
Структура экосистемы пространственная: Структура экосистем | Параграф 5. 4
Структура экосистем | Параграф 5. 4
«Биология. Общая биология. Базовый уровень. 10-11 классы». В.И. Сивоглазов (гдз)
Вопрос 1. Что такое биогеоценоз?
Биогеоценоз — это экологическая система (экосистема), границы которой определены растительным сообществом. Совокупность биогеоценозов земного шара образует глобальную систему, биосферу. Примерами биогеоценозов являются дубрава, луг, ельник, березовая роща и др.
Вопрос 2. Расскажите о пространственной структуре экосистемы.
Пространственная структура экосистемы определяется ярусным расположением растительности. Сверху вниз над землей выделяют налоговый (древесный), кустарниковый, травяной и надпочвенный (приземный) ярусы. В почве также идет разделение на уровни, образованные корнями растений разных видов. Такая пространственная организация позволяет растениям эффективно использовать свет и другие ресурсы, а животным — занимать разные экологические ниши и ослаблять конкуренцию между близкими видами.
Вопрос 3. Какие обязательные компоненты включает любая экосистема?
В любой экосистеме можно выделить два основных структурных компонента — биотоп и биоценоз. Биотоп — это комплекс факторов неживой природы, которые в сумме формируют определенные климатические, географические, почвенные и другие параметры экосистемы. Биоценоз — это совокупность всех живых организмов (популяций) экосистемы. Он разделяется на зооценоз (сообщество животных), фитоценоз (сообщество растений) и микробоценоз (сообщество микроорганизмов).
Вопрос 4. В каких отношениях находятся друг с другом обитатели биоценозов? Охарактеризуйте эти связи.
С точки зрения экологической структуры в экосистеме можно выделить три группы особей
1. Продуценты (производители) — автотрофные организмы, главным образом зеленые растения, синтезирующие органические вещества из неорганических. Их биомасса — это первичная продукция экосистемы, которая служит пищей и источником энергии для всех остальных организмов сообщества.
 Автотрофами являются растения, фотосинтезирующие и хемосинтезирующие прокариоты.
Автотрофами являются растения, фотосинтезирующие и хемосинтезирующие прокариоты.2. Консументы (потребители) — гетеротрофные организмы (растительноядные и плотоядные), потребители живого органического вещества. Консументами являются многие животные, паразитические бактерии, хищные растения.
Таким образом, продуценты используют неорганические соединения, создавая первичную продукцию. Далее часть консументов (растительноядные животные) используют первичную продукцию, создавая вторичную биомассу, которая используется другими консументами (хищники, паразиты). После смерти продуценты и консументы разлагаются редуцентами до минеральных соединений, которые вновь используются продуцентами.
Вопрос 5. Опишите видовой состав и пространственную структуру экосистемы дубравы.
Видовой состав дубравы очень разнообразен, что обеспечивает ее стабильность как экосистемы. Растения дубравы образуют четко разделенные ярусы. В верхнем древесном «этаже» расположены крупные многолетние дубы и липы. Второй ярус составляют низкорослые и менее светолюбивые груша, клен, яблоня. Далее следует кустарниковая растительность: лещина, бересклет, калина, боярышник, бузина. Травянистый ярус состоит из кустарничков, всходов деревьев, папоротников и различных трав (медуница, хохлатка, ветреница, иван-да-марья и т. д.). Приземный ярус представлен мхами, низкими травами; здесь также обитают лишайники и грибы. Животный мир дубравы не менее разнообразен. Членистоногие населяют все ее уровни. Из позвоночных в верхних ярусах мы встретим много птиц — сороку, зяблика, дроздов, синиц, ночных и дневных хищников. В кустарниках обитают зарянка, мухоловка-пеструшка, пеночки, славки, крапивник. В травянистом .слое живут мыши, землеройки, ежи. Некоторые животные, например серая белка, способны перемещаться почти по всем ярусам.
24. Структура экосистем. Биология. Общая биология. 11 класс. Базовый уровень
24. Структура экосистем
Вспомните!
Какие уровни организации живой природы вам известны?
Что такое экосистема?
Влияние абиотических факторов на живые организмы и взаимодействия между отдельными видами лежат в основе жизни любого сообщества. Сообщество, или биоценоз, – это совокупность сосуществующих популяций разных видов. Вместе с факторами неживой природы (абиотическими факторами) сообщество образует экосистему.
Экосистема – это очень широкое понятие. Дождевой тропический лес и болото, гниющий пень и муравейник, лужа посреди просёлочной дороги и одиноко стоящее дерево с его обитателями – это разные природные экосистемы. Существуют экосистемы искусственного происхождения, например сельскохозяйственные угодья, аквариум, ферма.
Любая экосистема имеет пространственную, видовую и экологическую структуры.
Пространственная структура экосистемы

Рис. 75. Пример ярусности экосистемы. Ярусная структура лиственного леса
В составе любой экосистемы можно выделить два основных структурных компонента: комплекс факторов неживой природы, так называемое абиотическое окружение, или биотоп, и совокупность всех живых организмов – биоценоз. В свою очередь, биоценоз можно подразделить на зооценоз (сообщество животных), фитоценоз (сообщество растений), микробоценоз (сообщество микроорганизмов). Биотоп – это тоже неоднородная система, он состоит из разнообразных абиотических факторов, которые в сумме формируют определённые климатические, географические, почвенные и другие параметры экосистемы.
Видовая структура экосистемы. Биоценоз любой экосистемы характеризуется определённым видовым разнообразием, т. е. числом видов, которые его образуют, и количественным соотношением особей этих видов. Видовое разнообразие обеспечивает стабильность экосистем. Высокая численность популяций, входящих в состав экосистемы, свидетельствует о том, что данные виды оптимально приспособлены к конкретным условиям и важны для стабильного существования этой экосистемы. Обычно общую численность особей в популяциях подсчитать достаточно сложно, поэтому при характеристике экосистем используют понятие
Обычно общую численность особей в популяциях подсчитать достаточно сложно, поэтому при характеристике экосистем используют понятие
Экологическая структура экосистемы. Несмотря на громадное разнообразие экосистем, все они имеют примерно одинаковую экологическую структуру. Экологическая структура – это соотношение групп видов, занимающих определённые экологические ниши и выполняющих определённые функции в сообществе. Наличие этих групп является обязательным условием стабильного существования любой экосистемы, потому что благодаря их взаимодействию обеспечивается главное свойство экосистем – способность к самоподдержанию. Эти обязательные компоненты любой экосистемы – продуценты, консументы и редуценты.
 Биомассу, образованную в экосистеме автотрофными организмами, называют первичной продукцией. Она служит пищей и источником энергии для остальных организмов сообщества.
Биомассу, образованную в экосистеме автотрофными организмами, называют первичной продукцией. Она служит пищей и источником энергии для остальных организмов сообщества.
Основными продуцентами являются зелёные растения, хотя свой вклад в образование первичной продукции экосистемы вносят также фотосинтезирующие и хемосинтезирующие бактерии. Для каждой крупной экосистемы или для любого биогеоценоза характерны свои специфические растения, осуществляющие фотосинтез, т. е. свои продуценты.
Консументы, или потребители, – это гетеротрофные организмы, которые используют синтезированную продуцентами биомассу для собственной жизнедеятельности. Съедая и перерабатывая растения, консументы получают энергию и образуют вторичную продукцию экосистемы.
Консументами являются самые разные живые организмы – от микроскопических бактерий до крупных млекопитающих, от простейших до человека. С точки зрения структуры экосистемы и той роли, которую играют разные консументы в поддержании её равновесного состояния, всех консументов можно подразделить на несколько подгрупп, что мы и сделаем несколько позже, когда будем разбирать пищевые связи экосистем.
Редуценты, или разлагатели, перерабатывают мёртвое органическое вещество (детрит) до минеральных соединений, которые снова могут быть использованы продуцентами. Многие организмы, такие как, например, дождевые черви, многоножки, термиты, муравьи и др., питаются растительными и животными остатками, а часть древесины гниёт и разлагается в процессе жизнедеятельности грибов и бактерий. Когда грибы и другие редуценты отмирают, они сами превращаются в детрит и служат пищей и источником энергии другим редуцентам.
Таким образом, несмотря на многообразие экосистем, все они обладают структурным сходством. В каждой способной к самостоятельному существованию экосистеме есть свои продуценты, различные виды консументов и редуцентов (рис. 76).
Экосистема дубравы. Рассмотрим в качестве примера дубраву – очень устойчивую наземную экосистему (рис. 77). Дубрава является типичным широколиственным лесом ярусной структуры, в котором совместно существуют многие сотни видов растений и несколько тысяч видов животных, микроорганизмов и грибов.
Верхний древесный ярус образуют крупные (до 20 м) многолетние дубы и липы. Эти светолюбивые растения, растущие достаточно свободно, создают благоприятные условия для формирования второго древесного яруса, представленного низкорослыми и менее светолюбивыми грушей, клёном, яблоней.
Рис. 76. Необходимые компоненты экосистемы
Под пологом двух ярусов формируется кустарниковая растительность. Лещина, бересклет, калина, боярышник, терновник, бузина, крушина – это далеко не полный перечень растений, которые образуют третий ярус до высоты 2–4 м.
Следующий, травянистый ярус составляют многочисленные кустарнички и полукустарнички, папоротники, всходы деревьев и разнообразные травы. Причём в течение года в дубраве происходит смена травянистого покрова. Весной, когда листвы на деревьях ещё нет и поверхность почвы ярко освещена, расцветают светолюбивые первоцветы: медуница, хохлатка, ветреница. Летом им на смену приходят теневыносливые растения.
В приземном ярусе, высота которого всего несколько сантиметров от поверхности почвы, растут лишайники, мхи, грибы, низкие травы.
Сотни видов растений (продуцентов), используя энергию солнца, создают зелёную биомассу дубравы. Дубравы очень продуктивны: в течение года на площади в 1 га они создают до 10 т прироста растительной массы.
Мёртвые корни и опавшие листья образуют подстилку, в которой обитают многочисленные редуценты: дождевые черви, личинки мух и бабочек, жуки-навозники и мертвоеды, мокрицы и многоножки, ногохвостки, клещи, нематоды. Питаясь, эти организмы не только преобразуют детрит, но и формируют почвенную структуру. Деятельность таких землероев, как кроты, мыши и некоторые крупные беспозвоночные, не даёт почве слёживаться. В каплях воды между частичками почвы обитают многочисленные почвенные простейшие, а грибы образуют симбиоз с корнями растений и участвуют в разложении детрита.
Рис. 77. Экосистема дубравы
Несмотря на то что ежегодно на 1 га поверхности почвы в дубраве поступает 3–4 т отмерших растений, почти вся эта масса разрушается в результате деятельности редуцентов.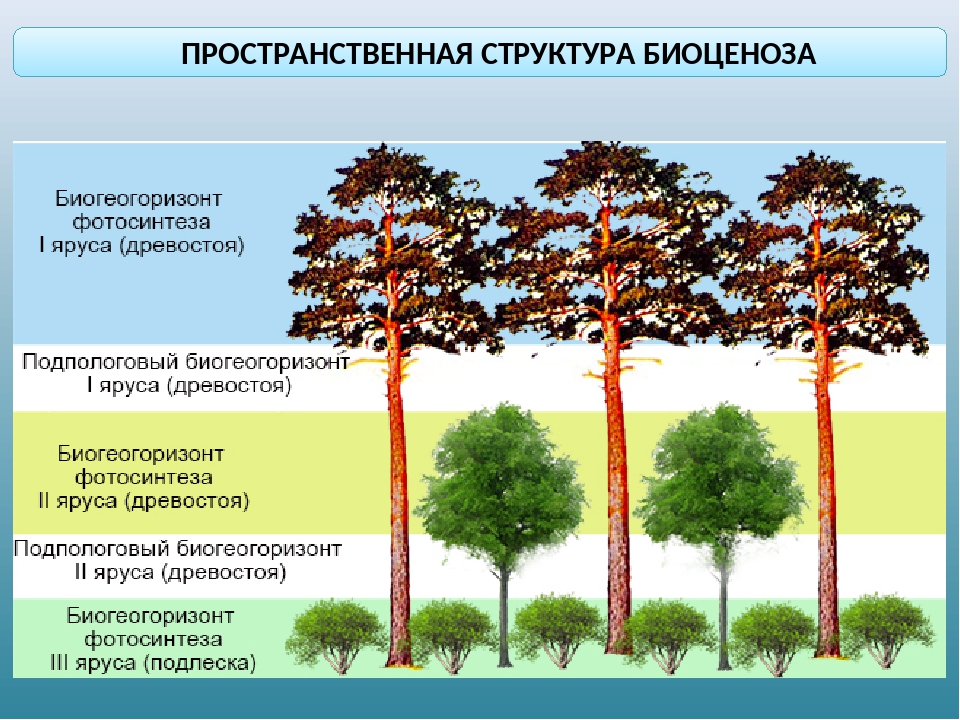 Особая роль в этой переработке принадлежит дождевым червям, которых в дубравах насчитывается огромное количество: несколько сот особей на 1 м2.
Особая роль в этой переработке принадлежит дождевым червям, которых в дубравах насчитывается огромное количество: несколько сот особей на 1 м2.
Разнообразен животный мир верхних ярусов дубравы. В кронах деревьев гнездятся десятки видов птиц. Вьют гнёзда сорока и галка, певчий дрозд и зяблик, большая синица и лазоревка. В дуплах выводят птенцов филин и обыкновенная неясыть. Чеглок и перепелятник наводят страх на мелких певчих птиц. В кустарниках обитают зарянка и чёрный дрозд, мухоловка-пеструшка и поползень. Ещё ниже находятся гнёзда славки и крапивника. По всем ярусам перемещается в поисках пищи серая белка. Бабочки, пчёлы, осы, мухи, комары, жуки – более 1600 видов насекомых тесно связаны с дубом! В травяном ярусе делят место под солнцем кузнечики и жуки, пауки и сенокосцы, мыши, землеройки и ежи. Самыми крупными консументами этой экосистемы являются косули, лани и кабаны.
Устойчивость этой и любой другой экосистемы обеспечивает сложная система взаимоотношений всех организмов, входящих в её состав.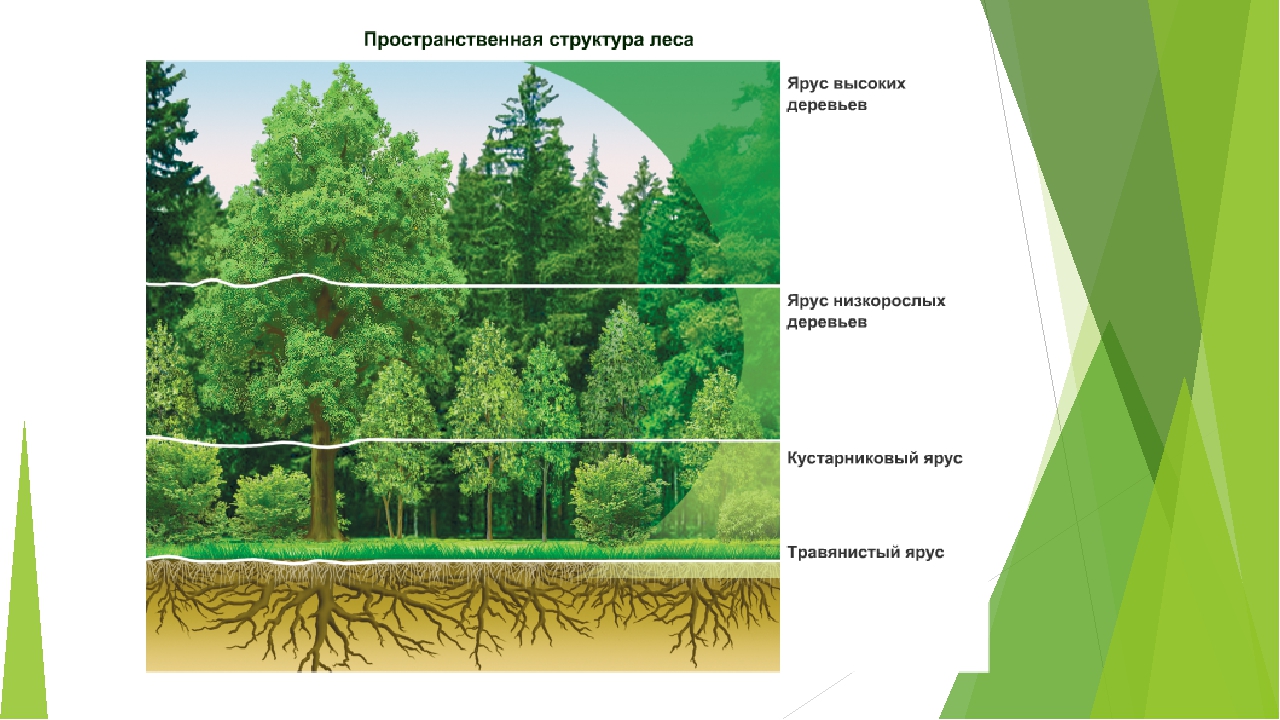
Вопросы для повторения и задания
1. Что такое биогеоценоз?
2. Расскажите о пространственной структуре экосистемы.
3. Какие обязательные компоненты включает любая экосистема?
4. В каких отношениях находятся друг с другом обитатели биоценозов? Охарактеризуйте эти связи.
5. Опишите видовой состав и пространственную структуру экосистемы дубравы.
Подумайте! Выполните!
1. Назовите общие черты биогеоценозов лиственного леса и пресноводного водоёма.
2. Возможно ли существование биоценоза, состоящего только из растений? Обоснуйте свою точку зрения.
3. Выполните исследование на тему «Моё жильё как пример экосистемы».
4. Разработайте экскурсионный маршрут, позволяющий продемонстрировать видовую, пространственную и экологическую структуры типичной экосистемы вашего региона (групповой проект).
Работа с компьютером
Обратитесь к электронному приложению. Изучите материал и выполните задания.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРесСТРУКТУРА ЭКОСИСТЕМЫ
- Каковы обязательные компоненты экосистемы?
- Чем определяются видовая и пространственная структуры экосистемы?
Структура (от лат. структура — строение, расположение) — это определённая взаимосвязь составных частей, обеспечивающая целостность объекта. Как было изложено ранее, в составе любой экосистемы можно выделить два основных структурных блока: комплекс факторов неживой природы, так называемое абиотическое окружение, или экотоп, и совокупность всех живых организмов — биоценоз. Экотоп — это не однородная система, он состоит из разнообразных абиотических факторов, которые в сумме определяют климатические, географические, почвенные и другие характеристики экосистемы. Однако это не единственная структурная характеристика экосистемы.
Однако это не единственная структурная характеристика экосистемы.
Экологическая ниша. Несмотря на то, что в пределах любой экосистемы на одной и той же территории обитает множество популяций, каждая из них адаптирована к строго определённым условиям существования (рис. 143). Как вы уже знаете, совокупность всех необходимых для данной популяции условий называют её экологической нишей. Часто говорят, что экологическая ниша вида определяется его «адресом», «профессией» и «стилем жизни». Ни в одной экологической нише не может одновременно существовать больше одного вида организмов. Существует и обратное правило: пустующая ниша обязательно будет заполнена. Популяции, занимающие одни и те же экологические ниши, никогда не смогут ужиться на одной территории. В своё время в Европе повсеместно была распространена чёрная крыса, однако серые крысы, приплывшие на кораблях из Китая, сильно потеснили своих чёрных собратьев, поскольку их экологические ниши в значительной степени совпадали.
В соответствии с числом и расположением экологических ниш популяций можно охарактеризовать структуру экосистемы.
Видовая структура экосистемы. Видовая структура экосистемы определяется числом видов, которые входят в её состав, и количественным соотношением особей этих видов. В благоприятных условиях может существовать большое количество экологических ниш, которые заселяются многочисленными видами. Обычно же в сообщество входит множество относительно малочисленных видов и сравнительно немного видов, общая численность и биомасса которых велики. Высокая численность популяций свидетельствует о том, что конкретные виды оптимально приспособлены к конкретным условиям и важны для стабильного существования данной экосистемы.
Видовое разнообразие обеспечивает стабильность экосистем. Чем больше видовое разнообразие, тем больше у экосистемы возможностей приспособиться к изменяющимся условиям, следовательно, тем больше её устойчивость. В дальнейшем мы увидим, что искусственные системы, создаваемые человеком, в которых специально поддерживается очень Цизкий уровень видового разнообразия, обладают и очень низкой устойчивостью.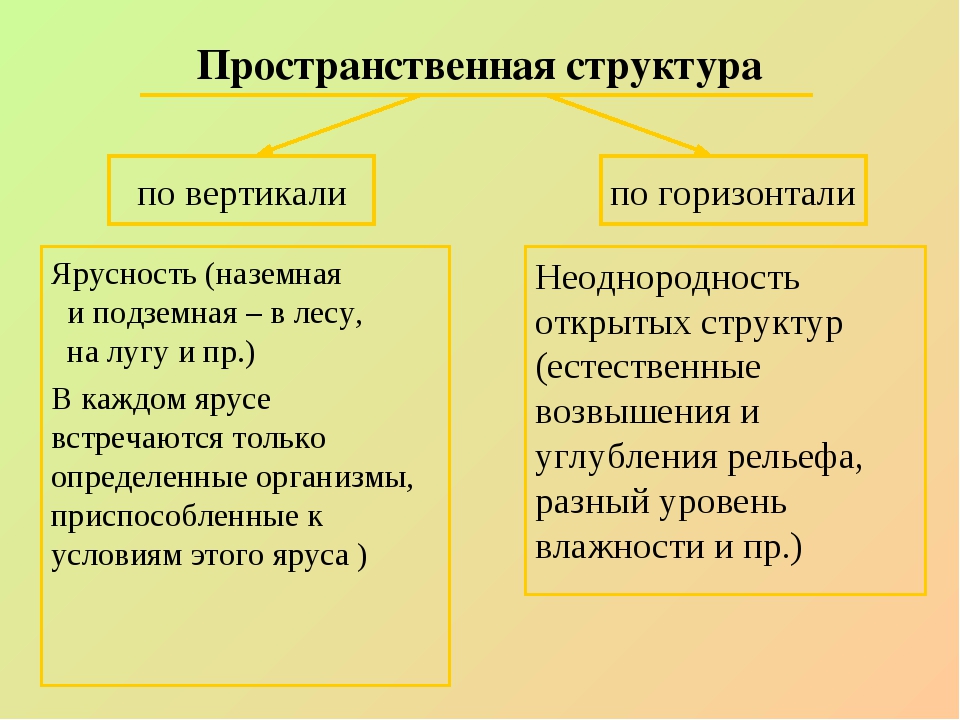
Пространственная структура экосистемы. Пространственная структура большинства биогеоценозов и, следовательно, экосистем определяется ярусным расположением растительности. Например, в типичном листопадном лесу выделяют древесный, кустарниковый, травяной и надпочвенный (приземный) ярусы (рис. 144). Углубляясь в почву, также можно обнаружить «этажи», образованные корнями определённых растений, где обитают разные виды подземных животных. Подобная пространственная организация экосистемы позволяет растениям эффективно использовать солнечный свет и ресурсы почвы, а многочисленным животным и микроорганизмам — сосуществовать вместе, занимая разнообразные экологические ниши. Даже такие подвижные обитатели леса, как птицы, предпочитают жить и тем более гнездиться на определённой, «своей» высоте. Часто родственные виды птиц обитают в различных ярусах леса для того, чтобы разделить свои экологические ниши.
Вертикальная ярусность существует и в водоёмах, где верхние слои воды имеют большую освещённость и более высокую температуру. На поверхностной плёнке водоёма обитают организмы, называемые нейстоном. Организмы, обитающие в толще воды и пассивно переносимые течением, — это планктон. Различают фитопланктон, состоящий из мелких водорослей, и зоопланктон, в состав которого входят простейшие животные, мелкие ракообразные, некоторые личинки и пр. Активно передвигающиеся в толще воды животные, такие как рыбы и китообразные, образуют нектон. И наконец, на самом дне или возле него обитают животные (кораллы, некоторые ползающие рыбы и моллюски), которых называют бентосом.
На поверхностной плёнке водоёма обитают организмы, называемые нейстоном. Организмы, обитающие в толще воды и пассивно переносимые течением, — это планктон. Различают фитопланктон, состоящий из мелких водорослей, и зоопланктон, в состав которого входят простейшие животные, мелкие ракообразные, некоторые личинки и пр. Активно передвигающиеся в толще воды животные, такие как рыбы и китообразные, образуют нектон. И наконец, на самом дне или возле него обитают животные (кораллы, некоторые ползающие рыбы и моллюски), которых называют бентосом.
Ещё одной важной характеристикой экосистемы является её трофическая структура, которую мы рассмотрим в следующем параграфе.
Запомнить: экологическая ниша; видовая структура экосистемы, пространственная структура экосистемы; ярусное расположение растительности; нейстон, планктон, нектон, бентос.
ВыводыВ соответствии с числом и расположением экологических ниш популяций можно охарактеризовать структуру экосистемы.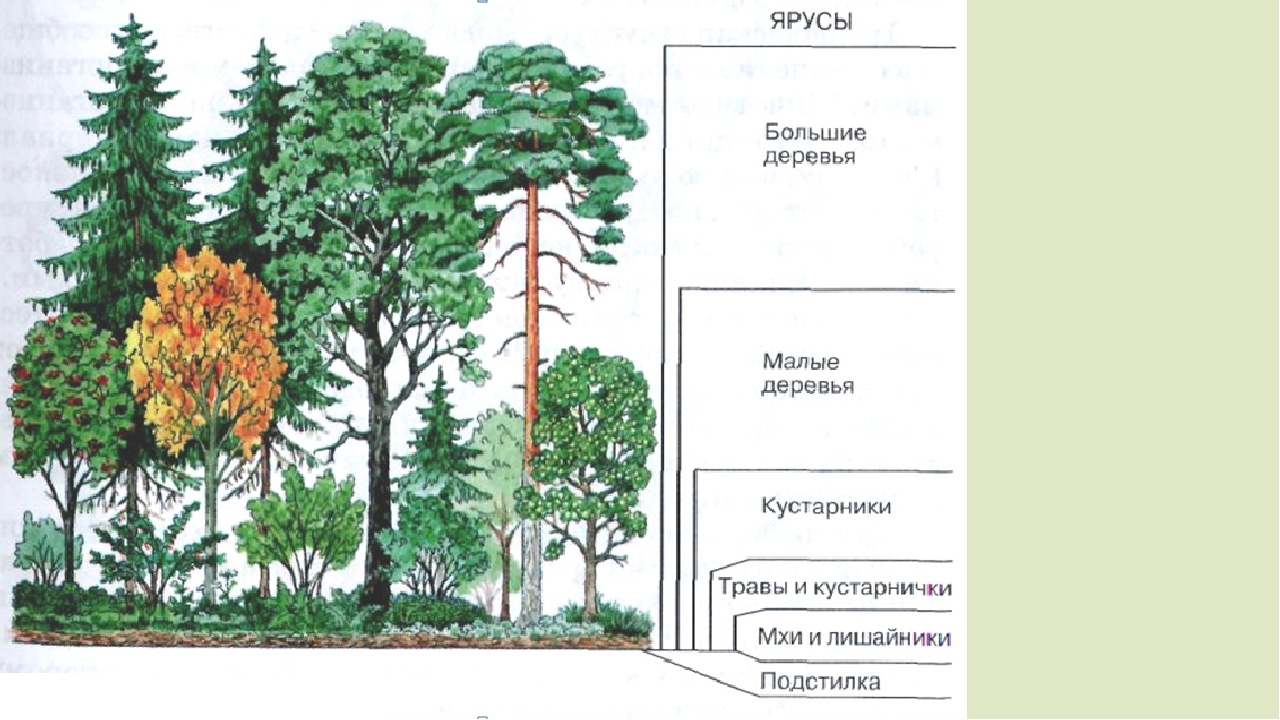 Видовая структура определяется числом видов, которые входят в состав экосистемы, и количественным соотношением особей этих видов. Пространственная структура большинства экосистем определяется ярусным расположением растительности.
Видовая структура определяется числом видов, которые входят в состав экосистемы, и количественным соотношением особей этих видов. Пространственная структура большинства экосистем определяется ярусным расположением растительности.
- Что такое экологическая ниша?
- Какова роль видового разнообразия в поддержании стабильного состояния экосистемы?
- Почему искусственные экосистемы менее устойчивые, чем природные?
- Чем определяется пространственная структура экосистем?
- Чем планктон отличается от нектона?
Выполни задания
- Перечислите основные ярусы растительности в экосистемах вашего края.
- Опишите видовой состав и пространственную структуру одной из экосистем вашего края.
Где больше видовое разнообразие: на отдельном острове или на архипелаге? Объясните почему.
Много ли пустующих экологических ниш можно обнаружить в экосистеме
что такое, трофические уровни, кто ввел термин в науку
История возникновения термина, определение понятия
Любая наука оперирует терминами и понятиями. Смысл их и содержание раскрывается, объясняется в определениях. Понятие «Экосистема» – одно из основных в экологии – науке о том, как живые организмы взаимодействуют между собой и с окружающей средой. Можно дать определение: «Что такое «Экосистема»?» после ответа на вопрос: «Кто термин «экосистема» ввел и для чего?».
Биолог Эрнст Геккель в 1866 году в своей работе «Общая морфология организмов» впервые употребил термин «Экология», описывая взаимоотношения живой природы с неживой. Через одиннадцать лет другой немецкий ученый, гидробиолог, К. Мебиус, описывая сообщество живых организмов в устричной банке, назвал его «Биоценоз». Год спустя американец С. Форбс называет совокупность живых организмов, обитающих в определенном месте, «микрокосм». Об этом он говорит в своей книге «Озеро как микрокосм». В науку ввел термин «Экосистема» Артур Тенсли. Это произошло в 1935 году, с того времени и существует современное понимание этого термина. Некоторые ученые, в том числе русские В.В. Докучаев и В.Н. Сукачев, в своих работах вышеприведенный термин предпочитают заменять другими – «Биоценоз» и «Биогеоценоз». Тоже касается и наук смежных с биологией, где понятие, имеющее подобное определение и содержание, называют «геосистема», «голоцен» или «биокосное тело». Последнее понятие было введено в том же 1944 году В.И. Вернадским, что и «биогеоценоз» В.Н. Сукачевым.
Об этом он говорит в своей книге «Озеро как микрокосм». В науку ввел термин «Экосистема» Артур Тенсли. Это произошло в 1935 году, с того времени и существует современное понимание этого термина. Некоторые ученые, в том числе русские В.В. Докучаев и В.Н. Сукачев, в своих работах вышеприведенный термин предпочитают заменять другими – «Биоценоз» и «Биогеоценоз». Тоже касается и наук смежных с биологией, где понятие, имеющее подобное определение и содержание, называют «геосистема», «голоцен» или «биокосное тело». Последнее понятие было введено в том же 1944 году В.И. Вернадским, что и «биогеоценоз» В.Н. Сукачевым.
Каких бы названий ни придерживались те или иные ученые, экологическая система или экосистема представляет собой совокупность сообщества живых организмов, среды, где они обитают, и связей и взаимодействий между ними, в том числе по обмену веществом и энергией.
Несколько шире понятие «Экосистемы», его можно определить следующим образом: под сообществом живых организмов подразумеваются все представители животного и растительного мира, от крупных млекопитающих, птиц, рыб, беспозвоночных до червей, личинок и микроорганизмов. Под средой обитания надо понимать конкретный ареал с лишь ему присущими химическими и физическими свойствами воды, почвы и воздуха, температурным, световым и климатическим режимом. А под связями и взаимодействиями — видовую продуктивность и кругооборот трофической цепи, в результате которых происходит обмен химическими веществами и соединениями и энергией.
Под средой обитания надо понимать конкретный ареал с лишь ему присущими химическими и физическими свойствами воды, почвы и воздуха, температурным, световым и климатическим режимом. А под связями и взаимодействиями — видовую продуктивность и кругооборот трофической цепи, в результате которых происходит обмен химическими веществами и соединениями и энергией.
Понятие «Экосистемы» дает наиболее общее представление о том, что биосистема имеет составные части.
Структура и ее виды
Эти части и есть структура экосистемы, которая может быть классифицирована на несколько категорий. Первая – наиболее общее разделение — живая и неживая природа. Вторая — роль живого организма в пищевой цепи. Третья – видовое наполнение каждого этапа этой цепи и пространственное их распределение.
Первая категория содержит два элемента. Биотический – живая природа и абиотический – неживая.
В свою очередь, биотический подразделяется на организмы, которые получая солнечную энергию, синтезируют из неорганических веществ органические.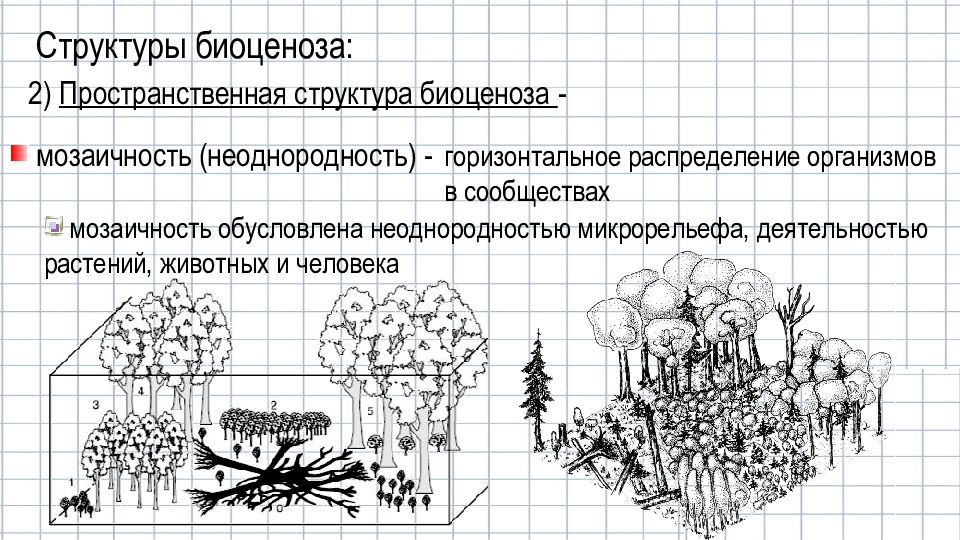 Такие организмы названы автотрофами или продуцентами, производителями. Благодаря им производится первичные органические вещества, необходимые для существования последующих звеньев пищевой цепи. Основной характерной их особенностью является способность к фотосинтезу. Такая способность обусловлена наличием у растений хлорофилла, а у бактерий еще и ородопсина. Эти фотосинтетические пигменты на свету из углекислого газа и воды образовывают органические вещества. Продуценты усваивают солнечную энергию с эффективностью 0,1 – 1%. Есть некоторые виды растений, у которых этот показатель значительно выше – до 4,5%.
Такие организмы названы автотрофами или продуцентами, производителями. Благодаря им производится первичные органические вещества, необходимые для существования последующих звеньев пищевой цепи. Основной характерной их особенностью является способность к фотосинтезу. Такая способность обусловлена наличием у растений хлорофилла, а у бактерий еще и ородопсина. Эти фотосинтетические пигменты на свету из углекислого газа и воды образовывают органические вещества. Продуценты усваивают солнечную энергию с эффективностью 0,1 – 1%. Есть некоторые виды растений, у которых этот показатель значительно выше – до 4,5%.
Автотрофные продуценты или производители могут считаться первым звеном пищевой цепи или ее первым уровнем. Без вырабатываемых ими органических веществ и соединений, в том числе кислорода, рождение и существование жизни на Земле было бы невозможно.
Вторым видом организмов, входящих в биотическую часть трофической структуры экосистемы, являются гетеротрофы. Это живые организмы, которые черпают энергию, необходимую им для существования, из органических веществ. К ним относятся два вида – консументы или потребители и редуценты или разрушители.
К ним относятся два вида – консументы или потребители и редуценты или разрушители.
Консументы или потребители – это живые организмы, которые питаются органическим веществом и другими живыми организмами. Они являются вторым звеном пищевой цепи.
В процессе существования живые организмы выделяют органику. Они и сами после гибели не перестают быть органическим веществом. Все эти выделения и останки опадают на землю и дно водных объектов, где становятся пищей для редуцентов, могильщиков или разрушителей. Это последнее звено пищевой цепи, потому что разрушители перерабатывают органическое вещество в неорганическое, минеральное. Которое вновь начинает свое движение в вышестоящие экосистемы трофические уровни.
Редуценты являются последним звеном пищевой цепи. Трофическая структура экосистемы, разделенная на уровни, с точки зрения временного фактора, ими замыкается.
Они же существуют в нижнем ярусе пространственной структуры экосистемы.
Пространственная и видовая
Пространственная структура подразумевает распределение живых организмов на определенной территории в вертикальной и горизонтальной плоскости.
Растения являются основным и определяющим фактором для формирования и существования любой экосистемы. Они же диктуют видовое распределение по вертикали или высоте. Растения одной высоты формируют ярусы. Высота этих ярусов может существенно различаться. Это зависит от многих факторов, и, прежде всего, от абиотических. Ярусы есть, как в высокорослых тропических лесах, так и зарослях кустарников степных зон и покрытых мхами болотах. В лесах они ярче выражены и их больше.
Верхний ярус занимают высокие деревья. Для некоторых климатических зон высота верхнего яруса может превышать 55 метров. Несколько ниже располагается второй ярус. Там деревья ниже. Третий – это кустарники. Четвертый – травы, мхи и лишайники. Почва является лишь условной границей, так сказать, видимой. Но и она имеет свою подземную, ярусность, которая связана с глубиной проникновения корней растений.
Существует также горизонтальное пространственное распределение. Оно еще имеет название «мозаичное». Факторами, формирующими такое своеобразие, может быть различный состав почвы, рельеф, влажность и тому подобное. Потому растения группируются на плоскости, приспосабливаясь к условиям своего произрастания и, таким образом, создают эффект сложенной мозаики или орнамента.
Потому растения группируются на плоскости, приспосабливаясь к условиям своего произрастания и, таким образом, создают эффект сложенной мозаики или орнамента.
Животные, птицы, насекомые и другие живые организмы распределяются по ярусам и в горизонтальной плоскости в зависимости от приспосабливаемости каждого их вида к созданным условиям. Это и есть видовая структура экосистемы.
Видовая структура экосистемы включает в себя разнообразие видов растительного и животного мира, численность особей в каждом и взаимосвязь между ними. Эти показатели зависят от множества факторов. Основным, из которых является влияние неживой природы – количество света, воды, состав почв, температура, направление и сила ветров, атмосферное давление и так далее. Второй по значимости влияния фактор – это сформировавшийся растительный мир на данной территории. И третий, которые ворвался в природу относительно недавно, — это человек, его деятельность, степень активности и направленность.
Например: на 100 м2 в тайге 30 видов растений, на лугу вдоль реки в два раза больше, а в Амазонском лесу — в сто. Вмешательство человека, приводит к сокращению видового разнообразия, количественного состава популяций, генетическим изменениям и даже, к вымиранию животного и растительного мира и гибели всей экосистемы.
Вмешательство человека, приводит к сокращению видового разнообразия, количественного состава популяций, генетическим изменениям и даже, к вымиранию животного и растительного мира и гибели всей экосистемы.
Видео — Водная экосистема
Структура экосистем 👍 | Школьные сочинения
Вопрос 1. Что такое биогеоценоз?
Биогеоценоз – это экологическая система (экосистема), границы которой определены растительным сообществом. Совокупность биогеоценозов земного шара образует глобальную систему, биосферу. Примерами биогеоценозов являются дубрава, луг, ельник, березовая роща и др.
Вопрос 2. Расскажите о пространственной структуре экосистемы.
Пространственная структура экосистемы определяется ярусным расположением растительности. Сверху вниз над землей выделяют пологовый (древесный), кустарниковый, травяной
и надпочвенный (приземный) ярусы. В почве также идет разделение на уровни, образованные корнями растений разных видов.
В почве также идет разделение на уровни, образованные корнями растений разных видов.Такая пространственная организация позволяет растениям эффективно использовать свет и другие ресурсы, а животным – занимать разные экологические ниши и ослаблять конкуренцию между близкими видами.
Вопрос 3. Какие обязательные компоненты включает любая экосистема?
В любой экосистеме можно выделить два основных структурных компонента – биотоп и биоценоз. Биотоп – это комплекс факторов неживой природы, которые в сумме формируют определенные климатические, географические,
почвенные и другие параметры экосистемы. Биоценоз – это совокупность всех живых организмов (популяций) экосистемы.Он разделяется на зооценоз (сообщество животных), фитоценоз (сообщество растений) и микробоценоз (сообщество микроорганизмов).
Вопрос 4. В каких отношениях находятся друг с другом обитатели биоценозов? Охарактеризуйте эти связи.
С точки зрения экологической структуры в экосистеме можно выделить три группы особей.
1) Продуценты, или производители, – автотрофы, синтезирующие органические вещества из неорганических. Их биомасса – это первичная продукция экосистемы, которая служит пищей и источником энергии для всех остальных организмов сообщества.
Автотрофами являются растения, фотосинтезирующие и хемосинтезирующие прокариоты.
2) Консументы, или потребители, – гетеротрофные организмы, использующие первичную биомассу или биомассу других консументов для собственной жизнедеятельности. Консументами являются многие животные, паразитические бактерии, хищные растения.
3) Редуценты, или разлагатели, – организмы, перерабатывающие мертвое органическое вещество (детрит) до минеральных соединений. Редуценты – это дождевые черви, многоножки, термиты, грибы, бактерии.
Таким образом, продуценты используют неорганические соединения, создавая первичную продукцию. Далее часть консументов (растительноядные животные) используют первичную продукцию, создавая вторичную биомассу, которая используется другими консументами (хищники, паразиты). После смерти продуценты и консументы разлагаются редуцентами до минеральных соединений, которые вновь используются продуцентами.
После смерти продуценты и консументы разлагаются редуцентами до минеральных соединений, которые вновь используются продуцентами.
Вопрос 5. Опишите видовой состав и пространственную структуру экосистемы дубравы.
Видовой состав дубравы очень разнообразен, что обеспечивает ее стабильность как экосистемы. Растения дубравы образуют четко разделенные ярусы. В верхнем древесном “этаже” расположены крупные многолетние дубы и липы. Второй ярус составляют низкорослые и менее светолюбивые груша, клен, яблоня.
Далее следует кустарниковая растительность: лещина, бересклет, калина, боярышник, бузина. Травянистый ярус состоит из кустарничков, всходов деревьев, папоротников и различных трав (медуница, хохлатка, ветреница, иван-да-марья и т. д.). Приземный ярус представлен мхами, низкими травами; здесь также обитают лишайники и грибы.
Животный мир дубравы не менее разнообразен. Членистоногие населяют все ее уровни. Из позвоночных в верхних ярусах мы встретим много птиц – сороку, зяблика, дроздов, синиц, ночных и дневных хищников.
В кустарниках обитают зарянка, мухоловка-пеструшка, пеночки, славки, крапивник. В травянистом слое живут мыши, землеройки, ежи. Некоторые животные, например серая белка, способны перемещаться почти по всем ярусам.
В подстилке преобладают редуценты: дождевые черви, мокрицы, личинки мух, жуки – навозники и мертвоеды, клещи, нематоды. В почве обитают различные животные, роющие землю (например, кроты).
Структура экосистем. Биогеоценоз. Компоненты экосистемы
Стр.146. Вспомните.
1. Какие уровни организации живой природы вам известны?
1. Молекулярный (Например молекулы жиров).
2. Клеточный (эритроцит – красная клетка крови).
3. Тканевый – это уже не одна клетка, а группа клеток сходных по строению, происхождению и выполняющих одинаковые функции. (Например у животных нервная, эпителиальная, жировая, кровь и лимфа, а у растений проводящая, покровная, механическая и образовательная).
4. Органный (Орган – это несколько тканей, соединенных между собой ) Например: сердце, печень, почки…
5. Организменный (Например, человек, собака, кошка).
6. Видовой (вид Homo Sapiens).
7. Популяционный (один вид на одной территории частично или полностью изоллированый от других таких популяций).
8. Биоценозный.
Иногда говорят, что существует и биосферный уровень, но на нем располагаются все живые организмы планеты и косное вещество (атмосфера, гидросфера и литосфера).
2. Что такое экосистема?
Экосистема – это биологическая система, состоящая из живых организмов (биоценоз), среды их обитания (биотоп), системы связей, осуществляющей обмен и энергией между ними.
Стр. 151. Вопросы для повторения и задания.
1. Что такое биогеоценоз?
Биогеоценоз – это экологическая система (экосистема), границы которой определены растительным сообществом. Совокупность биогеоценозов земного шара образует глобальную систему, биосферу. Примерами биогеоценозов являются дубрава, луг, ельник, березовая роща и др.
Совокупность биогеоценозов земного шара образует глобальную систему, биосферу. Примерами биогеоценозов являются дубрава, луг, ельник, березовая роща и др.
2. Расскажите о пространственной структуре экосистемы.
Пространственная структура экосистемы определяется ярусным расположением растительности. Сверху вниз над землей выделяют налоговый (древесный), кустарниковый, травяной и надпочвенный (приземный) ярусы. В почве также идет разделение на уровни, образованные корнями растений разных видов. Такая пространственная организация позволяет растениям эффективно использовать свет и другие ресурсы, а животным – занимать разные экологические ниши и ослаблять конкуренцию между близкими видами.
3. Какие обязательные компоненты включает любая экосистема?
В любой экосистеме можно выделить два основных структурных компонента – биотоп и биоценоз. Биотоп – это комплекс факторов неживой природы, которые в сумме формируют определенные климатические, географические, почвенные и другие параметры экосистемы. Биоценоз – это совокупность всех живых организмов (популяций) экосистемы. Он разделяется на зооценоз (сообщество животных), фитоценоз (сообщество растений) и микробоценоз (сообщество микроорганизмов).
Биоценоз – это совокупность всех живых организмов (популяций) экосистемы. Он разделяется на зооценоз (сообщество животных), фитоценоз (сообщество растений) и микробоценоз (сообщество микроорганизмов).
4. В каких отношениях находятся друг с другом обитатели биоценозов? Охарактеризуйте эти связи.
С точки зрения экологической структуры в экосистеме можно выделить три группы особей
1. Продуценты (производители) – автотрофные организмы, главным образом зеленые растения, синтезирующие органические вещества из неорганических. Их биомасса – это первичная продукция экосистемы, которая служит пищей и источником энергии для всех остальных организмов сообщества. Автотрофами являются растения, фотосинтезирующие и хемосинтезирующие прокариоты.
2. Консументы (потребители) – гетеротрофные организмы (растительноядные и плотоядные), потребители живого органического вещества. Консументами являются многие животные, паразитические бактерии, хищные растения.
3. Редуценты (разлагатели) – гетеротрофные организмы, которые разрушают мертвые остатки животных и растений и превращают их в простые соединения. Редуценты – это дождевые черви, многоножки, термиты, грибы, бактерии.
Таким образом, продуценты используют неорганические соединения, создавая первичную продукцию. Далее часть консументов (растительноядные животные) используют первичную продукцию, создавая вторичную биомассу, которая используется другими консументами (хищники, паразиты). После смерти продуценты и консументы разлагаются редуцентами до минеральных соединений, которые вновь используются продуцентами.
5. Опишите видовой состав и пространственную структуру экосистемы дубравы.
Видовой состав дубравы очень разнообразен, что обеспечивает ее стабильность как экосистемы. Растения дубравы образуют четко разделенные ярусы. В верхнем древесном «этаже» расположены крупные многолетние дубы и липы. Второй ярус составляют низкорослые и менее светолюбивые груша, клен, яблоня. Далее следует кустарниковая растительность: лещина, бересклет, калина, боярышник, бузина. Травянистый ярус состоит из кустарничков, всходов деревьев, папоротников и различных трав (медуница, хохлатка, ветреница, иван-да-марья и т. д.). Приземный ярус представлен мхами, низкими травами; здесь также обитают лишайники и грибы. Животный мир дубравы не менее разнообразен. Членистоногие населяют все ее уровни. Из позвоночных в верхних ярусах мы встретим много птиц – сороку, зяблика, дроздов, синиц, ночных и дневных хищников. В кустарниках обитают зарянка, мухоловка-пеструшка, пеночки, славки, крапивник. В травянистом .слое живут мыши, землеройки, ежи. Некоторые животные, например серая белка, способны перемещаться почти по всем ярусам. В подстилке преобладают редуценты: дождевые черви, мокрицы, личинки мух, жуки-навозники и мертвоеды, клещи, нематоды. В почве обитают различные животные, роющие землю (например, кроты).
Далее следует кустарниковая растительность: лещина, бересклет, калина, боярышник, бузина. Травянистый ярус состоит из кустарничков, всходов деревьев, папоротников и различных трав (медуница, хохлатка, ветреница, иван-да-марья и т. д.). Приземный ярус представлен мхами, низкими травами; здесь также обитают лишайники и грибы. Животный мир дубравы не менее разнообразен. Членистоногие населяют все ее уровни. Из позвоночных в верхних ярусах мы встретим много птиц – сороку, зяблика, дроздов, синиц, ночных и дневных хищников. В кустарниках обитают зарянка, мухоловка-пеструшка, пеночки, славки, крапивник. В травянистом .слое живут мыши, землеройки, ежи. Некоторые животные, например серая белка, способны перемещаться почти по всем ярусам. В подстилке преобладают редуценты: дождевые черви, мокрицы, личинки мух, жуки-навозники и мертвоеды, клещи, нематоды. В почве обитают различные животные, роющие землю (например, кроты).
Подумайте и выполните.
1. Назовите общие черты биогеоценозов лиственного леса и пресноводного водоёма.
Назовите общие черты биогеоценозов лиственного леса и пресноводного водоёма.
1. имеются продуценты, консументы, редуценты
2. имеются факторы неживой природы-свет, влажность, температура
3. биотические факторы (различные взаимоотношения организмов-симбиоз конкуренция, и т.д.)
4. есть влияние человека (антропогенные факторы)
5. есть цепи питания, пищевые сети
6. способны к саморегуляции (и лес и водоем)
7. они открытые системы, устойчивые
2. Возможно ли существование биоценоза, состоящего только из растений? Обоснуйте свою точку зрения.
Существование биоценоза, состоящего только из растений, невозможно. Пищевых цепей там быть не может, потому что не присутствуют ни консументы, ни редуценты, следовательно существование такого сообщества не возможно.
Понятие о биоценозе. Структура биоценоза (пространственная, видовая, экологическая, трофическая).
 Виды эдификаторы.
Виды эдификаторы.Биоценоз – это исторически сложившаяся организованная группа популяций растений, животных и микроорганизмов, живущих совместно в одних и тех же условиях среды, возникшая на основе биогенного круговорота и обеспечивающая его в конкретных условиях среды.
Видовая структура – это число видов, образующих данный
биоценоз. Видовую структуру рассматривают
также как соотношение видов по численности
или биомассе. Биоценозы в отношении
видового разнообразия бывают бедные и
богатые. Богатыми считаются тропические
леса, шельфовые зоны, некоторые островные
сообщества. Бедные биоценозы приурочены
к тундровым зонам, высокогорьям и к
пустынным районам Земли. Различают α-
и β-разнообразие.α-разнообразие– видовое
разнообразие в данном местообитании;
β-разнообразие– сумма всех видов всех
местообитаний в данном регионе. В каждом
биоценозе можно выделить виды доминанты – или виды, преобладающие по численности;
и виды эдификаторы – виды
средообразователи, или виды создающие
условия для жизни других видов.
В каждом
биоценозе можно выделить виды доминанты – или виды, преобладающие по численности;
и виды эдификаторы – виды
средообразователи, или виды создающие
условия для жизни других видов.
Пространственная структура – это распределение организмов в биоценозе по вертикали и по горизонтали. При вертикальной градации биоценоза выделяют ярусы. Ярусность может быть как надземная, так и подземная. В любом биоценозе могут быть и внеярусные растения, например лианы.
При горизонтальном
членении биоценоза можно говорить о
его мозаичности. Горизонтально
выделенные структурные элементы
биоценоза получили наименование синузий.
Они образуются потому, что растения,
распределяясь неравномерно, создают
то большие, то меньшие скопления,
придающие растительному покрову
мозаичный характер. Парцелла – это
структурная часть при горизонтальном
членении биоценоза, отличающаяся
составом и свойствами компонентов,
спецификой их связей и материально-энергетическим
обменом.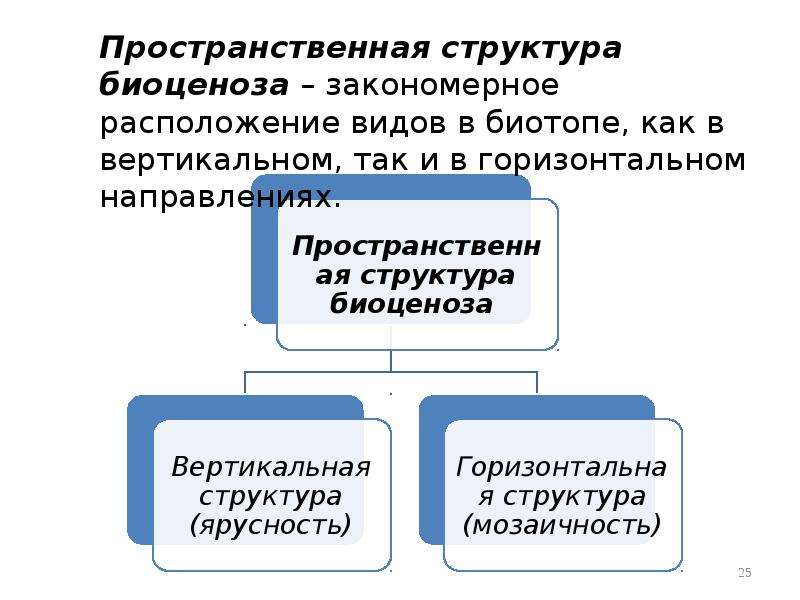
Экологическая структура – это состав биоценоза из экологических групп организмов, выполняющих в сообществе в каждой экологической нише определенные функции. Например, экологическую структуру биоценоза отражает соотношение групп организмов объединенных сходным типом питания. В лесах преобладают сапрофаги, в степях и полупустынях – фитофаги, в глубинах Мирового океана – хищники и детритоеды. Разные экологические группы по отношению к абиотическим факторам также преобладают в определенных типах сообществ. Например, на болоте чаще встретишь растения гигрофиты; в степях и полупустынях – ксерофиты; во влажном тропическом лесу – мезофиты, мезо-гигрофиты, гигро-гидрофиты и т.д.
БИЛЕТ № 5
Понятие экосистемы (а. Тенсли) и биогеоценоза (в.Н. Сукачев). Сходство и отличие понятий. Структура экосистемы (видовая, пространственная, трофическая, экологическая).
Любую совокупность
организмов и неорганических компонентов,
в которой может осуществляться круговорот
веществ, называют экосистемой. Термин
был предложен в 1935 г. английским экологом
А. Тенсли, который подчеркивал, что при
таком подходе неорганические и
органические факторы выступают как
равноправные компоненты.
Термин
был предложен в 1935 г. английским экологом
А. Тенсли, который подчеркивал, что при
таком подходе неорганические и
органические факторы выступают как
равноправные компоненты.
«Биогеоценоз – это совокупность на известном протяжении земной поверхности однородных природных явлений (атмосферы, горной породы, растительности, животного мира и мира микроорганизмов, почвы и гидрологических условий), имеющих свою специфику взаимодействия этих слагаемых ее компонентов и определенный тип обмена веществами и энергией между собой и другими явлениями природы и представляющая собой внутренне противоречивое единство, находящееся в постоянном движении, развитии» (В. Н. Сукачев, 1964).
«Экосистема» и «биогеоценоз» – близкие по сути понятия, но если первое из них приложимо для обозначения систем, обеспечивающих круговорот любого ранга, то «биогеоценоз» – понятие территориальное, относимое к таким участкам суши, которыезаняты определенными единицами растительного покрова – фитоценозами.
Видовая структура представляет собой совокупность всех видов, составляющих экосистему, их взаимосвязь между собой и соотношение численности. В одних случаях первенство — за животными, например, биоценоз кораллового рифа, в других ведущую роль играют растения (пойменные луга, дубовые и еловые леса, ковыльная степь). Видовая структура экосистемы отражает ее состав в том числе и по количеству видов. Он зависит главным образом от географического положения места. Наиболее известная закономерность заключается в том, что чем ближе к экватору, тем флора и фауна разнообразнее.
Популяции разных видов в экосистеме распределены определенным образом – образуют пространственную структуру. Различают вертикальную и горизонтальную структуры экосистемы. Основу вертикальной структуры формирует растительность.
Растительное сообщество определяет, как правило, облик экосистемы. Растения в значительной мере влияют на условия существования остальных видов. В лесу это крупные деревья, на лугах и в степях — многолетние травы, а в тундрах господствуют мхи и кустарнички.
Обитая совместно, растения одинаковой высоты создают своего рода этаж -ярусы. Благодаря вертикальной и горизонтальной структурам обитающие в экосистеме организмы более эффективно используют минеральные вещества почвы, влагу, световой поток.
Виды, входящие в состав экосистемы, связаны между собой пищевыми связями, так как служат объектами питания друг для друга. Благодаря пищевым взаимоотношениям в экосистеме осуществляется трансформация биогенных веществ и энергии. Последовательность питающихся друг другом организмов или пути, по которым осуществляется постоянный поток веществ и энергии, называют цепями питания или пищевой, трофической цепью. Этот термин был предложен Чарльзом Элтоном (1934).
Цепи питания составляют трофическую структуру любой экосистемы. Каждая цепь состоит из нескольких звеньев, которые называются трофическим уровнем, последовательность которых соответствует потоку энергии:
Первый трофический уровень − продуценты (автотрофные организмы, преимущественно зеленые растения).
Второй трофический уровень − консументы первого порядка(растительноядные животные).
Третий трофический уровень − консументы второго порядка (первичные хищники, питающиеся растительноядными животными).
Четвертый трофический уровень − консументы третьего порядка(вторичные хищники, питающиеся плотоядными животными).
Пространственная структура в экосистемах | Oikos Journal
Отправлено редактором 21 апреля 2015 г. Получите статью!Экологов давно интересовала пространственная структура сообществ, то есть любая неслучайная пространственная организация в распределении сообществ. Если сообщества пространственно структурированы, близлежащие участки более похожи по составу, чем более удаленные. Пространственная структура в экологических сообществах отражает ограничение распространения организмов, смертность, социальную организацию и пространственно структурированную изменчивость окружающей среды (Peres-Neto and Legendre 2010).
В недавней статье «Пространственная структура в экологических сообществах — количественный анализ», опубликованной в Early View, я представил результаты большого метаобзора о степени пространственной структуры в экологических сообществах (n = 322 набора данных). Эта статья принадлежит к серии исследовательских работ о пространственных закономерностях в экологических сообществах, таких как взаимосвязь видов и территорий (Drakare et al., 2006), уменьшение сходства сообществ на расстоянии (Soininen et al. 2007a), бета-разнообразие (Soininen et al.2007b) и сортировка видов по окружающей среде (Soininen 2014). Эти недавние метаобзоры существенно расширили наши знания о макроэкологических закономерностях в составе сообществ или биоразнообразии, но большинство из них не смогли должным образом учесть различия в неоднородности окружающей среды между исследованиями. Здесь я спросил, варьируется ли пространственная структура в биотических сообществах предсказуемо в зависимости от характеристик организма, таких как размер тела, трофическое положение или способ распространения, а также в разных типах экосистем (пресноводных, морских и наземных экосистемах).Я количественно оценил пространственную изменчивость как долю вариации сообщества, которая может быть объяснена пространственными переменными после учета вариации факторов окружающей среды.
Я показал, что в среднем 11,0% вариации в составе сообщества объяснялись пространственными переменными после того, как было учтено влияние переменных окружающей среды. Для всех таксонов пространственный компонент увеличивался с увеличением размера тела, что позволяет предположить, что более крупные таксоны могут иметь более ограниченное распространение по участкам, чем более мелкие.Пространственный компонент также значительно увеличился со степенью исследования, указывающей на то, что ограничение расселения усиливается с увеличением степени. Пространственный компонент был самым высоким среди наземных таксонов и выше у эктотермов, чем у гомойотерм. Пространственная структура также была выше у всеядных, чем у автотрофов.
Эти результаты предполагают, что степень пространственной структуры определяется как внешними факторами, такими как масштабы исследования и тип экосистемы, так и внутренними факторами, такими как размер тела, терморегуляция и взаимодействие между размером тела и режимом расселения.Количественная оценка основных закономерностей пространственной структуры экологических сообществ является не только важной задачей для фундаментальных исследований, но также имеет важное прикладное значение, поскольку кампании по отбору проб для программ биомониторинга или сохранения основываются на знаниях о степени естественных вариаций в природе. сообщества в космосе.
Ссылка, процитированная выше
Drakare, S. et al. 2006. Влияние географического, эволюционного и экологического контекста на взаимоотношения видов и ареалов.- Экол. Lett. 9: 215-227.
Перес-Нето П. Р. и Лежандр П. 2010. Оценка и контроль пространственной структуры при изучении экологических сообществ. — Global Ecol. Биогеогр. 19: 174-184.
Сойнинен, Дж. 2014. Количественный анализ сортировки видов по организмам и экосистемам. — Экология 95: 3284-3292.
Soininen, J. et al. 2007a. Дистанционный распад сходства в экологических сообществах. — Экография 30: 3-12
Soininen, J. et al. 2007b.Многомерный анализ бета-разнообразия организмов и сред. — Экология 88: 2830-2838.
Категории:
Исследования OikosКомментарии
Пространственная экология и сохранение | Изучайте науку в Scitable
Чейз, Дж. М. К действительно единой теории метасообществ. Функциональная экология 19, 182–186 (2005).
Коллиндж, С. К. Экология фрагментированных ландшафтов. Балтимор, Мэриленд: Джонс Хопкинс Издательство университета, 2009.
Делькур, Х. Р., Делькур, П. А. и др. . Динамическая экология растений: спектр растительных изменений в пространстве и времени. Обзоры четвертичной науки 1, 153–175 (1983).
Форман, Р. Т. Т. и Годрон М. Ландшафтная экология . Нью-Йорк, штат Нью-Йорк: Wiley, 1986.
.Фостер, Д. Р., Флюет М. и др. . Человек или естественное нарушение: ландшафтная динамика тропических лесов Пуэрто-Рико. Экологический Заявки 9 , 555–572 (1999).
Ганский, I. & Gilpin, M. E. eds. Биология метапопуляции . Сан-Диего, Калифорния: Academic Press, 1997.
Holyoak, M., Leibold, M.A. et al. ред. Метасообщества: Пространственные Динамика и экологические сообщества . Чикаго, Иллинойс: University of Chicago Press, 2005.
.Джулс, Э. С. и Шахани, П. А широкий экологический контекст фрагментации среды обитания: почему матричная среда обитания более важнее, чем мы думали. Журнал Наука о растительности 14, 459–464 (2003).
Кремен, К., Уильямс, Н. М. et al. Урожай опыление от местных пчел, подверженных риску интенсификации сельского хозяйства. Труды Национальной Академии наук 99, 16812-16816 (2002).
Leibold, M. A., Holyoak, M. et al. al. Концепция метасообщества: основа для многомасштабного сообщества экология. Письма по экологии 7, 601–613 (2005).
Левин, С. А. и Пейн, Р. Т. Нарушение, образование пятен и структура сообщества. Труды Национального Академия наук 71, 2744–2747 (1974).
Пикетт, С. Т. и Уайт, П. С. Экология естественных нарушений и динамика пятен. Нью-Йорк, Нью-Йорк: Academic Press, 1985.
Тильман, Д. и Кариава П., ред. Пространственная экология: роль пространства в динамике популяций и межвидовых взаимодействиях . Принстон, штат Нью-Джерси: Princeton University Press, 1997.
Тернер, М.Изд. Неоднородность ландшафта и Мудрость . Нью-Йорк, Нью-Йорк: Springer-Verlag, 1987.
Тернер, М.Г. Ландшафтная экология: влияние рисунка на процесс. Ежегодный обзор экологии и систематики 20, 171–197 (1989).
Урбан, Д. Л., О’Нил, Р. В. et al. Ландшафтная экология. BioScience 37, 119–127 (1987).
Вт, А.С. Закономерности и процессы в растительном сообществе. Экологический журнал 35, 1-22 (1947).
Wiens, J. A. Реакция населения на неоднородную среды. Ежегодный обзор экологии и систематика 7, 81–120 (1976).
Wiens, J. A. Возникающая роль неоднородности в биология сохранения. В г. Основа сохранения: неоднородность, экосистемы и биоразнообразие . Пикетт, S. T. A. и Ostfeld, R. S., et al. ред. (Нью-Йорк: Chapman & Hall, 1997): 93–107.
Винс, Дж. А., Мосс, М. R. et al. ред. Фундаментальные бумаги в пейзаже Экология . Новый York, NY: Columbia University Press, 2006.Пространственная структура ведет к экологическому разрушению и потере разнообразия
Proc Biol Sci. 7 июня 2009 г .; 276 (1664): 2065–2070.
Герда Саксер
1 Департамент зоологии, Университет Британской Колумбии, 6270 Университетский бульвар, Ванкувер, Британская Колумбия, Канада V6T 1Z4
2 Департамент биологии и биохимии, Университет Хьюстона, Хьюстон, Техас 77204 США
3 Экология и эволюционная биология, Университет Райса, MS 170, 6100 Мэйн Стрит, Хьюстон, Техас 77005, США
Майкл Добели
1 Кафедра зоологии, Университет Британской Колумбии, 6270 Университетский бульвар, Ванкувер , Британская Колумбия, Канада V6T 1Z4
4 Департамент математики, Университет Британской Колумбии, 1984 Mathematics Road, Ванкувер, Британская Колумбия, Канада V6T 1Z2
Michael Travisano
2 Департамент биологии и биохимии Университета Хьюстон, Хьюстон, Техас 77204, США
5 Департамент экологии, эволюции и поведения, Университет Миннесота a, 100 Ecology, 1987 Upper Buford Circle, St Paul, MN 55108, USA
1 Департамент зоологии, Университет Британской Колумбии, 6270 University Boulevard, Ванкувер, Британская Колумбия, Канада V6T 1Z4
2 Департамент Биология и биохимия, Хьюстонский университет, Хьюстон, Техас 77204, США
3 Экология и эволюционная биология, Университет Райса, MS 170, 6100 Main Street, Хьюстон, Техас 77005, США
4 Департамент математики, Университет Британской Колумбии, 1984 Mathematics Road, Ванкувер, Британская Колумбия, Канада V6T 1Z2
5 Департамент экологии, эволюции и поведения, Университет Миннесоты, 100 Ecology, 1987 Upper Buford Circle, St Paul, MN 55108, USA
* Автор и адрес для переписки: Ecology and Evolutionary Biology MS 170, Rice University, 6100 Main Street, Houston, TX 77005, USA (ude.ecir @ rexasg)Поступила в редакцию 9 декабря 2008 г .; Пересмотрено 4 февраля 2009 г .; Принято 5 февраля 2009 г.
Эта статья цитируется в других статьях в PMC.Abstract
Было установлено, что пространственная структура вносит основной вклад в поддержание разнообразия. Здесь мы показываем, что влияние пространственной структуры на разнообразие сильно зависит от экологических механизмов, поддерживающих разнообразие. В хорошо перемешанной, неструктурированной среде популяции микробов могут диверсифицироваться за счет производства метаболитов во время роста, обеспечивая дополнительные ресурсы для новых специалистов.Напротив, пространственно структурированная среда потенциально ограничивает такое облегчение из-за уменьшения диффузии метаболитов. Используя воспроизводимые микрокосмы, содержащие бактерию Escherichia coli , мы предсказали потерю разнообразия во время сдвига окружающей среды от пространственно неструктурированной среды к пространственно структурированным условиям. Хотя часто наблюдается, что пространственная структура является основным стимулом разнообразия, наши результаты показывают, что она также может оказывать негативное влияние на разнообразие.
Ключевые слова: эволюция разнообразия, пространственная структура, потеря разнообразия, содействие, экологические взаимодействия
1. Введение
Теория ниши утверждает, что видовое разнообразие развивается и поддерживается посредством компромиссов (Whittaker 1965), преимущественно опосредованным экологические взаимодействия, такие как конкуренция, хищничество или мутуализм (Doebeli & Dieckmann 2000; Schluter 2000). Пространственная структура может способствовать разнообразию за счет локализации воздействия организмов на окружающую их среду (Amarasekare 2003).Локально истощенные ресурсы и ограниченная диффузия ингибиторов создают неоднородную среду, в результате чего многочисленные ниши предоставляют экологические возможности для диверсификации (Chao & Levin 1981; Durrett & Levin 1994; Rainey & Travisano 1998; Czaran et al .2002; Greig & Travisano) 2004; Habets et al .2006). Однако пространственная структура может также потенциально ограничить создание ниши за счет снижения доступности ресурсов; в частности, ресурсы, предоставляемые посредством содействия (например, в качестве побочных продуктов потребления).В таком случае первичные потребители производят отходы, дающие новые ресурсы. В хорошо перемешанной среде такие ресурсы легко доступны для мусорщиков. Напротив, пространственная структура локализует ресурсы побочных продуктов в непосредственной среде основных потребителей, потенциально ограничивая доступность ресурсов для мусорщиков и тем самым ограничивая экологические возможности.
Лабораторные популяции Escherichia coli и других бактерий быстро адаптируются и диверсифицируются в пространственно неструктурированных микрокосмах (Friesen et al .2004), как предсказывает теория ниши, даже в средах с одним питательным веществом (Helling et al . 1987; Rozen & Lenski 2000). Во время роста на избытке глюкозы или других углеводов микробы выделяют в среду метаболиты, которые служат дополнительными субстратами для роста и, следовательно, могут вызвать эволюцию новых и сосуществующих типов, специализирующихся на этих метаболитах, как неоднократно демонстрировалось (Rosenzweig et al. . 1994; Тернер и др. . 1996; Дебели 2002; Пфайффер и Бонхёффер 2004; Вольф 2005).Поэтому мы предположили, что разнообразие, поддерживаемое таким посредничеством метаболитов, будет снижаться после перехода к пространственно структурированным условиям, которые ограничивают диффузию и, следовательно, доступность метаболитов.
2. Материал и методы
(a) Эволюционный эксперимент
Мы развили 12 популяций E. coli B в минимальной жидкой среде Дэвиса с добавлением 410 мкг / мл -1 глюкозы (DM + Glu) для 1000 поколений. путем ежедневного 100-кратного разбавления популяций стационарной фазы свежей средой.Популяции были основаны на двух изогенных наследственных генотипах, которые различались только селективно нейтральным маркером (ara — и ara + ; Levin et al . 1977; Lenski & Travisano 1994; Travisano & Lenski 1996), которые легко могут быть быть различимым на индикаторных пластинах тетразолий-арабинозы (ТА). Мы оценили размер колонии, высевая образец каждой популяции с низкой плотностью на чашки с агаром TA и инкубируя их в течение 48 часов перед измерением размера отдельной колонии (в пикселях) с помощью ImageJ (v.1,31 В, NIH). Разнообразие в популяции рассчитывалось как скорректированный коэффициент вариации (CV *) размера колонии как CV * = (1 + 1/4 n ) × CV, где n — размер выборки. Коэффициент вариации CV рассчитывается как CV = s × 100 / Y¯ с выборочной дисперсией s и выборочным средним Y¯ (Sokal & Rohlf 1995). Скорректированный коэффициент вариации корректирует небольшие размеры выборки по сравнению с коэффициентом вариации (Sokal & Rohlf 1995). Мы использовали CV * вместо типичных «биннинговых» мер разнообразия (например,грамм. Шеннон-Уивер или Симпсон), поскольку он предоставил беспристрастную альтернативу трудности выбора подходящего размера бункера. Мы проверили генетическую основу размера колонии, высеивая отдельные колонии, измеряя их размер на чашках до и после 1 дня роста в жидкой среде, а затем вычисляя коэффициент корреляции среднего размера колонии между двумя наборами чашек.
(b) Размножение в пространственно структурированной среде
Микрокосмы выращивали в виде газонов на 10 мл DM с добавлением 410 мкг мл -1 глюкозы и 14 мг мл -1 агара (DM + Glu + Agar) в маленькие чашки Петри (диаметр 62 мм).Каждый день микрокосмы размножались 100-кратным разбавлением микрокосмов стационарной фазы, выращенных на лужайке. Мы извлекли пробку диаметром 13 мм из чашки, диспергировали бактерии в 500 мл 0,85-процентного физиологического раствора и перенесли 116 мкл на свежую среду. Пробка занимала примерно одну двадцатую часть площади. Учитывая большой размер перенесенной популяции, эта процедура выборки сама по себе не привела к потере редких типов и, следовательно, к снижению разнообразия. На пространственно структурированной селективной среде (DM + Glu + Agar) разные специалисты были неотличимы при посеве с низкой плотностью, поскольку размер колоний не менялся в любое время (через 24 или 48 часов после посева).На чашках ТА размер колоний действительно различался. Чтобы оценить разнообразие, мы поместили образец каждого микрокосма с низкой плотностью на чашки ТА, измерили размер колонии после 48 часов роста на чашке и рассчитали скорректированный коэффициент вариации (CV *) как меру разнообразия размеров колоний.
(c) Частотный анализ
Два морфотипа колонии были выделены из каждого микрокосма и конкурировали посредством ежедневного переноса в свежую среду в течение 5 дней в жидкости и в течение 3 дней в твердой среде с разными начальными частотами (10:90 и 90:10). .Метод распространения в пространственно структурированной среде использовался, как описано выше. В пространственно структурированной среде анализы были остановлены до того, как один из морфотипов вымер. В начале и в конце соревнований частота двух спортсменов оценивалась и использовалась для расчета относительной физической формы за день (Ленски и др. . 1991).
Для изучения экологических взаимоотношений внутри культур мы выбрали наиболее расходящиеся морфы колоний из каждой популяции, чтобы эти морфы можно было надежно различить в последующих экспериментах.Впоследствии мы сгруппировали морфотипы как экотипы в зависимости от того, сохранился ли вариант колонии или исчез в пространственно структурированной среде.
(d) Анализ перекрестного кормления
Чтобы проверить, является ли перекрестное кормление возможным механизмом поддержания разнообразия, мы оценили рост различных морф колоний в селективных и обедненных средах. Мы создали обедненную среду, засевая 10 мл селективной среды DM + Glu 100 мкл стационарных кондиционированных культур каждой морфы колонии, выращенных индивидуально.После 7 и 24 часов роста мы собрали популяции и отделили бактерии от среды стерилизацией фильтрованием с использованием шприцевого фильтра PSE 0,22 мкм. Для анализа кривой роста мы инокулировали 200 мкл среды 2 мкл кондиционированной культуры в стационарной фазе и выращивали культуры в 96-луночном планшете в течение 24 часов на считывающем устройстве для планшетов. Каждый тип колонии выращивали трижды на трех разных средах: на селективной среде DM + Glu; среда истощена самостоятельно; и средние, обедненные сородичами.
Чтобы оценить максимальную скорость роста, мы преобразовали данные оптической плотности в журнал и определили максимальную скорость роста, установив линейный наклон в скользящем окне и определив максимальный наклон. Перед проверкой того, росли ли экотипы по-разному в обедненных средах, мы выполнили дисперсионный анализ (ANOVA) максимальной скорости роста в предковой среде, DM + Glu, с блоком и экотипом в качестве факторов. Экотип определялся в соответствии с характеристиками морфа колонии во время эксперимента на пространственно структурированной среде (сохраняется или исчезает).Мы не наблюдали значительного эффекта для блока и впоследствии выполнили полный факторный ANOVA с окружающей средой и экотипом в качестве фиксированных факторов. Чтобы проверить компромисс в быстром росте со специализацией на глюкозу и метаболиты, мы выполнили полный факторный дисперсионный анализ времени максимальной скорости роста с экотипом и блоком в качестве факторов.
(e) Индивидуальные анализы роста на пространственно структурированной среде
Чтобы проверить эксплуатационные различия в пространственно структурированной среде между различными морфами колоний, разные морфы выращивали индивидуально до стационарной фазы в жидкой среде, прежде чем они были перенесены в пространственно структурированную среду и выращивали в течение 24 часов в экспериментальных условиях.До и после роста на пространственно структурированной среде размеры популяции измеряли для обоих типов морфа колоний и сравнивали с использованием t -теста с равной дисперсией и восемью степенями свободы.
(f) Статистический анализ
Чтобы определить влияние пространственной структуры на разнообразие, мы выполнили REML ANCOVA (анализ ковариации с ограниченным максимальным правдоподобием) с использованием JMP (SAS Institute Inc., Кэри, Северная Каролина, США) по различиям между CV * неструктурированный (контроль) и CV * структурированный (лечение) в течение 7 дней с фиксированной точкой пересечения на нуле.Был проведен полный факторный анализ, в котором в качестве факторов использовались день (фиксированный), микрокосм (случайный) и блок (случайный). Чтобы проверить влияние начальной плотности, мы выполнили ANCOVA для разницы между CV * нормальной плотностью переноса и CV * 10-кратной более низкой плотности переноса с дневным (фиксированным) и микрокосмом (случайным) в качестве факторов. Как и раньше, мы устанавливаем точку пересечения на ноль. Поскольку мы проверили гипотезу направленности, мы преобразовали дневное значение F в одностороннее значение t .
3.Результаты
Во-первых, мы добились разнообразия в хорошо смешанных популяциях и подтвердили, что разнообразие поддерживается посредством фасилитации. Из одного клона E. coli мы инициировали 12 копий микрокосмов и размножили их во встряхиваемой среде с ограничением глюкозы в течение 1000 поколений. В ходе отбора популяция увеличилась в приспособленности на 23% по сравнению с их общим предком (средняя приспособленность = 1,23, 95% ДИ = 0,029, n = 12), что свидетельствует об адаптации к селективной среде.Прирост приспособленности был больше в течение первых 500 поколений, чем у вторых 500 поколений ( t 11 = 3,56, p <0,01), закономерность, часто наблюдаемая в экспериментах по селекции (Lenski & Travisano 1994). Точно так же фенотипическое разнообразие развивалось в течение первых 500 поколений и оставалось стабильным до конца эксперимента ( a ). После 350 поколений мы наблюдали множественные морфотипы колоний во всех популяциях, когда мы отбирали образцы микрокосмов на твердую, богатую питательными веществами индикаторную среду (Rosenzweig et al .1994; Тернер и др. . 1996; Рейни и Трэвисано 1998; Friesen и др. . 2004 г.). Поскольку все микрокосмы были засеяны от одного предкового клона, любое разнообразие внутри микрокосмов возникло в результате появления de novo мутаций в ходе селекционного эксперимента. Мы проверили генетическую основу этого фенотипического признака и наблюдали генетическую основу морфологии колонии, поскольку размеры различных вариантов колонии, выделенных после 1000 поколений, сильно коррелировали до и после 24 часов роста в жидкой среде ( r = 0.92, n = 27, p <0,0001).
Разнообразие эволюционировало и сохранилось в жидкой среде массового действия. ( a ) Вариация размера колонии в пределах микрокосма эволюционировала в ходе эксперимента по отбору, о чем свидетельствует увеличение среднего коэффициента вариации размера колонии (CV *) в микрокосмах (среднее значение с 95% доверительным интервалом). Фенотипическое разнообразие увеличивалось в течение первых 500 поколений ( F 1,29 = 38,36, p <0,0001) без значительных изменений между поколениями 500 и 1000 ( F 1,59 = 0.0013, р = 0,971). ( b ) Стабилизирующая частотная зависимость между сохраняющимся и исчезающим экотипом имела место в хорошо перемешанной среде. Относительная приспособленность исчезающего экотипа была высокой при низкой начальной частоте и низкой при высокой начальной частоте ( p <0,0001, парный t -тест). Линии, соединяющие значения пригодности на низких и высоких частотах, пересекают пунктирную линию, где оба спортсмена имеют одинаковую физическую форму. ( c ) Скорость роста двух экотипов в DM + Glu и в среде, истощенной в течение 7 или 24 часов самим собой или конспецифом (в среднем с 95% доверительным интервалом).Максимальная скорость роста была измерена в течение 24-часового цикла роста как для сохраняющегося (черные столбцы), так и для исчезающего (серые столбцы) экотипа. В среде, истощенной в течение 24 часов, экотипы не различались по скорости роста. В среде, истощенной в течение 7 часов, мы наблюдали значительный эффект окружающей среды (среда, истощенная самим собой или сородичами: F 1,104 = 29,84, p <0,0001) со значительным сроком взаимодействия между средой и экотипом ( F 1,104 = 6.96, p = 0,0096).
Частотно-зависимый отбор необходим для поддержания разнообразия, эволюционировавшего в результате экологической специализации (Levene 1953; Rainey & Travisano 1998; Friesen et al , 2004). Чтобы проверить частотно-зависимый отбор и нишевую специализацию среди представителей микрокосмов, мы выделили отдельные пары дивергентных морфотипов колоний из девяти микрокосмов в поколении 1000 и соревновались между ними. Мы исключили из анализа три микрокосма из-за трудности надежной дифференциации вариантов колоний.Эксперименты проводились как взаимное «вторжение из редких» анализов, причем один вариант колонии изначально был редким, а другой — обычным (Rainey & Travisano 1998). Для поддержания разнообразия посредством частотно-зависимого отбора особи данного типа должны иметь более высокую относительную приспособленность, когда они редки. Это избирательное преимущество должно в конечном итоге исчезнуть по мере того, как частота увеличивается. В течение 5 дней последовательного переноса варианты колоний имели высокую приспособленность при низкой начальной частоте и низкую приспособленность при высокой начальной частоте по сравнению с их аналогичными конкурентами (парные t -тест: t 8 = 11.71, p <0,0001; б ). Эта закономерность показывает, что разнообразие поддерживалось за счет стабилизации частотной зависимости в девяти микрокосмах после 1000 поколений отбора.
Разнообразие в средах с одним ресурсом может развиваться посредством разделения и упрощения ресурсов, как было показано в предыдущих экспериментах по селекции с E. coli (Розенцвейг и др. . 1994; Тернер и др. . 1996; Розен и Ленски 2000; Friesen et al .2004; Habets и др. . 2006 г.). Во время роста на глюкозе E. coli выделяет различные метаболиты в окружающую среду. Эти метаболиты поглощаются и метаболизируются, когда рост бактерий исчерпывает доступную в среде глюкозу. Специализация глюкозы приводит к компромиссу между быстрым ростом и использованием метаболитов посредством цикла трикарбоновой кислоты (TCA) (Helling et al .1987; Rosenzweig et al .1994; Turner et al .1996; Rozen & Lenski 2000; Doebeli 2002; Friesen et al .2004; Пфайффер и Бонхёффер 2004; Вулф 2005). Как следствие, метаболиты накапливаются в окружающей среде, обеспечивая ресурсы для эволюции специалистов по метаболитам (Turner et al . 1996). Чтобы проверить такое перекрестное кормление, мы измерили максимальную скорость роста различных вариантов колонии в среде, истощенной самим собой и другой морфой колонии ( c ). Мы собирали среду из двух временных точек: во время позднего экспоненциального роста и во время стационарной фазы, собирая среду после 7 и 24 часов инкубации, соответственно.Сначала мы проверили, различаются ли два экотипа по максимальной скорости роста в DM + Glu, и определили, что два экотипа не различаются по максимальной скорости роста в селективной среде при изолированном выращивании ( F 1,106 = 0,05, p = 0,82). Это указывает на то, что оба типа могли расти на среде с ограничением глюкозы. В соответствии с компромиссом между быстрым ростом глюкозы и специализацией в отношении метаболитов, два экотипа различались по времени максимальной скорости роста, при этом сохраняющиеся экотипы достигли максимального роста до исчезновения экотипов (экотип: F 1 102 = 10.93, p = 0,0013; блок: F 2,102 = 5,4, p = 0,006; экотип × блок: F 2,102 = 0,34, p = 0,714).
Мы также протестировали рост в среде, собранной во время стационарной фазы. Мы ожидали, что экотипы не будут различаться по максимальной скорости роста, так как среда была одинаково истощена. Как и ожидалось, мы не наблюдали значительного влияния окружающей среды или экотипа на максимальную скорость роста (экотип: F 1,104 = 0.0013, p = 0,97; среда: F 1,104 = 2,46, p = 0,119; экотип × среда: F 1,104 = 1,7, p = 0,19). Наконец, мы протестировали рост в среде, собранной во время поздней фазы экспоненциального роста. В этой среде мы ожидали разницы в росте. Мы ожидали, что исчезающий экотип сможет расти в среде, истощенной сохраняющимся экотипом, в то время как сохраняющийся экотип не будет расти в среде, истощенной исчезающим экотипом.Как и ожидалось, мы наблюдали значительный эффект окружающей среды ( F 1,104 = 29,84, p <0,0001) и, что более важно, значимое взаимодействие между окружающей средой и экотипом ( F 1,104 = 6,96, p ). = 0,0096), что указывает на то, что разные экотипы росли по-разному в средах, истощенных самими собой или сородичами. В целом, мы не наблюдали значимой разницы между экотипами ( F 1,104 = 0,144, p = 0.70). Эти результаты предполагают, что перекрестное кормление — весьма вероятный механизм, поддерживающий разнообразие в этих микрокосмах.
Получив разнообразие популяций, мы затем проверили влияние пространственной структуры на существующее разнообразие в пределах микромира. Разнообразие, поддерживаемое посредством фасилитации, должно уменьшаться, если диффузия метаболитов ограничена, поскольку это является основой для фасилитации в этой системе. Мы проверили устойчивость разнообразия во время экологических нарушений, сравнив разнообразие микрокосмов 1000-го поколения, поддерживаемых в хорошо перемешанной и пространственно структурированной среде в течение 7 дней.Каждый из девяти эволюционировавших микрокосмов использовался для инициирования нескольких новых микрокосмов, поддерживаемых по той же временной схеме ежедневного переноса, что и во время первоначального режима отбора 1000 поколений, но различающихся по пространственной структуре. Единственным различием между селективной средой и пространственно структурированной средой было добавление агара, инертного непитательного вещества, обычно используемого для отверждения жидких сред. Первый набор поддерживали в предыдущих хорошо перемешанных условиях встряхивания в жидкой среде, в то время как второй и третий наборы поддерживали на твердом агаре с исходной или в 10 раз меньшей плотностью при переносе, соответственно.Более низкая начальная плотность увеличивает расстояние между колониями и, следовательно, еще больше снижает возможность экологических взаимодействий. Независимо от плотности размножения, плотности на твердой среде были высокими, и бактерии росли как лужайка, а не как отдельные колонии. Наложение пространственной структуры вызвало 50-процентное сокращение разнообразия по сравнению с контролем в жидкой среде (день: F 1,116 = 46,73, p <0,0001; микрокосм: F 8,116 = 11.46, p <0,0001; блок: F 1,116 = 8,52, p = 0,004; a , b ), когда микрокосмы отбирались на твердой среде при низкой плотности. Как и ожидалось, большее сокращение экологического взаимодействия между специалистами из-за увеличения начального расстояния между колониями привело к еще большей потере разнообразия ( т 54 = 1,87, p = 0,039).
Разнообразие уменьшилось в пространственно структурированной среде.( a ) Разнообразие размеров колоний одной популяции (ara- популяция 5), перенесенных (i) в жидкости (DM + Glu) или (ii) в пространственно структурированной среде (DM + Glu + Agar), нанесенной на индикаторные пластины (TA ). К 7 дню небольшие колонии не присутствовали в популяции, перенесенной в пространственно структурированную среду, в то время как небольшие колонии сохранялись на протяжении всего эксперимента в популяциях, перенесенных в жидкую среду. Присутствие небольших колоний в пространственно структурированных популяциях к 3 дню указывает на то, что общее разнообразие было зафиксировано в режиме переноса, использованном для этого эксперимента.(b ) Разнообразие уменьшилось после перехода от хорошо перемешанной к пространственно структурированной среде как при нормальной плотности переноса (треугольники и сплошная линия), так и при 10-кратном снижении плотности переноса (квадраты и пунктирная линия). Изменение разнообразия рассчитывали как разницу в разнообразии в твердой и жидкой средах (точки представляют собой среднее значение двух измерений для девяти популяций при нормальной плотности переноса и среднее значение девяти популяций при плотности переноса в 10 раз ниже). Каждая линия была определена путем регрессии по независимо собранным данным.( c ) Стабилизирующая частотная зависимость между сохраняющимся и исчезающим экотипами не возникала в пространственно структурированной среде. Относительная приспособленность исчезающего экотипа была низкой, независимо от его первоначальной повторяемости. Ни одна из линий, соединяющих фитнес при высокой и низкой начальной частоте, не пересекала линию одинаковой пригодности (пунктирная линия).
Чтобы убедиться, что снижение экологических взаимодействий является причиной потери разнообразия, мы провели два дополнительных эксперимента.Во-первых, мы оценили частотную зависимость в пространственно структурированной среде с использованием тех же пар вариантов колонии, которые использовались в предыдущем частотном анализе в хорошо перемешанной среде. Конкуренция между сородичами в пространственно структурированной среде значительно отличалась от частотно-зависимого отбора, наблюдаемого в жидкой среде (парный t -тест на склонах: t 8 = -10,21, p <0,0001), с небольшим частотная зависимость в пространственно структурированной среде ( c ).Во-вторых, мы оценили способность каждой из различных морфов колоний расти в пространственной среде в отсутствие конкуренции и не обнаружили различий между разными морфами колоний в использовании окружающей среды в отсутствие конкуренции ( t 8 = 1,31, p = 0,23). Эти результаты убедительно подтверждают гипотезу о том, что снижение разнообразия пространственно структурированной среды происходит из-за прерывания экологических взаимодействий.
4.Обсуждение
Пространственная структура глубоко влияет на разнообразие. Пространственная структура может способствовать разнообразию, поддерживаемому различными экологическими взаимодействиями, включая конкуренцию (Czaran et al , 2002), взаимодействия хищник-жертва (Kerr et al , 2002) и даже некоторые формы содействия (Korona et al , 1994; Рейни и Трэвисано 1998). В этих системах наложение пространственной структуры приводит к ограниченному распространению ресурсов, ингибиторов, конкурентов или хищников.Ограниченное распространение способствует увеличению разнообразия из-за создания пространственно распределенных ниш, к которым адаптируются разные специалисты. Хотя часто наблюдается, что пространственная структура способствует диверсификации, она не является ни необходимой, ни полезной ни для эволюции, ни для поддержания разнообразия в нашей модельной системе. Разнообразие в этом исследовании сохраняется из-за сложности ресурсов, как это предусмотрено в нишевой модели Уиттакера (Whittaker, 1965), в которой компромиссы в использовании ниши не позволяют получить единственный наиболее подходящий генотип (Lenski & Hattingh 1986).В этом случае хорошо описанные компромиссы в центральных метаболических путях создают сложную ресурсную среду, на что указывает дифференциальный рост в обедненных средах и различное время до максимальной скорости роста в селективной среде. Наложение пространственной структуры изменяет сложность ресурсов в окружающей среде, перемещая ее в микросреду, в которой доминирует один генотип. Против отбираются мусорщики и специалисты по побочным продуктам.
Мы использовали богатые питательными веществами индикаторные пластины для различения генотипов внутри культур, аналогично подходу, принятому Рейни и Тревизано (1998).Анализ морфологии колоний на поверхности агара, богатого питательными веществами, был важен для нашего исследования, поскольку разные специалисты по питательным веществам формировали колонии, которые нельзя было различить при отборе образцов с низкой плотностью на среду с ограничением питательных веществ, используемую для селекционного эксперимента. Посеяв популяции с низкой плотностью на богатый питательными веществами ТА-агар, мы смогли различить различные морфы колоний и оценить разнообразие без выявления post hoc дискретных различий между колониями. К счастью, нам удалось идентифицировать разные экотипы, различающиеся морфологией колонии.Даже в этом случае наша оценка экотипов, вероятно, будет недооценкой фенотипического и генетического разнообразия, так как мы сосредоточились только на двух дивергентных морфотипах на культуру для проведения экспериментов по конкуренции.
Морфология колонии экотипа действительно варьировалась в зависимости от исходного маркерного генотипа: морфы больших колоний были потеряны в микрокосмах, состоящих из генотипов, способных использовать арабинозу, в то время как морфы малых колоний были потеряны в микрокосмах, состоящих исключительно из генотипов без арабинозы ( a ).Даже в этом случае маркер использования арабинозы влияет на утраченный морфотип, но не на потерю разнообразия. Арабиноза не предоставлялась ни разу во время отбора и присутствовала только в богатой питательными веществами среде, используемой для различения морфотипов. Наблюдаемый маркерный эффект предполагает, что специфическая для ниши адаптация влияет на использование углеводов в целом, даже на углеводы, отсутствующие во время отбора. Это также демонстрирует, что размер колонии в богатой питательными веществами среде, используемой для различения морфов, не был фактором, влияющим на потерю разнообразия во время отбора в пространственно структурированной среде.
Наши эксперименты не затрагивают вопрос о том, благоприятна ли пространственная структура для эволюции разнообразия или для поддержания разнообразия после эволюции в пространственно структурированной среде. Теоретические модели предполагают, что разнообразие, основанное на перекрестном кормлении, может поддерживаться в пространственно структурированной среде при определенных обстоятельствах (Krone & Guan 2006). Habets и др. . (2006) исследовали этот вопрос экспериментально, эволюционируя популяции в различно структурированных и неструктурированных средах.Они заметили, что пространственная структура может способствовать развитию разнообразия, если структура сохраняется, даже при перемещении. Такие условия, безусловно, будут способствовать развитию упрощения формальностей и позволят разнообразию развиваться. В наших экспериментах при переносе популяции смешивались. Предположительно, разнообразие в пространственно структурированной среде могло уменьшиться в результате смешения популяций при перемещении, что нарушило установившееся содействие. Две линии доказательств подтверждают наше утверждение о том, что пространственно структурированная среда больше способствовала потере разнообразия, чем смешение популяций при перемещении.Во-первых, разнообразие поддерживалось в жидкой, постоянно перемешанной среде. Во-вторых, когда мы перенесли наши разнообразные популяции на 250 поколений в пространственно структурированную среду, мы снова наблюдали первоначальное уменьшение разнообразия, за которым последовало увеличение (J. Ho, G. Saxer and M. Travisano 2003, неопубликованные данные), что указывает на то, что разнообразие могли развиваться, несмотря на смешение популяций при переносе. Однако это разнообразие, вероятно, поддерживалось другими механизмами, помимо содействия.
Наши результаты также показывают, что положительные экологические взаимодействия могут играть решающую роль в эволюции и поддержании видового разнообразия. Разнообразие в этой среде с одним ресурсом могло произойти только через некоторую форму разделения ниши (Stewart & Levin 1973). Потенциал для облегчения возникает как эволюционный ответ на конкуренцию за основное питательное вещество и адаптацию к быстрому росту за счет компромиссов глюкозы с ростом продуктов метаболизма гликолиза. Общность конкуренции за ресурсы и компромиссов в использовании ресурсов предполагает, что содействие, вероятно, будет частым результатом разделения ниши и, следовательно, общим механизмом возникновения и поддержания разнообразия (Dieckmann & Doebeli 1999; Doebeli 2002).
Благодарности
Мы признательны Р. Азеведо, К. Берчу, Б. Коулу, О. Гилберту, Э. Островски, М. Куансу, Р. Куансу, К. Раутеру, Дж. Раджерсу, Дж. Штрассманну, Д. • Вернашу, коллегам из лаборатории и многочисленным анонимным рецензентам за полезные комментарии. Дж. Хо и С. Дуррани помогали во время экспериментов и анализа изображений. Т. Купер любезно предоставил оборудование и советы для эксперимента по перекрестному вскармливанию. Доктор медицины выражает признательность за поддержку Швейцарскому национальному научному фонду Джеймса С.Фонд Макдоннелла (США) и NSERC (Канада). M.T. был поддержан Национальным научным фондом США.
Список литературы
- Амарасекаре П. Конкурентное сосуществование в пространственно структурированной среде: синтез. Ecol. Lett. 2003. 6: 1109–1122. DOI: 10.1046 / j.1461-0248.2003.00530.x [Google Scholar]
- Чао Л., Левин Б.Р. Структурированные среды обитания и эволюция антиконкурентных токсинов у бактерий. Proc. Natl Acad. Sci. США. 1981; 78: 6324–6328. DOI: 10.1073 / pnas.78.10.6324 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Czaran T.L., Hoekstra R.F., Pagie L. Химическая война между микробами способствует сохранению биоразнообразия. Proc. Natl Acad. Sci. США. 2002; 99: 786–790. doi: 10.1073 / pnas.012399899 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Дикманн У., Добели М. О происхождении видов по симпатрическому видообразованию. Природа. 1999; 400: 354–357. doi: 10.1038 / 22521 [PubMed] [Google Scholar]
- Doebeli M. Модель эволюционной динамики полиморфизмов перекрестного питания у микроорганизмов.Popul. Ecol. 2002; 44: 59–70. doi: 10.1007 / s101440200008 [Google Scholar]
- Doebeli M., Dieckmann U. Эволюционное ветвление и симпатрическое видообразование, вызванное различными типами экологических взаимодействий. Являюсь. Nat. 2000; 156: S77 – S101. doi: 10.1086 / 303417 [PubMed] [Google Scholar]
- Дарретт Р., Левин С. Важность дискретности (и пространственности) Теор. Popul. Биол. 1994; 46: 363–394. doi: 10.1006 / tpbi.1994.1032 [Google Scholar]
- Friesen M.L., Saxer G., Travisano M., Doebeli M.Экспериментальные доказательства симпатрической экологической диверсификации из-за частотно-зависимой конкуренции у Escherichia coli . Эволюция. 2004. 58: 245–260. doi: 10.1111 / j.0014-3820.2004.tb01642.x [PubMed] [Google Scholar]
- Грейг Д., Трэвисано М. Дилемма заключенного и полиморфизм генов дрожжей SUC . Proc. R. Soc. Лондон. Б. 2004; 271: S25 – S26. doi: 10.1098 / rsbl.2003.0083 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Habets M.G.J.L., Rozen D.E., Hoekstra R.Ф., де Виссер J.A.G.M. Влияние популяционной структуры на адаптивную радиацию микробных популяций, развивающихся в пространственно-структурированной среде. Ecol. Lett. 2006; 9: 1041–1048. doi: 10.1111 / j.1461-0248.2006.00955.x [PubMed] [Google Scholar]
- Хеллинг Р. Б., Варгас С. Н., Адамс Дж. Эволюция Escherichia coli во время роста в постоянной среде. Генетика. 1987. 116: 349–358. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Керр Б., Райли М.А., Фельдман М.В., Боханнан Б.Дж.М. Местное рассредоточение способствует сохранению биоразнообразия в реальной игре «камень-ножницы-бумага». Природа. 2002; 418: 171–174. DOI: 10.1038 / nature00823 [PubMed] [Google Scholar]
- Корона Р., Накацу К.Х., Форни Л.Дж., Ленски Р.Э. Доказательства множественных адаптивных пиков популяций бактерий, развивающихся в структурированной среде обитания. Proc. Natl Acad. Sci. США. 1994; 91: 9037–9041. doi: 10.1073 / pnas.91.19.9037 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Крон С. М., Гуань Ю. Пространственная самоорганизация в циклической модели ресурсов и видов.J. Theor. Биол. 2006; 241: 14–25. doi: 10.1016 / j.jtbi.2005.11.005 [PubMed] [Google Scholar]
- Ленски Р.Э., Хаттинг С.Е. Сосуществование двух конкурентов на одном ресурсе и одном ингибиторе: модель хемостата на основе бактерий и антибиотиков. J. Theor. Биол. 1986; 122: 83–93. doi: 10.1016 / S0022-5193 (86) 80226-0 [PubMed] [Google Scholar]
- Ленски Р.Е., Трависано М. Динамика адаптации и диверсификации: эксперимент с популяциями бактерий для 10 000 поколений. Proc. Natl Acad. Sci.США. 1994; 91: 6808–6814. doi: 10.1073 / pnas.91.15.6808 [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ленски Р.Э., Роуз М.Р., Симпсон С.С., Тадлер С.С. Долгосрочная экспериментальная эволюция Escherichia coli. I: адаптация и дивергенция в течение 2000 поколений. Являюсь. Nat. 1991; 138: 1315–1341. doi: 10.1086 / 285289 [Google Scholar]
- Левен Х. Генетическое равновесие, когда доступно более одной экологической ниши. Являюсь. Nat. 1953; 87: 331–333. DOI: 10.1086 / 281792 [Google Scholar]
- Левин Б.Р., Стюарт Ф.М., Чао Л. Рост, конкуренция и хищничество с ограниченными ресурсами: модель и экспериментальные исследования с бактериями и бактериофагами. Являюсь. Nat. 1977; 111: 3–24. doi: 10.1086 / 283134 [Google Scholar]
- Пфайфер Т., Бонхёффер С. Эволюция перекрестного кормления в микробных популяциях. Являюсь. Nat. 2004; 163: E126 – E135. doi: 10.1086 / 383593 [PubMed] [Google Scholar]
- Рейни П. Б., Тревизано М. Адаптивное излучение в гетерогенной среде. Природа. 1998. 394: 69–72. doi: 10.1038 / 27900 [PubMed] [Google Scholar]
- Розенцвейг Р.Ф., Шарп Р.Р., Тревес Д.С., Адамс Дж. Микробная эволюция в простой неструктурированной среде: генетическая дифференциация в Escherichia coli . Генетика. 1994; 137: 903–917. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Розен Д.Э., Ленски Р.Э. Долгосрочная экспериментальная эволюция Escherichia coli . VIII: динамика сбалансированного полиморфизма. Являюсь. Nat. 2000; 155: 24–35. DOI: 10.1086 / 303299 [PubMed] [Google Scholar]
- Schluter D. Oxford University Press; Оксфорд, Великобритания: 2000.Экология адаптивного излучения. [Google Scholar]
- Sokal R.R., Rohlf F.J. W. H. Freeman and Co; Нью-Йорк, Нью-Йорк: 1995. Биометрия. [Google Scholar]
- Стюарт Ф.М., Левин Б.Р. Разделение ресурсов и исход межвидовой конкуренции: модель и некоторые общие соображения. Являюсь. Nat. 1973; 107: 171–198. DOI: 10.1086 / 282825 [Google Scholar]
- Travisano M., Lenski R.E. Долгосрочная экспериментальная эволюция Escherichia coli IV: мишени отбора и специфичность адаптации.Генетика. 1996. 143: 15–26. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Turner P.E., Souza V., Lenski R.E. Тестирование экологических механизмов, способствующих стабильному сосуществованию двух генотипов бактерий. Экология. 1996; 77: 2119–2129. doi: 10.2307 / 2265706 [Google Scholar]
- Уиттакер Р. Доминирование и разнообразие сообществ наземных растений. Наука. 1965; 147: 250–260. [PubMed] [Google Scholar]
- Вулф А.Дж. Ацетатный переключатель. Microbiol. Мол. Биол. Ред. 2005; 69: 12–50. DOI: 10.1128 / MMBR.69.1.12-50.2005 [Бесплатная статья PMC] [PubMed] [Google Scholar]
Разделение пространственных, экологических и общественных факторов функционирования экосистемы
Ахти Т., Хэмет-Ахти Л., Джалас Дж. (1968) Зоны растительности и их секции в северо-западной Европе. Энн Бот Фенн 5 (3): 169–211
Google Scholar
Ashton KG, Tracy MC, de Queiroz A (2000) Верно ли правило Бергмана для млекопитающих? Am Nat 156 (4): 390–415
PubMed Статья Google Scholar
Benfield EF (1996) Распад листьев в экосистемах ручьев.В: Хауэр Ф. Р., Ламберти Г. А. (ред.) Методы экологии ручья. Academic Press, San Diego, pp 579–589
Google Scholar
Blanchet FG, Legendre P, Borcard D (2008) Прямой выбор независимых переменных. Экология 89 (9): 2623–2632
PubMed Статья Google Scholar
Borcard D, Legendre P (2002) Полномасштабный пространственный анализ экологических данных с помощью главных координат соседних матриц.Ecol Model 153 (1): 51–68
Артикул Google Scholar
Boyero L, Graca MAS, Tonin AM et al (2017) Качество подстилки прибрежных растений увеличивается с увеличением широты. Научный доклад 7 (1): 10562
PubMed PubMed Central Статья CAS Google Scholar
Brose U, Hillebrand H (2016) Биоразнообразие и функционирование экосистем в динамичных ландшафтах. Филос Транс Р Соц Б.https://doi.org/10.1098/rstb.2015.0267
Артикул Google Scholar
Браун Дж. Х., Гиллули Дж. Ф., Аллен А. П., Сэвидж В. М., Вест ГБ (2004) К метаболической теории экологии. Экология 85 (7): 1771–1789
Статья Google Scholar
Burdon FJ, Harding JS (2008) Связь между прибрежными хищниками и водными насекомыми по всему спектру ресурсов водотока.Freshw Biol 53 (2): 330–346
Google Scholar
Carlson PE, McKie BG, Sandin L, Johnson RK (2016) Сильное влияние землепользования на характер расселения взрослых речных насекомых: последствия для передачи субсидий на водные ресурсы наземным потребителям. Freshw Biol 61 (6): 848–861
Статья Google Scholar
Cornacchia L, van de Koppel J, van der Wal D, Wharton G, Puijalon S, Bouma T.J. (2018) Пейзажи облегчения: как самоорганизованная мозаичность водных макрофитов способствует разнообразию водотоков.Экология 99 (4): 832–847
PubMed Статья Google Scholar
Cottenie K (2005) Интеграция экологических и пространственных процессов в динамику экологического сообщества. Ecol Lett 8 (11): 1175–1182
PubMed Статья Google Scholar
Dangles O, Gessner MO, Guerold F, Chauvet E (2004) Влияние подкисления водотока на разрушение подстилки: последствия для оценки функционирования экосистемы.J Appl Ecol 41: 365–378
CAS Статья Google Scholar
Declerck SAJ, Coronel JS, Legendre P, Brendonck L (2011) Масштабная зависимость процессов, структурирующих метасообщества кладоцер во временных бассейнах водно-болотных угодий Высоких Анд. Экография 34 (2): 296–305
Статья Google Scholar
Des Roches S, Post DM, Turley NE et al (2018) Экологическое значение внутривидовой изменчивости.Nat Ecol Evol 2 (1): 57–64
PubMed Статья Google Scholar
Дитрих А.Л., Нильссон С., Янссон Р. (2013) Фитометры недостаточно используются для оценки экологического восстановления. Basic Appl Ecol 14 (5): 369–377
Статья Google Scholar
Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJ, Collen B (2014) Defaunation in the Anthropocene. Наука 345 (6195): 401–406
CAS PubMed Статья Google Scholar
Донг X, Фишер С.Г. (2019) Пространственная самоорганизация экосистемы: свободный порядок ни в чем? Ecol Complex 38: 24–30
Статья Google Scholar
Dray S, Dufour AB (2007) Пакет ade4: реализация диаграммы двойственности для экологов.J Stat Softw 22 (4): 1–20
Статья Google Scholar
Dray S, Legendre P, Blanchet FG (2013) packfor: forward Selection с перестановкой (Canoco, стр.46). Версия пакета R 0.0-8 / r109
Dray S, Legendre P, Peres-Neto PR (2006) Пространственное моделирование: комплексная основа для анализа основных координат соседних матриц (PCNM). Ecol Model 196 (3–4): 483–493
Артикул Google Scholar
Энквист Б.Дж., Норберг Дж., Бонсер С.П. и др. (2015) Масштабирование от черт к экосистемам: разработка общей теории драйверов черт путем интеграции теорий на основе черт и теории метаболического масштабирования.В: Pawar S, Woodward G, Dell AI (eds) Экология на основе черт — от структуры к функции. Успехи в экологических исследованиях. Academic Press, London, pp. 249–318
Chapter Google Scholar
Frainer A, McKie BG (2015) Изменения в разнообразии и составе потребительских качеств ограничивают влияние землепользования на функционирование экосистемы ручья. В: Pawar S, Woodward G, Dell AI (eds) Экология на основе черт — от структуры к функции.Успехи в экологических исследованиях. Academic Press, San Diego, pp. 169–200
Chapter Google Scholar
Frainer A, McKie BG, Malmqvist B, Woodward G (2014) Когда имеет значение разнообразие? Функциональное разнообразие видов и функционирование экосистемы в разных средах обитания и сезонах в полевом эксперименте. J Anim Ecol 83 (2): 460–469
PubMed Статья Google Scholar
Frainer A, Polvi LE, Jansson R, McKie BG (2017) Улучшение функционирования экосистемы после восстановления водотока: роль неоднородности среды обитания и особенности видов беспозвоночных.J Appl Ecol 55: 377–385
Статья Google Scholar
Гагич В., Бартомеус И., Йонссон Т. и др. (2015) Функциональная идентичность и разнообразие животных предсказывают функционирование экосистемы лучше, чем индексы, основанные на видах. Proc Biol Sci 282 (1801): 20142620
PubMed PubMed Central Статья Google Scholar
Gessner MO (2005) Эргостерин как мера биомассы грибов.В: Graça MAS, Bärlocher F, Gessner MO (eds) Методы изучения разложения подстилки: практическое руководство. Springer, Dordrecht, pp. 189–195
Глава Google Scholar
Gessner MO, Chauvet E (2002) Пример использования разбивки подстилки для оценки функциональной целостности потока. Ecol Appl 12 (2): 498–510
Статья Google Scholar
Gessner MO, Swan CM, Dang CK et al (2010) Разнообразие встречается с разложением.Trends Ecol Evol 25 (6): 372–380
PubMed Статья Google Scholar
Гёте Э., Ангелер Д. Г., Готшалк С., Лёфгрен С., Сандин Л. (2013) Влияние экологических, биотических и пространственных факторов на метасообщество диатомовых водорослей: структура в шведских верховьях. PLoS ONE 8 (8): e72237
PubMed PubMed Central Статья CAS Google Scholar
Grime JP (1998) Преимущества разнообразия растений для экосистем: немедленные, фильтрующие и основательные эффекты.J Ecol 86 (6): 902-910
Артикул Google Scholar
Грёнроос М., Хейно Дж., Сикейра Т., Ландейро В.Л., Котанен Дж., Бини Л.М. (2013) Структурирование метасообщества в потоковых сетях: роли режима рассредоточения, типа расстояния и регионального экологического контекста. Ecol Evol 3 (13): 4473–4487
PubMed PubMed Central Статья Google Scholar
Хеффернан Дж. Б., Соранно П. А., Ангиллетта М. Дж. И др. (2014) Экология макросистем: понимание экологических закономерностей и процессов в континентальном масштабе.Front Ecol Environ 12 (1): 5–14
Артикул Google Scholar
Хейно Дж., Мело А.С., Бини Л.М. и др. (2015) Сравнительный анализ показывает слабую взаимосвязь между экологическими факторами и бета-разнообразием метасообществ ручьевых насекомых на двух пространственных уровнях. Ecol Evol 5 (6): 1235–1248
PubMed PubMed Central Статья Google Scholar
Hildrew AG, Raffaelli DG, Edmonds-Brown R (2007) Размер тела: структура и функция водных экосистем.Издательство Кембриджского университета, Кембридж
Книга Google Scholar
Hillebrand H (2004) Об общности градиента широтного разнообразия. Am Nat 163 (2): 192–211
PubMed Статья Google Scholar
Hooper DU, Solan M, Symstad AJ et al (2002) Видовое разнообразие, функциональное разнообразие и функционирование экосистемы. В: Al Loreau M (ed) Биоразнообразие и функционирование экосистем, синтез и перспективы.Oxford University Press, Нью-Йорк, стр. 195–281
Google Scholar
Jeppesen E, Sondergaard M, Sondergaard M, Christoffersen K (1998) Структурирующая роль погруженных макрофитов в озерах. Спрингер, Нью-Йорк
Бронировать Google Scholar
Джонсон Р.К., Гёдкооп В., Сандин Л. (2004) Пространственный масштаб и экологические отношения между сообществами макробеспозвоночных в каменистых местообитаниях ручьев и озер.Freshw Biol 49 (9): 1179–1194
Статья Google Scholar
Джонсон Р.К., Геринг Д. (2010) Пространственная конгруэнтность сообществ донных диатомей, беспозвоночных, макрофитов и рыб в европейских водотоках. Ecol Appl 20 (4): 978–992
PubMed Статья Google Scholar
Jonsson M (2006) Влияние разнообразия видов на функционирование экосистемы увеличивается со временем в системе эфемерных ресурсов.Acta Oecol 29 (1): 72–77
Статья Google Scholar
Kéfi S, Berlow EL, Wieters EA et al (2012) Больше, чем еда… интеграция взаимодействий, не связанных с кормлением, в пищевые сети. Ecol Lett 15 (4): 291–300
PubMed Статья Google Scholar
Ламберти Г.А., Реш В.Х. (1983) Перифитон ручья и насекомые-травоядные: экспериментальное исследование выпаса популяции ручейников.Экология 64 (5): 1124–1135
Статья Google Scholar
Лаворель С., Гарнье Э. (2002) Прогнозирование изменений в составе сообщества и функционировании экосистемы на основе характеристик растений: возвращение к Святому Граалю. Funct Ecol 16: 545–556
Статья Google Scholar
Лаворел С., Григулис К., Макинтайр С. и др. (2008) Оценка функционального разнообразия в полевых условиях — методология имеет значение! Funct Ecol 22 (1): 134–147
Google Scholar
Layer K, Hildrew AG, Woodward G (2013) Выпас и детритворы в 20 цепочках питания ручья в широком градиенте pH.Oecologia 171 (2): 459–471
PubMed Статья Google Scholar
Lecerf A, Chauvet E (2008) Внутривидовая изменчивость признаков листьев сильно влияет на разложение листьев ольхи в ручье. Basic Appl Ecol 9 (5): 598–605
Статья Google Scholar
Legendre P, Gallagher ED (2001) Экологически значимые преобразования для упорядочивания данных по видам.Oecologia 129 (2): 271–280
PubMed Статья Google Scholar
Лежандр П., Лежандр Л. (2012) Многомасштабный анализ: пространственные собственные функции. В: Legendre P, Legendre L (eds) Развитие моделирования окружающей среды. Elsevier, Oxford, pp. 859–906
Google Scholar
Leibold MA, Holyoak M, Mouquet N et al (2004) Концепция метасообщества: основа для многомасштабной экологии сообщества.Ecol Lett 7 (7): 601–613
Статья Google Scholar
Logue JB, Mouquet N, Peter H, Hillebrand H (2011) Эмпирические подходы к метасообществам: обзор и сравнение с теорией. Trends Ecol Evol 26 (9): 482–491
PubMed Статья Google Scholar
Лоро М., Муке Н., Холт Р.Д. (2005) От метасообществ к метаэкосистемам. В: Дрисколл Д.А. (ред.) Метасообщества: пространственная динамика и экологические сообщества.Издательство Чикагского университета, Чикаго, стр. 418–438
Google Scholar
Макки Б.Г., Крэнстон П.С. (2005) Размер имеет значение: систематические и экологические последствия аллометрии в ответах морфологических соотношений хирономидных мошек на экспериментальные температурные манипуляции. Can J Zool 83 (4): 553–568
Статья Google Scholar
Макки Б.Г., Мальмквист Б. (2009) Оценка функционирования экосистемы в реках, затронутых лесопользованием: усиленное разложение листьев происходит без изменений в составе донных сообществ.Freshw Biol 54 (10): 2086–2100
Статья Google Scholar
McKie BG, Petrin Z, Malmqvist B (2006) Смягчение или нарушение? Влияние известкования на структуру сообщества макробеспозвоночных и разложение листового опада в гуминовых потоках северной Швеции. J Appl Ecol 43 (4): 780–791
Статья Google Scholar
Макки Б.Г., Шиндлер М., Гесснер М.О., Мальмквист Б. (2009) Размещение биоразнообразия и функционирования экосистемы в контексте: нарушения окружающей среды и эффекты богатства видов в эксперименте с полевым водотоком.Oecologia 160 (4): 757–770
PubMed Статья Google Scholar
Макки Б.Г., Вудворд Г., Хладиз С. и др. (2008) Экосистема, функционирующая в сообществах ручьев из разных регионов: противоположные реакции на изменение в составе, равномерности и плотности детритофагов. J Anim Ecol 77 (3): 495–504
CAS PubMed Статья Google Scholar
Мейер Э. (1989) Взаимосвязь между параметрами длины тела и сухой массой у проточных водных беспозвоночных.Arch Hydrobiol 117 (2): 191–203
Google Scholar
Наим С., Райт Дж. П. (2003) Выявление влияния биоразнообразия на функционирование экосистемы: поиск решений, казалось бы, непреодолимой проблемы. Ecol Lett 6: 567–579
Статья Google Scholar
Naturvårdsverket (2003) Handledning for miljöövervakning — Undersökningstyp: Makrofyter i vattendrag. Версия 1: 2, 2004-12-04
Naturvårdsverket (2010) Handledning for miljöövervakning — Undersökningstyp: Bottenfauna i sjöars litoral och vattendrag — tidsserier.Версия 1: 1, 2010-03-01
O’Connor NE, Donohue I (2013) Экологический контекст определяет мультитрофические эффекты потери видов-потребителей. Glob Change Biol 19 (2): 431–440
Статья Google Scholar
Оксанен Дж., Бланшет Ф.Г., Руланд Киндт Р и др. (2015) веган: пакет экологических исследований сообщества. Версия пакета 2.3-1
Perkins DM, Mckie BG, Malmqvist B, Gilmour SG, Reiss J, Woodward G (2010) Потепление окружающей среды и функционирование биоразнообразия-экосистемы в пресноводных микрокосмах: разделение эффектов видовой идентичности, богатства и метаболизма .Adv Ecol Res 43: 177–209
Статья Google Scholar
Pinel-Alloul B, Niyonsenga T, Legendre P (1995) Пространственные и экологические компоненты структуры пресноводного зоопланктона. Écoscience 2 (1): 1–19
Статья Google Scholar
Pringle RM, Doak DF, Brody AK, Jocqué R, Palmer TM (2010) Пространственный образец улучшает функционирование экосистемы в африканской саванне.PLoS Biol 8 (5): e1000377
PubMed PubMed Central Статья CAS Google Scholar
Raab B, Vedin H (1995) Sveriges Nationalatlas: klimat, sjöar och vattendrag. Бюстгальтер böcker, Höganäs
Google Scholar
Рафаэлли Д., Ван дер Путтен WH, Перссон Л. и др. (2002) Мульти-трофическая динамика и экосистемные процессы. В: Loreau M, Naeem S, Inchausti P (eds) Биоразнообразие и функционирование экосистем, синтез и перспективы.Oxford University Press, Oxford, pp. 147–154
Google Scholar
R Core Team (2018) R: язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. https://www.R-project.org/
Reice SR (1994) Неравновесные детерминанты структуры биологического сообщества. Am Sci 82 (5): 424–435
Google Scholar
Schmidt-Kloiber A, Hering D (2015) www.freshwaterecology.info — онлайн-инструмент, который объединяет, стандартизирует и систематизирует более 20 000 европейских пресноводных организмов и их экологические предпочтения. Ecol Indic 53: 271–282
Статья Google Scholar
Schmitz OJ (2010) Пространственная динамика и функционирование экосистемы. PLoS Biol 8 (5): e1000378
PubMed PubMed Central Статья CAS Google Scholar
Спонселлер Р.А., Темнеруд Дж., Бишоп К., Лаудон Х. (2014) Модели и факторы речного азота (N) в альпийской, субарктической и северной Швеции.Биогеохимия 120 (1): 105–120
CAS Статья Google Scholar
Шривастава Д.С., Велленд М. (2005) Исследование функций биоразнообразия и экосистемы: актуально ли это для сохранения? Annu Rev Ecol Evol Syst 36: 267–294
Статья Google Scholar
Tachet H, Bournaud M, Richoux P, Usseglio-Polatera P (2010) Invertébrés d’eau douce — systématique, biologie, écologie.Издания CNRS, Париж
Google Scholar
Törnroos A, Nordström MC, Aarnio K, Bonsdorff E (2015) Экологический контекст и пластичность трофических признаков у ключевого вида, моллюска-теллинида Macoma balthica L. J Exp Mar Biol Ecol 472: 32–40
Статья Google Scholar
Тручи А., Анджелер Д.Г., Спонселлер Р.А., Джонсон Р.К., Макки Б.Г. (2015) Глава вторая — увязка биоразнообразия, функционирования экосистем и услуг, а также экологической устойчивости: на пути к интеграционной структуре для улучшенного управления.В: Guy W, David AB (eds) Достижения в экологических исследованиях. Academic Press, Лондон, стр. 55–96
Google Scholar
van Donk E, Gulati RD, Iedema A, Meulemans JT (1993) Связанные с макрофитами сдвиги в содержании азота и фосфора на различных трофических уровнях в мелководном озере с биоманипуляциями. Hydrobiologia 251 (1): 19–26
Статья Google Scholar
Venail PA, Maclean RC, Meynard CN, Mouquet N (2010) Dispersal увеличивает взаимосвязь между биоразнообразием и продуктивностью в экспериментальном метасообществе источник-поглотитель.Proc R Soc Lond B 277 (1692): 2339–2345
Статья Google Scholar
Violle C, Navas M-L, Vile D et al (2007) Пусть концепция черты будет функциональной! Oikos 116 (5): 882–892
Статья Google Scholar
Фон Шиллер Д., Марте Э., Риера Дж.Л., Рибо М., Маркс Дж. К., Сабатер Ф. (2008) Влияние землепользования на функцию экосистемы водотока в водосборе Средиземного моря.Freshw Biol 53 (12): 2600–2612
Статья CAS Google Scholar
Widenfalk LA, Leinaas HP, Bengtsson J, Birkemoe T (2018) Возраст и уровень самоорганизации влияют на мелкомасштабное распространение коллембол (Collembola). Экосфера 9 (1): e02058
Статья Google Scholar
Woodward G, Gessner MO, Giller PS et al (2012) Воздействие загрязнения питательными веществами на функционирование экосистемы водотока в континентальном масштабе.Наука 336 (6087): 1438–1440
CAS PubMed PubMed Central Статья Google Scholar
Янг Р.Г., Коллиер К.Дж. (2009) Противопоставление реакции на изменение водосбора ряда функциональных и структурных показателей здоровья речной экосистемы. Freshw Biol 54 (10): 2155–2170
CAS Статья Google Scholar
Пространственная структура и экологический анализ
Allen, T.Ф.Х., Бартелл, С. И Кунсе, Дж. Ф. 1977. Множественные стабильные конфигурации в распределении скоростей изменения сообщества фитопланктона. Экология 58: 1076–1084.
Google Scholar
Allen, T.F.H. И Старр, Т. 1982. Иерархия — перспективы экологической сложности. Univ. Чикаго Пресс, Чикаго.
Google Scholar
Андерссон, П.-А. 1988. Упорядочение и классификация операционных географических единиц на юго-западе Швеции.Vegetatio 74: 95–106.
Google Scholar
Белл М.А. и Лежандр П. 1987. Многосимвольная хронологическая кластеризация в последовательности ископаемых колючек. Syst. Zool. 36: 52–61.
Google Scholar
Беннетт Р.Дж., Хайнинг Р.П. и Гриффит Д.А. 1984. Проблема отсутствия данных на пространственных поверхностях. Аня. Доц. Являюсь. Геогр. 74: 138–156.
Google Scholar
Биванд, Р.1980. Исследование методом Монте-Карло оценки коэффициента корреляции с пространственно автокоррелированными наблюдениями. Квест. Геогр. 6: 5–10.
Google Scholar
Ботинки, B.N. 1977. Свойства контактного номера в исследовании сотовых сетей. Геогр. Анальный. 9: 379–387.
Google Scholar
Бушар А., Бержерон Ю., Камире К., Ганглофф П. и Гариепи М. 1985.Proposition d’une méthodologie d’inventaire et de cartographie écologique: le cas de la MRC du Haut-Saint-Laurent. Да. Géogr. Qué. 29: 79–95.
Google Scholar
Box, G.E.P. И Дженкинс, Г. 1970. Анализ временных рядов, прогнозирование и управление. Холден-Дэй, Сан-Франциско.
Google Scholar
Burgman, M.A. 1987. Анализ распределения растений на обнажениях гранита в южной части Западной Австралии с использованием тестов Mantel.Vegetatio 71: 79–86.
Google Scholar
Берроу, П.А. 1986. Принципы географических информационных систем для оценки земельных ресурсов. Кларендон Пресс, Оксфорд.
Google Scholar
Берроу, П.А. 1987. Пространственные аспекты экологических данных. В: Jongman, R.H.G., terBraak, C.J.F. & vanTongeren, O.F.R. (ред.), Анализ данных в сообществе и ландшафтной экологии, стр. 213–251.Пудок, Вагенинген.
Google Scholar
Carpenter, S.R. И Чейни, Дж. Э. 1983. Масштаб пространственного рисунка: сравнение четырех методов. Vegetatio 53: 153–160.
Google Scholar
Cicéri, M.-F., Marchand, B. & Rimbert, S. 1977. Introduction à l’analyse de l’espace. Применимо Collection de Géographie. Массон, Париж.
Google Scholar
Клифф, А.Д. и Орд, Дж. К. 1981. Пространственные процессы: модели и приложения. Пион Лимитед, Лондон.
Google Scholar
Cochran, W.G. 1977. Методы отбора проб, 3-е изд. John Wiley & Sons, Нью-Йорк.
Google Scholar
Купер Д.У. 1968. Уровень значимости в нескольких тестах, выполненных одновременно. Наследственность 23: 614–617.
Google Scholar
Кертис, Д.J. & Bignal, E.M. 1985. Количественное описание физиогномики растительности с использованием вертикальных квадратов. Vegetatio 63: 97–104.
Google Scholar
Дарвин, К. 1881. Образование плесени в овощах под действием червей. Джон Мюррей, Лондон.
Google Scholar
Дэвид М. 1977 г. Геостатистическая оценка запасов руды. Развитие геоматематики, 2.Эльзевир, Амстердам.
Google Scholar
Дэвис, Дж. К. 1986. Статистика и анализ данных в геологии, 2-е изд. John Wiley & Sons, Нью-Йорк.
Google Scholar
Дитц, Э.Дж. 1983. Перестановочные тесты для ассоциации между двумя матрицами расстояний. Syst. Zool. 32: 21–26.
Google Scholar
Доу, М.М. и Чеверуд, Дж. М. 1985. Сравнение матриц расстояний в исследованиях популяционной структуры и генетической микродифференциации: квадратичное присвоение. Являюсь. J. Phys. Антрополь. 68: 367–373.
Google Scholar
Edgington, E.S. 1987. Рандомизационные тесты, 2-е изд. Марсель Деккер Инк., Нью-Йорк.
Google Scholar
Edwards, D. & Coull, B.C. 1987. Анализ тенденций авторегрессии: пример с использованием долгосрочных экологических данных.Ойкос 50: 95–102.
Google Scholar
Estabrook, G.F. И Гейтс Б. 1984. Анализ характера комплекса Banisteriopsis campestris (Malpighiaceae) с использованием пространственной автокорреляции. Таксон 33: 13–25.
Google Scholar
Estabrook, G.F. И Роджерс, Д. 1966. Общий метод таксономического описания для вычисляемой меры сходства.BioScience 16: 789–793.
Google Scholar
Фишер, У.Д. 1958. О группировке для максимальной однородности. J. Amer. Стат. Жопа. 53: 789–798.
Google Scholar
Ford, E.D. 1976. Полог сосны обыкновенной: описание поверхности сложной шероховатости. Agric. Meteorol. 17: 9–32.
Google Scholar
Ford, E.Д. и Реншоу, Э. 1984. Интерпретация процесса по образцу с использованием двухмерного спектрального анализа: моделирование моделей отдельных видов в растительности. Vegetatio 56: 113–123.
Google Scholar
Fortin, M.-J. 1985. Анализируйте пространственное разделение экологических феноменов: методы пространственного анализа, теория л’эчантильоннажа. Mémoire de Maîtrise, Université de Montréal.
Габриэль К.Р. и Сокал, Р. Р. 1969. Новый статистический подход к анализу географических вариаций. Syst. Zool. 18: 259–278.
Google Scholar
Галиано, E.F. 1982. Обнаружение закономерностей в популяциях растений посредством анализа расстояний от растения до всех растений. Vegetatio 49: 39–43.
Google Scholar
Галиано, E.F. 1983. Обнаружение многовидовых моделей в популяциях растений.Vegetatio 53: 129–138.
Google Scholar
Галзин Р. и Лежандр П. 1988. Рыбные сообщества трансекты коралловых рифов. Pac. Sci. 41: 158–165.
Google Scholar
GauchJr., H.G. 1982. Многомерный анализ в общественной экологии. Cambridge Univ. Press, Кембридж.
Google Scholar
Гири, Р.С. 1954. Коэффициент смежности и статистическое отображение. Incorp. Статист. 5: 115–145.
Google Scholar
Getis, A. & Boots, B. 1978. Модели пространственных процессов: подход к изучению точечных, линейных и площадных структур. Cambridge Univ. Press, Кембридж.
Google Scholar
Гиттинс Р. 1968. Анализ поверхности тренда экологических данных. J. Ecol. 56: 845–869.
Google Scholar
Goodall, D.W. 1974. Новый метод анализа пространственной картины путем случайного объединения квадратов в пары. Vegetatio 29: 135–146.
Google Scholar
Гордон, А. Д. 1973. Классификация при наличии ограничений. Биометрия 29: 821–827.
Google Scholar
Гордон, А.Д. и Биркс, Х.Дж. 1972. Численные методы в четвертичной палеоэкологии. I. Зонирование пыльцевых диаграмм. Новый Фитол. 71: 961–979.
Google Scholar
Gordon, A.D. & Birks, H.J.B. 1974. Численные методы в четвертичной палеоэкологии. II. Сравнение диаграмм пыльцы. Новый Фитол. 73: 221–249.
Google Scholar
Гауэр, Дж. К. и Лежандр, П. 1986.Метрические и евклидовы свойства коэффициентов несходства. J. Classif. 3: 5–48.
Google Scholar
Gower, J.C. & Ross, G.J.S. 1969. Минимальные остовные деревья и кластерный анализ единственной связи. Прил. Статист. 18: 54–64.
Google Scholar
Грин, Р.Х. 1979. Дизайн выборки и статистические методы для экологов-биологов. John Wiley & Sons, Нью-Йорк.
Google Scholar
Hassell, M.P. И Мэй, Р. 1974. Агрегация хищников и насекомых-паразитов и ее влияние на устойчивость. J. Anim. Ecol. 43: 567–594.
Google Scholar
Хокинс, Д.М. И Мерриам, Д.Ф. 1973. Оптимальное зонирование оцифрованных последовательных данных. J. Int. Доц. Математика. Геология 5: 389–395.
Google Scholar
Хокинс, Д.М. и Мерриам, Д.Ф. 1974. Зонирование многомерных последовательностей оцифрованных геологических данных. J. Int. Доц. Математика. Геология 6: 263–269.
Google Scholar
Hensen, V. 1884. Ueber die Bestimmung der Planktons oder des im Meer triebenden Materials an Pflanzen und Tieren. Бер. Comm. Wiss. Unters. Deutsch. Meere 5.
Hubert, L.J. 1985. Комбинаторный анализ данных: ассоциация и частичная ассоциация. Психометрика 50: 449–467.
Google Scholar
Hudon, C. & Lamarche, G. в печати. Разделение ниши между американским лобстером ( Homarus americanus ) и каменным крабом ( Cancer irroratus ). Mar. Ecol. Прог. Сер.
Хаффакер, К. Б. 1958. Экспериментальные исследования хищничества: факторы дисперсии и колебания хищник-жертва. Хильгардия 27: 343–383.
Google Scholar
Ибанез, Ф.1981. Немедленное обнаружение неоднородностей в непрерывных многомерных океанографических записях. Приложение к анализу временных рядов изменений в заливе Вильфранш-сюр-Мер. Лимнол. Oceanogr. 26: 336–349.
Google Scholar
Journel, A.G. & Huijbregts, C. 1978. Горная геостатистика. Academic Press, Лондон.
Google Scholar
Джумарс, П.А., Thistle, D. & Jones, M.L. 1977. Обнаружение двумерной пространственной структуры в биологических данных. Oecologia (Berl.) 28: 109–123.
Google Scholar
Кракхардт, Д. 1988. Прогнозирование с помощью сетей: непараметрический множественный регрессионный анализ двоичных данных. Социальные сети 10: 359–381.
Google Scholar
Krige, D.G. 1966. Двумерные взвешенные поверхности тренда скользящего среднего для оценки руды.Jour. S. Afr. Inst. Мин. Металл. 66: 13–38.
Google Scholar
Лам, Н.С. 1983. Методы пространственной интерполяции: обзор. Являюсь. Картогр. 10: 129–149.
Google Scholar
Lebart, L. 1978. Program d’agrégation avec contraintes (C.A.H. contiguïté). Да. Анальный. Données 3: 275–287.
Google Scholar
Ли, П.J. 1981. Самый предсказуемый метод картирования поверхности (MPS) при разведке нефти. Бык. Может. Бензин. Геол. 29: 224–240.
Google Scholar
Лефкович Л.П. 1978. Генерация и группировка кластеров с использованием математического программирования. Математика. Biosci. 41: 91–110.
Google Scholar
Лефкович, Л.П. 1980. Условная кластеризация. Биометрия 36: 43–58.
Google Scholar
Лежандр, Л.И Демерс, С. 1985. Вспомогательная энергия, эргоклины и водная биологическая продукция. Натуралист может. (Qué.) 112: 5–14.
Google Scholar
Лежандр, Л. и Лежандр, П., 1983a Численная экология. Развитие моделирования окружающей среды, 3. Elsevier, Амстердам.
Google Scholar
Legendre, L. & Legendre, P. 1983b. Разделение упорядоченных переменных на дискретные состояния для дискриминантного анализа экологических классификаций.Может. J. Zool. 61: 1002–1010.
Google Scholar
Legendre, L. & Legendre, P. 1984a. Écologie numérique, 2ième éd. Том 2: La structure des données écologiques. Masson, Paris et les Presses de l’Université du Québec.
Лежандр, П. 1987. Ограниченная кластеризация. В: Лежандр, П. и Лежандр, Л. (ред.), Развитие численной экологии. Серия НАТО ASI, Vol. G 14, стр. 289–307. Спрингер, Берлин.
Google Scholar
Лежандр П., Далло С., Лежандр Л. 1985. Последовательность видов в сообществе: хронологическая кластеризация с применением к морскому и пресноводному зоопланктону. Являюсь. Nat. 125: 257–288.
Google Scholar
Лежандр, П. и Лежандр, В. 1984b. Постледниковое расселение пресноводных рыб на полуострове Квебек. Джан Дж. Фиш. Акват.Sci. 41: 1781–1802.
Google Scholar
Лежандр П. и Трусселье М. 1988. Водные гетеротрофные бактерии: моделирование в присутствии пространственной автокорреляции. Лимнол. Oceanogr. 33: 1055–1067.
Google Scholar
Левин Л.А. 1984. Жизненный цикл и характер расселения в плотном инфаунальном сообществе полихет: структура сообщества и реакция на нарушение.Экология 65: 1185–1200.
Google Scholar
MacDonald, G.M. & Waters, N.M. 1988. Использование наиболее предсказуемых поверхностей для классификации и картирования ассоциаций таксонов. Vegetatio 74: 125–135.
Google Scholar
Маккуин, Дж. Б. 1967. Некоторые методы классификации и анализа многомерных наблюдений. В: LeCam, L.M. & Neyman, J. (eds), Proc.Пятый симпозиум Беркли. Математика. Стат. Вероятно. Vol. 1. С. 281–297. Калифорнийский университет Press, Беркли.
Google Scholar
Manly, B.F.J. 1986. Методы рандомизации и регрессии для проверки ассоциаций с географическими, экологическими и биологическими расстояниями между популяциями. Res. Popul. Ecol. 28: 201–218.
Google Scholar
Mantel, N. 1967.Обнаружение кластеризации болезней и обобщенный регрессионный подход. Cancer Res. 27: 209–220.
Google Scholar
Matheron, G., 1962. Traité de géostatistique appliquée. Том 1. Издательство Technip, Париж.
Google Scholar
Матерон, Г. 1973. Внутренние случайные функции и их приложения. Adv. Прил. Вероятно. 5: 439–468.
Google Scholar
Мэй, р.М. 1974. Устойчивость и сложность в модельных экосистемах, 2-е изд. Princeton Univ. Press, Принстон, Нью-Джерси.
Google Scholar
McBratney, A.B. & Webster, R. 1981. Определение рисунка гребней и борозд с помощью спектрального анализа урожайности сельскохозяйственных культур. Int. Стат. Откр. 49: 45–52.
Google Scholar
McCune, B. & Allen, T.F.H. 1985. Будет ли подобный лес развиваться на подобных участках? Может.J. Bot. 63: 367–376.
Google Scholar
Miles, R.E. 1970. Об однородном плоском точечном процессе Пуассона. Математика. Biosci. 6: 85–127.
Google Scholar
MillerJr, R.G. 1977. Развитие множественных сравнений. J. Amer. Стат. Жопа. 72: 779–788.
Google Scholar
Moran, P.A.P.1950. Заметки о непрерывных случайных явлениях. Биометрика 37: 17–23.
Google Scholar
Ньюбери, округ Колумбия, Реншоу, Э. и Брюниг, Э.Ф. 1986 Пространственный узор деревьев в лесу керангас, Саравак. Vegetatio 65: 77–89.
Google Scholar
Oden, N.L. 1984. Оценка значимости пространственной коррелограммы. Геогр. Анальный. 16: 1–16.
Google Scholar
Оден, Н.Л. и Сокал, Р. Р. 1986. Направленная автокорреляция: расширение пространственных коррелограмм до двух измерений. Syst. Zool. 35: 608–617.
Google Scholar
Openshaw, S. 1974. Программа регионализации для больших наборов данных. Компьютерное приложение 3–4: 136–160.
Google Scholar
Орлоци Л. 1978. Многомерный анализ в исследованиях растительности, 2-е изд. Джанк, Гаага.
Google Scholar
Филлипс, Дж. Д. 1985. Измерение сложности градиентов окружающей среды. Vegetatio 64: 95–102.
Google Scholar
Пиелу Э.С. 1977 Математическая экология, 2-е изд. J. Wiley & Sons, Нью-Йорк.
Google Scholar
Pielou, E.C.1984. Интерпретация экологических данных.Букварь по классификации и рукоположению. John Wiley & Sons, Нью-Йорк.
Google Scholar
Пристли, М. 1964. Анализ двумерных стационарных процессов с разрывными спектрами. Биометрика 51: 195–217.
Google Scholar
Ray, D.M. И Берри, Б.Дж.Л. 1966. Многомерная социально-экономическая регионализация: экспериментальное исследование в центральной Канаде.В: Остри С. и Раймс Т. (редакторы), Документы по региональным статистическим исследованиям, стр. 75–130. Univ. Торонто Пресс, Торонто.
Google Scholar
Rayner, J.N. 1971. Введение в спектральный анализ. Пион Лтд., Лондон.
Google Scholar
Renshaw, E. & Ford, E.D. 1983. Интерпретация процесса по образцу с помощью двумерного спектрального анализа: методы и проблемы интерпретации.Прил. Статист. 32: 51–63.
Google Scholar
Renshaw, E. & Ford, E.D. 1984. Описание пространственной картины с помощью двухмерного спектрального анализа. Vegetatio 56: 75–85.
Google Scholar
Рипли, Б.Д. 1981. Пространственная статистика. John Wiley & Sons, Нью-Йорк.
Google Scholar
Рипли, Б.Д. 1987. Анализ пространственных точек в экологии. В: Лежандр, П. и Лежандр, Л. (ред.), Развитие численной экологии. Серия НАТО ASI, Vol. G 14, стр. 407–429, Springer-Verlag, Berlin.
Google Scholar
Сакаи, А.К. И Оден, Н. 1983. Пространственный образец выражения пола у серебряного клена ( Acer saccharinum L.): индекс Мориситы и пространственная автокорреляция. Являюсь. Nat. 122 489–508.
Google Scholar
Шеррер, Б.1982. Techniques de sondage en écologie. В: Frontier, S. (ed.), Stratégies d’échantillonnage en écologie, стр. 63–162. Collection d’Écologie, 17. Masson, Paris et les Presses de l’Université Laval, Квебек.
Google Scholar
Silverman, B.W. 1986. Оценка плотности для статистики и анализа данных. Чепмен и Холл, Лондон.
Google Scholar
Smouse, P.Э., Лонг, Дж. К. и Сокал, Р. Р. 1986. Множественные регрессионные и корреляционные расширения теста Мантеля на соответствие матриц. Syst. Zool. 35: 627–632.
Google Scholar
Sneath, P.H.A. 1966. Сравнение различных методов кластеризации применительно к случайно расположенным точкам. Классификация Soc. Бык. 1: 2–18.
Google Scholar
Сокал Р.Р., 1979. Экологические параметры, полученные на основе пространственных коррелограмм.В: Патил, Г. И Розенцвейг, М.Л., (ред.), Современная количественная экология и родственная экометрика. Статистическая серия по экологии, Vol. 12. С. 167–196. International Co-operative Publ. Дом, Фэрленд, Мэриленд.
Google Scholar
Сокал Р.Р. 1983. Анализ вариаций характера в географическом пространстве. В: Фельзенштейн, Дж. (Ред.), Числовая таксономия. Серия НАТО ASI, Vol. G1, стр. 384–403. Springer-Verlag, Берлин.
Google Scholar
Сокаль, р.Р. 1986. Анализ пространственных данных и исторические процессы. В: Diday, E. et al. (ред.), Анализ данных и информатика, IV. Proc. Четвертый Int. Symp. Data Anal. Информатика, Версаль, Франция, 1985, стр. 29–43. Северная Голландия, Амстердам.
Google Scholar
Sokal, R.R. & Oden, N.L. 1978. Пространственная автокорреляция в биологии. 1. Методология, биол. J. Linnean Soc. 10: 199–228.
Google Scholar
Сокаль, р.Р. и Томсон, Дж. Д. 1987. Применение пространственной автокорреляции в экологии. В: Лежандр, П. и Лежандр, Л. (ред.), Развитие численной экологии. Серия НАТО ASI, Vol. G 14, стр. 431–466. Springer-Verlag, Берлин.
Google Scholar
Тапиа, Р.А. И Томпсон, Дж. 1978. Непараметрическая оценка плотности вероятности. Johns Hopkins Univ. Пресса, Балтимор.
Google Scholar
терБраак, К.J.F. 1986. Канонический анализ соответствий: новый метод собственных векторов для многомерного прямого градиентного анализа. Экология 67: 1167–1179.
Google Scholar
terBraak, C.J.F. 1987. Анализ взаимосвязи растительности и окружающей среды с помощью анализа канонических соответствий. Vegetatio 69: 69–77.
Google Scholar
Аптон, Г.Дж.Г. И Финглтон Б. 1985.Анализ пространственных данных на примере. Vol. 1: Точечный рисунок и количественные данные. Джон Вили и сыновья, Чичестер.
Google Scholar
Wartenberg, D. 1985a. Канонический анализ поверхности тренда: метод описания географических закономерностей. Syst. Zool. 34: 259–279.
Google Scholar
Wartenberg, D. 1985b. Многомерная пространственная корреляция: метод исследовательского географического анализа.Геогр. Анальный. 17: 263–283.
Google Scholar
Уотсон, Д.Ф. 1981. Вычисление n-мерной мозаики Делоне с применением к многоугольникам Вороного. Компьютер J. 24: 167–172.
Google Scholar
Webster, R. 1973 Автоматическое определение границы почвы по данным разреза. J. Int. Доц. Математика. Геол. 5: 27–37.
Google Scholar
Вебстер, Р.И Берроу П.А. 1972. Компьютерное картирование почв небольших территорий по выборочным данным. I. Многомерная классификация и ординация. II. Классификация сглаживания. J. Почвоведение. 23: 210–221, 222–234.
Google Scholar
Границы | Влияние пространственных и временных масштабов нарушений на стабильность экосистемы
1. Введение
Понимание стабильности экосистем, то есть их способности восстанавливаться и сохраняться перед лицом естественных и антропогенных нарушений, имеет фундаментальное значение для экологии и сохранения (May, 1973; Neubert and Caswell, 1997; Loreau and de Mazancourt, 2013) .Экосистемы пространственно расширены, состоят из множества взаимодействующих сообществ в разных местах, и поэтому важным фактором в понимании их стабильности является их пространственная структура (Levin, 1992; Peterson et al., 1998; Wang and Loreau, 2016). Однако, хотя влияние космоса на такие свойства, как биоразнообразие и структура пищевой сети, интенсивно исследовалось (Loreau et al., 2001; Chase and Leibold, 2002; Montoya and Sol, 2002; McCann et al., 2005), основные вопросы относительно пространственной устойчивости остаются открытыми.В частности, несмотря на то, что большинство возмущений (например, пожары, нашествия вредителей, загрязняющие стоки) сильно неоднородны в пространстве, влияние их пространственной структуры на стабильность в значительной степени неизвестно. Точно так же их временное измерение, например, их временной промежуток или частота их появления, имеет решающее значение. Взятые вместе, эти измерения охватывают огромное пространство возможных нарушений, с которыми могут столкнуться экосистемы (например, пожары и штормы). Это отчасти объясняет, почему достижение четкого понимания стабильности экосистемы оказалось чрезвычайно сложной задачей.
Исследования стабильности экосистем имеют долгую историю в области экологии, и многочисленные исследования изучали, как различные свойства нарушений влияют на реакцию экосистем. В частности, важность пространственных свойств возмущений оценивалась несколькими исследованиями динамики регенерации при повторяющихся, пространственно структурированных возмущениях (Turner et al., 1993; Moloney, Levin, 1996; Fraterrigo, Rusak, 2008). Эти исследования представили концепцию ландшафтного равновесия и продемонстрировали, как пространственные и временные масштабы нарушений могут порождать различные модели устойчивости.Однако в этих исследованиях прямо не рассматривается важность динамики спасения, происходящей в региональном масштабе, когда локальные процессы восстановления слишком медленны или вообще не работают. Это может произойти в достаточно связанных экосистемах после сильных нарушений (Foster et al., 1998; Fraterrigo and Rusak, 2008). Фактически, восстановление после нарушения является следствием как местных, так и региональных процессов. Локальные процессы приводят к восстановлению благодаря внутренней динамике местных сообществ (например,g., рождение и смерть особей), в то время как региональные процессы приводят к восстановлению, привлекая особей из соседних сообществ через расселение (Turner, 1989; Leibold et al., 2004). Эти два процесса опосредуют реакцию крупномасштабной системы на возмущение, и их соответствующие части в этом ответе должны сильно зависеть от пространственной связи системы и, что важно, от пространственной структуры возмущений.
Недавняя работа сделала эту взаимосвязь более явной, определив три различных режима восстановления после одного пространственно неоднородного возмущения: Изолированный, Спасательный и Смешивающий (Zelnik et al., 2018). Если система сильно связана из-за сильного расселения организмов, то она находится в режиме смешивания, и поведение системы в больших масштабах, по сути, является расширенной версией локальной системы (Durrett and Levin, 1994). С другой стороны, если рассеяние невелико и, следовательно, каждый сайт действует отдельно со своей собственной локальной динамикой, тогда система находится в изолированном режиме, а ее крупномасштабное поведение представляет собой совокупность множества независимых небольших систем (Tilman et al., 1998; Ячи и Лоро, 1999).Между этими двумя крайностями находится режим спасения, в котором системы с промежуточным подключением демонстрируют крупномасштабную динамику спасения из-за взаимодействия между ограниченным рассредоточением и поведением системы в локальном масштабе (Peterson, 2000; Dai et al., 2013; Wang et al., др., 2017). Например, в исследовании Dai et al. (2013), метапопуляция дрожжей демонстрирует фронтальную структуру, которая возникает из-за взаимодействия дисперсии с нелинейным локальным поведением дрожжей. Другой пример можно найти в работе Wang et al.(2017), где корреляции между местными популяциями птиц, опосредованные расселением, приводят к закону пространственного масштабирования изменчивости популяций по всей Северной Америке.
В то время как пространственная структура как системы, так и возмущения не играет роли в режиме перемешивания, при более слабом рассредоточении она играет: как в изолированном режиме, так и в режиме спасения пространственная структура возмущения имеет значительные эффекты, поскольку может инициировать качественно разные реакции, которые вовлекают как местные, так и региональные процессы (Zelnik et al., 2018). Так обстоит дело в экспериментальном исследовании системы хищник-жертва-протист, в которой на локальные вымирания отвечают процессы спасения, которые препятствуют синхронизации региональной метапопуляции (Fox et al., 2017). Поэтому мы рассмотрим системы с промежуточным рассредоточением и сосредоточимся на влиянии пространственной структуры возмущений, а также на их временных свойствах.
Количественная оценка воздействия возмущений сводится к определению соответствующих мер устойчивости. Если возмущение является изолированным событием, естественной мерой, которую следует учитывать, является время возврата в невозмущенное состояние (May, 1973; Neubert and Caswell, 1997).С другой стороны, в режиме повторяющихся возмущений (например, климатических явлений) обычно используются меры временной изменчивости (Tilman et al., 2006). При наличии альтернативных стабильных состояний эти повторяющиеся возмущения могут вызвать переключение режима из одного состояния в другое. Одним из хорошо известных примеров является эвтрофикация озера (Карпентер, 2005) из-за нарушения стока удобрений. Здесь интересующей мерой стабильности обычно является устойчивость, то есть вероятность того, что система останется в желаемом состоянии (Holling, 1973; Pimm, 1984).Важно отметить, что эти меры устойчивости отражают не только пространственные и временные свойства возмущения, но и динамические особенности нарушенной экосистемы. Изучение этого взаимодействия является целью нашего исследования, которое мы рассмотрим, рассмотрев три пространственные модели экосистемы с возрастающей нелинейной локальной динамикой, от логистического роста до бистабильности. При различных сценариях возмущений мы будем измерять их стабильность, используя время возврата, изменчивость и постоянство.
Мы начнем с рассмотрения восстановления экосистемы после единичного нарушения и покажем, что изменение пространственной структуры нарушения выявляет две основные траектории восстановления: изолированную и спасательную.Изолированные траектории восстановления отражают локальную устойчивость системы, в то время как траектории спасения связаны с пространственными процессами, и их доминирование сигнализирует о сбое локальных процессов. Таким образом, мы утверждаем, что взаимосвязь между пространственной структурой и восстановлением содержит существенную информацию о локальной динамике системы, как близкой к равновесию, так и вдали от нее. Мы продолжаем исследовать временную ось возмущений и демонстрируем прямую связь между временем возврата (после отдельного возмущающего события) и временной изменчивостью (в режиме повторяющихся возмущений).Мы обнаружили, что для частот с низким уровнем помех образцы изменчивости не содержат дополнительной информации по сравнению с образцами времени возврата. Однако после порога частоты (который зависит от внутренней динамики системы) паттерны изменчивости изменяются. Как мы будем утверждать, это сигнализирует о наступлении нового динамического режима, вызванного возмущениями, который может привести к смене режима — в нашем случае переходу от заселенного к голому состоянию (вымирание).
Наша работа демонстрирует, что пространственное измерение нарушений может быть использовано для выявления информации о внутреннем поведении экосистемы.Кроме того, наши результаты показывают, что сочетание пространственных и временных свойств возмущений может привести к непредвиденным динамическим реакциям с серьезными экологическими последствиями.
2. Методы
2.1. Модели
Мы предполагаем для простоты, что динамика местного сообщества может быть описана одной переменной состояния N , которая представляет локальную плотность биомассы экосистемы. Мы изучаем динамику в нескольких точках пространства, используя уравнения в частных производных.Мы определяем три разные модели, которые различаются по своей локальной динамике, но имеют одинаковое распространение в пространстве с линейной диффузией. Во всех моделях локальная биомасса может достигать пропускной способности K , так что N = K (заселенное состояние) является стабильным устойчивым состоянием во всех трех моделях. Дополнительное решение существует для N = 0 (голое состояние), причем его свойства устойчивости различаются между моделями.
Первая и простейшая модель (LG) описывает локальный логистический рост в сочетании с рассредоточением:
Nt = rN (1-N / K) + d∇2N, (1), где N t — это изменение локальной биомассы во времени, а ∇ 2 N — вторая производная в пространстве от N (диффузионный член).Здесь r — характерная локальная динамическая скорость роста, в то время как скорость распространения за счет рассеивания определяется величиной d . В этой модели голое состояние N = 0 является нестабильным решением. Это классическая модель роста популяции (Hall, 1988), которая, как было показано, надлежащим образом изображает динамику различных биологических систем, от роста одноклеточных организмов (Gause, 1934) до человеческих популяций (Marchetti et al., 1996).
Вторая модель (AE) описывает динамику видов с сильным эффектом Алли (Kramer et al., 2009), так что низкие плотности биомассы нежизнеспособны. Такая динамика была обнаружена у множества видов, от непарного шелкопряда до лесного карибу (Kramer et al., 2009). Модель читает
Nt = rN (1-N / K) (N / α-1) + d∇2N, (2), где α — порог жизнеспособности, т.е. минимальное количество биомассы N , необходимое для положительного роста. Эта модель имеет два альтернативных стабильных состояния ( N = 0, N = K ) и одно нестабильное состояние ( N = α), и мы предполагаем, что 0 <α < K .Это простейшая модель динамики с альтернативными стабильными состояниями, свойство, которое было обнаружено во многих экосистемах (Scheffer, 2009), таких как озера (Carpenter, 2005) и коралловые рифы (Nyström et al., 2000).
Наконец, наша третья модель (SR) описывает динамику с медленным восстановлением после интенсивных возмущений и является промежуточным звеном между двумя предыдущими моделями. Это поможет нам прояснить различие между сильной нелинейностью и бистабильностью. Его главная особенность состоит в том, что, хотя существует только одно устойчивое равновесие при N = K , вдали от этого равновесия скорость возврата очень мала по сравнению со скоростью возврата, близкой к равновесной.Это могло бы моделировать динамику сукцессии, для которой восстановление после сильных нарушений (например, сплошных рубок) происходит очень медленно, так как оно включает последовательную колонизацию различными видами, а не просто возобновление роста нарушенных видов (Uhl, 1987) или слабое Эффект Аллее, преобладающий признак в динамике популяции (Kramer et al., 2009). Модель есть:
Nt = rN (1-N / K) (N / K) γ + d∇2N, (3), где γ управляет нелинейностью динамики, так что при высоких значениях γ локальное восстановление происходит очень медленно после возмущений высокой интенсивности.
Для каждой модели мы можем определить локальный потенциал (см. Левые панели рисунка 2), так что его производная по N соответствует производной N по времени, то есть локальной динамике. Это означает, что локальная динамика следует за наклоном этого потенциала, так что плотность биомассы можно рассматривать как шар, движущийся от пиков к долинам в ландшафте, который определяет потенциал. В обеих моделях LG и SR существует только одно устойчивое равновесие, но скорость возврата к равновесию может быть намного медленнее при низкой плотности биомассы в модели SR.В модели AE существует два стабильных состояния (заполненное состояние и голое состояние).
Изменяя масштаб времени, пространства и биомассы, мы можем эффективно уменьшить пространство параметров и установить r = 1, d = 1 и K = 1. Таким образом, наши результаты справедливы для любых значений этих трех параметров. Мы устанавливаем α = 0,4, чтобы убедиться, что модель AE восстанавливается после одиночного возмущения (см. Следующий подраздел), и γ = 4, чтобы убедиться, что время возврата вдали от равновесия модели SR является достаточно медленным.Мы фокусируемся на одномерных системах, поскольку их проще анализировать, но качественные результаты справедливы для других типов пространственных структур, таких как двумерные системы (см. Приложение D). Мы используем размер системы L = 500, который достаточно велик, чтобы позволить пространственной динамике проявить себя (так что система не находится в режиме смешивания Zelnik et al., 2018) с периодическими граничными условиями. Для наглядности на рисунках 3 и S2 мы показываем снимки двумерной системы размером 200 × 200.
2.2. Пространственное измерение возмущений
Мы определяем нарушение как изменение переменной состояния, которое воздействует на экосистему. Мы рассматриваем импульсное возмущение, возникающее в данный момент времени, и его полное действие применяется в это время. Это предположение подходит для многих типов нарушений, которые происходят быстрее, чем динамика экосистемы, и поддается математическому анализу. Мы выбираем возмущение, которое удаляет биомассу (уменьшает N ), так что нарушение силы s уменьшит общую биомассу экосистемы на sK (но любые отрицательные значения N будут установлены на 0 для последовательность).Как только происходит нарушение, экосистема может восстановиться до исходного состояния или может произойти смена режима, если экосистема является бистабильной. Здесь нас интересует динамика устойчивости и восстановления, поэтому мы сосредоточены на значениях параметров, для которых одно возмущение не может привести к смене режима.
Поскольку возмущение не обязательно должно происходить равномерно в пространстве, мы изменяем пространственную протяженность возмущения σ, сохраняя при этом его общую интенсивность с постоянной. Нарушение осуществляется путем выбора его локуса и удаления некоторой биомассы в области размера σ с центром вокруг локуса.Мы можем изменять пространственную протяженность от σ = 1 для однородного возмущения в пространстве до σ = с для локализованного возмущения.
Для измерения восстановления мы используем время возврата T , определяемое как время, необходимое экосистеме для восстановления 90% биомассы, потерянной в результате нарушения. Хотя выбор порога является произвольным, его конкретное значение не оказывает существенного влияния на результаты, если оно не слишком близко ни к 0%, ни к 100% (что примерно соответствует реактивности и асимптотической устойчивости, соответственно, Arnoldi et al., 2016). Избегая этих экстремальных значений, мы просто подчеркиваем роль, которую играет общая динамика восстановления, а не первоначальная реакция системы или окончательная конвергенция.
2.3. Временной размер нарушений
Мы рассматриваем режим возмущения, многократно применяя возмущения с заданной средней частотой f в течение периода времени τ. Для простоты мы предполагаем отсутствие корреляции ни в пространстве, ни во времени, так что время между возмущениями берется из экспоненциального распределения с некоторой средней частотой (процесс Пуассона, подробности см. В Приложении B), а местоположение центра возмущения берется из равномерное распределение.
Мы используем две меры стабильности для системы, которая постоянно нарушается, т. Е. Изменчивость, которая измеряет, насколько далеко система отклоняется от своего среднего значения, и устойчивость, которая измеряет, насколько вероятно, что она переместится в область притяжения другого равновесие. Мы определяем изменчивость V как изменение во времени общей биомассы системы, учитывая режим повторяющихся нарушений. Чтобы пренебречь влиянием переходных процессов, мы вычисляем В, по последним 80% симуляций, которые длятся 10 000 временных шагов.Мы определяем вероятность коллапса C как вероятность того, что система будет в голом состоянии в конце моделирования, так что C = 0 означает отсутствие шансов на коллапс системы, а C = 1 означает, что коллапс неизбежен. Мы используем более длительное время моделирования (100 000 временных шагов) для вычисления C , поскольку мы заинтересованы в прогнозировании коллапса до того, как оно произойдет. Для каждого из этих вычислений мы запускаем 100 симуляций с различными рандомизациями местоположения и времени возмущений.
3. Результаты
3.1. Расширение пространственного измерения нарушений выявляет динамику локальной экосистемы
Мы начнем с изучения реакции экосистемы на единичное нарушение с различной пространственной протяженностью σ. Мы сосредоточимся на возмущениях с фиксированной общей силой с = с 0 для простоты и ясности, и ослабим это предположение в обсуждении. Таким образом, глобальное возмущение σ = 1 (рисунок 1, правые панели) возникает, когда N уменьшается на с 0 K во всей системе, в то время как локальное возмущение σ = с 0 (рисунок 1, левые панели) возникает, когда N установлено равным нулю в области относительного размера s 0 .
Рисунок 1 . Динамика восстановления после локализованного и глобального возмущения (левая и правая панели соответственно) для бистабильной модели АЭ (см. Основной текст). Верхние панели: снимки в разное время (t) вдоль траекторий восстановления, каждый снимок показывает пространственный профиль биомассы. Нижние панели показывают изменение общей биомассы с течением времени после нарушения, где пунктирная линия обозначает порог, после которого система считается восстановленной, а красные кружки соответствуют снимкам.Обратите внимание, что время возврата T из локализованного возмущения намного больше, чем время возврата из глобального возмущения. Параметры возмущения: с = 0,1, при этом σ = 0,1 для локализованного возмущения и σ = 1 для глобального.
Реакция на нарушение может принимать две возможные формы: изолированное восстановление из-за местных процессов и аварийное восстановление из-за поступления биомассы из-за пределов нарушенного региона. Изолированное восстановление доминирует над реакцией системы, когда каждый сайт восстанавливается без помощи соседних сайтов (рис. 1, правые панели).Напротив, аварийное восстановление происходит, когда нарушенная область не может восстановиться без остальной системы, или когда основная часть восстановления происходит из-за такой пространственной динамики (Рисунок 1, левые панели).
Связь локальной динамики и рассеивания приводит к различным процессам восстановления в трех моделях, как показано траекториями на диаграммах фазового пространства в среднем столбце рисунка 2. На этих панелях мы разворачиваем восстановление в двух измерениях: по горизонтальной оси обозначает размер нарушенной области в данный момент времени, а вертикальная ось показывает плотность биомассы в нарушенной области.Сразу после возмущения система движется вдоль пунктирной черной кривой, а затем изменяется с течением времени, пока не войдет в заштрихованную область, где считается, что она восстановилась.
Рисунок 2 . Вклад изолированного и аварийного восстановления в зависимости от пространственной протяженности возмущения для трех моделей, представленных в основном тексте. В левом столбце показаны локальные потенциалы, определяющие локальные процессы. Верхний ряд: модель Logistic (LG); Средний ряд: сильно нелинейная модель SR; Нижний ряд: бистабильная модель АЭ.Средний столбец: изолированное восстановление по оси y и аварийное восстановление по оси x. Черная пунктирная линия показывает одинаковую используемую силу возмущения с = 0,2 для разной степени возмущения σ. Синие линии — это траектории восстановления, где восстановление считается завершенным, когда траектории достигают серой заштрихованной области. Для моделей SR и AE по мере того, как возмущения становятся более локализованными, наблюдается переход от доминирующего изолированного восстановления (восходящие траектории) к доминирующему аварийному восстановлению (траектории влево), влияя на время возврата (правый столбец).Знаки «x» синего цвета соответствуют различным траекториям, показанным в средних столбцах. Зеленые и пурпурные круги показывают начальные состояния после двух разных возмущений (левый и средний столбцы) и связанные с ними времена возврата (правый столбец). Пунктирная линия (нижний ряд) показывает локальный переломный момент бистабильной модели АЭ, за пределами которого локальная динамика коллапсирует до голого состояния.
Если большая часть траектории во время восстановления является горизонтальной, это означает, что нарушенная область сокращается из-за аварийного восстановления, что указывает на отсутствие локальной устойчивости, которая в противном случае позволила бы иметь место изолированному восстановлению.Такое поведение отражает сильную нелинейность локальной динамики, которую можно увидеть по изменяющейся кривизне локального потенциала (рис. 2, левый столбец). Мы можем видеть, что для модели AE (рисунок 2, нижний ряд) восстановление происходит вдоль горизонтальной линии для сценариев восстановления с достаточно малой пространственной протяженностью, так что региональные процессы вызывают восстановление. Напротив, восстановление полностью связано с локальными процессами в модели LG, поскольку локальная динамика здесь намного быстрее, в то время как для промежуточной модели SR можно видеть, что имеет место смесь двух процессов.
Эти различия выражаются в заметно разных значениях времени возврата T (Рисунок 2, правые панели). Траектории вертикального восстановления, которые следуют за всеми возмущениями в модели LG и возмущениями большого размера в других моделях, указывают на изолированное восстановление и, следовательно, на небольшие значения T . Для промежуточной модели SR локализованные возмущения приводят к большему вкладу спасательного восстановления, что приводит к сигмовидной форме T в зависимости от степени возмущения σ.Модель AE показывает аналогичное поведение более крупного T после локализованных возмущений, но тренд здесь показывает максимум для возмущений среднего размера. Это происходит потому, что в бистабильных системах наиболее эффективный способ нарушить работу системы — это локально удалить биомассу ниже порога жизнеспособности, а затем позволить системе локально разрушиться. Такое возмущение имеет эффект, эквивалентный более сильному возмущению, которое может удалить всю биомассу на большей территории. Процесс пространственного восстановления займет больше времени, что даст большие значения времени возврата (подробности см. В Приложении A).Это объясняет горбатую форму времени возврата в зависимости от степени возмущения. Поскольку бистабильность является достаточным условием возникновения горбовидной связи, последняя может использоваться как индикатор бистабильности. Это иллюстрирует более общую идею о том, что рассмотрение пространственного измерения возмущений может позволить нам исследовать локальную динамику пространственно протяженной экосистемы.
3.2. От изменчивости к коллапсу при увеличении частоты возмущений
Природные экосистемы постоянно нарушаются, поэтому мы должны учитывать временные рамки нарушений, а именно их среднюю частоту.Поэтому мы переводим результаты предыдущего раздела о реакции на одиночное возмущение (рис. 3, вверху) для понимания временной изменчивости при повторяющихся возмущениях (рис. 3, внизу). Фактически, существует прямая связь между реакцией на единичное нарушение и временной изменчивостью в ответ на повторяющиеся нарушения. Действительно, колебания биомассы являются результатом прошлых нарушений, поскольку они объединяют краткосрочные и долгосрочные реакции экосистемы на отдельные нарушения (Arnoldi et al., 2016). Изменчивость является статистикой этих колебаний и, следовательно, является функцией как интегрированной реакции на одиночное возмущение, так и средней частоты f возмущений. Точнее, если g ( t ) отслеживает изменение общей биомассы во времени после импульсного возмущения за время t = 0, то изменчивость V может быть выражена как V = f∫0∞g2 (t ) dt (см. уравнение S15 в Приложении B). Однако это тождество предполагает отсутствие взаимодействия в пространстве между различными возмущениями и, следовательно, не должно сохраняться при высокой частоте возмущений.
Рисунок 3 . Режимы одиночных и множественных возмущений и взаимосвязь между временем возврата и изменчивостью. На левой панели показаны временные ряды общей биомассы, а на правой панели — пространственные снимки соответствующих временных рядов (красные точки на левой панели). Реакция на одиночное нарушение отображается в верхней левой панели. Мы сосредоточимся на двух его характеристиках: время возврата T и интегральная мера переходного процесса g ( t ) (см. Основной текст).Реакция на множественные помехи, возникающие случайным образом на средней частоте f , показана на нижней левой панели. Резюмируется его изменчивость V (дисперсия общей биомассы). В пределе низкого значения f существует внутренняя взаимосвязь между временем возврата и изменчивостью в том смысле, что V может быть аппроксимировано величиной f∞0∞g2 (t) dt. Расчеты проводились с использованием модели SR со значениями параметров: с = 0,1, σ = 0,11, f = 0.025 и γ = 2. Случайный равномерный шум был добавлен в левые панели, чтобы продемонстрировать, как могут выглядеть реалистичные временные ряды.
Как и ожидалось, при низкой частоте возмущений аналитическое приближение достаточно хорошо согласуется с численным моделированием для всех трех моделей (рис. 4, второй столбец). Для более высоких частот (рис. 4, третий столбец), когда несколько возмущений часто имеют место в одном временном интервале, мы видим небольшую недооценку аналитического приближения, хотя общая тенденция хорошо прослеживается.Важно отметить, что изменчивость и время возврата показывают одно и то же поведение. Мы видим влияние региональных процессов на изменчивость для более локализованных возмущений как в модели SR, так и в модели AE, где первая имеет сигмовидную форму, а вторая — форму горба, что является следствием бистабильности в модели AE. Мы отмечаем, что эти тенденции сохраняются в более общих сценариях, таких как возмущения со случайной протяженностью или следование сезонным моделям (Приложение D).
Рисунок 4 .Время возврата, изменчивость и вероятность обрушения как функция пространственной протяженности возмущения для трех моделей (сверху вниз: LG, SR, AE). Левый столбец показывает время возврата (как на рисунке 2), в то время как средние столбцы показывают изменчивость при низкой и высокой частоте возмущений, а правый столбец показывает вероятность коллапса. Черная пунктирная (сплошная) линия представляет собой численное (аналитическое предсказание) значение изменчивости, серой штриховкой отмечена оценка ошибки. Отклонение от этого прогноза предполагает некоторую степень взаимодействия между возмущениями.Время возврата и изменчивость качественно аналогичны с низкой зависимостью пространственной протяженности возмущения для модели LG, но гораздо более сильной зависимостью, когда локальная динамика сильно нелинейна (модели SR и AE). В случае бистабильной модели АЭ мы распознаем немонотонную «горбовидную» зависимость от степени возмущения, при этом возмущения среднего размера вызывают наиболее серьезную реакцию. Параметры возмущения составляли с = 0,1, σ = 0,1, а для низкой частоты: f = 0,002, а для высокой частоты: f = 0.02.
На этом этапе может показаться, что временное измерение возмущений f не так информативно для поведения экосистемы, как пространственное измерение возмущений σ. Однако по мере дальнейшего увеличения f расхождение между изменчивостью и ее предсказанием, основанным на восстановлении после единственного возмущения, начинает расти. Это сигнализирует о том, что возмущения начинают взаимодействовать друг с другом — явление, которое не улавливается нашим приближением. Нарушения начинают накапливаться в космосе, что может существенно увеличить изменчивость (Приложение C) из-за больших отклонений в сторону низких уровней общей биомассы.Для бистабильных систем, таких как модель AE, такие отклонения могут привести к коллапсу всей системы. Это очевидно в двух последних столбцах рисунка 4, где мы видим для модели AE, что значения σ, для которых несоответствие изменчивости является наибольшим, точно соответствуют значениям σ, для которых вероятность коллапса наиболее значительна. . Таким образом, на высоких частотах возмущения одинаковой силы, но разной пространственной протяженности приводят к совершенно разным ответам. Этот пример подчеркивает тот факт, что сочетание пространственных и временных параметров возмущений может иметь решающее влияние на стабильность экосистемы.
4. Обсуждение
Исследуя роль пространственных и временных измерений нарушений в стабильности экосистемы, мы получили четыре основных результата: (1) По сравнению с глобальным нарушением, локализованное нарушение такой же силы может инициировать фундаментально иную и гораздо более медленную экосистему. отклик, особенно когда локальная динамика нелинейна. (2) Время возврата от одиночного возмущения и временная изменчивость, вызванная повторяющимися возмущениями, демонстрируют одинаковые тенденции, даже для локально интенсивных (и, следовательно, нелинейных) возмущений.(3) Связь между реакцией системы и пространственной протяженностью возмущений, которые она испытывает, выявляет лежащую в ее основе динамику. Например, горбовидная зависимость между временем возврата и пространственной протяженностью возмущений может указывать на бистабильность. (4) Соответствие между временем возврата и изменчивостью нарушается для высоких частот возмущений. Это несоответствие сигнализирует о возникновении пространственных взаимодействий между возмущенными областями, что, в свою очередь, может привести к смене режима.
Хотя мы рассматривали простые пространственно однородные модели, наши результаты должны применяться к широкому кругу экосистем. Леса, саванны и кустарники могут быть хорошими примерами экосистем, к которым применимы наши модели, поскольку нарушения, такие как пожары и выпас скота, происходят часто и часто локализованы, а восстановление растительных сообществ часто следует за сложной динамикой сукцессии, управляемой пространственными процессами (Adler et al. , 2001; Тернер, 2010; Ставер, Левин, 2012). Наши результаты, однако, не должны ограничиваться такими пространственно однородными системами.Хотя мы построили нашу теорию, используя пространственно однородные модели, эта упрощающая особенность не является существенной для наших аргументов, которые требуют только понятия локальности. Следовательно, наша теория может иметь отношение и к менее однородным экосистемам, таким как сети горных озер, коралловые рифы и речные системы. Действительно, такие экосистемы подвергаются различным нарушениям, которые часто сильно локализованы, и их динамика может быть достаточно нелинейной (Knowlton, 1992; Campbell Grant et al., 2007; Forrest and Arnott, 2007).
В рамках нашей работы мы рассматривали системы, которые локально отодвинуты от своего равновесия и даже попали в другую область притяжения. В контексте морской экосистемы это могут быть коралловые рифы (Nyström et al., 2000; Adjeroud et al., 2009) или скалистые приливные системы (Sousa, 1979; Paine and Levin, 1981), которые часто подвергаются интенсивным нарушениям (например, урон от шторма). Эти локально интенсивные нарушения могут позволить восстановлению, опосредованному расселением, доминировать над реакцией экосистемы.В случае бистабильной (AE) модели этот проблеск за пределы области притяжения населенного государства является прямой причиной горбовидных тенденций изменчивости и времени возврата в зависимости от степени возмущения. Фактически, переднее распространение, которое приводит к спасению, содержит информацию о бассейнах аттракционов экосистемы, отражающую существование альтернативных стабильных состояний и переходную динамику между ними. Таким образом, наблюдая за реакцией экосистемы на локальные нарушения, восстановление после спасения позволяет нам исследовать динамику экосистемы, далекую от равновесия.Например, сравнение различных нарушенных морских экосистем может дать дополнительные доказательства того, что некоторые из них имеют альтернативные состояния (например, коралловые рифы), в то время как для других динамика показывает процесс сукцессии (например, каменистые приливные системы). Это рассуждение можно продолжить, сосредоточив внимание на регионах, где происходит спасательное восстановление, например, анализируя структуру растительного сообщества в переходных зонах между пастбищами и лесами в экосистеме саванны (Augustine, 2003).
Расширение пространственного измерения возмущений может, таким образом, позволить нам обнаруживать нелинейности в поведении экосистемы, проявляющиеся в увеличивающейся локальной интенсивности возмущений (см. Рисунок 2).Можно ожидать, что по временному измерению возмущений увеличение их средней частоты также может выявить нелинейные эффекты, поскольку экосистема становится более нарушенной. Фактически, увеличение частоты имеет лишь тривиальный линейный эффект, о чем свидетельствует обнаруженная нами связь между временем возврата и изменчивостью (см. Рисунок 3). Однако за пределами некоторого порога возникает реакция другого типа из-за пространственного взаимодействия между нарушенными областями, которые собираются в потенциально крупномасштабные кластеры.Это вызывает более высокую изменчивость, чем ожидалось, и, следовательно, может вызвать глобальную потерю устойчивости или смену режима. Взяв еще раз пример с коралловыми рифами, мы могли бы спросить, как воздействие как естественных, так и антропогенных нарушений приводит к фазовому сдвигу от доминирования твердых кораллов к преобладанию мясистых водорослей. Сдвиг режима из-за скопления невосстановленных областей будет происходить не как типичный переломный момент из-за потери устойчивости (например, из-за изменения температуры), а скорее из-за пересечения порога частоты возмущений.Важно отметить, что в таком сценарии два измерения, пространственное и временное, должны рассматриваться во взаимосвязи. Порог, за которым происходит агрегация, сильно зависит от пространственной протяженности возмущений, и, следовательно, соответствующий ответ не является простым суперпозицией ответов на одиночные возмущения. Другими словами, это открытие подчеркивает и объясняет, как взаимодействие между пространственными и временными измерениями нарушений может иметь серьезные экологические последствия, такие как потеря устойчивости.Поскольку наши результаты являются чисто теоретическими, было бы полезно выяснить преобладание этого взаимодействия в эмпирических системах, которые претерпели смену режима (например, фазовые сдвиги в коралловых рифах Nyström et al., 2000 или опустынивание некогда зеленой Сахары Ортис). и др., 2000).
Как упоминалось ранее, в бистабильных системах взаимосвязь между временем возврата (а также изменчивостью) и пространственной протяженностью возмущений имеет форму горба. Это соотношение можно использовать в качестве индикатора бистабильности, оцениваемого эмпирически путем сравнения временных рядов одной и той же экосистемы в разных регионах с оценками интенсивности отдельных нарушений.Его последствия для управления экосистемой зависят от типа рассматриваемых нарушений. Антропогенные нарушения, которые в значительной степени контролируются, такие как вырубка леса (Chazdon, 2003) или крупномасштабный рыбный промысел (Kaiser et al., 2006), можно лучше спланировать, чтобы избежать как непредсказуемого вылова из-за высокой изменчивости, так и общего коллапса. Для многих естественных нарушений контроль невозможен и нежелателен (например, пожары в полузасушливых экосистемах, необходимые для прорастания растений, Wellington and Noble, 1985), но прогнозирование их последствий и возможности смены режима имеет первостепенное значение (Kéfi et al., 2007).
Чтобы сосредоточить внимание на роли пространственных свойств возмущений и дать более ясное представление, мы провели наш анализ, предполагая возмущения постоянной общей силы. Нетрудно расширить анализ до более общих параметров, таких как случайная степень возмущений и сезонные закономерности (подробности см. В Приложении D). Особенно интересно рассмотреть случай различных значений мощности возмущения с . Как показано на рисунке 5, если мы случайным образом выберем набор точек с разными значениями силы s и протяженности σ, мы сможем использовать их для восстановления нормализованной версии зависимости различных мер устойчивости от степени возмущения.Таким образом, мы можем использовать различные явления, описанные ранее, такие как соотношение формы горба в качестве индикатора бистабильности, в более общих условиях, тем самым делая нашу теорию более эмпирически доступной.
Рисунок 5 . Реконструкция кривой зависимости времени возврата от степени возмущения из более общего пространства параметров свойств возмущения. Верхние (нижние) панели соответствуют модели SR (AE). Левый столбец показывает время возврата в пространстве параметров степени возмущения σ (ось x) и силы возмущения с (ось y).Правый столбец показывает соответствующую реконструкцию кривой времени возврата с использованием 100 случайно выбранных точек (красные звездочки) в пространстве параметров. Значения времени возврата нормированы на мощность возмущения с , в то время как нормализованная степень возмущения определяется как σ ~ = 1-с / σ. Обратите внимание, что форма горба (сигмовидной) кривой для модели AE (SR) легко распознается по этим реконструкциям.
Наша работа — это шаг к количественному учету пространственных и временных размеров нарушений и их взаимодействия с местной и региональной динамикой экосистем.Это важная цель в контексте глобальных изменений. Нарушения происходят все чаще и в разных масштабах (что очевидно, например, в коралловых рифах Jackson, 1991 и лесах Turner et al., 1993), в то время как пространственная структура самих экосистем изменяется из-за изменений в землепользовании, часто вызывая фрагментацию. ландшафта (Харрисон и Бруна, 1999). Таким образом, важно создать структуру, в которой мы сможем понять и предсказать экологические последствия этого сложного взаимодействия.
Заявление о доступности данныхРезультаты, показанные в этом исследовании, основаны на коде Matlab, который можно найти в репозитории Github по адресу https://github.com/yzelnik/stddes-scripts.
Авторские взносы
YZ, J-FA и ML разработали исследование. YZ и J-FA провели исследование. YZ, J-FA и ML написали рукопись.
Финансирование
Эта работа была поддержана лабораторией передового опыта TULIP (ANR-10-LABX-41) и расширенным грантом BIOSTASES, финансируемым Европейским исследовательским советом в рамках программы исследований и инноваций Европейского Союза Horizon 2020 (соглашение о гранте № 666971).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы хотим поблагодарить Барта Хэгемана и Матье Барбье за полезные обсуждения и комментарии к предыдущей версии рукописи. Мы также хотели бы поблагодарить двух рецензентов за их полезные комментарии к рукописи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fevo.2018.00224/full#supplementary-material
Список литературы
Аджеро, М., Мишонно, Ф., Эдмундс, П. Дж., Шансерель, Ю., де Лома, Т. Л., Пенин, Л. и др. (2009). Периодические нарушения, траектории восстановления и устойчивость коралловых сообществ на рифе южной части центральной части Тихого океана. Коралловые рифы 28, 775–780. DOI: 10.1007 / s00338-009-0515-7
CrossRef Полный текст | Google Scholar
Арнольди, Ж.-Ф., Лоро, М., и Хэгеман, Б. (2016). Устойчивость, реактивность и изменчивость: математическое сравнение показателей экологической устойчивости. J. Theor. Биол. 389, 47–59. DOI: 10.1016 / j.jtbi.2015.10.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Августин, Д. Дж. (2003). Пространственная неоднородность травяного яруса полузасушливой экосистемы саванны. Завод Ecol. 167, 319–332. DOI: 10.1023 / A: 1023927512590
CrossRef Полный текст | Google Scholar
Баччелли, Ф., и Блащишин, Б. (2010). Стохастическая геометрия и беспроводные сети: Том I Теория. Найдено. Тенденции. Сеть. , 3, 249–449. DOI: 10.1561 / 1300000006
CrossRef Полный текст
Кэмпбелл Грант, Э. Х., Лоу, У. Х. и Фаган, У. Ф. (2007). Жизнь в ветвях: динамика популяций и экологические процессы в дендритных сетях. Ecol. Lett. 10, 165–175. DOI: 10.1111 / j.1461-0248.2006.01007.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаздон, Р.Л. (2003). Восстановление тропических лесов: наследие антропогенного воздействия и стихийных бедствий. Перспектива. Завод Ecol. Evol. Syst. 6, 51–71. DOI: 10.1078 / 1433-8319-00042
CrossRef Полный текст | Google Scholar
Дарретт Р. и Левин С. (1994). Важность дискретности (и пространственности). Теор. Popul. Биол. 46, 363–394. DOI: 10.1006 / TPBI.1994.1032
CrossRef Полный текст | Google Scholar
Форрест Дж. И Арнотт С. Э. (2007).Изменчивость и предсказуемость в сообществе зоопланктона: роль нарушения и распространения. Ecoscience 14, 137–145. DOI: 10.2980 / 1195-6860 (2007) 14 [137: ВАПИАЗ] 2.0.CO; 2
CrossRef Полный текст | Google Scholar
Фостер Д. Р., Найт Д. Х. и Франклин Дж. Ф. (1998). Ландшафтные узоры и наследие, возникшие в результате редких крупных нарушений леса. Экосистемы 1, 497–510. DOI: 10.1007 / s100219
6
CrossRef Полный текст | Google Scholar
Фокс, Дж.W., Vasseur, D., Cotroneo, M., Guan, L., and Simon, F. (2017). Вымирание популяций может увеличить устойчивость метапопуляции. Nat. Ecol. Evol. 1, 1271–1278. DOI: 10.1038 / s41559-017-0271-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фратерриго, Дж. М., и Русак, Дж. А. (2008). Изменения в изменчивости экологических моделей и процессов, вызванные нарушениями. Ecol. Lett. 11, 756–770. DOI: 10.1111 / j.1461-0248.2008.01191.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холл, С.А. (1988). Оценка нескольких исторически наиболее влиятельных теоретических моделей, используемых в экологии, и данных, представленных в их поддержку. Ecol. Модель. 43, 5–31. DOI: 10.1016 / 0304-3800 (88) -1
CrossRef Полный текст | Google Scholar
Харрисон, С., и Бруна, Э. (1999). Фрагментация среды обитания и широкомасштабное сохранение: что мы знаем наверняка? Экография 22, 225–232. DOI: 10.1111 / j.1600-0587.1999.tb00496.x
CrossRef Полный текст | Google Scholar
Холлинг, К.С. (1973). Устойчивость и устойчивость экологических систем. Annu. Rev. Ecol. Syst. 4, 1–23. DOI: 10.1146 / annurev.es.04.110173.000245
CrossRef Полный текст | Google Scholar
Джексон, Дж. Б. (1991). Адаптация и разнообразие рифовых кораллов. BioScience 475–482.
Google Scholar
Кайзер, М., Кларк, К., Хинц, Х., Остин, М., Сомерфилд, П., и Каракассис, И. (2006). Глобальный анализ реакции и восстановления бентической биоты на промысел. Mar. Ecol. Прог. Сер. 311, 1–14. DOI: 10.3354 / meps311001
CrossRef Полный текст | Google Scholar
Kéfi, S., Rietkerk, M., Alados, C.L., Pueyo, Y., Papanastasis, V.P., ElAich, A., et al. (2007). Структура пространственной растительности и неизбежное опустынивание в средиземноморских засушливых экосистемах. Природа 449, 213–217. DOI: 10.1038 / nature06111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ноултон, Н. (1992). Пороги и множественные стабильные состояния в динамике сообщества коралловых рифов. г. Zool. 32, 674–682. DOI: 10.1093 / icb / 32.6.674
CrossRef Полный текст | Google Scholar
Крамер А. М., Деннис Б., Либхольд А. М. и Дрейк Дж. М. (2009). Доказательства эффектов Аллее. Население. Ecol. 51, 341–354. DOI: 10.1007 / s10144-009-0152-6
CrossRef Полный текст | Google Scholar
Лейболд, М.А., Холиоак, М., Муке, Н., Амарасекаре, П., Чейз, Дж. М., Хупес, М. Ф. и др. (2004). Концепция метасообщества: основа для многомасштабной экологии сообщества. Ecol. Lett. 7, 601–613. DOI: 10.1111 / j.1461-0248.2004.00608.x
CrossRef Полный текст | Google Scholar
Левин, С. А. (1992). Проблема структуры и масштаба в экологии: лекция премии Роберта Макартура. Экология 73, 1943–1967. DOI: 10.2307 / 1941447
CrossRef Полный текст | Google Scholar
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J., Hector, A., et al. (2001). Биоразнообразие и функционирование экосистем: текущие знания и будущие задачи. Наука 294, 804–808. DOI: 10.1126 / science.1064088
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маркетти К., Мейер П. С. и Осубель Дж. Х. (1996). Возвращение к динамике населения с помощью логистической модели: что можно смоделировать и спрогнозировать? Technol. Прогноз. Soc. Изменить 52, 1–30. DOI: 10.1016 / 0040-1625 (96) 00001-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мэй, Р. М. (1973). Стабильность и сложность модельных экосистем .Номер 6 в Монографиях по популяционной биологии. Принстон, Нью-Джерси: Издательство Принстонского университета.
PubMed Аннотация | Google Scholar
Макканн, К. С., Расмуссен, Дж. Б., и Умбанховар, Дж. (2005). Динамика пространственно связанных пищевых сетей: пространственно связанных пищевых сетей. Ecol. Lett. 8, 513–523. DOI: 10.1111 / j.1461-0248.2005.00742.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерон, Э. (2015). Нелинейная физика экосистем .Бока-Ратон, Флорида: CRC Press.
Молони К.А., Левин С.А. (1996). Влияние архитектуры нарушений на динамику населения на ландшафтном уровне. Экология 77, 375–394. DOI: 10.2307 / 2265616
CrossRef Полный текст | Google Scholar
Neubert, M. G., and Caswell, H. (1997). Альтернативы устойчивости для измерения реакции экологических систем на возмущения. Экология 78, 653–665. DOI: 10.1890 / 0012-9658 (1997) 078 [0653: ATRFMT] 2.0.CO; 2
CrossRef Полный текст | Google Scholar
Нюстрём М., Фольке К. и Моберг Ф. (2000). Нарушение коралловых рифов и устойчивость в среде, где доминирует человек. Trends Ecol. Evol. 15, 413–417. DOI: 10.1016 / S0169-5347 (00) 01948-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ортис, Дж., Гильдерсон, Т., Адкинс, Дж., Сарнтейн, М., Бейкер, Л., Ярусинский, М., и др. (2000). Внезапное начало и окончание влажного периода в Африке: быстрая реакция климата на постепенное воздействие инсоляции. Quat. Sci. Ред. 19, 347–361. DOI: 10.1016 / S0277-3791 (99) 00081-5
CrossRef Полный текст | Google Scholar
Пейн Р. Т. и Левин С. А. (1981). Приливные пейзажи: нарушение и динамика рисунка. Ecol. Monogr. 51:35. DOI: 10.2307 / 2937261
CrossRef Полный текст | Google Scholar
Петерсон, Г., Аллен, К. Р., и Холлинг, К. С. (1998). Экологическая устойчивость, биоразнообразие и масштаб. Экосистемы 1, 6–18.DOI: 10.1007 / s100219
2
CrossRef Полный текст | Google Scholar
Петерсон, Г. Д. (2000). Масштабирование экологической динамики: самоорганизация, иерархическая структура и экологическая устойчивость. Клим. Чанг. 44, 291–309. DOI: 10.1023 / A: 1005502718799
CrossRef Полный текст | Google Scholar
Шеффер, М. (2009). Критические переходы в природе и обществе . Принстон учится по сложности. Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Соуза, В. П. (1979). Нарушение морских приливных валунов: неравновесное поддержание видового разнообразия. Экология 60: 1225. DOI: 10.2307 / 1936969
CrossRef Полный текст | Google Scholar
Тилман Д., Леман К. Л. и Бристоу К. Э. (1998). Отношения между разнообразием и стабильностью: статистическая неизбежность или экологические последствия? г. Nat. 151, 277–282.
Google Scholar
Тернер, М.Г. (1989). Ландшафтная экология: влияние рисунка на процесс. Annu. Rev. Ecol. Syst. 20, 171–197. DOI: 10.1146 / annurev.es.20.110189.001131
CrossRef Полный текст | Google Scholar
Тернер, М. Г., Ромм, В. Х., Гарднер, Р. Х., О’Нил, Р. В., и Кратц, Т. К. (1993). Пересмотренная концепция ландшафтного равновесия: нарушение и стабильность масштабных ландшафтов. Landsc. Ecol. 8, 213–227. DOI: 10.1007 / BF00125352
CrossRef Полный текст | Google Scholar
Уль, К.(1987). Факторы, влияющие на наследование после подсечно-огневого земледелия в Амазонии. J. Ecol. 75: 377. DOI: 10.2307 / 2260425
CrossRef Полный текст | Google Scholar
Ван С., Лоро М., Арнольди Ж.-Ф., Фанг Дж., Рахман К. А. и Тао С. (2017). Взаимосвязь неизменности и площади проливает новый свет на пространственное масштабирование экологической стабильности. Nat. Commun. 8: 15211. DOI: 10.1038 / ncomms15211
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Веллингтон, А.Б. и Нобл И. Р. (1985). Постпожарный набор и смертность в популяции Mallee Eucalyptus Incrassata в полузасушливых районах Юго-Восточной Австралии. J. Ecol. 73: 645. DOI: 10.2307 / 2260501
CrossRef Полный текст | Google Scholar
Ячи С. и Лоро М. (1999). Биоразнообразие и продуктивность экосистем в изменчивой среде: страховая гипотеза. Proc. Natl. Акад. Sci. США, 96, 1463–1468. DOI: 10.1073 / pnas.96.4.1463
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пространственная динамика и функционирование экосистем
Образец цитирования: Schmitz OJ (2010) Пространственная динамика и функционирование экосистем.PLoS Biol 8 (5): e1000378. https://doi.org/10.1371/journal.pbio.1000378
Опубликовано: 25 мая 2010 г.
Авторские права: © 2010 Oswald J. Schmitz. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Специального финансирования по этой статье получено не было.
Конкурирующие интересы: Автор заявил, что конкурирующих интересов не существует.
Классическая теория динамики видов в экосистемах построена на концепции однородного взаимного взаимодействия. Эта концепция заимствована из раздела физики и химии, изучающего кинетику реакций молекул в хорошо перемешанных газах и жидкостях. Он идеализирует отдельные сущности — уже не молекулы, а теперь особей вида — как взаимодействующие друг с другом, со своими хищниками или конкурентами таким образом, что каждый индивидуум имеет равную вероятность взаимодействия с каждым другим индивидуумом в системе.В системе нет пространственной структуры; фактически считается, что пространство не имеет значения для системной динамики.
Но любой чуткий наблюдатель природы может заплакать. В отличие от упрощенной теоретической концепции природные экосистемы характеризуются сложной и неоднородной пространственной структурой. Растения собраны в группы. Соответственно, травоядные животные, поедающие их, и хищники, поедающие травоядных, располагаются в пространстве одинаково [1]. Это наблюдение не было упущено экологическими теоретиками, которые в 1980-х и 1990-х годах начали более подробно рассматривать пространственную неоднородность [1], [2].Эта новая экологическая теория, по существу построенная на дополнительных концепциях физики и химии (например, [3]), эффективно разделяет динамику системы на две фазы: фазу реакции, в которой особи видов взаимодействуют локально, и фазу диффузии, в которой особи расходятся после происходят локальные взаимодействия. Распространение активируется (положительная обратная связь) в фазе реакции такими факторами, как интенсивная конкуренция или риск нападения хищников, которые заставляют людей перемещаться в менее конкурентные или более безопасные места.Распространение становится сдерживаемым (отрицательная обратная связь) всякий раз, когда попытки людей переехать получают отпор со стороны людей, уже занимающих новые места.
Этот основной механизм реакции-диффузии был использован для разработки двух различных классов теории динамики видов и экосистем. Теории фундаментально различаются в предположениях о пространственной структуре и в том, как активирующие и тормозящие обратные связи действуют на ландшафт. Один из видов теории (известный теперь как теория метапопуляций и метасообществ) расширяет классическую теорию, налагая пространственную структуру пятен как физическое состояние системы (рис. 1), и переделывает параметры, описывающие процессы рождения и смерти населения с точки зрения пространственного двигательные процессы [4].Затем он исследует последствия этой структуры для динамики системы посредством анализа взаимодействий внутри участков и распределения видов между участками [4]. Теория другого типа (известная сейчас как теория самоорганизованных систем) начинается с чистого листа и исследует, как пространственная структура возникает как следствие взаимодействий и движений видов [5]. В теории метасистем диффузия по ландшафту сдерживается локальной отрицательной обратной связью внутри участка (рис. 1). То есть положительная и отрицательная обратная связь действуют внутри локальных патчей [4].В теории самоорганизованных систем положительная обратная связь является локальной, а отрицательная обратная связь проявляется как напряжение между локальными скоплениями видов, которое предотвращает дальнейшее расселение (рис. 1). Таким образом, мозаичная структура ландшафта возникает в теории самоорганизованных систем как следствие локальной положительной обратной связи и тормозящей или отрицательной обратной связи в масштабе ландшафта [5].
Рисунок 1. Два альтернативных представления теории пространственной динамики экосистем.
Верхние рамки представляют механизмы, определяющие перемещение людей между отдельными участками ландшафта.В простейшей идеализации теории индивиды внутри дискретных участков принадлежат к местному населению. Местные популяции связаны друг с другом индивидуальным рассредоточением (диффузией), которое активируется взаимодействиями (реакцией) между людьми в пределах участка. Колонизация другого участка отдельными особями тормозится всякий раз, когда плотные местные популяции противодействуют рассредоточенным особям. Рассеивание связывает динамику местного населения, создавая грандиозную «метапопуляцию». Нижние рамки представляют механику движения людей из локальной концентрации вовне (диффузия), снова активируемую локальным взаимодействием между людьми.В простейшей идеализации теории полное случайное рассредоточение индивидов запрещается всякий раз, когда индивиды сталкиваются и взаимодействуют с представителями других локальных скоплений в пределах общих буферных зон. Это заставляет регулярно расположенные группы людей самоорганизовываться в ландшафте.
https://doi.org/10.1371/journal.pbio.1000378.g001
Теория метасистем получила значительное распространение в экологии, поскольку она перекликается с нашим интуитивным пониманием текущего состояния многих экосистем [6], [7] .Например, небольшие пруды представляют собой естественные дискретные участки в наземных ландшафтах, что приводит к характерным моделям изобилия видов в местном и ландшафтном масштабе и функционирования экосистем [8]. Человеческая деятельность также искусственно наложила пространственную структуру на многие экосистемы путем фрагментации ранее сплошных ландшафтов на отдельные участки среды обитания. Это привело к предсказуемой трансформации сообществ видов и связанного с ними функционирования из-за различных способностей видов жить на участках определенного размера и рассредоточиваться между ними [9].Теория метасистем имеет ясные и глубокие последствия для сохранения биоразнообразия [6], [7], [10].
Применимость теории самоорганизованных систем имеет тенденцию быть менее ясной, потому что это более абстрактная конструкция, чем теория метасистем. Более того, существует разделенное мнение относительно того, является ли предсказанная динамика эмерджентности, основанная на довольно простых математических правилах взаимодействия видов, устойчивой к изменениям в допущениях, отражающих реальные экологические условия [11].Эти дебаты, однако, по-прежнему носят в основном академический характер, поскольку окончательный арбитр — обширный массив эмпирических свидетельств явных проверок теории — еще не накоплен [5]. Несомненно, существует множество предполагаемых примеров самоорганизующихся крупномасштабных структур, во многом благодаря достижениям в области спутниковых изображений [5]. И были попытки разрешить механизмы, управляющие формированием самоорганизованных паттернов в популяциях видов [12], [13]. Но доказательства того, что такая пространственная организация на популяционном уровне влияет на функционирование всей экосистемы, остается недостающей частью головоломки.
Тестирование теории самоорганизованных систем в контексте всей экосистемы в природе не является делом для тех, кому дано проводить исследования, дающие быстрые и простые ответы. В отличие от теории метасистем не существует простого и быстрого способа очертить структуру системы. Границы участков самоорганизованных систем имеют тенденцию быть нечеткими [5], что требует сложных статистических методов для разрешения пространственного паттерна. Успех такого анализа зависит от получения обширного набора данных с высоким разрешением.Однако перед этим нужно решить, что такое патч и что управляет структурой патча. Например, возникает ли структура участков из пространственных градиентов концентраций питательных веществ в почве, которые затем вызывают пространственное скопление растений и наращивание пищевых цепей? Или же пространственная структура возникает в результате взаимодействий хищник-жертва, из-за которых виды уходят от локального точечного источника? Скорее всего, это комбинация двух, и поэтому их относительная важность должна быть решена посредством стратегических экспериментов и отбора проб биоты и физических условий.Наконец, необходимо найти точки пространственного напряжения и разрешить механизмы, очерчивающие границы участков. Сложность может вызывать недоумение, приводя к окончательному априорному вопросу: с чего начать?
В этом выпуске PLoS Biology , Pringle et al. [14] отвечают на эти вопросы, предпринимая титанические усилия по объяснению пространственного построения экосистемы африканской саванны. Прорыв в нашем понимании экологических систем часто происходит благодаря хорошему пониманию естественной истории рассматриваемой системы и вниманию к подсказкам, которые дает природа [15].Действительно, Pringle et al. [14] извлекают выгоду из важных предшествующих подсказок естественной истории о том, что термиты имеют тенденцию демонстрировать локально неперекрывающиеся кормовые территории вокруг своих колоний [16]. Движение термитов из колонии, по-видимому, активируется необходимостью найти пищу, и движение в конечном итоге тормозится, когда отдельные термиты сталкиваются и соревнуются с членами другой колонии [16]. Удивительно, но такое поведение может привести к довольно регулярному расположению колоний термитов по ландшафту [14].
Более важным для структуры и функционирования всей экосистемы саванны является то, что покрытые травой насыпи, созданные колониями термитов, более песчаные, чем окружающие почвы. Это обеспечивает большую инфильтрацию воды, аэрацию и накопление питательных веществ на насыпях по сравнению с окружающими почвами [14]. Курганы термитов представляют собой «оазисы» влаги и питательных веществ в матрице засушливых земель. Концентрированная влага и питательные вещества, соответственно, способствуют росту древесных пород на окраинах колонии, причем самые толстые деревья располагаются по периметру насыпи, а постепенно более тонкие деревья выходят за пределы периметра и переходят в тонкие деревья, выходящие из других холмов [14].Питательные вещества, поставляемые насыпями деревьям, также способствуют наращиванию пищевых цепочек, состоящих из насекомых-травоядных и хищников насекомых-пауков и ящериц.
Эта возникающая структура также ведет к параллельной динамике активации и ингибирования между видами в пищевой цепи [14]. Травоядные насекомые сконцентрированы на более толстых деревьях возле насыпей и уменьшаются на более тонких удаленных деревьях. Ящерицы и пауки также рассредоточены, и полевые эксперименты показали, что это отчасти объяснялось тем, что более толстые деревья предлагали лучшие места для охоты, а отчасти потому, что плотность добычи была самой высокой на толстых деревьях, которые, как правило, служили убежищами от холмов.Связанное с этим исследование [17] показывает, что содержание азота в растениях также выше вблизи курганов термитов, а это означает, что как количество, так и качество пищи выше возле курганов, что, вероятно, вносит свой вклад во все эти закономерности.
Сочетание поступления питательных веществ для первичного производства растений и преобразования питательных веществ для растений во вторичную продукцию травоядных и хищников означает, что термитники также становятся горячими точками продуктивности экосистемы. Эти горячие точки сохраняются благодаря взаимодействию между активацией и ингибированием пространственного движения всех компонентов экосистемы.Таким образом, на ландшафте виден регулярный рисунок высокой и низкой продуктивности, который отражает регулярный рисунок термитников. Дальнейшее статистическое моделирование предполагает, что эта форма неоднородности приводит к большей чистой продуктивности, чем можно было бы ожидать, если бы термитники были неравномерно сгруппированы по ландшафту [14]. Это происходит из того статистического свойства, что при регулярном расположении пятен ни одна точка не находится очень далеко от холма, поэтому продуктивность всех точек при усреднении выше, чем в случае, когда участки сильно сгруппированы или рассредоточены случайным образом [14].Конечно, было бы чрезвычайно трудно провести окончательную экспериментальную проверку этого утверждения, которая потребовала бы изменения пространственной конфигурации термитников. Это, пожалуй, самая большая ахиллесова пята любой эмпирической попытки проверить теорию самоорганизованных систем в реальной экосистеме. Тем не менее, исследование [14] является образцовым в том смысле, что оно ближе всего подходит к удовлетворению эмпирических условий, необходимых для демонстрации существования самоорганизованной экосистемы [5].Собирая данные о поведении животных, популяции животных и экосистемах, авторы, таким образом, создают достаточно последовательную картину пространственных механизмов, управляющих структурой и функционированием экосистемы.
Интересно то, что если вместо того, чтобы сосредоточиться на окрестностях курганов термитов, мы сделаем вид с высоты птичьего полета на пейзаж, нас можно обмануть, сделав вывод, что саванна — довольно однородный ландшафт. И действительно, такой взгляд на структуру системы вполне правдоподобен, учитывая все более широкое и широкое применение современных спутниковых изображений для изучения структур саванн и других экосистем [5], [18], [19].С другой стороны, если мы слишком внимательно сфокусируемся на термитнике и только его ближайшем окружении, наша точка зрения может быть настолько подавлена высокоразрешенными взаимодействиями местных видов, что мы рискуем вообще не увидеть пространственную структуру. Искусство эмпирического разрешения структуры и динамики самоорганизованных экосистем заключается в выборе подходящей шкалы разрешения для исследования [1], [5]. Это нетривиальное упражнение, потому что для этого требуются годы бесстрашных полевых исследований, направленных на понимание как естественной истории системы, так и измерения пространственных закономерностей и динамических процессов во многих различных, но дополняющих друг друга пространственных перспективах.Это вполне может быть единственной наиболее важной причиной, по которой еще нет полевых данных о самоорганизованных системах.
Исследование Pringle et al. [14] прекрасно показывает, что теория самоорганизующихся систем — это не просто феномен виртуального компьютерного мира. Действительно, есть основа в «надежной реальности» [11]. Как и в теории метасистем, значение теории самоорганизованных систем для сохранения, как показано в исследовании [14], является глубоким. В этом конкретном случае очень нехаризматичный вид культивирующих грибы термитов, который живет преимущественно под землей, кажется, создает биофизические и биотические условия, которые приводят к эволюции надземной трофической структуры и параллельной самоорганизованной динамике на высших трофических уровнях.Это, в свою очередь, предполагает, что потеря любой из частей приведет к быстрому коллапсу параллельной динамики, поддерживающей общее функционирование экосистемы. Это усиливает необходимость рассмотрения того, как характер взаимодействия видов связан с функционированием всей экосистемы при разработке стратегий сохранения биоразнообразия [7], [15], [20].