень большом грузе, который мышца неспособна поднять, работа равна 0.
Внешняя механическая работа мышцы (А) возможна только в изотоническом режиме работы с грузом при условии, что вес груза меньше общей силы мышцы. В этом случае сократительный аппарат сначала растягивает и напрягает эластический элемент – сухожилие, а затем поднимает подвешенный к нему груз.
Внешняя механическая работа мышцы максимальна при средних нагрузках. Это называется законом средних нагрузок.
Наибольшую работу мышца совершает при некоторых средних нагрузках. В данном случае 200-250 г.
Мощность мышцы, измеряемая величиной работы в единицу времени,
также достигает максимальной величины при средних нагрузках.
Работа мышцы, при которой происходит перемещение, волокна развивают напряжение, но почти не укорачиваются (мышца при этом сокращается в изометрическом режиме) называется статической.
Статическая работа более утомительна, чем
динамическая
Например, при более динамическом сокращении мышцы могут эффективно выполнять свою функцию, если статическое напряжение других мышц обеспечивает сохранение покоя.
Динамометрия – метод измерения мышечной силы.
Эргография – метод определения мышечной работоспособности.
Регистрирует работу группы мышц, совместно выполняющих данные движения. Так, эргограф Моссо записывает движения нагруженного пальца и общего разгибателя всех пальцев руки. Форма кривой утомления и величина произведенной работы широко варьируют у разных людей и зависят от разнообразных факторов внешней среды.
Функциональные особенности гладких мышц
Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эффективными волокнами симпатической и парасимпатической НС, а также местными гуморальными и физическими воздействиями.
Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствии их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчерченности.
Гладкомышечные клетки имеют веретенообразную форму, l =
500-400 мкм. и толщину 2-10 мкм. Отделены друг от друга узкими щелями (60-150 нм).
Гладкая мышца с морфологической точки зрения является не истинным, а функциональным синцитием. Клетки в гладкой мышце расположены хаотично, неравномерно, иногда они собраны в пучки или тяжы. Их окружает соединительная ткань.
studfiles.net
Общая физиология возбудимых тканей
2-1. Как изменится мембранный потенциал, если поток натрия внутрь клетки увеличится, а количество калия останется прежним?
2-2. Как изменится мембранный потенциал нервного волокна, если закрыть натриевые каналы?
2-3. Как изменится мембранный потенциал, если заблокировать работу Nа-K-зависимой АТФ-азы?
2-4. Порог раздражающего тока 3 в. Ткань раздражается током в 10 в., но возбуждения не возникает. В каком случае это может наблюдаться?
2-5. Возникает ли распространяющееся возбуждение в нерве, если известно, что мембранный потенциал равен 90 мв., критический уровень деполяризации на 30% ниже, а раздражающий ток сдвигает мембранный потенциал в одном случае на 10 мв., в другом — на 30 мв.?
2-6. Как изменится возбудимость ткани, если при гиперполяризации мембраны критический уровень деполяризации остается прежним?
2-7. Как изменится возбудимость ткани, если мембранный потенциал вырос на 20%, а критический уровень деполяризации — на 30%? Исходные величины : Ео=90 мв., Ек = 60 мв.
2-8. В результате длительного раздражения постоянным током критический уровень деполяризации (Ек) упал на 20%. Величина деполяризации — 10% от уровня мембранного потенциала (Ео). Исходные величины Ео = 100 мв7, Ек=70 мв. Как изменится возбудимость нерва в данном случае?
2-9. Каким образом и на какую величину должен сдвинуться критический уровень деполяризации, чтобы на аноде возникло возбуждение при размыкании постоянного тока, который увеличит Ео на 10 мв.? Принять Ео =100 мв., Ек = 70 мв.
2-10. Порог раздражения под анодом при размыкании 2 в. Сократится ли мышца при замыкании и размыкании, если раздражать нервно-мышечный препарат восходящим током в 1,9 в.?
2-11. Реобаза размыкательного удара 3 в. Нерв раздражается током в 10 в. Направление тока нисходящее. Что произойдет с мышцей при размыкании цепи раздражающего тока?
2-12. Нерв между раздражающими электродами перевязан. При действии тока мышца данного нервно-мышечного препарата сокращалась только в момент замыкания. Какой электрод находится ближе к мышце?
2-13. Какой электрод находится ближе к мышце, если при действии сильного тока сокращение возникает только при замыкании?
2-14. Схема какого процесса приведена ниже? Добавьте недостающие звенья.
Раздражение нерва —- ? —— вход Na внутрь клетки —— ? — генерация ПД и перезарядка мембраны — ? — увеличение К — проницаемости —— реполяризация мембраны—— активация Na-K-зависимой АТФ-азы — — восстановление мембранного потенциала.
2-15. Мембранный потенциал нервного волокна равен 100 мв. Критический уровень деполяризации отличается от мембранного потенциала на 30%. Какова реобаза нерва, если 1 в. раздражающего тока сдвигает Ео на 5 мв.?
2-16. Постройте кривую силы-длительности по следующим данным:
0,5 в 1000 мсек
1,0 в 80 мсек
1,2 в 40 мсек
2,0 в 25 мсек
3,0 в 10 мсек
4,0 в 9 мсек
Определите ориентировочные параметры возбудимости данной ткани.
2-17. Правильно и представлена последовательность событий, которые приводят к возбуждению нерва? Под каким электродом это происходит?
Пассивная деполяризация —- усиление потока Na в клетку —- повышение натриевой проницаемости —- локальный ответ —- активная деполяризация —- потенциал действия.
2-18. Нерв раздражается электрическими стимулами разной формы:
1 2 3 4
Укажите, при какой форме импульса порог раздражения будет наименьшим и почему?
2-19. Нарисуйте кривые тока действия при следующих методах отведения:
1 2
2-20. Какой ответ дает возбудимая мембрана на раздражение, близкое к пороговой силе?
2-21. Изменится ли величина потенциала покоя, если искусственно снизить на 30% концентрацию ионов К внутри нервного волокна?
2-22. Какая из перечисленных возбудимых структур характеризуется наибольшей возбудимостью: нерв, синапс или мышца ? У какой структуры лабильность наименьшая?
БА
2-23. Какая из структур, указанных на схеме ( А,Б,С,Д), обладает повышенной химической чувствительностью. Замените буквы обозначениями..Б
С
Д
2-24. Нарисуйте схему соотношения кривых потенциала действия, возбудимости и одиночного сокращения скелетной мышцы.
2-25. Какие процессы протекают в мышце во время латентного периода при непрямом раздражении?
2-26 Проверьте, правильно ли расставлены частоты раздражения на миограммах, приведенных ниже:
5 гц 80 гц 60 гц 20 гц
2-27. Какая из представленных кривых утомления получена при раздражении нормальной мышцы? В каком случае может быть получена вторая кривая? Почему?
1 2
2-28. Добавьте недостающие звенья в цепи следующих процессов:
Нервный импульс — высвобождение ацетилхолина — ? — повышение ионной проницаемости постсинаптической мембраны — ? — возникновение ПД и распространение его по мышечному волокну.
2-29. Длительность рефрактерности мышцы 10 мсек. Длительность одиночного сокращения 200 мсек. Назовите интервал частот раздражения, при которых данная мышца будет сокращаться в режиме гладкого тетануса.
2-30. Скорость проведения возбуждения по мембране мышечного волокна равна 5 мсек. Чему равна скорость проведения волны сокращения? Через какое время волна сокращения дойдет от одного конца мышцы до другого, если длина мышечного волокна 10 см.?
2-31. Длительность потенциала действия мышцы 10 мсек. Какую частоту раздражения следует дать, чтобы каждый импульс попадал в фазу супернормальной возбудимости?
2-32. Схема какого процесса приведена ниже? Добавьте недостающие звенья.
Раздражение — возникновение ПД — проведение его вглубь волокна — — ? — взаимодействие актина и миозина — ? — активация Са-насоса — — ? — расслабление мышцы.
2-33. При раздражении нерва нервно-мышечного препарата мышца доведена до утомления. Что произойдет, если в это вре- мя подключить раздражение мышцы?
2-34. Мышца сокращается тетанически. Как изменится ритм ее сокращения, если в перфузируемый раствор ввести атропин?
2-35. При каждом импульсе возбуждения в межфибриллярное пространство выходит 1 функциональная условная единица Са. Весь Са возвращается в ретикулюм с постоянной скоростью за 10 мсек. При какой частоте раздражения будет происходить суммация сокращений? Через сколько циклов при частоте 50 герц амплитуда сокращения достигнет максимума, если одна функциональная единица Са соответствует критической концентрации Са?
2-36. Рефрактерный период мышцы равен 5 мсек. Лабильность мышцы в 4 раза меньше теоретической. Нарисуйте миограммы при следующих частотах раздражения: 10 герц, 50 герц, 100 герц.
2-37. В эксперименте на нервно-мышечном препарате было определено, что при неизменной силе тока пессимум наступает при частоте 150 герц. При какой частоте раздражения можно получить на этом препарате состояние оптимума? Какова лабильность нервно-мышечных синапсов данного препарата?
2-38. Площадь физиологического поперечного сечения мышцы 25 кв. см. Рассчитайте удельную силу мышцы, если она в состоянии поднять максимально 200 кг.?
2-39. Перед Вами два нерва. Как будет проходить по ним возбуждение, если нарушить целостность мембраны в указанных участках? Раздражение наносится в точку А.
1
2
2-40. Сколько времени потребуется на регенерацию нерва, если его длина до травмы была равна 45 см.? Скорость регенерации принять за 3 мм/сутки. Нерв перерезан на границе верхней и средней его трети.
2-41. Каким будет время проведения возбуждения по нерву типа А, если расстояние между раздражающими и регистрирующими электродами 10 см.?
2-42. Каким будет время проведения возбуждения по волокну типа В, если расстояние между раздражающими и отводящими электродами равно 8 см.?
2-43. Через какое время волна возбуждения достигнет отводящих электродов, если они наложены на нервное волокно типа С на расстоянии 5 см. от раздражающих электродов.
2-44. К какому типу относится нервное волокно, если при межэлектродном расстоянии в 5 см. время проведения возбуждения равно 0,05 сек.?
2-45. На рисунке изображена серия потенциалов действия при частом и длительном раздражении. Что произойдет с нервом, если теперь нанести новое пороговое раздражение в период, отмеченный стрелкой?. Как называется это явление?.
. . . .
2-46. Каково время проведения возбуждения по мякотному волок ну, если между отводящими и регистрирующими электродами находится 15 перехватов Ранвье?
2-47. Определите, сколько перехватов Ранвье находится между электродами, если известно, что возбуждение проходит это расстояние за 140 мсек.
2-48. Определите центральное время рефлекса в сложной рефлекторной дуге, если в ее составе 15 синапсов (без учета времени распространения возбуждения по нервам).
2-49. Сколько синапсов входит в состав центральной части рефлекторной дуги рефлекса, если его центральное время равно 100 миллисекундам?
2-50. Параметры возбудимости ткани: реобаза, хронаксия, лабильность. Все ли параметры перечислены?
2-51. Что покажет гальванометр, если: а) микроэлектрод проколол мембрану; б) введен глубоко вглубь клетки?
2-52. Если бы клеточная мембрана была абсолютно непроницаема для ионов, как бы изменилась величина потенциала покоя?
2-53. Тетродотоксин — яд, блокирующий натриевые каналы. Как влияет этот яд на величину потенциала покоя?
2-54. Батрахотоксин — сильный яд, который значительно увеличивает натриевую проницаемость мембраны в покое. Как этот яд повлияет на величину ПП?
2-55. Гигантский аксон кальмара поместили в среду которая по своему составу соответствовала межклеточной жидкости. При раздражении в аксоне возник ПД. Затем концентрацию ионов натрия в среде уравняли с их концентрацией в аксоне и повторили раздражение. Что обнаружили?
2-56. Как изменится кривая ПД при замедлении процесса инактивации натриевых каналов?
2-57. Возбудимость нервных волокон выше, чем мышечных. Почему?
2-58. Почему гиперполяризация мембраны приводит к снижению возбудимости?
2-59. Что произойдет с нервной клеткой, если ее обработать цианидами?
2-60. Нерв раздражают с частотой 10, 100 и 1000 раз в секунду. Сколько ПД будет возникать в каждом случае?
2-61. Концентрацию ионов натрия внутри нервной клетки повысили. Как это повлияет на возникновение ПД?
2-62. Может ли какое-либо вещество повлиять на состояние нервной клетки, если это вещество не способно пройти через клеточную мембрану?
2-63. Если обработать нерв протеолитическими ферментами, то пострадают ли при этом механизмы, связанные с генерацией ПД?
2-64. Два человека случайно подверглись кратковременному действию переменного тока одинаково высокого напряжения, но разной частоты. В одном случае частота тока составляла 50 гц, в другом — 500000 гц. Один человек не пострадал, другой получил электротравму. Какой именно?
2-65. Может ли воздействие на человека высокочастотного тока, который не вызывает возбуждения из-за кратковременности действия каждого колебания тока, вызвать, тем не менее, патологический эффект?
2-66. Почему возбуждение, переходя в участок, соседний с возбужденным, не возвращается в уже пройденную точку?
2-67. При перерезке двигательного нерва мышца, которую он иннервировал, атрофируется. Чем можно это объяснить?
2-68. Мышцу нервно-мышечного препарата подвергают непрямому раздражению. Через некоторое время амплитуда сокращений начинает уменьшаться. Означает ли это, что в мышце наступило утомление? Как проверить это предположение?
2-69. К покоящейся мышце подвесили груз. Как при этом изменится ширина Н-зоны саркомера?
2-70. Почему быстрые мышцы при сокращении потребляют в единицу времени больше энергии АТФ, чем медленные?
studfiles.net
Методика проведения кистевой динамометрии
Для работы необходимы: кистевой динамометр, секундомер, весы для измерения массы тела. Объект исследования–человек.
Проведение работы : Измерения рекомендуется проводить на нескольких испытуемых, поскольку в этом случае будет заметна разница выраженности индивидуальных реакций. Регистрацию каждого показателя экспериментатор проводит с обеих сторон и отмечает его выраженность и симметричность.1) Для определения абсолютного показателя силы мышц-сгибателей кисти испытуемый в положении стоя отводит вытянутую руку с динамометром (подвижной частью к пальцам) под прямым углом к туловищу (на уровне плеча). Вторая, свободная рука, опущена и расслаблена. По сигналу экспериментатора испытуемый дважды выполняет максимальное усилие на динамометре (максимально его сжимает) на каждой руке. Фиксируется лучший результат. 2) Для определения среднего показателя силы мышц (Р), который отражает уровень работоспособности, испытуемый в исходном положении выполняет 10-кратные усилия с частотой 1 раз в 5 сек. Результаты записывают и по формуле вычисляют Р = (f1+f2+f3+…+fn) / n, где Р – средний показатель силы мышц, f1,f2,f3 – показатели динамометра при отдельных мышечных усилиях, n – количество попыток. 3)Показатель силы руки (ПСР) выражают в % и рассчитывают по формуле: ПСР (%) = абсолютная сила мышц (кг) х 100% / масса тела (кг). 4) Снижение уровня работоспособности рассчитывают по формуле: S=[(f1-fmin)/f max] х 100, где S –показатель снижения силы мышц, f1 – величина начального мышечного усилия, fmin – минимальная величина усилия, f max – максимальная величина усилия.
Оформление результатов и их оценка: запишите в протокол абсолютные показатели силы, вычислите уровень работоспособности (Р), ПСР и показатель снижения работоспособности мышц по результатам 10—кратных усилий. Начертите график, который выявит характер снижения работоспособности мышц: на оси абсцисс отложите порядковые номера усилий, на оси ординат–показатели динамометра при каждом усилии. Сравните результаты у нескольких испытуемых. Средние величины абсолютной мышечной силы кисти у человека правши составляют: правая кисть – м-35-45 кг, ж-25-33 кг; левая кисть – на 5-10 кг меньше. В среднем ПСР у м = 60-70%, у ж = 45-50 %.
ТЕСТОВЫЕ ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ УРОВНЯ знаний:
1. В эксперименте под действием химического вещества в мышцах ослаблена работа Са2 — насоса. Какие явления будут развиваться при этом?
А. Снижение скорости распространения ПД
В. Активация натрий-калиевого насоса
С. Увеличение продолжительности расслабления
D. Увеличение длительности ПД
Е. Снижение потенциала покоя
2. У спортсменов за счет тренировок может увеличится объем мышц. Какое вещество является непосредственным источником энергии мышечного сокращения?
А. Аденозинфосфат
В. Аденозинтрифосфат
С. Молочная кислота
D. Нейтральные жирные кислоты
Е. Креатинфосфат
3. Какой вид сокращения мышц верхней конечности имеет место при попытке поднять непосильный груз?
А. Фазический
В. Ауксотонический
С. Изотонический
D. Изометрический
Е. Все ответы верны
4. Если мышца развивает силу и при этом ее длина не уменьшается, то такой вид сокращения будет называться:
А. Изотоническим
В. Изомерическим
С. Ауксотоническим
D. Изометрическим
Е. Тетаническим
5. Если взаимодействие между актином и миозином в скелетных мышцах при патологическом процессе изменится таким образом, что связи могут образовываться, но не разрываться, то мышца:
А. Будет напряженной и неэластичной
B. Сократится с повышенной скоростью
С. При стимуляции гидролиз АТФ будет происходить на повышенном уровне
D. Во время стимуляции сократится и расслабится как обычно
Е. Утратит поперечную исчерченность
6. Исследуемый получил задание выполнить на протяжении 1 часа 1200 кГм2 работы. Какие условия работы обеспечивают выполнение этой работы?
А. Оптимальная нагрузка и максимальный ритм работы
В. Оптимальная нагрузка и оптимальный ритм работы
С. Максимальная нагрузка и оптимальный ритм работы
D. Максимальная нагрузка и максимальный ритм работы
Е. Минимальная нагрузка и максимальный ритм работы.
7. Повышение содержания ионов кальция в саркоплазме мышцы приводит к ее сокращению. Укажите возможную причину этого.
А. Влияние кальция на саркоплазматический ретикулум
В. Активация кальциевого насоса
С. Блокада миозиновой АТФ-азы
D. Активация активных центров актина
Е. Изменение структуры молекулы тропомиозина.
8. При раздражении скелетной и гладкой мышцы с одной и той же частотой гладкая мышца отвечает тетаническим сокращением, а скелетная — одиночными сокращениями. Какими особенностями гладкой мышцы это обусловлено?
А. Рефрактерность гладкой мышцы больше
В. Лабильность гладкой мышцы больше
С. Хронаксия гладкой мышцы меньше
D. Длительность сокращения гладкой мышцы меньше
Е. В гладкой мышце сильнее развит саркоплазматический ретикулум.
9. В мышце фармакологическим методом заблокирована АТФ-аза, после чего она утратила свойство сократимости. Какая возможная причина этого?
А. Открытие кальциевых каналов ретикулума
В. Открытие калиевых каналов поверхностной мембраны
С. Остановка Nа-К- насоса поверхностной мембраны
D. Натриевая инактивация
Е. Активация кальциевого насоса ретикулума.
10. При фосфоглюконатном пути окисления глюкозы энергия акумулируется:
А. АТФ
В. АДФ
С. В креатинфосфате
D. НАДФ
Е. Ц-АМФ
Ответы: 1.С, 2.В, 3.D, 4.D, 5.A, 6.B, 7.D, 8.A, 9.C, 10.D.
ТЕСТОВЫЕ ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ по программе «Крок-1»:
1. После забега на длинную дистанцию у спортсмена возникла контрактура икроножных мышц (мышц нижних конечностей). Накопление какого продукта метаболизма, наиболее вероятно, вызвало это состояние?
А. Мочевины
В. Мочевой кислоты
С. Креатинина
D. Пировиноградной кислоты
Е. Молочной кислоты
2. После тренировки у штангиста возникла контрактура трёхглавой мышцы. Уменьшение концентрации в мышцах какого вещества, наиболее вероятно, вызвало это состояние?
А.АТФ
В. Пировиноградной кислоты
С. Молочной кислоты
D. Креатинина
Е. Глюкозы
3. В эксперименте к мышце взятой из мочеточника животного подвешивают груз. Мышца растягивается и остаётся в таком положении после снятия груза. Какое свойство мышечной ткани демонстрирует данный опыт?
А. Растяжимости
В. Эластичности
С. Пластичности
D. Автоматии
Е. Сократимости
4.Тетаническое сокращение скелетной мышцы возникает, если интервал между раздражающими импульсами …
А. Меньше, чем продолжительность одиночного сокращения
В. Больше в 2 раза, чем продолжительность одиночного сокращения
С. Больше в 5 раз, чем продолжительность одиночного сокращения
Д. Равно продолжительности одиночого сокращения
Е. Больше в 7 раз, чем подолжительность одиночного сокращения
5. Са2+ в миоплазме необходим для осуществления процесса…
А. Замыкания акто-миозиновых мостиков
В.Размыкания акто-миозиновых мостиков
С. Формирования головок миозина
Д. Формирования тропомиозина
Е. Распространения ПД
6. Серия одиночных сокращений скелетных мышц возникает, если интервал между раздражениями…
А.Меньше, чем продолжительность одиночного сокращения
В. Больше в 2 раза, чем продолжительность одиночного сокращения
С. Больше в 5 раз, чем продолжительность одиночного сокращения
Д. Равно продолжительности одиночого сокращения
Е. Больше в 7 раз, чем подолжительность одиночного сокращения
7. Роль Са2+ в сокращении скелетной мышце состоит в …
А. Конформационном изменении тропонина
В.Конформационном изменении актина
С.Обеспечении движения головки миозина
Д.Блокировании активного центра миозина
Е. Блокировании активного центра актина
8. При условии действия на скелетную мышцу яда, угнетающего синтез АТФ, нарушится процесс … в период мышечного сокращения
А. Скольжения актина и миозина
В. Открытия активных центров актина
С. Выхода Ca2+ из саркоплазматического ретикулума
Д. Закрытия активных центров актина
Е. Изменения конформации тропонина
9. Зубчатый тетанус мышц возникает, если следующее раздражение попадает на…
А. Фазу расслабления предыдущего сокращения
В. Фазу укорочения предыдущего сокращения
С. Латентный период предыдущего сокращения
Д. Период покоя
Е. Любую фазу предыдущего сокращения
10. При действии на скелетную мышцу яда, угнетающего синтез АТФ, серия последующих раздражений пиведет к тому, что концентрация Са2+ в саркоплазме …
А. Увеличится
В. Уменьшится
С. Не изменится
Д. Исчезнет
Е. Конформируется
Ответы: 1.E, 2.A, 3.C, 4.A, 5.A, 6.B, 7.A, 8.A, 9.A, 10.A.
Ситуационные задачи:
1. Длительность рефрактерности мышцы 10 мсек. Длительность фазы сокращения 200 мсек. Рассчитайте интервал частот при которых данная мышца будет сокращаться в режиме гладкого тетануса.
2. Схема какого процесса приведена ниже? Добавьте недостающие звенья: Раздражение клеточной мембраны — возникновение ПД — проведение его вдоль волокна по Т-системе —? — взаимодействие актина и миозина —? — активация Са-насоса —? — расслабление мышцы.
3. При каждом импульсе возбуждения в межфибриллярное пространство выходит 1 функциональная Са++ единица. Весь кальций возвращается в ретикулюм с постоянной скоростью 10 м/сек. При какой частоте раздражения будет происходить суммация.
4. Рефрактерный период мышцы равен 5 мсек. Лабильность мышцы в 4 раза менше теоретической. Нарисуйте миограммы следующих частотах раздражения; 10 Гц, 50 Гц, 100 Гц.
5. Площадь физиологического поперечного сечения мышцы 25 см2. Рассчитайте удельную силу мышцы, если она в состоянии поднять максимально 200 кг?
6. Рассчитайте с какой частотой надо раздражать скелетную мышцу для получения зубчатого и гладкого тетануса, если одиночное сокращение её продолжается 0,1с (100мсек).
7. Рассчитайте с какой частотой надо раздражать скелетную мышцу для получения зубчатого и гладкого тетануса, если одиночные сокращения её продолжаются 0,05с (50мсек).
8. Рассчитайте сколько израсходует икроножная мышца лягушки АТФ за 5 секунд зубчатого и гладкого тетануса при условиях: А. одиночное сокращение длится 100мс; б) зубчатый тетанус – при частоте раздражения 15 в 1”; В. гладкий тетанус – при частоте 30 в 1”; г) на одно мышечное сокращение расходуется 0,3 мкМоль АТФ на 1,0г массы мышцы; D. масса икроножной мышцы лягушки 12,0г; Е. содержание АТФ на 1,0г мышечной массы = 3 мкМоль.
ответы к Ситуационным задачам:
1. Для гладкого тетанического сокращения необходимо, чтобы интервал между раздражениями был длиннее рефрактерного периода, но короче всей длительности сокращения. В данном случае этот интервал лежит в пределах от 10 до 70 мсек, значит при частоте от 15 до 100 Гц будет наблюдаться тетанус. При меньшей частоте будут одиночные сокращения, при большей — пессимум.
2. Приведена схема электромеханического сопряжения: Раздражение клеточной мембраны – возникновение ПД – проведение его вдоль волокна по Т-системе –освобождение кальция из саркоплазматического ретикулюма – взаимодействие актина и миозина – сокращение мышечного волокна – активация Са-насоса – возвращение кальция в цистерны – расслабление мышцы.
3. Если весь Са++ возвращается в ретикулюм за 100 мсек, значит, суммация сокращения и зубчатый тетанус будут возникать при частоте больше 10 Гц. При частоте раздражения 50 Гц перерыв между импульсами в 5 раз короче, и за это время в ретикулюм вернется уже не 1 функциональная единица Са++, а только 1/5 единицы. 4/5 же остаются в межфибриллярном пространстве и накапливаются там. Поскольку максимальная концентрация Са+ (5 х 10 мэкв/л) в 10 раз больше критической (0,5 х 106 мэкв/л), то такое количество Са++ накопится в пространстве через 10:4/5 = 12,5 импульсов. Это значит, что в ответ на 13-й импульс мышца даст максимальную высоту сокращения.
4. В данном случае теоретически ткань могла бы воспроизводить 1000:5 = 200 импульсов. В условии сказано, что истинна лабильность в 4 раза меньше, т. е. равна 50 Гц. Значит, при частоте раздражения 10 Гц мышца будет отвечать одиночными сокращ-ниями или зубчатым тетанусом, при 50 Гц — гладким, а при частоте более 50 Гц возникнет пессимум частоты.
5. Удельная сила мышцы равна отношению максимального груза к площади физиологического поперечного сечения. В данном случае она равна 8 кг/см2. По-видимому, это двуглавая мышца плеча человека.
6. Для получения зубчатого тетануса указанной мышцы нужна частота 11-19 в 1”, т.к. при частоте 10 в 1“ получим 10 одиночных сокращений. При этом каждое следующее раздражение падает на мышцу сразу после её расслабления – интервал между раздражениями 100 мс. При частоте 20 в 1“ получим гладкий тетанус, т.к. каждое раздражение будет заставать мышцу еще в состоянии сокращения, интервал между раздражениями 50 мс. Для получения гладкого тетануса частота раздражений должна быть 20 и более в 1“.
7. Для получения зубчатого тетануса указанной мышцы частота раздражений должна быть 21-39 в 1 “. Для получения гладкого тетануса – 40 и более в 1 сек.
8. В икроножной мышце лягушки массой 12г содержится 36 мкмоль АТФ. 1,0 г – 3 мкМоль АТФ 3 мкМоль х 12 = 36 мкМоль АТФ. При зубчатом тетанусе, вызванном частотой 15 в сек расходуется АТФ 4,5 мкмоль в 1 “ : 0,3 мкМоль АТФ х 15 = 4,5 мкМоль АТФ в 1 сек. Т.к. сокращение длится 5 сек, то 4,5мкМоль АТФ в 1 “ х 5 = 22,5 мкМоль АТФ. При гладком тетанусе частотой 30 в сек расходуется АТФ 9 мкмоль в 1 сек. Расчет: 0,3 мкМоль АТФ х 30 = 9,0 мкМоль АТФ, за 5 сек гладкого тетануса мышца израсходует 45 мкМоль АТФ.
infopedia.su
Лекция 4. Физиология мышечного сокращения
4-1. Скелетная мышца: строение мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы, физиологические свойства скелетной мышцы и ее функции.
Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц – мышечных волокон или мышечных клеток. Они имеют цилиндрическую форму и расположены параллельно друг другу. Волокна состоят из большого количества миофибрилл, создающих характерную поперечно-полосатую исчерченность. Миофибрилла включает белковые нити двух типов – актиновые (тонкие) и миозиновые (толстые). Между миофибриллами находится множество митохондрий. Цитоплазма волокна или саркоплазма содержит сеть внутренних полостей, окруженных мембранами – саркоплазматический ретикулум (СПР). Отдельная замкнутая везикула СПР часто называется цистерной.
Поперек волокна между миофибриллами проходит система трубочек, представляющих впячивания сарколеммы, называемые Т-системой. В определенных местах трубочки располагаются между двумя цистернами. Комплекс, состоящий из одной Т-трубочки и двух, тесно примыкающих к ней цистерн, представляет триаду. Трубочки и цистерны соединены между собой поперечными мембранными мостиками. Саркоплазматический ретикулум, являясь хранилищем Са2+, выполняет функцию регулятора уровня свободных катионов Са2+ в саркоплазме.
В световой микроскоп видна только поперечная исчерченность миофибрилл, включающая светлый диск I, темный диск А и Z–линию, разделяющую зону I. Участок между двумя полосками Z называется саркомером. При гораздо большем увеличении, которое дает электронный микроскоп, видно, что такая картина полосатости обусловлена определенным расположением нитей актина и миозина.
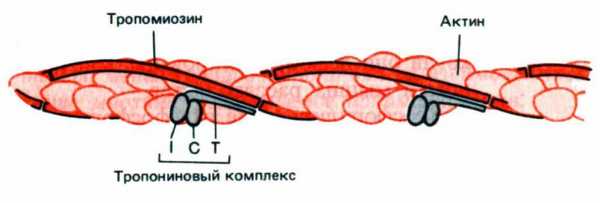
Рис. 6. Схематическое изображение тонкой актиновой нити , показывающее локализацию тропомиозина и тропонина на его поверхности. I, C, T — белок тропонин, состоящий из трех полипептидных цепей (тропонины I, C, T)
Молекула миозина состоит из длинного участка – хвоста и глобулярной части из двух головок. Молекулы миозина так расположены в миозиновой нити, что головки равномерно распределены по ее длине, кроме небольшого участка в середине, где они отсутствуют. Для механизма сокращения принципиально, что в местах перекрытия нитей актина и головок миозина они могут взаимодействовать между собой, вызывая укорочение мышцы. Энергия для совершения такой работы черпается из АТФ, поэтому неудивительно, что миозиновые головки обладают АТФ-азной активностью.
Тропонин участвует в Са2+ — зависимой регуляции сокращения. Тропонин Т имеет участок для связывания тропомиозина и ответственен за прикрепление всего комплекса к актиновому филаменту. Если к тропонину Т и тропомиозину добавить тропонин I, образуется комплекс, препятствующий взаимодействию актина с миозином даже в присутстви катионов Са2+. Дополненный тропонином С, этот комплекс продолжает блокировать актин-миозиновое взаимодействие, но только в отсутствии Са2+ ; при наличии в среде свободных катионов кальция тропонин С связывает четыре катиона Са2+ и таком состоянии снимает блокаду. именно этот эффект и лежит в основе индукции мышечного сокращения катионами кальция.
Актиновые филаменты имеют сложный состав и структуру. Каждая актиновая нить состоит из двух спиральных тяжей, каждый из которых образован молекулами глобулярного G-актина. Комплекс из двух нитей называется F-актином. Кроме того, в актиновом филаменте содержатся вспомогательные регуляторные белки тропомиозин и тропонин.
Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные поперечно-полосатые мышцы подразделяют на тонические и фазные волокна. Тонические волокна имеют множественную иннервацию, т.е. на одном волокне по всей его длине располагаются несколько синапсов. Каждый импульс приводит к освобождению только небольшого количества ацетилхолина, поэтому степень деполяризации зависит от частоты стимуляции, а механический ответ носит градуальный характер. Тонические волокна участвуют в сохранении позы.
Фазные волокна, наоборот, сокращаются по принципу «все или ничего». Как только нервный импульс достигнет надпороговой силы, происходит генерация распространяющегося потенциала действия, максимальное освобождение Са2+ из везикул и максимальное (одиночное) по силе сокращение, которое уже не возрастает при увеличении интенсивности раздражения.
Вместе с тем, при электрическом раздражении целой мышцы (при наложении на нее электродов) сила сокращения мышцы зависит от интенсивности стимула. Стимул, едва превышающий порог, вызовет ответ по принципу «все или ничего» только в ближайших волокнах, где плотность тока максимальна. При этом при небольшой силе раздражения возбуждаются более возбудимые волокна. Для возбуждения всех волокон требуется много большая интенсивность стимула. Причина такого положения в том, что каждое волокно электрически изолировано друг от друга.
Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна I типа и «быстрые» – волокна II типа. Различие между быстрыми и медленными волокнами легко заметно даже гистологически: как правило, быстрые волокна содержат мало митохондрий и очень мало миоглобина, поэтому окрашены слабо. В силу этого их часто называют «белыми». Поскольку мощность ресинтеза АТФ в них невелика (преобладает анаэробный гликолиз), то они быстро утомляются, но способны развивать большое напряжение.
Быстрые волокна подразделяются как минимум также на две группы. Одна группа – это быстрые гликолитические волокна II B типа, вторая – быстрые окислительные волокна II A типа, способные использовать как окислительное фосфорилирование, так и гликолиз. Волокна типа II А присутствуют у некоторых млекопитающих, но у человека имеются в небольших количествах. Медленные волокна содержат много митохондрий и миоглобина, имеют больше капилляров, вследствие этого имеют выраженное красноватое окрашивание и называются «красными». В силу большой мощности аэробного ресинтеза АТФ в окислительном фосфорилировании утомляются мало, но развивают относительно небольшое напряжение.
У млекопитающих и некоторых других видов часть мышц представлена почти исключительно волокнами одного типа. Например, длинный разгибатель пальца включает только быстрые волокна, а камбаловидная мышца – только медленные. У человека же мышцы, включая отмеченные, преимущественно смешанные. Но все же в длинном разгибателе пальца доминируют быстрые, а в камбаловидной мышце медленные волокна.
4-2. Современная теория мышечного сокращения и расслабления. Электромеханическое сопряжение.
Механизм мышечного сокращения. В присутствии АТФ, но при низкой концентрации Са2+, составляющей 10–8 моль/л и меньше, волокно находится в расслабленном состоянии, т.е. головки миозина не образуют связи с актином. В момент увеличения концентрации Са2+ вблизи головок до 10–6-10–5 моль/л ферментативная активность головок многократно увеличивается и в той же мере возрастает гидролиз АТФ. В скелетных мышцах тяжи тропомиозина закрывают активные участки актинового филамента, препятствуя взаимодействию миозиновых головок с мономерами актина, тем самым предотвращая сокращение. Повышение концентрации Са2+ сопровождается его связыванием с тропонином. При этом молекулы комплекса тропонин-тропомиозин изменяют свое расположение таким образом, что обнажают активные участки на актиновом филаменте.
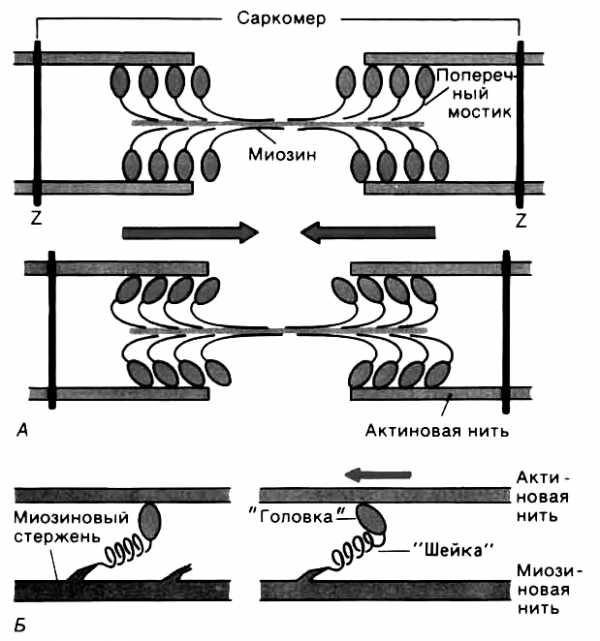
Рис. 7. Функция поперечных мостиков. А. Модель механизма сокращения: миозиновая нить с поперечными мостиками, прикрепленными к соседним актиновым нитям; вверху — до, внизу — после «гребка» мостиков (на самом деле они функционируют асинхронно). Б. Модель генерирования силы поперечными мостиками; слева — до, справа — после «гребка». Поперечные мостики химически соответствуют субфрагменту миозина — «тяжелому меромиозину», который состоит из субфрагментов I (головка) и II (ножка).
За счет освобождающейся энергии происходит связывание головок с нитью актина, изгибание «ножки» и вследствие этого актиновые филаменты продольно перемещаются относительно центрального миозинового стержня. Уменьшение же концентрации Са2+ до прежнего минимального уровня вызывает размыкание поперечных мостиков и возвращение
Описанное взаимодействие глобулярной головки миозина и активным участком молекулы актина называют циклом поперечного мостика. Один цикл поперечного мостика происходит в следующей последовательности:
активация миозиновой головки;
обнажение активного участка молекулы актина в присутствии кальция;
самопроизвольное формирование поперечного мостика;
поворот глобулярной головки, сопровождающийся продвижением актиновой нити и укорочением саркомера;
разрыв поперечного мостика.
Цикл после завершения может повториться или остановиться. Если бы все поперечные мостики в отдельной мышце прошли одновременно только один цикл, мышца укоротилась бы всего на 1% от ее длины в состоянии покоя. Однако многие мышцы способны укорачиваться до 60% длины покоя. Это показывает, что цикл поперечного мостика происходит в них многократно. При этом каждый раз миозин захватывает и тянет новый участок актина. Именно такая многократность повторения цикла замыкания-размыкания мостика вызывает более выраженное укорочение и генерирует бόльшую силу.
В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются к ее центру. Прочная соединительнотканная фасциальная оболочка, прикрепляющая мышцу посредством сухожилия к костям, передает вызываемую сокращением силу костям. Линейно направленная мышечная сила тянет кость и вызывает ее вращение в плоскости, перпендикулярной оси сустава. Поскольку мышца обычно проходит, по крайней мере, через один сустав, более свободный сегмент костного скелета будет перемещаться, вызывая вращательное движение в суставе.
Электромеханическое сопряжение. Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала о начале сокращения от возбужденной сарколеммы, называется электромеханическим сопряжением.
Электромеханическое сопряжение происходит посредством распространения возбуждения (потенциала действия) по системе поперечных трубочек внутрь волокна. При этом информация о возбуждении (фактически о перезарядке мембраны) очень быстро достигает продольной системы СПР, в конечном итоге вызывая открытие в мембране цистерн Са2+ -каналов и залповый выброс катиона, быстро диффундирующего к миофибриллам. Ключевое значение в процессе передачи этого сигнала имеет разница электрических потенциалов по разные стороны мембраны Т-трубочки. Предполагают, что развивающаяся при передаче потенциала действия по Т-трубочке деполяризация вызывает конформационные изменения белков ножки концевой цистерны. Эти конформационные изменения приводят к открытию Са2+ -каналов в СПР и обеспечивают выход ионов Са2+ в цитоплазму по электрохимическому градиенту.
Остановка сокращения. Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного импульса в двигательной концевой пластинке нервно-мышечного соединения. Прекращение импульса означает устранение сигнала к высвобождению Са2+, которое прекращается. Однако действие остающегося в саркоплазме Са2+ может все еще продолжаться. Поэтому при одиночном нервном импульсе и соответственно одиночном сокращении генерация силы (укорочение) саркомером только останавливается. Чтобы сила, генерируемая саркомером, уменьшилась до величины в состоянии покоя, и произошло расслабление саркомера, необходимо многократное уменьшение Са2+ вблизи миозиновых головок. Это происходит вследствие откачки Са2+, усиливающейся по мере его выхода в саркоплазму, поскольку катионы Са2+ все сильнее начинают активировать Са2+ -насос (Са2+ -АТФазу) мембран саркоплазматического ретикулума. В результате «накачивания» Са2+ в цистерны СПР его концентрация вблизи головок резко уменьшается и поперечные мостики размыкаются – мышечное волокно расслабляется.
Циклическое образование поперечных мостиков продолжается до тех пор, пока в волокне имеются в достаточном количестве свободных катионов кальция и АТФ. Все, что снижает концентрацию Са2+ и АТФ, будет ослаблять мышечное сокращение, что происходит при утомлении мышцы. Таковыми факторами, например, является низкая концентрация калия, яды и токсины, нарушающие выработку АТФ в окислительном фосфорилировании.
Полное отсутствие АТФ вызывает устойчивое сокращение, называемое трупным окоченением. Такое состояния устойчивого сокращения обусловлено двумя обстоятельствами. Во-первых, в отсутствие АТФ невозможна откачка Са2+ и саркоплазмы в везикулы СПР. Во-вторых, без АТФ не может произойти повторная активация миозиновой головки, требующаяся для разрыва поперечного мостика между актином и миозином. В итоге поперечные мостики остаются замкнутыми, а мышца в сокращенном состоянии.
4-3. Виды и режимы мышечного сокращения. Одиночное сокращение. понятие о тетанусе. Механизм длительного сокращения в естественных условиях.
Типы сокращения скелетных мышц. В ответ на раздражение мышца развивает напряжение и сокращается. Возможны два случая – сокращение с укорочением и без укорочения. Если мышца развивает напряжение без изменения длины, то развивается изометрическое сокращение. Такое сокращение возникает в двух случаях: когда оба конца мышцы жестко закреплены; когда закреплен один конец, но развиваемая мышцей сила недостаточна для передвижения нагрузки, прикрепленной к другому концу. В отсутствие укорочения работа в физическом смысле не совершается, поскольку отсутствует передвижение. Но в физиологическом отношении мышца работу совершает, так как в ней происходят изменения, характеризующие физиологическую работу: происходит гидролиз АТФ, повышается температура и совершается ряд других изменений.
Когда мышца укорачивается, преодолевая постоянную нагрузку, говорят об изотоническом сокращении. Большинство сокращений в организме представляют комбинацию изотонического и изометрического компонентов. Изометрическая фаза продолжается до тех пор, пока мышца не разовьет силу, достаточную для перемещения нагрузки. С этого момента начинается изотоническая фаза, и мышца сокращается с постоянной силой, перемещая нагрузку. С увеличением нагрузки скорость и степень укорочения мышцы во время изотонического сокращения уменьшается, а продолжительность изометрической фазы сокращения увеличивается.
Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50 мс, называемое латентным периодом, вызывает сокращение мышцы, и в ней развивается напряжение. Фаза сокращения длится примерно 100 мс. У быстрых фазных мышц она намного короче, чем у медленных. Например, время сокращения быстрых волокон наружной головки четырехглавой мышцы бедра молодых мужчин составляет 30 мс, а медленных – 80 мс. Вслед за сокращением наступает более медленная фаза расслабления, длящаяся до 200 мс.
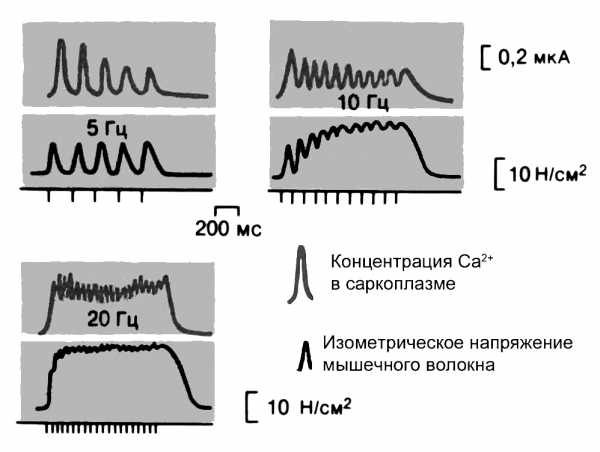
Рис. 8. Суммация и слияние одиночных сокращений до тетануса (зубчатого) при повышении частоты раздражения. Частота толчков тока 5, 10 и 20 Гц. Изометрическое напряжение калибровано в Н/см2 площади поперечного сечения мышцы.
Если следующий нервный импульс приходит к волокну раньше, чем закончилось расслабление предыдущего одиночного сокращения, происходит суперпозиция (наложение) механических ответов. При ритмической стимуляции мышца все время будет развивать силу, но она будет переменной, так как за межимпульсные интервалы будет происходить расслабление за счет депонирование Са2+ в СПР. Такая форма сокращения называется зубчатым тетанусом (рис. 8). На внутриволоконном уровне это означает, что выходящий из саркоплазматического ретикулума Са2+ только частично успевает вновь «закачаться» Са2+-насосом в цистерны. Явление последовательного увеличения амплитуды (силы) сокращения скелетной мышцы до величины плато во время частой стимуляции после периода покоя называется эффектом лестницы
При достаточно высокой частоте стимуляции выходящий из ретикулума Са2+ не успевает вернуться в цистерны, а мышца поэтому будет постоянно находиться в состоянии максимальной активации. Возникающий при этом вид сокращения называется гладким тетанусом. Существенно, что повышение частоты стимуляции сопровождается увеличением силы, развиваемой мышцей. При гладком тетанусе напряжение, развиваемое мышцей, обычно в 2-4 раза больше, чем при одиночном сокращении. В естественных условиях для скелетных мышц характерны режимы тетанических, а не одиночных сокращений.
Сила сокращения скелетных мышц. Двигательная единица. Под двигательной единицей понимают каждую группу мышечных волокон вместе с иннервирующим ее аксоном. Количество мышечных волокон, входящих в двигательную единицу, неодинаково в разных мышцах. Например, у человека двигательная единица передней большеберцовой мышцы содержит 500-600 волокон, двигательная единица внутренней головки икроножной мышцы – 1500-2000 волокон, мышцы кисти – 100-300, глазные мышцы от 13 до 20 волокон.
Тонкое управление мышечной активностью достигается регуляцией напряжения, развиваемого каждой отдельной мышцей. Такая регуляция осуществляется одним из двух способов (или обоими одновременно):
— изменением числа мышечных волокон, возбуждающихся в данный момент. Развиваемая мышцей сила будет тем больше, чем больше мышечных волокон (и, следовательно, двигательных единиц) будет стимулировано;
— изменением частоты нервных импульсов. Чем выше частота приходящих нервных импульсов, тем больше развиваемая сила.
Максимальная сила мышц. Абсолютная сила, развиваемая разными мышцами, неодинакова. Она определяется: 1) поперечным сечением мышцы, 2) ее длиной, 3) типом сокращения, 4) характером сокращения.
Для сравнения силы разных мышц используют понятие относительной максимальной силы. Ее измеряют при изометрическом тетанусе небольшой продолжительности (1,5-5 с) и выражают в килограммах (кг) или ньютонах (Н) на площадь поперечного сечения. Этот показатель определяется числом миофибрилл на площади поперечного сечения и количеством поперечных мостиков, т.е. исходной длиной саркомера, точнее толстого миозинового филамента. Максимальная сила у разных мышц разных животных достигает верхнего предела в 4-6 кг/см2, хотя имеются мышцы и со значительно меньшей силой. Очень большую силу развивает аддуктор устрицы – 12 кг/см2. В то же время сердечная мышцы крысы развивает силу всего до 0,016 кг/см2. В относительном масштабе максимальная сила целой мышцы ниже таковой отдельного мышечного волокна (на единицу площади сечения).
studfiles.net
2. Физиология мышечного сокращения (Задачи №№ 2-1 – 2- 2 из сd- приложения к Учебнику «Физиология м основами морфологии» под ред. Судакова к.В., и в.Ф. Волкова [ 3 ])
2-1. Препарат ЭДТА связывает ионы кальция. Как изменится процесс синаптической передачи, если ввести ЭДТА в область нервно-мышечного синапса скелетной мышцы?
2-2. Препарат гемихолиний угнетает обратный захват из синаптической щели в пресинаптическую область продукта гидролиза ацетилхолина (АХ) – холина Как изменится процесс синаптической передачи, если ввести гемохолиний в область нервно-мышечного синапса скелетной мышцы?
(Задачи №№ 2-3 – 2-10 из Сборника задач под ред. Г.И. Косицкого [ 1 ])
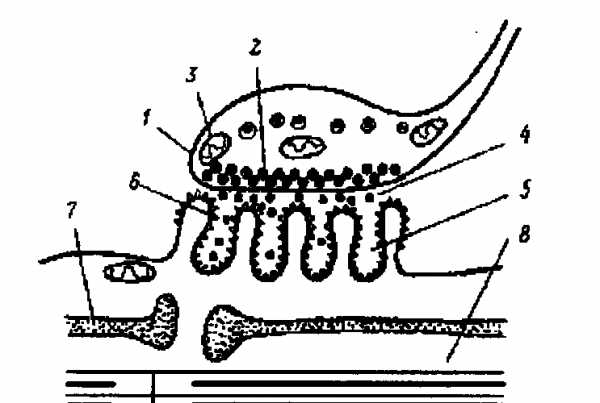
2-3. Какая структура изображена на рисунке? Назовите обозначенные цифрами элементы. |
| |
2-4. Правильно ли расставлены частоты раздражения на рисунке? |
| |
2-5. Какая из изображенных на схеме мышц обладает большей абсолютной силой? |
| |
2-6. Какие виды сокращения мышцы представлены на рисунке? Какими раздражениями они вызваны? |
| |
2-7. Потенциалы волокон каких мышц зарегистрированы на рисунках А, Б, В? ПД мышечных волокон различных мышц. |
| |
2-8. Какие выводы Вы можете сделать, рассмотрев рисунок? Сокращение (а) икроножной мышцы лягушки при раздражении седалищного нерва;. ПД(б). |
| |
2-9. Мышца какого нервно-мышечного препарата не будет отвечать на раздражение нерва и почему? | В | |
2-10. Назовите структурные единицы миофибриллы, обозначенные цифрами. Какой из дисков уменьшается при сокращении мышцы? |
| |

 раствор яда кураре погружена
мышца нервно-мышечногопрепарата А и
нервпрепарата
Б
раствор яда кураре погружена
мышца нервно-мышечногопрепарата А и
нервпрепарата
Б
2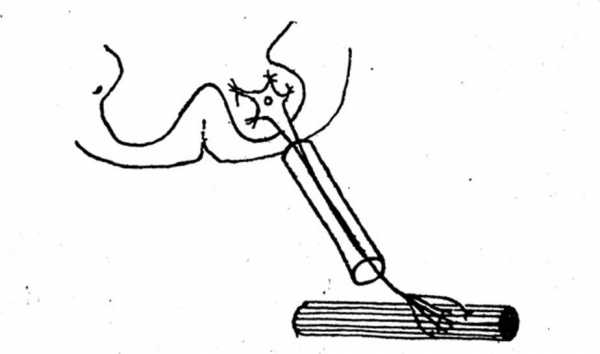 -11.
Схематическое изображение какой
структурной единицы представлено
на рисунке? Назовите элементы,
входящие в состав этой структурной
единицы.
-11.
Схематическое изображение какой
структурной единицы представлено
на рисунке? Назовите элементы,
входящие в состав этой структурной
единицы.
2-12. При перерезке двигательного нерва мышца, которую он иннервировал, атрофируется. Чем можно это объяснить?
2-13. Мышцу нервно-мышечного препарата подвергают непрямому раздражению. Через некоторое время амплитуда сокращений начинает уменьшаться. Означает ли это, что в мышце наступило утомление? Как проверить это предположение?
2-14. К покоящейся мышце подвесили груз. Как при этом изменится ширина Н-зоны саркомера?
2-15. Почему быстрые мышцы при сокращении потребляют в единицу времени больше энергии АТФ, чем медленные?
2-16. Нарисуйте схему соотношения кривых потенциала действия, возбудимости и одиночного сокращения скелетной мышцы.
2-17. Какие процессы протекают в мышце во время латентного периода при непрямом раздражении?
2-18. Проверьте, правильно ли расставлены частоты раздра-жения на миограммах, приведенных ниже: | 5 гц 80 гц 60 гц 20 гц |
2-19. Какая из представленных кривых утомления получена при раздражении нормальной мышцы? В каком случае может быть получена вторая кривая? Почему? | 1 2 |
2-20. Длительность рефрактерности мышцы 10 мсек. Длительность одиночного сокращения 200 мсек. Назовите интервал частот раздражения, при которых данная мышца будет сокращаться в режиме гладкого тетануса.
2-21. Скорость проведения возбуждения по мембране мышечного волокна равна 5 мсек. Чему равна скорость проведения волны сокращения? Через какое время волна сокращения дойдет от одного конца мышцы до другого, если длина мышечного волокна 10 см.?
2-22. Длительность потенциала действия мышцы 10 мсек. Какую частоту раздражения следует дать, чтобы каждый импульс попадал в фазу супернормальной возбудимости?
2-23. Схема какого процесса приведена ниже? Добавьте недостающие звенья.
Раздражение — возникновение ПД — проведение его вглубь волокна — — ? — взаимодействие актина и миозина — ? — активация Са-насоса — — ? — расслабление мышцы.
2-24. При раздражении нерва нервно-мышечного препарата мышца доведена до утомления. Что произойдет, если в это время подключить раздражение мышцы?
2-25. Мышца сокращается тетанически. Как изменится ритм ее сокращения, если в перфузируемый раствор ввести атропин?
2-26. При каждом импульсе возбуждения в межфибриллярное пространство выходит 1 функциональная условная единица Са++. Весь Са++ возвращается в ретикулюм с постоянной скоростью за 10 мсек. При какой частоте раздражения будет происходить суммация сокращений? Через сколько циклов при частоте 50 герц амплитуда сокращения достигнет максимума, если одна функциональная единица Са++ соответствует критической концентрации Са++?
2-27. Рефрактерный период мышцы равен 5 мсек. Лабильность мышцы в 4 раза меньше теоретической. Нарисуйте миограммы при следующих частотах раздражения: 10 герц, 50 герц, 100 герц.
2-28. В эксперименте на нервно-мышечном препарате было определено, что при неизменной силе тока пессимум наступает при частоте 150 герц. При какой частоте раздражения можно получить на этом препарате состояние оптимума? Какова лабильность нервно-мышечных синапсов данного препарата?
2-29. Площадь физиологического поперечного сечения мышцы 25 кв. см. Рассчитайте удельную силу мышцы, если она в состоянии поднять максимально 200 кг?
studfiles.net
ФИЗИОЛОГИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ
884.Частота сердечных сокращений у плода 140-160в минуту.
885.Частота сердечных сокращений уноворожденного 120-140в минуту.
886.Частота сердечных сокращений уребенка 1 года110-120с минуту.
887.Тахикардия — это учащение сердечных сокращений.
888.Брадикардияэто урежение частоты сердечных сокращений.
889.Синокардиальный узел рассоложен между устьем верхней полой вены и ушком правого предсердия.
890.Пейсмекер сердцаэто группа атипических клеток миокарда, задающих ритм сердца.
891.Градиент автоматииэто убывание степени автоматии участков проводящей системы по мере удаления от синоатриального узла.
892.Водителем ритма первого порядка является, синоатриальныйузел.
893.Водителем ритма второго порядка является атриовентрикулярный узел.
894.Водителем ритма третьего порядка является пучок Гиса.
895.Не обладает автоматией нижняя треть желудочков сердца.
896.Возбуждение по миокардураспространяется со скоростью 0,9-1,0м/’сек.
897.По пучкуГиса возбуждение распространяется со скоростью4-5м/сек.
898.В атриовентрикулярном узле возбуждение распространяется со скоростью 0,02-0,05м/сек.
899.Задержка проведения возбуждения в атриовентрикулярном узле имеет физиологическое значение координациисокращений предсердий и желудочков.
900.Абсолютная рефрактерность сердечной мышцыэто время, в течение которого сердечная мышца не отвечает ни на какие раздражители.
901.Относительная рефрактерность сердечной мышцы -этовремя, в течение которого сердечная мышца отвечает только на сверхпороговые раздражители.
902.Длительность периода абсолютной рефрактерности миокарда- 0,27 сек.
903.Длительность периода относительной рефрактерности миокарда- 0,03 сек.
904.Закон Франка-Старлингахарактеризует сократимость миокарда.
905.Закон Франка-Старлингазаключается в том, что увеличение растяжения сердца во время диастолы приводит к усилению его сокращенияво время систолы.
906.При раздражении блуждающего нерва возбудимость миокарда понижается.
907.При раздражении симпатического нерва возбудимость миокарда повышается.
908.При раздражении блуждающего нерва проводимость миокарда понижается.
909.При раздражении симпатического нерва проводимость миокарда повышается.
910.При раздражении блуждающего нерва сократимость миокарда понижается.
911.При раздражении симпатического нерва сократимость миокарда повышается.
912.При раздражении блуждающего нерва частота сокращений миокарда понижается.
913.При раздражении симпатического нерва частота сокращений миокарда повышается.
914.Звуковые характеристики первого тона сердца: низкий, протяжный, глухой.
915.Звуковые характеристики второго тона сердца:высокий, звонкий, короткий.
916.Компоненты первого тона сердца: вибрация при сокращении миокарда, закрытие атриовентрикулярных клапанов, открытие полулунных клапанов, сосудистый шум.
917.Компонентывтороготонасердца:вибрацияприрасслаблениимиокарда,закрытие полулунных клапанов, открытие атриовентрикулярных клапанов, сосудистый шум.
918.Причина возникновения третьего тона сердца: наполнение желудочков кровью во время их диастолы.
919.Причина возникновения четвертого тона сердца:систола предсердий.
920.Функцией сердца является насосная функция.
921.Насосная функция сердца осуществляется за счет периодического повторения систолы и диастолы.
922.Каждый отдельный цикл сердечной деятельности начинается с систолы предсердий.
923.С окончанием систолы предсердий начинается систола желудочков.
924.За окончанием систолы желудочков следует диастола желудочков.
925.Односторонний ток крови по направлению “предсердияжелудочкиаорта” обеспечивается наличием клапанного аппарата сердца.
studfiles.net