
Сократительный аппарат мышечного волокна – 31. Сократительный аппарат мышечного волокна – строение. Способность к регенерации различных мышечных тканей.
31. Сократительный аппарат мышечного волокна – строение. Способность к регенерации различных мышечных тканей.
Сократительным аппаратом скелетного мышечного волокна являются продольно ориентированные миофибриллы. Построенные из сократительных белков, они занимают большую часть волокна, оттесняя ядра на периферию. Диаметр миофибрилл около 1 – 2 мкм. Миофибриллы состоят из чередующихся темных и светлых полос (дисков). Структурной единицей миофибриллы является саркомер. В миофибрилле они расположены, следуя друг за другом. Саркомер это участок миофибриллы, состоящий из линии Z (для двух соседних саркомеров), половины диска I, диска А с линией Н, половины следующего диска I, линии Z (для двух соседних саркомеров). Диски саркомер состоят из миофиламентов (белок миозин – толстые миофилламенты; актин, тропомиозин В, тропин – тонкие миофилламенты. Участок А диска, состоящий только из миозиновых миофиламентов, называют Н линией. Линия Z состоит из Z-филаментов. В них выявлены белки тропомиозин-В, а-актин. Z-филаменты формируют решетку, к котрой прикреплены тонкие актиновые филаменты полосок I двух соседних саркомеров.
Сократительный аппарат в сердечной мышце так же, как в скелетной мышечной ткани, состоит из миофибрилл, которые занимают периферическую часть клетки. По своему строению миофибриллы сходны с таковыми скелетной мышечной ткани. Они также построены из анизотропных (полосы А) и изотропных дисков (полосы I). Элементы саркоплазматической сети окружают миофибриллы. Характерное свойство сердечных миоцитов – отсутствие терминальных цистерн, а поэтому триад.
Сократительным аппаратом миоцита являются миофилламенты. Они расположены в цитоплазме на периферии клетки и имеют продольную ориентацию. Миофилламенты: миозиновые (толстые), актиновые (тонкие) и промежуточные.
Процесс регенерации, протекающий в скелетной мышечной ткани, сходен с гистогенезом; в нем выявлены те же миобластическая стадия, стадия мышечных трубочек и стадия мышечных волокон. Регенерация скелетной мышечной ткани
Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами.
Восстановление любых тканей организма может осуществляется за счет двух механизмов: гипертрофии и гиперплазии. Под гипертрофией подразумевают компенсаторное увеличение объема самого симпласта, в т.ч. за счет увеличения количества миофибрилл. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани.
Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается (рабочая гипертрофия клеток). Не исключена, однако, и пролиферация клеток (т.е. гиперплазия).
studfiles.net
Сократительным аппаратом мышечной ткани является
Сократительный аппарат скелетного и сердечного мышечных волокон
Ø тонких (актиновых)
Ø толстых (миозиновых)
Тонкие или актиновые филаменты состоят из трех типов белков, расположенных определенным образом относительно друг друга (рис. 8):
Ø G-актина (глобулярный белок), который полимеризуется с образованием фибриллярного (нитчатого) актина (F-актина), имеющего вид спирали с желобком. В составе каждой актинового филамента имеются две спирали F-актина, в желобках которых, располагаются:
Ø молекулы нитчатого белка тропомиозина– белка, в составе которого две субъединицы, переплетающиеся между собой в α-спиралевидные фибриллярные структуры. Тропомиозин связывается в единый комплекс с F-актином в области изгиба молекулы, обеспечивая его стабильность. По длине тропомиозин равен 7 субъединицам G-актина, при этом контактирует только с одной из нитевидных структур F-актина. Кроме этого, тропомиозин совместно с тропонином участвует в регуляции взаимодействия актина с миозином,
Ø молекулы глобулярного тропонина,состоящего из трех белков:
ü тропонина C (мол. м. 20 тыс.) связывает ионы Ca2+
ü тропонина I (мол. м. 37 тыс.) связывается с тропомиозином, образуя с ним тропонин-тропомиозиновый комплекс,
ü тропонина T (мол. м. 25 тыс.) связывается с актином в тонких филаментах, удерживая таким образом тропонин-тропомиозиновый комплекс на месте, препятствует взаимодействию актина и миозина, когда ионы кальция не связаны с тропонином C.
Толстые или миозиновые филаменты (диаметр – 10нм, длина – 1,6мкм) образованы преимущественно
В строении молекулы миозина выделяют (рис. 10):
Ø 2 головки (в образовании каждой принимают участие 2 субъединицы), обладающие способностью расщеплять молекулу АТФ, т.е. обладают АТФазной активностью;
Ø шейку(образована 1 субъединицей)
Ø собственно хвост(образован 1 субъединицей) – концевая часть нитчатых субъединиц миозиновой молекулы.
В области присоединения головки миозиновой молекулы к шейке и шейки – к хвосту имеются по шарнирному участку (всего в молекуле их получается два), позволяющих миозиновой молекуле изменять свою конформацию (пространственную структуру).
Тонкие (актиновые) и толстые (миозиновые) миофиламенты уложены друг относительно друга в составе миофибриллы, таким образом, что образуются регулярно чередующиеся по длине миофибриллы темные (анизотропные, А-диски)
Светлые I-диски состоят только из параллельно уложенных актиновых филаментов. В центре каждого светлого диска проходит Z-мембрана – узкая полоска белковой природы, к которой крепятся одним своим концом актиновые нити; другой конец актиновых филаментов направлен в пространства между миозиновыми нитями.
Темные А-диски состоят из параллельно уложенных миозиновых филаментов, между которыми располагаются участки актиновых филаментов. В центре темных дисков находится М-линия – нить белковой природы, которая объединяет центральные части миозиновых нитей друг с другом.
В середине темных дисков, располагается зона, состоящая только из миозиновых нитей, которая носит название H-зоны. При сокращении мышечного волокна изменяется ширина этой зоны.
Структурно-функциональной единицей миофибриллы является саркомер – участок миофибриллы, расположенный между двумя соседними Z-мембранами. Т.е., саркомер состоит из:
½ I-диска + А-диска + ½ I-диска.
Все миофибриллы в составе волокна уложены таким образом, что темные диски одной находятся строго под темными дисками другой миофибриллы (рис. 12).
Каждая миофибрилла окружена цистернами СР таким образом, что место контакта Т-трубочки с боковыми цистернами СР у млекопитающих животных и человека соответствует границе темных и светлых дисков.
Дата публикования: 2014-12-30; Прочитано: 222 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.001 с)…
Мышечными тканями (лат. textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Мышечные ткани состоят из вытянутых клеток, которые принимают раздражение от нервной системы и отвечают на него сокращением. Они обеспечивают перемещения в пространстве организма в целом, его движение органов внутри организма (сердце, язык, кишечник и др.) и состоят из мышечных волокон. Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Основные морфологические признаки элементов мышечных тканей: удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы — миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина — при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин — белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко падает).
Свойства мышечной ткани:
- Возбудимость
- Проводимость
- Сократимость
- Лабильность
Виды мышечной ткани:
1. Гладкая мышечная ткань
Гладкая мышечная ткань состоит из одноядерных клеток — миоцитов веретеновидной формы длиной 20—500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Эта мышечная ткань обладает особыми свойствами: она медленно сокращается и расслабляется, обладает автоматией, является непроизвольной (то есть ее деятельность не управляется по воле человека). Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта (сокращение стенок желудка и кишечника).
2. Поперечно-полосатая скелетная мышечная ткань
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) и диаметр 50—100 мкм; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование тёмных и светлых полосок. Свойствами этой мышечной ткани является высокая скорость сокращения, расслабления и произвольность (то есть её деятельность управляется по воле человека). Эта мышечная ткань входит в состав скелетных мышц, а также стенки глотки, верхней части пищевода, ею образован язык, глазодвигательные мышцы. Волокна длиной от 10 до 12 см.
3.
Сократительный аппарат поперечно-полосатой мышечной ткани
Поперечно-полосатая сердечная мышечная ткань
Состоит из 1 или 2-х ядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы(по периферии цитолеммы). Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма.Существует также другой межклеточный контакт- аностамозы(впячивание цитолеммы одной клетки в цитолемму другой) Этот вид мышечной ткани образует миокард сердца. Развивается из миоэпикардальной пластинки (висцерального листка спланхнотома шеи зародыша) Особым свойством этой ткани является автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках(типичные кардиомиоциты). Эта ткань является непроизвольной(атипичные кардиомиоциты).
Существует 3-й вид кардиомиоцитов- секреторные кардиомиоциты (в них нет фибрилл) Они синтезируют гормон тропонин, понижающий АД и расширяющий стенки кровеносных сосудов.
Мышечная оболочка сердца (миокард) — образована мышечными полостными клетками, которые с помощью вставочных дисков соеденяются в мышечные комплексы или сердечные мышечные волокна. Мышечная ткань имеет свойства восстанавливаться, она защищена соеденительной тканью, образующий рубец.
Лекция добавлена 05.02.2014 в 10:26:09
Прежде всего следует отметить, что гладкая мышечная ткань состоит из клеток (гладкие миоциты, лейомиоциты), различных по своему происхождению. Лейомиоциты представляют собой гетерогенную и изменчивую популяцию клеток, длина которых колеблется от 20 до 200 мкм (но в матке во время беременности может достигать 500 мкм), в поперечнике их размеры достигают 4-8 мкм.
Клетки обычно собираются в пучки, являющиеся функциональными единицами ткани. Механизм объединения гладкомышечных клеток многообразен. На их концах имеются пальцевидные впячивания с десмосомами и нексусами. Пучки объединяются коллагеновыми и эластическими волокнами и гликопротеиновым промежуточным веществом. Все эти вещества вырабатываются в основном самими лейомиоцитами. На поверхности клеток находятся многочисленные мелкие впячивания (кавеолы), а вблизи плазмолеммы – пиноцитозные пузырьки.
В клетках располагаются миофиламенты, содержащие актин и миозин, образующие пучки, ориентированные вдоль или косо по отношению к длинной оси лейомиоцита. Строение сократительного аппарата гладкомышечных клеток имеет значительные отличия от такового в попечечно-полосатой мышечной ткани.
На внутренней стороне плазмолеммы гладкомышечных клеток располагаются пластинки прикрепления (плотные пластинки). Они имеют вид длинных (возможно, непрерывных) «ребер», идущих параллельно друг другу вдоль лейомиоцита. Периферическая часть этих пластинок, прилежащая к сарколемме, образована филаментами немышечного актина (нмАКТ). В глубоких слоях пластинок прикрепления к молекулам немышечного актина прикрепляются филаменты мышечного актина (мАКТ) с помощью связующих белков.
Плотные тельца (аналоги Z-мембран поперечно-полосатой мышечной ткани) располагаются в цитоплазме не свободно, как считалось до сих пор, а в виде цепочек, соединяясь друг с другом нитями нмАКТ. Пучки филаментов мышечного актина проникают в плотные тельца под углом (рис. 14).
Таким образом, получается, что основная масса цитоплазмы гладкомышечных клеток занята актиновыми и миозиновыми белками, которые, взаимодействуя друг с другом, образуют линейные структуры – «сократимые единицы». Актиновые филаменты связываются на концах сократимых единиц с плотными тельцами (см. рис. 14), вследствие чего каждая сократимая единица закрепляется в двух точках поверхностного аппарата клетки.
Предполагают, что примембранные плотные пластинки расположены по спирали и поэтому сократимые единицы оказываются под некоторым углом к длинной оси клетки. При сокращении лейомиоцита вся поверхность клетки приобретает бугристый и спиральный вид благодаря образованию глубоких впячиваний в местах прикрепления сократимых единиц.
Каждая сократимая единица представляет собой структуру, аналогичную миофибрилле поперечнополосатого волокна, но в ней нет правильного саркомерного распределения тонких и толстых миофиламентов. Протофибриллы лежат более или менее параллельно, однако на поперечном срезе их расположение неупорядоченное.
Основу толстых филаментов составляют хвосты молекул миозина в комплексе с другими дополнительными белками. Головки миозиновых молекул располагаются с внешней стороны этих комплексов, по спирали, рядами. Взаимодействие их с актиновыми филаментами, вероятно, происходит в разных направлениях. Одна миозиновая нить взаимодействует с 10-15 актиновыми.
Сократительный аппарат гладкой мышечной ткани
Миозин гладких мышц отличается от миозина соматической мускулатуры. Он имеет в 10 раз более низкую активность АТРазы головок, обладает большей чувствительностью к рН, ионному составу и концентрации кальция.
Актин тонких филаментов лейомиоцитов также своеобразен и отличается несколько иным аминокислотным составом. В тонких нитях этих клеток находится тропомиозин, а тропонин в них не обнаружен.
Дата публикования: 2014-12-30; Прочитано: 305 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.001 с)…
МЫШЕЧНЫЕ ТКАНИ
А) миофибриллы Б) тонофибриллы
В) нейрофибриллы г) псевдоподии
Способность сердца сокращаться под действием собственных импульсов
А) систола б) диастола в) автоматизм г) рефрактерность
76. Расположение органа ближе к задней поверхности тела
А) медиальное Б) латеральное
В) поверхностное Г) дорсальноеД) вентральное
Кость растет в ширину за счет
А) суставного хряща Б) надкостницы
В) метафизарного хряща Г) костного мозга
Скелет туловища образован
А) грудиной, позвоночником
Б) грудная клетка, позвоночник
В) позвоночник, ребра, кости таза
Околосердечная сумка называется
А) эпикард Б) перикард
В) миокард Г) эндокард
Агглютиногены 1 группы крови
А) А; Б) В; В) А и В; Г) все не верно.
Процесс склеиваяния эритроцитов
А) гемолиз в) агглютинация
Б) гемопоэз г) гемостаз
Ткань, выстилающая внутреннюю поверхность полых органов
А) мышечная в) эпителиальная
Б) нервная г) рыхлая соединительная
83. Белок, участвующий в образовании тромба
А) альбумин в) фибриноген
Б) глобулин г) фибрин
Продолжительность сердечного цикла
А) 0,5 сек б) 0,6 сек в) 0,7 сек г) 0,8 сек.
Малый круг кровообращения начинается
А) с правого предсердия в) с левого желудочка
Б) с правого желудочка г) с левого предсердия
86.Концентрациягипотонического раствора
А) 0,9% б) больше 0,9% в) меньше 0,9%
Кость снаружи покрыта веществом
А) губчатым в) хрящом д) серозной оболочкой
Б) компактным г) надкостницей
Подвижная кость черепа
А) верхняя челюсть в) нижняя челюсть
Б) височная г) скуловая
Основная функция миоцита
А) секреция в) саморегуляция
Б) сокращения г) раздражение
Мышца, предохраняющая вены шеи
А) грудино-ключично-сосцевидная
Б) поверхностная мышца шеи (платизма)
В) двубрюшная
Агглютиногены находятся в
А) плазме крови б) сыворотке крови
В) эритроцитах в) в стандартных сыворотках
Большой круг кровообращения закачивается
а) в правом желудочке в) в левом предсердии
б) в правом предсердии г) в левом желудочке
Средняя часть трубчатой кости называется
А) эпифиз б) диафиз
В) метафиз г) апофиз
Морфоструктурная единица соединительной ткани
А) ретикулоцит б) нейроцит В) фибробласт г) симпласт
Клетка тела, не имеющая ядра и митохондрий
А) симпласт б) миоцит
В) эритроцит в) лейкоцит
Продолжительность жизни тромбоцитов
А) 20 – 30 дней б) 3-7 дней в) 120 дней
К центральным органам иммунной системы относятся
А) красный костный мозг б) миндалины
В) селезенка в) лимфатические узлы
Лимфоэпителиальное кольцо Пирогова-Вальдейера образуют миндалины
А) нёбные, глоточная, трубные, аппендикулярные
Б) нёбные, глоточная, трубные, язычная
В) глоточная, трубные, аппендикулярнык
Фактор, активизирующий почти все фазы свертывания крови
А) ионы Са б) ионы Na в) ионы Mg
Ррезус-принадлежность крови определяют
А) плазма крови б) тромбоциты
В) лейкоциты в) эритроциты
Венозному возврату крови в правое предсердие способствует
А) толстая стенка артериальных сосудов в) клапаны верхней и нижней полых вен
Б) присасывающая способность сердца
102. Что обозначает зубец QRS на ЭКГ
А) систолу предсердий б) систолу желудочков
В) диастолу предсердий г) диастолу желудочков
Какой сосуд имеет полулунные клапаны
А) аорта б) плечевая артерия
В) общая сонная артерия г) плечеголовной ствол
От какой части собирает кровь нижняя полая вена
А) верхней половины тела б) грудной клетки
В) головы и шеи г) нижней половины тела
Концевые окончания волокна Пуркинье заканчиваются на клетках миокарда
А) обоих желудочков в) предсердий и желудочков
Б) обоих предсердий г) в левом желудочке
Где расположена селезенка
А) в правом подреберье б) в левом подреберье
В) в эпигастральной области г) в левой подвздошной области
Лимфатические капилляры начинаются от
А) межклеточных щелей б) лимфоузлов
В) селезенки в) красного костного мозга
Строму лимфатического узла составляет ткань
А) соединительная б) рыхлая волокнистая
В) ретикулярная в) пигментная
Грудной лимфатический проток начинается на уровне позвонка
А) 2- го поясничного б) 12- го грудного
В) 3 – 4 поясничных г) 7 – 8 грудных
Внутренняя оболочка сердца называется
А) эпикард в) миокард
Б) эндокард г) перикард
Зрелая клетка костной ткани называется
А) остеокласт в) остеоцит
Б) остебласт г) остеон
Морфоструктурная единица костной ткани называется
А) нейрон б) остеон в) нефрон г) оссеин
Уровень расположения лопаток
А) 1- 6 ребро в) 1- 7 ребро
Б) 2- 6 ребро г) 2- 8 ребро
⇐ Предыдущая123
magictemple.ru
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ СКЕЛЕТНОЙ — МегаЛекции
МЫШЕЧНОЙ ТКАНИ
Скелетная (соматическая) мышечная ткань образована пучками поперечнополосатых мышечных волокон, являющихся ее структурно-функциональными единицами. Всего в скелетных мышцах человека содержится порядка 300 млн. мышечных волокон (рис. 7.1, 7.2).
| А | Б |
Рис. 7.2. На микрофотографии (х 320) изображена: Скелетная мышца А – продольный разрез, Б – поперечный разрез Поперечно-полосатые волокна имеют цилиндрическую форму, расположены параллельно друг другу, ядра лежат под сарколеммой. Волокна имеют поперечную исчерченность. Между волокнами – эндомизий. С – разрезы капилляров, Р – перимизий.
МЫШЕЧНОЕ ВОЛОКНО скелетной (соматической) мышечной ткани представляет собой цилиндрическое образование диаметром 10-100 мкм (в среднем —
50 мкм) вариабельной длины (до 10-30 см). Мышечные волокна в мышцах образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму (рис 7.2).
Миосимпластическая часть мышечного волокнавключает от нескольких сотен до нескольких тысяч ядер, лежащих на периферии под сарколеммой, и саркоплазму, образующую его центральную часть.
Саркоплазма миосимпласта содержит все органеллы общего значения (за исключением центриолей) и некоторые специальные органеллы, а также включения. Эти структуры образуют несколько функциональных аппаратов: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат мышечного волокнапредставлен миофибриллами – специальными органеллами, которые располагаются продольно в центральной части саркоплазмы и отделяются друг от друга рядами вытянутых митохондрий и цистерн саркоплазматической сети.
Миофибриллы имеют вид нитей диаметром 1-2 мкм и длиной, сопоставимой с протяженностью волокна. Их количество в отдельном волокне варьирует в широких пределах (от нескольких десятков до 2000 и более). Они обладают собственной поперечной исчерченностью. А- и I-диски одних миофибрилл точно совпадают с аналогичными дисками других, обусловливая поперечную исчерченность всего волокна. Структурно-функциональной единицей миофибриллы является саркомер (миомер).
Поперечная исчерченностъ скелетных мышечных волокон обусловлена чередованием темных А-дисков (анизотропных, обладающих двойным лучепреломлением в поляризованном свете) и светлых 1-дисков (изотропных, не обладающих двойным лучепреломлением). Каждый диск I рассекается надвое тонкой темной Z, называемой также телофрагмой. В середине А-диска определяется светлая зона – полоска Н, через центр которой проходит М-линия — мезофрагма (рис. 7.3).
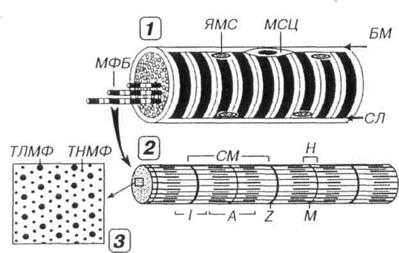
Рис. 7.3. Строение скелетного мышечного волокна (1), миофибриллы (2) и расположение в последней миофиламентов (3). В мышечном волокне (MB) в целом и каждой миофибрилле (МФ), входящей в его состав, выявляются чередующиеся темные анизотропные А-диски (А) и светлые изотропные I-диски (I). Последние рассекаются надвое телофрагмой, или Z-линией (Z), а в середине первых определяется светлая полоска Н (Н), через центр которой проходит М-линия (М). СМ — саркомер, ЯМС — ядра миосимпласта, МСЦ — миосателлитоцит, БМ — базальная мембрана, СЛ — сарколемма. В пределах СМ каждый толстый миофиламент (ТЛМФ) окружен шестью тонкими миофиламентами (ТНМФ).
Саркомер (миомер) представляет собой участок миофибриллы, расположенный между двумя телофрагмами (Z-линиями) и включающий А-диск и две половины 1-дисков — по одной половине с каждой стороны (см. рис. 7.3; 7.6). В расслабленной мышце длина саркомера составляет около 2-3 мкм, а ширина его участков выражается соотношением Н : А : I = 1 : 3 : 2; при сокращении мышцы саркомер укорачивается до 1.5 мкм. Миофибрилла типичного мышечного волокна человека длиной около 5 см насчитывает порядка 20 тыс. последовательно расположенных саркомеров.
Структура саркомера представлена упорядоченной системой толстых и тонких белковых нитей (миофиламентов). Толстые нити (диаметром около 10-12 нм и длиной 1.5-1.6 мкм) связаны с мезофрагмой и сосредоточены в А-диске, а тонкие (диаметром 7-8 нм и длиной 1 мкм) прикреплены к телофрагмам, образуют I-диски и частично проникают в А-диски между толстыми нитями (более светлый участок А-диска, свободный от тонких волокон, называется полоской Н). В саркомере насчитывается несколько сотен толстых нитей. По сечению саркомера толстые и тонкие нити располагаются высокоорганизованно в узлах гексагональной решетки. Каждая толстая нить окружена шестью тонкими, каждая из тонких нитей частично входит в окружение трех соседних толстых (см. рис. 7.3).
Толстые нити (миофиламенты) образованы упорядоченно упакованными молекулами фибриллярного белка миозина, на который приходится около 54% всех белков миофибриллы. Молекула миозина имеет вид нити длиной 150 нм и толщиной 2 нм. На одном из концов эта молекула содержит две округлые головки длиной около 20 нм и шириной около 4 нм (рис. 7.4). Протеолитическими ферментами миозин расщепляется на две фракции – легкий меромиозин («стержень» молекулы миозина) и тяжелый меромиозин (участки головок и шейки, связывающие их со стержневой частью). Молекула миозина может сгибаться, как на шарнирах, в месте соединения тяжелого меромиозина с легким и в области прикрепления головки. Стержневые части молекул миозина собраны в пучки (численностью до 200 и более) (рис. 7.4 (2)). Такие пучки, соединенные зеркально концами друг с другом в области М-линии, формируют толстые нити с центральной гладкой частью длиной около 0.2 мкм и двумя периферическими участками, в которых от центрального стержня отходят миозиновые головки (около 500). Миозин головок обладает АТФазной активностью (способностью осуществлять гидролиз АТФ), однако в отсутствие его взаимодействия с актином скорость гидролиза АТФ ничтожно мала.
Рис. 7.4. Строение толстых миофиламентов (по К.Де Дюву, 1987, с изменениями). 1 — молекула миозина имеет вид нити с двумя головками (Г) на одном конце. Миозин включает легкий меромиозин (ЛММ), образующий стержневую часть молекулы, и тяжелый меромиозин (ТММ), соответствующий участкам Г и связующей шейки. Участки сгибания молекулы миозина показаны стрелками. 2 — стержневые части молекул миозина собраны в пучки (П), снаружи которых располагаются миозиновые Г. 3 — толстые миофиламенты (ТЛМФ) образованы П молекул миозина, соединенными зеркально концами друг с другом. Центральная часть ТЛМФ — гладкая, периферическая содержат многочисленные миозиновые Г.
Тонкие нити (миофиламенты)содержат сократимый белок актин (на него приходится 20% белков миофибриллы) и два регуляторных белка – тропонин (около 2%) и тропомиозин (около 7%). Последние формируют функционально единый тропонин-тропомиозиновый комплекс.
Актин в мономерной форме представлен полярными глобулярными субъединицами диаметром 4-5 нм (G-актин), которые имеют активные центры, способные связываться с молекулами миозина. G-актин агрегирует с образованием полимерного фибриллярного актина (F-актина), молекула которого имеет вид двух скрученных нитей толщиной 7 нм и вариабельной длины (рис. 7.5).
Рис. 7.5. Строение тонких миофиламентов. Тонкие миофиламенты (ТНМФ) содержат сократимый белок актин (АКТ) и два регуляторных белка — тропонин (ТРН) и тропомиозин (ТРМ). Глобулярные субъединицы АКТ (G-AKT), агрегируют с образованием фибриллярного АКТ (F-AKT), молекула которого имеет вид двух скрученных нитей. ТРМ образован нитевидными молекулами, соединяющимися своими концами и образующими тяж, лежащий в борозде молекулы F-AKT. ТРН — глобулярный белок, связанный с молекулой ТРМ и формирующий с ней функционально единый комплекс ТРН-ТМ.
Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуя длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Так как таких борозд на молекуле актина две, то и тропомиозиновых нити тоже две. Всего в состав тонкой нити входит примерно 50 молекул тропомиозина.
Тропонин представляет собой глобулярный белок, каждая его молекула располагается на тропомиозиновой молекуле вблизи ее конца. Тропонин состоит из трех субъединиц: ТпС – связывающей кальций, ТпТ – прикрепляющейся к тропомиозину, и Tnl – ингибирующей связывание миозина с актином.
Механизм мышечного сокращенияописывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что тонкие нити вдвигаются в промежутки между толстыми без изменения их длины (рис. 7.6).
Скольжение нитей в саркомере и усилие, развиваемое мышцей, обеспечиваются благодаря циклической активности миозиновых мостиков, которые при сокращении повторно прикрепляются к актину, обеспечивают усилие тяги, а затем открепляются от него (рис. 7.7). В этом механизме АТФ играет двойную роль, обеспечивая энергию, необходимую как для осуществления сокращения, так и для открепления мостиков.
Рис. 7.6. Механизм мышечного сокращения в соответствии с теорией скользящих нитей. Укорочение саркомеров (СМ) при сокращении (2) по сравнению с их состоянием в покое (1) происходит благодаря тому, что тонкие миофиламенты (ТНМФ) вдвигаются в промежутки между толстыми (ТЛМФ) без изменения их длины. Остальные обозначения — как на рис. 13-3.
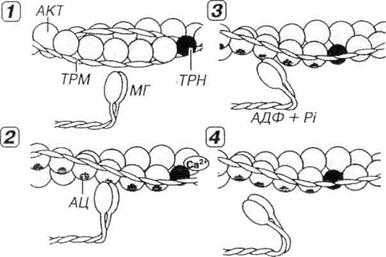
Рис. 7.7. Молекулярные механизмы мышечного сокращения.,1 — в покое миозиновые головки (МГ), с которыми связаны молекулы АТФ, неспособны взаимодействовать с активными центрами (АЦ) на молекуле актина (АКТ), потому что последние прикрыты комплексом тропонин-тропомиозин (ТРН-ТРМ). 2 — мышечное сокращение начинается вследствие повышения концентрации Са2+, который воздействует на ТРН. Возникающее изменение конформации ТРН и смещение молекулы связанного с ним ТРМ демаскирует АЦ на молекуле АКТ, с которыми связываются МГ, образуя поперечные мостики. 3 — за счет сгибания МГ в области их прикрепления к молекуле АКТ развивается усилие, смещающее тонкие миофиламенты (ТНМФ) вдоль толстых (ТЛМФ) к центру саркомера (см. рис. 13-06). АТФ при этом гидролизуется до АДФ и фосфата (Pi). 4 — размыкание мостика и его отделение от ТНМФ наступают вследствие связывания с ним новой молекулы АТФ. Далее мостик принимает исходное положение (перпендикулярное ТНМФ) и начинается новый цикл сокращения. Циклическое взаимодействие МГ и ТНМФ будет продолжаться при сохранении высокой концентрации ионов Са2+ и наличии АТФ.
В покое (при очень низкой концентрации ионов Са2+) в миофибрилле расслабленного мышечного волокна толстые и тонкие нити не соприкасаются. Миозиновые головки (с которыми связаны молекулы АТФ) не могут взаимодействовать с активными центрами (участками связывания миозина) на молекуле актина, потому что последние прикрыты тропонин-тропомиозиновым комплексом. Толстые и тонкие филаменты беспрепятственно скользят друг относительно друга. При этом мышечные волокна почти не сопротивляются пассивному растяжению.
Мышечное сокращение вызывается резким повышением концентрации ионов Са2+ в области миофиламентов и включает несколько этапов (см. рис. 7.7).
А. Связывание ионов Са2+ с тропонином и освобождение активных центров на молекуле актина. При этом тропонин изменяет свою конформацию, смещает молекулы тропомиозина и открывает активные центры (участки связывания миозина) на молекуле актина.
Б. Связывание миозина и актина (формирование поперечных мостиков). Миозиновые головки связываются с активными центрами на молекуле актина, формируя мостики, расположенные перпендикулярно продольной оси нити. Менее чем через 1 мс после этого под влиянием актомиозинового комплекса происходит гидролиз АТФ и отщепление его продуктов (АДФ и неорганического фосфата). При этом угол наклона мостика относительно продольной оси нити изменяется до 40°. Такой конформационный переход, происходящий в области прикрепления головки миозиновой молекулы, обусловливает развитие усилия и смещение тонких филаментов к центру саркомера. Предполагается, что «рабочий ход» миозинового мостика составляет около 10 нм. Таким образом, за один цикл мостик вызывает относительное перемещение тонких нитей на расстояние, равное примерно 1/200 длины саркомера.
B. Размыкание мостика. Связывание новой молекулы АТФ с мостиком вызывает его отделение от тонкого филамента. Мостик размыкается, возвращаясь в прежнее положение относительно миозиновой нити и может прийти в замыкание со следующим активным центром на тонкой. Каждый цикл замыкания-размыкания сопровождается расщеплением молекулы АТФ. В живой мышце это осуществляется с интервалом в несколько десятков миллисекунд после присоединения новой молекулы АТФ. В трупной мышце, где АТФ отсутствует, мостик не может разомкнуться, и мышца переходит в состояние трупного окоченения.
При сокращении мышцы не происходит одновременного замыкания всех мостиков – их число нарастает по ходу его развития. При последующем расслаблении мышцы число мостиков снижается.
Аппарат передачи возбуждения (саркотубулярная система)необходим для того, чтобы распространяющаяся по сарколемме волна деполяризации могла вызвать срабатывание сократительного аппарата миофибрилл. В мышечном волокне связь между возбуждением и сокращением выполняют две специализированные мембранные системы — саркоплазматическая сеть и поперечные (Т-) трубочки (от англ. transverse — поперечный), образующие функционально единую саркотубулярную систему (рис. 7.8)

Рис. 7.8. На электронной микрофотографии (x 44.000) изображен: Поперечный срез скелетной мышцы сделанный на уровне соединений А и J дисков. Хорошо видны детали строения СПР (SR). SR располагается между миофибриллами. ICS – intracellular space – внутриклеточное пространство, L – наружная пластинка, Т – трубочки, М – митохондрии.
Саркоплазмашическая сеть — система уплощенных, вытянутых и анастомозирующих мембранных трубочек и мешочков, которая окружает каждый саркомер, миофибриллы наподобие муфты. В области наружных отделов А- и I-дисков трубочки сливаются, образуя пары плоских терминальных цистерн (на каждый саркомер приходится по две такие пары). Саркоплазматическая сеть обладает выраженной способностью депонировать и выделять ионы кальция. Ее мембрана содержит высокие концентрации интегральных белков, являющихся кальциевыми насосами, а на внутренней поверхности находится белок кальсеквестрин, связывающий ионы Са2+.
Поперечные (Т-) трубочки представляют собой впячивания сарколеммы, отходящие от нее под прямым углом к оси волокна и расположенные у млекопитающих вблизи границы I- и А- дисков. Ветви соседних Т-трубочек опоясывают каждый саркомер и анастомозируют друг с другом. Конечные участки Т-трубочек проникают в промежуток между двумя терминальными цистернами саркоплазматической сети (см. рис. 7.7), формируя вместе с ними особые структуры — триады. В области триады между параллельно лежащими мембранами Т-трубочки и терминальных цистерн, разделенными узкой щелью, имеются специализированные контакты, которые образованы рядами плотных частиц (ножек), предположительно служащие каналами выделения кальция.
Выделение кальцияпроисходит после того, как волна деполяризации с поверхности сарколеммы по Т-трубочкам распространяется вглубь волокна. В области триад возбуждение передается на мембрану саркоплазматической сети и вызывает повышение ее проницаемости. Это приводит кбыстрому выделению из ее элементов ионов кальция (главным образом, в области терминальных цистерн). Выделившийся Са2+ диффундирует в миофибриллы, где он, присоединяясь ктропонину, запускает механизм взаимодействия актина и миозина. Концентрация Са2+ вокруг миофиламентов при этом резко повышается с 10-7М до 10-5М.
Активный обратный транспорт кальция в саркоплазматическую сеть (секвестрация кальция) происходит наряду с его выбросом, который представляет собой кратковременный процесс.
Опорный аппарат мышечного волокнавключает особые элементы цитоскелета, обеспечивающие высокоупорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также связанную с ними сарколемму и базальную мембрану, соединяющие мышечное волокно с сухожилием, на которое передается усилие, развиваемое волокном при сокращении. К опорным элементам относятся телофрагма и мезофрагма.
Телофрагма (Z-линия) — область прикрепления тонких миофиламентов двух соседних саркомеров; она имеет вид плотной полоски шириной 30-100 нм без резких границ.
Мезофрагма (М-линия) — плотная линия шириной 75-85 нм, расположенная в центре А-диска и являющаяся областью закрепления толстых (миозиновых) филаментов в саркомере.
Структура краевых участков мышечных волокон. На концах мышечных волокон сарколемма, покрытая базальной мембраной, образует многочисленные глубокие впячивания, в которые вдаются коллагеновые волокна сухожилия, вплетающиеся в базальную мембрану и прочно связывающие сухожилие с мышечными волокнами.
Энергетический аппарат мышечных волоконпредставлен митохондриями, вырабатывающими энергию, необходимую для осуществления мышечной работы, синтетических, транспортных и других процессов жизнеобеспечения, а также трофическими включениями (гликоген, липиды, миоглобин), содержащими вещества, расщепление которых служит источником энергии.
Синтетический аппарат мышечного волокнапредставлен свободными рибосомами и полирибосомами (особенно многочисленными под сарколеммой в области I-диска и вблизи ядер), цистернами грЭПС и комплексом Гольджи, элементы которого в виде сотен или тысяч стопок мешочков рассеяны по саркоплазме миосимпласта.
Лизосомальный аппарат (аппарат внутриклеточного переваривания) в мышечных волокнахнеобходим для обеспечения постоянно протекающего процесса обновления его структурных компонентов. Содержание лизосом связано с функциональной активностью мышцы и возрастом человека. Остаточные тельца лизосомального генеза, содержащие липофусцин, становятся многочисленными при старении и, в особенности, при резком снижении функциональной активности мышцы.
Типы мышечных волокон
Выделяют три основных типа мышечных волокон, между которыми существуют переходные варианты (рис. 7.9): тип I (красные), тип IIВ (белые) и тип IIА (промежуточные).
Тип I — красные, медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами, высокой активностью окислительных ферментов, преобладанием аэробных процессов, высоким содержанием миоглобина (определяющим их красный цвет). Численно преобладают в мышцах, выполняющих длительные тонические нагрузки.

Рис. 7.9. Типы мышечных волокон в скелетной мышце. На препарате – поперечном срезе мышечных волокон – проведено гистохимическое выявление фермента сукцинатдегидрогеназы (СДГ). Плотность продукта гистохимической реакции пропорциональна активности СДГ. Представлены три основных типа мышечных волокон: тип I (I) — красные (с высокой активностью СДГ), тип ИВ (ИВ) – белые (с низкой активностью СДГ) и тип НА (НА) – промежуточные (с умеренной активностью СДГ).
Тип IIВ — белые, быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, низким содержанием липидов и миоглобина (определяющим их светлый цвет), Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип IIА — промежуточные, быстрые, устойчивые к утомлению, с большой силой, оксилительно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и ПВ.
СКЕЛЕТНАЯ МЫШЦА КАК ОРГАН
Скелетная мышца состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов.
Количество мышечных волокон в отдельных мышцах человека варьирует в широких пределах. Так, в четырехглавой мышце бедра (m. quadriceps femoris) оно составляет около 1.7 млн., икроножной мышце (m. gastrocnemius) — 1-1.5 млн., портняжной мышце (m. sartorius) – 100-200 тыс., в двуглавой и трехглавой мышцах плеча (m. biceps и т. triceps brachii) оно примерно одинаково и колеблется в пределах 200-600 тыс.
Соединительнотканные компоненты мышцы представлены эпимизием, перимизием и эндомизием (рис. 7.10, 7.11). Общее содержание соединительной ткани в скелетной мышце очень значительно – в зависимости от типа мышцы коллаген соединительной ткани составляет от 3 до 30% ее белков.

Рис. 7.10. Структурная организаций скелетной мышцы. Мышца (М) покрыта эпимизием (ЭПМ), отдающим вглубь более тонкие соединительнотканные перегородки — перимизий (ПМ), который образует оболочки пучков мышечных волокон (ПМВ). От ПМ внутрь ПМВ отходят тончайшие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно (MB), именуемые эндомизием (ЭНМ). MB покрыто сарколеммой (СЛ) и базальной мембраной (не показана) и обладает поперечной исчерченностью; в его центральной части содержатся поперечно исчерченные миофибриллы (МФБ), образованные системой тонких и толстых миофиламентов — МФЛ.
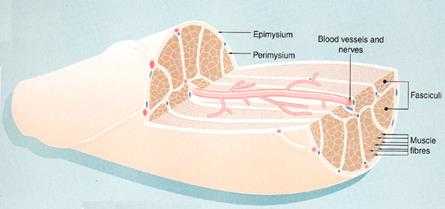
Рис. 7.11. Строение мышцы как органа. Мышечные волокна (Muscle fibers) образуют пучки (Fasciculi), каждый из которых соединен рыхлой соединительной тканью – перимизием (Perimysium), снаружи мышца покрыта соединительной тканью, образующей эпимизий (Epimysium). Между волокнами расположены прослойки соединительной ткани – эндомизий (Endomysium). В эндо-, пери- и эпимизии проходят сосуды (blood vessels) и нервы (nerves).
Эпимизий — тонкий, прочный и гладкий снаружи чехол из плотной волокнистой соединительной ткани, окружающий всю мышцу.
Перимизий — тонкие, разветвляющиеся и не всегда четко очерченные соединительнотканные перегородки, отходящие от внутренней поверхности эпимизия вглубь мышцы. Он образует оболочки отдельных пучков мышечных волокон, численностью 10-100 (наиболее часто — около 20) волокон.
Эндомизий — тончайшие прослойки рыхлой волокнистой соединительной ткани, отходящие от перимизия внутрь пучков мышечных волокон и окружающие каждое мышечное волокно. Соединительнотканные волокна эндомизия вплетаются в базальную мембрану мышечных волокон.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Сократительный аппарат мышечных волокон — Справочник химика 21
Принято считать, что процессом, непосредственно связанным с работающим механизмом поперечно-полосатого мышечного волокна, является распад АТФ с образованием АДФ и неорганического фосфата. Возникает вопрос каким образом мышечная клетка может обеспечить свой сократительный аппарат достаточным количеством энергии в форме АТФ, т.е. каким образом в процессе мышечной деятельности происходит непрерывный ресинтез этого соединения [c.654]Поперечно-полосатая структура мышечных волокон может наблюдаться под обычным микроскопом. Отдельное мышечное волокно имеет диаметр 20 — 80 мкм и окружено плазматической мембраной толщиной 10 нм. Каждое отдельное волокно -это сильно вытянутая клетка. Длина отдельных волокон (клеток) может существенно варьироваться, в зависимости от вида мышцы, от сотен микрон до нескольких сантиметров. Внутри волокна, кроме известных органелл (ядро, ядрышко, митохондрии, аппарат Гольджи и др.), находятся сократительный аппарат клетки, состоящий из 1000 — 2000 параллельно расположенных миофибрилл диаметром 1-2 мкм, а также клеточные органеллы саркоплазматический ретикулум и система поперечных трубочек — Т-система. [c.144]
Сократительный аппарат мышечных клеток — это не однородная система. Следует различать области, содержащие только актин или только миозин, и области, в которых волокна этих двух белков находятся во взаимном зацеплении. Более того, вода сама представляет собой непрерывную фазу, пронизывающую всю систему. Природа структуры воды, окружающей белки, обсуждается Берналом [67] представляется, что в ближайшем к белкам слое толщиной ЮЛ вода находится в льдообразном состоянии. [c.289]
Мышечными называют все типы клеток, функция которых состоитв сокращении, хотя в остальном эти клетки могут быть мало сходны между собой. Как уже говорилось в гл. 11, сократительный аппарат, включающий актин и миозин, — это фундаментальная особенность животных клеток вообще, но в мышечных клетках он особенно сильно развит. У млекопитающих имеются четыре главных типа клеток, специально приспособленных для сокращения волокна скелетных мышц, клетки сердечной мышцы, гладкомышечные и миоэпителиальные клетки (рис. 17-36). Они различаются по функции, структуре и пути развития. [c.189]
Клетки скелетных мышц, сократительный аппарат которых детально рассмотрен в гл. И, ответственны практически за все произвольные движения. Эти клетки могут иметь огромные размеры (до полуметра в длину и до 100 мкм в диаметре у взрослого человека) и за свою форму получили также название мышечных волокон. Каждая такая клетка представляет собой синцитий, содержащий много ядер в общей цитоплазме. В отличие от этого мышечные клетки трех других типов имеют более обычное строение — в них только по одному ядру. Клетки сердечной мышцы сходны с волокнами скелетной мускулатуры в том отношении, что нити актина и миозина в них образуют упорядоченные системы, придающие клетке исчерченный вид. Гладкомышечные клетки получили свое название потому, что они, напротив, не выглядят исчерченными. Функции у гладкой мускулатуры весьма разнообразны — от проталкивания пищи по пищеварительному тракту до поднятия шфсти дыбом при холоде или страхе. Миоэпителиальные клетки (тоже лишенные исчфченности) в отличие от клеток трех других типов лежат в эпителии и происходят из эктодермы. Эти клетки образуют мускулатуру радужной оболочки глаза, расширяющую зрачок, а также используются для выдавливания слюны, пота и молока из соответствующих желез (см. рис. 17-36, Д). [c.190]
www.chem21.info
Вопрос 2. Организация мышечного волокна.
Структурно-функциональной единицей этой ткани является мышечное волокно. Это длинный цитоплазматический тяж со множеством ядер, которые лежат сразу под плазмолеммой. Мышечное волокно в эмбриогенезе образуется при слиянии клеток – миобластов, т.е., представляет собой клеточное производное –симпласт.
Мышечное волокно сохраняет общий план клеточной организации. В нем есть все органеллы общего значения, много включений, а также органеллы специального значения. Все компоненты волокна адаптированы для выполнения главной функции – сокращения – и подразделяются на несколько аппаратов.
Сократительный аппарат состоит из миофибрилл. Это органеллы, которые тянутся вдоль всего волокна и занимают большую часть всего объема цитоплазмы. Они способны значительно изменять свою длину.
Аппарат белкового синтеза представлен, в основном, свободными рибосомами и специализирован на выработке белков для построения миофибрилл.
Аппарат передачи возбуждения образован саркотубулярной системой. Она включает гладкую ЭПС и Т-трубочки. Гладкая ЭПС (саркоплазматическая сеть) имеет вид плоских цистерн, которые оплетают все миофибриллы. Она служит для накопления Сa2+. Ее мембраны способны быстро выпускать кальций наружу, что необходимо для укорочения миофибрилл, а затем активно закачивает его внутрь. Наружная мембрана мышечного волокна (сарколемма) образует многочисленные трубчатые впячивания, пронизывающие все волокно в поперечных направлениях. Их совокупность называют Т-системой. Т-трубочки тесно контактируют с мембранами ЭПС, образуя единую саркотубулярную систему. К каждой Т-трубочке …..
Энергетический аппарат составлен митохондриями и включениями. Митохондрии крупные вытянутые и лежат, в основном цепочками, заполняя все пространство между миофибриллами. Субстратами для получения АТФ служит гликоген и липидные капли. Включения миоглобина – специфического мышечного пигмента, обеспечивают волокна кислородом в случае длительной и напряженной работы мышц.
Лизосомальный аппарат развит слабо. Служит, главным образом, для процессов внутриклеточной регенерации.
Вопрос 3 Механизм мышечного сокращения. Для его понимания необходимо ознакомиться с молекулярной организацией миофибрилл – органелл, специализированных на сокращении. Это длинные тяжи, образующие продольные пучки по тысяче и более миофибрилл, которые почти полностью заполняют цитоплазму волокна.
Каждая миофибрилла построена из огромного числа актиновых и миозиновых филаментов.
Тонкие актиновые нити построены из глобулярных молекул белка актина, которые объединяются в две спирально закрученные цепочки. Более толстая миозиновая нить построена из 300-400 молекул белка миозина. Каждая молекула включает длинный хвост, к которому с одного края прикреплена подвижная головка. Головки могут менять угол своего наклона. Хвосты множества молекул укладываются плотным пучком, формируя стержень филамента. Головки при этом остаются на поверхности. На двух краях нити головки лежат разнонаправленно.
Благодаря дополнительным белкам, миофиламенты имеют стабильный диаметр и стабильную длину около 1 мкм. Филаменты одного вида образуют аккуратно подогнанные пучки или стопки. Миофибриллы образованы из многократно чередующихся пучков актиновых и миозиновых нитей. Высокая упорядоченность в расположении миофиламентов достигается с помощью различных белков цитоскелета. Например, белок актинин формирует Z-линию (телофрагму), к которой с обеих сторон присоединяются края тонких актиновых нитей. Так образуется актиновая стопка. Другие белки организуют в стопку толстые миозиновые нити, прошнуровывая их в середине. Они образуют М-линию (мезофрагму). При чередовании стопок свободные концы тонких и толстых нитей заходят друг за друга, обеспечивая взаимное скольжение друг относительно друга в момент сокращения. В результате такой организации в миофибрилле многократно повторяются светлые участки, называемые I-дисками (изотропные), и темные участки, называемые А-дисками (анизотропные). Это и создает эффект поперечной исчерченности. Изотропные участки соответствуют центральной части актиновой стопки и содержат только тонкие нити. Анизотропные диски соответствуют целой миозиновой стопке, и в них входят чисто миозиновая часть (Н-полоска) и те участки, где концы тонких и толстых нитей перекрываются.
Участок между двумя Z-линиями называют саркомером. Саркомер является структурной единицей миофибриллы. (20 тысяч саркомеров на миофибриллу). Строгая организация миофибрилл обеспечивается широким набором различных белков цитоскелета.
При сокращении длина миофибриллы уменьшается за счет одновременного укорочения всех I-дисков. Длина каждого саркомера при этом уменьшается в 1,5-2 раза. Процесс сокращения объясняется теорией скольжения Хаксли, согласно которой в момент сокращения свободные, заходящие друг за друга концы актиновых и миозиновых нитей вступают в молекулярные взаимодействия и глубже задвигаются друг относительно друга. Скольжение начинается с того, что выступающие миозиновые головки образуют мостики с активными центрами актиновых филаментов. Затем угол наклона головки уменьшается, мостики совершают как бы гребковые движения, смещая и актиновую нить. После этого мостик размыкается, что сопровождается гидролизом 1 молекулы АТФ. Связывание миозиновых головок с актиновой нитью регулируется специальными белками. Это тропонин и тропомиозин, которые встроены в актиновую нить, и препятствуют контакту с миозиновыми головками. При повышении в гиалоплазме концентрации Са2+ происходит изменение конформационного состояния этих регуляторных белков и их блокирующее действие снимается. Гребковые движения повторяются сотни раз за одно мышечное сокращение. Расслабление наступает только после того, как снизится концентрация Ca2+.
studfiles.net
3. Сократительный аппарат гладких миоцитов образован:
а) актиновыми миофиламентами
б) белком -актинина
в) белком винкулина
г) миозиновыми миофиламентами
д) белком цитокератином
4. Гладкие миоциты могут синтезировать и выделять в межклеточное
вещество:
а) эластин б) протеогликаны в) гликозаминогликаны | г) гликопротеины д) фосфолипиды |
К трофическому аппарату гладкого миоцита относятся:
а) ядро б) митохондрии в) гранулярная ЭПС | г) комплекс Гольджи д) миофибриллы |
Для регенерации гладкой мышечной ткани характерны явления:
а) внутриклеточной регенерации
б) регенерационной гипертрофии
в) пролиферации миоцитов
г) рубцевания зоны повреждения
д) пролиферации миосателлитоцитов
Миоэпителиальные клетки входят в состав:
а) потовых желез б) молочных желез в) слюнных желез | г) слезных желез д) сальных желез |
В ходе гистогенеза сердечной мышечной ткани образуются следующие
типы кардиомиоцитов:
а) сократительные б) проводящие в) переходные | г) секреторные д) транспортные |
Гладкие миоциты образуют друг с другом следующие типы контактов:
а) щелевые в) десмосомы б) нексусы | г) простые д) синаптические |
Вставочные диски между кардиомиоцитами содержат следующие типы
контактов:
а) десмосомы в) нексусы б) интердигитации | г)зоны прикрепления миофиламентов д) синаптические контакты |
Проводящие миоциты отличаются от сократительных:
а) более крупными размерами
б) малочисленностью миофибрилл
в) обилием включений гликогена
г) упрощенными вставочными дисками
д) отсутствием L-системы
12. Для секреторных кардиомиоцитов характерны:
а) хорошо развитый аппарат Гольджи
б) хорошо развитая гранулярная ЭПС
в) обилие секреторных гранул
г) признаки экзоцитоза гранул
д) развитый сократительный аппарат
13. Для регенерации сердечной мышечной ткани характерны процессы:
а) внутриклеточной регенерации
б) регенерационной гипертрофии
в) рубцевания зоны повреждения
г) увеличения количества внутриклеточных органелл
д) размножения стволовых клеток
Основные особенности, отличающие структуру скелетной
мышечной ткани от сердечной:
а) структурной единицей является мышечное волокно
б) эмбриональный источник — миотомы сомитов
в) наличие миосателлитоцитов
г) наличие большого количества ядер на периферии волокна
д) наличие саркоплазматической сети
Скелетные мышечные волокна сходны с сердечными миоцитами
следующими признаками:
а) структурой эндомизия
б) наличием многочисленных митохондрий
в) наличием развитой саркоплазматической сети
г) саркомерной организацией миофибрилл
д) наличием вставочных дисков
Предсердные кардиомиоциты отличаются от желудочковых:
а) присутствием элементов гранулярной ЭПС
б) развитым аппаратом Гольджи
в) присутствием секреторных гранул
г) наличием L-системы д) наличием Т-системы
Скелетная мышечная ткань отличается от гладкой:
а) источниками эмбрионального развития
б) иннервацией
в) строением сократительного аппарата
г) межклеточными контактами
д) наличием сократительных миофиламентов
Иннервация скелетной мышечной ткани осуществляется:
а) эфферентными (двигательными)нервными волокнами
б) афферентными (чувствительными)нервными волокнами
в) двигательными (моторными) бляшками
г) чувствительными нервными окончаниями (мышечными веретенами)
д) вегетативными ганглиями
В состав миофибрилл входят следующие белки:
а) актин б) миозин в) тропонин | г) тропомиозин д) фибронектин |
Для регенерирующей скелетной мышечной ткани характерны процессы:
а) внутриклеточной регенерации
б) регенерационной гипертрофии
в) формирования рубца на месте повреждения
г) размножения миосателлитоцитов
д) полноценного (эпиморфного) заживления места повреждения
В саркомере миофибрилл:
а) толстые филаменты состоят из миозина и С-белка
б) тонкие филаменты состоят из актина, тропомиозина, тропонина
в) обнаруживается один А-диск и две половины I-диска
г) при сокращении уменьшается ширина I-диска
д) в середине I-диска проходит М-линия
В эмбриональном соматическом миогенезе:
а) миобласты происходят из клеток миотомов сомитов мезодермы
б) часть пролиферирующих миобластов обособляется в качестве клеток-сателлитов
в) в мышечных трубочках начинается сборка миофибрилл
г) ядра перемещаются на периферию симпласта
д) во время митозов дочерние клетки G1-миобластов остаются связанными цитоплазматическими мостиками, образуя мышечные трубочки
При расслабление гладкомышечной клетки:
а) в саркоплазме снижается концентрация Са2+
б) активируется аденилатциклаза
в) фосфатаза лёгких цепей миозина дефосфорилирует миозин
г) происходит дезинтеграция миозиновых миофиламентов
д) увеличивается содержание инозилтрифосфата
Гладкая мышечная ткань обладает следующими морфофункциональными
особенностями:
а) непроизвольная мышечная ткань
б) находится под контролем вегетативной нервной системы
в) формирует мышечную оболочку полых и трубчатых органов
г) способна к регенерации
д) сократительная активность не зависит от гормональных влияний
Поперечнополосатая мышца характеризуется следующими признаками:
а) наличием прослоек соединительной ткани между мышечными пучками
б) способностью к произвольному сокращению
в) наличием моторных бляшек и нервно-мышечных веретен
г) наличием клеток-сателлитов
д) клеточным строением
studfiles.net
Мышечные ткани
мт№1
Морфо-функциональная характеристика и классификация мышечных тканей. Гладкая мышечная ткань: источник развития, строение, иннерва5ция. Структурные основы сокращения гладких мышечных клеток. Регенерация.
КЛАССИФИКАЦИЯ: в зависимости от структуры органоидов сокращения делят на:
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ:
НЕЙРАЛЬНОГО ПРОИСХОЖДЕНИЯ
ЭПИДЕРМАЛЬНЫЕ – клетки имеют звездчатую форму, называются миоэпителиоциты (корзинчатые клетки). Своими отростками охватывают концевые отделы желез. При сокращении способствуют выведению секрета.
МЕЗЕНХИМНЫЕ – образуют сократительный аппарат всех внутренних органов. Структурно-функциональной единицей является гладкая мышечная клетка. Имеет веретеновидную форму. L до 200 мкм (в матке до 500). На концах клеток – пальцевидные впячивания. На боковых поверхностях – десмосомы, встречаются нексусы. Основную цементирующую роль играет межклеточное вещество, синтезированное самими гладкомышечными клетками. Поверхность клеток неровная, имеются пузырьковидные впячивания – кавиолы (содержат кальций). В молодых клетках хорошо развита гранулярная ЭПС (синтез межклеточного вещества), ядро овальной формы в центре клетки.
СОКРАТИТЕЛЬНЫЙ АППАРАТ – представлен актиновыми и миозиновыми миофиламентами. Актиновые располагаются продольно или под углом. Образуют трехмерную сеть. В месте их контакта др с др и с цитолеммой образуются электронно плотные тельца, состоящие из α-актина. Миозин в виде мономеров находится между актиновыми фибриллами. Под воздейтвием ПД происходит высвобождение кальция из кавеол и полимеризация миозина. Происходит смещение актиновых нитей относительно миозиновых, благодаря этому меняется форма клетки. Цитоскелет в гладких мышечных клетках развит хорошо, образован промежуточными филаментами – десминами.
РЕГЕНЕРАЦИЯ: кроме внутриклеточной регенерации клетки способны к пролиферации. Кроме этого миофибробласты способны дифференцироваться в миоциты.
МТ №2
Морфо-функциональная характеристика и классификация мышечных тканей. Исчерченная скелетная мышечная ткань: источник развития, строение, иннервация. Структурные основы сокращения мышечного волокна. Типы мышечных волокон. Регенерация.
КЛАССИФИКАЦИЯ: в зависимости от структуры органоидов сокращения делят на:
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ: развивается из мезенхимы, которая выселяется из миотома сомитов→миобласты (активно делятся и накапливают актин и миозин), сливаются→миотубы (ядра лежат в центре, а по периферии миофибриллы)→мышечное волокно (возрастает количество миофибрилл, они занимают центральное положение, а ядра смещаются на периферию).
Основная сруктурно-функциональная единица – мышечное волокно (симпласт), достигает длинны до 12 см, содержит до нескольких десятков тысяч ядер. Выделяют две части: симпласт и миосателит. Между волокнами проходят прослойки соединительной ткани – эндомизий, группы волокон окружены перимизием, снаружи эпимизий. Снаружи волокно покрыто базальной мембраной, которая окружает миосимпласт и миосателитоцит. Собственно миосимпласт покрыт плазмолеммой. Между ними лежат сателлиты. Базальная мембрана + плазмолемма = сарколемма. Содержит белок миоглобин.
По количеству белка волокна делятся на: белые быстрые, красные медленные, промежуточные
Мышечные волокна: ядра по периферии, миофибриллы в центре, слабо развиты КГ и рибосомы, много митохондрий и гладкой ЭПС, которые образуют L-каналы (депо кальция). T–каналы – впячивания плазмолеммы
Сократительный аппарат: представлен миофибриллами: светлые (изотропные) диски, темные (анизотропные) диски. Темные – обладают двойным лучепреломлением. Светлые – состоят в основном из актина, посередине Z-линия (образована α-актином). Темный диск – в основном миозин, есть актин, посередине M-линия (образована миомизином). Структурно-функциональная единица миофибрилл – саркомер – участок между двумя Z-дисками.
Титин – фиксирует миозин к Z-линиям
Фибриллярный актин – двунитчатая спираль.
Тропомиозин – располагается в желобках двунитчатой актиновой спирали (в покоящейся мышце закрывает активные центры в молекуле актина)
Тропонин – состоит из 3 субъединиц: 1 – связана с актином, 2 – с тропомиозином, 3 – с ионами кальция
Небулин – фибриллярный белок, связанный с тонкими нитями. Проходит от Z-линий до свободного конца тонких нитей и контролирует их длину.
Формула саркомера: Z+1/2 I+1/2A+M+1/2A+1/2I+Z
I – светлый диск, A – темный
На поперечном срезе соотношение тонких и толстых нитей 2:1 Сокращение построено на принципе скольжения нитей относительно др др. При обычных условиях саркомер укорачивается на 20%.
РЕГЕНЕРАЦИЯ: протекает активно за счет миосателитоцитов
ИННЕРВАЦИЯ: двигательные, чувствительные и вегетативные волокна, отросток нервной клетки ветвится в перимизии, его ветви на поверхности симпласта (плазмолемме) образуют терминали, участвуя в организации моторной бляшки. Ю Выделяется АХ→возбуждение.
МТ №3
Морфо- функциональная характеристика и классификация мышечных тканей. Источники развития. Мышца как орган: строение, васкуляризация, эфферентная и афферентная иннервация. Связь мышцы с сухожилием.
КЛАССИФИКАЦИЯ: в зависимости от структуры органоидов сокращения делят на:
Передача усилий сокращения на скелет осуществляется посредством сухожилия и прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна, спирально оплетенные ретикулярными волокнами. Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани – эндомизий. Группы волокон окружены перимизием. Снаружи – эпимизий (соединительная ткань)
ВАСКУЛЯРИЗАЦИЯ: артерии всупают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви 5-6 порядка в перимизии образуют артериолы. В эндомизии расположены капилляры. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости стенки сосуда.
ИННЕРВАЦИЯ: эфферентная (двигательная) каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапиляров, образуя мионом. Группа мышечных волокон, иннервируемая одним мотонейроном называется нервно-мышечная единица. Чувствительная (афферентная) чувствительные нервные окончания располагаются в мышечных веретенах (интерфузальные мышечные волокна), расположенных в перимизии. Интерфузальные мышечные волокна – два вида: волокна с ядерной сумкой, волокна с ядерной цепочкой. Ядра округлые, расположены в толще симпласта. В волокнах с ядерной сумкой ядра образуют скопление в его утолщенной средней части. В волокнах с ядерной цепочкой утолщение не образуется. Ядра лежат продольно, одно за другим. Рядом со скоплениями ядер располагаются органеллы общего значения. Миофибриллы находятся в концах симпластов. Сарколемма соединяется с капсулой нервно-мышечного веретена. Интерфузальные мышечные волокна постоянно находятся в напряжении.
МТ №4
Морфо-функциональная характеристика и классификация мышечных тканей. Исчерченная сердечная мышечная ткань: источник развития, структурно-функциональная характеристика. Регенерация.
КЛАССИФИКАЦИЯ: в зависимости от структуры органоидов сокращения делят на:
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ. Встречается в стенке сердца, проксимальной части аорты, в верхней полой вене.
Структурная единица – КМЦ.
3 популяции КМЦ: сократительные, проводящие и секреторные
СОКРАТИТЕЛЬНЫЕ КМЦ: призматической Фомы, могут ветвиться, образуя сеть. 1-2 ядра в центре к-ки. Объединяются в волокна с помощью вставочных дисков (плазмолеммы соседних к-к). Выпячивания (интердигитации) соединены межклеточными контактами (щелевые, десмосомы,fascia adherens – миофибрилла не прерываясь продолжается в соседнюю к-ку). В цитоплазме КМЦ много гликогена и липидов, из органоидов развиты митохондрии Есть анастомозы(мостики), которые перекидываются от одного волокна к другому. Особенность работы : кальций поступает извне к-ки.
2 группы КМЦ: — предсердные – мелкие, слабо развиты Т-трубочки.
— желудочковые – крупнее, развита Т – система
ПРОВОДЯЩИЕ: — водители ритма (пейсмейкерные к-ки) – небольшого размера, мало гликогена, миофибриллы по периферии. Ф-ия – генерация нервного импульса
— проводящие – проводят импульсы от узла к миокарду
Пучок Гиса – к-ки содержат длинные миофибриллы и мелкие митохондрии, мало гликогена.
Волокна Пуркинье – содержат самые крупные к-ки, в которых редкая неупорядоченная сеть миофибрилл, много мелких митохондрий, гликогена, нет Т-трубочек.
РЕГЕНЕРАЦИЯ: при усиленной работе происходит рабочая гипертрофия КМЦ. СК или к-ок предшественников нет→ не восстанавливаются.
studfiles.net