
Трофические цепи и сети: Пищевая сеть трофическая
Трофические цепи: примеры и описание
Пища — это то, без чего не может жить и развиваться ни один живой организм. Растения — это такие живые организмы, которые могут самостоятельно создавать для себя еду практически из ничего. Другие же питаются тем, что было создано без их участия.
Что такое трофическая цепь?
Биосфера огромна и многогранна. Она включает в себя самые разные создания. И прекрасные, и ужасные. Но все эти существа связаны друг с другом. Порой эти связи незаметны, а порой от одного вида зависит существование другого. Самой явной связью между живыми существами является пищевая. Пищевые цепи также называются трофическими.
Растения — это существа, которые способны к фотосинтезу. Это значит, что если будет углекислый газ, солнце и вода, то они смогут выжить. Животные на такое не способны. Потому подавляющая часть трофических цепей начинается с растений.
Трофические цепи бывают разные. Их разделяют на две группы. Те трофические цепи, которые начинаются с продуцента, называются пастбищными. Но есть и те, которые начинаются с отходов. Например, с трупов животных. Такие цепи называются детритными.
Но есть и те, которые начинаются с отходов. Например, с трупов животных. Такие цепи называются детритными.
Среди живых организмов выделяют продуценты, консументы и редуценты.
Продуценты
Многие трофические цепи начинаются с продуцентов. Именно так называют все те организмы, которые самостоятельно производят для себя питательные вещества. Самыми известными продуцентами являются зеленые растения, которые способны к фотосинтезу. Но также к ним можно отнести и бактерии-хемотрофы, которые могут обойтись даже без участия солнечного света.
Консументы
Но не все участники трофических цепей могут обходиться минимумом. Большинству нужно для жизни то, что было произведено другими. Такие потребители называются консументами. Но и среди них можно выделить разные по типу питания группы.
Некоторые животные питаются растениями. Они также попадают в трофические цепи. Трофический уровень их ненамногим выше растения. В свою очередь они тоже станут чьим-то ужином.
Те, кому для жизни нужно мясо, называются хищниками. Это новый уровень. Они вынуждены выходить на охоту для того, чтобы найти себе пропитание. Для того чтобы получить пищу, они должны применить все свои уловки, чтобы выследить и поймать жертву. У каждого хищника есть целый список предпочтений. Это помогает разным видам выживать даже в самые трудные времена. Особенно это важно сейчас, потому что многие виды находятся на грани вымирания.
Это новый уровень. Они вынуждены выходить на охоту для того, чтобы найти себе пропитание. Для того чтобы получить пищу, они должны применить все свои уловки, чтобы выследить и поймать жертву. У каждого хищника есть целый список предпочтений. Это помогает разным видам выживать даже в самые трудные времена. Особенно это важно сейчас, потому что многие виды находятся на грани вымирания.
Редуценты
Трофические цепи включают в себя и такие необычные организмы, как редуценты. Это те живые существа, которых называют «могильщиками природы». Они помогают отходам и трупам разложиться на более простые компоненты. Редуценты питаются тем, что больше никому не интересно.
Труп или отход, разложиться которому помогает редуцент, называется детритом. Потому пищевая цепь, которая начинается столь необычно, является детритной.
Существует большое количество разных редуцентов. Среди них выделяют грибы, бактерии, сапрофаги, некрофаги и копрофаги.
Сапрофаги — это те редуценты, которые питаются трупами других живых существ. Некрофаги также питаются трупами и падалью. Копрофаги питаются органическими отходами живых существ. Все эти организмы помогают сохранить чистоту в природе, дать место для жизни и роста новым живым организмам. Несмотря на то что некоторые считают этих живых существ отвратительными, без них невозможно представить здоровую биосферу.
Некрофаги также питаются трупами и падалью. Копрофаги питаются органическими отходами живых существ. Все эти организмы помогают сохранить чистоту в природе, дать место для жизни и роста новым живым организмам. Несмотря на то что некоторые считают этих живых существ отвратительными, без них невозможно представить здоровую биосферу.
Примеры трофических цепей
Непросто изучить трофические цепи. Примеры помогут понять, кто и кем питается. Самой привычной является пастбищная трофическая цепь. Она начинается с растения. Итак, можно начать со злаков. Они входят в список пристрастий зайцев. Зайцы — это консументы первого порядка. Зайцами питаются волки. Волки — это консументы второго порядка.
Мыши также любят злаки. Мыши в этой трофической цепи становятся консументами первого порядка. Порой мыши становятся жертвами ежей. Ежи — консументы второго порядка. Они также не могут быть спокойны за свою жизнь, ведь на них ведут охоту лисы. Последние станут консументами третьего порядка.
Трофические цепи можно обнаружить и в воде. Начать можно с фитопланктона, который способен самостоятельно прокормить себя. Но им питается зоопланктон. Зоопланктон — пища небольших рыбешек. А они же, в свою очередь, становятся любимым лакомством щуки. Но она тоже может стать чьей-то пищей.
Самые длинные трофические цепи можно найти в океане. В таких цепях встречаются даже консументы пятого и шестого порядка. Каждый может провести небольшое исследование. Это очень интересно и позволяет лучше понять окружающий мир.
Детритные трофические цепи не включают в себя так много участников. Обычно в их составе — только отход или труп и существо, которое им питается. Самыми известными редуцентами являются бактерии и грибы.
Трофические сети
Трофические цепи не способны полностью отразить всю жизнь биосферы. Ведь если допустить, что вымерли все зайцы, волки и лисы не погибнут. Они просто будут питаться другими животными, хотя и численность этих хищников сократится. Такой природный баланс сохраняется благодаря трофическим сетям.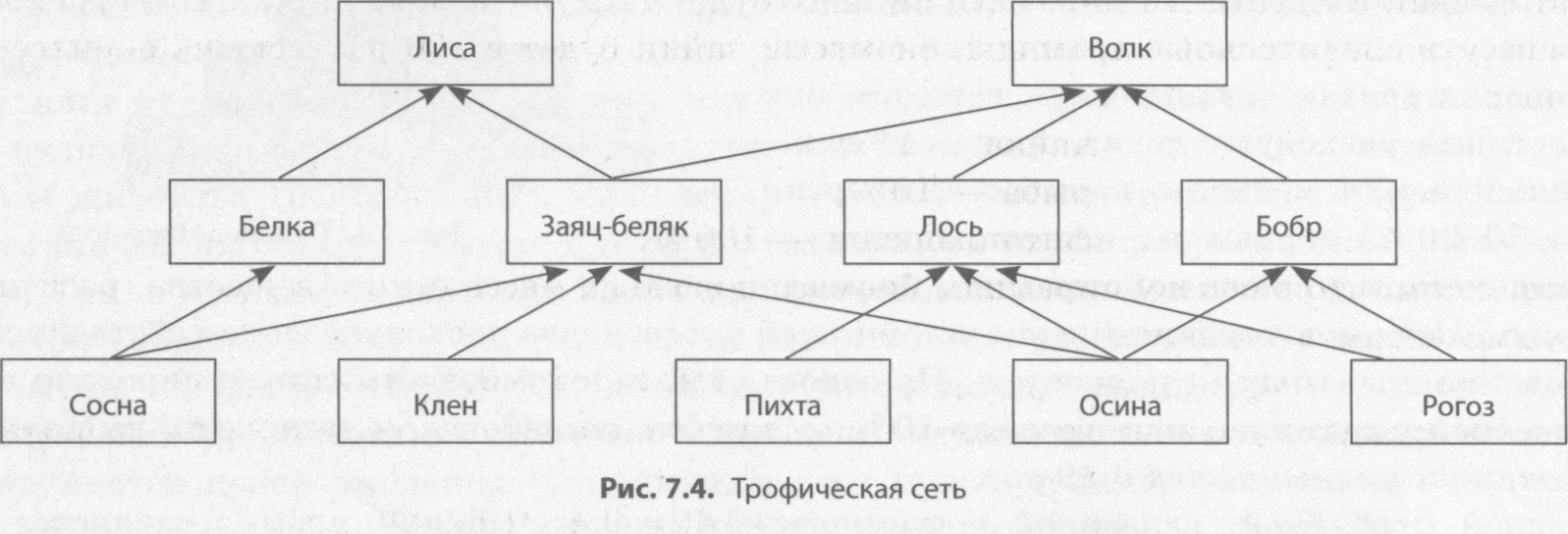
Один и тот же заяц может питаться разными растениями. Он может есть злаки, овес, лишайник, клевер и многое другое. Волк также может есть разных животных, которые отличаются друг от друга даже по размерам. Трофические цепи и сети помогают природе установить такой баланс, чтобы живые существа смогли пережить тот случай, если какой-то вид исчезнет с лица Земли.
Трофические сети порой очень и очень запутаны. Непросто разобраться, как много животных желают полакомиться каким-то растением и от каких особей прячется какой-либо небольшой зверек. Можно составить большое количество различных пищевых сетей. Особенно они богаты там, где бок о бок соседствует большое количество живых существ.
Мир, окружающий человека, хранит много загадок и интересных моментов. Одним из них являются трофические цепи и сети. Тщательно продуманные природой цепи питания помогают самым разным растениям и животным сосуществовать в относительном мире друг с другом, находить пищу для себя и своих детенышей, а также очищать свою среду обитания от того, что отжило свой срок.
Пищевые цепи и сети экосистем
Непрерывный поток солнечной энергии воспринимается молекулами живых клеток растений и в результате фотосинтеза преобразуется в энергию химических связей создаваемого органического вещества. В отличие от растений животные не способны к реакциям фото- или хемосинтеза, поэтому вынуждены использовать солнечную и очень незначительно химическую энергию опосредовано – питаясь органическим веществом, созданным растениями. Животные-консументы 1-го уровня питаются растительной пищей и одновременно служат пищей животных-консументов 2-го уровня и т. д. Таким образом в экосистемах образуется цепь последовательной передачи вещества и эквивалентной ему энергии от одних организмов к другим, которая называется трофической или пищевой.
При каждом очередном переносе большая часть (более 90 %) потенциальной энергии пищи теряется, переходя в тепло. Данное обстоятельство ограничивает возможное число звеньев пищевой цепи до 4 – 5. Чем короче пищевая цепь или чем ближе к её началу место, занимаемое данным организмом, тем больше доступной пищи и энергии.
Пищевые цепи можно разделить на два основных типа: пастбищные цепи, которые начинаются с зелёного растения и идут дальше к пасущимся фитофагам, а затем к хищникам; детритные цепи, которые от мёртвого органического вещества идут к детритофагам, а затем к хищникам.
Пастбищные цепи, в свою очередь, объединяют пищевые цепи хищников и пищевые цепи паразитов.
Пищевые цепи хищников идут от продуцентов к травоядным, поедаемым мелкими плотоядными, а они служат пищей более крупным хищникам и т.д. По мере продвижения по цепи хищников животные увеличиваются в размерах и уменьшаются в числе.
Трофические цепи бывают простыми и короткими, например, «осина – заяц – лиса», и более сложными и длинными, например, «сосна — тли — божьи коровки — пауки – насекомоядные птицы — хищные птицы».
Удлинение цепи происходит благодаря участию в ней хищников.
Пищевые цепи паразитов, наоборот, ведут к организмам, которые всё более уменьшаются в размерах и увеличиваются численно. Примером может служить цепь: «трава — травоядные млекопитающие — блохи – жгутиконосцы».
Примером может служить цепь: «трава — травоядные млекопитающие — блохи – жгутиконосцы».
В детритных цепях консументами являются детритофаги, относящиеся к различным систематическим группам: мелкие животные, преимущественно беспозвоночные, которые живут в почве и питаются опавшей листвой, или бактерии и грибы, разлагающие органические вещества по схеме: «детрит — бактерии, грибы» или «детрит – мелкие животные – хищники».
В большинстве случаев деятельность обеих групп детритофагов характеризуется строгой согласованностью: животные создают условия для работы микроорганизмов, разделяя трупы животных и мёртвые растения на мелкие части.
Пищевые цепи, начинающиеся с зеленых растений и от мёртвого органического вещества, чаще всего представлены в экосистемах совместно, но почти всегда одна из них доминирует над другой. Тем не менее в некоторых специфических средах (например, подземной), где существование организмов с хлорофиллом невозможно из-за отсутстыия света, сохраняются пищевые цепи только детритного типа. Пищевые цепи не изолированы одна от другой, а тесно переплетены. Они составляют так называемые пищевые сети. Принцип образования пищевых сетей состоит в следующем. Каждый продуцент имеет не одного, а несколько консументов. В свою очередь, консументы, среди которых преобладают полифаги, пользуются не одним, а несколькими источниками питания.
Пищевые цепи не изолированы одна от другой, а тесно переплетены. Они составляют так называемые пищевые сети. Принцип образования пищевых сетей состоит в следующем. Каждый продуцент имеет не одного, а несколько консументов. В свою очередь, консументы, среди которых преобладают полифаги, пользуются не одним, а несколькими источниками питания.
Необходимо подчеркнуть, что трофическая классификация делит на группы не сами виды, а типы их жизнедеятельности. Популяция одного вида может занимать один или более трофических уровней, в зависимости от того, какие источники энергии эти виды используют. Точно так же любой трофический уровень представлен не одним, а несколькими видами, в результате чего цепи питания сложно переплетены.
В процессе жизнедеятельности всех уровней трофической цепи возникают обходы. Зелёные растения частично или полностью сбрасывают листву, значительная часть организмов постоянно отмирает. В конечном счёте, созданное органическое вещество частично или полностью заменяется. Эта замена происходит благодаря особому звену трофической цепи – звену редуцентов. Это звено составляют бактерии, грибы, простейшие, мелкие беспозвоночные организмы. Редуценты разлагают органические остатки всех трофических уровней продуцентов и консументов на минеральные компоненты. Минеральные вещества и выделяющийся при дыхании редуцентов диоксид углерода (CO2) вновь возвращаются к продуцентам.
Эта замена происходит благодаря особому звену трофической цепи – звену редуцентов. Это звено составляют бактерии, грибы, простейшие, мелкие беспозвоночные организмы. Редуценты разлагают органические остатки всех трофических уровней продуцентов и консументов на минеральные компоненты. Минеральные вещества и выделяющийся при дыхании редуцентов диоксид углерода (CO2) вновь возвращаются к продуцентам.
Рекомендуем прочитать:
Конспект по экологии
СТРУКТУРА ЭКОСИСТЕМЫ ПИЩЕВЫЕ ЦЕПИ И СЕТИ ВОПРОСЫ
СТРУКТУРА ЭКОСИСТЕМЫ. ПИЩЕВЫЕ ЦЕПИ И СЕТИ
ВОПРОСЫ ЛЕКЦИИ I. Экосистема, ее компоненты: продуценты, консументы, редуценты, их роль II. Видовая и пространственная структуры экосистемы III. Трофические уровни. Цепи и сети питания, их звенья IV. Экологические пирамиды
КЛАССИФИКАЦИЯ ЭКОСИСТЕМ Э К О С И С Т Е М А ГЛОБАЛЬНАЯ ЭКОСИСТЕМА (биосфера) МАКРОЭКОСИСТЕМА (например, континент, океан и т. д. ) МЕЗОЭКОСИСТЕМА (например, лес, пруд и т. д. ) МИКРОЭКОСИСТЕМА (например, ствол гниющего дерева)
д. ) МЕЗОЭКОСИСТЕМА (например, лес, пруд и т. д. ) МИКРОЭКОСИСТЕМА (например, ствол гниющего дерева)
КЛАССИФИКАЦИЯ ЭКОСИСТЕМА НАЗЕМНЫЕ ЭКОСИСТЕМЫ (БИОМЫ) Тундра ПРЕСНОВОДНЫЕ ЭКОСИСТЕМЫ Лотические (реки, ручьи и т. д. ) Тайга Широколиственные леса Степи Саванны Пустыни и т. д. Лентические (озера, пруды, водохранилища) Заболоченные угодья (болота, болотистые леса) МОРСКИЕ ЭКОСИСТЕМЫ Открытый океан Прибрежные воды шельфа Районы апвеллинга Эстуарии Глубоководные рифовые зоны
КОМПОНЕНТЫ ЭКОСИСТЕМЫ Э К О С И С Т Е М А НЕОРГАНИЧЕСКИЕ ВЕЩЕСТВА (С, N, CO 2, H 2 O), включающиеся в круговороты ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ (белки, углеводы, липиды, гумусовые вещества и т. д. ) ВОЗДУШНАЯ, ВОДНАЯ, СУБСТРАТНАЯ СРЕД , включая А климатический режим и другие физические факторы АВТОТРОФЫ (ПРОДУЦЕНТЫ) –зеленые растения, сине-зеленые водоросли, фото- хемосистезирующие бактерии ГЕТЕРОТРОФЫ – животные, грибы, большая часть бактерий, подразделяются на: q консументы (растительноядные, плотоядные, паразиты, симбиотрофы) q дентритофаги (сапрофаги) q редуценты
ПРОДУЦЕНТЫ – это автотрофные организмы, вырабатывающие органические вещества из неорганических растения цианобактерии Хемоситезирующие бактерии
КОНСУМЕНТЫ — гетеротрофные организмы, главным образом животные, питающиеся другими организмами или частицами органического вещества Консументы I порядка (растительноядные ) Консументы II порядка (хищные) Консументы III порядка (падальщики)
РЕДУЦЕНТЫ — гетеротрофные организмы, в основном бактерии и грибы, получающие энергию путем разложения мертвых тканей или поглощения растворенного органического вещества бактерии грибы
ПРОСТРАНСТВЕННАЯ СТРУКТУРА ЭКОСИСТЕМЫ
ТРОФИЧЕСКИЕ УРОВНИ ПИЩЕВОЙ ЦЕПИ IV УРОВЕНЬ вторичные хищники (консументы третьего порядка) и паразиты вторичных консументов III УРОВЕНЬ хищники, питающиеся растительноядными животными (консументы второго порядка), и паразиты первичных консументов II УРОВЕНЬ растительноядные животные (фитофаги, консументы первого порядка) автотрофы, зеленые растения (продуценты), первичные потребители солнечной энергии
ТРОФИЧЕСКИЕ УРОВНИ ПИЩЕВОЙ ЦЕПИ
ПРИМЕР ПРОСТОЙ ПИЩЕВОЙ ЦЕПИ
КЛАССИФИКАЦИЯ ПИЩЕВЫХ ЦЕПЕЙ Пищевые цепи ДЕТРИТНЫЕ ПАСТБИЩНЫЕ (автотрофные), в которых в качестве первого звена выступают растения (гетеротрофные), в которых первое звено представлено мертвым органическим веществом, служащим пищей детритофагам
ПАСТБИЩНЫЕ (АВТОТРОФНЫЕ) ПИЩЕВЫЕ ЦЕПИ заяц лиса фитопланктон зоопланктон окунь щука
ДЕТРИТНАЯ (ГЕТЕРОТРОФНАЯ) ПИЩЕВАЯ ЦЕПЬ Опавшие листья Дождевой червь Скворец Сокол
Пищевые сети — это разветвленные пищевые цепи, поскольку особи одного вида могут питаться разнообразной пищей, а особи разных видов одинаковой
ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ.
ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ. ПИРАМИДА БИОМАССЫ Пирамида биомассы наземной экосистемы Пирамида биомасс океана
ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ. ПИРАМИДА ЭНЕРГИЙ
РАСХОД ЭНЕРГИИ В ЗВЕНЬЯХ ПИЩЕВЫХ ЦЕПЕЙ
ПРАКТИЧЕСКАЯ ЧАСТЬ
Задания на выбор одного правильного ответа из предложенных четырех
Задание 1. К продуцентам можно отнести: 1) водоросли 2) грибы 3) сапротрофные бактерии 4) плотоядных животных
Задание 2. Какую роль играют консументы в трофической структуре экосистемы? 1) потребляют готовые органические вещества 2) разлагают остатки органических веществ 3) создают биологическую продукцию, используя потоки солнечной энергии 4) не используют органические вещества
Задание 3. Какую роль выполняют продуценты в экосистеме дубравы? 1) поглощают готовые органические вещества 2) образуют органические вещества 3) разлагают органические вещества 4) выполняют все перечисленные функции
Задание 4. Число трофических уровней пищевой цепи ограничено вследствие: 1) большой биомассы редуцентов; 2) небольшой биомассы консументов; 3) передачи небольшого количества энергии (10%) с одного трофического уровня на другой; 4) большой эффективности передачи энергии с одного трофического уровня на другой (90%)
Число трофических уровней пищевой цепи ограничено вследствие: 1) большой биомассы редуцентов; 2) небольшой биомассы консументов; 3) передачи небольшого количества энергии (10%) с одного трофического уровня на другой; 4) большой эффективности передачи энергии с одного трофического уровня на другой (90%)
Задание 5. Устойчивость лесной экосистемы обеспечивается: 1) высокой численностью грызунов; 2) большой биомассой продуцентов; 3) наличием разветвленных сетей питания; 4) отсутствием редуцентов
Задание 6. Из перечня организмов к детритофагам не относятся: 1) дождевые черви 2) двупарноногие многоножки 3) пескожил 4) личинки капустной белянки
Задание 7. К макроэкосистемам относится 1) березовый лес 2) Тихий океан 3) биосфера 4) небольшой водоем
Задание 8. В цепи выедания размеры организмов при переходе от одного трофического уровня к другому 1) остаются приблизительно одинаковыми 2) постепенно уменьшаются 3) постепенно увеличиваются 4) могут как уменьшаться, так и увеличиваться
Задание 9. Детритная пищевая цепь может начинаться 1) с опавших листьев 2) с зеленых растений 3) с дождевого червя 4) с придонных организмов фильтраторов
Детритная пищевая цепь может начинаться 1) с опавших листьев 2) с зеленых растений 3) с дождевого червя 4) с придонных организмов фильтраторов
Задание 10. В трофической цепи паразитов размеры организмов при переходе с одного пищевого звена к другому 1) остаются приблизительно одинаковыми 2) постепенно уменьшаются 3) постепенно увеличиваются 4) могут как увеличиваться, так и уменьшаться
Задание 11. Какая трофическая сеть составлена правильно: 1) семена ели – мышь – ёж – лисица 2) мышь – семена ели – ёж – лисица 3) лисица – ёж – семена ели – мышь 4) семена ели – ёж – лисица – мышь
Задание 12. В прудовых хозяйствах выгодно выращивать толстолобиков, а не щук, так как: 1) толстолобики быстрее растут 2) щуки чаще гибнут от болезней и неблагоприятных условий 3) толстолобики питаются энергетически дешевой растительной пищей, а щуки дорогой животной 4) мясо толстолобика вкуснее мяса щуки
Задание 13. Трофический уровень определяется как совокупность: 1) продуцентов, консументов и редуцентов 2) организмов, объединенных одним типом питания 3) автотрофов и гетеротрофов 4) организмов, питающихся другом
Трофический уровень определяется как совокупность: 1) продуцентов, консументов и редуцентов 2) организмов, объединенных одним типом питания 3) автотрофов и гетеротрофов 4) организмов, питающихся другом
Задание 14. В пищевой цепи хвоя сосны – сосновый шелкопряд – большая синица – ястреб – пухоед трофических уровней насчитывается 1) 3 2) 4 3) 5 4) 6
Задание 15. Волк и лисица находятся на трофическом уровне, потому что одном 1) поедают растительноядных животных 2) используют свою пищу приблизительно на 10% 3) имеют сходные размеры 4) их кормовой рацион разнообразен
Задание 16. Согласно правилу биологического усиления концентрация токсических веществ в пищевой цепи: 1) уменьшается при переходе от одного уровня к другому в 5 раз 2) уменьшается при переходе от одного уровня к другому в 10 раз 3) увеличивается при переходе от одного уровня к другому в 5 раз 4) увеличивается при переходе от одного уровня к другому в 10 раз трофического
Задание 17. Согласно правилу экологической пирамиды биомассы 1) общее число особей, участвующих в цепях питания с каждым звеном уменьшается 2) на каждом предыдущим трофическом уровне количество биомассы, создаваемой за единицу времени, меньше чем на последующих 3) суммарная масса растений сообщества оказывается больше, чем биомасса всех фитофагов 4) на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени больше, чем на последующих
Согласно правилу экологической пирамиды биомассы 1) общее число особей, участвующих в цепях питания с каждым звеном уменьшается 2) на каждом предыдущим трофическом уровне количество биомассы, создаваемой за единицу времени, меньше чем на последующих 3) суммарная масса растений сообщества оказывается больше, чем биомасса всех фитофагов 4) на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени больше, чем на последующих
Задание 18. Согласно правилу пирамиды чисел 1) общее количество особей, участвующих в цепях питания, с каждым звеном уменьшается 2) на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени меньше, чем на последующем 3) суммарная масса растений сообщества оказывается больше, чем биомасса всех фитофагов 4) общее число особей, участвующих в цепях питания, с каждым звеномувеличивается
Задание 19. Для океана правило пирамиды биомассы имеет «перевернутый вид» , так как: 1) на низших трофических преобладает тенденция к накоплению биомассы 2) на низших трофических преобладает тенденция к накоплению биомассы 3) на высших трофических преобладает тенденция к накоплению биомассы 4) на высших трофических преобладает тенденция к уменьшению биомассы
Задание 20. Хищники в биоценозе выполняют функцию 1) продуцентов 2) редуцентов 3) консументов 2 го порядка 4) консументов 1 го порядка
Хищники в биоценозе выполняют функцию 1) продуцентов 2) редуцентов 3) консументов 2 го порядка 4) консументов 1 го порядка
Задание 21. Продуцентами в биоценозе пресного проточного водоема являются: 1) Бактерии гниения 2) Водные насекомые 3) Растительноядные рыбы 4) Водоросли
Задание 22. Редуцентами в биоценозе пресного проточного водоема являются: 1) Водные насекомые 2) Бактерии гниения 3) Растительноядные рыбы 4) водоросли
Задание 23. Определите редуцента в цепи питания листья растений – гусеницы бабочки – славка – ястреб – бактерии гниения 1) Бактерии гниения 2) Листья растения 3) Гусеницы бабочки 4) Славка
Задание 24. Из представленных ниже вариантов выберите экосистему, которая относится к морским: 1) Эстуарии 2) Болотистые леса 3) Ручьи 4) Пруды
Задание 25. Определите консумента второго порядка в цепи питания листья растения – гусеницы бабочки – насекомоядная птица – ястреб – бактерии гниения 1) Бактерии гниения 2) Листья растения 3) Гусеницы бабочки 4) Насекомоядная птица
Задание 26. Показателем разнообразия экосистемы служит: 1) Небольшое количество видов, имеющих высокую численность 2) Большое разнообразие видов растений и животных 3) Небольшое количество доминирующих видов 4) Большое число популяций доминирующего вида
Показателем разнообразия экосистемы служит: 1) Небольшое количество видов, имеющих высокую численность 2) Большое разнообразие видов растений и животных 3) Небольшое количество доминирующих видов 4) Большое число популяций доминирующего вида
Задание 27. Наземная ярусность растений служит приспособлением к 1) Поглощению воды из почвы 2) Поглощению минеральных веществ 3) Оптимальному использованию лучистой энергии 4) Использованию углекислого газа из атмосферы
Задания на выбор трех верных ответов из представленных шести
Задание 28.
Задание 29.
Задание 30.
Задание 31.
Задание 32.
Задание 33.
Задание 34.
Задание 35.
Задание 36.
Задание 37.
Задание 38. К детритным пищевым цепям относятся: 1) Диатомовые водоросли – личинки поденки – личинки ручейника 2) Коровий помет – личинка мухи – скворец – ястреб перепелятник 3) Листовая подстилка – дождевой червь – землеройка – горностай 4) Нектар – муха – паук – землеройка – сова 5) Мертвое животное – личика падальной мухи – травяная лягушка – обыкновенный уж 6) листья растений – гусеницы бабочки – славка – ястреб – бактерии гниения
Задание 39. Из представленных ниже вариантов выберите лентические экосистемы: 1) саванны 2) озера 3) Эстуарии 4) пруды 5) Заболоченные угодья 6) водохранилища
Задания на установление правильной последовательности биологических процессов, явлений, событий.
Задание 40.
Задание 41.
Задание 42.
Задание 43.
Задание 44. Установите последовательность групп организмов в порядке уменьшения их численности в экосистеме в соответствии с правилом экологической пирамиды. 1) растительноядные насекомые 2) хищные насекомые 3) хищные млекопитающие 4) травянистые растения 5) насекомоядные млекопитающие
Установите последовательность групп организмов в порядке уменьшения их численности в экосистеме в соответствии с правилом экологической пирамиды. 1) растительноядные насекомые 2) хищные насекомые 3) хищные млекопитающие 4) травянистые растения 5) насекомоядные млекопитающие
Задание 45. Установите, в какой последовательности будут распределяться организмы по трофическим уровням в пастбищной цепи питания. 1) кузнечик 2) растение 3) ящерица 4) ястреб
Задание 46. Укажите, в какой последовательности в пищевой цепи должны располагаться указанные объекты: 1) горностай 2) растительный опад 3) землеройка 4) дождевой червь
Задания на установления соответствия
Задание 47.
Задание 48.
Задание 49.
Задание 50.
Задание 51.
Задание 52.
Задания с развернутым ответом
Задание 53. Что служит основой формирования разнообразных сетей питания в экосистемах?
Задание 54. К каким изменениям в экосистеме озера может привести сокращение численности хищных рыб?
Задание 55. Почему природная экосистема смешанного леса является стабильной?
Задание 56. Перечислите основные виды пищевых отношений, в которые вступают бактерии в биоценозах
Задание 57. Укажите роль сапротрофных бактерий в биоценозах
Задание 58. Почему растения (продуценты) считают начальным звеном круговорота веществ и превращения энергии в экосистеме?
Задание 59. Почему широколиственный лес считают более устойчивой экосистемой, чем разнотравный луг?
Задание 60. Почему цепи питания не бывают длинными и обычно состоят из 4— 5 звеньев?
Задание 61. Объясните, как осуществляется саморегуляция в водоеме на примере соотношения численности щук и плотвы.
Объясните, как осуществляется саморегуляция в водоеме на примере соотношения численности щук и плотвы.
СПАСИБО ЗА ВНИМАНИЕ!!!
Ответы № вопроса ответ 1 1 16 4 31 1, 3, 6 46 2, 4, 3, 1 2 1 17 4 32 2, 3, 5 47 1, 2, 2, 1, 1, 2 3 2 18 1 33 1, 4, 6 48 2, 1, 2, 2, 1, 1 4 3 19 3 34 1, 4, 6 49 1, 2, 1, 2 5 3 20 3 35 1, 4, 5 50 1, 1, 2, 1, 1, 2 6 4 21 4 36 1, 3, 6 51 1, 2, 2, 1, 1, 2 7 2 22 2 37 1, 3, 4 51 1, 3, 2, 1, 3, 3 8 4 23 1 38 2, 3, 5 9 1 24 1 39 2, 4, 6 10 2 25 4 40 3, 2, 1, 4 11 1 26 2 41 4, 3, 2, 1, 5 12 3 27 3 42 4, 3, 1, 2 13 2 28 1, 2, 6 43 4, 2, 1, 3 14 3 29 1, 5, 6 44 4, 1, 2, 5, 3 15 1 30 1, 2, 6 45 2, 1, 3, 4
Ответы 53. Элементы ответа: 1. Разнообразие видов, наличие среди них продуцентов, консументов, редуцентов. 2. Питание видов разнообразной пищей (широкая пищевая специализация). 54. Элементы ответа: 1. К увеличению численности растительноядных организмов 2. К уменьшению численности растений 55. Элементы ответа: 1. Разнообразие видов растений, животных и других организмов 2. Разветвленные цепи (сети) питания, наличие нескольких трофических уровней 3. Сбалансированный круговорот веществ 56. Элементы ответа: 1. Хемотрофы используют для производства органических соединений энергию окислительно восстановительных реакций 2. Фототрофы синтезируют органические вещества, используя энергию солнечного света 3. Сапротрофы поглощают органические вещества отмерших тел живых организмов или их выделения. 4. Паразиты питаются органическими веществами живых тел
Элементы ответа: 1. Разнообразие видов растений, животных и других организмов 2. Разветвленные цепи (сети) питания, наличие нескольких трофических уровней 3. Сбалансированный круговорот веществ 56. Элементы ответа: 1. Хемотрофы используют для производства органических соединений энергию окислительно восстановительных реакций 2. Фототрофы синтезируют органические вещества, используя энергию солнечного света 3. Сапротрофы поглощают органические вещества отмерших тел живых организмов или их выделения. 4. Паразиты питаются органическими веществами живых тел
Ответы 57. Элементы ответа: 1. Гниение – превращение погибших растений и животных в перегной 2. Минерализация – превращение перегноя в минеральные вещества, всасываемыми корнями растений 3. Клубеньковые бактерии на корнях бобовых растений помагают усваивать азот из воздуха. 58. Элементы ответа: 1. создают органические вещества из неорганических; 2. аккумулируют солнечную энергию; 3. обеспечивают органическими веществами и энергией организмы других звеньев экосистемы. 59. Элементы ответа: 1. в широколиственном лесу большее разнообразие видов, чем на лугу; 2. многоярусность и большое разнообразие экологических ниш; 3. разветвленные сети питания обеспечивают более полный круговорот веществ.
59. Элементы ответа: 1. в широколиственном лесу большее разнообразие видов, чем на лугу; 2. многоярусность и большое разнообразие экологических ниш; 3. разветвленные сети питания обеспечивают более полный круговорот веществ.
Ответы 60. Элементы ответа: При передаче вещества и энергии значительная часть энергии теряется (см. справочный материал). Поэтому каждому новому звену пищевой цепи достается все меньше и меньше энергии. Полная потеря энергии прекращает пищевую цепь. Причина в нехватке энергии, которая теряется в каждом звене пищевой цепи. 61. Элементы ответа: Саморегуляция в экосистемах основана на пищевых связях. В данном примере щука хищник, потребляющий плотву в пищу. Поэтому 1) при увеличении численности плотвы увеличивается чис ленность щук; 2) увеличение численности щук приводит к сокращению численности плотвы; 3) таким образом осуществляется саморегуляция рыб в водоеме
Урок №28. Пищевые цепи, пищевые сети и трофические уровни
Методическое пособие уроков с видеоматериалами
Тип урока — комбинированный
Методы: частично-поисковый, проблемного изложения, репродуктивный, объяснительно-иллюстративный.
Цель:
— осознание учащимися значимости всех обсуждаемых вопросов, умение строить свои отношения с природой и обществом на основе уважения к жизни, ко всему живому как уникальной и бесценной части биосферы;
Задачи:
Образовательные: показать множественность факторов, действующих на организмы в природе, относительность понятия «вредные и полезные факторы», многообразие жизни на планете Земля и варианты адаптаций живых существ ко всему спектру условий среды обитания.
Развивающие: развивать коммуникативные навыки, умения самостоятельно добывать знания и стимулировать свою познавательную активность; умения анализировать информацию, выделять главное в изучаемом материале.
Воспитательные:
Воспитывать культуру поведения в природе, качества толерантной личности, прививать интерес и любовь к живой природе, формировать устойчивое положительное отношение к каждому живому организму на Земле, формировать умение видеть прекрасное.
УУД
Личностные: познавательный интерес к экологии.. Понимание необходимости получения знаний о многообразии биотических связей в природных сообществах для сохранения естественных биоценозов. Способность выбирать целевые и смысловые установки в своих действиях и поступках по отношению к живой природе. Потребность в справедливом оценивании своей работы и работы одноклассников
Познавательные: умение работать с различными источниками информации, преобразовывать её из одной формы в другую, сравнивать и анализировать информацию, делать выводы, готовить сообщения и презентации.
Регулятивные: умение организовать самостоятельно выполнение заданий, оценивать правильность выполнения работы, рефлексию своей деятельности.
Коммуникативные: участвовать в диалоге на уроке; отвечать на вопросы учителя, товарищей по классу, выступать перед аудиторией, используя мультимедийное оборудование или другие средства демонстрации
Планируемые результаты
Предметные: знать — понятия «среда обитания», «экология», «экологические факторы» их влияние на живые организмы, «связи живого и неживого»;. Уметь — определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.
Уметь — определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.
Личностные: высказывать суждения, осуществлять поиск и отбор информации; анализировать связи, сопоставлять, находить ответ на проблемный вопрос
Метапредметные: связи с такими учебными дисциплинами как биология, химия, физика, география. Планировать действия с поставленной целью; находить необходимую информацию в учебнике и справочной литературе; осуществлять анализ объектов природы; делать выводы; сформулировать собственное мнение.
Форма организации учебной деятельности – индивидуальная, групповая
Методы обучения: наглядно-иллюстративный, объяснительно-иллюстративный, частично-поисковый, самостоятельная работа с дополнительной литературой и учебником, с ЦОР.
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Изучение нового материала
Пищевые цепи, пищевые сети и трофические уровни
Пищевой цепью называется перенос энергии, заключенной в растительной пище — поскольку лишь растения создают органическое вещество из неорганического — через ряд организмов в процессе их поедания друг другом
Организмы, получающие свою энергию от Солнца через одинаковое число ступеней, принадлежат к одному трофическому уровню (от греч. ггорке — питание, пища), т. е. это совокупность организмов, занимающих определенное положение в общей цепи питания. Зеленые растения (автотрофы) занимают первый трофический уровень (уровень продуцентов), травоядные — второй (уровень первичных консументов), первичные хищники, поедающие травоядных, — третий (уровень вторичных консументов), вторичные хищники — четвертый (уровень третичных консументов).
Различают два типа пищевых цепей: пастбищная и детритная.
Пастбищная цепь
зеленое растение -> растительноядное животное -» хищник
Детритная цепь
мертвое -» микроорганизмы
органическое (бактерии, грибы)
вещество (детрит) -> детритофаги -» хищники детрито
Пищевые цепи не изолированы одна от другой, а тесно переплетены. Они составляют так называемые пищевые сети. Принцип образования пищевых сетей состоит в следующем. Каждый продуцент имеет не одного, а несколько консументов. В свою очередь, консументы, среди которых преобладают полифаги (от греч. ро1у — много), пользуются не одним, а несколькими источниками питания.
Вопросы и задания
1.Что называется пищевой цепью? Приведите 2-3 примера пищевых цепей в наземно-воздушной и водной средах жизни.
2.Приведите пример пищевой сети в наземно-воздушной среде жизни.
3.К какому трофическому уровню относятся следующие организмы: заяц-беляк, лисица обыкновенная, лось, лесные травы?
4.Приведите 2-3 примера организмов, относящихся к первому, второму и третьему трофическому уровню водной экосистемьГ
Пищевые цепи питания в экологии
Цепи питания
Презентация к уроку биологии по теме: Цепи питания
 youtube.com/embed/KhySWj0EYrM»>
youtube.com/embed/KhySWj0EYrM»>
Ресурсы:
С. В. Алексеев. Экология: Учебное пособие для учащихся 9 класса общеобразовательных учреждений разных видов. СМИО Пресс, 1997. — 320 с
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
— http://ppt4web.ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
Трофические цепи и трофические уровни. Понятие о трофических уровнях
Знакомы ли вы с такими понятиями, как консументы, редуценты и продуценты? Если нет, то наша статья — для вас. На самом деле эти организмы хорошо известны каждому. Кто же это такие? Давайте разберемся вместе.
Понятие о трофической цепи
Все компоненты экосистемы тесно взаимосвязаны между собой. Благодаря этому в природе и формируются различные сообщества. В структуру любой экосистемы входит абиотическая и биотическая часть. Первая представляет собой совокупность живых организмов. Она называется биоценозом. К абиотической части относятся минеральные и органические соединения.
Первая представляет собой совокупность живых организмов. Она называется биоценозом. К абиотической части относятся минеральные и органические соединения.
Функционирование любой экосистемы связано с преобразованием энергии. Ее основным источником служит солнечный свет. Фотосинтезирующие организмы используют его для синтеза органических веществ. Гетеротрофы получают энергию при расщеплении органических веществ. Для роста используется только ее незначительная часть. А остальная расходуется для осуществления процессов жизнедеятельности.
В результате образуются очередности, в которых особи одних видов, их остатки или продукты жизнедеятельности являются источником питания других. Они называются трофическими или цепями питания.
Трофические уровни
Каждая цепь питания состоит из определенного количества звеньев. Установлено, что при переходе от одного к другому часть энергии постоянно утрачивается. Поэтому количество звеньев обычно равно 4-5. Положение популяции отдельных видов в цепи питания называется трофическим уровнем.
Что такое консументы
Все организмы объединяют в группы. К ним относятся представители абсолютно всех царств живой природы, независимо от уровня их организации. Рассмотрим каждую из них.
Консументы: порядки
Гетеротрофы в пищевой цепи занимают разные уровни. Все растительноядные виды являются Следующий уровень — это хищники. Они уже консументы второго порядка.
Рассмотрим эту иерархию на конкретном примере. Допустим, трофическая сеть имеет вид: комар, лягушка, аист. Кто из них консумент первого порядка? Это лягушка. Тогда консументом второго порядка является аист. В природе встречаются гетеротрофы, которые питаются и растениями, и животными. Такие консументы могут одновременно находиться на нескольких трофических уровнях.
Продуценты
Говоря о том, что такое консументы, мы обратили внимание на тип их питания. Рассмотрим в этом ракурсе и другую группу трофической сети. Продуценты — это группа организмов, которые являются автотрофами. Они способны синтезировать органические вещества из минеральных.
Они способны синтезировать органические вещества из минеральных.
Различают продуценты двух видов: авто — и хемотрофы. Первые используют для создания органики энергию солнечного света. Это растения, цианобактерии, некоторые простейшие животные. Хемотрофы обладают способностью окислять различные химические соединения. При этом вырабатывается энергия, которую они используют для осуществления продуктов жизнедеятельности. К ним относятся азотфиксирующие, серо-, железобактерии.
Наличие продуцентов является необходимым условием развития любой экосистемы. Этот факт объясняется тем, что фотосинтезирующие организмы являются источником поступления энергии.
Редуценты
Еще одна роль в экосистеме принадлежит гетеротрофным организмам, которые питаются органическими веществами остатков или продуктов жизнедеятельности других видов, которые они разлагают до минеральных веществ. Эту функцию и выполняют редуценты. Представителями этой группы являются бактерии и грибы.
Именно на уровне продуцентов в экосистеме происходит накопление энергии. Затем она проходит через консументы и продуценты, где расходуется. На каждом последующем трофическом уровне часть энергии рассеивается в виде тепла.
Затем она проходит через консументы и продуценты, где расходуется. На каждом последующем трофическом уровне часть энергии рассеивается в виде тепла.
Виды цепей питания
Энергия в экосистеме разделена на два потока. Первый направлен к консументам от продуцентов, в второй — от мертвой органики. В зависимости от этого различают трофические сети пастбищного и детритного типа. В первом случае начальным трофическим уровнем являются продуценты, которые передают энергию консументам разных уровней. Завершается пастбищная цепь редуцентами.
Детритная цепь начинается мертвой органикой, а продолжается сапротрофами, которые являются представителями консументов. Последним звеном в этой цепи также являются редуценты.
В пределах любой экосистемы одновременно существует множество трофических цепей. Все они неотделимы друг от друга и тесно переплетаются. Так происходит, поскольку представители одного вида могут одновременно являться звеньями разных цепей. Благодаря этому и формируются трофические сети. И чем они разветвленнее, тем устойчивее экосистема.
И чем они разветвленнее, тем устойчивее экосистема.
Перенос энергии
пищи от её источника — автотрофов
(растений) — через ряд организмов,
происходящий путём поедания одних
организмов другими, называется пищевой
цепью. При
каждом переносе большая часть (80-90%) потенциальной
энергии теряется, переходя в тепло.
Поэтому, чем короче пищевая цепь (чем
ближе организм к её началу) , тем больше
количество энергии, доступной для
популяции. Пищевые цепи можно разделить
на два основных типа: пастбищная
цепь, которая
начинается с зелёного растения и идёт
далее к пасущимся растительноядным
животным (т.е. к организмам, поедающим
живые растительные клетки или ткани) и
к хищникам (организмам, поедающим
животных), и детритная
цепь , которая
от мёртвого органического вещества
идёт к микроорганизмам, а затем к
детритофагам и к их хищникам. Пищевые
цепи не изолированы одна от другой, а
тесно переплетаются друг с другом,
образуя, так называемые пищевые
сети. В
сложных природных сообществах организмы,
получающие свою энергию от Солнца через
одинаковое число ступеней, считаются
принадлежащими к одному трофическому
уровню. Так,
зелёные растения занимают первый
трофический уровень (уровень продуцентов),
травоядные — второй (уровень первичных
консументов), первичные хищники, поедающие
травоядных, — третий (уровень вторичных
консументов), а вторичные хищники —
четвёртый (уровень третичных консументов).
Так,
зелёные растения занимают первый
трофический уровень (уровень продуцентов),
травоядные — второй (уровень первичных
консументов), первичные хищники, поедающие
травоядных, — третий (уровень вторичных
консументов), а вторичные хищники —
четвёртый (уровень третичных консументов).
Пищевые цепи
знакомы каждому из нас: человек съедает
крупную рыбу, а она ест мелких рыб,
поедающих зоопланктон, который питается
фитопланктоном, улавливающим солнечную
энергию, или же человек может употреблять
в пищу мясо коров, которые едят траву,
улавливающую солнечную энергию, он
может использовать и гораздо более
короткую пищевую цепь, питаясь зерновыми
культурами, которые улавливают солнечную
энергию. В последнем случае человек
является первичным консументом на
втором трофическом уровне. В пищевой
цепи трава — коровы — человек он является
вторичным консументом на третьем
трофическом уровне. Но чаще человек
является одновременно и первичным и
вторичным консументом, так как в его
диету обычно входит смесь растительной
и животной пищи.
При каждом переносе пищи часть потенциальной энергии теряется. Прежде всего, растения фиксируют лишь малую долю поступающей энергии солнечного излучения. Поэтому число консументов (например, людей), которые могут прожить при данном выходе первичной продукции, сильно зависит от длины цепи, переход к каждому следующему звену в нашей традиционной сельскохозяйственной пищевой цепи уменьшает доступную энергию примерно на порядок величины (т.е. в 10 раз). Поэтому если в рационе увеличивается содержание мяса, то уменьшается число людей, которых можно прокормить. Если окажется, что на основе имеющейся первичной продукции придётся кормить очень много новых ртов, то нужно вовсе отказываться от мяса или резко снизить его потребление.
Некоторые вещества по мере продвижения по цепи не рассеиваются, а наоборот накапливаются. Это так называемое концентрирование в пищевой цепи (биоконцентрирование) нагляднее всего демонстрируют устойчивые радионуклиды и пестициды.
Тенденция некоторых
радионуклидов, побочных продуктов
деления ядра атома увеличивать свою
концентрацию с каждым этапом пищевой
цепи была обнаружена в 50-ых годах. Крайне
малые (следовые) количества радиоактивного
J,
P,
Cs,
Se
в реке Колумбия концентрировались в
тканях рыб и птиц. Было обнаружено, что
коэффициент накопления (соотношение
количества вещества в тканях и окружающей
среде) радиоактивного фосфора в яйцах
гусей равен 2 млн. Таким образом, безопасные
выбросы в реку могут стать крайне
опасными для высших звеньев пищевой
цепи.
Крайне
малые (следовые) количества радиоактивного
J,
P,
Cs,
Se
в реке Колумбия концентрировались в
тканях рыб и птиц. Было обнаружено, что
коэффициент накопления (соотношение
количества вещества в тканях и окружающей
среде) радиоактивного фосфора в яйцах
гусей равен 2 млн. Таким образом, безопасные
выбросы в реку могут стать крайне
опасными для высших звеньев пищевой
цепи.
Пример: ДДТ (4,4 — дихлордифенил трихлорметилметан).
Чтобы сократить численность комаров
на Лонг-Айленде, болота много лет опыляли
ДДТ. Специалисты по борьбе с насекомыми
не применяли таких концентраций, которые
были бы непосредственно летальны для
рыбы и других животных, но они не учли
экологических процессов и длительного
сохранения остатков ДДТ. Вместо того,
чтобы смываться в море, ядовитые остатки
адсорбированные на детринге,
концентрировались в тканях детрингофагов
и мелких рыб и далее — в хищниках высшего
порядка (рыбоядные птицы). Коэффициент
концентрации (отношение содержания ДДТ
в организме к содержанию в воде, выраженное
в частях на миллион) составляет для
рыбоядных животных около 500 000. У рыб и
птиц накоплению способствует значительные
жировые накопления, в которых
концентрируется ДДТ. Птицы особенно
чувствительны к отравлению ДДТ, т.к.
этот яд (и др. инсектициды, представляющие
собой хлорированные углеводороды)
посредством снижения в крови концентрации
стероидных гормонов нарушает образование
яичной скорлупы; тонкая скорлупа лопается
ещё до того, как разовьётся птенец. Таким
образом, очень малые дозы, неопасные
для особи, оказываются летальными для
популяции.
У рыб и
птиц накоплению способствует значительные
жировые накопления, в которых
концентрируется ДДТ. Птицы особенно
чувствительны к отравлению ДДТ, т.к.
этот яд (и др. инсектициды, представляющие
собой хлорированные углеводороды)
посредством снижения в крови концентрации
стероидных гормонов нарушает образование
яичной скорлупы; тонкая скорлупа лопается
ещё до того, как разовьётся птенец. Таким
образом, очень малые дозы, неопасные
для особи, оказываются летальными для
популяции.
Принципы биологического накопления надо учитывать при любых решениях, связанных с поступлением загрязнений в среду. Многие небиологические факторы, однако, могут уменьшать или увеличивать коэффициент концентрации. Так, человек получает меньше ДДТ, чем птица, т.к. при обработке и варке пищи часть этого вещества удаляется.
Трофический уровень — это совокупность организмов, занимающих определённое место в пищевой сети.
I трофический уровень — всегда растения,
II трофический уровень — первичные консументы
III
трофический уровень
— вторичные
консументы и т. д.
д.
Детритофаги могут находиться на II и выше трофическом уровне.
Обычно в экосистеме насчитывается 3-4 трофических уровня.
Трофическую структуру можно измерить и выразить либо урожаем на корню (на единицу площади), либо количеством энергии, фиксируемой на единице площади за единицу времени на последовательных трофических уровнях.
Трофическую структуру и трофическую функцию можно изобразить графически в виде экологических пирамид , основанием которых служит первый уровень (уровень продуцентов), а последующие уровни образуют этажи и вершину пирамиды. Экологические пирамиды можно отнести к трём основным типам:
пирамида чисел, отражающая численность отдельных организмов;
пирамида биомассы , характеризующая общую сухую массу, калорийность или другую меру общего количества живого вещества;
пирамида энергии показывающая величину потока энергии
и «продуктивность» на последовательных
трофических уровнях. С каждым переходом
из одного трофического уровня в другой
в пределах пищевой цепи или сети
совершается работа и в окружающую среду
выделяется тепловая энергия, а количество
энергии высокого качества, используемой
организмами следующего трофического
уровня, снижается. Процентное содержание
энергии высокого качества, переходящей
из одного трофического уровня в другой
колеблется от 2 до 30%. Большая часть
энергии теряется в окружающей среде
как тепловая энергия низкого качества.
Чем длиннее пищевая цепь, тем больше
теряется полезной энергии. Пирамида
энергетических потоков объясняет,
почему можно прокормить большее
количество людей, если сократить пищевую
цепь до прямого потребления зерновых
(рис – человек), чем если в качестве
пищи использовать животных, потребляющих
зерно. Чтобы избежать белкового
(протеинового) недоедания, вегитарианское
питание должно состоять из разнообразных
растений.
Процентное содержание
энергии высокого качества, переходящей
из одного трофического уровня в другой
колеблется от 2 до 30%. Большая часть
энергии теряется в окружающей среде
как тепловая энергия низкого качества.
Чем длиннее пищевая цепь, тем больше
теряется полезной энергии. Пирамида
энергетических потоков объясняет,
почему можно прокормить большее
количество людей, если сократить пищевую
цепь до прямого потребления зерновых
(рис – человек), чем если в качестве
пищи использовать животных, потребляющих
зерно. Чтобы избежать белкового
(протеинового) недоедания, вегитарианское
питание должно состоять из разнообразных
растений.
Пирамиды чисел Можно
собрать все образцы организмов в
экосистеме и подсчитать численность
всех видов, обнаруженных на каждом
трофическом уровне. Такая информация
необходима для создания пирамиды
численностей. Например, миллион особей
фитоплангтона в небольшом пруду может
прокормить 10 000 особей зооплангтона,
которые в свою очередь прокормят 100
окуней, которых будет достаточно, чтобы
прокормиться одному человеку в течение
месяца.
Рис. 3.2 Пирамида чисел
Но для некоторых экосистем пирамиды численностей имеют другую форму. Например, в лесу небольшое количество больших деревьев, таких как секвойя вечнозеленая, снабжает пищей огромное количество небольших по размеру насекомых-фитофагов и птиц – консументов первого порядка.
Пирамида биомассы , характеризующая массу живого вещества (на ед. площади или объема). Каждый трофический уровень пищевой цепи или сети содержит определённое количество биомассы. В наземных экосистемах действует следующее правило пирамиды биомасс : суммарная масса растений превышает массу всех травоядных, а их масса превышает всю биомассу хищников.
Для океана правило
пирамиды биомасс недействительно –
пирамида имеет перевернутый
(обращенный) вид .
Для экосистемы океана характерно
накаливание биомассы на высоких уровнях,
у хищников. Хищники живут долго, и
скорость оборота их регенерации мала,
но у продуцентов – фитопланктонных
водорослей оборачиваемость в сотни раз
превышает запас биомассы.
Рис. 3.3 Пирамида биомассы
Пирамиды чисел и биомассы могут быть обращёнными, (или частично обращёнными), т.е. основание может быть меньше, чем один или несколько верхних этажей. Так бывает, когда средние размеры продуцентов меньше размеров консументов. Напротив, энергетическая пирамида всегда будет сужаться к верху, при условии, что мы учитываем все источники пищевой энергии в системе.
Устойчивые биогеохимические циклы вещества и энергии в биосфере нашей планеты формируются вследствие биологического разнообразия потребляемого организмами набора веществ и выделяемых в природную среду продуктов жизнедеятельности. Базу биологического круговорота веществ составляют трофические уровни , которые представлены конкретными видами живых организмов, делящимися на три основные группы: продуценты, консументы и редуценты. Трофический уровень составляют популяции организмов, выполняющих в экосистеме одинаковые трофические функции и имеющих различный видовой состав (от греч. trophe — «питание»).
trophe — «питание»).
Первый трофический уровень — уровень первичной продукции — образуют автотрофы. Это организмы, которые синтезируют органические вещества (углеводы, жиры, белки, нуклеиновые кислоты) из неорганических соединений, используя энергию Солнца. Первичная продукция — это биомасса растительных тканей. Первичные продуценты — растения, фотоавтотрофные бактерии и хемосинтезирующие бактерии (хемотрофы). Хемотрофы — микроорганизмы, синтезирующие органическое вещество за счет энергии окисления аммиака, сероводорода и других веществ, имеющихся в воде и почве.
Второй трофический уровень представляют консументы (гетеротрофы) :
1) первого порядка — фитофаги — используют в качестве пищи растения;
2) второго порядка — питаются животной пищей.
На третьем трофическом уровне — редуценты . Это организмы, разлагающие до минеральных веществ, диоксида углерода и воды отходы жизнедеятельности и отмершие организмы. Консументы также участвуют в минерализации органических веществ.
Все организмы используют в пищу биомассу предыдущих трофических уровней, теряя энергию с потерями на дыхание, обогрев тела, на различные формы деятельности, на выделение экскрементов.
Между видами разных трофических уровней существуют взаимоотношения, образующие систему трофических цепей (цепей питания). Использование ресурсов на каждом трофическом уровне зависит от видового разнообразия экосистемы.
Видовое разнообразие может снижаться в зонах загрязнения, вызывая упрощение трофической структуры.
Сегодня фиксируются нарушения структуры биоценозов вследствие загрязнения окружающей природной среды. Токсиканты передаются по цепям питания и способствуют гибели животных, птиц, гидробионтов, а также накапливаются в пищевых продуктах, потребляемых человеком.
| Предыдущие материалы: |
Огромную роль в воспроизводстве жизни играет энергия Солнца. Количество этой энергии очень велико (примерно 55 ккал на 1 см 2 в год). Из этого количества продуценты — зеленые растения — в результате фотосинтеза фиксируют не более 1-2 % энергии, а пустыни и океан — сотые доли процента.
Число звеньев в пищевой цепи может быть различным, но обычно их 3-4 (реже 5). Дело в том, что к конечному звену пищевой цепи поступает так мало энергии, что ее не хватит в случае увеличения числа организмов.
Рис. 1. Пищевые цепи в наземной экосистеме
Совокупность организмов, объединенных одним типом питания и занимающих определенное положение в пищевой цепи, носит название трофический уровень. К одному трофическому уровню принадлежат организмы, получающие свою энергию от Солнца через одинаковое число ступеней.
Простейшая пищевая цепь (или цепь питания) может состоять из фитопланктона, затем идут более крупные травоядные планктонные ракообразные (зоопланктон), а заканчивается цепь китом (или мелкими хищниками), которые фильтруют этих ракообразных из воды.
Природа сложна. Все ее элементы, живые и неживые, — одно целое, комплекс приспособленных друг к другу, взаимодействующих и взаимосвязанных явлений и существ. Это звенья одной цепи. И если удалить из общей цепочки хотя бы одно такое звено, результаты могут быть непредвиденными.
Особенно негативно разрыв цепей питания может сказаться на леса — будь то лесные биоценозы умеренной зоны либо отличающиеся богатым видовым разнообразием биоценозы тропического леса. Многие виды деревьев, кустарников или травянистых растений пользуются услугами определенного опылителя — пчелы, осы, бабочки или колибри, обитающих в пределах ареала данного растительного вида. Как только погибнет последнее цветущее дерево или травянистое растение, опылитель вынужден будет покинуть данное местообитание. В результате погибнут питающиеся этими растениями или плодами дерева фитофаги (травоядные). Без пиши останутся охотившиеся на фитофагов хищники, а далее изменения последовательно коснутся остальных звеньев пищевой цепи. В итоге они скажутся и на человеке, поскольку у него есть свое определенное место в пищевой цепи.
Пищевые цепи можно разделить на два основных типа: пастбищную и детритную. Пищевые цени, которые начинаются с автотрофных фотосинтезирующих организмов, называются пастбищными, или цепями выедания. На вершине пастбищной цепи стоят зеленые растения. На втором уровне пастбищной цепи обычно находятся фитофаги, т.е. животные, питающиеся растениями. Примером пастбищной пищевой цепи могут служить взаимоотношения между организмами на пойменном лугу. Начинается такая цепь с лугового цветкового растения. Следующее звено — бабочка, питающаяся нектаром цветка. Затем идет обитатель влажных местообитаний — лягушка. Ее покровительственная окраска позволяет ей подстеречь жертву, но не спасает от другого хищника — обыкновенного ужа. Цапля, поймав ужа, замыкает пищевую цепь на пойменном лугу.
На вершине пастбищной цепи стоят зеленые растения. На втором уровне пастбищной цепи обычно находятся фитофаги, т.е. животные, питающиеся растениями. Примером пастбищной пищевой цепи могут служить взаимоотношения между организмами на пойменном лугу. Начинается такая цепь с лугового цветкового растения. Следующее звено — бабочка, питающаяся нектаром цветка. Затем идет обитатель влажных местообитаний — лягушка. Ее покровительственная окраска позволяет ей подстеречь жертву, но не спасает от другого хищника — обыкновенного ужа. Цапля, поймав ужа, замыкает пищевую цепь на пойменном лугу.
Если пищевая цепь начинается с отмерших остатков растений, трупов и экскрементов животных — детрита, она называется детритной , или цепью разложения. Термин «детрит» означает продукт распада. Он позаимствован из геологии, где детритом называют продукты разрушения горных пород. В экологии детрит — это органическое вещество, вовлеченное в процесс разложения. Такие цепи характерны для сообществ дна глубоких озер, океанов, где многие организмы питаются за счет оседания детрита, образованного отмершими организмами верхних освещенных слоев водоема.
В лесных биоценозах детритная цепь начинается с разложения мертвого органического вещества животными-сапрофагами. Наиболее активное участие в разложении органики здесь принимают почвенные беспозвоночные животные (членистоногие, черви) и микроорганизмы. Присутствуют и крупные сапрофаги — насекомые, которые готовят субстрат для организмов, осуществляющих процессы минерализации (для бактерий и грибов).
В отличие от пастбищной цепи размеры организмов при движении вдоль детритной цепи не возрастают, а, наоборот, уменьшаются. Так, на втором уровне могут стоять насекомые-могильщики. Но наиболее типичными представителями детритной цепи являются грибы и микроорганизмы, питающиеся мертвым веществом и довершающие процесс разложения биоорганики до состояния простейших минеральных и органических веществ, которые затем в растворенном виде потребляются корнями зеленых растений на вершине пастбищной цепи, начиная тем самым новый круг движения вещества.
В одних экосистемах преобладают пастбищные, в других — детритные цепи. Например, лес считается экосистемой с преобладанием детритных цепей. В экосистеме гниющего пня пастбищная цепь вообще отсутствует. В то же время, например, в экосистемах поверхности моря практически все продуценты, представленные фитопланктоном, потребляются животными, а их трупы опускаются на дно, т.е. уходят изданной экосистемы. В таких экосистемах преобладают пастбищные пищевые цепи, или цепи выедания.
Например, лес считается экосистемой с преобладанием детритных цепей. В экосистеме гниющего пня пастбищная цепь вообще отсутствует. В то же время, например, в экосистемах поверхности моря практически все продуценты, представленные фитопланктоном, потребляются животными, а их трупы опускаются на дно, т.е. уходят изданной экосистемы. В таких экосистемах преобладают пастбищные пищевые цепи, или цепи выедания.
Общее правило , касающееся любой пищевой цепи, гласит: на каждом трофическом уровне сообщества большая часть поглощаемой с пищей энергии тратится на поддержание жизнедеятельности, рассеивается и больше не может быть использована другими организмами
. Таким образом, потребленная пища на каждом трофическом уровне ассимилируется не полностью. Значительная ее часть расходуется на обмен веществ. При переходе к каждому последующему звену пищевой цепи общее количество пригодной для использования энергии, передаваемой на следующий, более высокий трофический уровень, уменьшается.
Энергия передается от организма к организму, создающих пищевую или трофическую (греч. trophe-пища) цепь от автотрофов, продуцентов (создателей) к гетеротрофам, консументам (пожирателям) и так 4-6 раз с одного трофического уровня на другой.
Трофический уровень — это место каждого звена в пищевой цепи. Первый трофический уровень — это продуценты, все остальные — консументы. Второй трофический уровень — это растительноядные консументы; третий — плотоядные консументы, питающиеся растительноядными формами; четвертый — консументы, потребляющие других плотоядных, и т.д. Следовательно, можно и консументов разделить по уровням: консументы первого, второго, третьего и т.д. порядков.
Четко распределяются по уровням лишь консументы, специализирующиеся на определенном виде пиши. Однако есть виды, которые питаются мясом и растительной пищей (человек, медведь и др.), которые могут включаться в пищевые цепи на любом уровне.
Пища, поглощаемая консументом, усваивается не полностью — от 12 до 20% у некоторых растительноядных, до 75% и более у плотоядных. Энергетические затраты связаны прежде всего с поддержанием метаболических процессов, траты на дыхание, оцениваемая общим количеством СО 2 , выделенного организмом. Значительно меньшая часть идет на образование тканей и некоторого запаса питательных веществ, т.е. на рост. Остальная часть пищи выделяется в виде экскрементов. Кроме того, значительная часть энергии рассеивается в виде тепла при химических реакциях в организме и особенно при активной мышечной работе. В конечном итоге вся Энергия, использованная на метаболизм, превращается в тепловую и рассеивается в окружающей среде.
Энергетические затраты связаны прежде всего с поддержанием метаболических процессов, траты на дыхание, оцениваемая общим количеством СО 2 , выделенного организмом. Значительно меньшая часть идет на образование тканей и некоторого запаса питательных веществ, т.е. на рост. Остальная часть пищи выделяется в виде экскрементов. Кроме того, значительная часть энергии рассеивается в виде тепла при химических реакциях в организме и особенно при активной мышечной работе. В конечном итоге вся Энергия, использованная на метаболизм, превращается в тепловую и рассеивается в окружающей среде.
Таким образом, большая часть энергии при переходе с одного трофического уровня на другой, более высокий, теряется. Приблизительно потери составляют около 90%. На каждый следующий уровень передается не более 10% энергии от предыдущего уровня. Так, если калорийность продуцента 1000 Дж, то при попадании в тело фитофага остается 100 Дж, в теле хищника уже 10 Дж, а если этот хищник будет съеден другим, то на его долю останется лишь 1 Дж, т. е. 0,1 % от калорийности растительной пищи.
е. 0,1 % от калорийности растительной пищи.
Однако такая строгая картина перехода энергии с уровня на уровень не совсем реальна, поскольку трофические цепи экосистем сложно переплетаются, образуя трофические сети. Но конечный итог: рассеивание и потеря энергии, которая, чтобы существовала жизнь, должна возобновляться.
Нельзя забывать еще и мертвую органику, которой питается значительная часть гетеротрофов. Среди них есть и сапрофаги и сапрофиты (грибы), использующие энергию, заключенную в детрите. Поэтому различают два вида трофических цепей:
- Пастбищные (цепи выедания) — начинаются с продуцентов; для таких цепей при переходе с одного трофического уровня на другой характерно увеличение размеров особей при одновременном уменьшении плотности
Разница между пищевой цепью и пищевой сетью
Пищевая цепочка может быть названа прямым и единым путем для потока энергии в экосистеме через различные виды организмов. Пищевая сеть, с другой стороны, определяется как извилистый или сложный путь экосистемы, состоящий из многочисленных пищевых цепей различного трофического уровня, через которые проходит поток энергии.
Пищевая сеть, с другой стороны, определяется как извилистый или сложный путь экосистемы, состоящий из многочисленных пищевых цепей различного трофического уровня, через которые проходит поток энергии.
Пищевые цепочки, пищевая сеть — это часть экосистемы, которую можно описать как сообщество самых маленьких микроорганизмов, растений высших животных, которые питаются, живут, размножаются, взаимодействуют и умирают в одной и той же области или среде. Все экосистемы имеют иерархию питания, которая включает солнце (источник энергии), производителя, потребителя и разложителя.
Но в экосистеме невозможно иметь единую пищевую цепочку. Чтобы выжить, организмы питаются на другом трофическом уровне. Таким образом, в пищевой сети различные пищевые цепи связаны друг с другом, а также перекрещиваются друг с другом, образуя сложную сеть. Пищевая сеть повышает конкурентоспособность и адаптивность среди организмов, но ее не видно в пищевой цепи.
В этой статье мы сосредоточимся на некоторых важных моментах, которые отличают пищевую цепочку и пищевую сеть.
Сравнительная таблица
| Основа для сравнения | Пищевая цепочка | Пищевая сеть |
|---|---|---|
| Смысл | Поток энергии через один прямой путь от нижнего трофического уровня к более высокому трофическому уровню называется пищевой цепью. | Взаимосвязанные многочисленные пищевые цепи, через которые поток энергии в экосистеме называется пищевой сетью. |
| Количество цепей | Он состоит только из одной прямой цепи. | Он состоит из множества взаимосвязанных пищевых цепочек. |
| стабильность | Нестабильность увеличивается из-за увеличения числа отдельных и замкнутых пищевых цепей. | Стабильность увеличивается за счет наличия сложных пищевых цепей. |
| Нарушения возникают из-за | Даже если нарушится одна группа организма, будет затронута вся цепь. | Пищевая сеть не нарушается удалением одной группы организмов. |
| Питаться | Обычно член более высокого трофического уровня зависит или питается одним типом организмов более низкого трофического уровня. | В пищевой сети члены более высокого трофического уровня зависят или питаются многими различными типами организма более низкого трофического уровня. |
| Трофический уровень | Пищевая цепочка состоит всего из 4-6 трофических уровней разных видов. | Пищевая сеть содержит многочисленные трофические уровни и различных популяций видов. |
| Типы | 1.Загрязнение пищевой цепи. 2. Пищевая цепь детрита. | Нет типа |
Определение пищевой цепи
Поток энергии в одном процессе и последовательность, в которой энергия от нижнего уровня организмов к более высокому уровню организмов называется пищевой цепью. Пищевая цепочка бывает двух видов — пастбищная пищевая цепочка и пищевая цепочка Detritus.
1. Пищевая цепь выпаса: Пищевая цепь выпаса состоит в основном из Производителя (зеленые растения), Первичного потребителя (Травоядных), Вторичного потребителя (Плотоядных). Цепочка начинается с зеленых растений, также известных как автотрофы. Они могут готовить пищу с помощью солнечного света (основного источника энергии), воды и воздуха.
Они могут готовить пищу с помощью солнечного света (основного источника энергии), воды и воздуха.
Это известно как первый трофический уровень. Эти растения съедаются первым потребителем (травоядными), и в конечном итоге эти (травоядные) съедаются другим вторичным потребителем (плотоядными).
2. Пищевая цепь детрита. Она начинается с мертвого органического вещества (детрита), которое образовалось в пищевой цепочке. Отходы, такие как трупы животных или опавшие листья, съедаются разложителями или детритиворами. Наконец органическое вещество разлагается их хищниками. Энергия течет в большом количестве в этой пищевой цепи.
ПРИМЕР :
Солнце дает энергию для зеленых растений или травы.
Эту ТРАВУ съедает КУБОК.
Далее, ТРАВЯНОЙ МАШИНЕ съедает Лягушка.
Лягушка съедена ЗМЕЕЙ.
Ястреб ест змею.
Определение пищевой сети
Когда несколько пищевых цепочек связаны или соединены вместе, образуя сеть, это называется пищевой сетью. Существует участие различных. Организмы разных видов популяции. Во всем есть одна общая вещь — потребность в энергии, чтобы выполнять свою деятельность. Солнце считается основным источником энергии на Земле. Эта энергия используется зелеными растениями (производителем) для производства пищи. Как только энергия была захвачена, она теперь пройдет через многие фазы различных организмов определенной области, это называется пищевой сетью.
Существует участие различных. Организмы разных видов популяции. Во всем есть одна общая вещь — потребность в энергии, чтобы выполнять свою деятельность. Солнце считается основным источником энергии на Земле. Эта энергия используется зелеными растениями (производителем) для производства пищи. Как только энергия была захвачена, она теперь пройдет через многие фазы различных организмов определенной области, это называется пищевой сетью.
Ключевые различия между пищевой цепью и пищевой сетью
Ниже приводятся важные моменты, которые отличают Food Chain и Food Web:
- Пищевая цепь может быть названа как единственный прямой путь, по которому идет поток энергии от нижнего трофического уровня к более высокому трофическому уровню. Пищевая сеть может быть определена как сложная взаимосвязь многочисленных пищевых цепей, через которые поток энергии в экосистеме.
- Пищевая цепь состоит только из одной прямой цепи, в то время как пищевая сеть имеет множество взаимосвязанных пищевых цепей .

- По сравнению с пищевой сетью в пищевой цепи существует большая нестабильность, и это связано с увеличением числа отдельных и замкнутых пищевых цепей. Принимая во внимание, что в пищевой сети есть стабильность, и она увеличивается из-за наличия сложных пищевых цепей.
- Как и в пищевой цепи, существует 4-6 трофических уровней только разных видов, и любое нарушение на любом уровне может нарушить всю цепь. С другой стороны, в пищевой сети участвуют многочисленные трофические уровни различных популяций вида, и поэтому она не влияет на пищевую сеть, если происходит удаление какой-либо группы организмов на любом трофическом уровне.
- В пищевой цепи обычно член с более высоким трофическим уровнем зависит или питается одним типом организмов с более низким трофическим уровнем.
Напротив, в пищевой сети представители более высокого трофического уровня зависят или питаются многими различными типами организма более низкого трофического уровня.


Вывод
Все формы жизни нуждаются в энергии, прямо или косвенно. Поток энергии и питательных веществ между различными организмами представляет собой экосистему, называемую пищевой цепью, тогда как естественное связывание или взаимосвязь многочисленных пищевых цепочек того, что ест, и того, что в сообществе, называется пищевой сетью.
Но наблюдается, что пищевая сеть более реалистична, чем пищевая цепь, поскольку она показывает множественные взаимосвязи того, как взаимодействия происходят между организмами. Поэтому пищевая сеть считается более сложной, чем пищевая цепочка.
Ответы | Практ. 2. Решение задач по теме «Цепи и сети питания» — Биология, 11 класс
1.
Последовательность организмов, в которой происходит поэтапный перенос вещества и энергии от источника (предыдущего звена) к потребителю (последующему звену), называется трофической цепью. Совокупность организмов, которые в зависимости от способа их питания и вида корма составляют определенное звено пищевой цепи, называется трофическим уровнем.
2.
б) трава → кузнечик → лягушка → уж;
3.
Дубы → гусеницы → синицы → коршуны.
4.
а) аскарида; б) волк; в) гнилостная бактерия; г) дятел; д) жук-навозник; е) жук-олень; ж) ель; з) клещ; и) подосиновик; к) сосна; л) сосновая пяденица; м) хлорелла; н) ястреб.
- Продуценты: ж, к, м;
- Редуценты: в, д, з;
- Консументы: б, г, з, е, л, и, а;
Составьте цепь питания из пяти трофических уровней, выбрав необходимые организмы из приведенного выше перечня.
Сосна → сосновая пяденица → дятел → ястреб → клещ.
5.
- Чертополох → стрекоза → лягушка → уж → змееяд
- Ель → белка → куница
6.
- а) тимофеевка луговая → бабочка → лягушка → змея;
- б) опавшая листва → червь → грибы → бактерии.
К какому типу относится каждая из цепей питания?
- а) пастбищная цепь питания
- б) детритная цепь питания
7.

Если исключить моллюсков из цепочки, начнут вымирать рыбы, а дафнии неограниченно размножаться. Если исключить инфузории, начнут вымирать дафнии, рыбы и моллюски, а бактерии будут размножаться без сдерживающих факторов.
Присоединяйтесь к Telegram-группе @superresheba_11, делитесь своими решениями и пользуйтесь материалами, которые присылают другие участники группы!Сеть анализа пищевой цепи — ОЭСР
Сеть анализа пищевых цепочек (FCAN) — это экспертная группа ОЭСР, специализирующаяся на анализе пищевых систем. FCAN исследует вопросы, связанные со сбором и анализом данных о продовольственных системах, с целью информирования многостранового диалога и поддержки разработки более эффективных политик.
Сеть анализа пищевых цепей работает с Секретариатом над выработкой общего понимания упрощенных схем маркировки пищевых продуктов В 2021 году FCAN окажет помощь Секретариату ОЭСР в выработке общего понимания упрощенной политики маркировки пищевых продуктов с учетом быстрого распространения этих схем по всему миру. На недавней встрече FCAN, состоявшейся 24 июня 2021 года, эксперты по продовольственной политике стран поделились своим национальным опытом разработки политики в этой области, сосредоточив внимание на том, как разработать согласованную политику, а также на пробелах в знаниях и передовой практике для устранения этих пробелов.
На недавней встрече FCAN, состоявшейся 24 июня 2021 года, эксперты по продовольственной политике стран поделились своим национальным опытом разработки политики в этой области, сосредоточив внимание на том, как разработать согласованную политику, а также на пробелах в знаниях и передовой практике для устранения этих пробелов.
Эксперты предоставили информацию по трем всеобъемлющим вопросам, связанным со схемами:
- Эффективность: Как маркировка влияет на изменение поведения в продовольственных системах? Какая информационная база необходима для измерения этих воздействий?
- Процесс определения, внедрения и мониторинга: Как можно привлечь заинтересованные стороны продовольственных систем и потребителей? Как может быть организовано управление? Какие сопутствующие меры необходимы?
- Потенциальная потребность в гармонизации: Каково потенциальное влияние таких схем на торговлю? Как можно добиться большей согласованности?
Дискуссия FCAN о том, как эффективно внедрить упрощенную политику маркировки продуктов питания, подчеркнула необходимость устранения разрозненности между государственными учреждениями, субъектами частного сектора и потребителями, чтобы обеспечить согласованность политики в рамках всей продовольственной системы.
Использование такого подхода улучшает понимание практических аспектов реализации схем маркировки. Координация также может помочь в устранении пробелов в данных, связанных с измерением эффективности схем изменения поведения потребителей, поскольку разные участники имеют доступ к разным данным. Также важно учитывать последствия для международной торговли.
Для более глубокого изучения трех тем Секретариат разработал анкету, которую 14 стран вызвались заполнить к сентябрю.Консультации с национальными заинтересованными сторонами будут организованы виртуально в октябре и ноябре 2021 года.
FCAN соберется снова 2-3 декабря 2021 года, чтобы обсудить предварительные результаты этого этапа сбора информации.
интегрированных пищевых сетей | Технологический институт цепочек поставок и логистики Джорджии
Существует острая общая потребность как внутренних, так и международных продовольственных цепей воплотить инженерные системы, которые интегрируют поток продуктов питания через различные компоненты цепочек. Большинство пищевых цепочек представляют собой слабосвязанные объекты с ограниченной координацией, несовершенной согласованностью и непоследовательной инфраструктурой и процессами. Результатом являются огромные потери и чрезмерный риск для безопасности в сочетании с потерей продукта и чрезмерными затратами на логистику. Интеграция пищевой цепочки требует, чтобы субъекты в цепочке поставок пищевых продуктов сотрудничали для достижения определенных желаемых результатов. Основой для интеграции является то, что каждый компонент в пищевой цепи должен работать эффективно. Однако для эффективной интеграции компании должны учитывать как свои собственные потребности, так и потребности нижестоящих участников при управлении своей частью холодовой цепи.
Большинство пищевых цепочек представляют собой слабосвязанные объекты с ограниченной координацией, несовершенной согласованностью и непоследовательной инфраструктурой и процессами. Результатом являются огромные потери и чрезмерный риск для безопасности в сочетании с потерей продукта и чрезмерными затратами на логистику. Интеграция пищевой цепочки требует, чтобы субъекты в цепочке поставок пищевых продуктов сотрудничали для достижения определенных желаемых результатов. Основой для интеграции является то, что каждый компонент в пищевой цепи должен работать эффективно. Однако для эффективной интеграции компании должны учитывать как свои собственные потребности, так и потребности нижестоящих участников при управлении своей частью холодовой цепи.
Стремясь выявить и решить критические проблемы интеграции в сквозной пищевой цепи и способствовать эффективной интеграции в пищевой цепи, Институт цепочки поставок и логистики Технологического института Джорджии и Sterling Solutions LLC создали Центр интегрированной пищевой цепи (IFC) как совместные усилия академических, правительственных и промышленных кругов. Основное внимание в исследованиях уделяется разработке, анализу и постоянному совершенствованию холодильных цепей для скоропортящихся пищевых продуктов с целью гарантировать, что производители, переработчики, розничные торговцы и поставщики логистических услуг могут выполнять свои «обещания качества» высокоэффективными способами за счет использования систематических и синхронизированные связи по всей их цепочке поставок/холода.Центр IFC обеспечивает стратегическое руководство и направление сотрудничества между всеми заинтересованными сторонами в пищевой цепочке для понимания проблем и определения их приоритетности; исследовать и определить методологии, процессы и технологии для решения этих проблем; и использовать образование для обеспечения человеческого капитала, необходимого для планирования и реализации исключительных пищевых цепей.
Основное внимание в исследованиях уделяется разработке, анализу и постоянному совершенствованию холодильных цепей для скоропортящихся пищевых продуктов с целью гарантировать, что производители, переработчики, розничные торговцы и поставщики логистических услуг могут выполнять свои «обещания качества» высокоэффективными способами за счет использования систематических и синхронизированные связи по всей их цепочке поставок/холода.Центр IFC обеспечивает стратегическое руководство и направление сотрудничества между всеми заинтересованными сторонами в пищевой цепочке для понимания проблем и определения их приоритетности; исследовать и определить методологии, процессы и технологии для решения этих проблем; и использовать образование для обеспечения человеческого капитала, необходимого для планирования и реализации исключительных пищевых цепей.
Агропродовольственные сети и сети для развития
Нажмите, чтобы рассмотреть
Об этой книге Содержание Отзывы клиентов Связанные названия
Об этой книге
Агропродовольственные цепи и сети играют все более важную роль в обеспечении доступа на рынки для производителей из развивающихся стран. В развивающихся странах компании интегрируются в географически рассредоточенные сети поставок, которые связывают производителей, торговцев и переработчиков с юга с розничными торговцами и потребителями в городских центрах и на севере. Глобализация торговли и интеграция цепочек поставок приводят к новым требованиям в отношении качества, безопасности и здоровья пищевых продуктов.
В развивающихся странах компании интегрируются в географически рассредоточенные сети поставок, которые связывают производителей, торговцев и переработчиков с юга с розничными торговцами и потребителями в городских центрах и на севере. Глобализация торговли и интеграция цепочек поставок приводят к новым требованиям в отношении качества, безопасности и здоровья пищевых продуктов.
Компании, участвующие в глобальных цепочках и сетях продовольственного и сельскохозяйственного бизнеса, сталкиваются с быстрыми изменениями на рынке и в бизнес-среде. Появляются новые процедуры и методы организации сетей снабжения продовольствием, основанные на совместных инновациях между первичными производителями, переработчиками и розничными торговцами, чтобы справиться с качеством и стандартами пищевых продуктов.Оптимизация производительности цепочки требует сотрудничества между всеми участниками цепочки поставок. Агропродовольственные компании доводят требования потребителей до мелких землевладельцев, чтобы гарантировать стабильный доступ к рынку. Государственные и добровольные организации могут внести важный вклад в укрепление среды цепочки поставок.
Государственные и добровольные организации могут внести важный вклад в укрепление среды цепочки поставок.
В этом томе теоретические выводы сочетаются с большим количеством бизнес-кейсов, что позволяет лучше понять возможности и ограничения, которые может предложить интеграция цепочки поставок для стимулирования развития сельских районов.
Содержимое
Предисловие.- Введение и аналитическая основа: 1. Агропродовольственные цепи и сети для развития: вопросы, подходы и стратегии.- Цепная интеграция и развитие: 2. Сельскохозяйственное развитие и либерализация торговли.- 3. Участие мелких землевладельцев в международной торговле. — 4. Сети и сети для развития: определение заинтересованных сторон в международной торговле.- 5. Источники овощей в Африке: опыт Freshmark.- 6. Создание партнерских отношений для создания добавленной стоимости: роль агробизнеса в развитии торговли.- 7. Вклад справедливой торговли в доступ на рынок для мелких производителей бананов. — 8. быстрый рост супермаркетов и использование частных стандартов в их системах закупок пищевых продуктов в развивающихся странах.- 9. Агропродовольственные цепочки и устойчивые средства к существованию: тематическое исследование маркетинга маниоки в Нигерии.- Бизнес-кейсы: 10. Развитие цепочки поставок для свежих фруктов и овощей в Таиланде.- 11. FRUITFUL: интегрированная информационная система цепочки поставок фруктовой продукции между Южной Африкой и Нидерландами.- 12. Brascan: как получить ценность в цепочке производства говядины.- 13. Рыба в Кении: цепочка нильского окуня.- 14 Сеть производства органического какао в целях развития: пример Ассоциации мелких фермеров Таламанки.- 15. Проект Novella: создание устойчивой цепочки поставок масла Allanblackia.- 16. Развитие устойчивой цепочки поставок лекарственных растений в Индии: объединение людей, рынков. и ценности.- Резюме и выводы: 17.Устойчивые агропродовольственные цепочки: проблемы для исследований и разработок.- 18. Исключение мелких фермеров из скоординированных цепочек поставок: сбой рынка, провал политики или просто эффект масштаба.
— 8. быстрый рост супермаркетов и использование частных стандартов в их системах закупок пищевых продуктов в развивающихся странах.- 9. Агропродовольственные цепочки и устойчивые средства к существованию: тематическое исследование маркетинга маниоки в Нигерии.- Бизнес-кейсы: 10. Развитие цепочки поставок для свежих фруктов и овощей в Таиланде.- 11. FRUITFUL: интегрированная информационная система цепочки поставок фруктовой продукции между Южной Африкой и Нидерландами.- 12. Brascan: как получить ценность в цепочке производства говядины.- 13. Рыба в Кении: цепочка нильского окуня.- 14 Сеть производства органического какао в целях развития: пример Ассоциации мелких фермеров Таламанки.- 15. Проект Novella: создание устойчивой цепочки поставок масла Allanblackia.- 16. Развитие устойчивой цепочки поставок лекарственных растений в Индии: объединение людей, рынков. и ценности.- Резюме и выводы: 17.Устойчивые агропродовольственные цепочки: проблемы для исследований и разработок.- 18. Исключение мелких фермеров из скоординированных цепочек поставок: сбой рынка, провал политики или просто эффект масштаба. — 19. Продовольственные цепочки и сети в целях развития: уроки и перспективы .- Вагенингенская декларация
— 19. Продовольственные цепочки и сети в целях развития: уроки и перспективы .- Вагенингенская декларация
Отзывы клиентов
Уязвимость цепочек поставок продуктов питания в Соединенном Королевстве, подверженных воздействию COVID-19
Термин «цепочка поставок» предполагает линейную цепочку поставщиков, где одна организация поставляет ресурсы или материалы другой.Однако, в частности, применительно к современным цепочкам поставок более точно рассматривать сети множественных и двунаправленных взаимозависимостей между организациями (узлами в сети) 9 .
Сложные сети цепочек поставок также можно рассматривать как по существу открытые системы, а не как изолированные системы с четко определенными границами. Неограниченная взаимозависимость между организациями в цепочке поставок бросает вызов теоретическому пониманию того, как работают цепочки поставок, поскольку трудно понять концептуально или вычислительно, какими могут быть последствия этой связанности 10 , особенно с точки зрения устойчивости к системным потрясениям.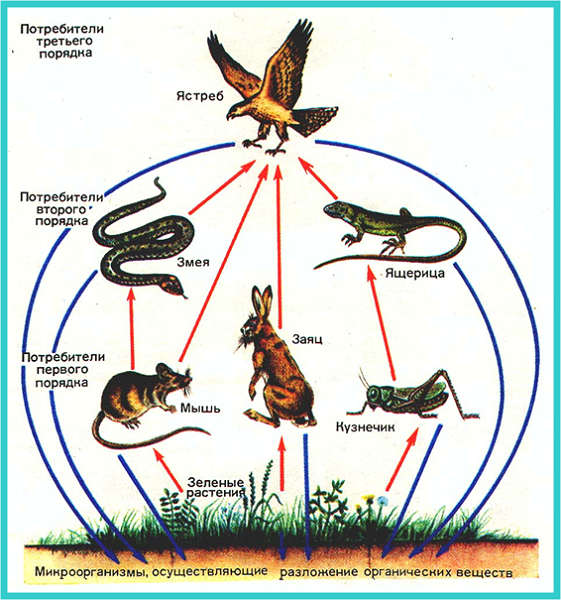
В ситуациях, когда цепочки поставок работают нормально, поток материалов и ресурсов может быть близок к беспрепятственному 11 , а в сложных сетях цепочек поставок станет максимально оптимизированным для максимизации потока ресурсов через систему. Эта оптимизация может привести к небольшой избыточности или ее полному отсутствию на любом узле сети. Хотя эти условия часто рассматриваются как благоприятные с точки зрения эффективности, сбой в одной части сети может распространяться по всей (сложной) сети, подвергая цепочку поставок волновому эффекту 12 .
Сильно связанные сети могут выйти из строя по разным причинам, некоторые из которых могут быть следствием самих взаимозависимостей 13 . Сбой в сложных сетях цепочек поставок может также проявляться как форма тупика, когда все части сети останавливаются в ожидании ресурсов, а сложные взаимозависимости между частями (организациями и процессами в цепочках поставок) сети делают выход из тупика крайне сложно (рис. 1).Это проблема, которую часто можно наблюдать в вычислительных системах, таких как параллельные системы, где процессы могут зайти в тупик, ожидая друг друга 14 .
1).Это проблема, которую часто можно наблюдать в вычислительных системах, таких как параллельные системы, где процессы могут зайти в тупик, ожидая друг друга 14 .
Все части системы ждут друг друга. Ситуация, которая не может быть разрешена без внешнего вмешательства, чтобы выйти из тупика.
В цепочках поставок продуктов питания характер зависимости между организациями и процессами различается. Некоторые цепочки поставок продуктов питания имеют высокую степень вертикальной координации 15 , чтобы как можно быстрее перемещать продукт от источника к потребителю, хотя они по-прежнему работают внутри более крупной организационной сети и зависят от нее.Многие продукты питания поставляются по всему миру (а не сосредоточены в каком-либо одном регионе или стране), и, даже если один продукт вертикально скоординирован, организации в разных странах зависят от нескольких хорошо скоординированных глобальных цепочек поставок. Эти цепочки и сети уязвимы для сбоев в более широкой сети цепочек поставок из-за потери доступа к ресурсам или другим вспомогательным функциям.
Эти цепочки и сети уязвимы для сбоев в более широкой сети цепочек поставок из-за потери доступа к ресурсам или другим вспомогательным функциям.
Цепочки поставок продовольствия полагаются на высокую степень временной координации, основанную на принципах своевременной доставки для работы вертикально скоординированных цепочек от производителя к потребителю.Системы розничных заказов JiT для свежих фруктов и овощей из ЕС работают по алгоритмам, созданным системой электронной торговой точки (EPOS) в магазине. Как правило, это создает заказ утром, который затем размещается в штаб-квартире супермаркета, например, в Великобритании, а затем до 12 часов того же дня передается одному из их сторонних поставщиков в ЕС27, например, в Испании. Рабочие, используемые для сбора и упаковки продукта, часто передаются по субподряду. Затем супермаркет ожидает доставку этого заказа в какой-то момент на следующий день.Это называется «система первого дня для второго дня» 8 . Эффективность этой системы в значительной степени зависит от беспрепятственной торговли, особенно в контексте ЕС. Таким образом, сети цепочки поставок продуктов питания также уязвимы во времени — как с точки зрения скорости, с которой супермаркеты полагаются на доставку продуктов для пополнения полок, так и с точки зрения блокировок в цепочке поставок, которые могут сделать продукт непригодным для использования или несъедобным (или и тем, и другим).
Таким образом, сети цепочки поставок продуктов питания также уязвимы во времени — как с точки зрения скорости, с которой супермаркеты полагаются на доставку продуктов для пополнения полок, так и с точки зрения блокировок в цепочке поставок, которые могут сделать продукт непригодным для использования или несъедобным (или и тем, и другим).
Цепочки поставок также не являются единственным источником системной уязвимости продовольственных систем.Изменение климата оказывает и будет оказывать в будущем огромное влияние на производство и доступность продуктов питания, влияя на предложение, место производства и повышая риски, связанные с экстремальными погодными условиями, как в Великобритании 16 , так и во всем мире 17,18 . Политические сбои, такие как Brexit в Великобритании 2 , также могут выявить уязвимые места в системах, например, зависимость от небольшого числа поставщиков (см. ниже). Однако политические риски влияют не только на предложение, но и на другие области, такие как цены на продукты питания 2 . Поэтому стоит подумать о том, как крупномасштабные системные потрясения, такие как COVID-19, могут накладываться друг на друга и вызывать потрясения в других частях системы, потенциально обнажая множественные одновременные системные уязвимости и вызывая возмущения в продовольственной системе 19 .
Поэтому стоит подумать о том, как крупномасштабные системные потрясения, такие как COVID-19, могут накладываться друг на друга и вызывать потрясения в других частях системы, потенциально обнажая множественные одновременные системные уязвимости и вызывая возмущения в продовольственной системе 19 .
Модели экологических сетей связывают разнообразие, структуру и функции пищевой сети планктона
Хатчинсон, Г. Е. Парадокс планктона. Являюсь. Нац. 95, 137–145 (1961).
Google Scholar
Лоро, М.От загрязнений к экосистемам: теоретические основы нового экологического синтеза. моногр. Народ. биол. (Издательство Принстонского университета, 2010 г.).
Червь, Б. и др. Воздействие утраты биоразнообразия на экосистемные услуги океана. Наука 314, 787–90 (2006).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Хаттон, И. а. и другие. Закон силы хищник-жертва: масштабирование биомассы в наземных и водных биомах. Наука (80-.). 349, (2015).
Закон силы хищник-жертва: масштабирование биомассы в наземных и водных биомах. Наука (80-.). 349, (2015).
Bascompte, J., Melián, C.J. & Sala, E. Комбинации силы взаимодействия и перелов. проц. Натл. акад. науч. США 102, 5443–5447 (2005).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Бойт, А., Мартинес, Н.Д., Уильямс, Р.Дж. и Гаедке, У. Механистическая теория и моделирование сложной динамики пищевых сетей в Боденском озере. Экол. лат. 15, 594–602 (2012).
Google Scholar
Хейрстон, Н.Г. Дж. и Хейрстон, Н. Г. С. Причинно-следственные связи в потоке энергии, трофической структуре и межвидовых взаимодействиях. Являюсь. Нац. 142, 379–411 (1993).
Google Scholar
Thompson, R. M. et al. Пищевые сети: согласование структуры и функции биоразнообразия. Тенденции Экол. Эвол. 27, 689–97 (2012).
Google Scholar
де Варгас, К. и др. Разнообразие эукариотического планктона в освещенном солнцем океане.Наука (80-х гг.). 348, 1–12 (2015).
и др. Разнообразие эукариотического планктона в освещенном солнцем океане.Наука (80-х гг.). 348, 1–12 (2015).
Google Scholar
Д’Алелио, Д. и др. Время для секса: двухгодичный жизненный цикл морских планктонных диатомей. Лимнол. океаногр. 55, 106–114 (2010).
ОБЪЯВЛЕНИЕ Google Scholar
Мандала Вятта Т. Маргалефа и стратегии цветения фитопланктона. Глубокое море Res. Часть II Верх. Стад. океаногр. 1–18 (2012).
Маццокки, М.Г., Ликандро П., Дуброка Л., Ди Капуа И. и Саджиомо В. Ассоциации зоопланктона в долгосрочных временных рядах Средиземноморья. Дж. Планктон Рез. 33, 1163–1181 (2011).
Google Scholar
Лима-Мендез, Г. и др. Детерминанты структуры сообщества в глобальном интерактоме планктона. Наука (80-х гг.). 348, 1–10 (2015).
Google Scholar
Steele, J. A. et al.Ассоциативные сети морских бактерий, архей и протистов выявляют экологические связи. ISME J. 5, 1414–1425 (2011).
A. et al.Ассоциативные сети морских бактерий, архей и протистов выявляют экологические связи. ISME J. 5, 1414–1425 (2011).
ПабМед ПабМед Центральный Google Scholar
Митра, А. и др. Роль миксотрофных протистов в биологическом углеродном насосе. Биогеонауки 11, 995–1005 (2014).
ОБЪЯВЛЕНИЕ Google Scholar
Беренфельд М.Дж. и Босс Э.С.Возрождение экологических основ цветения океанического планктона. Аня. Преподобный Мар. 1–28 (2013).
Беренфельд, М. Дж., Босс, Э., Сигел, Д. а. & Shea & DM. Продуктивность океана на основе углерода и физиология фитопланктона из космоса. Глобальная биогеохимия. Циклы 19, 1–14 (2005).
Google Scholar
Митра, А. и др. Преодоление разрыва между морскими биогеохимическими науками и науками о рыболовстве; настройка ссылки на зоопланктон.прог.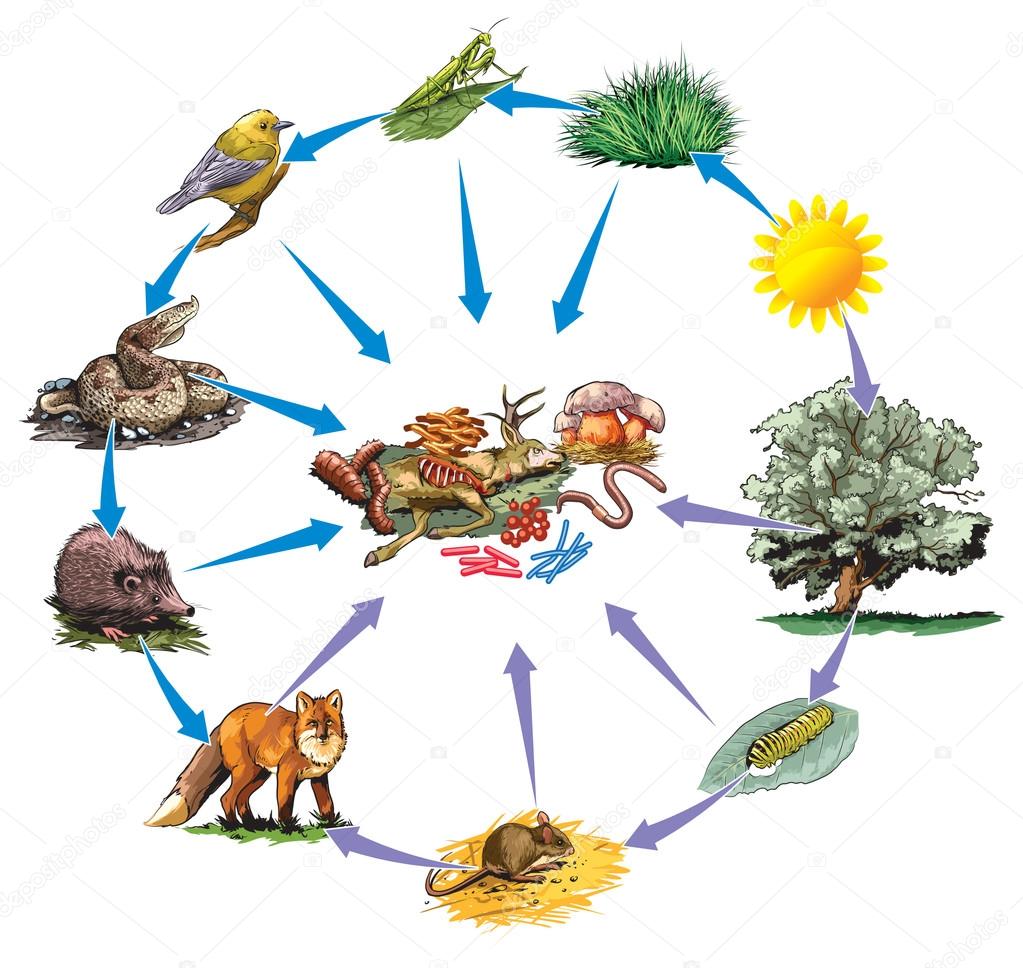 океаногр. 129, 176–199 (2014).
океаногр. 129, 176–199 (2014).
ОБЪЯВЛЕНИЕ Google Scholar
Franks, PJS Модели динамики планктона NPZ: их построение, связь с физикой и применение. Ж. океаногр. 58, 379–387 (2002).
Google Scholar
Личман Э., Оман М. Д. и Киорбо Т. Подходы к сообществам зоопланктона на основе признаков. Дж. Планктон Рез. 35, 473–484 (2013).
Google Scholar
Флинн, К. Дж. и др. Неправильное использование дихотомии фитопланктон-зоопланктон: необходимость относить организмы к миксотрофам внутри функциональных типов планктона. Дж. Планктон Рез. 35, 3–11 (2012).
Google Scholar
Стокер, Д. К., Джонсон, М. Д., Варгас, К. де и Нот, Ф. Приобретенная фототрофия у водных простейших. 1–10 (2009).
Шмокер, К., Эрнандес-Леон, С. и Калбет, А. Микрозоопланктон, пасущийся в океанах: воздействие, изменчивость данных, пробелы в знаниях и будущие направления. Дж. Планктон Рез. 35, 691–706 (2013).
Дж. Планктон Рез. 35, 691–706 (2013).
Google Scholar
Иригойен, X. Цветение фитопланктона: «лазейка» в воздействии выпаса микрозоопланктона? Дж. Планктон Рез. 27, 313–321 (2005).
Google Scholar
Боун, К. и другие.Биология пелагических оболочников. (Издательство Оксфордского университета, Оксфорд, 1998).
Кальбет А., Карлотти Ф. и Гауди Р. Экология питания копепод Centropages typicus (Kröyer). прог. океаногр. 72, 137–150 (2007).
ОБЪЯВЛЕНИЕ Google Scholar
Паффенхофер, Г.-А., Маццокки, М. Г. и Ценг, М. В. Жизнь на краю: питание субтропических веслоногих ракообразных открытого океана. Мар. Экол. 27, 99–108 (2006).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Дуро, А. и Саиз, Э. Распределение и трофическая экология хетогнат в западной части Средиземного моря в зависимости от прибрежного и прибрежного градиента. Дж. Планктон Рез. 22, 339–361 (2000).
Дж. Планктон Рез. 22, 339–361 (2000).
Google Scholar
Уланович, Р. Э. О важности моделей более высокого уровня в экологии. Экол. Модель. 43, 45–56 (1988).
Google Scholar
Фатх Б.Д., Шарлер, У. М., Уланович, Р. Э. и Хэннон, Б. Анализ экологических сетей: построение сетей. Экол. Модель. 208, 49–55 (2007).
Google Scholar
Зорах, А. К. и Уланович, Р. Э. Количественная оценка сложности сетей потоков: сколько существует ролей? Сложность 8, 68–76 (2003).
MathSciNet Google Scholar
Абарка-Аренас, Л. Г. и Уланович, Р.E. Влияние таксономической агрегации на сетевой анализ. Экол. Модель. 149, 285–296 (2002).
Google Scholar
Уланович, Р. Э. Количественные методы анализа экологических сетей. вычисл. биол. хим. 28, 321–39 (2004).
вычисл. биол. хим. 28, 321–39 (2004).
КАС МАТЕМАТИКА Google Scholar
Кристенсен В. и Уолтерс С. Дж. Ecopath с Ecosim: методы, возможности и ограничения.Экол. Модель. 172, 109–139 (2004).
Google Scholar
Колл М., Либралато С., Тудела С., Паломера И. и Пранови Ф. Перелов экосистемы в океане. PLoS One 3 (2008 г.).
Рибера д’Алькала, М. и др. Сезонные закономерности в сообществах планктона в многолетних временных рядах на прибрежном участке Средиземного моря (Неаполитанский залив): попытка выявить повторения и тенденции. науч. 68 марта, 65–83 (2004).
Google Scholar
Д’Алелио, Д. и др. Зелено-синее колебание: пластичность пищевых сетей планктона в ответ на прибрежную океанографическую динамику. Мар. Экол. 36, 1155–1170 (2015).
ОБЪЯВЛЕНИЕ Google Scholar
Crawford, D. W. Mesodinium rubrum : фитопланктер, которого не было. Мар. Экол. прог. сер. 58, 161–174 (1989).
W. Mesodinium rubrum : фитопланктер, которого не было. Мар. Экол. прог. сер. 58, 161–174 (1989).
ОБЪЯВЛЕНИЕ Google Scholar
Стич, Х.-Б. & Lampert, W. Уклонение от хищников как объяснение суточной вертикальной миграции зоопланктона. Природа 293, 396–398 (1981).
ОБЪЯВЛЕНИЕ Google Scholar
Пауэр, М. Э., Тилман, Д., Эстес, Дж. А., Менге, Б. А. и Бонд, В. Дж. Проблемы в поисках краеугольных камней. Биологические науки 46, 609 (1996).
Google Scholar
Либралато С., Кристенсен В. и Поли Д.Метод определения ключевых видов в моделях пищевых сетей. Экол. Модель. 195, 153–171 (2006).
Google Scholar
Уланович, Р. Э. и Норден, Дж. С. Симметричные накладные расходы в потоковых сетях. Междунар. Дж. Сист. науч. 21, 429–437 (1990).
МАТЕМАТИКА Google Scholar
Уланович, Р. Э. Экология, восходящая перспектива. (Издательство Колумбийского университета, 1997).
Линдеман Р.Л. Трофодинамический аспект экологии. Экология 23, 399–417 (1942).
Google Scholar
Либралато, С. и др. Особенности пищевой сети охраняемых и эксплуатируемых районов Адриатического моря. биол. Консерв. 143, 2182–2194 (2010).
Google Scholar
Saiz, E. & Calbet, A. Питание веслоногих ракообразных в океане: масштабирование, состав их рациона и погрешность оценок из-за выпаса микрозоопланктона во время инкубации.Гидробиология 666, 181–196 (2010).
Google Scholar
Saiz, E. & Calbet, A. Масштабирование питания морских каланоидных копепод. Лимнол. океаногр. 52, 668–675 (2007).
ОБЪЯВЛЕНИЕ Google Scholar
Mazzocchi, MG, Dubroca, L. , García-Comas, C., Capua, I. Di & Ribera d’Alcalà, M. Стабильность и устойчивость прибрежных скоплений веслоногих ракообразных: случай долговременной экологической исследования на Станции MC (LTER-MC).прог. океаногр. 97–100, 135–151 (2012).
, García-Comas, C., Capua, I. Di & Ribera d’Alcalà, M. Стабильность и устойчивость прибрежных скоплений веслоногих ракообразных: случай долговременной экологической исследования на Станции MC (LTER-MC).прог. океаногр. 97–100, 135–151 (2012).
ОБЪЯВЛЕНИЕ Google Scholar
Хартманн, М. и др. Миксотрофная основа атлантических олиготрофных экосистем. проц. Натл. акад. науч. США 109, 5756–5760 (2012).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Ландри, М. и Калбет, А. Производство микрозоопланктона в океанах. ICES J. Mar. Sci. 61, 501–507 (2004).
Google Scholar
Шейнберг Р., Лэндри, М. и Калбет, А. Выпас двух обыкновенных аппендикулярий на естественной добыче тропической прибрежной экосистемы. Мар. Экол. прог. сер. 294, 201–212 (2005).
ОБЪЯВЛЕНИЕ Google Scholar
Сан-Мартин, Э.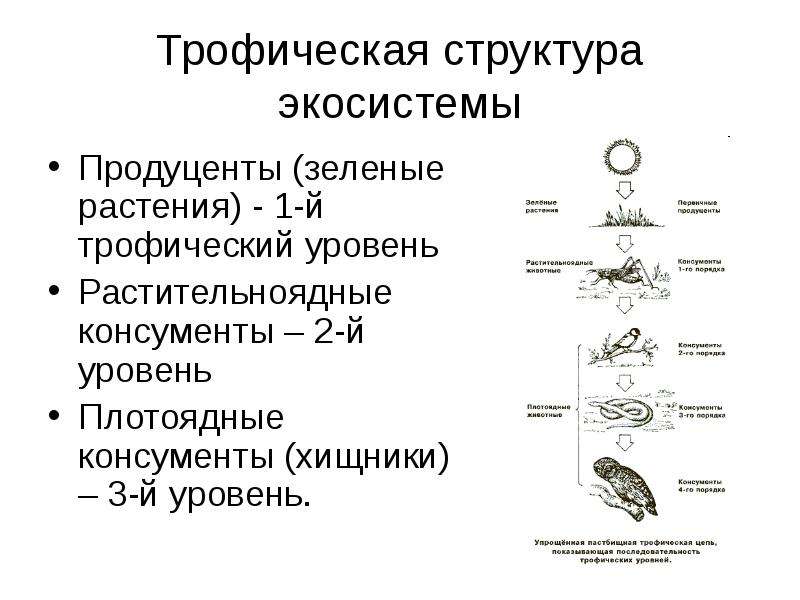 и др. Изменение переноса энергии в морском планктоне по градиенту продуктивности в Атлантическом океане. Лимнол. океаногр. 51, 2084–2091 (2006).
и др. Изменение переноса энергии в морском планктоне по градиенту продуктивности в Атлантическом океане. Лимнол. океаногр. 51, 2084–2091 (2006).
ОБЪЯВЛЕНИЕ Google Scholar
Поли Д.и Кристенсен, В. Первичное производство, необходимое для поддержания глобального рыболовства. Природа 374, 255–257 (1995).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Одум, Э.П., Одум, Х.Т. и Эндрюс, Дж. Основы экологии. 3 (Сондерс, Филадельфия, 1971).
Либралато, С., Колл, М., Тудела, С., Паломера, И. и Пранови, Ф. Новый индекс для количественной оценки воздействия рыболовства на экосистему как удаление вторичной продукции.Мар. Экол. прог. сер. 355, 107–129 (2008).
ОБЪЯВЛЕНИЕ Google Scholar
Heymans, J.J. et al. Глобальные закономерности экологических показателей морских пищевых сетей: подход к моделированию.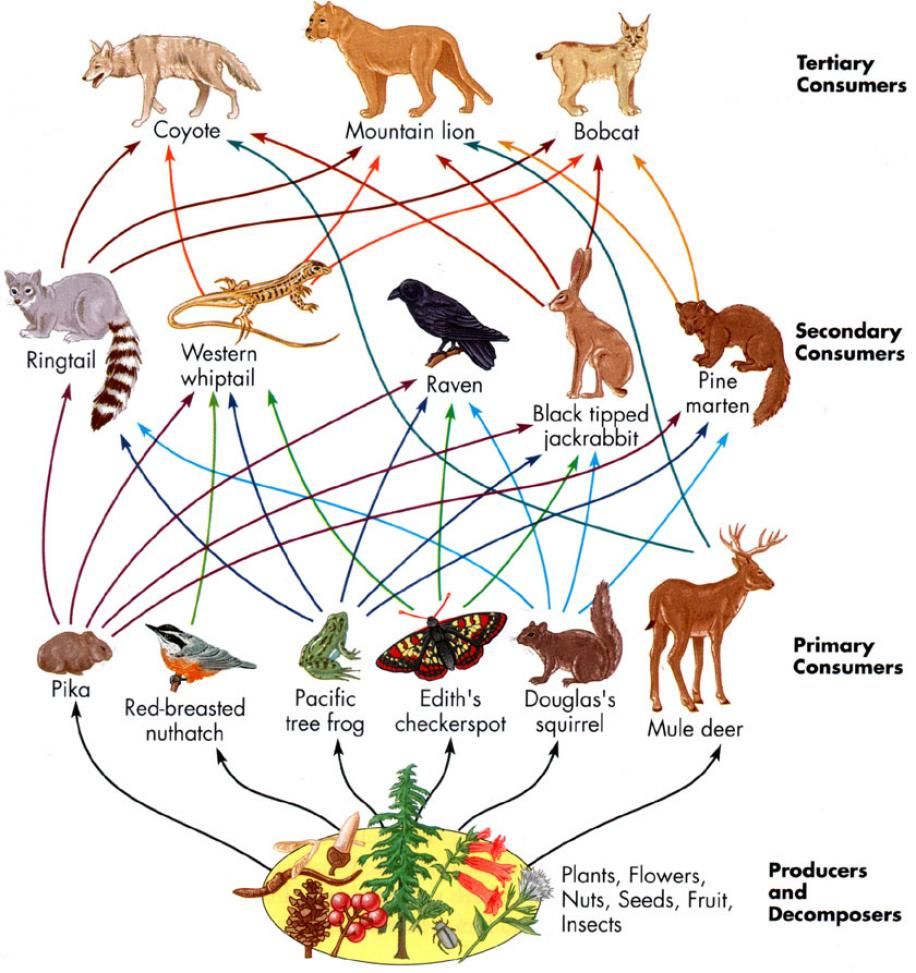 PLoS One 9, e95845 (2014 г.).
PLoS One 9, e95845 (2014 г.).
ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Google Scholar
Берглунд Дж., Мюрен У., Бамштедт У. и Андерссон А. Эффективность пищевой сети на основе фитопланктона и бактерий в пелагической морской системе.Лимнол. океаногр. 52, 121–131 (2007).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Gaedke, U. & Straile, D. Сезонные изменения эффективности трофического переноса в пищевой цепи планктона, полученные на основе распределения размеров биомассы и сетевого анализа. Экол. Модель. 75–76, 435–445 (1994).
Google Scholar
Franzé, G. & Modigh, M. Экспериментальные доказательства внутреннего хищничества в сообществах микрозоопланктона.Мар биол. 160, 3103–3112 (2013).
Google Scholar
СТЕКФ. Научно-технический и экономический комитет по рыболовству (STECF) – Оценка запасов Средиземного моря – часть 1 (STECF-14-17, 2014).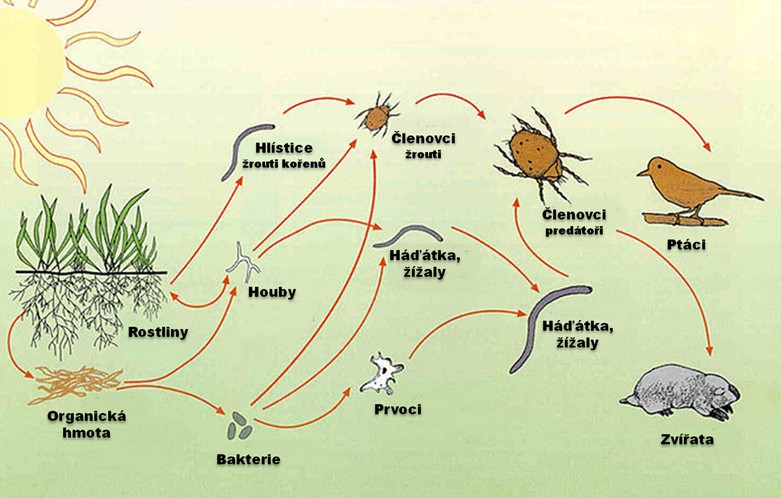 Бюро публикаций Европейского Союза, Люксембург, EUR 26955 EN, JRC 93120 . (2014).
Бюро публикаций Европейского Союза, Люксембург, EUR 26955 EN, JRC 93120 . (2014).
Линк, Дж. С. и др. Эмерджентные свойства очерчивают возмущение и восстановление морской экосистемы. Тенденции Экол.Эвол. 30, 649–661 (2015).
Google Scholar
Lazzari, P. et al. Сезонная и межгодовая изменчивость хлорофилла и первичной продукции планктона в Средиземном море: подход к моделированию. Биогеонауки 9, 217–233 (2012).
КАС ОБЪЯВЛЕНИЯ Google Scholar
ван дер Линген, К., Хатчингс, Л. и Филд, Дж. Сравнительная трофодинамика анчоуса Engraulis encrasicolus и сардины Sardinops sagax в южной части Бенгелы. Африканский Дж.мар. 28, 465–477 (2006).
Google Scholar
Hairston, N.G.J. & Hairston, N.G.S. Устраняет ли сложность пищевой сети динамику трофического уровня? Являюсь.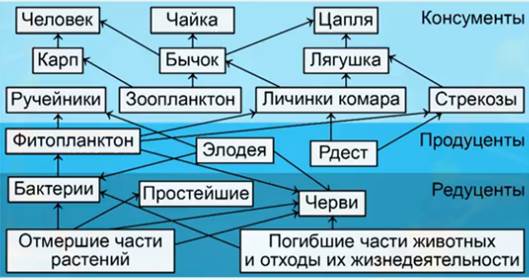 Нац. 149, 1001–1007 (1997).
Нац. 149, 1001–1007 (1997).
Google Scholar
Лефчек, Дж. С. и др. Биоразнообразие повышает многофункциональность экосистем на всех трофических уровнях и средах обитания. Нац. коммун. 6, 1–7 (2015).
Google Scholar
Леман, К.Л. и Тилман Д. Биоразнообразие, стабильность и продуктивность в конкурентных сообществах. Являюсь. Нац. 156, 534–552 (2000).
Google Scholar
Вейтч, Дж., Пенвен, П. и Шиллингтон, Ф. Бенгела: лаборатория для исследований сравнительного моделирования. прог. океаногр. 83, 296–302 (2009).
ОБЪЯВЛЕНИЕ Google Scholar
Шурин Дж. Б., Грунер Д. С. и Хиллебранд Х.Весь мокрый или высохший? Реальные различия между водными и наземными пищевыми сетями. проц. Р. Соц. В 273, 1–9 (2006).
Google Scholar
Вандер Занден, М. Дж. и Фетцер, В. В. Глобальные закономерности длины пищевой цепи в воде. Ойкос 116, 1378–1389 (2007).
Дж. и Фетцер, В. В. Глобальные закономерности длины пищевой цепи в воде. Ойкос 116, 1378–1389 (2007).
Google Scholar
Райтер, Дж. Х. Фотосинтез и производство рыбы в море. Наука (80-х гг.). 166, 72–76 (1969).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Ломбард, Ф. и др. Экспериментальные и модельные доказательства аппендикулярно-инфузорных взаимодействий. Лимнол. океаногр. 55, 77–90 (2010).
ОБЪЯВЛЕНИЕ Google Scholar
Клерн, Дж. Э. Наша эволюционирующая концептуальная модель проблемы прибрежной/ревтрофикации. Мар. Экол. прог. сер. 210, 223–253 (2001).
КАС ОБЪЯВЛЕНИЯ Google Scholar
Чуст, Г.и другие. Изменения биомассы и трофическое усиление планктона в более теплом океане. Глоб. Чанг. биол. 20, 2124–39 (2014).
ОБЪЯВЛЕНИЕ Google Scholar
Либралато С. , Каччин А. и Пранови Ф. Моделирование инвазий видов с использованием динамики термических и трофических ниш в условиях изменения климата. Фронт. мар. 2, 29 (2015).
, Каччин А. и Пранови Ф. Моделирование инвазий видов с использованием динамики термических и трофических ниш в условиях изменения климата. Фронт. мар. 2, 29 (2015).
Google Scholar
Кавана, П., Ньюлендс, Н., Кристенсен В. и Поли Д. Автоматическая оптимизация параметров для моделей экосистемы Ecopath. Экол. Модель. 172, 141–149 (2004).
Google Scholar
Повышение устойчивости сети по отношению к большим пищевым сетям
Резюме
Стабильность является ключевым свойством сложных пищевых сетей, которое долгое время находилось в центре внимания исследований. Оставался интригующим вопрос, насколько сохраняются большие и сложные пищевые сети, если более мелкие и простые имеют тенденцию быть более стабильными, по крайней мере, с математической точки зрения.Предполагая, что с увеличением размеров пищевых сетей растет и их устойчивость, мы проанализировали зависимость между числом узлов в пищевых сетях и их устойчивостью на основе 450 пищевых сетей с числом узлов от нескольких до 200. Наши результаты показывают, что стабильность увеличивается нелинейно с размером пищевой сети, основываясь как на времени возврата после нарушения, так и на устойчивости, рассчитанной на основе скорости вторичного вымирания более высоких трофических уровней. В качестве методологического новшества мы учитывали время образования пищевой сети в процессе расчета времени возврата.Наши результаты способствуют объяснению устойчивости больших и сложных пищевых сетей: несмотря на то, что с увеличением количества видов устойчивость пищевых сетей снижается при малых числах узлов, наблюдается постоянное повышение устойчивости на большом интервале увеличения размера пищевой сети. . Поэтому в исследованиях стабильности пищевой сети мы подчеркиваем использование времени генерации пищевой сети.
Наши результаты показывают, что стабильность увеличивается нелинейно с размером пищевой сети, основываясь как на времени возврата после нарушения, так и на устойчивости, рассчитанной на основе скорости вторичного вымирания более высоких трофических уровней. В качестве методологического новшества мы учитывали время образования пищевой сети в процессе расчета времени возврата.Наши результаты способствуют объяснению устойчивости больших и сложных пищевых сетей: несмотря на то, что с увеличением количества видов устойчивость пищевых сетей снижается при малых числах узлов, наблюдается постоянное повышение устойчивости на большом интервале увеличения размера пищевой сети. . Поэтому в исследованиях стабильности пищевой сети мы подчеркиваем использование времени генерации пищевой сети.
Введение
В последние десятилетия значительно возросло значение экологических сетей 1,2 .С одной стороны, благодаря научным достижениям в теории сетей 3 , а с другой — из-за возросшей потребности в понимании закономерностей сохранения живых сообществ 4 . Из-за значительного сокращения биологического разнообразия и угрозы дальнейшего ускорения этого сокращения мотивация изучения устойчивости и устойчивости экологических сетей приобрела большее значение, чем когда-либо 5 .
Из-за значительного сокращения биологического разнообразия и угрозы дальнейшего ускорения этого сокращения мотивация изучения устойчивости и устойчивости экологических сетей приобрела большее значение, чем когда-либо 5 .
Уменьшение биологического разнообразия вызвано несколькими причинами, такими как деятельность человека в результате утраты и фрагментации среды обитания и изменение климата, ускоренное крупномасштабной деятельностью человека 6 .Климатические изменения вызовут сдвиги ареалов для многих видов, поэтому целые сообщества столкнутся с вымиранием и внедрением 7 . Эти смещающиеся виды и сообщества изменят экосистемы, вызывая либо сокращение, либо рост, тогда как в некоторых случаях это может привести к коллапсу сети 8 .
Из-за коллапса и вымирания целых сообществ характеристики стабильности, такие как устойчивость и устойчивость экологических сетей, стали обычным предметом изучения 9 .Стойкость описывает время, в течение которого сообщества будут выживать в изменяющихся условиях окружающей среды 10 . Устойчивость описывает время, необходимое системе, в данном случае сообществу, для восстановления стабильной структуры, подобной той, которая существовала до возмущений 10 . Стабильность экологических сетей вызвала интенсивные и продолжительные дебаты, известные как дебаты о сложности и стабильности 11–13 . Основной вопрос дискуссии: каким образом могут сохраняться сложные экологические сети, в то время как редуцированные сети кажутся более стабильными 14,15 ? Дебаты о сложности и стабильности преобладали в течение последних пяти десятилетий, предположительно не имея простого решения из-за своей сложности.Предположительно может существовать несколько хороших разрешений 9,16,17 .
Устойчивость описывает время, необходимое системе, в данном случае сообществу, для восстановления стабильной структуры, подобной той, которая существовала до возмущений 10 . Стабильность экологических сетей вызвала интенсивные и продолжительные дебаты, известные как дебаты о сложности и стабильности 11–13 . Основной вопрос дискуссии: каким образом могут сохраняться сложные экологические сети, в то время как редуцированные сети кажутся более стабильными 14,15 ? Дебаты о сложности и стабильности преобладали в течение последних пяти десятилетий, предположительно не имея простого решения из-за своей сложности.Предположительно может существовать несколько хороших разрешений 9,16,17 .
Теоретически прогнозируется, что сложные экологические сети будут более хрупкими, чем простые, поскольку сложность влияет на вероятность того, что виды будут находиться в стабильном равновесии 18 . Более того, изменчивость плотности населения во времени также влияет на постоянство этого состава сообщества 18 . Но эта взаимосвязь между сложностью и хрупкостью в некоторых случаях кажется непостоянной, поэтому отсутствует общее консенсусное соглашение 9 .Пространственность может иметь более выраженный стабилизирующий эффект на сложные пищевые сети, чем это было предсказано на основе экологической теории 16,19 . Более того, в эмпирических пищевых сетях, по-видимому, отсутствует связь стабильности со сложностью, поскольку эмпирические пищевые сети обладают неслучайными свойствами 17 .
Но эта взаимосвязь между сложностью и хрупкостью в некоторых случаях кажется непостоянной, поэтому отсутствует общее консенсусное соглашение 9 .Пространственность может иметь более выраженный стабилизирующий эффект на сложные пищевые сети, чем это было предсказано на основе экологической теории 16,19 . Более того, в эмпирических пищевых сетях, по-видимому, отсутствует связь стабильности со сложностью, поскольку эмпирические пищевые сети обладают неслучайными свойствами 17 .
Однако до 1970-х годов экологи поддерживали идею о том, что сложные сообщества более устойчивы, чем простые, учитывая, что естественные сообщества развиваются в устойчивые системы посредством сукцессионной динамики 9 .Затем работа Мэя (1972, 2001), также вызвавшая большую критику, показала, что случайно собранные малые сообщества более устойчивы, чем большие 21,22 . С тех пор впоследствии было продемонстрировано, что экосистемы реального мира значительно отличаются от случайно собранных модельных экосистем, принимая во внимание их многогранные структурные модели 17,22,23
потенциальное улучшение с помощью молекулярных методов (например,г. метабаркодирование или метагеномика) 24 . В недавних исследованиях, придерживающихся дебатов о сложности и стабильности, используются эмпирические экосистемы разных размеров, но большинство из них по-прежнему работает в основном с небольшими (S<50, S = число видов) пищевыми сетями 17 . Исследования, посвященные большим (S>100) пищевым цепям, относительно немногочисленны 25 , поэтому обобщения в отношении экосистем реального мира все еще сомнительны.
метабаркодирование или метагеномика) 24 . В недавних исследованиях, придерживающихся дебатов о сложности и стабильности, используются эмпирические экосистемы разных размеров, но большинство из них по-прежнему работает в основном с небольшими (S<50, S = число видов) пищевыми сетями 17 . Исследования, посвященные большим (S>100) пищевым цепям, относительно немногочисленны 25 , поэтому обобщения в отношении экосистем реального мира все еще сомнительны.
Мы проанализировали изменение устойчивости крупных пищевых сетей с помощью широкого диапазона доступных эмпирических данных, чтобы найти подтверждение исходному предположению первых экологов о том, что сложные сообщества должны быть более стабильными, чем простые 9 , и получить новое понимание реалистичной части дискуссии о сложности и стабильности.Наша исследовательская гипотеза заключалась в том, что стабильность пищевых сетей растет по мере усложнения. Чтобы проанализировать эту гипотезу, мы выбрали сетевые показатели, основанные на различных методологиях: i) устойчивость, измеренная временем возврата от события возмущения 26 и ii) надежность сети на основе вторичных кривых исчезновения 27,28 . Выбранные показатели приводят к сопоставимым результатам как с подходом Мэя, так и с более производными подходами, например. те, которые использовались во многих экологических недавних исследованиях. Мы сформулировали следующий прогноз: как устойчивость, так и надежность (стабильность) пищевой сети увеличиваются с размером сети (сложностью) независимо от типа среды обитания.
Выбранные показатели приводят к сопоставимым результатам как с подходом Мэя, так и с более производными подходами, например. те, которые использовались во многих экологических недавних исследованиях. Мы сформулировали следующий прогноз: как устойчивость, так и надежность (стабильность) пищевой сети увеличиваются с размером сети (сложностью) независимо от типа среды обитания.
Результаты
Описательная статистика пищевых сетей
Наш проанализированный набор данных включал в общей сложности 450 количественных пищевых сетей с номерами узлов от 3 до 263. Из проанализированных пищевых сетей в общей сложности 85 были больше 50 узлов (18,88%). , а 34 состояли из более чем 100 узлов (7,55%). Из проанализированных наборов данных 258 пищевых сетей были водными (57,33%), из которых 206 были собраны в пресной воде, а 52 — в морских местообитаниях. Из оставшихся пищевых сетей 130 были наземными (28.88%) и 62 смешанных (13,77%). Смешанные пищевые сети включали эстуарии, болота, берега и пляжи; экотонные зоны, границы морских и наземных местообитаний.
Все используемые пищевые сети сравнивались с топологической точки зрения с использованием различных сетевых показателей, чтобы убедиться, что они репрезентативны для всего диапазона реальных пищевых сетей (рис. 1). Типы местообитаний показали существенные различия, за исключением количества узлов сети (S) (Gamma GLM: χ 2 = 5,38, df = 3, p = 0.14). В случае плотности связей пищевые сети морских местообитаний имели значительно более высокую плотность связей, чем другие, в то время как наземные имели значительно меньшую плотность связей (Gamma GLM: χ 2 = 43,02, df = 3, p <0,001). Для наименьшего среднего расстояния пресноводные и наземные пищевые сети имели меньшие средние расстояния, чем морские и смешанные (Gamma GLM: χ 2 = 22,47, df = 3, p <0,001). В случае связанности пищевые сети морских местообитаний имели значительно большую связанность, чем другие, в то время как наземные имели значительно меньшую плотность связи (Gamma GLM: χ 2 =40.75, df=3, p<0,001).
Рисунок 1.Сравнение пищевых сетей из различных типов местообитаний, на основе часто используемых показателей сети: номер узла (S), плотность связи, наименьшее среднее расстояние, связь.
Показатели стабильности
Для расчета времени возврата мы выполнили моделирование методом Монте-Карло на матрицах смежности загруженных пищевых сетей (дополнительный материал S1, таблица 1). Для одной пищевой сети сначала мы установили силы взаимодействия узлов, как определено Pimm & Lawton 14 . Во-вторых, мы случайным образом изменили эти силы взаимодействия 20 000 раз, после чего мы вычислили ведущие собственные значения каждой рандомизированной матрицы.Если пищевая сеть показывала ведущие собственные значения с отрицательными действительными частями, то считалось, что она сходится к локальной устойчивости. Если бы ведущие собственные значения имели только положительные действительные части, то сходимость пищевых сетей к локальной устойчивости не происходила бы. Затем мы взяли времена возврата как отрицательную обратную величину ведущих собственных значений с отрицательными действительными частями. Исходя из этого, мы рассчитали моду их 5% усеченного нормализованного частотного распределения, которое использовалось в качестве репрезентативного времени возврата соответствующей пищевой сети.Мы применили эту процедуру для каждой загруженной пищевой сети. Проанализированные пищевые сети с положительными ведущими собственными значениями (N = 34) были исключены из анализа времени возврата.
Для подтверждения использования нормализованных режимов частотного распределения мы также рассчитали ненормализованные режимы частотного распределения для 5% сокращенного времени возврата. Отношения между режимами ненормированного и нормализованного времени возврата в зависимости от размера пищевой сети были противоположными: режимы ненормированного времени возврата значительно увеличивались, в то время как нормированное время возврата значительно уменьшалось с размером пищевой сети, независимо от экотипов (рис. 2).Однако снижение нормированных сроков возврата наиболее выражено в морских экосистемах и слабее всего в наземных (табл. 1).
Рисунок 2. Взаимосвязь между ненормализованным и нормализованным режимами времени возврата (RT) и размерами пищевой сети.Ненормализованное среднее время возврата увеличивалось с увеличением размера пищевой сети, в то время как нормализованное время возврата уменьшалось с увеличением размера пищевой сети, независимо от экотипов (N=416, морской: N=42, смешанный: N=57, пресноводный: N=190, наземный : N=127). Координаты RT представлены с эффектом джиттера.
Таблица 1. Противоположные отношения между ненормализованным и нормализованным средним временем возвращения в зависимости от размера пищевой сети.Для всех проанализированных сетей, независимо от экотипов, взаимосвязь между ненормализованным временем возврата и размерами пищевой сети для небольшого диапазона размеров (N = 56) показала увеличение как при линейной, так и при LOESS-подгонке (рис. 3a). Кроме того, для всего диапазона размеров пищевой сети (N = 416) режимы ненормализованного времени возврата увеличились как для линейной, так и для LOESS-подгонки (рис. 3b). Линейная зависимость между режимами ненормализованного времени возврата и небольшим размером пищевой сети (диапазон S: 3–10 узлов, N = 56 пищевых сетей) показала увеличение времени возврата с увеличением размера пищевых сетей (линейная LS: пересечение = 0). .96, SE=0,22, t=4,45, p<0,001; наклон = 0,38, SE = 0,03, t = 14,27, р <0,001; F=203,68, p<0,01, N=54) (рис. 3а). Для всего диапазона размеров пищевой сети (диапазон S: 3–164, N = 416) линейная зависимость между ненормализованными режимами времени возврата и размером пищевой сети также показала непрерывное увеличение во всем диапазоне размеров (линейная LS: точка пересечения = 3,76, SE=0,34, t=11,08, p<0,001; наклон=0,22, SE=0,01, t=32,19, p<0,001; F=1035,97, p<0,001, N=362) (рис. 3b).
Рисунок 3. Взаимосвязь между временем возврата и размером пищевой сети.a) Для небольших диапазонов размеров пищевой сети (N = 56) ненормализованные режимы времени возврата увеличились как для логарифмической, так и для LOESS-подгонки. b) Для всего диапазона размеров пищевой сети (N = 416) ненормализованные режимы времени возврата увеличились как для логарифмической, так и для LOESS-подгонки. c) Для небольших диапазонов размеров пищевой сети (N = 56) режимы нормализованного времени возврата увеличились как для логарифмической, так и для LOESS-подгонки. d) Для всего диапазона размеров пищевой сети (N = 416) режимы нормализованного времени возврата постоянно уменьшались при логарифмической подгонке и продемонстрировали снижение с локальным увеличением при подгонке LOESS.Координаты RT представлены с эффектом джиттера.
Опять же, для всех проанализированных сетей, независимо от экотипов, взаимосвязь между нормализованным временем возврата и размерами пищевой сети для небольшого диапазона размеров (N = 56) показала увеличение как для логарифмических, так и для LOESS-подгонок (рис. 3c). В то время как для всего диапазона размеров (N = 416) режимы нормализованного времени возврата постоянно уменьшались как для логарифмических, так и для LOESS-подгонок (за исключением вышеупомянутого диапазона размеров малых пищевых сетей S = 3-10) (рис. 3d). Логарифмическая зависимость между нормализованными режимами времени возврата и небольшим размером пищевой сети (диапазон S: 3-10 узлов, N = 56 пищевых сетей) показала увеличение времени возврата с увеличением размера пищевых сетей (логарифмический NLS: пересечение = 3.23, SE=1,42, t=2,28, p=0,03; наклон = 2,00, SE = 0,68, t = 2,92, р = 0,005; F=8,55, p=0,005, N=54) (рис. 3в). Для всего диапазона размеров пищевой сети (диапазон S: 3–164, N = 416) логарифмическая зависимость между нормализованными режимами времени возврата и размером пищевой сети показала непрерывное уменьшение во всем диапазоне размеров (логарифмический NLS: пересечение = 9,78, SE = 0,54, t=18,17, p<0,001; наклон=-1,13, SE=0,15, t=-7,27, p<0,001; F=52,9, p<0,001, N=362) (рис. 3d).
На основе процедуры взвешенной полиномиальной регрессии LOESS для аппроксимации кривой нормированные значения режима времени возврата начинают резко уменьшаться после того, как размер пищевой сети достигает S=10, и после небольшого увеличения около S~50 оно продолжает снижаться для оставшийся диапазон размеров (S max = 164) (рис. 3d).
Для расчета устойчивости мы использовали площадь под вторичной кривой ослабления 28 , также известную как кривая устойчивости к атакам 27 . Кривая толерантности показывает, сколько вымираний (исчезновений видов) может произойти на уровне добычи в пищевой сети, не достигая полной потери уровня хищника. Высокие значения устойчивости представляют собой более стабильную пищевую сеть, в которой резкое уменьшение количества хищников происходит только после большого количества жертв. Для низких значений надежности резкое снижение происходит уже после нескольких потерь молитв.Таким образом, низкое значение устойчивости представляет собой нестабильную пищевую сеть. Значения устойчивости были установлены между 0 и 1. Значения устойчивости не могли быть рассчитаны в случае двух пищевых сетей из-за небольшого размера и небольшого количества соединений. Таким образом, набор данных устойчивости содержал N = 448 проанализированных пищевых сетей.
Логарифмическая зависимость между надежностью и небольшим размером пищевой сети (диапазон S: 3-10 узлов, N = 52 пищевых сети) показала незначительное увеличение устойчивости с увеличением размера пищевой сети (логарифмический NLS: пересечение = 0.53, SE=0,09, t=5,84, p<0,01; наклон = 0,05, SE = 0,04, t = 1,26, p = 0,22; ANOVA log(x): F=1,58, p=0,21, N=52) (рис. 5а). Логарифмическая зависимость между устойчивостью и размером пищевой сети показала значительное увеличение во всем диапазоне размеров (диапазон S: 3–164, N = 448) (логарифмический NLS: пересечение = 0,49, SE = 0,02, t = 29,46, p <0,001; наклон = 0,07, SE = 0,005, t = 14,05, p <0,001, ANOVA log (x): F = 197,28, p <0,001, N = 448) (рис. 5b).
Рисунок 5. Взаимосвязь между надежностью и размерами пищевой сети.а) Для небольших диапазонов размеров пищевой сети (N = 52) надежность увеличилась как для логарифмической, так и для LOESS-подгонки.b) Для всего диапазона размеров пищевой сети (N = 448) надежность непрерывно увеличивалась при логарифмической подгонке и также продемонстрировала увеличение с локальным снижением при подгонке LOESS.
На основе процедуры взвешенной полиномиальной регрессии LOESS для подбора кривой значения устойчивости показали тенденцию к увеличению размера пищевой сети S = 10 и, а также снова показали увеличение для остального диапазона размеров (S макс = 263) . Значения устойчивости по типам среды обитания также демонстрируют тенденцию к увеличению с увеличением размера пищевой сети (рис. 6).
Рисунок 6. Взаимосвязь между устойчивостью и размерами пищевых сетей для типов местообитаний.Значения устойчивости увеличивались с увеличением размера пищевой сети в четырех типах среды обитания: пресноводной (N=186), морской (N=47), смешанной (N=56) и наземной (N=127).
Обсуждение
Наше исследование является одним из немногих анализов, которые также включали эмпирические системы с более чем 200 видами 9,16,17,19,29 . Несмотря на то, что мы проанализировали обширный набор эмпирических данных, большинство предыдущих исследований стабильности сетей касались либо теоретических сетей; или, если использовались наборы эмпирических данных, они, как правило, содержали несколько небольших сетей 15,26,30,31 .Насколько нам известно, до сих пор это самый большой проанализированный набор эмпирических данных с точки зрения стабильности пищевой сети.
Мы проанализировали взаимосвязь между сложностью и стабильностью в большом диапазоне эмпирических размеров пищевых сетей по времени возврата и надежности. Наши результаты привели нас к предположению, противоречащему предположению о том, что стабильность должна снижаться с увеличением сложности пищевой сети: большие пищевые сети имеют более короткое время возврата, чем маленькие, если время возврата масштабируется (нормализовано от минимума до максимума).Объяснение нашего предположения основано на следующем. Крупные пищевые сети имеют более высокие трофические уровни, которые обычно включают более крупные виды 32,33 , у которых увеличилось среднее время генерации 34,35 . Время генерации влияет на динамику пищевой сети, оказывая значительное влияние на время возвращения 36 . Таким образом, при сравнении пищевых сетей разного размера с использованием их времени возврата требуется нормированное минимальное и максимальное масштабирование для исправления систематической ошибки из-за различий во времени образования и их влияния на время возвращения.Таким образом, мы фактически получили с помощью нормализации минимума и максимума время генерации без времени возврата. Наши результаты пищевых сетей без времени генерации (рис. 3а) также подтверждают результаты литературы: для сетей с 10 или менее узлами типичное время возврата увеличивается с номерами узлов 15,26 . Это означает, что в диапазоне малых размеров более крупным системам требуется в среднем больше времени для возврата в устойчивое состояние после возникновения случайного возмущения.
С точки зрения структуры пищевой сети и других свойств собранные сети были разнообразными, но достаточно похожими, чтобы их можно было сравнивать на основе топологических параметров.Судя по изученным пищевым цепям, морские экосистемы отличаются от других местообитаний и в среднем по всему набору данных в нескольких случаях. Эти явления можно отнести к особенностям данного типа среды обитания. Наш результат согласуется с другими результатами анализа пищевой сети 35 . Однако наши результаты были постоянными независимо от показателей и типов среды обитания. Независимо от типов местообитаний время возвращения уменьшалось с увеличением размера сети, и аналогичные результаты наблюдались с точки зрения надежности.Это имеет два следствия: с одной стороны, повышение стабильности по отношению к крупным сетям не зависит от среды обитания; с другой стороны, независимая метрика, которая работает с другими математическими методами, дала тот же результат (надежность).
Использование простой метрики, такой как R, основанный на вторичных вымираниях, привело к увеличению стабильности в зависимости от размера пищевой сети. Используя нормализованное ВУ, рассчитанное на основе 95% их распределения, в отличие от методологии Мэя, где использовалось только ВУ в интервале 0–150, мы получили те же результаты, что и R.Помимо этого, мы также обнаружили, что предположения Мэя справедливы как для нормализованного времени, так и для интервала 0–150 RT только для небольших пищевых сетей, в то время как для более крупных пищевых сетей положительная связь между сложностью и стабильностью является правилом.
Надежность часто используется для мутуалистических и небольших пищевых сетей 1,27,28,37 . Наши результаты подтверждают результаты Dunne et al. 38 : надежность увеличивается с размером сети и, как следствие, сложностью. Для оценки надежности мы применили их методы (оценка по вторичным исключениям), и диапазон размеров их сетей (S = 25-172) хорошо перекрывается с нашим набором данных, хотя количество исследованных сетей было относительно небольшим (N = 16). .
Хотя мы подтвердили, что и надежность, и время возврата одинаково полезны для описания стабильности сети (эти два подхода взаимно дополняют друг друга), мы рекомендуем использовать время возврата, поскольку оно может предоставить временную информацию для стабильности. Кроме того, наши результаты проливают свет на новые четкие представления о взаимосвязи «сложность-стабильность». В соответствии с предыдущими предположениями, дебаты, направленные на отношение «сложность-стабильность», вероятно, будут разрешены не одной простой теорией, а комбинацией нескольких теорий и подходов.Соответственно, полученные нами результаты потенциально могут преодолеть разрыв между предыдущими сильными математическими теориями и противоречивыми эмпирическими исследованиями.
Время возврата является привлекательной метрикой с математической точки зрения. Исследования устойчивости, основанные на собственных значениях матриц систем, имеют долгую историю как среди математиков, так и среди физиков. В результате вычислительные методы и теории, основанные на них, усложнились и завоевывают популярность в других областях исследований.Эта метрика также интересна тем, что она может определить время, необходимое системе для достижения равновесия. Он также обеспечивает интуитивную индикацию стабильности. С момента появления противоречивых результатов 15,20,26,39 число исследований, основанных на показателях времени возврата, увеличилось. Хотя метод един, результаты исследований часто противоречивы. Одно из значений наших результатов заключается в том, что они продемонстрировали стабильность больших сетей с точки зрения времени возврата и надежности, учитывая противоречия, обнаруженные в литературе.Они до сих пор остаются нерешенными из-за различных аналитических методов и используемых критериев 9,12 .
Хотя мы обнаружили значительную связь между сложностью и стабильностью, Jacquet et al. 17 показали, что эмпирические пищевые сети обладают несколькими неслучайными свойствами, которые способствуют отсутствию взаимосвязи сложность-стабильность. Однако в их исследовании стабильность зависела не от размера сети, а от ее сложности. Более того, хотя они проанализировали 116 сетей, их самый большой размер сети включал только 50 видов.На рисунке 2 видно, что увеличение стабильности в наших данных проявляется при числе узлов 50, и увеличение является постоянным для более высоких размеров, основываясь также на кривой LOESS. Другие недавние подходы 16,19 пытались объяснить стабильность больших сетей пространственными характеристиками. Таким образом, наше исследование несопоставимо с ними, но используемая методология основана либо на времени возврата, либо на использовании матриц Якобиана, что дополнительно может доказать эффективность этих методов.Однако отрицательная корреляция между стабильностью и сложностью ранее также противопоставлялась тому ограничению, что более длинные пищевые цепи могут иметь более короткое время возврата, чем более короткие, если их поток энергии достаточно быстр 40,41 .
Таким образом, наш подход исправляет взаимосвязь между стабильностью пищевых сетей и их размером, правильно обрабатывая влияние времени образования на время возврата. Этот результат позволяет по-новому взглянуть на противоречивую взаимосвязь стабильности и сложности пищевых сетей.
Материалы и методы
Анализ данных
Данные
На протяжении всего настоящего исследования мы использовали набор из 450 эмпирических пищевых сетей, собранных из различных онлайновых и общедоступных источников. Большинство пищевых сетей (~80%) были созданы на сайте www.globalwebdb.com, который представляет собой онлайн-базу данных, содержащую большой набор (359) опубликованных эмпирических пищевых сетей. Часть используемых пищевых сетей (51) доступна через базу данных ECOWeb 42 . Другая часть пищевых цепей (26) поступает из Ференджордана.База данных webnode.hu. Скомпилированный набор данных содержит широкий спектр пищевых сетей, происходящих из разных типов местообитаний, охватывающих широкий диапазон размеров (3–263 S) и имеющих разные уровни плотности связей (0,67–36,02 связи/вид) (дополнительный материал S1, таблица 1). .
Мы классифицировали каждую пищевую сеть на основе типа среды обитания, из которой она возникла, и определили 4 категории: водные среды обитания, которые можно далее разделить на пресноводные и морские среды обитания; наземные среды обитания, содержащие в основном неводные виды; и смешанные типы местообитаний, представляющие собой экотонные зоны, расположенные на границе наземных и различных водных типов местообитаний (напр.эстуарии, болота, болота, берега и пляжи, водно-болотные угодья, трясины, илистые отмели и т. д.).
Мы сравнили скомпилированные пищевые сети с топологической точки зрения, чтобы убедиться, что характеристики стабильности пищевых сетей являются репрезентативными и не являются побочным продуктом различий в топологической структуре. Мы рассчитали 4 широко используемые топологические характеристики для каждой пищевой сети (таблица 2, дополнительный материал S1, таблица 1) и использовали обобщенные линейные модели для оценки различий в топологии между 4 различными типами местообитаний (таблица 1).
Таблица 2. Топологические метрики и их формулы, используемые при сравнении пищевых сетей.Мы изучили стабильность пищевых сетей, используя два показателя стабильности: время возврата (RT) и надежность (R). Основной причиной выбора этих метрик являются принципиальные различия в том, как они рассчитываются, и как они оценивают стабильность. Используя метрики, существенно отличающиеся как по концепции, так и по способу их расчета, мы снижаем риск получения результатов, необъективных из-за используемых метрик.
Время возврата (RT)
RT получено из анализа линейной стабильности матрицы взаимодействия пищевой сети 43 . Мы получаем такую матрицу путем линеаризации системы уравнений Лотки–Вольтерра вида N в точке равновесия. Каждая запись a ij матрицы взаимодействия количественно определяет изменение скорости роста популяции видов i , вызванное небольшим отклонением численности видов j от равновесной численности.Такую матрицу взаимодействия можно рассматривать как матрицу Якоби системы, и она описывает динамику численности каждого вида в течение непрерывного времени. Мы можем определить (локальную) устойчивость равновесия системы, взглянув на ее собственные значения, полученные из матрицы Якоби. Если ведущее собственное значение (собственное значение с наибольшей действительной частью) системы имеет отрицательные действительные части, то равновесие устойчиво (малые возмущения в равновесии со временем затухают). Если ведущее собственное значение системы имеет положительные действительные части, то равновесие неустойчиво (малые возмущения в равновесии со временем усиливаются).Если равновесие устойчиво, мы можем использовать отрицательную инверсию действительной части ведущего собственного значения, чтобы определить время, необходимое системе для возврата в состояние равновесия после небольшого возмущения. Вышеупомянутое время, необходимое для возврата к равновесию, равно RT 9,15,20,39 . Целью нашего исследования не было продемонстрировать достоверность показателя стабильности Мэя 15,20 , и наш анализ не включает этот критерий стабильности, поскольку его достоверность обсуждается 44 .
На протяжении всего моделирования мы считаем, что каждая пищевая сеть находится в состоянии локального равновесия. Силы взаимодействия по умолчанию для каждой матрицы Якоби были установлены на основе Pimm & Lawton 26 . Значения в верхнем треугольнике матрицы (влияние хищника на жертву) были установлены на -1, а значения в нижнем треугольнике матрицы (влияние жертвы на хищника) были установлены на +0,1. Диагонали каждой матрицы (внутривидовые взаимодействия) также были установлены равными -1. Мы моделировали возмущения, умножая каждый ненулевой элемент матрицы Якоби на случайные числа, полученные из распределения U(0, 1).После рандомизации вычислялись ведущие собственные значения и определялись их знаки. Мы рассчитали RT для каждого случая, в котором были получены ведущие собственные значения с отрицательными действительными pats. Мы повторили эти шаги для каждой пищевой сети 20 000 раз. После удаления пищевых сетей, которые не давали ведущих собственных значений с отрицательными действительными частями, мы получили набор из 416 пищевых сетей.
После выполнения моделирования мы получаем число RT от 0 до 20 000 для каждой пищевой сети со значениями в диапазоне от ∼1 до >2e+09 (самое большое встречающееся RT).Этот большой диапазон порождает потенциальную проблему. Значения верхнего предела диапазона ВУ могут быть слишком большими, чтобы их можно было применить в реальной жизни (системе потребуется слишком много времени, чтобы вернуться в состояние локального равновесия, чтобы ее можно было применить). Pimm & Lawton 26 справляется с этим большим диапазоном, исключая все RT, превышающие 150, и работая только со значениями, которые меньше этого порога. Этот метод действителен для небольших пищевых сетей (цепей), но приводит к другой проблеме: хотя все пищевые сети дают правонаправленное распределение RT, а некоторые моды (и, следовательно, большинство значений) очень близки или даже превышают этот порог, удаляя значения, близкие к исходным (неусеченным и ненормализованным) режимам распределений, приводит к искусственному сдвигу в сторону более высоких значений.Мы решаем эту проблему, выполняя 5-процентную подстройку в верхней части нашего диапазона ВУ. Подстройки на 5% достаточно, чтобы избавиться от длинного хвоста распределения с экстремальными значениями, сохранив при этом наиболее частые значения RT вокруг мод.
Другая поправка, которую мы использовали для каждой пищевой сети, связана со временем их образования. Если две пищевые сети имеют одинаковый размер, но различаются средним временем генерации (например, две пищевые сети с 10 узлами, одна из которых состоит из трав, насекомых и насекомоядных птиц, а другая — из деревьев, млекопитающих, травоядных и хищников), их RT после возмущений будут пропорциональны среднему времени их генерации.Это означает, что пищевые сети с большим средним временем генерации будут иметь в среднем более высокие значения RT. Чтобы оценить взаимосвязь между RT и размером пищевой сети, нам пришлось внести поправку на среднее время генерации. Чтобы скорректировать среднее время генерации пищевых сетей, мы выполнили минимальную и максимальную нормализацию RT каждой пищевой сети. Таким образом, мы масштабировали диапазон RT каждой пищевой сети до [0, 1], что сделало распределения напрямую сопоставимыми.
Мы определили репрезентативную RT пищевой сети, создав ячейки на интервале [0, 1] с шагом 0.01 (всего 100 ячеек в случае каждой пищевой сети) и подсчет количества RT в каждой ячейке. Мы взяли режим распределения ВУ для этих ячеек (интервал 1%) в качестве репрезентативного ВУ (1% наиболее часто встречающегося ВУ в случае пищевой сети). Только в 7 случаях мы получили две моды распределений ВУ. В этих случаях мы выбирали режимы с более высокими значениями RT.
Дальнейший анализ чувствительности, нацеленный на построение случайных матриц Якоби, может привести к лучшему пониманию динамики стабильности, путем извлечения множителей силы взаимодействия из других распределений, отличных от U(0, 1), и за счет учета коррелированных сил взаимодействия.
Надежность (R)
Основная причина использования R заключалась в том, чтобы иметь независимую меру стабильности, которую можно было бы использовать для сравнения и проверки результатов анализа ВУ. R привлекателен, потому что это простая метрика стабильности, которую легче интерпретировать, чем RT.
Взаимодействия в пищевой сети можно рассматривать как двусторонние взаимодействия, где мы рассматриваем взаимодействия пар видов (хищник и жертва) по отдельности. Этот подход может быть полезен, потому что, рассматривая эти системы как двудольные, мы можем использовать метрики, разработанные для характеристики стабильности двудольных систем.Одним из таких показателей является скорость вторичного вымирания (SER) верхнего уровня. В этом случае хищники находятся на верхнем уровне, а жертвы — на нижнем. Для более длинных пищевых цепей промежуточные виды включаются отдельно как хищники и жертвы. Для определения SER верхних уровней случайным образом по одному удаляем виды с нижних уровней и подсчитываем количество хищников, оставшихся без добычи в конце каждого шага. Предполагается, что такие хищники вымрут из-за отсутствия добычи, и с этого момента они исключаются из моделирования.Этот процесс повторяется до тех пор, пока в системе не останется ни одного вида 27,28 .
Если мы нанесем количество выживших видов после этапов вымирания, мы получим так называемую кривую устойчивости к атакам, которая характеризует выживаемость системы 27 . Этот подход может дать хорошее представление о потенциальных каскадах вымирания, которые могут быть вызваны случайным вымиранием определенных видов в данной сети. SER можно смоделировать, применив сплайн-интерполяцию к кривой толерантности к атаке с помощью функции f(x) = 1-x a , где показатель степени a представляет вторичные скорости исчезновения 28 .
Значения R получены из SER 27,28 и определены как площадь под кривой устойчивости к атаке. Мы выполнили минимальную и максимальную нормализацию по осям x и y, чтобы значения находились в диапазоне от 0 до 1, и, таким образом, площадь под кривой устойчивости к атаке (R) также попадает в этот диапазон. Это полезно, потому что интуитивно значение R около 0 характеризует нестабильную, менее надежную систему, а значение около 1 характеризует стабильную, гораздо более надежную систему. Это связано с тем, что для возникновения низких значений R количество видов-жертв, которые необходимо удалить для массового вымирания, невелико.Это означает, что система развалится после относительно небольшого количества вымираний, что указывает на ее хрупкость. Противоположное верно в случае высоких значений R: необходимо большее количество вымираний, чтобы система развалилась, что указывает на то, что система более стабильна 27,28 .
Моделирование SER проводилось 1000 раз для каждой отдельной пищевой сети, а среднее значение R рассчитывалось и использовалось для анализа в случае каждой пищевой сети.
Программное и аппаратное обеспечение
Статистический анализ и моделирование проводились с использованием статистической среды R 45 . GLM оценивались с использованием пакета «автомобиль» 46 . Моделирование устойчивости проводилось с использованием модифицированной (распараллеленной) версии функции « Robustness » из «bipartite» пакета 47 . Базы данных SQLite использовались для хранения больших таблиц данных, созданных в результате моделирования, и управления ими. Моделирование проводилось на сервере «Котыс» Университета Бабель Дж. Бойяи 48 .
Благодарности
Мы благодарны Джоэлу Э. Коэну за его ценные и критические комментарии к рукописи. Мы благодарим команду управления кластером «Котыс» в BBU Cluj за техническую поддержку во время моделирования. Мы также благодарим нескольких коллег за ценные комментарии к нашему анализу и проекту рукописи.
Библиография
- 2.↵
- 3.↵
- 3.↵
- 4.↵
- 5.↵
- 6. ↵
- 7. ↵
- 8.↵
Hillebrand, H. et al. Изменение климата: воздействие потепления на морское биоразнообразие. в Справочнике по защите морской среды 353–373 (Springer International Publishing, 2018). doi: 10.1007/978-3-319-60156-4_18
- 9.↵
- 10.↵
- 11.↵
Dell, AI, Kokkoris, GD, Bana, , М. Как пищевые сети сохраняются в природе? в Dynamic Food Webs□: многовидовые сообщества, развитие экосистем и изменение окружающей среды 425–436 (Academic Press, Elsevier, 2005).
- 12.↵
- 13.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21 .
- 22.↵
- 23.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 27.↵
- 29.↵
- 30.↵
- 31 .
- 32.↵
- 33.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
Cohen, J.E. Co-Operative Web Bank. Версия 1.1. Машиночитаемая база данных пищевых сетей. Нью-Йорк: Университет Рокфеллера. (2018).
- 43.↵
- 44.↵
- 45.↵
- 46.↵
John, F. & Sanford, W. R Companion to Applied Regression Third Edition. SAGE Publications, Inc. (2019). doi: 10.1017/CBO9781107415324.004
- 47.↵
- 48.↵
Пищевая цепь и пищевая сеть: различия и важность
Мы все зависим от еды, чтобы выжить. Энергия необходима для роста живых существ. Но знаете ли вы, что существует процесс создания этой еды? Есть также ряд различных растений и животных , которые зависят от этого творения, чтобы выжить. Для этого есть имена, и узнать больше — отличный урок, который может вас удивить.
Пищевая цепь — это система, используемая для передачи энергии от одного организма к другому.Когда эти системы используются в экологии, они известны как пищевая сеть. Пищевая цепь описывает метод, с помощью которого конкретный организм собирает пищу. Другими словами, пищевая цепь — это последовательность того, кто кого ест в биологическом сообществе или экосистеме для получения пищи. Пищевая цепь — это способ изображения потока энергии от одного организма к другому, к следующему и так далее. Всем нужна энергия, передаваемая по пищевой цепи, чтобы выжить.
Пищевая цепь в основном показывает, как живые существа получают пищу и как энергия и питательные вещества передаются от существа к существу.
Согласно Википедии,
«Пищевая цепь представляет собой линейную сеть звеньев в пищевой сети, начинающуюся с организмов-продуцентов (таких как трава или деревья, которые используют солнечное излучение для производства пищи) и заканчивая высшими видами хищников (такими как медведи гризли или косатки). ), детрифагов (таких как дождевые черви или мокрицы) или разлагающих видов (таких как грибы или бактерии)».
Солнечный свет основной источник энергии для всех живых организмов. Он необходим для того, чтобы снабжать растения необходимой им пищей, а также давать людям и другим людям энергию, которая поддерживает их здоровье и избавляет от болезней.Есть четыре основных звена пищевой цепи. Эти четыре части:
- Продуценты: Продуценты, или растения, получили это название в связи с тем, что они являются единственными организмами, производящими себе пищу. Растения также производят пищу для других организмов.
- Потребители: Потребитель — это организм, который питается чем-либо, например животными или растениями. Есть как хищники, так и травоядные, паразиты и падальщики.
- Поглотители: Поглотителями являются бактерии или грибы.Хотя мы не слишком любим падальщиков, они важны в пищевой цепочке из-за того, что мертвое вещество превращается в азот и углерод, которые выбрасываются в нашу атмосферу.
- Разлагатели: Разлагатели вступают в игру, чтобы закончить там, где остановился мусорщик.
Давайте более подробно рассмотрим каждую из этих частей пищевой цепи и узнаем, как они работают.
Производители
Производители дают нам необходимую энергию посредством процесса, известного как фотосинтез.Этот процесс в сочетании с минералами и питательными веществами из почвы позволяет растению расти и цвести, а затем размножаться. Продукты, которые производятся с помощью этого процесса, включают:
- Яблоки
- Пекан
- Персики
- Морковь
- Картофель
Растения производят большое количество энергии, необходимой для питания. Они используют солнечный свет, чтобы помочь им расти. Да, все те растения, которые вы видите там, используют солнечный свет, чтобы производить себе пищу и помогать себе расти.
Выживание многих животных зависит от этих растений. Это подводит нас к первичным потребителям.
Потребители
Первичные потребители
Первичные потребители известны как травоядные в мире животных. Этим потребителям дано название, потому что они первыми съедают растение. Как только растение съедено, энергия передается и затем используется этим животным. Например, основным потребителем является кузнечик, который ест траву.
Вторичные потребители
Вторичные потребители — более крупные животные, которые поедают более мелких животных и насекомых.В этом случае вторичным потребителем является крупное животное, поедающее животное, поедающее растение. Энергия передается вторичному потребителю так же, как и первичному потребителю. Например, вторичный потребитель — это крыса, которая ест кузнечика.
Третичные потребители
Третичные консументы — это плотоядные, которые потребляют других плотоядных. Человек считается третичным потребителем, как и ряд других крупных животных, включая тигров, лосей, слонов, львов, медведей и акул.В большинстве случаев у третичного потребителя нет хищника, но это не всегда так. Когда они умирают, они становятся вкусной едой для многих животных. Как правило, это не беспокоит человека, к счастью, но есть много животных, которые беспокоятся. Третичные потребители, такие как змея, поедают вторичных потребителей.
Мусорщики и разлагатели
Последнее звено в цепи — падальщики и разлагатели. Когда животное проживает свою жизнь и умирает естественной смертью, оно оставляет свой труп. Редуценты расщепляют питательные вещества в мертвом «веществе» и возвращают его в почву.Животное оставляет после себя огромное количество энергии. Животное начинает гнить, и это становится очень грубой сценой. Чтобы этого не произошло, существуют падальщики и разлагатели.
Мусорщик обладает следующими качествами:
- Мусорщик — это животное, которого привлекает запах мертвого животного
- Мусорщики едят мертвое животное
- В большинстве случаев именно запах предупреждает падальщика о присутствии мертвого животного. Однако иногда падальщик получает подсказки от других групп падальщиков, пришедших насладиться пиршеством .
- Падальщики включают лисиц, медведей, койотов и грифов-индюков.
- У падальщиков острое обоняние
- Эти животные едят животное, оставляя его тушу
Однако ничего не пропадает даром, и с оставшейся частью животного в игру вступает разлагатель. Когда животное поедают падальщики, появляются насекомые и ломают животное даже дальше, чем падальщик. Личинки, муравьи и жуки являются распространенными редуцентами. Съев животное, они откладывают яйца. Рождающиеся личинки продолжают поедать разлагающееся тело.Бактериальные и грибковые вещества завершают процесс разложения, пока он не станет полностью единым. Энергия, которая была у животного, передается животным и насекомым, которые его съели.
Еще несколько фактов, которые важно знать:
- Бактерии и грибки возвращают вещество в почву. Затем он повторно используется для роста растений и получения энергии.
- Все это кажется неприятным процессом, и вы, конечно, не хотите стоять в стороне, когда это происходит. Но подумайте об этом так: в каком мире мы жили бы без падальщиков и разлагателей?
Взгляд на пищевую сеть
Пищевая сеть — это практически то же самое, что и пищевая цепь, только в гораздо более упрощенном виде.Сеть из множества пищевых цепей называется пищевой сетью. Пищевая сеть обеспечивает обильный запас пищи, чтобы все растения, животные и насекомые имели достаточное количество. Если растение одного определенного типа погибнет из-за болезни, пострадают кузнечики и многие другие насекомые. Тогда это вызовет страдания других звеньев пищевой цепи. Пищевая сеть гарантирует, что этого не произойдет.
Люди всеядны, то есть питаются животными и растениями. Таким образом, человек является как первичным, так и вторичным потребителем пищи.Однако в большинстве случаев третичным потребителем является человек. Когда вы едите овощи, вы являетесь их основным потребителем. Когда вы едите мясо, вы вторичный потребитель. Но как третичный потребитель пищи человек обычно является основным потребителем мяса и растений.
Разница между пищевой сетью и пищевой цепью
Вот несколько фактов о пищевой сети и пищевой цепи, которые важно понимать.
- Пищевая сеть представляет собой группу различных пищевых цепей, которые работают вместе, чтобы создать экосистему.
- Пищевые цепи в пищевой сети взаимосвязаны в той или иной точке.
- Пищевая цепь представляет собой модель, по которой энергия передается от производителей к разлагателям.
- Пищевая цепь движется только по одному пути, когда животные находят пищу.
- Пищевая сеть показывает множество различных путей, которыми связаны растения и животные.
- Пищевая сеть представляет собой несколько пищевых цепей, соединенных вместе.
- Пищевая цепь всегда начинается с растения и заканчивается животным.
- Всем живым существам для выживания нужна энергия. Эта пищевая сеть и процессы пищевой цепи позволяют получить эту энергию.
- В мире существует более 100 000 различных разлагателей
- Существует четыре типа потребителей: хищник не ест ничего, кроме животных; травоядное не ест ничего, кроме растений; всеядное животное питается как растениями, так и животными; падальщики едят мертвых животных и их туши.
- Необходимы все звенья пищевой цепочки.Даже если один из звеньев пищевой цепи будет устранен, это может привести к катастрофическим последствиям.
- Растения находятся в начале пищевой цепи, а люди в конце.
Важность пищевых цепочек
Пищевые цепи важны, потому что они показывают сложные взаимосвязи в экосистемах. Видно, что в пищевых цепях каждый организм занимает определенную нишу в экосистеме. Пищевые цепи показывают, как каждый организм зависит от кого-то другого в своем выживании.
1.Пищевая цепь уравновешивает экосистемуПищевые цепи отображают обстоятельства, при которых производитель или потребитель теряется при любом происшествии. Целые сообщества могут развалиться. Таким образом, пищевые цепи помогают ученым больше узнать об экосистемах и помогают им сохранять равновесие.
Выживание каждого вида растений и животных, больших или малых, в некоторой степени зависит от другого вида растений или животных. Это могут быть пчелы, собирающие пыльцу с цветов, фотосинтез растений, олени, поедающие листья кустарников, или львы, поедающие оленей.Если хоть одна точка будет потеряна, баланс в экосистеме будет нарушен.
2. Питательные сети показывают, что растения являются основой
Пищевые сети являются важным инструментом для понимания того, что растения являются основой всех экосистем и пищевых цепей. Фотосинтез — это только начало пищевой цепи. Растения, у которых есть фотосинтез, поставляют нам первый продукт пищевой цепи для питания.
Растения являются источником пищи, которую мы едим, кислорода для выживания и размножения, нашей одежды и даже нашей мебели, многих других бесчисленных вещей, обеспечивающих устойчивость.Растения также удаляют из воздуха парниковые газы, обеспечивают среду обитания для многих животных. Поэтому жизненно важно понимать экологию окружающей среды по отношению к растениям.
3. Естественный отбор
Пищевые сети помогают понять естественный отбор, показывая иерархию видов, с плотоядными, всеядными и третичными животными на вершине всех пищевых цепей. Это физическая и поведенческая адаптация растений и животных, представляющая модель потребления, основанную на естественных атрибутах и инстинктах выживания.
Пищевые сети также объясняют, как нехватка продовольствия из-за чрезмерной охоты, браконьерства, глобального потепления и разрушения среды обитания разрушает популяции, приводя к их вымиранию.
4. Движение энергии и питательных веществ
Пищевая цепь описывает, как энергия, питательные вещества и органические соединения перемещаются по экосистеме. В основе лежат растения, производящие энергию; затем он переходит к организмам более высокого уровня, таким как травоядные. После этого, когда хищники поедают травоядных, энергия передается от одного к другому.
5. Пищевая цепь определяет трофический уровень организмов
В зависимости от пищевой цепи организм можно рассматривать как находящийся более чем на одном трофическом уровне. Например, тюленей можно считать высшими хищниками на самом высоком трофическом уровне в определенных средах, где они поедают рыбу, которая является первичным или вторичным потребителем.
Однако в других сообществах, где тюлени становятся добычей акул, их можно считать находящимися на более низком трофическом уровне. Эти отношения легче увидеть в пищевых сетях и труднее заметить в пищевых цепях или пирамидах.
Заключительные мысли
Очень интересно изучать процесс пищевой сети и пищевой цепи. Мы часто просто садимся и едим, а не думаем о том, откуда берется наша еда или о том, как она попала на наши столы. Когда вы узнаете больше о пищевой сети и пищевой цепи, вы легко сможете больше заботиться о своих продуктах.
.