
Роль ассоциативных зон коры в формировании замысла движений: Repository of Kharkiv National Medical University: Invalid Identifier
Возрастные изменения в системе регуляции движений — Возрастная физиология
Возрастные изменения в системе регуляции движений
Возрастные изменения включенности в деятельность и роли различных структур системы регуляции движений определяются постепенным и гетерохронным созреванием всех ее компонентов.
К моменту рождения дифференцированы все корковые зоны, но наиболее развиты проекционные области. В этом возрасте в общих чертах завершается дифференцировка подкорковых структур, ядерных образований и проводящих путей спинного мозга (за исключением пирамидного тракта). Сложное строение имеют мышечные, суставные и сухожильные рецепторы.
Среди анализаторных систем
головного мозга, имеющих непосредственное отношение к регуляции движений,
более зрелой и готовой к функционированию является система кожного и двигательного
анализаторов. Раннее созревание этих систем позволяет наблюдать и вызывать у
новорожденных целый комплекс общих и локальных двигательных реакций,
обеспечивающих функции питания, защиты и т.
В первые 3—4 мес жизни ребенка постепенно усиливается участие коры больших полушарий в регуляции движений. Идет созревание мозжечка, полосатого тела и других структур мозга, что способствует снижению общего тонического напряжения мышц, установлению баланса активности мышц верхних конечностей и увеличению амплитуды движений рук.
Первые условные рефлексы (пассивное разгибание ножки ребенка в коленном суставе) появляются на 3—4-й неделе жизни. Эти рефлексы также приобретают относительное постоянство к 3—4-му месяцу жизни.
Во втором полугодии жизни продолжается созревание всех структур системы регуляции движений.
Структурное созревание коры больших полушарий и
интра-кортикальных путей в постнатальный период создает широкие возможности для установления межанализаторных
отношений и участия многих отделов головного мозга в регуляции движений.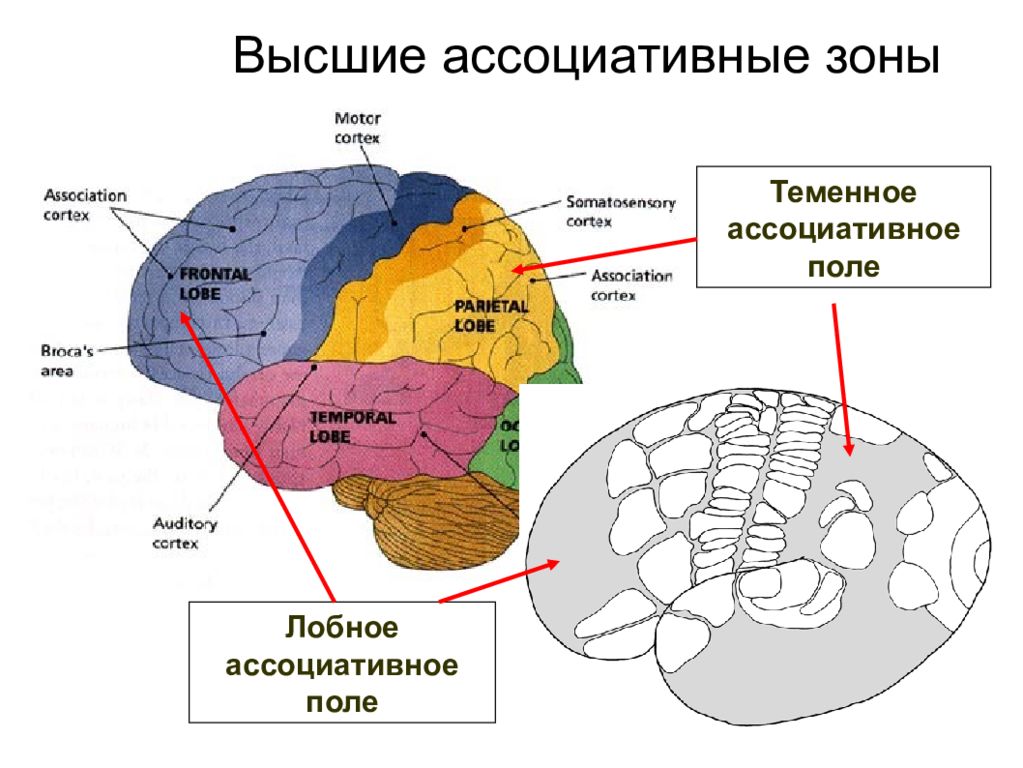
По мере развития все большее значение в процессах формирования сложных реакций, обеспечивающих адекватность взаимодействия организма с окружающим миром, приобретают центральные отделы двигательной системы. Электрофизиологические исследования показывают, что нейроны сенсомоторной области коры начинают проявлять способность к межсенсорной конвергенции уже на ранней стадии постнатального онтогенеза.
Во втором полугодии жизни ребенка межапализаторные функциональные связи становятся более упорядоченными и эффективными.
В этот период появляются
направленные движения руки к определенному предмету, его захват и удержание.
Повышается число движений, когда кисть руки раскрывается до захватывания предмета. Идет перестройка петлеобразных хватательных движений с частыми промахами и
превращение их в движения с «прямым плановым» приближением к предмету. С 10-го
месяца жизни наблюдается предварительное приспособление пальцев руки к форме
объекта, который ребенок намеревается схватить, т.е. отмечаются элементы
специфической готовности к движению, а значит формируется «замысел действия». В
этот же период отмечаются хватательные движения вслепую за счет
предварительного нацеливания на предмет. Возникновение в течение второго
полугодия жизни подобных усложненных форм двигательной активности, по-видимому,
свидетельствует о начальных этапах становления способности центральной нервной системы
к программированию движения.
Идет перестройка петлеобразных хватательных движений с частыми промахами и
превращение их в движения с «прямым плановым» приближением к предмету. С 10-го
месяца жизни наблюдается предварительное приспособление пальцев руки к форме
объекта, который ребенок намеревается схватить, т.е. отмечаются элементы
специфической готовности к движению, а значит формируется «замысел действия». В
этот же период отмечаются хватательные движения вслепую за счет
предварительного нацеливания на предмет. Возникновение в течение второго
полугодия жизни подобных усложненных форм двигательной активности, по-видимому,
свидетельствует о начальных этапах становления способности центральной нервной системы
к программированию движения.
В этом же возрасте (7—10 мес)
обнаруживается способность детей совершать (при их пассивной полдержке под
руки) попеременные шагательныс движения по движущемуся тредбану. Структура
этих движений, включая закономерности изменений суставных углов,
межконечностную и межсуставную координацию, сравнительно совершенна.
Фактически в течение второго полугодия происходит постепенное включение в систему регуляции движений всех компонентов структуры от ассоциативных зон коры до м ото нейронов спинного мозга. Это обеспечивает совершенствование движений рук, йог, туловища ребенка, однако практически все движения еще нс очень точны, так как не сформирован батане мышц-антагонистов, несовершенна нервно-мышечная регуляция, недостаточно сформированы механизмы поддержания позы.
Именно с этого возраста начинается активное формирование нового типа движений — произвольных, целенаправленных.
Изменение роли отдельных
структур системы регуляции движений в процессе возрастного развития отчетливо
видно на примере формирования сложных произвольных движений, таких как ходьба
и письмо.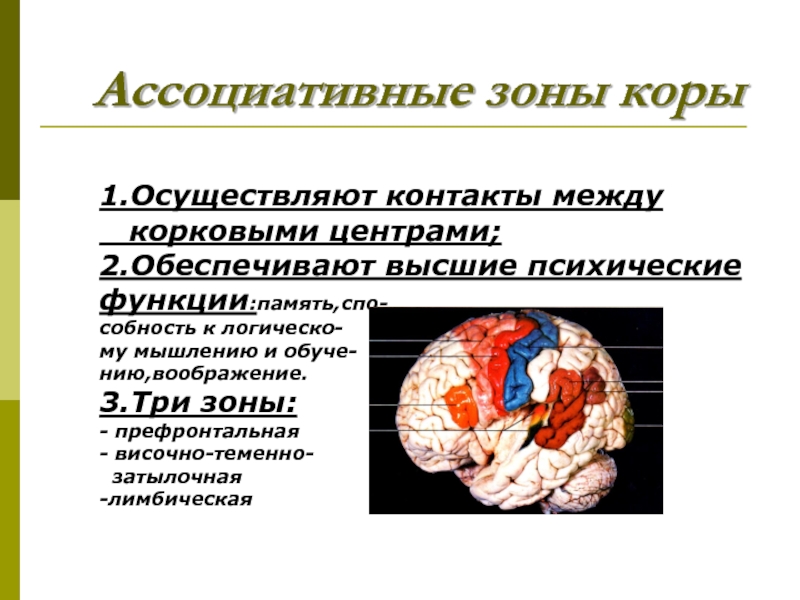
Ребенок начинает ходить в возрасте 9—15 мес. Однако вплоть до 4 лет формируется временная структура акта ходьбы. К 4 годам сформированной оказывается лишь структура каждого отдельного шага, а система шагов, т.е. структура серии движений при ходьбе, еще не сформирована, движения неритмичны, нестабильны, вариативны. Фактически еще не сложилась программа серии движений, каждый шаг корригируется по ходу деятельности.
Следующий этап формирования ходьбы — 4—7 лет. В этом
возрасте заметно расширяются связи двигательной области головного мозга с
одним из важных центров регуляции движений — мозжечком и подкорковыми
образованиями, в частности с красным ядром, т.е. созревают механизмы регуляции
позы и механизмы реализации последовательности серийных движений.
В этом же возрасте продолжается совершенствование серии отдельных шагов при ходьбе, но они нестабильны, вариативны, еще отсутствует зависимость между темпом ходьбы и длиной шага.
Новым этаном в формировании акта ходьбы является возраст 9—10 лет. В этом возрасте показатели структуры шага и ходьбы близки к показателям взрослых. Темп ходьбы равномерный, дайна шага стабильна, ускорение темпа связано с удлинением шага. В этом возрасте структура регуляции движений устойчива к влиянию дополнительных нагрузок и различных «шумов», мешающих реализации этого движения. Значит, мы можем говорить о том, что при выполнении движения реализуются все функции системы регуляции:
принятие решения
і
выбор двигательной программы і
достижение «полезного результата» или решение «задачи
действия».
Таким образом, только к 10 голам реализация движения (ходьбы) обеспечивается включенностью и взаимодействием всех звеньев системы регуляции движения, а формирование этого движения идет почти 10 лет.
Графические движения проходят те же этапы формирования, что и ходьба: в 1 — 1,5 года — это нестабильные, плохо регулируемые движения, от 2 до 4 лет формируется структура графических движений, к 5 годам сформированы основные компоненты графических движений — вертикальные и горизонтальные линии, овалы, круги, но они еще нестабильны, плохо регулируются их размерность, соотношение элементов, еще недостаточно четки линии, много дополнительных штрихов, которые свидетельствуют о корректировке по ходу движения, а значит, о том, что программа движения еще не сформирована.
В 6—7 лет начинается собственно формирование навыка
письма, а все графические движения и действия, которые ребенок освоил до этого
возраста, служат базисом для их развития.
Основные характеристики письма у детей этого возраста: нестабильность, неровность, нечеткость штрихов, сильное мышечное напряжение (часты жалобы детей — «болит рука», «устала рука»), несовершенны механизмы регуляции позы (ребенок очень быстро нарушает правильную позу при письме), фактически каждое движение в серии выполняется отдельно, нет плавности и связности движений. Это тот этап, когда ребенок еше не может произвольно менять скорость письма, сохраняя качество (движения могут быть медленными и правильными или быстрыми, но неправильными). Все это свидетельствует о том, что моторная программа еще только формируется и необходима постоянная коррекция и оценка по ходу движения. Так же как и при формировании акта ходьбы, в 9—10 лет при письме существенно изменяются центральные механизмы регуляции движений.
На этом этапе уже сформирована и совершенствуется
моторная программа, резко сокращается пауза между отдельными двигательными
актами, движения становятся четкими, стабильными, менее напряженными, ребенок
способен произвольно регулировать основные параметры движения, варьировать
темп, скорость, качество серии движений, т. е. показатели временной структуры
письма близки к показателям взрослых.
е. показатели временной структуры
письма близки к показателям взрослых.
Одним из факторов совершенствования центральных механизмов регуляции движений к 9—10 годам, по-видимому, является созревание лобных зон коры, которым отводится роль принятия решения и выбора программы действия.
Нельзя не отметить, что в период с И —12 до 14—15 лет ухудшается синхронность движений в суставах, повышается мышечное напряжение, замедляется развитие функций управления пространственно-временными параметрами точностных действий и снижаются резервные возможности совершенствования движений под влиянием упражнения. Все это, по-видимому, связано с существенным изменением механизмов центральной регуляции движений, ослаблением роли регуляторных влияний лобной коры в период полового созревания.
Интересно, что, по данным нейрофизиологических
исследований, регрессивные изменения активности коры, повышение активности
подкорковых структур, менее дифференцированное вовлечение структур неокортскса
в процессы организации деятельности характерны для подростков на II и III
стадиях полового созревания. Это свидетельствует об определенных регрессивных
изменениях ряда функций центральной нервной системы в период полового
созревания, затрагивающих не только сенсорные процессы восприятия, организации
внимания, но и центрально-эф-фекторные механизмы регуляции моторикой, и
процессы организации двигательной деятельности.
Это свидетельствует об определенных регрессивных
изменениях ряда функций центральной нервной системы в период полового
созревания, затрагивающих не только сенсорные процессы восприятия, организации
внимания, но и центрально-эф-фекторные механизмы регуляции моторикой, и
процессы организации двигательной деятельности.
Но уже после 14—15 лет эти явления становятся менее заметными и исчезают, совершенствуется регуляция сложнокоордини-рованных движений, усиливается их интеграция и в то же время автономность, возрастают точность управления пространственными параметрами движений, улучшается качество выполнения двигательных действий при разных скоростных режимах, вновь повышается чувствительность двигательных функций к тренировочным воздействиям.
Таким образом, возрастное
развитие движений — это путь от недифференцированных, нестабильных, напряженных
отдельных движений, очень зависящих от влияния различных факторов (внешних —
условий выполнения и внутренних — состояния организма), к более
дифференцированным и интегрированным в серии, плавным, точным, стабильным и
менее зависимым от влияния различных факторов.
Вопросы и задания
1. Какие движения называются произвольными, непроизвольными? В чем их различие?
2. Какие основные структуры включены в систему организации и регуляции движения? Нарисуйте схему, покажите пути нервных импульсов от коры к мышцам.
3. Что означает соматотопический принцип организации двигательных зон коры? Какая часть тела в двигательной коре наиболее широко представлена? Как вы думаете, почему?
4. Каковы функции двигательной коры в организации и регуляции движений?
5. Каковы функции базальных ганглиев в организации и регуляции движений?
6. Каковы функции мозжечка в организации и регуляции движений?
7. Выделите функции ствола мозга в организации и регуляции движений.
8. Выделите функции спинного мозга в организации и регуляции движений.
9. Назовите методы изучения организации и регуляции движений. Какой уровень регуляции движений позволяет изучать каждый из перечисленных методов?
10. Опишите возрастные изменения в структуре
организации и регуляции движений. Как изменяются при этом характер движений,
их качество и эффективность?
Опишите возрастные изменения в структуре
организации и регуляции движений. Как изменяются при этом характер движений,
их качество и эффективность?
79
Двигательные программы — PDF Free Download
Физиология поведения
Частное учреждение образования «Минский институт управления» Физиология поведения Курс лекций 1-23.01.04 — ПСИХОЛОГИЯ 4 издание переработанное и дополненное Минск Изд-во МИУ 2013 Лекция 22 Управление движениями
ПодробнееСПИННОЙ МОЗГ. СТРОЕНИЕ
СПИННОЙ МОЗГ. СТРОЕНИЕ Спинной мозг лежит в позвоночном канале представляет собой длинный тяж (его длина у взрослого человека около 45 см), несколько сплющенный спереди назад. Вверху он переходит в продолговатый
СТРОЕНИЕ Спинной мозг лежит в позвоночном канале представляет собой длинный тяж (его длина у взрослого человека около 45 см), несколько сплющенный спереди назад. Вверху он переходит в продолговатый
Алексеева Елена Владимировна
Алексеева Елена Владимировна «Собраться вместе это начало. Держаться вместе это прогресс. Работать вместе это успех» Генри Форд Нервная система это совокупность специальных структур. Объединяющая и координирующая
ПодробнееТема: Рефлекс. Рефлекторная дуга
Глава II. Нервно-гуморальная регуляция физиологических функций На дом: 8 Тема: Рефлекс. Рефлекторная дуга Задачи: Дать характеристику рефлексам, рефлекторным дугам Пименов А.В. Рефлексом называется ответная
ПодробнееФункции нервной системы
Нервная система Нервная система Это совокупность специальных структур, объединяющая и координирующая деятельность всех органов и систем организма в постоянном взаимодействии с внешней средой Функции нервной
ПодробнееИППОКРАТ Т.
 А.,КУВАЕВ
А.,КУВАЕВИППОКРАТ Т.А.,КУВАЕВ Т. В. Алейникова, В. Н. Думбай, Г. А. Кураев, Г. Л. Фельдман ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ Учебное пособие Издание второе, дополненное и исправленное Научный редактор доктор
ПодробнееЧувствительные проводящие пути
Проводящие пути Чувствительные проводящие пути Сознательная проприоцептивная чувствительность Сознательное положение тела и его частей в пространстве Чувство стереогноза узнавание предмета на ощупь Нарушения
ПодробнееНЕВРОЛОГИЯ ПРОВОДЯЩИЕ ПУТИ МОЗГА
НЕВРОЛОГИЯ ПРОВОДЯЩИЕ ПУТИ МОЗГА Типы проводящих путей Проводящие пути пучки нервных волокон, содержащие функционально однородные участки серого вещества в ЦНС, занимающие в белом веществе головного и
ПодробнееМОЗЖЕЧКОВАЯ СТИМУЛЯЦИЯ КАК
МОЗЖЕЧКОВАЯ СТИМУЛЯЦИЯ КАК СОСТАВНАЯ ЧАСТЬ НЕЙРОПСИХОЛОГИЧЕСКОЙ КОРРЕКЦИИ В ДЕТСКОМ ВОЗРАСТЕ ОТДЕЛЕНИЕ МЕДИКО-СОЦИАЛЬНОЙ РЕАБИЛИТАЦИИ ИНДУСТРИАЛЬНОГО Р-НА Г. ПЕРМИ Клинический психолог, детский нейропсихолог
ПЕРМИ Клинический психолог, детский нейропсихолог
ФИЗИОЛОГИЯ СПИННОГО МОЗГА
ФИЗИОЛОГИЯ СПИННОГО МОЗГА 1. Функциональная организация спинного мозга 2. Проводниковые функции спинного мозга 3. Рефлексы спинного мозга Вопрос_1 Функциональная организация спинного мозга В строении спинного
ПодробнееОБЩАЯ НЕЙРОФИЗИОЛОГИЯ
Учебное пособие написано в соответствии с требованиями государственного образовательного стандарта к курсу физиологии центральной нервной системы по специальности 020400 «Психология». В нем рассматриваются
ПодробнееФИЗИОЛОГИЯ СТРУКТУР СТВОЛА МОЗГА
ФИЗИОЛОГИЯ СТРУКТУР СТВОЛА МОЗГА 1. Функции продолговатого мозга 2. Функции моста заднего мозга 3. Функции среднего мозга Вопрос_1 Функции продолговатого мозга Продолговатый мозг является частью головного
ПодробнееНервные узлы.
 Спинной мозг.
Спинной мозг.Модуль IV «Частная гистология (часть первая)». Нервные узлы. Спинной мозг. доцент кафедры гистологии, цитологии и эмбриологии, к.б.н. Е.В. Блинова г. Оренбург, 2016 Функции нервной системы 1. Обеспечение
ПодробнееНервная система Спинной мозг.
Нервная система Функции нервной системы. Особо важную роль в жизнедеятельности организма человека играет нервная система совокупность различных структур нервной ткани. Функциями нервной системы являются:
ПодробнееПРИМЕР РАБОТЫ СО СХЕМАМИ И ДЕФИНИЦИЯМИ
ПРИМЕР РАБОТЫ СО СХЕМАМИ И ДЕФИНИЦИЯМИ 1.1. ОТРАСЛИ ПСИХОЛОГИИ Психология труда Социальная психология Возрастная психология Юридическая психология Медицинская психология Ощущения Восприятие Память Мышление
ПодробнееЧАСТНАЯ ГИСТОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Профессор С. Ю.Виноградов ЧАСТНАЯ ГИСТОЛОГИЯ НЕРВНОЙ СИСТЕМЫ (факультативная лекция для студентов стоматологического факультета) Частная гистология наука о микроскопическом строении, развитии и функциях
Ю.Виноградов ЧАСТНАЯ ГИСТОЛОГИЯ НЕРВНОЙ СИСТЕМЫ (факультативная лекция для студентов стоматологического факультета) Частная гистология наука о микроскопическом строении, развитии и функциях
Задачи для экзамена по физиологии.
Задачи для экзамена по физиологии. Порог раздражения электрическим током у одной мышцы 2в, у другой 3в. У какой из мышц возбудимость выше? Известно, что возбуждение нерва или мышцы можно вызвать применяя
Подробнее1.7. Анализаторы человека
1 1.7. Анализаторы человека 1.7.1. Устройство анализатора. Зрительный анализатор Изменение условий окружающей среды и состояние внутренней среды человека воспринимается нервной системой, которая регулирует
ПодробнееНоцицепция. а2-рецепторы
Конференция и круглый стол по анестезиологии собак и кошек Ноцицепция.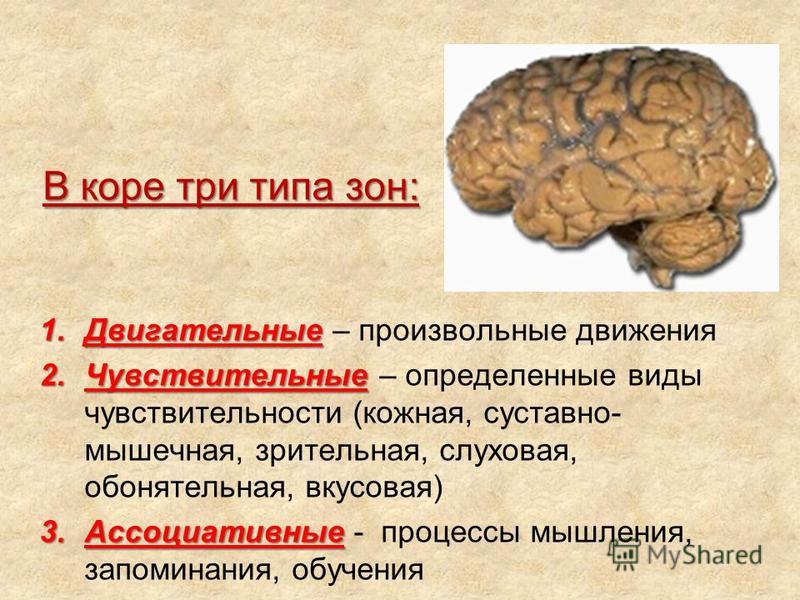 а2-рецепторы Юрий Дородных Клиника «Белый клык» 8 декабря 2016 г., Москва. Понятие «боль» Боль — неприятное сенсорное и эмоциональное
а2-рецепторы Юрий Дородных Клиника «Белый клык» 8 декабря 2016 г., Москва. Понятие «боль» Боль — неприятное сенсорное и эмоциональное
ОБЩАЯ ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
Лекция 1 ОБЩАЯ ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ Объективная и субъективная сторона восприятия Специфичность сенсорных систем Закон специфических энергий Строение сенсорной системы Принципы организации сенсорных
ПодробнееЭкстрапирамидные нарушения движений
Экстрапирамидные нарушения движений Двигательный акт формируется в результате последовательного, согласованного по силе и длительности включения отдельных нейронов корково- мышечного пути и большого комплекса
ПодробнееВопрос 1. Виды памяти
Тема: Физиологические основы психических процессов. Вопрос: Психофизиология памяти Вопросы: 1. Виды памяти. 2. Временная организация памяти и механизмы запечатления. 3. Теория кратковременной и долговременной
Виды памяти. 2. Временная организация памяти и механизмы запечатления. 3. Теория кратковременной и долговременной
ÀÍÀÒÎÌÈß ÖÅÍÒÐÀËÜÍÎÉ ÍÅÐÂÍÎÉ ÑÈÑÒÅÌÛ
Ñ. Þ. Êèñåëåâ ÀÍÀÒÎÌÈß ÖÅÍÒÐÀËÜÍÎÉ ÍÅÐÂÍÎÉ ÑÈÑÒÅÌÛ Ó ÅÁÍÎÅ ÏÎÑÎÁÈÅ ÄËß ÂÓÇÎÂ Ðåêîìåíäîâàíî ìåòîäè åñêèì ñîâåòîì ÓðÔÓ â êà åñòâå ó åáíî-ìåòîäè åñêîãî ïîñîáèÿ äëÿ ñòóäåíòîâ, îáó àþùèõñÿ ïî ïðîãðàììàì áàêàëàâðèàòà
ПодробнееОБЩИЙ ПЛАН СТРОЕНИЯ МОЗГА
ОБЩИЙ ПЛАН СТРОЕНИЯ МОЗГА Усложнение структуры нервной системы беспозвоночные позвоночные Макроанатомический уровень организации нервной системы: Ядра Слои Тракты Заварзин А.А. Ядерные центры: скопление
Подробнее4. ДВИГАТЕЛЬНЫЕ ПРОГРАММЫ. Основы психофизиологии
4. ДВИГАТЕЛЬНЫЕ ПРОГРАММЫ
Управление движениями немыслимо без согласования активности большого количества мышц. Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан воды, то ЦНС должна располагать информацией о положении стакана относительно тела и об исходном положении руки. Однако чтобы движение было успешным, кисть заранее раскрылась на величину, соответствующую размеру стакана, чтобы сгибатели пальцев сжимали стакан с силой, достаточной для предотвращения проскальзывания, чтобы приложенная сила была достататочной для плавного подъёма, но не вызывала резкого отрыва, чтобы ориентация стакана в кисти после захвата всё время была вертикальной. Таким образом, чтобы реализация движения соответствовала двигательной задаче, необходимы не только данные о пространственных соотношениях, но и сведения о свойствах объекта манипулирования. Многие из этих сведений не могут быть получены в ходе самого движения посредством обратных связей, а должны быть предусмотрены на этапе планирования. Следовательно, для осуществления движения должна быть сформирована двигательная программа.
Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан воды, то ЦНС должна располагать информацией о положении стакана относительно тела и об исходном положении руки. Однако чтобы движение было успешным, кисть заранее раскрылась на величину, соответствующую размеру стакана, чтобы сгибатели пальцев сжимали стакан с силой, достаточной для предотвращения проскальзывания, чтобы приложенная сила была достататочной для плавного подъёма, но не вызывала резкого отрыва, чтобы ориентация стакана в кисти после захвата всё время была вертикальной. Таким образом, чтобы реализация движения соответствовала двигательной задаче, необходимы не только данные о пространственных соотношениях, но и сведения о свойствах объекта манипулирования. Многие из этих сведений не могут быть получены в ходе самого движения посредством обратных связей, а должны быть предусмотрены на этапе планирования. Следовательно, для осуществления движения должна быть сформирована двигательная программа. Двигательную или центральную программу рассматривают как заготовленный набор базовых двигательных команд, а также набор готовых корректирующих подпрограмм, обеспечивающих реализацию движения с учётом текущих афферентных сигналов и информации, поступающей от других частей ЦНС.
Двигательную или центральную программу рассматривают как заготовленный набор базовых двигательных команд, а также набор готовых корректирующих подпрограмм, обеспечивающих реализацию движения с учётом текущих афферентных сигналов и информации, поступающей от других частей ЦНС.
Зарождение побуждения к движению связано с активностью подкорковых и корковых мотивационных зон. Замысел движения формируется в ассоциативных зонах коры. Далее происходит формирование программы движения с участием базальных ганглиев и мозжечка, действующих на двигательную кору через ядра таламуса. За реализацию программы отвечает двигательная кора и нижележащие стволовые и спинальные двигательные центры.
Предполагается, что двигательная память содержит обобщённые классы двигательных программ, из числа которых в соответствии с двигательной задачей выбирается нужная. Программа модифицируется применительно к ситуации: однотипные движения могут выполняться быстрее или медленнее, с большей или меньшей амплитудой. Интересно, что одна и та же программа может быть реализована разными наборами мышц. Так, почерк человека сохраняет характерные черты при письме правой и левой рукой и даже карандашом, зажатым в зубах или прикреплённым к носку ботинка. Такой межконечностный перенос навыка возможен потому, что система управления движениями является многоуровневой (уровень планирования движения и уровень его исполнения в ней не совпадают). Действительно, произвольное движение планируется в терминах трёхмерного евклидового пространства. Для исполнения этого плана необходимо перевести линейные перемещения в соответствующие угловые переменные (изменения суставных углов), определить, какие мышечные моменты необходимы для этих угловых перемещений и, наконец, сформировать двигательные команды, которые вызовут активацию мышц, дающую необходимые значения моментов.
Интересно, что одна и та же программа может быть реализована разными наборами мышц. Так, почерк человека сохраняет характерные черты при письме правой и левой рукой и даже карандашом, зажатым в зубах или прикреплённым к носку ботинка. Такой межконечностный перенос навыка возможен потому, что система управления движениями является многоуровневой (уровень планирования движения и уровень его исполнения в ней не совпадают). Действительно, произвольное движение планируется в терминах трёхмерного евклидового пространства. Для исполнения этого плана необходимо перевести линейные перемещения в соответствующие угловые переменные (изменения суставных углов), определить, какие мышечные моменты необходимы для этих угловых перемещений и, наконец, сформировать двигательные команды, которые вызовут активацию мышц, дающую необходимые значения моментов.
Двигательная программа может быть реализована различными способами. В простейшем случае ЦНС посылает к мышцам заранее сформированную последовательность команд, не подвергающуюся во время реализации никакой коррекции. В этом случае говорят о разомкнутой системе управления. Подобное управление используется при осуществлении быстрых, так называемых «баллистических» движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов от многочисленных рецепторов, и в реализуемую программу вносятся нужные коррекции. Это замкнутая система управления с обратными связями. Однако и такое управление имеет недостатки. В связи с относительно малыми скоростями проведения сигналов, значительными задержками в центральном звене обратной связи и значительным временем, необходимым для развития усилия мышцей, коррекция движения по сигналу обратной связи может запаздывать. Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение ещё до того, как оно успело вызвать отклонение. Такое управление называют управлением по возмущению.
В этом случае говорят о разомкнутой системе управления. Подобное управление используется при осуществлении быстрых, так называемых «баллистических» движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов от многочисленных рецепторов, и в реализуемую программу вносятся нужные коррекции. Это замкнутая система управления с обратными связями. Однако и такое управление имеет недостатки. В связи с относительно малыми скоростями проведения сигналов, значительными задержками в центральном звене обратной связи и значительным временем, необходимым для развития усилия мышцей, коррекция движения по сигналу обратной связи может запаздывать. Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение ещё до того, как оно успело вызвать отклонение. Такое управление называют управлением по возмущению.
Другим способом уменьшения влияния задержек является антиципация. Во многих случаях ЦНС способна предусмотреть в двигательной программе появление возмущений ещё до их возникновения.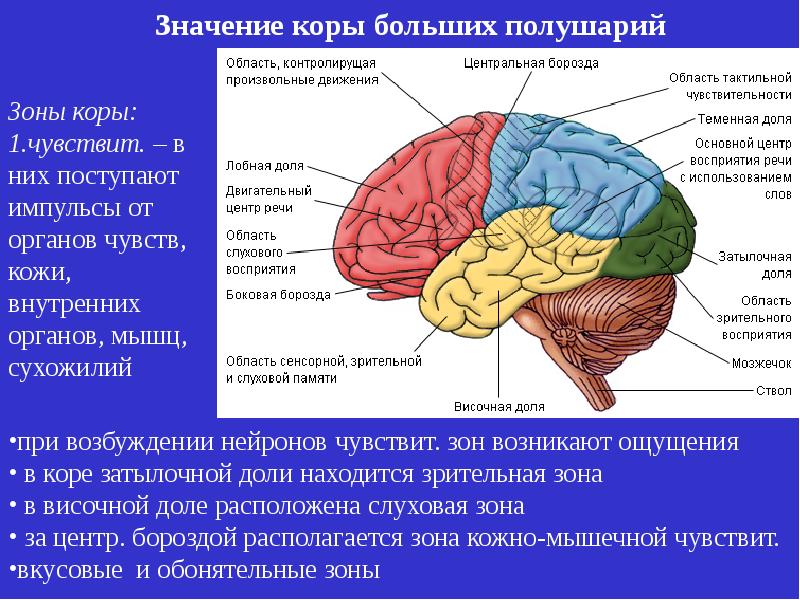 Примечательно, что эта упреждающая «позная» активность (антиципация) осуществляется автоматически с очень короткими центральными задержками. Роль упреждающей активности в стабилизации положения звеньев тела иллюстрирует простой пример. Если официант удерживает на ладони вытянутой руки поднос с бутылкой шампанского и рюмками, а другой человек внезапно снимет бутылку с подноса, то рука резко подпрыгнет вверх с соответствующими последствиями. Если же он сам снимет бутылку свободной рукой, то рука с подносом останется на прежнем уровне.
Примечательно, что эта упреждающая «позная» активность (антиципация) осуществляется автоматически с очень короткими центральными задержками. Роль упреждающей активности в стабилизации положения звеньев тела иллюстрирует простой пример. Если официант удерживает на ладони вытянутой руки поднос с бутылкой шампанского и рюмками, а другой человек внезапно снимет бутылку с подноса, то рука резко подпрыгнет вверх с соответствующими последствиями. Если же он сам снимет бутылку свободной рукой, то рука с подносом останется на прежнем уровне.
Двигательные программы | Студент-Сервис
Управление движениями немыслимо без согласования активности большого количества мышц.
Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан воды, то ЦНС должна располагать информацией о положении стакана относительно тела и об исходном положении руки.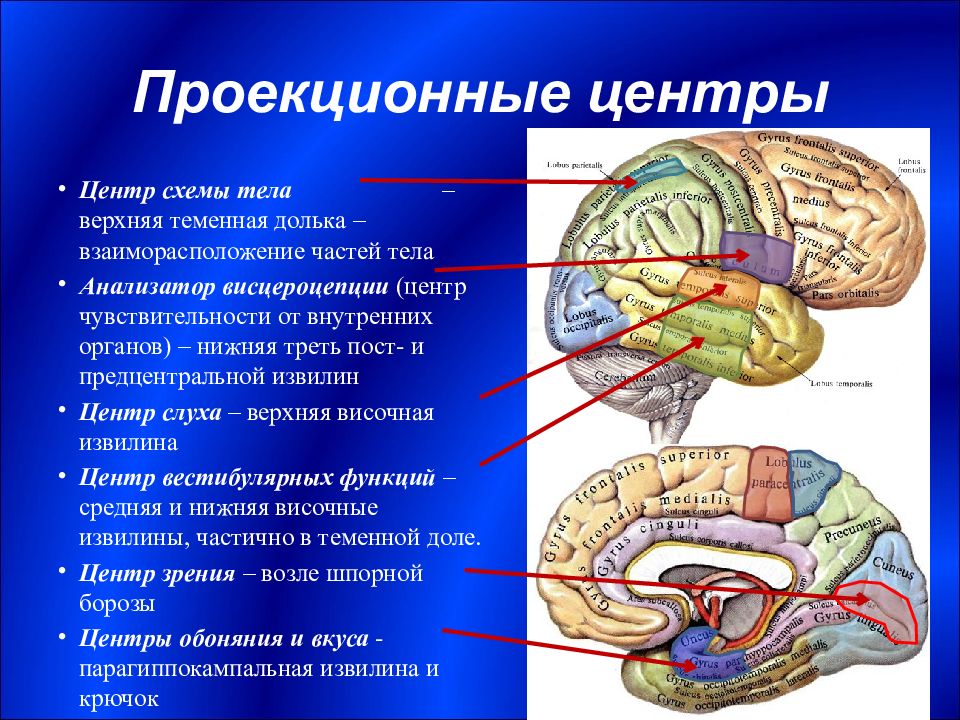
Однако чтобы движение было успешным, кисть заранее раскрылась на величину, соответствующую размеру стакана, чтобы сгибатели пальцев сжимали стакан с силой, достаточной для предотвращения проскальзывания, чтобы приложенная сила была достататочной для плавного подъёма, но не вызывала резкого отрыва, чтобы ориентация стакана в кисти после захвата всё время была вертикальной.
Таким образом, чтобы реализация движения соответствовала двигательной задаче, необходимы не только данные о пространственных соотношениях, но и сведения о свойствах объекта манипулирования.
Многие из этих сведений не могут быть получены в ходе самого движения посредством обратных связей, а должны быть предусмотрены на этапе планирования. Следовательно, для осуществления движения должна быть сформирована двигательная программа.
Двигательную или центральную программу рассматривают как заготовленный набор базовых двигательных команд, а также набор готовых корректирующих подпрограмм, обеспечивающих реализацию движения с учётом текущих афферентных сигналов и информации, поступающей от других частей ЦНС.
Зарождение побуждения к движению связано с активностью подкорковых и корковых мотивационных зон. Замысел движения формируется в ассоциативных зонах коры. Далее происходит формирование программы движения с участием базальных ганглиев и мозжечка, действующих на двигательную кору через ядра таламуса. За реализацию программы отвечает двигательная кора и нижележащие стволовые и спинальные двигательные центры.
Предполагается, что двигательная память содержит обобщённые классы двигательных программ, из числа которых в соответствии с двигательной задачей выбирается нужная. Программа модифицируется применительно к ситуации: однотипные движения могут выполняться быстрее или медленнее, с большей или меньшей амплитудой.
Интересно, что одна и та же программа может быть реализована разными наборами мышц. Так, почерк человека сохраняет характерные черты при письме правой и левой рукой и даже карандашом, зажатым в зубах или прикреплённым к носку ботинка.
Такой межконечностный перенос навыка возможен потому, что система управления движениями является многоуровневой (уровень планирования движения и уровень его исполнения в ней не совпадают).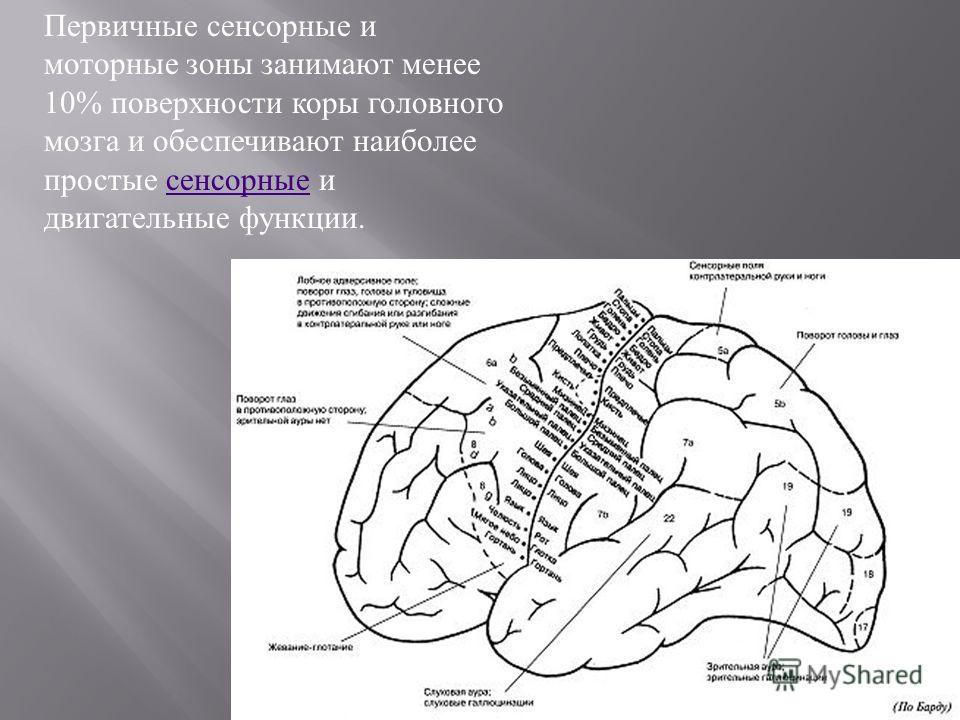 Действительно, произвольное движение планируется в терминах трёхмерного евклидового пространства.
Действительно, произвольное движение планируется в терминах трёхмерного евклидового пространства.
Для исполнения этого плана необходимо перевести линейные перемещения в соответствующие угловые переменные (изменения суставных углов), определить, какие мышечные моменты необходимы для этих угловых перемещений и, наконец, сформировать двигательные команды, которые вызовут активацию мышц, дающую необходимые значения моментов.
Двигательная программа может быть реализована различными способами. В простейшем случае ЦНС посылает к мышцам заранее сформированную последовательность команд, не подвергающуюся во время реализации никакой коррекции. В этом случае говорят о разомкнутой системе управления.
Подобное управление используется при осуществлении быстрых, так называемых «баллистических» движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов от многочисленных рецепторов, и в реализуемую программу вносятся нужные коррекции.
Это замкнутая система управления с обратными связями. Однако и такое управление имеет недостатки. В связи с относительно малыми скоростями проведения сигналов, значительными задержками в центральном звене обратной связи и значительным временем, необходимым для развития усилия мышцей, коррекция движения по сигналу обратной связи может запаздывать.
Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение ещё до того, как оно успело вызвать отклонение. Такое управление называют управлением по возмущению.
Другим способом уменьшения влияния задержек является антиципация. Во многих случаях ЦНС способна предусмотреть в двигательной программе появление возмущений ещё до их возникновения. Примечательно, что эта упреждающая «позная» активность (антиципация) осуществляется автоматически с очень короткими центральными задержками.
Роль упреждающей активности в стабилизации положения звеньев тела иллюстрирует простой пример.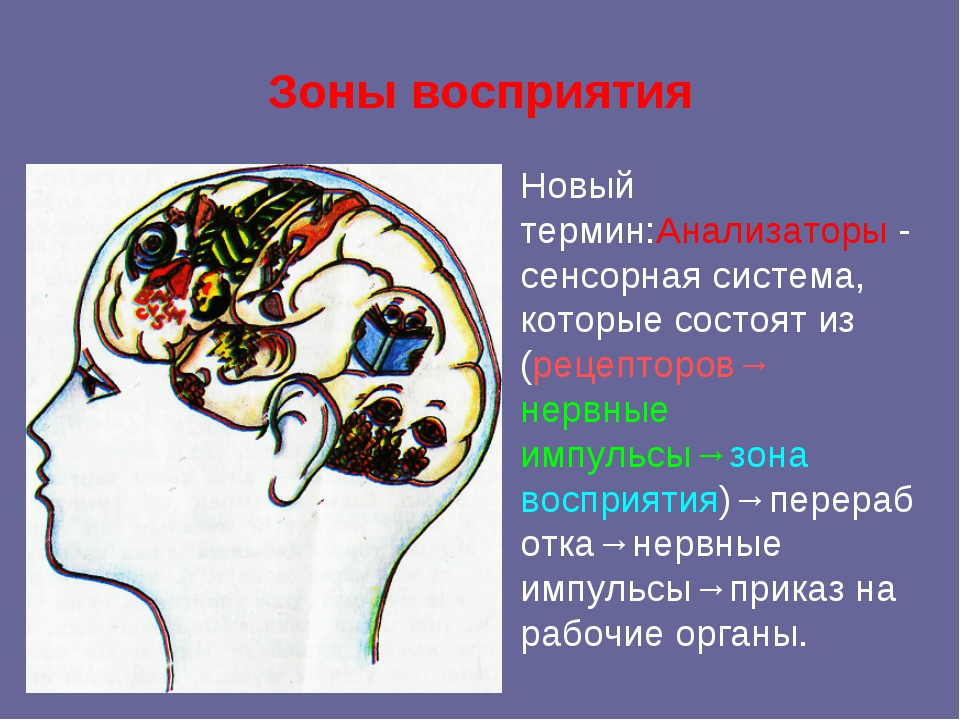 Если официант удерживает на ладони вытянутой руки поднос с бутылкой шампанского и рюмками, а другой человек внезапно снимет бутылку с подноса, то рука резко подпрыгнет вверх с соответствующими последствиями. Если же он сам снимет бутылку свободной рукой, то рука с подносом останется на прежнем уровне.
Если официант удерживает на ладони вытянутой руки поднос с бутылкой шампанского и рюмками, а другой человек внезапно снимет бутылку с подноса, то рука резко подпрыгнет вверх с соответствующими последствиями. Если же он сам снимет бутылку свободной рукой, то рука с подносом останется на прежнем уровне.
Association Cortex — обзор
Болезнь Альцгеймера
Болезнь Альцгеймера — распространенное дементирующее заболевание, характеризующееся прогрессирующей дегенерацией нейронов, глиозом, заметным истощением ацетилхолина и других нарушений нейротрансмиттеров, а также накоплением сенильных (амилоидных) бляшек и нейлесофибриллярных области базального переднего мозга, гиппокампа и ассоциативной коры. На рубеже тысячелетий появились многообещающие сообщения о том, что эстрогены играют важную роль в нормальном познании человека, благотворно влияют на проявления болезни Альцгеймера и могут даже защищать от развития этого нейродегенеративного расстройства у женщин.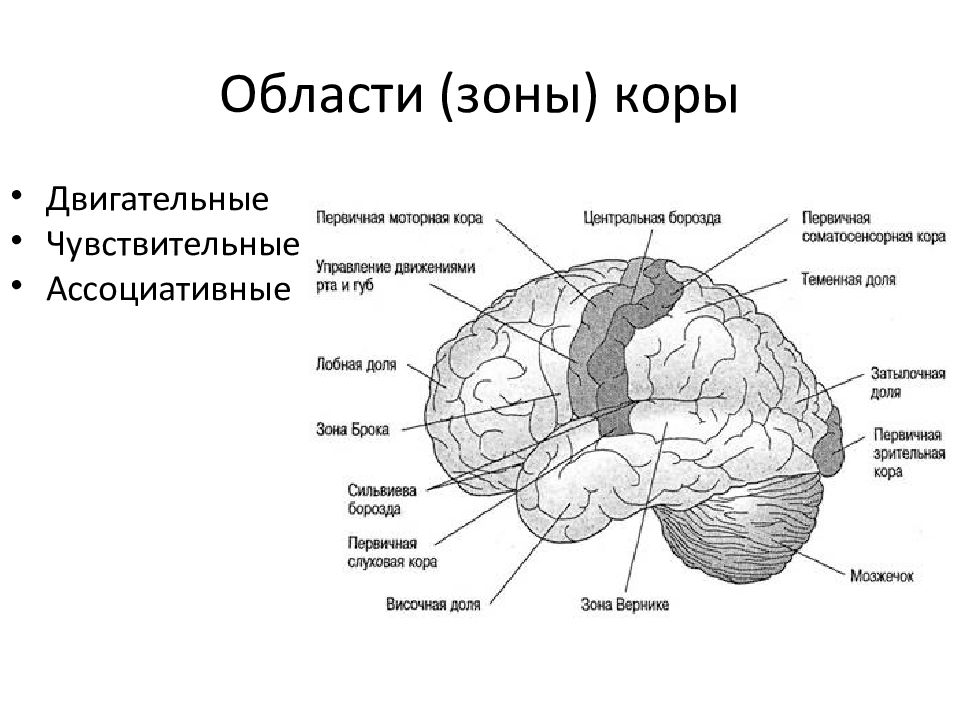 Фундаментальные исследования показывают, что эстрогены оказывают трофическое влияние на холинергические нейроны базального переднего мозга грызунов, индуцируют дендритные шипы (синапсы) и функциональные рецепторы N -метил-d-аспартата (NMDA) (важные для памяти) в гиппокампе взрослых крыс и вызывают массивный рост нейритов в гипоталамических эксплантатах грызунов. Кроме того, было показано, что эстрогены проявляют антиоксидантные свойства, уменьшают отложение фибриллярного β-амилоида, модулируют экспрессию аполипопротеина E, подавляют воспалительные реакции, участвующие в формировании нейритных бляшек, увеличивают церебральный кровоток и утилизацию глюкозы (которые недостаточны у субъектов с болезнью Альцгеймера. ) и стабилизируют паттерны экспрессии микроРНК и биоэнергетику митохондрий. 5 Также накапливаются данные о том, что эстрогены улучшают когнитивное поведение у крыс и обезьян; что на психометрические характеристики женщин влияет фаза менструального цикла; что трансгендерная гормональная терапия влияет на познавательные способности мужчин и женщин-транссексуалов; и что заместительная терапия эстрогенами увеличивает показатели вербальной памяти у нормальных женщин в менопаузе.
Фундаментальные исследования показывают, что эстрогены оказывают трофическое влияние на холинергические нейроны базального переднего мозга грызунов, индуцируют дендритные шипы (синапсы) и функциональные рецепторы N -метил-d-аспартата (NMDA) (важные для памяти) в гиппокампе взрослых крыс и вызывают массивный рост нейритов в гипоталамических эксплантатах грызунов. Кроме того, было показано, что эстрогены проявляют антиоксидантные свойства, уменьшают отложение фибриллярного β-амилоида, модулируют экспрессию аполипопротеина E, подавляют воспалительные реакции, участвующие в формировании нейритных бляшек, увеличивают церебральный кровоток и утилизацию глюкозы (которые недостаточны у субъектов с болезнью Альцгеймера. ) и стабилизируют паттерны экспрессии микроРНК и биоэнергетику митохондрий. 5 Также накапливаются данные о том, что эстрогены улучшают когнитивное поведение у крыс и обезьян; что на психометрические характеристики женщин влияет фаза менструального цикла; что трансгендерная гормональная терапия влияет на познавательные способности мужчин и женщин-транссексуалов; и что заместительная терапия эстрогенами увеличивает показатели вербальной памяти у нормальных женщин в менопаузе.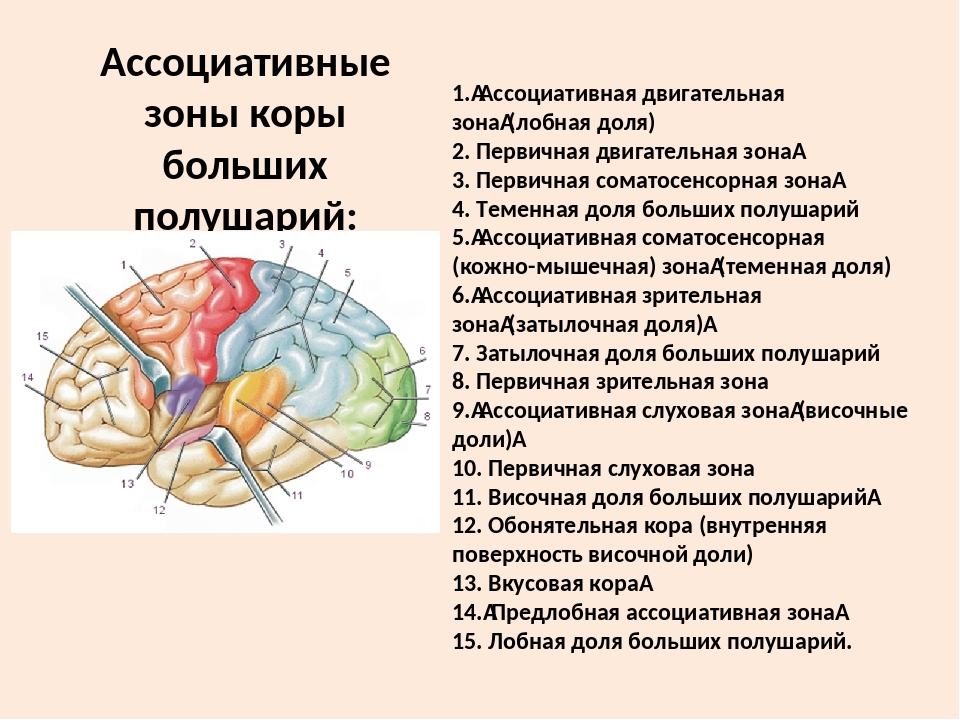 Более того, ранние клинические исследования показали, что заместительная терапия эстрогенами может улучшить когнитивные функции, особенно речевую функцию, вербальную память и внимание, у женщин в менопаузе с болезнью Альцгеймера и повысить вероятность положительного ответа на ингибиторы ацетилхолинэстеразы у пострадавших женщин.В нескольких исследованиях заместительная терапия эстрогенами в постменопаузе, по-видимому, связана со значительным снижением риска развития болезни Альцгеймера. Были некоторые указания на то, что заместительная эстрогеновая терапия в постменопаузе защищает от развития деменции у женщин с болезнью Паркинсона и что лечение андрогенами (тестостероном) или эстрогенами дает когнитивные преимущества у пожилых мужчин с болезнью Альцгеймера или умеренными когнитивными нарушениями. Более низкие уровни эстрадиола в постменопаузе также были связаны с повышенным риском деменции у людей с синдромом Дауна.
Более того, ранние клинические исследования показали, что заместительная терапия эстрогенами может улучшить когнитивные функции, особенно речевую функцию, вербальную память и внимание, у женщин в менопаузе с болезнью Альцгеймера и повысить вероятность положительного ответа на ингибиторы ацетилхолинэстеразы у пострадавших женщин.В нескольких исследованиях заместительная терапия эстрогенами в постменопаузе, по-видимому, связана со значительным снижением риска развития болезни Альцгеймера. Были некоторые указания на то, что заместительная эстрогеновая терапия в постменопаузе защищает от развития деменции у женщин с болезнью Паркинсона и что лечение андрогенами (тестостероном) или эстрогенами дает когнитивные преимущества у пожилых мужчин с болезнью Альцгеймера или умеренными когнитивными нарушениями. Более низкие уровни эстрадиола в постменопаузе также были связаны с повышенным риском деменции у людей с синдромом Дауна.
Результаты других крупных рандомизированных плацебо-контролируемых проспективных исследований, оценивающих потенциальные преимущества половой ЗГТ в предотвращении деменции, вызванной болезнью Альцгеймера, неутешительны. Скорректированные по возрасту показатели когнитивной функции у женщин с ишемической болезнью сердца, получавших эстроген и прогестин, не отличаются от контрольной группы, получавшей плацебо. Некоторые исследования даже продемонстрировали немного более высокий риск деменции при ЗГТ по сравнению с контрольной группой, получавшей плацебо. Следует отметить, что определенные полиморфизмы рецептора фолликулостимулирующего гормона могут обеспечивать защиту от заболевания у женщин (но не у мужчин).
Скорректированные по возрасту показатели когнитивной функции у женщин с ишемической болезнью сердца, получавших эстроген и прогестин, не отличаются от контрольной группы, получавшей плацебо. Некоторые исследования даже продемонстрировали немного более высокий риск деменции при ЗГТ по сравнению с контрольной группой, получавшей плацебо. Следует отметить, что определенные полиморфизмы рецептора фолликулостимулирующего гормона могут обеспечивать защиту от заболевания у женщин (но не у мужчин).
Третья канадская консенсусная конференция по диагностике и лечению деменции (2006) рекомендовала отказаться от использования заместительной терапии эстрогенами / прогестинами для снижения риска деменции у женщин в постменопаузе. Был также сделан вывод о недостаточности доказательств в пользу или против назначения заместительной терапии андрогенами при когнитивной дисфункции у пожилых мужчин. После публикации этих рекомендаций была высказана гипотеза, что может существовать критическое перименопаузальное «окно», в течение которого ЗГТ может защитить от развития болезни Альцгеймера. 5 Важно отметить, что якобы благотворное влияние эстрогена на когнитивные способности и объемы гиппокампа может быть компенсировано у стареющих женщин, несущих одну или две копии аллеля аполипопротеина E ε4. Что касается андрогенов, более высокие уровни биодоступного тестостерона в сыворотке крови в пожилом возрасте предсказывают снижение риска развития болезни Альцгеймера у мужчин, возможно, из-за индуцированной андрогеном подавляющей модуляции отложения β-амилоида в головном мозге. Могут потребоваться проспективные клинические испытания, достаточные для определения эффективности лечения андрогенами в предупреждении деменции у мужчин (и, возможно, женщин).
5 Важно отметить, что якобы благотворное влияние эстрогена на когнитивные способности и объемы гиппокампа может быть компенсировано у стареющих женщин, несущих одну или две копии аллеля аполипопротеина E ε4. Что касается андрогенов, более высокие уровни биодоступного тестостерона в сыворотке крови в пожилом возрасте предсказывают снижение риска развития болезни Альцгеймера у мужчин, возможно, из-за индуцированной андрогеном подавляющей модуляции отложения β-амилоида в головном мозге. Могут потребоваться проспективные клинические испытания, достаточные для определения эффективности лечения андрогенами в предупреждении деменции у мужчин (и, возможно, женщин).
Нейростероидогенез также вовлечен в патофизиологию болезни Альцгеймера. Имеются сообщения о снижении уровней DHEA-S в плазме и спинномозговой жидкости и концентрации аллопрегнанолона в префронтальной коре головного мозга людей с болезнью Альцгеймера. Аллопрегнанолон может способствовать когнитивному благополучию, поскольку он подавляет нейровоспаление (активацию микроглии), уменьшает патологию β-амилоида, увеличивает нейрогенез в гиппокампе и обращает вспять дефицит обучения и памяти в моделях болезни Альцгеймера на животных.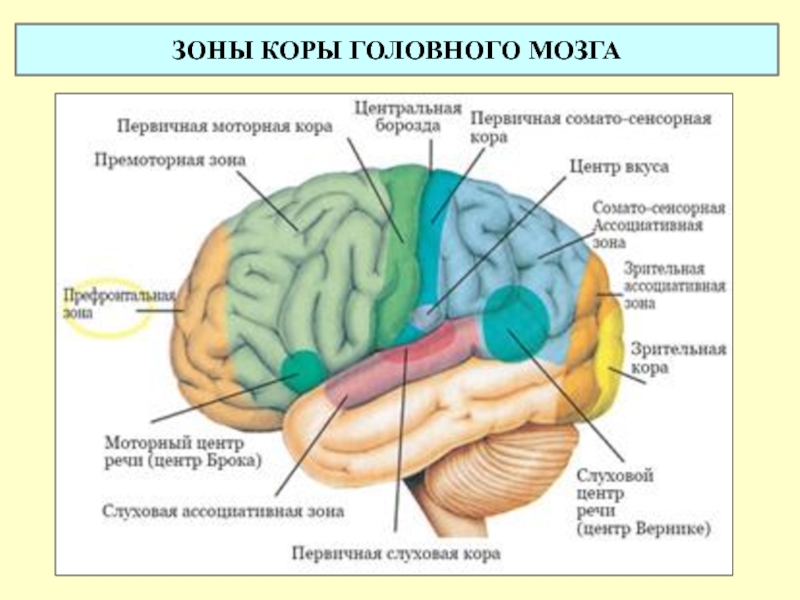
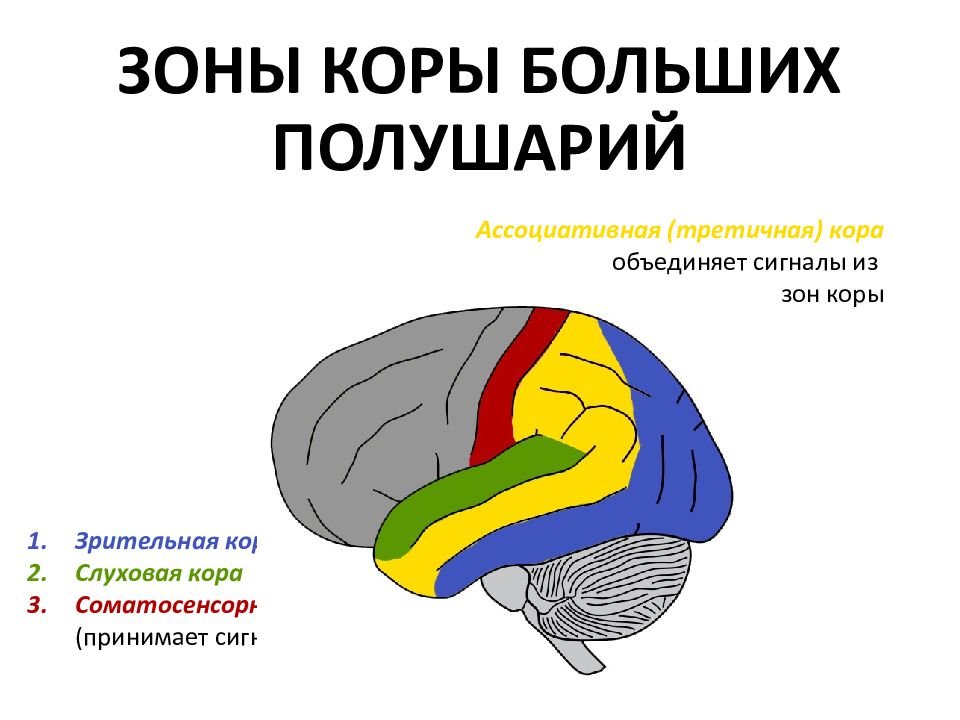
Высшие корковые функции: ассоциативная и исполнительная обработка (Раздел 4, Глава 9) Нейронаука в Интернете: Электронный учебник для нейронаук | Кафедра нейробиологии и анатомии
Интегративные области коры высшего порядка, называемые ассоциативными областями, вмешиваются между сенсорными входами и моторными выходами. Иерархическая организация коры была предложена в 1870-х годах известным британским неврологом Джоном Хьюлингсом Джексоном. Эти области ассоциации являются участком когнитивной обработки и являются фокусными областями в быстро развивающейся области когнитивной нейробиологии.
Рисунок 9.1 |
Три мультимодальных ассоциативных области будут обсуждаться вместе с тремя унимодальными ассоциативными областями и тремя первичными сенсорными областями.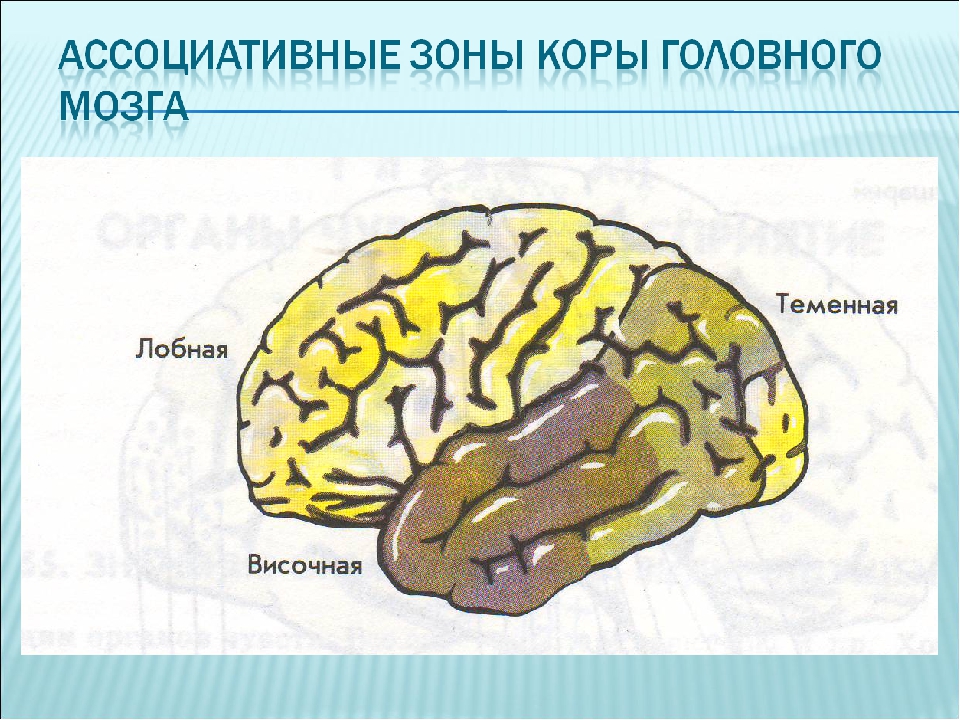
- Зона лимбической ассоциации
Располагается в передневентральной части височной доли, парагиппокампальной извилине.
Связывает эмоции со многими сенсорными сигналами.
Важен в обучении и памяти. - Зона задней ассоциации
Находится на стыке затылочной, височной и теменной долей.
Связывает информацию из первичных и одномодальных сенсорных областей
Важен в восприятии и языке. - Зона передней ассоциации
Расположен в префронтальной коре.
Ссылки на информацию из других областей ассоциации.
Важен в памяти, планировании и формировании концепций высшего порядка.
Три унимодальных ассоциативных области расположены рядом с их соответствующими первичными сенсорными областями коры, как показано на рисунке. Общий принцип, который следует усвоить, состоит в том, что по мере удаления от основных сенсорных областей ассоциативные функции становятся более общими.
Сенсорная информация обрабатывается и отправляется от рецепторов параллельными путями через первичную сенсорную кору и унимодальную ассоциативную кору в заднюю мультимодальную ассоциативную кору каждого полушария — заднюю теменную и височную коры.
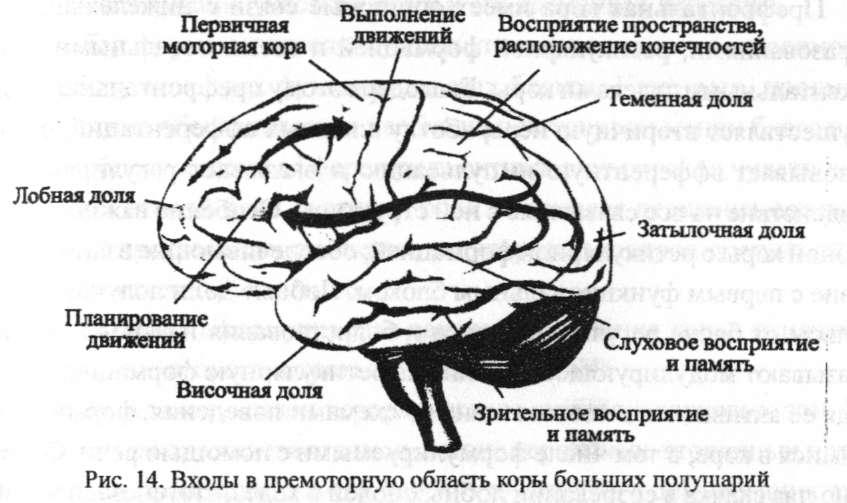
Задняя мультимодальная ассоциативная кора тесно связана с передними ассоциативными областями, которые, в свою очередь, отвечают за концептуальные когнитивные функции и планирование двигательных действий.
После планирования двигательных действий в области передних ассоциаций фактическая обработка выходных данных двигательного ответа является обратной обработкой в сенсорной (входной) системе. Премоторная кора головного мозга является ростральной по отношению к моторной коре — области 6 и 8 Бродмана.
Рисунок 9.2 | |
Извлечение информации, которая была изучена, очевидно, также является обратной по отношению к путям и структурам, используемым при хранении этой информации.
Рисунок 9.3. |
9.1 Зона задней ассоциации
Агнозия
Повреждение задних ассоциативных областей, также иногда включающих части одномодальных ассоциативных областей, может привести к агнозии, греческому слову, означающему «незнание». Поражение области задней зрительной ассоциации может привести к неспособности узнавать знакомые лица или изучать новые лица, в то же время оставляя без изменений другие аспекты визуального распознавания — дефицит, называемый прозопагнозией.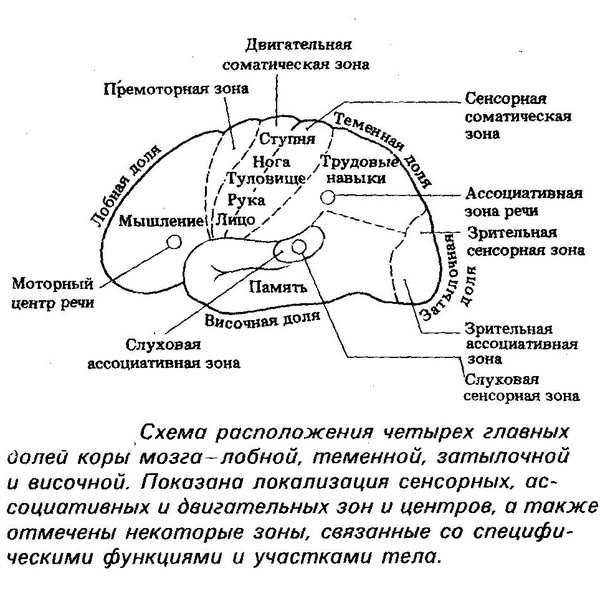 Эти пациенты могут идентифицировать лицо как лицо, его части и определенные эмоции по выражению лица, но они не могут идентифицировать лицо как конкретного человека. Они часто не могут узнать родственников, таких как родители или дети, а в некоторых случаях не могут узнать собственное лицо. Они не потеряли знания о своих близких родственниках и друзьях, потому что используют звук голоса и другие сигналы, чтобы узнать их. Поражения, вызывающие прозопагнозию, всегда двусторонние на нижней поверхности обеих затылочных долей и распространяются вперед на внутреннюю поверхность височных долей.
Эти пациенты могут идентифицировать лицо как лицо, его части и определенные эмоции по выражению лица, но они не могут идентифицировать лицо как конкретного человека. Они часто не могут узнать родственников, таких как родители или дети, а в некоторых случаях не могут узнать собственное лицо. Они не потеряли знания о своих близких родственниках и друзьях, потому что используют звук голоса и другие сигналы, чтобы узнать их. Поражения, вызывающие прозопагнозию, всегда двусторонние на нижней поверхности обеих затылочных долей и распространяются вперед на внутреннюю поверхность височных долей.
Пациенты с поражениями задней мультимодальной ассоциативной коры (т. Е. Задней теменной коры) могут воспринимать и рисовать объекты, но не могут их назвать — дефицит, называемый ассоциативной агнозией. Они не забыли названия этих предметов, потому что могут правильно назвать их на ощупь. Другое повреждение затылочных долей и окружающей области может привести к тому, что пациенты не смогут рисовать предметы, но, что примечательно, все еще могут их называть — дефицит, называемый апперцептивной агнозией.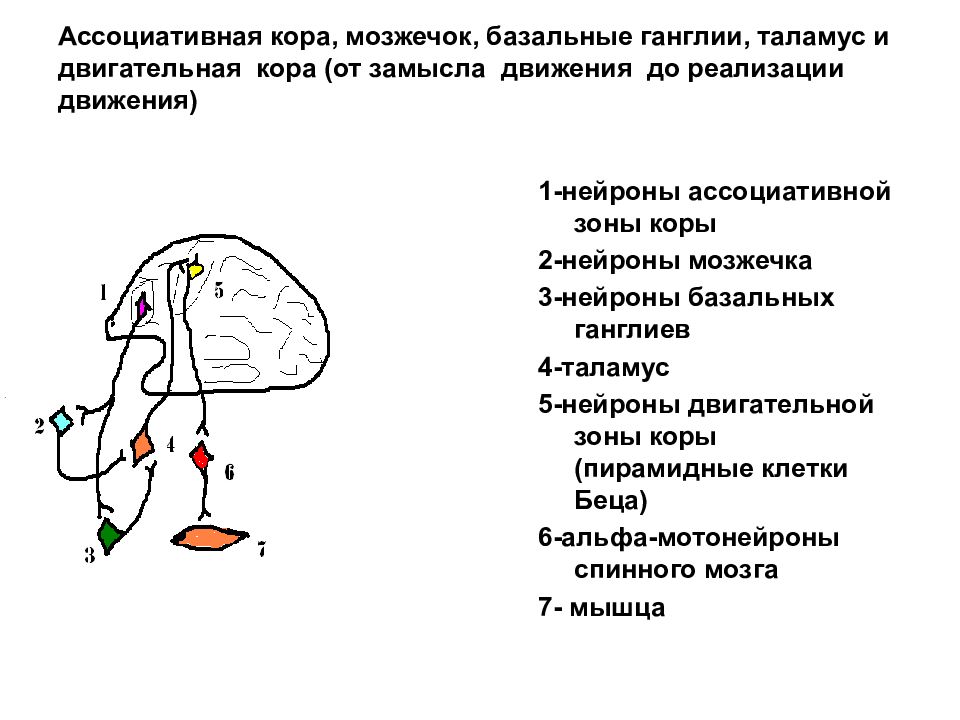
Рисунок 9.4 | Рисунок 9.5 |
Есть и другие случаи, когда поражения в аналогичных областях правого и левого полушария вызывают почти дополнительные различия в восприятии.Повреждение правого полушария нарушает восприятие глобальных объектов, а повреждение левого полушария нарушает восприятие локальных объектов. Это обнаруживается только в острых стадиях сразу после инсульта и только при поражениях, сосредоточенных в височно-теменном соединении (TPJ).
9.2 Контралатеральное пренебрежение
Рис. 9.6. |
Агнозии правой задней теменной зрительной коры — одни из самых замечательных. Мультимодальная задняя ассоциативная зона получает сигналы от зрительной и слуховой систем и от гиппокампа. Они возникают в результате неспособности воспринимать объекты, несмотря на нормально функционирующие сенсорные системы. Эти пациенты игнорируют вещи слева, включая левую половину предметов и левую половину своего тела. Это называется синдромом контралатерального пренебрежения.
Контралатеральное пренебрежение — это дефицит самооценки левой стороны своего тела и дефицит восприятия мира левой стороной. Как правило, у пациентов, страдающих синдромом контралатерального пренебрежения, наблюдается паралич левой стороны.
Некоторые пациенты не моют и не одевают левый бок — синдром пренебрежения личным вниманием. Это пренебрежение может распространяться на отказ от левой части тела. Они будут делать такие замечания, как «Кто положил эту руку мне в кровать», когда ссылаются на свою левую руку.
Пренебрежение может распространяться не только на личное пространство, но и на мир вокруг них. Они могут нарисовать цветок, например, с лепестками только с правой стороны, или цифрами часов только с правой стороны и т. Д.
Небольшая мысль убедит вас, что это не сенсорная проблема. Не только правая половина всех объектов в окружающей среде имеет вход в такую сенсорную систему пациента. Большинство объектов состоит из нескольких частей. Вы можете думать об этих частях как об объектах.Так, например, каждый лепесток цветка — это объект. Они не «видят» только правую половину каждого лепестка. Это проблема бесконечного регресса.
Чтобы показать вам, насколько сложной и сложной может стать проблема игнорирования зрения, в Милане, Италия, было проведено исследование группы пациентов с игнорированием зрения.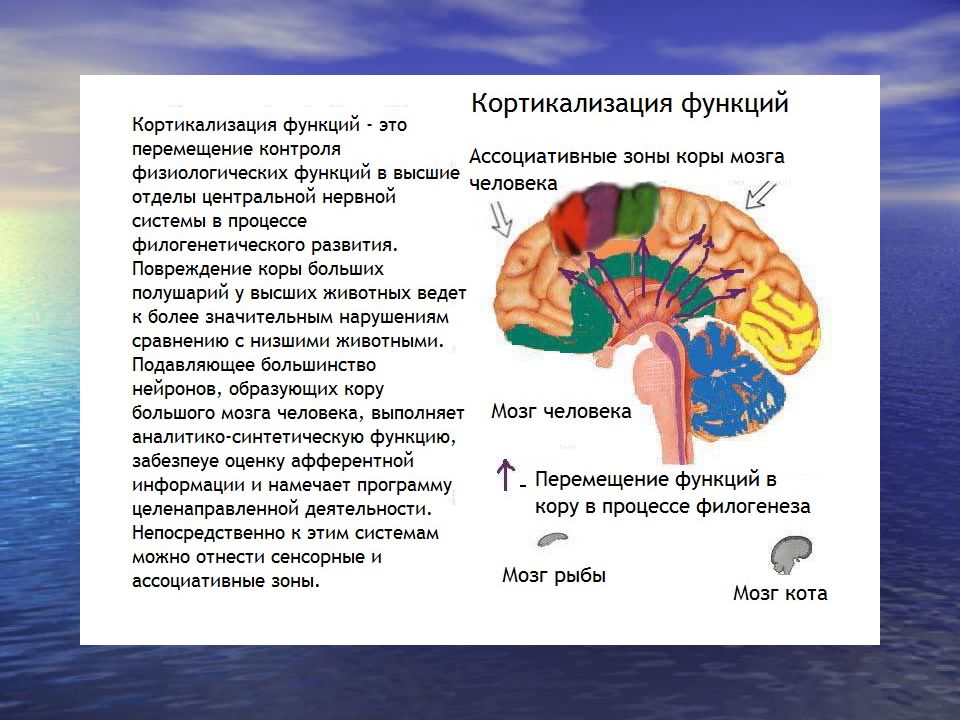 В медицинском кабинете их попросили представить известную общественную площадь — Пьяцца дель Дуомо. Им сказали, что они должны представить, что стоят лицом к собору. Затем их попросили вспомнить все здания на площади.Вспомнили только здания справа от их (воображаемой). Затем им сказали, что они стоят на ступенях собора и представляют, что смотрят в противоположную сторону. В очередной раз их попросили вспомнить все постройки на площади. Что они сделали, так это вспомнили все здания, которые они не смогли вспомнить во время первого отзыва, потому что здания, которые раньше были слева от них, теперь находятся справа от них. Помните, что все это в их воображении — в их памяти.
В медицинском кабинете их попросили представить известную общественную площадь — Пьяцца дель Дуомо. Им сказали, что они должны представить, что стоят лицом к собору. Затем их попросили вспомнить все здания на площади.Вспомнили только здания справа от их (воображаемой). Затем им сказали, что они стоят на ступенях собора и представляют, что смотрят в противоположную сторону. В очередной раз их попросили вспомнить все постройки на площади. Что они сделали, так это вспомнили все здания, которые они не смогли вспомнить во время первого отзыва, потому что здания, которые раньше были слева от них, теперь находятся справа от них. Помните, что все это в их воображении — в их памяти.
| Рисунок 9.7 Собор Пьяцца дель Дуомо в Милане, Италия. | ||
Это наглядное свидетельство дефицита обработки. Эти пациенты помнят о площади Пьяцца дель Дуомо; у них нет проблем с памятью ни о какой части квадрата. У них также есть полный доступ к своей памяти о площади. Но в зависимости от их воображаемой перспективы они не могут вспомнить объекты в левой части их системы координат.Более того, они явно не подозревают об этом дефиците. Их система координат сосредоточена на их теле, в памяти, а также в реальной жизни. По-видимому, доступ к воспоминаниям, таким как сцены из реального мира, осуществляется через контралатеральное полушарие. Дело не в том, что воспоминания были заложены с отсутствующими объектами слева, потому что они выросли, зная этот квадрат, задолго до того, как произошло поражение их мозга. Это поднимает вопросы сознания, самосознания, исполнительного контроля памяти.
У них также есть полный доступ к своей памяти о площади. Но в зависимости от их воображаемой перспективы они не могут вспомнить объекты в левой части их системы координат.Более того, они явно не подозревают об этом дефиците. Их система координат сосредоточена на их теле, в памяти, а также в реальной жизни. По-видимому, доступ к воспоминаниям, таким как сцены из реального мира, осуществляется через контралатеральное полушарие. Дело не в том, что воспоминания были заложены с отсутствующими объектами слева, потому что они выросли, зная этот квадрат, задолго до того, как произошло поражение их мозга. Это поднимает вопросы сознания, самосознания, исполнительного контроля памяти.
Что еще больше усложняет ситуацию, можно сосредоточить одностороннее игнорирование зрения по объекту, а не по телу пациента.Пациент с объектно-ориентированным односторонним игнорированием зрения. Слева: пациент заботится о левой руке исследователя даже при повороте. Справа: пациент пренебрегает правой рукой экзаменатора, даже если его тело повернуто на 90 градусов.
Двустороннее повреждение теменной поверхности может вызвать так называемый синдром Балинта. Это интересный синдром, потому что вместо того, чтобы игнорировать обе стороны объектов и ничего не видеть, как можно было бы ожидать при двустороннем пренебрежении полушарием, они видят по одному объекту за раз — одновременная агнозия.Эти пациенты сообщают, что объект появляется автоматически и случайным образом заменяется другим объектом, и они не могут контролировать, какой объект будет восприниматься. У этих пациентов есть проблемы с ADL, включая потерю, неспособность брать предметы, и они не могут есть, одеваться или передвигаться по комнате без посторонней помощи. Однако они могут правильно касаться частей своего тела.
9.3 Зона лимбической ассоциации
Область лимбической ассоциации получает информацию практически из любой другой области ассоциации и, следовательно, может связывать все стимулы события, включая его эмоциональный контекст.Эмоция, связанная с событием, может определить, запомнится ли оно и как долго.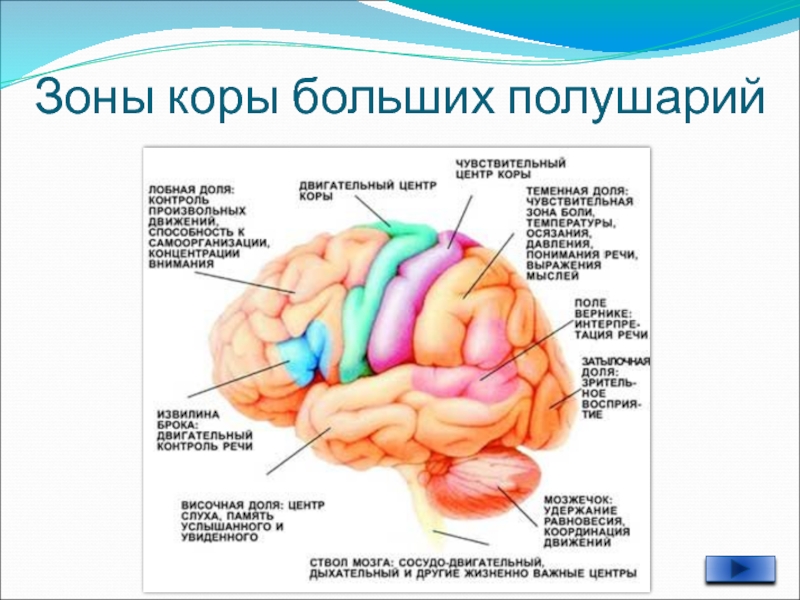 Это важно для выживания всех организмов. В самом деле, это то, что подразумевается под обучением. Вы не можете учиться без памяти. Когда вы голодны, найти пищу становится большим облегчением, и тогда у человека появляется больше шансов вспомнить место еды позже. Когда кто-то чудом избегает опасности, у него больше шансов избежать таких хищников и мест, где они обитают. У пациента HM были удалены области лимбической ассоциации с двух сторон, он не мог формировать какие-либо новые явные воспоминания, которые полагались бы на контекстную память, включая место, время и эмоции.
Это важно для выживания всех организмов. В самом деле, это то, что подразумевается под обучением. Вы не можете учиться без памяти. Когда вы голодны, найти пищу становится большим облегчением, и тогда у человека появляется больше шансов вспомнить место еды позже. Когда кто-то чудом избегает опасности, у него больше шансов избежать таких хищников и мест, где они обитают. У пациента HM были удалены области лимбической ассоциации с двух сторон, он не мог формировать какие-либо новые явные воспоминания, которые полагались бы на контекстную память, включая место, время и эмоции.
9.4 Зона передней ассоциации
Рисунок 9.8 |
Передняя ассоциативная зона находится в лобных долях. Это рострально по отношению к постцентральным извилинам, роландической щели и премоторным областям. Его задней границей является сильвиева трещина. Это называется префронтальной корой.
Его задней границей является сильвиева трещина. Это называется префронтальной корой.
Раннее свидетельство роли префронтальной коры было получено в случае с Финеасом Т. Гейджем. Гейдж был прорабом на строительстве железной дороги в середине 19 века. Он был надежным, хорошо организованным и трудолюбивым. Он забивал взрывчатку в яму, просверленную в скале с помощью 13-фунтового железного стержня длиной 3½ фута. Заряд неожиданно взорвался, когда он укладывал заряд взрывчатого вещества. Произошло то, что трамбующий стержень вылетел из отверстия и прошел через переднюю часть его головы, разрушив большие участки его префронтальной коры.После того, как он выздоровел физически (что примечательно всего за несколько недель), его личность изменилась. Коллеги сообщили, что «Гейдж не был Гейджем». Он был ненадежным. Его действия были импульсивными, без особого внимания к последствиям. Он стал алкоголиком и бродягой.
Префронтальная лоботомия была проведена в первой половине двадцатого века для облегчения психотических симптомов. Сейчас они заменены лекарственной терапией. При односторонней или двусторонней префронтальной лоботомии не хватает способности запоминать и связывать вещи с течением времени.Отсроченное вознаграждение сильнее сказывается на обучении. Продолжительность внимания и способность концентрироваться значительно уменьшаются. Абстрактное мышление в значительной степени исчезает. Префронтальная кора получает массивные сигналы от сенсорных ассоциаций коры (соматосенсорной, зрительной и слуховой), а также от дорсомедиального ядра таламуса. Поражение дорсомедиального ядра таламуса может вызывать многие из тех же симптомов, что и при префронтальной лоботомии.
Сейчас они заменены лекарственной терапией. При односторонней или двусторонней префронтальной лоботомии не хватает способности запоминать и связывать вещи с течением времени.Отсроченное вознаграждение сильнее сказывается на обучении. Продолжительность внимания и способность концентрироваться значительно уменьшаются. Абстрактное мышление в значительной степени исчезает. Префронтальная кора получает массивные сигналы от сенсорных ассоциаций коры (соматосенсорной, зрительной и слуховой), а также от дорсомедиального ядра таламуса. Поражение дорсомедиального ядра таламуса может вызывать многие из тех же симптомов, что и при префронтальной лоботомии.
Рисунок 9.9 |
Префронтальная кора подразделяется на три области.
- Дорсальная (верхняя) префронтальная кора
- Медиальная (окружающая главную борозду) префронтальная кора
- Вентрально-орбитофронтальная (или нижняя) кора
А.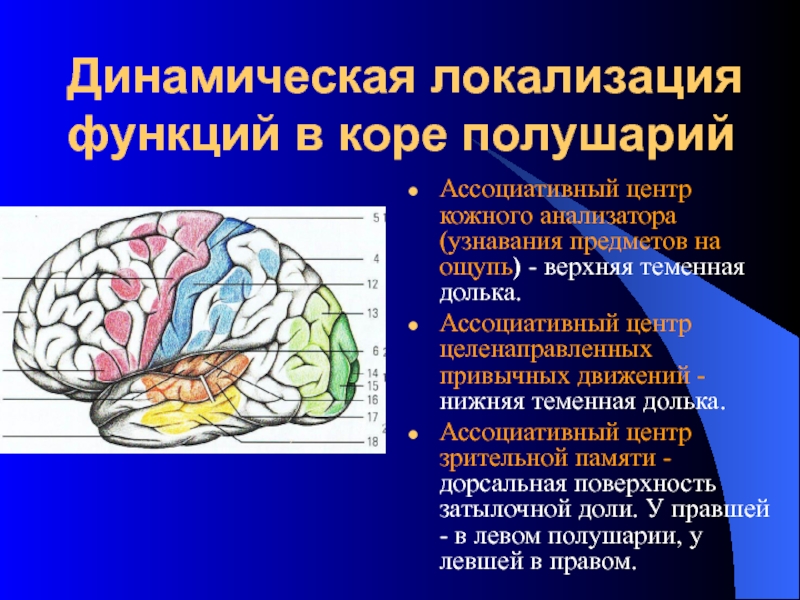 | Б. | |
Рисунок 9.10. | ||
Префронтальный синдром (или фронтальный синдром) у пациентов обычно относится к повреждению дорсальной (верхней) области префронтальной ассоциации. Большинство исследований дорсальной префронтальной области сосредоточено в дорсолатеральной префронтальной области.Это область (46) на приведенной выше диаграмме чуть дорсальнее основной борозды (рис. 9.10A). Это очень важная область для многих когнитивных способностей более высокого порядка. Дорсолатеральная префронтальная область вместе с поясной корой участвует в обработке внимания, планировании, обучении правилам и памяти. Например, при попытке выполнить арифметику пациент может успешно начать последовательное вычитание 7 из 100, но затем необъяснимо сдвинуться и сказать: «100, 93, 93, 73… 63 вместо 100, 93, 86, 79, 72 и т. Д. .Когда последовательности изучаются, дорсолатеральная префронтальная область и поясная извилина кора очень активны. Но когда обучение завершено и автоматизировано, эти области перестают быть активными.
Д. .Когда последовательности изучаются, дорсолатеральная префронтальная область и поясная извилина кора очень активны. Но когда обучение завершено и автоматизировано, эти области перестают быть активными.
Многие нейроны в дорсолатеральной префронтальной области и нейроны в самой главной борозде (то есть в дорсальной и медиальной префронтальной коре) опосредуют память о ГДЕ расположен объект. Эти нейроны получают сигналы от дорсального пути через заднюю теменную кору. Записи на обезьянах показывают, что нейроны «где» реагируют только на стимулы в определенной позиции в поле зрения и продолжают активироваться в течение периода задержки — предположительно, воспоминания о событии.Когда время от времени они прекращают стрельбу в течение периода задержки, это обычно сигнализирует о том, что обезьяна забыла о местонахождении. Устройства слежения за глазами используются для записи того, куда смотрит обезьяна, и обезьяна обучается смотреть в сторону определенного запомненного места.
В отличие от дорсолатеральной префронтальной области, нейроны в небольшой области (46) на рисунке 9.10A, расположенной вентрально от основной борозды, опосредуют память о том, ЧТО представляет собой объект, его форму и цвет. Это цель вентрального зрительного пути через нижнюю височную долю.
Рисунок 9.11. Нажмите на ярлыки, чтобы увидеть пути. |
Орбитофронтальная кора и медиальная префронтальная кора имеют прямые связи с миндалевидным телом и поясной корой лимбической системы и, таким образом, обеспечивают эмоциональный компонент запланированного поведения и памяти.
Доминирующим нейромедиатором в префронтальной области является дофамин.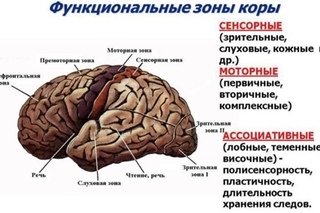 Истощение дофамина может вызвать поражение, подобное симптомам. Считается, что нарушения дофаминергической системы способствуют развитию симптомов шизофреников, и у многих шизофреников наблюдается гипофункция префронтальных областей коры головного мозга. Такие симптомы, как управление чужими голосами, указывают на дисфункцию исполнительной системы управления у шизофреников.
Истощение дофамина может вызвать поражение, подобное симптомам. Считается, что нарушения дофаминергической системы способствуют развитию симптомов шизофреников, и у многих шизофреников наблюдается гипофункция префронтальных областей коры головного мозга. Такие симптомы, как управление чужими голосами, указывают на дисфункцию исполнительной системы управления у шизофреников.
Например, если пациенты с шизофренией проходят тестирование с помощью Висконсинского теста сортировки карт, кровоток в префронтальных областях намного меньше, чем у нормальных людей.Показано, что приток крови к префронтальным областям является функциональной частью точного выполнения этой задачи, потому что, когда шизофреники награждаются за их правильные ответы, кровоток увеличивается намного больше, чем при нормальном, и они демонстрируют улучшенную работоспособность.
Еще одним доказательством тесного взаимодействия между префронтальной корой и поясной корой является то, что нейровизуализация шизофреников показывает нарушение активации поясной коры при выполнении когнитивных задач, а посмертный гистологический анализ выявил аномалии в поясной коре головного мозга шизофреников.
Некоторые последние мысли о латерализации когнитивных функций
Другой способ определить, являются ли функции латеральными в одном полушарии или представлены в обоих полушариях, — это тестирование пациентов с расщепленным мозгом, пациентов, у которых было рассечено мозолистое тело. Рассечение мозолистого тела — последнее средство лечения трудноизлечимой эпилепсии. Затем стимулы можно вводить только в одно полушарие, при этом другое не осознает эти стимулы. Пациентов можно обследовать, чтобы увидеть, как они обрабатывают эти раздражители.
Рисунок 9.12. | Рисунок 9. |
 13
13 Тахистоскопически стимулы предъявляются только к одному полушарию. Пациенты сосредотачиваются на точке прямо в центре экрана. Если стимул появляется слева, он попадает в правое полушарие. Если стимул появляется справа, он попадает в левое полушарие. Итак, в ответ на конус, представленный в правом (левом полушарии), пациент говорит «конус». Но когда он представлен слева, пациент отрицает, что что-либо видит, а когда его заставляют дать ответ, пациент начинает болтать.Итак, когда стимул конуса находится слева, не способен ли пациент распознавать визуальный стимул как конус?
На самом деле пациент способен распознавать раздражитель как конус. Но нужно быть умным, чтобы проявить эту способность. Пациент может на ощупь правильно выбрать нужный объект. Или пациент может правильно определить его, указав на него, но только левой рукой.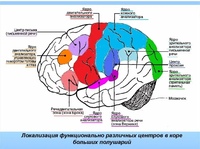
Можно сказать, что это просто сопоставление — даже перекрестное сопоставление (от визуального к тактильному) — но не язык.Но вы можете убедить себя, что это действительно язык, потому что, если вы направите буквы D-O-G в правое полушарие, пациенты смогут выбрать модель собаки — конечно, только левой рукой.
Другие результаты пациентов с расщепленным мозгом показали, что для большинства людей левое полушарие является доминирующим для математики, а также для языка.
Рисунок 9.14. |
Правое полушарие является доминирующим для музыки, распознавания лиц и всего, что связано с пространственными отношениями.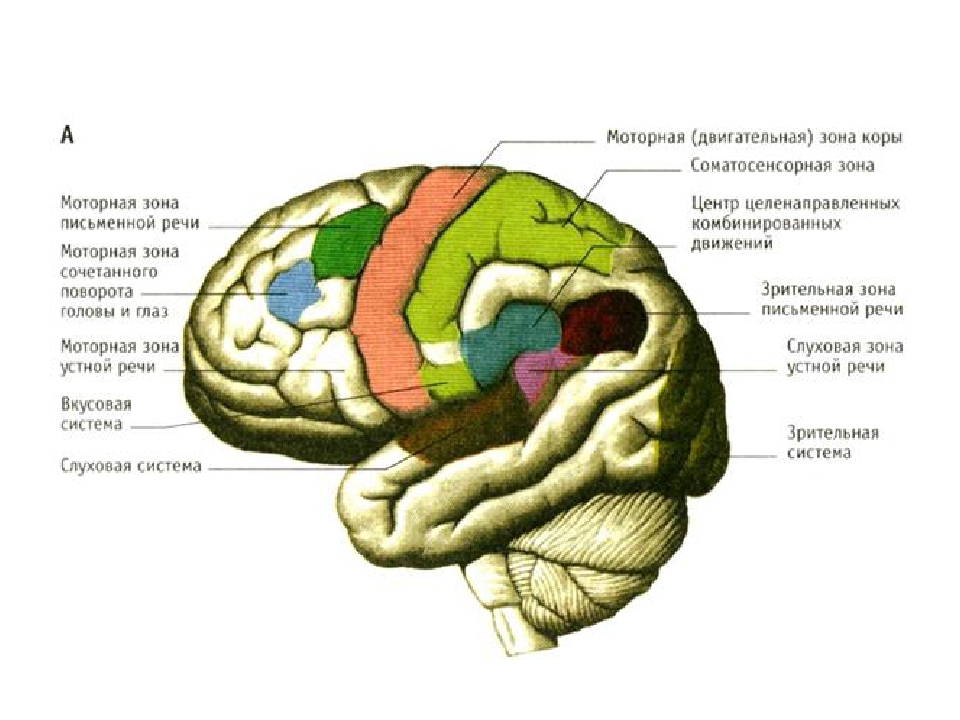
Например, даже пациенты-правши после резекции мозолистого тела (расщепление мозга) могут лучше рисовать левой рукой, чем правой, потому что левую руку контролирует правое полушарие.
Таким образом, правое полушарие выполняет некоторые функции, которые превосходят функции левого полушария, а правое полушарие не похоже на левое полушарие без языка. Другими примерами превосходства правого полушария может быть то, что пациенты с расщепленным мозгом могут соединять вместе деревянные блоки разных цветов, чтобы их левая рука получилась лучше, чем правой, снова показывая, что правое полушарие превосходит пространственное восприятие. задания.
В заключение следует отметить, что функции мозга, локализованные в определенных областях мозга, имеют большое клиническое значение.Локализация функции может объяснить, почему определенные синдромы характерны для болезни в определенных областях мозга. Тем не менее, ни одна часть мозга не работает изолированно. Каждая часть мозга работает согласованно со всеми остальными частями.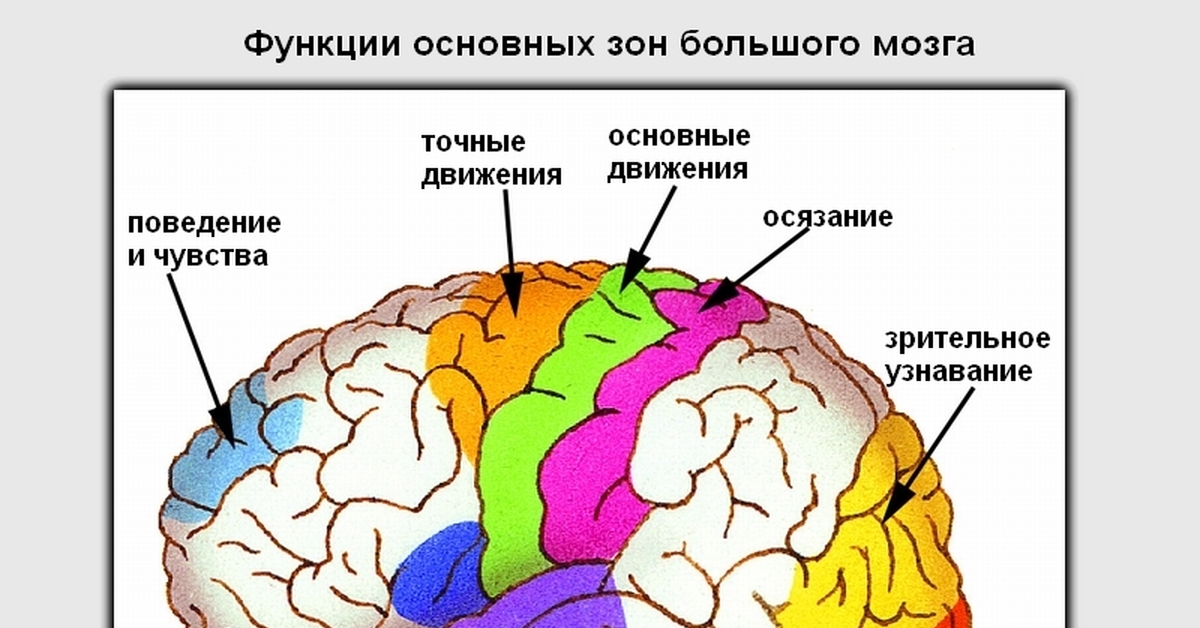 Когда часть мозга удаляется, результирующее поведение может больше отражать скорректированные возможности оставшихся «частей», чем удаленная часть.
Когда часть мозга удаляется, результирующее поведение может больше отражать скорректированные возможности оставшихся «частей», чем удаленная часть.
Проверьте свои знания
У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения».Какая область головного мозга была поражена наиболее вероятно?
A. Недоминантная теменно-затылочная кора
Б. Доминантная теменно-затылочная кора
C. Дорсальная префронтальная кора
D. Доминирующая височно-затылочная кора
E. Недоминантная височно-затылочная кора
У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения». Какая область головного мозга была поражена наиболее вероятно?
Какая область головного мозга была поражена наиболее вероятно?
A. Недоминантная теменно-затылочная кора. Ответ ПРАВИЛЬНЫЙ!
Поражение правой задней теменной зрительной коры (т. Е. Теменно-затылочной коры), скорее всего, вызывает игнорирование зрения. Это будет в правом полушарии для правшей, вызывающих пренебрежение левым зрением. Хотя иногда поражения доминирующего теменно-затылочного полушария (например,, левое полушарие у правшей) может вызвать визуальное игнорирование доминирующего участка, это менее вероятно, чем правильный ответ.
Б. Доминантная теменно-затылочная кора
C. Дорсальная префронтальная кора
D. Доминирующая височно-затылочная кора
E. Недоминантная височно-затылочная кора
У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения». Какая область головного мозга была поражена наиболее вероятно?
Какая область головного мозга была поражена наиболее вероятно?
A. Недоминантная теменно-затылочная кора
B. Доминирующая теменно-затылочная кора Этот ответ НЕПРАВИЛЬНЫЙ.
Эта область мозга обычно не связана с визуальным пренебрежением.
C. Дорсальная префронтальная кора
D. Доминирующая височно-затылочная кора
E.Недоминантная височно-затылочная кора
У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения». Какая область головного мозга была поражена наиболее вероятно?
A. Недоминантная теменно-затылочная кора
Б. Доминантная теменно-затылочная кора
C.
Эта область мозга обычно не связана с визуальным пренебрежением.
D. Доминирующая височно-затылочная кора
E. Недоминантная височно-затылочная кора
 Дорсальная префронтальная кора Этот ответ НЕПРАВИЛЬНЫЙ.
Дорсальная префронтальная кора Этот ответ НЕПРАВИЛЬНЫЙ.У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения». Какая область головного мозга была поражена наиболее вероятно?
А.Недоминантная теменно-затылочная кора
Б. Доминантная теменно-затылочная кора
C. Дорсальная префронтальная кора
D. Доминирующая височно-затылочная кора Этот ответ НЕПРАВИЛЬНЫЙ.
Эта область мозга обычно не связана с визуальным пренебрежением.
E. Недоминантная височно-затылочная кора

У 43-летнего пациента с церебральным сосудистым нарушением была диагностирована стойкая односторонняя пространственная агнозия или «игнорирование зрения».Какая область головного мозга была поражена наиболее вероятно?
A. Недоминантная теменно-затылочная кора
Б. Доминантная теменно-затылочная кора
C. Дорсальная префронтальная кора
D. Доминирующая височно-затылочная кора
E. Недоминантная височно-затылочная кора Этот ответ НЕПРАВИЛЬНЫЙ.
Эта область мозга обычно не связана с визуальным пренебрежением.
Motor Cortex (Раздел 3, Глава 3) Нейронауки в Интернете: Электронный учебник для нейронаук | Кафедра нейробиологии и анатомии
3. 1 Введение
1 Введение
В предыдущих главах обсуждались нижние уровни моторной иерархии (спинной мозг и ствол мозга), которые участвуют в низкоуровневой обработке «гаек и болтов», которая контролирует активность отдельных мышц.Отдельные альфа-моторные нейроны контролируют силу, действующую на конкретную мышцу, а спинномозговые цепи могут управлять сложными и сложными формами поведения, такими как ходьба и рефлекторные действия. Однако типы движений, контролируемые этими контурами, не инициируются сознательно. Произвольные движения требуют участия третьего и четвертого уровней иерархии: моторной коры и ассоциативной коры. Эти области коры головного мозга планируют произвольные действия, координируют последовательность движений, принимают решения о правильных поведенческих стратегиях и вариантах выбора, оценивают целесообразность конкретного действия с учетом текущего поведенческого или окружающего контекста и передают команды соответствующим наборам нижних мотонейронов. для выполнения желаемых действий.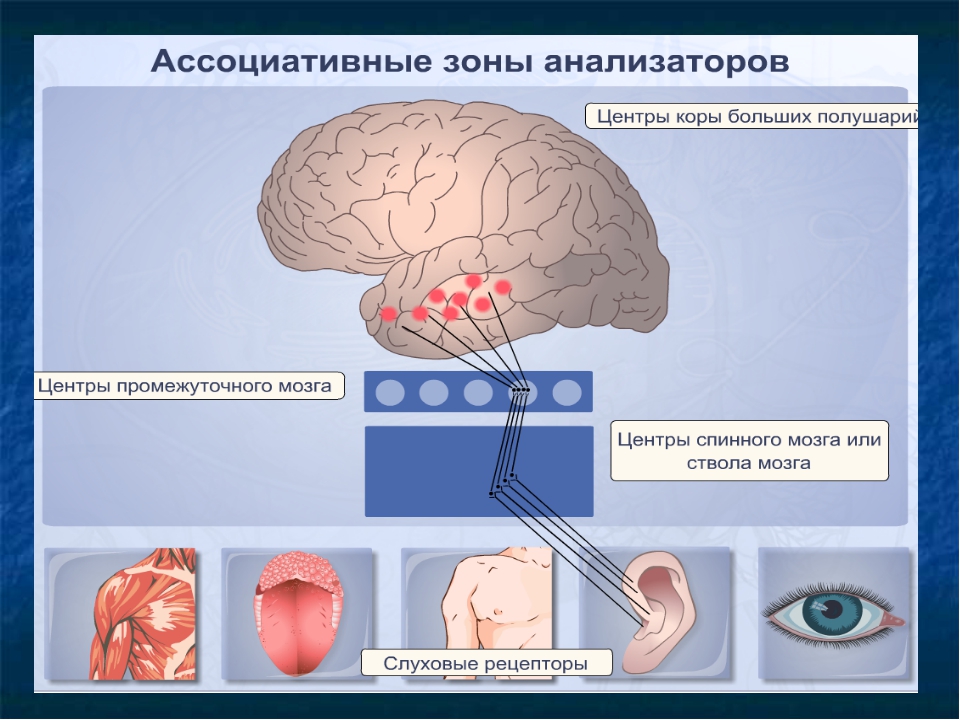
3.2 Моторная кора состоит из первичной моторной коры, премоторной коры и дополнительной моторной области
Рис. 3.1. |
Моторная кора состоит из трех различных областей лобной доли, непосредственно перед центральной бороздой .Этими областями являются первичная моторная кора (зона Бродмана 4), премоторная кора и дополнительная моторная зона (рис. 3.1). Электростимуляция этих областей вызывает движения определенных частей тела. Первичная моторная кора, или M1 , расположена на прецентральной извилине и на передней парацентральной доле на медиальной поверхности головного мозга. Из трех областей моторной коры для стимуляции первичной моторной коры требуется наименьшее количество электрического тока, чтобы вызвать движение.Низкие уровни кратковременной стимуляции обычно вызывают простые движения отдельных частей тела. Стимуляция премоторной коры или дополнительной моторной области требует более высоких уровней тока для вызова движений и часто приводит к более сложным движениям, чем стимуляция первичной моторной коры. Стимуляция в течение более длительных периодов времени (500 мсек) у обезьян приводит к перемещению определенной части тела в стереотипную позу или положение, независимо от начальной начальной точки этой части тела (рис.2). Таким образом, премоторная кора и дополнительные моторные области оказываются областями более высокого уровня, которые кодируют сложные паттерны моторной отдачи и выбирают соответствующие моторные планы для достижения желаемых конечных результатов.
Из трех областей моторной коры для стимуляции первичной моторной коры требуется наименьшее количество электрического тока, чтобы вызвать движение.Низкие уровни кратковременной стимуляции обычно вызывают простые движения отдельных частей тела. Стимуляция премоторной коры или дополнительной моторной области требует более высоких уровней тока для вызова движений и часто приводит к более сложным движениям, чем стимуляция первичной моторной коры. Стимуляция в течение более длительных периодов времени (500 мсек) у обезьян приводит к перемещению определенной части тела в стереотипную позу или положение, независимо от начальной начальной точки этой части тела (рис.2). Таким образом, премоторная кора и дополнительные моторные области оказываются областями более высокого уровня, которые кодируют сложные паттерны моторной отдачи и выбирают соответствующие моторные планы для достижения желаемых конечных результатов.
Рис. |
 3.2.
3.2. Как и соматосенсорная кора постцентральной извилины, первичная моторная кора организована соматотопически (рис.3). Стимуляция передней парацентральной доли вызывает движения противоположной ноги. По мере того, как стимулирующий электрод перемещается через прецентральную извилину от дорсомедиальной к вентролатеральной, движения прогрессивно вызываются от туловища, руки, кисти и лица (наиболее латерально).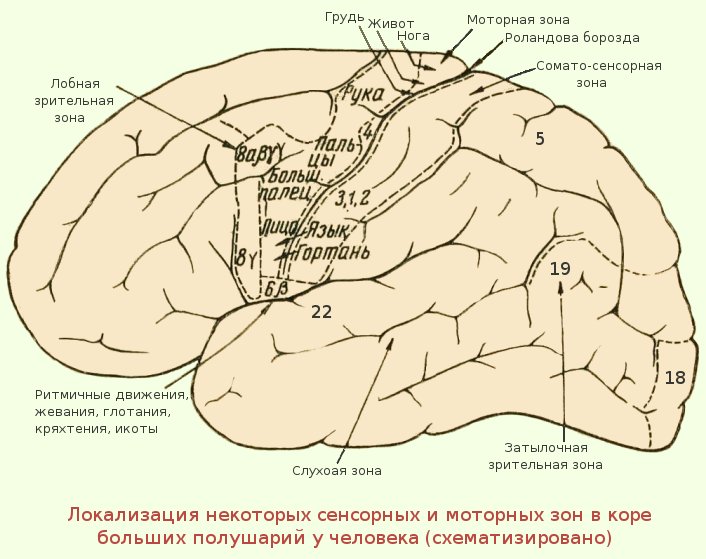 Представления частей тела, которые выполняют точные, деликатные движения, таких как руки и лицо, непропорционально велики по сравнению с изображениями частей тела, которые выполняют только грубые, необработанные движения, таких как туловище или ноги.Премоторная кора и дополнительная моторная область также содержат соматотопические карты.
Представления частей тела, которые выполняют точные, деликатные движения, таких как руки и лицо, непропорционально велики по сравнению с изображениями частей тела, которые выполняют только грубые, необработанные движения, таких как туловище или ноги.Премоторная кора и дополнительная моторная область также содержат соматотопические карты.
Рис. 3.3. |
Можно предсказать, что моторная кора « homunculus » возникает из-за того, что нейроны, управляющие отдельными мышцами, сгруппированы вместе в коре. То есть все нейроны, которые контролируют двуглавую мышцу, могут быть расположены вместе, и все нейроны, которые контролируют трицепс, могут быть сгруппированы поблизости, а нейроны, которые контролируют камбаловидную мышцу, могут быть сгруппированы в более удаленной области. Однако электрофизиологические записи показали, что это не так. Движения отдельных мышц коррелируют с активностью широко распространенных частей первичной моторной коры. Точно так же стимуляция небольших участков первичной моторной коры вызывает движения, требующие активности множества мышц. Таким образом, первичный гомункул моторной коры не отражает активность отдельных мышц. Скорее, он, по-видимому, представляет собой движения отдельных частей тела, которые часто требуют скоординированной активности больших групп мышц по всему телу.
Однако электрофизиологические записи показали, что это не так. Движения отдельных мышц коррелируют с активностью широко распространенных частей первичной моторной коры. Точно так же стимуляция небольших участков первичной моторной коры вызывает движения, требующие активности множества мышц. Таким образом, первичный гомункул моторной коры не отражает активность отдельных мышц. Скорее, он, по-видимому, представляет собой движения отдельных частей тела, которые часто требуют скоординированной активности больших групп мышц по всему телу.
3.3 Кортикальные аференты и эфференты
Моторная кора оказывает влияние на мышцы множеством нисходящих маршрутов (рис. 3.4). На некоторые из нисходящих путей, рассмотренных в предыдущей главе, может влиять продукция моторной коры. Таким образом, помимо прямой корковой иннервации альфа-мотонейронов через кортикоспинальный тракт, следующие корковые эфферентные пути влияют на остальные нисходящие пути:
- кортикорубральный тракт позволяет коре головного мозга модулировать руброспинальный тракт
- Кортикотектальный тракт позволяет коре головного мозга модулировать тектоспинальный тракт
- Кортикоретикулярный тракт позволяет коре головного мозга модулировать ретикулоспинальные тракты
Рисунок 3. |
 4
4 Кора головного мозга также может влиять на обработку боковых петель моторной иерархии. Кортикостриатный тракт иннервирует хвостатое ядро и скорлупу базальных ганглиев. Кортикопонтинный тракт и кортико-желудочный тракт иннервируют важные входы в мозжечок.Наконец, области коры могут влиять на другие области коры, напрямую через кортикокортикальные пути и косвенно через кортикоталамические пути (рис. 3.5). Большинство этих путей двунаправлены. Таким образом, моторная кора получает входные данные из других областей коры, прямо или косвенно, через таламус, и получает входные данные от мозжечка и базальных ганглиев, всегда через таламус.
Рисунок 3. |
 5
5 3.4 Цитоархитектура моторной коры
Как и все части неокортекса, первичная моторная кора состоит из шести слоев (рис.6). В отличие от первичных сенсорных областей, первичная моторная кора — это агранулярная кора; то есть он не имеет гранулированного слоя с ячейками (слой 4). Вместо этого наиболее отличительным слоем первичной моторной коры является нисходящий выходной слой (слой 5), который содержит гигантские клетки Беца. Эти пирамидные клетки и другие проекционные нейроны первичной моторной коры составляют ~ 30% волокон кортикоспинального тракта. Остальные волокна поступают из премоторной коры и дополнительной моторной области (~ 30%), соматосенсорной коры (~ 30%) и задней теменной коры (~ 10%).
Остальные волокна поступают из премоторной коры и дополнительной моторной области (~ 30%), соматосенсорной коры (~ 30%) и задней теменной коры (~ 10%).
Рисунок 3.6 |
3.5 Кодирование движения моторной корой
Cortex первичного двигателя
Как обсуждалось выше, первичная моторная кора обычно не контролирует отдельные мышцы напрямую, а скорее, по-видимому, контролирует отдельные движения или последовательности движений, которые требуют активности нескольких групп мышц.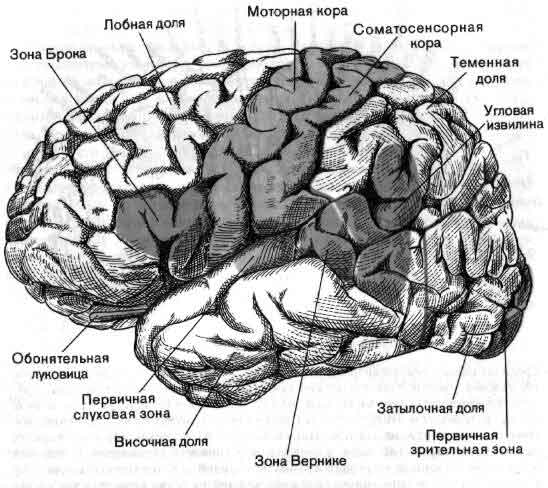 Альфа-мотонейроны в спинном мозге, в свою очередь, кодируют силу сокращения групп мышечных волокон, используя код скорости и принцип размера.Таким образом, в соответствии с концепцией иерархической организации моторной системы информация, представленная моторной корой головного мозга, представляет собой более высокий уровень абстракции, чем информация, представленная моторными нейронами спинного мозга.
Альфа-мотонейроны в спинном мозге, в свою очередь, кодируют силу сокращения групп мышечных волокон, используя код скорости и принцип размера.Таким образом, в соответствии с концепцией иерархической организации моторной системы информация, представленная моторной корой головного мозга, представляет собой более высокий уровень абстракции, чем информация, представленная моторными нейронами спинного мозга.
Что кодируется нейронами первичной моторной коры? Ключ к разгадке пришел из записи активности этих нейронов, когда экспериментальные животные выполняли различные двигательные задачи. В общем, первичная моторная кора кодирует параметры, которые определяют отдельные движения или простые последовательности движений.
- Первичные нейроны моторной коры срабатывают за 5–100 мсек до начала движения. Таким образом, вместо того, чтобы срабатывать в результате мышечной активности, эти нейроны участвуют в передаче моторных команд альфа-мотонейронам, которые в конечном итоге заставляют соответствующие мышцы сокращаться.
- Первичная моторная кора головного мозга кодирует силу движения. Сила, необходимая для поднятия руки из одного места в другое, намного больше, если вы держите шар для боулинга, чем если бы вы держали воздушный шар.Многие нейроны первичной моторной коры кодируют силу, необходимую для такого движения (рис. 3.7). Обратите внимание на различие между силой движения и силой мышц. В то время как меньшая часть нейронов первичной моторной коры кодирует индивидуальную мышечную силу, большее число кодирует количество силы, необходимой для конкретного движения, независимо от того, какие отдельные мышцы используются. Альфа-моторные нейроны, в свою очередь, транслируют команды нейронов моторной коры и контролируют количество силы, генерируемой отдельными мышцами для выполнения этого движения, в соответствии с принципами кода скорости и принципа размера.
Рисунки 3.7A, 3.7B и 3.7C
Моторная кора головного мозга кодирует силу, необходимую для движения. (Эвартс 1968)Рисунок 3.7A. При небольшой нагрузке мотонейрон в первичной моторной коре головного мозга, который контролирует разгибание запястья, срабатывает, когда запястье разгибается. Моторный нейрон, контролирующий сгибание запястья, не меняет своей низкой активности. Обратите внимание, что мотонейрон-удлинитель начинает запускать спайки еще до начала движения.
текст идет сюдаРисунок 3.7B. Когда на левый шкив помещается груз весом 5 фунтов, необходимо приложить больше силы, чтобы сначала удержать груз в устойчивом положении, а затем поднять его. Расширяющийся мотонейрон в первичной моторной коре срабатывает сильнее, создавая большую силу.
текст идет сюдаРисунок 3.
7C. Когда на правый шкив прикладывается нагрузка в 5 фунтов, нагрузка ложится на сгибатель. Таким образом, первичные нейроны моторной коры для сгибания активируются, чтобы поддерживать стабильный вес. Когда запястье разгибается, нейроны работают тише, поскольку сила движения фактически создается самим весом. (Обратите внимание, что моторная кора кодирует силу движения, такого как разгибание запястья или более сложные многосуставные движения. Сила отдельных мышц кодируется альфа-мотонейронами в спинном мозге и стволе головного мозга.)
здесь идет текст - Первичная моторная кора головного мозга кодирует направление движения. Многие нейроны первичной моторной коры избирательны в отношении определенного направления движения. Например, одна ячейка может сильно срабатывать при перемещении руки влево, тогда как при перемещении руки вправо она будет подавлена (рис. 3.8).
Рисунок 3.
8
Направленная настройка нейронов моторной коры. Ячейка срабатывает максимально, когда рука перемещается в направлениях 135º или 180º, умеренно, когда рука движется в направлениях 90º и 225º, и молчит, когда рука движется в противоположных направлениях (0º, 45º, 270º и 315º) ( Георгопулос и др., 1982). - Первичная моторная кора головного мозга кодирует степень движения. Срабатывание некоторых нейронов коррелирует с расстоянием движения.Обезьяна была обучена перемещать руку к разным целевым точкам, которые различались по направлению и расстоянию от центра. Возбуждение многих нейронов коррелировало с направлением движения (как в точке 3), тогда как возбуждение других нейронов коррелировало с расстоянием движения. Интересно, что некоторые нейроны коррелировали с взаимодействием определенного расстояния и направления; то есть они были соотнесены с определенной целевой позицией.
- Первичные нейроны моторной коры кодируют скорость движения. Почти все целевые движения следуют типичной колоколообразной кривой скорости как функции от расстояния (рис. 3.9). Например, когда рука перемещает объект, такой как чашка кофе, из одного места в другое (цель), рука ускоряется в течение первой половины движения, достигает максимальной скорости примерно на полпути к цели, а затем замедляется до тех пор, пока не достигнет цели. достигает цели. Скорость возбуждения некоторых первичных нейронов моторной коры у обезьян коррелирует с этим колоколообразным профилем скорости, демонстрируя, что информация о скорости движения содержится в цепочках шипов этих нейронов.

 7C. Когда на правый шкив прикладывается нагрузка в 5 фунтов, нагрузка ложится на сгибатель. Таким образом, первичные нейроны моторной коры для сгибания активируются, чтобы поддерживать стабильный вес. Когда запястье разгибается, нейроны работают тише, поскольку сила движения фактически создается самим весом. (Обратите внимание, что моторная кора кодирует силу движения, такого как разгибание запястья или более сложные многосуставные движения. Сила отдельных мышц кодируется альфа-мотонейронами в спинном мозге и стволе головного мозга.)
7C. Когда на правый шкив прикладывается нагрузка в 5 фунтов, нагрузка ложится на сгибатель. Таким образом, первичные нейроны моторной коры для сгибания активируются, чтобы поддерживать стабильный вес. Когда запястье разгибается, нейроны работают тише, поскольку сила движения фактически создается самим весом. (Обратите внимание, что моторная кора кодирует силу движения, такого как разгибание запястья или более сложные многосуставные движения. Сила отдельных мышц кодируется альфа-мотонейронами в спинном мозге и стволе головного мозга.) 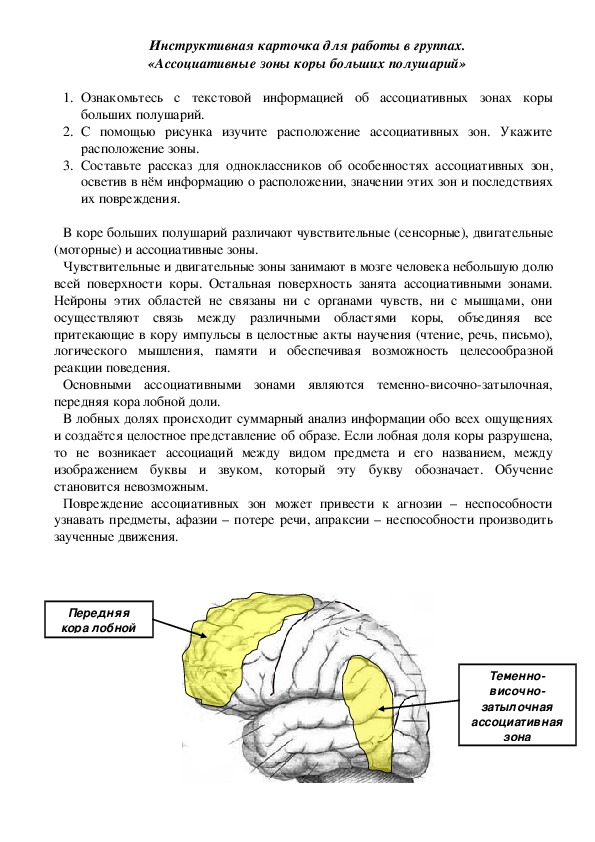 8
8  Почти все целевые движения следуют типичной колоколообразной кривой скорости как функции от расстояния (рис. 3.9). Например, когда рука перемещает объект, такой как чашка кофе, из одного места в другое (цель), рука ускоряется в течение первой половины движения, достигает максимальной скорости примерно на полпути к цели, а затем замедляется до тех пор, пока не достигнет цели. достигает цели. Скорость возбуждения некоторых первичных нейронов моторной коры у обезьян коррелирует с этим колоколообразным профилем скорости, демонстрируя, что информация о скорости движения содержится в цепочках шипов этих нейронов.
Почти все целевые движения следуют типичной колоколообразной кривой скорости как функции от расстояния (рис. 3.9). Например, когда рука перемещает объект, такой как чашка кофе, из одного места в другое (цель), рука ускоряется в течение первой половины движения, достигает максимальной скорости примерно на полпути к цели, а затем замедляется до тех пор, пока не достигнет цели. достигает цели. Скорость возбуждения некоторых первичных нейронов моторной коры у обезьян коррелирует с этим колоколообразным профилем скорости, демонстрируя, что информация о скорости движения содержится в цепочках шипов этих нейронов. Рисунок 3.9 |
Премоторный кортекс
Премоторная кора посылает аксоны в первичную моторную кору, а также непосредственно в спинной мозг. Он выполняет более сложную, связанную с задачами обработку, чем первичная моторная кора. Стимуляция премоторных областей у обезьяны сильным током создает более сложные позы, чем стимуляция первичной моторной коры.Премоторная кора, по-видимому, участвует в выборе подходящих двигательных планов для произвольных движений, тогда как первичная моторная кора участвует в выполнении этих произвольных движений.
Стимуляция премоторных областей у обезьяны сильным током создает более сложные позы, чем стимуляция первичной моторной коры.Премоторная кора, по-видимому, участвует в выборе подходящих двигательных планов для произвольных движений, тогда как первичная моторная кора участвует в выполнении этих произвольных движений.
- Нейроны премоторной коры сигнализируют о подготовке к движению. Обезьяны были обучены совершать определенное движение в ответ на визуальный сигнал с переменной задержкой между началом сигнала и началом движения (рис. 3.10). Записи премоторной коры показали, что многие нейроны избирательно срабатывают в интервале задержки, за много секунд до начала движения.Определенный нейрон срабатывает, когда обезьяна готовится сделать движение влево, например, но будет молчать, когда обезьяна готовится сделать движение вправо. Таким образом, возбуждение этого типа нейрона не вызывает самого движения, но, по-видимому, участвует в подготовке обезьяны к правильному движению при подаче сигнала «Вперёд». Этот тип нейрона называется нейроном с двигательным набором , поскольку он срабатывает, когда обезьяна готовится или собирается совершить движение.
Рис. 3.10.
Подготовка сигнала нейронов премоторной коры к движению. Обезьяну приучают готовиться к движению вправо или влево в зависимости от команды, но с задержкой движения до тех пор, пока не будет дан сигнал «Движение» (Weinrich and Wise, 1982). Некоторые нейроны срабатывают выборочно, когда животное готовится сделать движение вправо (Play Prepare right cell). Другие нейроны будут избирательно срабатывать, когда животное готовится сделать движение влево (Play Prepare left cell).Обратите внимание, что ячейки срабатывают в интервале между инструкциями Prepare и Move, но они не срабатывают во время самого движения. - Нейроны премоторной коры сигнализируют о различных сенсорных аспектах, связанных с определенными двигательными актами. Некоторые премоторные нейроны активируются, когда животное выполняет определенное действие, например ломает арахис (рис. 3.11). Интересно, что тот же нейрон избирательно срабатывает, когда животное видит, что другая обезьяна или человека ломают арахис.Он также срабатывает избирательно под звук ломающейся арахисовой скорлупы, даже без какой-либо зрительной или двигательной активности. Эти нейроны называются «зеркальными» нейронами , потому что они реагируют не только на конкретное действие обезьяны, но и на взгляд (или звук) другого человека, выполняющего то же действие. (Интересное видео PBS о зеркальных нейронах см. На http://www.pbs.org/wgbh/nova/sciencenow/3204/01.html.)
Рисунок 3.11
Зеркальный нейрон в премоторной коре головного мозга реагирует на действия обезьяны, а также на восприятие обезьяной человека, выполняющего то же действие (Kohler et al., 2002). - Премоторная кора головного мозга чувствительна к поведенческому контексту определенного движения. Премоторная кора головного мозга людей была визуализирована с помощью функциональной МРТ, когда они наблюдали видео, на котором рука держит чашку (рис. 3.12). В одном случае чаша была полна и окружена полными тарелками с едой; Подразумевалось, что человек схватился за чашку, чтобы сделать глоток.В другом случае чашка была пуста и окружена грязной посудой; Подразумевалось, что человек схватился за чашку, чтобы убрать со стола. В этом эксперименте премоторная кора головного мозга была более активной, когда испытуемые просматривали первое видео, чем второе, хотя движения были одинаковыми. Таким образом, нейроны премоторной коры чувствительны к предполагаемым намерениям движения, а не только к самому движению, как следует из поведенческого контекста, в котором движение произошло.
Рисунок 3.12
Активность премоторной коры различает одно и то же движение на основе поведенческого контекста движения (Iacoboni et al. , 2005). Когда субъект видел руку, движущуюся, чтобы взять чашку для питья (ИГРАТЬ вверху), активность в премоторной коре головного мозга была выше, чем когда он видел руку, движущуюся, чтобы взять чашку, чтобы убрать со стола после еды (ИГРАТЬ внизу) . Обратите внимание, что сила активности коры головного мозга (обозначается яркостью активированной кортикальной области) в верхней части анимации выше, чем в нижней. - Премоторная кора сигнализирует о правильных и неправильных действиях. Люди были изучены в эксперименте фМРТ, поскольку они наблюдали видеоклипы различных правильных и неправильных двигательных действий. Правильное действие — это действие, при котором движение и связанный с ним объект выполнялись правильно, например, установка времени на часах. Ошибка объекта — это ошибка, при которой действие было правильным, но объект был неправильным, например, полировка коричневой обуви черным кремом для обуви.Ошибка движения — это ошибка, при которой объект был правильным, но движение было неправильным, например, попытка положить монету в копилку, когда монета была ориентирована перпендикулярно прорези для монет. В этом эксперименте премоторная кора головного мозга активировалась с двух сторон во время испытаний правильных действий и ошибок движения; при испытаниях ошибок объекта преимущественно активировалась только премоторная кора левого полушария.
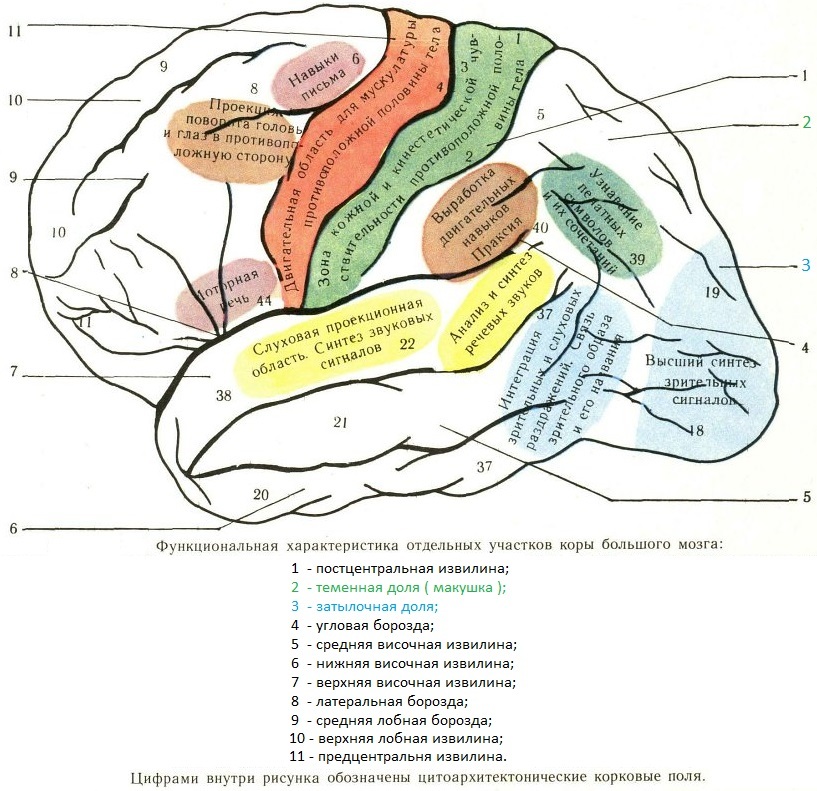 Этот тип нейрона называется нейроном с двигательным набором , поскольку он срабатывает, когда обезьяна готовится или собирается совершить движение.
Этот тип нейрона называется нейроном с двигательным набором , поскольку он срабатывает, когда обезьяна готовится или собирается совершить движение.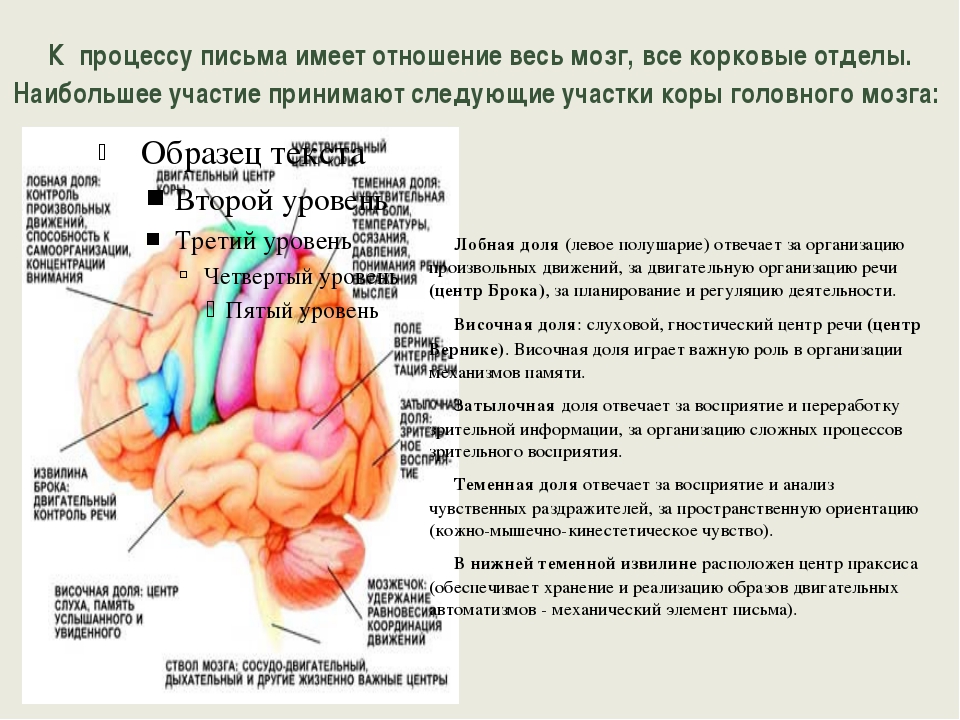 Некоторые премоторные нейроны активируются, когда животное выполняет определенное действие, например ломает арахис (рис. 3.11). Интересно, что тот же нейрон избирательно срабатывает, когда животное видит, что другая обезьяна или человека ломают арахис.Он также срабатывает избирательно под звук ломающейся арахисовой скорлупы, даже без какой-либо зрительной или двигательной активности. Эти нейроны называются «зеркальными» нейронами , потому что они реагируют не только на конкретное действие обезьяны, но и на взгляд (или звук) другого человека, выполняющего то же действие. (Интересное видео PBS о зеркальных нейронах см. На http://www.pbs.org/wgbh/nova/sciencenow/3204/01.html.)
Некоторые премоторные нейроны активируются, когда животное выполняет определенное действие, например ломает арахис (рис. 3.11). Интересно, что тот же нейрон избирательно срабатывает, когда животное видит, что другая обезьяна или человека ломают арахис.Он также срабатывает избирательно под звук ломающейся арахисовой скорлупы, даже без какой-либо зрительной или двигательной активности. Эти нейроны называются «зеркальными» нейронами , потому что они реагируют не только на конкретное действие обезьяны, но и на взгляд (или звук) другого человека, выполняющего то же действие. (Интересное видео PBS о зеркальных нейронах см. На http://www.pbs.org/wgbh/nova/sciencenow/3204/01.html.)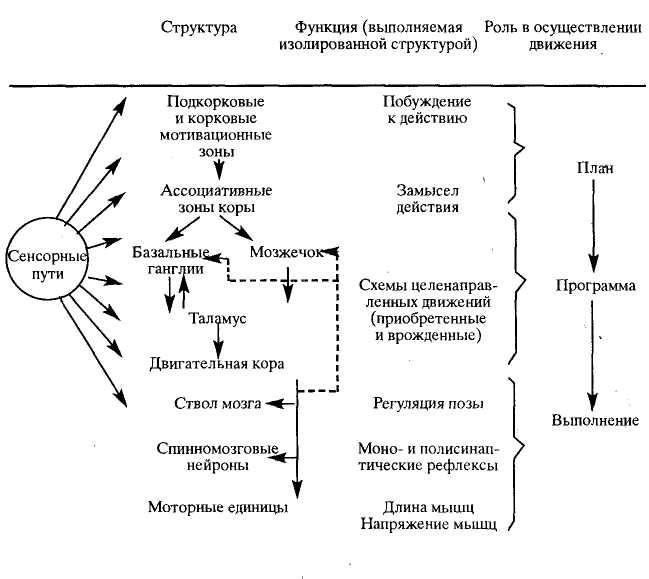 Премоторная кора головного мозга людей была визуализирована с помощью функциональной МРТ, когда они наблюдали видео, на котором рука держит чашку (рис. 3.12). В одном случае чаша была полна и окружена полными тарелками с едой; Подразумевалось, что человек схватился за чашку, чтобы сделать глоток.В другом случае чашка была пуста и окружена грязной посудой; Подразумевалось, что человек схватился за чашку, чтобы убрать со стола. В этом эксперименте премоторная кора головного мозга была более активной, когда испытуемые просматривали первое видео, чем второе, хотя движения были одинаковыми. Таким образом, нейроны премоторной коры чувствительны к предполагаемым намерениям движения, а не только к самому движению, как следует из поведенческого контекста, в котором движение произошло.
Премоторная кора головного мозга людей была визуализирована с помощью функциональной МРТ, когда они наблюдали видео, на котором рука держит чашку (рис. 3.12). В одном случае чаша была полна и окружена полными тарелками с едой; Подразумевалось, что человек схватился за чашку, чтобы сделать глоток.В другом случае чашка была пуста и окружена грязной посудой; Подразумевалось, что человек схватился за чашку, чтобы убрать со стола. В этом эксперименте премоторная кора головного мозга была более активной, когда испытуемые просматривали первое видео, чем второе, хотя движения были одинаковыми. Таким образом, нейроны премоторной коры чувствительны к предполагаемым намерениям движения, а не только к самому движению, как следует из поведенческого контекста, в котором движение произошло. , 2005). Когда субъект видел руку, движущуюся, чтобы взять чашку для питья (ИГРАТЬ вверху), активность в премоторной коре головного мозга была выше, чем когда он видел руку, движущуюся, чтобы взять чашку, чтобы убрать со стола после еды (ИГРАТЬ внизу) . Обратите внимание, что сила активности коры головного мозга (обозначается яркостью активированной кортикальной области) в верхней части анимации выше, чем в нижней.
, 2005). Когда субъект видел руку, движущуюся, чтобы взять чашку для питья (ИГРАТЬ вверху), активность в премоторной коре головного мозга была выше, чем когда он видел руку, движущуюся, чтобы взять чашку, чтобы убрать со стола после еды (ИГРАТЬ внизу) . Обратите внимание, что сила активности коры головного мозга (обозначается яркостью активированной кортикальной области) в верхней части анимации выше, чем в нижней. В этом эксперименте премоторная кора головного мозга активировалась с двух сторон во время испытаний правильных действий и ошибок движения; при испытаниях ошибок объекта преимущественно активировалась только премоторная кора левого полушария.
В этом эксперименте премоторная кора головного мозга активировалась с двух сторон во время испытаний правильных действий и ошибок движения; при испытаниях ошибок объекта преимущественно активировалась только премоторная кора левого полушария.Дополнительная моторная зона
Дополнительная моторная зона (SMA) участвует в программировании сложных последовательностей движений и координации двусторонних движений.В то время как премоторная кора, по-видимому, участвует в выборе моторных программ на основе зрительных стимулов или абстрактных ассоциаций, дополнительная моторная область, по-видимому, участвует в выборе движений на основе запомненных последовательностей движений.
- SMA реагирует на последовательности движений и на мысленную репетицию последовательности движений (рис. 3.13). Активность мозга измерялась с помощью ПЭТ-сканера, в то время как испытуемые выполняли простые и сложные последовательности движений. Когда движения были простыми, такими как повторяющееся движение одного пальца, первичная моторная кора и первичная соматосенсорная кора были активированы на контралатеральном полушарии. Когда испытуемого просили выполнить сложную последовательность движений пальцев, SMA активировалась с обеих сторон, в дополнение к активации контрлатеральной первичной моторной и соматосенсорной коры. Наконец, когда испытуемого попросили оставаться на месте, но мысленно репетировать сложную последовательность действий, SMA все еще была активна, хотя первичная моторная и соматосенсорная области коры головного мозга молчали. Таким образом, SMA, по-видимому, участвует в двусторонних движениях и в мысленной репетиции этих движений.
Рис. 3.13. Исследование
позитронно-эмиссионной томографией (ПЭТ) простых и сложных движений пальцев (Roland et al., 1980). SMA активируется с обеих сторон, когда субъекты выполняют сложные движения, и даже когда они только воображают выполнение этих движений. - SMA участвует в преобразовании кинематической информации в динамическую. Движения могут быть определены в терминах динамики , (количество силы, необходимой для выполнения движения) и кинематики , (расстояние и углы, которые определяют конкретное движение в пространстве). Многие планы движения представлены в кинематических терминах (например, «Переместите руку влево»). Однако двигательная система должна в конечном итоге преобразовать это в представление, основанное на динамике, чтобы дать указание соответствующим мышцам сокращаться с соответствующей силой. Записи, сделанные обезьянами, показали, что во время подготовительной задержки перед тем, как обезьяна совершит проинструктированное движение, некоторые нейроны SMA меняют свои корреляты срабатывания с кинематического представления на динамическое представление, предполагая, что SMA играет жизненно важную роль в этом преобразовании.
 Когда испытуемого просили выполнить сложную последовательность движений пальцев, SMA активировалась с обеих сторон, в дополнение к активации контрлатеральной первичной моторной и соматосенсорной коры. Наконец, когда испытуемого попросили оставаться на месте, но мысленно репетировать сложную последовательность действий, SMA все еще была активна, хотя первичная моторная и соматосенсорная области коры головного мозга молчали. Таким образом, SMA, по-видимому, участвует в двусторонних движениях и в мысленной репетиции этих движений.
Когда испытуемого просили выполнить сложную последовательность движений пальцев, SMA активировалась с обеих сторон, в дополнение к активации контрлатеральной первичной моторной и соматосенсорной коры. Наконец, когда испытуемого попросили оставаться на месте, но мысленно репетировать сложную последовательность действий, SMA все еще была активна, хотя первичная моторная и соматосенсорная области коры головного мозга молчали. Таким образом, SMA, по-видимому, участвует в двусторонних движениях и в мысленной репетиции этих движений.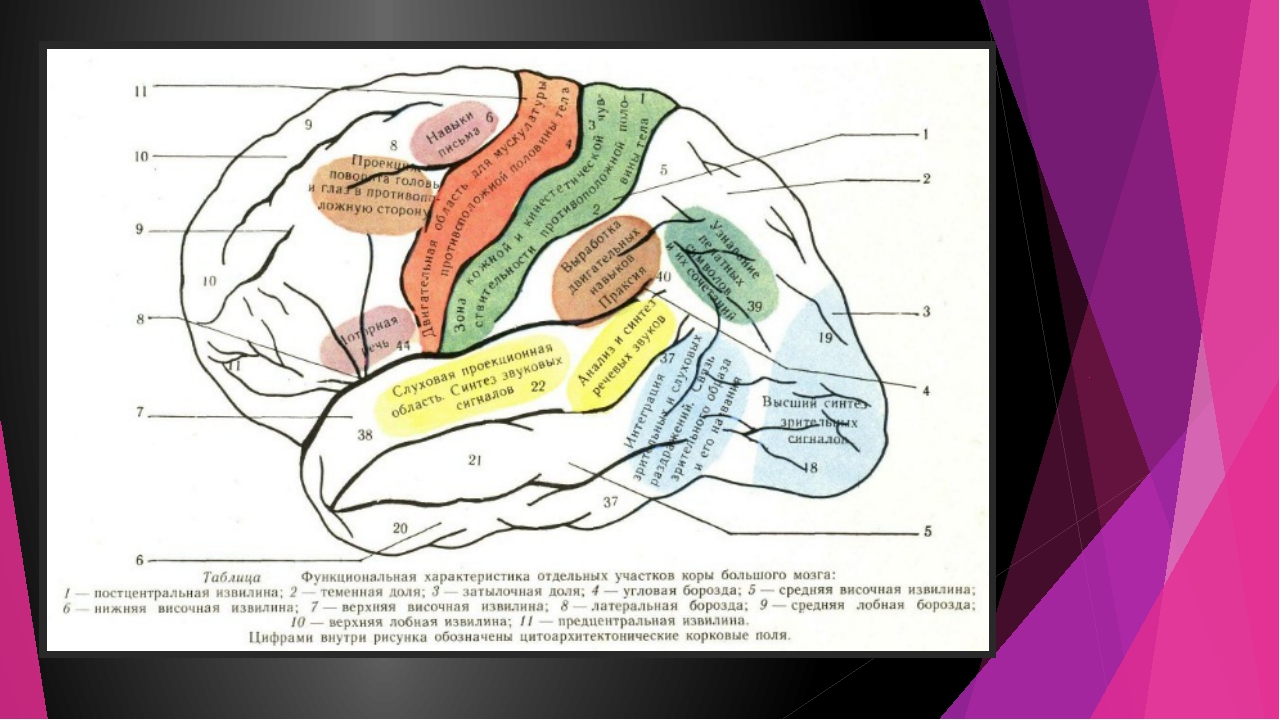 Движения могут быть определены в терминах динамики , (количество силы, необходимой для выполнения движения) и кинематики , (расстояние и углы, которые определяют конкретное движение в пространстве). Многие планы движения представлены в кинематических терминах (например, «Переместите руку влево»). Однако двигательная система должна в конечном итоге преобразовать это в представление, основанное на динамике, чтобы дать указание соответствующим мышцам сокращаться с соответствующей силой. Записи, сделанные обезьянами, показали, что во время подготовительной задержки перед тем, как обезьяна совершит проинструктированное движение, некоторые нейроны SMA меняют свои корреляты срабатывания с кинематического представления на динамическое представление, предполагая, что SMA играет жизненно важную роль в этом преобразовании.
Движения могут быть определены в терминах динамики , (количество силы, необходимой для выполнения движения) и кинематики , (расстояние и углы, которые определяют конкретное движение в пространстве). Многие планы движения представлены в кинематических терминах (например, «Переместите руку влево»). Однако двигательная система должна в конечном итоге преобразовать это в представление, основанное на динамике, чтобы дать указание соответствующим мышцам сокращаться с соответствующей силой. Записи, сделанные обезьянами, показали, что во время подготовительной задержки перед тем, как обезьяна совершит проинструктированное движение, некоторые нейроны SMA меняют свои корреляты срабатывания с кинематического представления на динамическое представление, предполагая, что SMA играет жизненно важную роль в этом преобразовании.Ассоциация Cortex
Четвертый уровень моторной иерархии — это ассоциативная кора , в частности префронтальная кора и задняя теменная кора (рис. 3.14). Эти области мозга не являются моторными в строгом смысле слова. Их активность не коррелирует в точности с отдельными двигательными актами, и стимуляция этих областей не приводит к двигательной активности. Однако эти области необходимы, чтобы гарантировать, что движения адаптируются к потребностям организма и соответствуют поведенческому контексту.
3.14). Эти области мозга не являются моторными в строгом смысле слова. Их активность не коррелирует в точности с отдельными двигательными актами, и стимуляция этих областей не приводит к двигательной активности. Однако эти области необходимы, чтобы гарантировать, что движения адаптируются к потребностям организма и соответствуют поведенческому контексту.
Рисунок 3.14 |
- Задняя теменная кора участвует в обеспечении точного нацеливания движений на объекты во внешнем пространстве. Эта область участвует в обработке пространственных отношений объектов в мире и в построении представления внешнего пространства, которое не зависит от положения глаз или положения тела наблюдателя.Такие представления позволяют стабильно воспринимать мир, не зависящее от ориентации зрителя, а также отображать желаемые траектории в пространстве, не зависящие от положения тела. Повреждение задней теменной коры может привести к ряду апраксий, , то есть к невозможности совершать сложные скоординированные движения. Например, пациент с конструкционной апраксией не может воспроизвести конфигурацию набора блоков в правильной последовательности, даже если пациент может маневрировать каждым блоком индивидуально с ловкостью.
- Префронтальная кора участвует в выборе подходящих действий для определенного поведенческого контекста. Он также участвует в оценке последствий определенного образа действий. Пациенты с повреждением префронтальной коры имеют проблемы с исполнительной обработкой. Они принимают неправильные поведенческие решения и часто не могут предвидеть вероятных последствий своих действий. Они демонстрируют импульсивное поведение, часто демонстрируя неспособность отложить мгновенное вознаграждение ради более долгосрочного вознаграждения.
 Повреждение задней теменной коры может привести к ряду апраксий, , то есть к невозможности совершать сложные скоординированные движения. Например, пациент с конструкционной апраксией не может воспроизвести конфигурацию набора блоков в правильной последовательности, даже если пациент может маневрировать каждым блоком индивидуально с ловкостью.
Повреждение задней теменной коры может привести к ряду апраксий, , то есть к невозможности совершать сложные скоординированные движения. Например, пациент с конструкционной апраксией не может воспроизвести конфигурацию набора блоков в правильной последовательности, даже если пациент может маневрировать каждым блоком индивидуально с ловкостью. Проверьте свои знания
Ячейки Беца наиболее многочисленны в слое . ..
..
A. IV соматосенсорной коры.
Б. В. соматосенсорной коры.
C. IV моторной коры.
D. V моторной коры.
E. III моторной коры.
Ячейки Беца наиболее многочисленны в слое…
A. IV соматосенсорной коры. Это НЕПРАВИЛЬНЫЙ ответ.
клеток Беца нет в соматосенсорной коре.
Б. В. соматосенсорной коры.
C. IV моторной коры.
D. V моторной коры.
E. III моторной коры.
Ячейки Беца наиболее многочисленны в слое …
А.IV соматосенсорной коры.
Б. В. соматосенсорной коры. Это НЕПРАВИЛЬНЫЙ ответ.
клеток Беца нет в соматосенсорной коре.
C.
D. V моторной коры.
E. III моторной коры.
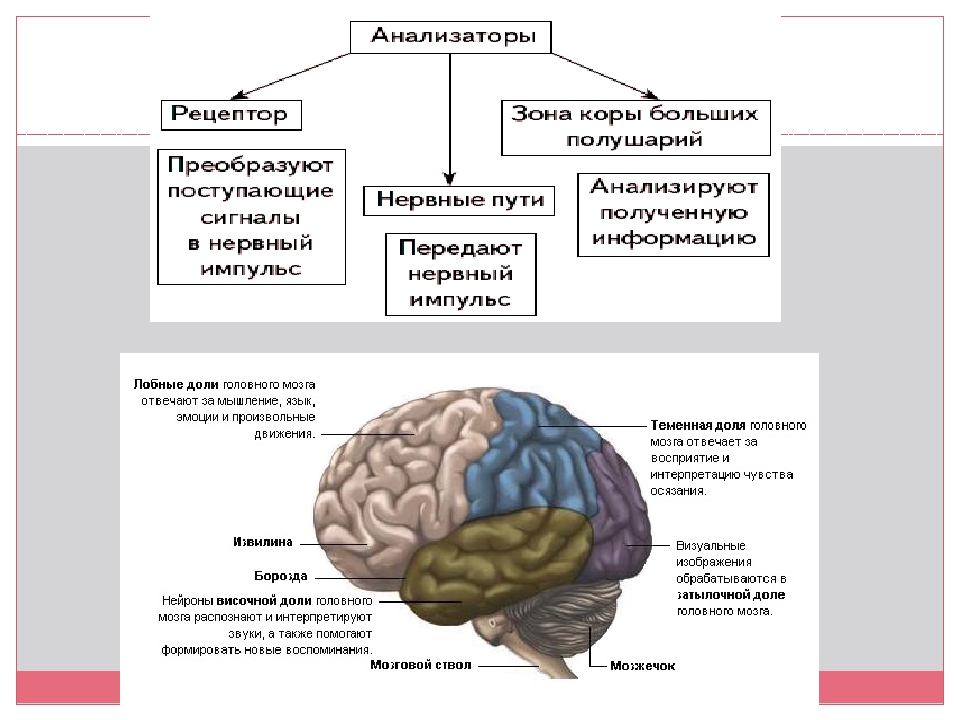 IV моторной коры.
IV моторной коры.Ячейки Беца наиболее многочисленны в слое …
A. IV соматосенсорной коры.
Б. В. соматосенсорной коры.
C. IV моторной коры. Это НЕПРАВИЛЬНЫЙ ответ.
клетки Беца не находятся в слое IV.
D. V моторной коры.
E. III моторной коры.
Ячейки Беца наиболее многочисленны в слое …
A. IV соматосенсорной коры.
Б.V соматосенсорной коры.
C. IV моторной коры.
D. V моторной коры. Это ПРАВИЛЬНЫЙ ответ!
E. III моторной коры.
Ячейки Беца наиболее многочисленны в слое .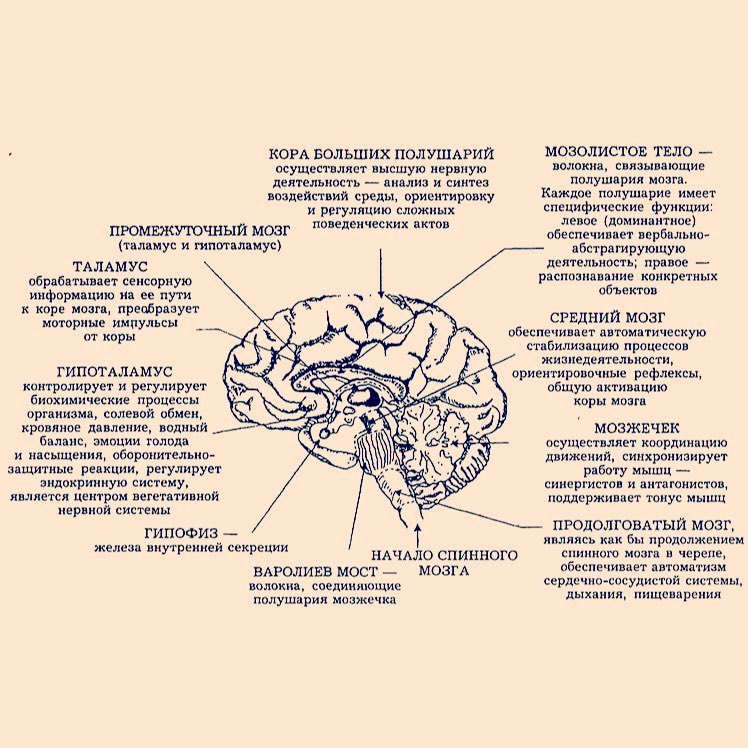 ..
..
A. IV соматосенсорной коры.
Б. В. соматосенсорной коры.
C. IV моторной коры.
Д.V моторной коры.
E. III моторной коры. Это НЕПРАВИЛЬНЫЙ ответ.
Ячейки Беца не находятся в слое III.
Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
А.Проецируйте на несколько пулов двигательных нейронов в спинном мозге.
B. Участвовать в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E. Код скорости движения.
Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
А.
Это ИСТИНА. На множество различных групп мышц влияет активность отдельных нейронов моторной коры.
B. Участвовать в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E. Код скорости движения.
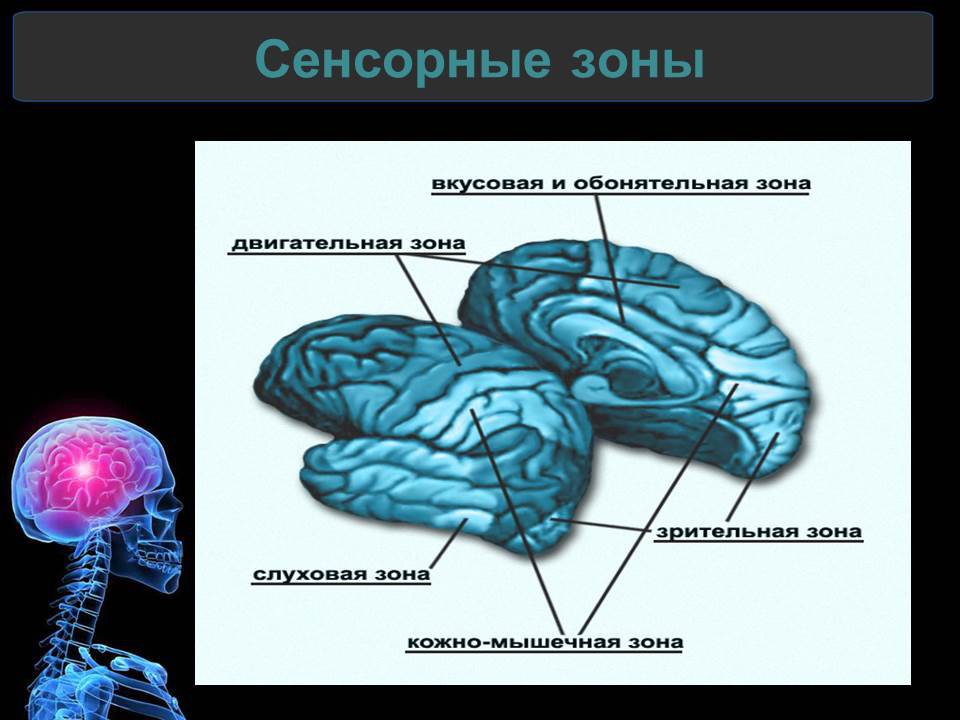 Проецируйте на несколько пулов двигательных нейронов в спинном мозге. Это НЕПРАВИЛЬНЫЙ ответ.
Проецируйте на несколько пулов двигательных нейронов в спинном мозге. Это НЕПРАВИЛЬНЫЙ ответ.Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
A. Проект для нескольких пулов двигательных нейронов в спинном мозге.
B. Участвовать в инициации движения. Это НЕПРАВИЛЬНЫЙ ответ.
Это ИСТИНА.
C. Код силы отдельных мышц.
Д.Код направления движения.
E. Код скорости движения.
Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
A.
B. Участвовать в инициации движения.
C. Код силы отдельных мышц.Это ПРАВИЛЬНЫЙ ответ!
Это ЛОЖНЫЙ оператор. Нейроны моторной коры кодируют силу отдельных движений, а не отдельных мышц. Нижние двигательные нейроны (альфа-двигательные нейроны) кодируют силу отдельных мышц.
D. Код направления движения.
E. Код скорости движения.
 Проект для нескольких пулов двигательных нейронов в спинном мозге.
Проект для нескольких пулов двигательных нейронов в спинном мозге.Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
А.Проецируйте на несколько пулов двигательных нейронов в спинном мозге.
B. Участвовать в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения. Это НЕПРАВИЛЬНЫЙ ответ.
Это ИСТИНА.
E.
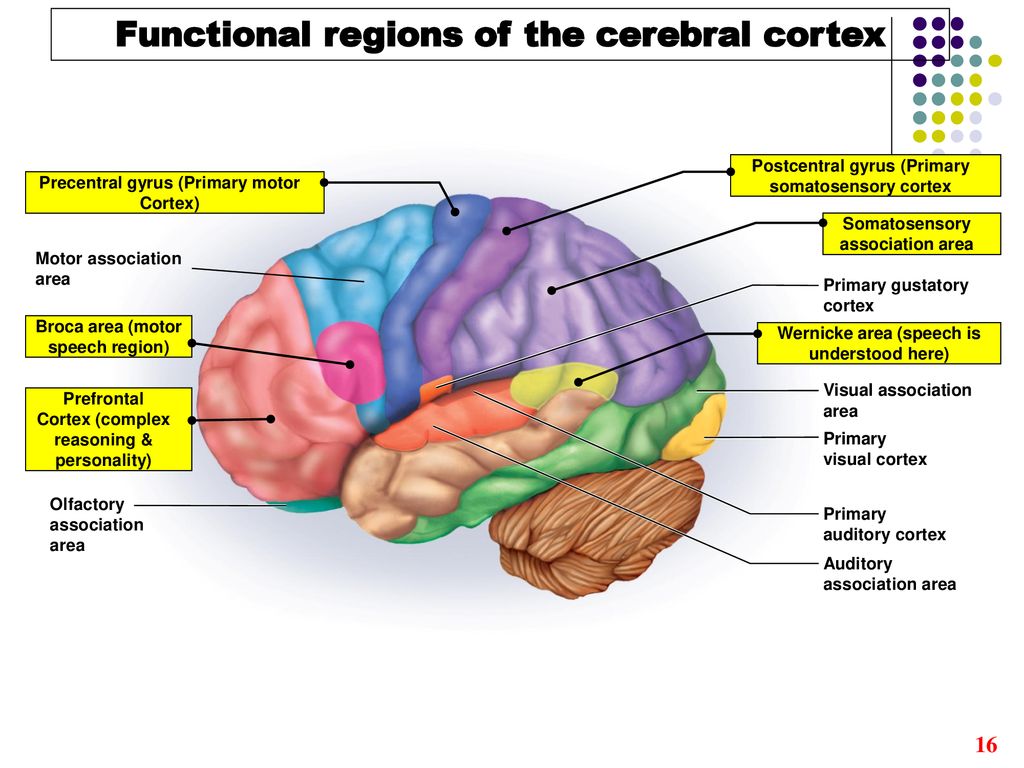 Код скорости движения.
Код скорости движения.Кортикоспинальный нейрон в первичной моторной коре может выполнять все следующие функции, ЗА ИСКЛЮЧЕНИЕМ:
А.Проецируйте на несколько пулов двигательных нейронов в спинном мозге.
B. Участвовать в инициации движения.
C. Код силы отдельных мышц.
D. Код направления движения.
E. Код скорости движения. Это НЕПРАВИЛЬНЫЙ ответ.
Это ИСТИНА.
Ассоциативное обучение и гиппокамп
Каждый день мы изучаем и запоминаем огромное количество новой информации, начиная с имен новых людей, с которыми мы встречаемся, и заканчивая лучшим блюдом, которое можно заказать в определенном ресторане, и местом расположения новой велосипедной дорожки. Эта новая информация сначала приобретается, и, если она усиливается посредством процесса, называемого консолидацией, в конечном итоге сохраняется в долговременной памяти. Структуры медиальной височной доли необходимы для этой способности приобретать новые долговременные воспоминания о фактах и событиях. Эта форма памяти называется декларативной памятью у людей и реляционной памятью у животных. Сходящиеся результаты нейропсихологических исследований на людях, вместе с экспериментальными исследованиями поражений и нейроанатомическими исследованиями на животных показали, что ключевые медиальные структуры височных долей, важные для декларативной / реляционной памяти, включают гиппокамп вместе с окружающей энторинальной, периринальной и парагиппокампальной корой.Хотя ясно, что структуры медиальной височной доли важны для приобретения новых декларативных / реляционных воспоминаний, у нас все еще есть только рудиментарное понимание нормальных паттернов нейронной активности, которые лежат в основе этой способности.
Эта новая информация сначала приобретается, и, если она усиливается посредством процесса, называемого консолидацией, в конечном итоге сохраняется в долговременной памяти. Структуры медиальной височной доли необходимы для этой способности приобретать новые долговременные воспоминания о фактах и событиях. Эта форма памяти называется декларативной памятью у людей и реляционной памятью у животных. Сходящиеся результаты нейропсихологических исследований на людях, вместе с экспериментальными исследованиями поражений и нейроанатомическими исследованиями на животных показали, что ключевые медиальные структуры височных долей, важные для декларативной / реляционной памяти, включают гиппокамп вместе с окружающей энторинальной, периринальной и парагиппокампальной корой.Хотя ясно, что структуры медиальной височной доли важны для приобретения новых декларативных / реляционных воспоминаний, у нас все еще есть только рудиментарное понимание нормальных паттернов нейронной активности, которые лежат в основе этой способности. Чтобы ответить на этот вопрос, моя лаборатория записала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной / реляционной памяти, называемой ассоциативной памятью, которая определяется как способность узнавать и запоминать отношения между несвязанными элементами, такими как имя человека, которого мы только что встретили, или аромат определенных духов.В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению. Наша долгосрочная цель — понять эволюцию сигналов, связанных с обучением, в средней височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
Чтобы ответить на этот вопрос, моя лаборатория записала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной / реляционной памяти, называемой ассоциативной памятью, которая определяется как способность узнавать и запоминать отношения между несвязанными элементами, такими как имя человека, которого мы только что встретили, или аромат определенных духов.В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению. Наша долгосрочная цель — понять эволюцию сигналов, связанных с обучением, в средней височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
Задача, требующая нового ассоциативного подчинения
Чтобы изучить закономерности нейронной активности во время формирования ассоциативной памяти, мы обучили двух обезьян выполнять задачу ассоциации место-сцена. В этом задании животные должны были изучить новые ассоциации между определенными сложными визуальными «сценами» и конкретными целевыми локациями, за которые вознаграждались. Мы знаем, что медиальная височная доля играет важную роль в нормальном выполнении этой задачи, поскольку повреждение этой области у обезьян приводит к значительному ухудшению способности изучать новые ассоциации местоположения и сцены (Brasted et al., 2003; Брастед и др., 2002; Мюррей и др., 2000; Мудрый и Мюррей, 1999; Мюррей и Уайз, 1996; Рупняк и Гаффан, 1987). Схематическое изображение задачи показано на Рисунке 1 . В каждом испытании обезьянам сначала показывают 4 идентичных целевых стимула, наложенных на сложную визуальную сцену (обычно изображение реальной сцены на открытом воздухе). После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают команду сделать одно движение глаза к одной из четырех периферийных целей на экране.Для каждой визуальной сцены только одна из 4 целей связана с наградой за сок. Каждый день животные методом проб и ошибок выучивали 2–4 новых ассоциации между местом и сценой. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонными» ассоциациями, которые животные наблюдали за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с разным награжденным целевым местоположением (т. Е. Севером, югом, востоком или западом). Ответы на эталонные сцены использовались для контроля возможной двигательной активности клеток гиппокампа.
После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают команду сделать одно движение глаза к одной из четырех периферийных целей на экране.Для каждой визуальной сцены только одна из 4 целей связана с наградой за сок. Каждый день животные методом проб и ошибок выучивали 2–4 новых ассоциации между местом и сценой. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонными» ассоциациями, которые животные наблюдали за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с разным награжденным целевым местоположением (т. Е. Севером, югом, востоком или западом). Ответы на эталонные сцены использовались для контроля возможной двигательной активности клеток гиппокампа.
Паттерны активности гиппокампа во время изучения новых ассоциаций локации и сцены
В наших первоначальных исследованиях мы сосредоточились на гиппокампе, структуре медиальной височной доли, долгое время участвовавшей в ассоциативном обучении и памяти (Eichenbaum and Cohen, 2001; Squire and Zola , 1996; Сковилл, Милнер, 1957). Сначала мы спросили, реагируют ли клетки гиппокампа по-разному на различные визуальные сцены, используемые в задаче. Мы обнаружили, что 61% изолированных клеток гиппокампа были задействованы в этой задаче, поскольку они по-разному реагировали на разные сцены (т.е., визуально избирательный ответ). Более того, в соответствии с нашей рабочей гипотезой, мы обнаружили, что 28% избирательно отвечающих клеток (18% от всей зарегистрированной популяции клеток гиппокампа) демонстрируют изменения в нейронной активности в ходе испытаний, которые в значительной степени коррелировали с кривой поведенческого обучения животного для конкретного случая. место действия. Мы назвали эти клетки «изменяющимися клетками». Наблюдались две категории изменяющихся ячеек. Устойчиво изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении с изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа).Многие из этих клеток продемонстрировали резкое повышение нейронной активности, которое соответствовало кривой поведенческого обучения животного для этой ассоциации (рис.
Сначала мы спросили, реагируют ли клетки гиппокампа по-разному на различные визуальные сцены, используемые в задаче. Мы обнаружили, что 61% изолированных клеток гиппокампа были задействованы в этой задаче, поскольку они по-разному реагировали на разные сцены (т.е., визуально избирательный ответ). Более того, в соответствии с нашей рабочей гипотезой, мы обнаружили, что 28% избирательно отвечающих клеток (18% от всей зарегистрированной популяции клеток гиппокампа) демонстрируют изменения в нейронной активности в ходе испытаний, которые в значительной степени коррелировали с кривой поведенческого обучения животного для конкретного случая. место действия. Мы назвали эти клетки «изменяющимися клетками». Наблюдались две категории изменяющихся ячеек. Устойчиво изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении с изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа).Многие из этих клеток продемонстрировали резкое повышение нейронной активности, которое соответствовало кривой поведенческого обучения животного для этой ассоциации (рис. 2A ). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что изменяющаяся ячейка обычно меняла свою активность только для одной конкретной заученной сцены, в то время как реакции на другие заученные сцены не менялись с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением.Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т. Е. Обучением реагировать на север). Согласно этой интерпретации, ранние правильные испытания, возможно, не вызвали большой активности, если бы движения делались ориентировочно, но сильная двигательная активность наблюдается, когда животное начинает последовательно реагировать на север. Если бы эта основанная на моторе интерпретация была правильной, мы бы ожидали увидеть аналогичные уровни активности в этой ячейке в ответ на эталонную сцену с таким же целевым местоположением, награждаемым севером.
2A ). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что изменяющаяся ячейка обычно меняла свою активность только для одной конкретной заученной сцены, в то время как реакции на другие заученные сцены не менялись с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением.Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т. Е. Обучением реагировать на север). Согласно этой интерпретации, ранние правильные испытания, возможно, не вызвали большой активности, если бы движения делались ориентировочно, но сильная двигательная активность наблюдается, когда животное начинает последовательно реагировать на север. Если бы эта основанная на моторе интерпретация была правильной, мы бы ожидали увидеть аналогичные уровни активности в этой ячейке в ответ на эталонную сцену с таким же целевым местоположением, награждаемым севером. Такого не было. Фактически, смена ячеек обычно реагировала с небольшой активностью или отсутствием активности на эталонную сцену с тем же целевым местоположением с вознаграждением. Эти результаты подтверждают идею о том, что изменяющаяся деятельность связана с изучением новой ассоциации между сценой и движением цели / глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные усвоили 2 последовательных набора новых ассоциаций между местом и сценой.Мы обнаружили, что изменение ячеек, идентифицированных в первом наборе ассоциаций изученного местоположения и сцены, никогда не сигнализировало об обучении второго нового набора ассоциаций местоположения и сцены, даже если полученное целевое местоположение было таким же (то есть, северная цель была вознаграждена).
Такого не было. Фактически, смена ячеек обычно реагировала с небольшой активностью или отсутствием активности на эталонную сцену с тем же целевым местоположением с вознаграждением. Эти результаты подтверждают идею о том, что изменяющаяся деятельность связана с изучением новой ассоциации между сценой и движением цели / глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные усвоили 2 последовательных набора новых ассоциаций между местом и сценой.Мы обнаружили, что изменение ячеек, идентифицированных в первом наборе ассоциаций изученного местоположения и сцены, никогда не сигнализировало об обучении второго нового набора ассоциаций местоположения и сцены, даже если полученное целевое местоположение было таким же (то есть, северная цель была вознаграждена).
Остальные 45% изменяющихся клеток продемонстрировали другую модель учебной деятельности. Эти изменяющиеся клетки начинались с избирательной реакции во время сцены или периода задержки задачи в начале сеанса, задолго до того, как животное узнало ассоциацию. Эти клетки сигнализировали об обучении, возвращаясь к исходной активности, и это возвращение к исходному уровню обычно было антикоррелировано с кривой обучения животного для этой конкретной сцены (рис. 2B ). Мы назвали эти клетки базовыми устойчивыми изменяющимися клетками. Важно отметить, что изменения в нейронной активности, наблюдаемые в базовых устойчивых клетках, были столь же избирательными для конкретной усвоенной сцены, как и в устойчиво изменяющихся клетках. Подобные модели активности никогда не наблюдались для эталонной сцены с соответствующим вознагражденным целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Эти клетки сигнализировали об обучении, возвращаясь к исходной активности, и это возвращение к исходному уровню обычно было антикоррелировано с кривой обучения животного для этой конкретной сцены (рис. 2B ). Мы назвали эти клетки базовыми устойчивыми изменяющимися клетками. Важно отметить, что изменения в нейронной активности, наблюдаемые в базовых устойчивых клетках, были столь же избирательными для конкретной усвоенной сцены, как и в устойчиво изменяющихся клетках. Подобные модели активности никогда не наблюдались для эталонной сцены с соответствующим вознагражденным целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Таким образом, как устойчивые изменяющиеся ячейки, так и базовые устойчивые изменяющиеся ячейки обеспечивают высокоселективный сигнал, когда изучается конкретная сцена. Мы предполагаем, что эти избирательные увеличения и уменьшения нейронной активности, которые происходят в популяции гиппокампа, могут составлять сигнал обучения гиппокампальной сети.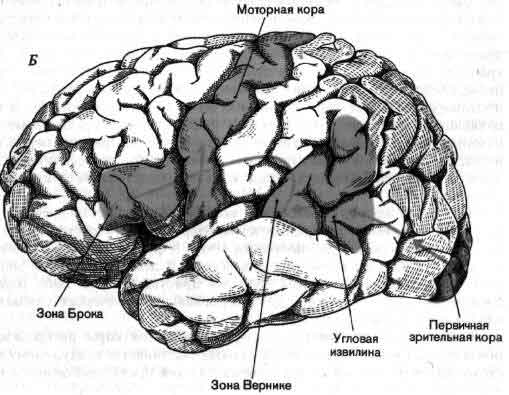 Будет важно изучить обучающие сигналы по большему количеству клеток гиппокампа, записываемых одновременно, чтобы лучше понять взаимодействия между клетками во время обучения.
Будет важно изучить обучающие сигналы по большему количеству клеток гиппокампа, записываемых одновременно, чтобы лучше понять взаимодействия между клетками во время обучения.
Время активности гиппокампа во время изучения новых ассоциаций местоположения и сцены
Критическим вопросом для любого исследования, изучающего нейронные корреляты поведения, является определение причинной связи между наблюдаемыми паттернами нейронной активности и поведенческим выходом. Является ли наблюдаемая нейронная активность движущей силой поведения или она происходит ниже по течению от критических исходных узлов? Для решения этой проблемы использовались самые разные подходы. Например, как упоминалось выше, исследования поражений могут вовлекать конкретную область мозга в нормальное выполнение задачи, хотя одни исследования поражений не могут определить паттерны нейронной активности, лежащие в основе этой функции.Исследования электрической стимуляции использовались для проверки влияния прямой стимуляции на выбор животного при проблемах сенсорной дискриминации (Salzman et al. , 1992; Salzman et al., 1990), но этот подход не использовался в исследованиях функции гиппокампа. Третий метод, который использовался для исследования взаимосвязи между нейронной активностью и поведением, заключается в изучении точного времени изменений нейронной активности относительно поведенческого обучения. Мы предполагаем, что те избирательные нейронные изменения, которые происходят до того, как проявится поведенческое обучение, могут быть задействованы в управлении поведенческими изменениями, в то время как те, которые происходят после поведенческого обучения, могут играть роль в укреплении вновь сформированной ассоциации.Чтобы ответить на этот вопрос, для всех изменяющихся клеток мы вычислили пробное число обучения и сравнили его с вычисленным пробным числом нейронных изменений (подробное описание используемых поведенческих и нейронных алгоритмов см. В Wirth et al., 2003). Мы обнаружили, что изменяющие гиппокамп клетки могут предшествовать (14 примеров) параллельному (4 примера) и запаздывать (19 примеров) поведенческому обучению (рис. 3 ). Эти данные свидетельствуют о том, что гиппокамп участвует во всех этапах процесса обучения из нескольких испытаний до того, как будет выражено поведенческое обучение, когда наблюдаемая деятельность может быть задействована в управлении поведенческими изменениями, лежащими в основе обучения, до нескольких испытаний после обучения, когда может быть задействована деятельность. в укреплении новообразованного объединения.
Резюме и дальнейшие направления
Мы показали, что клетки гиппокампа обеспечивают сильные связанные с обучением паттерны нейронной активности, которые участвуют в начальном формировании новых ассоциативных воспоминаний. Поскольку эти изменения могут происходить до, в то же время или после обучения, эти результаты предполагают, что может происходить постепенное привлечение сети нейронов гиппокампа во время формирования новых ассоциативных воспоминаний. Предыдущие исследования показали, что помимо гиппокампа клетки в нескольких других областях мозга, включая префронтальную кору (Asaad et al.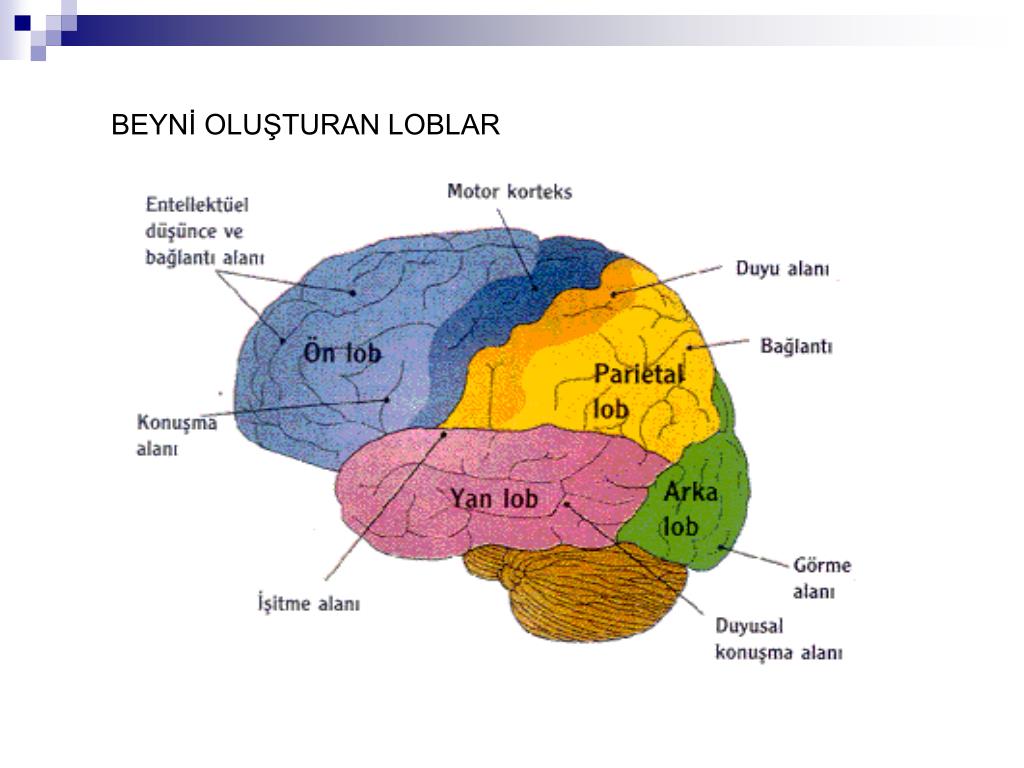 , 1998), лобные двигательные области (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и полосатое тело (Brasted and Wise, 2004) демонстрируют похожие модели обучения — сопутствующая деятельность во время аналогичных ассоциативных учебных задач. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с двигателем областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также для раннего усиления и консолидации нового ассоциативного обучения.
, 1998), лобные двигательные области (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и полосатое тело (Brasted and Wise, 2004) демонстрируют похожие модели обучения — сопутствующая деятельность во время аналогичных ассоциативных учебных задач. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с двигателем областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также для раннего усиления и консолидации нового ассоциативного обучения.
Асаад В. Ф., Райнер Г., Миллер Е. К. (1998) Нейронная активность в префронтальной коре приматов во время ассоциативного обучения. Нейрон 21: 1399-1407.
Brasted P. J., Bussey TJ, Murray EA, Wise SP (2002) Перерезка Fornix нарушает условное зрительно-моторное обучение при выполнении задач, связанных с несправедливо дифференцированными ответами. J Neurophysiol 87: 631-633.
Brasted P. J., Bussey TJ, Murray EA, Wise SP (2003) Роль системы гиппокампа в ассоциативном обучении за пределами пространственной области.Мозг 126: 1202-1223.
J., Bussey TJ, Murray EA, Wise SP (2003) Роль системы гиппокампа в ассоциативном обучении за пределами пространственной области.Мозг 126: 1202-1223.
Брастед П. Дж., Уайз С. П. (2004) Сравнение связанной с обучением активности нейронов в дорсальной премоторной коре и полосатом теле. Eur J Neurosci 19: 721-740.
Chen L. L., Wise S. P. (1995a) Активность нейронов в дополнительном поле глаза во время приобретения условных глазодвигательных ассоциаций. J Neurophys 73: 1101-1121.
Chen L. L., Wise S. P. (1995b) Дополнительное глазное поле контрастирует с лобным полем глаза во время получения условных глазодвигательных ассоциаций.Дж. Neurophys 73: 1122-1134.
Эйхенбаум Х., Коэн Н. Дж. (2001) От обусловленности к сознательному воспоминанию. Нью-Йорк: Издательство Оксфордского университета.
Миц А. Р., Годшалк М., Вайз С. П. (1991) Зависимая от обучения нейронная активность в премоторной коре: активность во время приобретения условных моторных ассоциаций.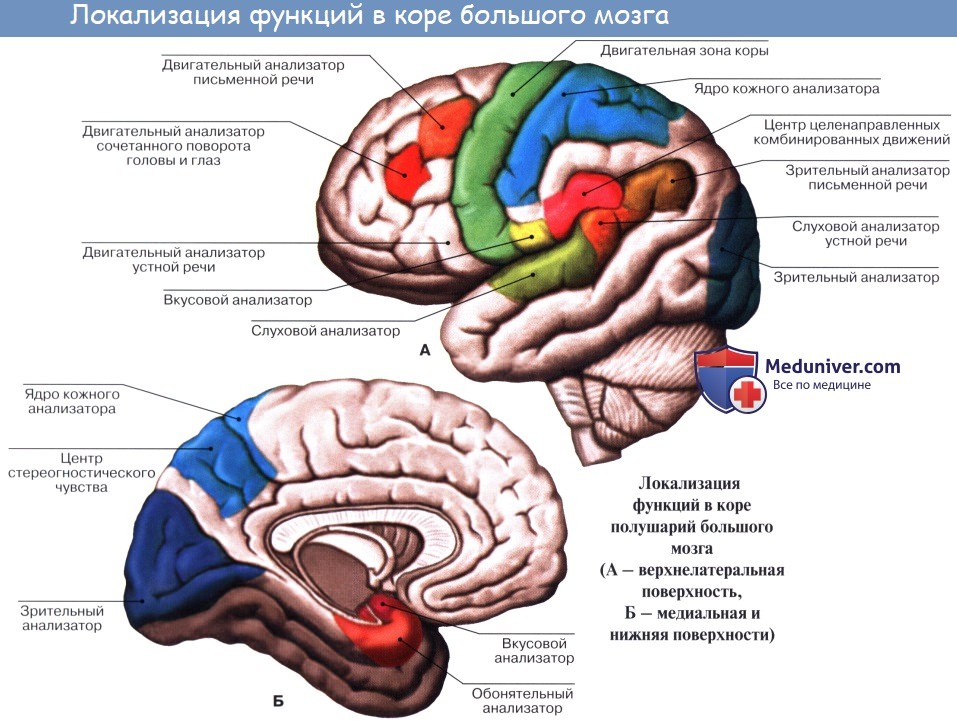 Дж. Neurosci 11: 1855-1872.
Дж. Neurosci 11: 1855-1872.
Мюррей Э. А., Бусси Т. Дж., Уайз С. П. (2000) Роль префронтальной коры в сети для произвольного зрительно-моторного картирования. Exp Br Res 133: 114-129.
Мюррей Э. А., Уайз С. П. (1996) Роль гиппокампа и прилежащей коры, но не миндалины, в зрительно-моторном условном обучении у макак-резусов. Behav Neurosci 110: 1261-1270.
Рупняк Н. М., Гаффан Д. (1987) Гиппокамп обезьяны и изучение пространственно направленных движений. J Neurosci 7: 2331-2337.
Зальцман К. Д., Бриттен К. Х., Ньюсом В. Т. (1990) Корковая микростимуляция влияет на перцепционные суждения о направлении движения. Nature 346: 174-177.
Зальцман К.Д., Мурасуги К. М., Бриттен К. Х., Ньюсом В. Т. (1992) Микростимуляция в визуальной области МТ: влияние на способность распознавания направления. J. Neurosci 12: 2331-2355.
Сковилл В. Б., Милнер Б. (1957) Потеря недавней памяти после двусторонних поражений гиппокампа. J. Neurol Neurosurg Psych 20: 11-21.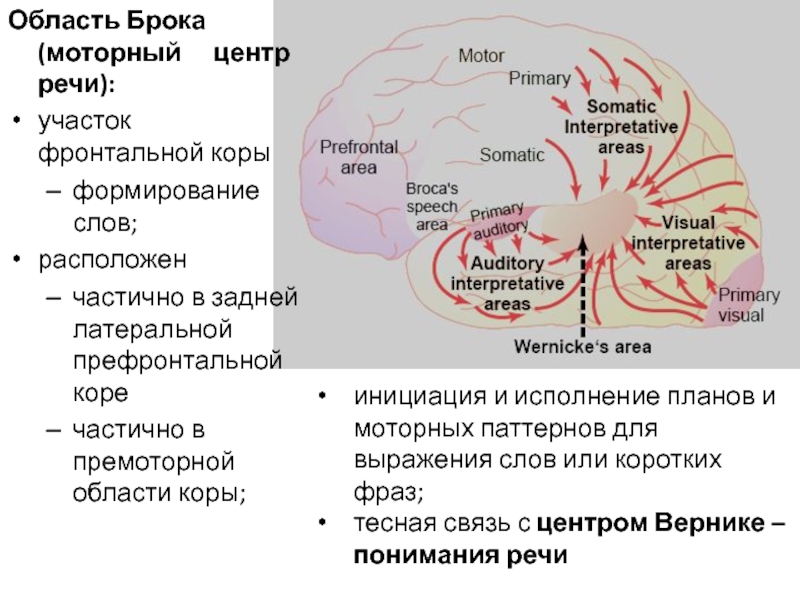
Сквайр Л. Р., Зола С. М. (1996) Структура и функции декларативных и недекларативных систем памяти. Proc Natl Acad Sci 93: 13515-13522.
Вирт С., Янике М., Франк Л.М., Смит А. С., Браун Е. Н., Судзуки В. А. (2003) Отдельные нейроны в гиппокампе обезьян и обучение новым ассоциациям. Science 300: 1578-1581.
Уайз С. П., Мюррей Э. А. (1999) Роль системы гиппокампа в условном моторном обучении: отображение предшествующих действий. Гиппокамп 9: 101-117.
Об авторе
Венди А. Сузуки получила докторскую степень в области нейробиологии в Калифорнийском университете в Сан-Диего в 1993 году и работала после получения докторской степени в лаборатории нейропсихологии Национального института психического здоровья.В настоящее время она является адъюнкт-профессором неврологии и психологии Нью-Йоркского университета. Ее исследования сосредоточены на понимании мозговой основы памяти. Чтобы ответить на этот вопрос, она использует поведенческую нейрофизиологию, регистрируя активность отдельных нейронов, когда обезьяны выполняют различные задачи, требующие памяти. Ее недавняя работа была сосредоточена на том, как клетки гиппокампа сигнализируют как о начальном формировании, так и о долгосрочном представлении ассоциативной памяти. Судзуки — лауреат премии Линдсли в области поведенческой нейробиологии (1994) и премии Троланда в области экспериментальной психологии (2004 год).
Ее недавняя работа была сосредоточена на том, как клетки гиппокампа сигнализируют как о начальном формировании, так и о долгосрочном представлении ассоциативной памяти. Судзуки — лауреат премии Линдсли в области поведенческой нейробиологии (1994) и премии Троланда в области экспериментальной психологии (2004 год).
Надпочечники | Johns Hopkins Medicine
Анатомия надпочечников
Надпочечник состоит из двух основных частей:
Кора надпочечника — это внешняя область, а также самая большая часть надпочечника. Он разделен на три отдельные зоны: клубочковая зона, фасцикулярная зона и ретикулярная зона. Каждая зона отвечает за выработку определенных гормонов.
мозговое вещество надпочечника находится внутри коры надпочечников в центре надпочечника.Он производит «гормоны стресса», в том числе адреналин.
Кора надпочечников и мозговой слой надпочечников заключены в жировую капсулу , которая образует защитный слой вокруг надпочечника.
Гормоны надпочечников
Роль надпочечников в организме заключается в высвобождении определенных гормонов непосредственно в кровоток. Многие из этих гормонов связаны с тем, как организм реагирует на стресс, а некоторые жизненно важны для существования.Обе части надпочечников — кора надпочечников и мозговой слой надпочечников — выполняют различные и отдельные функции.
Каждая зона коры надпочечников секретирует определенный гормон. Ключевые гормоны, вырабатываемые корой надпочечников, включают:
Кортизол
Кортизол — это глюкокортикоидный гормон, вырабатываемый фасциальной зоной, которая играет в организме несколько важных ролей. Он помогает контролировать использование организмом жиров, белков и углеводов; подавляет воспаление; регулирует артериальное давление; повышает уровень сахара в крови; а также может уменьшить образование костей.
Этот гормон также контролирует цикл сна / бодрствования. Он высвобождается во время стресса, чтобы помочь вашему телу получить заряд энергии и лучше справиться с чрезвычайной ситуацией.
Как надпочечники вырабатывают кортизол
Надпочечники вырабатывают гормоны в ответ на сигналы от гипофиза в головном мозге, который реагирует на сигналы от гипоталамуса, также расположенного в головном мозге. Это называется осью гипоталамуса и гипофиза надпочечников. Например, для выработки кортизола надпочечниками происходит следующее:
Гипоталамус производит кортикотропин-рилизинг-гормон (CRH), который стимулирует секрецию гипофиза гормона адренокортикотропина (ACTH).
- Затем
АКТГ стимулирует надпочечники производить и высвобождать гормоны кортизола в кровь.
Обычно и гипоталамус, и гипофиз могут определять, есть ли в крови необходимое количество циркулирующего кортизола. Если кортизола слишком много или слишком мало, эти железы соответственно изменяют количество выделяемых CRH и ACTH. Это называется петлей отрицательной обратной связи.
Избыточное производство кортизола может происходить из-за узелков в надпочечниках или из-за избыточного производства АКТГ из опухоли в гипофизе или из другого источника.

Альдостерон
Минералокортикоидный гормон, продуцируемый клубочковой оболочкой, играет центральную роль в регулировании артериального давления и некоторых электролитов (натрия и калия). Альдостерон посылает сигналы почкам, в результате чего почки поглощают больше натрия в кровоток и выделяют калий с мочой. Это означает, что альдостерон также помогает регулировать pH крови, контролируя уровень электролитов в крови.
ДГЭА и андрогенные стероиды
Эти гормоны, вырабатываемые сетчатой оболочкой, являются слабыми мужскими гормонами.Это гормоны-предшественники, которые в яичниках превращаются в женские гормоны (эстрогены), а в яичках — в мужские гормоны (андрогены). Однако эстрогены и андрогены вырабатываются яичниками и семенниками в гораздо большем количестве.
Адреналин (адреналин) и норэпинефрин (норадреналин)
мозговое вещество надпочечника , внутренняя часть надпочечника, контролирует гормоны, которые запускают реакцию «беги или сражайся». К основным гормонам, секретируемым мозговым веществом надпочечников, относятся адреналин (адреналин) и норадреналин (норадреналин), выполняющие схожие функции.
К основным гормонам, секретируемым мозговым веществом надпочечников, относятся адреналин (адреналин) и норадреналин (норадреналин), выполняющие схожие функции.
Среди прочего, эти гормоны способны увеличивать частоту сердечных сокращений и силу сердечных сокращений, увеличивать приток крови к мышцам и мозгу, расслаблять гладкие мышцы дыхательных путей и способствовать метаболизму глюкозы (сахара). Они также контролируют сжатие кровеносных сосудов (сужение сосудов), помогая поддерживать кровяное давление и повышая его в ответ на стресс.
Как и некоторые другие гормоны, вырабатываемые надпочечниками, адреналин и норадреналин часто активируются в ситуациях, вызывающих физический и эмоциональный стресс, когда вашему организму требуются дополнительные ресурсы и энергия, чтобы выдержать необычное напряжение.
Заболевания надпочечников
Два распространенных способа, которыми надпочечники вызывают проблемы со здоровьем, — это производство слишком малого или слишком большого количества определенных гормонов, что приводит к гормональному дисбалансу.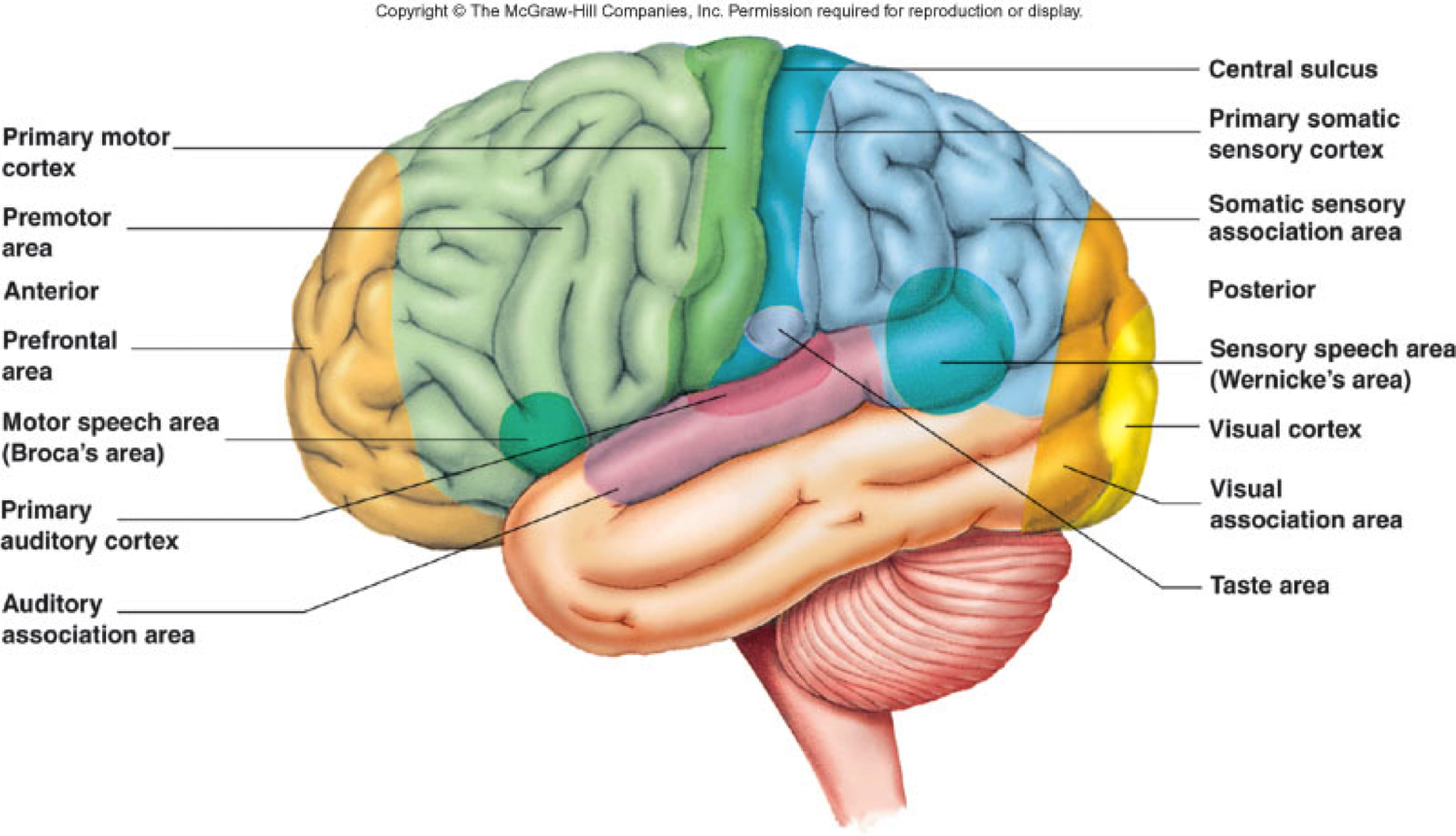 Эти нарушения функции надпочечников могут быть вызваны различными заболеваниями надпочечников или гипофиза.
Эти нарушения функции надпочечников могут быть вызваны различными заболеваниями надпочечников или гипофиза.
Надпочечниковая недостаточность
Надпочечниковая недостаточность — редкое заболевание. Это может быть вызвано заболеванием надпочечников (первичная надпочечниковая недостаточность, болезнь Аддисона) или заболеваниями гипоталамуса или гипофиза (вторичная надпочечниковая недостаточность).Он противоположен синдрому Кушинга и характеризуется низким уровнем гормонов надпочечников. Симптомы включают потерю веса, плохой аппетит, тошноту и рвоту, утомляемость, потемнение кожи (только при первичной надпочечниковой недостаточности), боль в животе и другие.
Причины первичной недостаточности надпочечников могут включать аутоиммунные заболевания, грибковые и другие инфекции, рак (редко) и генетические факторы.
Хотя надпочечниковая недостаточность обычно развивается со временем, она также может проявляться внезапно как острая надпочечниковая недостаточность (надпочечниковый криз). У него похожие симптомы, но последствия более серьезные, включая опасный для жизни шок, судороги и кому. Они могут развиться, если не лечить заболевание.
У него похожие симптомы, но последствия более серьезные, включая опасный для жизни шок, судороги и кому. Они могут развиться, если не лечить заболевание.
Врожденная гиперплазия надпочечников
Надпочечниковая недостаточность также может быть результатом генетического заболевания, называемого врожденной гиперплазией надпочечников. У детей, рожденных с этим заболеванием, отсутствует важный фермент, необходимый для выработки кортизола, альдостерона или того и другого. В то же время они часто испытывают избыток андрогенов, что может привести к появлению мужских качеств у девочек и преждевременному половому созреванию у мальчиков.
Врожденная гиперплазия надпочечников может оставаться невыявленной в течение многих лет в зависимости от степени недостаточности ферментов. В более тяжелых случаях младенцы могут страдать от неоднозначных гениталий, обезвоживания, рвоты и задержки развития.
Гиперактивные надпочечники
Иногда в надпочечниках могут образовываться узелки, вырабатывающие слишком много определенных гормонов. Узелки размером 4 сантиметра и более, а также узелки, которые показывают определенные особенности на изображении, увеличивают подозрение на злокачественное новообразование.Как доброкачественные, так и злокачественные узелки могут производить чрезмерное количество определенных гормонов, что называется функциональным узлом. Функциональные опухоли, злокачественные опухоли или узелки размером более 4 сантиметров рекомендуется направлять на хирургическое обследование.
Избыток кортизола: синдром Кушинга
Синдром Кушинга возникает в результате чрезмерного производства кортизола надпочечниками. Симптомы могут включать увеличение веса и жировые отложения в определенных областях тела, таких как лицо, ниже задней части шеи, называемой буйволиным горбом, и в области живота; истончение рук и ног; фиолетовые растяжки на животе; волосы на лице; усталость; мышечная слабость; кожа легко ушибается; высокое кровяное давление; сахарный диабет; и другие проблемы со здоровьем.
Избыточное производство кортизола также может быть вызвано перепроизводством АКТГ доброкачественной опухолью в гипофизе или опухолью в другом месте тела. Это известно как болезнь Кушинга. Другой частой причиной синдрома Кушинга является чрезмерное и продолжительное употребление внешних стероидов, таких как преднизон или дексаметазон, которые назначают для лечения многих аутоиммунных или воспалительных заболеваний (например, волчанки, ревматоидного артрита, астмы, воспалительного заболевания кишечника, рассеянного склероза и т. Д.)
Избыток альдостерона: гиперальдостеронизм
Гиперальдостеронизм возникает в результате перепроизводства альдостерона одним или обоими надпочечниками. Это характеризуется повышением артериального давления, для контроля которого часто требуется прием многих лекарств. У некоторых людей может развиться низкий уровень калия в крови, что может вызвать мышечные боли, слабость и спазмы. Когда причиной является гиперсекреция надпочечников, заболевание называется синдромом Конна.
Избыток адреналина или норадреналина: феохромоцитома
Феохромоцитома — это опухоль, которая приводит к избыточному производству адреналина или норадреналина мозговым веществом надпочечников, что часто случается всплесками.Иногда ткань нервного гребня, которая имеет ткань, аналогичную мозговому веществу надпочечников, может быть причиной гиперпродукции этих гормонов. Это известно как параганглиома.
Феохромоцитомы могут вызывать стойкое или спорадическое высокое кровяное давление, которое трудно контролировать с помощью обычных лекарств. Другие симптомы включают головные боли, потливость, тремор, беспокойство и учащенное сердцебиение. Некоторые люди генетически предрасположены к развитию этого типа опухоли.
Рак надпочечников
Злокачественные опухоли надпочечников (рак надпочечников), такие как карцинома коры надпочечников, встречаются редко и часто к моменту постановки диагноза распространились на другие органы и ткани.Эти опухоли имеют тенденцию к увеличению и могут достигать нескольких дюймов в диаметре.
Раковые опухоли надпочечников могут быть функциональными и выделять избыток одного или нескольких гормонов, что сопровождается соответствующими симптомами, как указано выше. Пациенты также могут испытывать боль в животе, боль в боку или чувство переполнения живота, особенно когда опухоль надпочечников становится очень большой.
Не все виды рака, обнаруживаемые в надпочечниках, происходят от самой железы. Большинство опухолей надпочечников являются метастазами или распространяются раком из другой первичной опухоли в другом месте тела.
Мозговые механизмы в моторном контроле во время выполнения движений: переход функциональной связности в соответствии с состояниями движения
Кандел, Э. Р., Шварц, Дж. Х., Джессел, Т. М., Сигельбаум, С. и Хадспет, А. Дж. Принципы неврологии . 5-е изд, (McGraw-Hill, 2013).
Георгопулос, А. П., Каласка, Дж. Ф., Каминити, Р. и Мэсси, Дж. Т. О взаимосвязи между направлением двумерных движений рук и разрядом клеток в моторной коре головного мозга приматов. Журнал нейробиологии: официальный журнал Общества нейробиологии 2 , 1527–1537 (1982).
Артикул CAS Google Scholar
Wessberg, J. et al. . Предсказание траектории руки в реальном времени ансамблями корковых нейронов приматов. Nature 408 , 361–365, https://doi.org/10.1038/35042582 (2000).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Тейлор Д. М., Тиллери С. И. и Шварц А. Б. Прямой кортикальный контроль трехмерных нейропротезных устройств. Наука 296 , 1829–1832, https://doi.org/10.1126/science.1070291 (2002).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Веллист, М., Перель, С., Сполдинг, М. К., Уитфорд, А. С. и Шварц, А. Б. Кортикальный контроль протеза руки для самостоятельного кормления. Природа 453 , 1098–1101, https: // doi.org / 10.1038 / nature06996 (2008).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Schalk, G. et al. . Расшифровка двумерных траекторий движения человека с помощью электрокортикографических сигналов. Журнал нейронной инженерии 4 , 264–275, https://doi.org/10.1088/1741-2560/4/3/012 (2007).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Truccolo, W., Friehs, G.M., Donoghue, J.P. & Hochberg, L.R. Настройка первичной моторной коры головного мозга на предполагаемую кинематику движения у людей с тетраплегией. Журнал нейробиологии: официальный журнал Общества нейробиологии 28 , 1163–1178, https://doi.org/10.1523/JNEUROSCI.4415-07.2008 (2008).
Артикул CAS Google Scholar
Брэдберри, Т. Дж., Джентили, Р. Дж. И Контрерас-Видаль, Дж.L. Восстановление трехмерных движений рук по неинвазивным электроэнцефалографическим сигналам. Журнал нейробиологии: официальный журнал Общества нейробиологии 30 , 3432–3437, https://doi.org/10.1523/JNEUROSCI.6107-09.2010 (2010).
Артикул CAS Google Scholar
Йом, Х. Г., Ким, Дж. С. и Чанг, К. К. Оценка скорости и траектории трехмерных движений на основе неинвазивных сигналов магнитоэнцефалографии. Журнал нейронной инженерии 10 , 026006, https://doi.org/10.1088/1741-2560/10/2/026006 (2013).
Артикул ОБЪЯВЛЕНИЯ PubMed Google Scholar
Хохберг, Л. Р. и др. . Дотягивайтесь до людей с тетраплегией с помощью нейронно-управляемой роботизированной руки. Природа 485 , 372–375, https://doi.org/10.1038/nature11076 (2012).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed PubMed Central Google Scholar
Коллингер, Дж. Л. и др. . Высокоэффективный нейропротезный контроль у человека с тетраплегией. Lancet 381 , 557–564, https://doi.org/10.1016/S0140-6736(12)61816-9 (2013).
Артикул PubMed PubMed Central Google Scholar
Бутон К. Э. и др. . Восстановление коркового контроля функционального движения у человека с квадриплегией. Природа 533 , 247–250, https: // doi.org / 10.1038 / nature17435 (2016).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Шварц, А. Б. Движение: как мозг общается с миром. Ячейка 164 , 1122–1135, https://doi.org/10.1016/j.cell.2016.02.038 (2016).
Артикул CAS PubMed PubMed Central Google Scholar
Colebatch, J.G. Bereitschafts: потенциальные и связанные с движением потенциалы: происхождение, значение и применение при нарушениях движения человека. Двигательные расстройства: официальный журнал Общества двигательных расстройств 22 , 601–610, https://doi.org/10.1002/mds.21323 (2007).
Артикул Google Scholar
Янкеловиц, С. К. и Колебатч, Дж. Г. Связанные с движениями потенциалы, связанные с самостоятельным темпом, заданными и воображаемыми движениями рук. Экспериментальное исследование мозга 147 , 98–107, https://doi.org/10.1007/s00221-002-1220-8 (2002).
Артикул CAS PubMed Google Scholar
Пфурчеллер Г. и Лопес да Силва Ф. Х. Синхронизация и десинхронизация ЭЭГ / МЭГ, связанные с событиями: основные принципы. Clin Neurophysiol 110 , 1842–1857 (1999).
Артикул CAS PubMed Google Scholar
Waldert, S. и др. . Обзор информации о направлении в нейронных сигналах для интерфейсов мозг-машина. Журнал физиологии, Париж 103 , 244–254, https://doi.org/10.1016/j.jphysparis.2009.08.007 (2009).
Артикул PubMed Google Scholar
Jurkiewicz, M. T., Gaetz, W. C., Bostan, A. C. и Cheyne, D. Постдвигательный бета-отскок генерируется в моторной коре головного мозга: данные нейромагнитных записей. NeuroImage 32 , 1281–1289, https://doi.org/10.1016/j.neuroimage.2006.06.005 (2006).
Артикул PubMed Google Scholar
Pfurtscheller, G. Графическое отображение и статистическая оценка десинхронизации, связанной с событиями (ERD). Электроэнцефалография и клиническая нейрофизиология 43 , 757–760 (1977).
Артикул CAS PubMed Google Scholar
Pfurtscheller, G. & Aranibar, A. Связанная с событием корковая десинхронизация, обнаруженная с помощью измерений мощности ЭЭГ кожи головы. Электроэнцефалография и клиническая нейрофизиология 42 , 817–826 (1977).
Артикул CAS PubMed Google Scholar
Pfurtscheller, G. Синхронизация, связанная с событием (ERS): электрофизиологический коррелят корковых областей в состоянии покоя. Электроэнцефалография и клиническая нейрофизиология 83 , 62–69 (1992).
Артикул CAS PubMed Google Scholar
Бланкерц, Б. и др. . Берлинский мозг-компьютер. Интерфейс: точная производительность с первого сеанса у субъектов, не знающих BCI. Транзакции IEEE по биомедицинской инженерии 55 , 2452–2462, https://doi.org/10.1109/TBME.2008.923152 (2008).
Артикул PubMed Google Scholar
Юань, Х. и Хе, Б. Интерфейсы мозг-компьютер с использованием сенсомоторных ритмов: текущее состояние и перспективы на будущее. Транзакции IEEE по биомедицинской инженерии 61 , 1425–1435, https://doi.org/10.1109/TBME.2014.2312397 (2014).
Артикул PubMed PubMed Central Google Scholar
Павела К. и Бисвал Б. Связь между мозгом: выходит новый журнал. Мозговая связь 1 , 1–2, https: // doi.org / 10.1089 / brain.2011.0020 (2011).
Артикул PubMed PubMed Central Google Scholar
Герлофф К. и др. . Функциональная связь и региональная активация моторных областей коры головного мозга человека во время простых движений пальцев с внутренним и внешним стимулированием. Мозг: журнал неврологии 121 (Pt 8), 1513–1531, https://doi.org/10.1093/brain/121.8.1513 (1998).
Артикул Google Scholar
Ван, Б.А. и др. . Частотно-зависимая модуляция связности в ипсилатеральной сенсомоторной коре с помощью различных форм инициации движения. NeuroImage 159 , 248–260, https://doi.org/10.1016/j.neuroimage.2017.07.054 (2017).
Артикул PubMed Google Scholar
Chung, J. W., Ofori, E., Misra, G., Hess, C. W. & Vaillancourt, D. E. Активность бета-диапазона и связность в сенсомоторной и теменной коре важны для точной двигательной активности. NeuroImage 144 , 164–173, https://doi.org/10.1016/j.neuroimage.2016.10.008 (2017).
Артикул PubMed Google Scholar
Ву, Дж., Кнапп, Ф., Крамер, С. К. и Сринивасан, Р. Электроэнцефалографические показатели связности предсказывают обучение задаче определения последовательности движений. Журнал нейрофизиологии 119 , 490–498, https://doi.org/10.1152/jn.00580.2017 (2018).
Артикул PubMed Google Scholar
Dal Maso, F., Desormeau, B., Boudrias, M. H. & Roig, M. Острые сердечно-сосудистые упражнения способствуют функциональным изменениям в кортико-моторных сетях на ранних этапах консолидации моторной памяти. NeuroImage 174 , 380–392, https://doi.org/10.1016/j.neuroimage.2018.03.029 (2018).
Артикул PubMed Google Scholar
Herz, D. M. et al. . Специфическая модуляция эффективной связи во время двух простых однообразных двигательных задач: 122-канальное исследование ЭЭГ. NeuroImage 59 , 3187–3193, https://doi.org/10.1016/j.neuroimage.2011.11.042 (2012).
Артикул PubMed Google Scholar
ван Вейк, Б. К. М., Бик, П. Дж. И Даффертсхофер, А. Нейронная синхронизация в двигательной системе: что мы узнали на данный момент? Границы нейробиологии человека 6 , https://doi.org/10.3389/fnhum.2012.00252 (2012).
Heinrichs-Graham, E.И Уилсон, Т. В. Сложность кодирования в двигательной цепи человека. Hum Brain Mapp 36 , 5155–5167, https://doi.org/10.1002/hbm.23000 (2015).
Артикул PubMed PubMed Central Google Scholar
Ханакава, Т., Димьян, М. А. и Халлетт, М. Планирование моторики, образы и исполнение в распределенной моторной сети: исследование динамики движения с функциональной МРТ. Cereb Cortex 18 , 2775–2788, https: // doi.org / 10.1093 / cercor / bhn036 (2008).
Артикул PubMed PubMed Central Google Scholar
van den Heuvel, M. P. & Sporns, O. Сетевые узлы в человеческом мозге. Тенденции в когнитивных науках 17 , 683–696, https://doi.org/10.1016/j.tics.2013.09.012 (2013).
Артикул PubMed Google Scholar
Аллен, Э.А. и др. . Отслеживание динамики подключения всего мозга в состоянии покоя. Cereb Cortex 24 , 663–676, https://doi.org/10.1093/cercor/bhs352 (2014).
Артикул PubMed Google Scholar
Бернс, С. П. и др. . Сетевая динамика мозга и влияние зоны начала эпилептического припадка. Труды Национальной академии наук Соединенных Штатов Америки 111 , E5321–5330, https: // doi.org / 10.1073 / pnas.1401752111 (2014).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed PubMed Central Google Scholar
Калхун, В. Д., Миллер, Р., Перлсон, Г. и Адали, Т. Хроннектом: изменяющиеся во времени сети связи как следующий рубеж в открытии данных фМРТ. Нейрон 84 , 262–274 (2014).
Артикул CAS PubMed PubMed Central Google Scholar
Скотт, С. Х. Оптимальное управление с обратной связью и нейронная основа произвольного моторного управления. Природные обзоры. Неврология 5 , 532–546, https://doi.org/10.1038/nrn1427 (2004).
Артикул CAS PubMed Google Scholar
Скотт, С. Х. Вычислительная и нейронная основа произвольного моторного контроля и планирования. Тенденции в когнитивных науках 16 , 541–549, https: // doi.org / 10.1016 / j.tics.2012.09.008 (2012).
Артикул PubMed Google Scholar
Медведь М. Ф., Коннорс Б. В. и Парадизо М. А. Неврология: исследование мозга . Четвертый выпуск. edn, (Wolters Kluwer, 2016).
Олдфилд, Р. К. Оценка и анализ руки: Эдинбургская инвентаризация. Neuropsychologia 9 , 97–113 (1971).
Артикул CAS Google Scholar
Таулу, С. и Симола, Дж. Метод пространственно-временного разделения пространства сигналов для подавления соседних помех при измерениях МЭГ. Физика в медицине и биологии 51 , 1759–1768, https://doi.org/10.1088/0031-9155/51/7/008 (2006).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Таулу, С. и Хари, Р. Удаление магнитоэнцефалографических артефактов с временным разделением сигналов и пространств: демонстрация с помощью однократных попыток слуховых вызванных ответов. Hum Brain Mapp 30 , 1524–1534, https://doi.org/10.1002/hbm.20627 (2009).
Артикул PubMed Google Scholar
http://neuro.imm.dtu.dk/wiki/Automated_Anatomical_Labeling.
Каффин Б. Н. и Коэн Д. Сравнение магнитоэнцефалограммы и электроэнцефалограммы. Электроэнцефалография и клиническая нейрофизиология 47 , 132–146 (1979).
Артикул CAS PubMed Google Scholar
Ким, Дж. С. и др. . Спектральные аспекты мощности сети режима по умолчанию при шизофрении: исследование MEG. Bmc Neurosci 15 , 104, https://doi.org/10.1186/1471-2202-15-104 (2014).
Артикул PubMed PubMed Central Google Scholar
Паскуаль-Марки, Р.D. Стандартизированная электромагнитная томография головного мозга с низким разрешением (sLORETA): технические детали. Методы и результаты экспериментальной и клинической фармакологии 24 (Дополнение D), 5–12 (2002).
PubMed Google Scholar
Джин, С. Х., Чон, В., Сеол, Дж., Квон, Дж. И Чунг, К. К. Функциональные корковые центры в закрытых глазах в состоянии покоя человеческого мозга с электрофизиологической точки зрения с использованием магнитоэнцефалографии. PloS one 8 , https://doi.org/10.1371/journal.pone.0068192 (2013).
Рубинов, М. и Спорнс, О. Комплексные сетевые меры связности мозга: использование и интерпретация. NeuroImage 52 , 1059–1069, https://doi.org/10.1016/j.neuroimage.2009.10.003 (2010).
Артикул PubMed Google Scholar
Ся, М., Ван, Дж. И Хе, Ю. BrainNet Viewer: инструмент сетевой визуализации для коннектомики человеческого мозга. PloS one 8 , e68910, https://doi.org/10.1371/journal.pone.0068910 (2013).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed PubMed Central Google Scholar
Waldert, S. et al. . Направление движения руки расшифровывается по данным МЭГ и ЭЭГ. Journal of Neuroscience 28 , 1000–1008, https://doi.org/10.1523/Jneurosci.5171-07.2008 (2008).
Артикул CAS PubMed Google Scholar
Тода, А., Имамизу, Х., Кавато, М. и Сато, М. А. Реконструкция двумерных траекторий движения на основе выбранных магнитоэнцефалографических кортикальных токов комбинированными разреженными байесовскими методами. NeuroImage 54 , 892–905, https://doi.org/10.1016/j.neuroimage.2010.09.057 (2011).
Артикул PubMed Google Scholar
Гордон, Дж., Гиларди, М. Ф. и Гез, К. Точность движений на плоскости.I. Независимость направления и изменчивости степени. Экспериментальное исследование мозга 99 , 97–111 (1994).
Артикул CAS PubMed Google Scholar
Эбнер, Т. Дж. И Палар, С. Церебеллум предсказывают будущее моторное состояние. Мозжечок 7 , 583–588, https://doi.org/10.1007/s12311-008-0059-3 (2008).
Артикул PubMed PubMed Central Google Scholar
Ито, М. Управление умственной деятельностью с помощью внутренних моделей в мозжечке. Природные обзоры. Неврология 9 , 304–313, https://doi.org/10.1038/nrn2332 (2008).
Артикул CAS PubMed Google Scholar
Manto, M. et al. . Консенсусный документ: роль мозжечка в моторном контроле — разнообразие представлений о вовлечении мозжечка в движение. Мозжечок 11 , 457–487, https: // doi.org / 10.1007 / s12311-011-0331-9 (2012).
Артикул PubMed PubMed Central Google Scholar
Хохберг, Л. Р. и др. . Контроль нейронного ансамбля протезных устройств у человека с тетраплегией. Природа 442 , 164–171, https://doi.org/10.1038/nature04970 (2006).
Артикул ОБЪЯВЛЕНИЯ CAS PubMed Google Scholar
Желязник, Х. З., Хокинс, Б. и Киссельбург, Л. Быстрая обработка визуальной обратной связи при движении с одним прицелом. Журнал моторного поведения 15 , 217–236 (1983).
Артикул CAS PubMed Google Scholar
Пистоль, Т., Болл, Т., Шульце-Бонхаге, А., Аэрцен, А. и Меринг, К. Прогнозирование траекторий движения рук на основе записей ЭКоГ у людей. Журнал методов нейробиологии 167 , 105–114, https: // doi.org / 10.1016 / j.jneumeth.2007.10.001 (2008).
Артикул PubMed Google Scholar
Attal, Y., Maess, B., Friederici, A. & David, O. Модели головы и динамическое причинно-следственное моделирование подкорковой активности с использованием данных магнитоэнцефалографии / электроэнцефалографии. Обзоры в неврологии 23 , 85–95, https://doi.org/10.1515/rns.2011.056 (2012).
Артикул PubMed Google Scholar
Речь и язык | Центр памяти и старения
Речевые и языковые трудности обычно возникают у людей с деменцией и другими неврологическими заболеваниями.Пациенты могут испытывать дефицит вербального выражения (например, трудности с поиском слов) или понимания (например, трудности с пониманием речи).
Язык
Язык состоит из общественно общих правил, которые включают следующее:
- Семантика или значение (например, «корма» может означать «строгость поведения» или «хребет лодки»)
- Как составить новые слова (например, друг, дружелюбный, недружелюбный)
- Грамматика (e.г., «Я пошел в новый ресторан», а не «пошел в новый ресторан»)
- Социальный контекст (например, «Не могли бы вы открыть окно?» Вместо «Эй, откройте окно сейчас же!»)
Выступление
Речь — это словесное средство общения. Речь состоит из следующего:
- Артикуляция: Как воспроизводятся звуки речи
- Голос: Использование голосовых связок и дыхания для создания звука (например, охриплость, одышка, проецирование)
- Беглость и просодия: Ритм, интонация, ударение и связанные с ними атрибуты речи
Когда кому-то трудно понимать других людей (восприимчивый язык) или объяснять мысли, идеи и чувства (выразительный язык), это языковое расстройство.
Когда кто-то не может правильно или плавно воспроизводить речевые звуки или имеет проблемы с голосом, это нарушение речи.
Анатомия языка
Есть несколько областей мозга, которые играют решающую роль в речи и языке.
- Зона Брока, расположенная в левом полушарии, связана с производством речи и артикуляцией. Наша способность формулировать идеи, а также точно использовать слова в устной и письменной речи была приписана этой важной области.
- Область Вернике — критическая языковая область в задней верхней височной доле, соединяется с областью Брока нервным путем. Область Вернике в первую очередь связана с пониманием. Исторически эта область была связана с обработкой речи, будь то письменная или устная.
- Угловая извилина позволяет нам связывать несколько типов языковой информации, будь то слуховая, визуальная или сенсорная. Он расположен в непосредственной близости от других критических областей мозга, таких как теменная доля, обрабатывающая тактильные ощущения, затылочная доля, участвующая в визуальном анализе, и височная доля, обрабатывающая звуки.Угловая извилина позволяет связать воспринимаемое слово с разными образами, ощущениями и представлениями.
Нарушения речи и языка
Афазия — это термин, используемый для описания приобретенной потери речи, которая вызывает проблемы с любым или всеми из следующих: говорение, аудирование, чтение и письмо. У некоторых людей с афазией возникают проблемы с использованием слов и предложений (экспрессивная афазия). У некоторых есть проблемы с пониманием других (рецептивная афазия). Другим, страдающим афазией, не удается использовать слова и понимание (глобальная афазия).Афазия может вызвать проблемы с разговорной речью (разговор и понимание) и письменной (чтение и письмо). Как правило, чтение и письмо ухудшаются больше, чем речь или понимание. Выраженность афазии зависит от размера и локализации поражения головного мозга.
Broca’s (выразительный или моторный) Aphasia
Было показано, что повреждение дискретной части мозга в левой лобной доле (область Брока) доминирующего речевого полушария существенно влияет на использование спонтанной речи и управление моторной речью.Слова могут произноситься очень медленно и плохо артикулироваться. Речь может быть трудной и состоять в основном из существительных, глаголов или важных прилагательных. Речь принимает телеграфный характер. Люди, страдающие афазией Брока, испытывают большие трудности с повторением и серьезными затруднениями при письме. Однако у некоторых пациентов понимание устной и письменной речи может быть относительно хорошо сохраненным. Нелегкий вариант первичной прогрессирующей афазии (nfvPPA) представляет собой тип экспрессивной афазии.
Глобальная афазия
Если повреждение охватывает как области Вернике, так и области Брока, может возникнуть глобальная афазия.В этом случае затрагиваются все аспекты речи и языка. Пациенты могут сказать максимум несколько слов и понять только несколько слов и фраз. Обычно они не могут выполнять команды или присваивать имена объектам. Они не могут читать, писать или повторять сказанные им слова.
Логопеническая первичная прогрессирующая афазия (lvPPA)
Дегенерация угловой извилины височной доли и нижней теменной доли может привести к lvPPA. Типичные симптомы включают замедленную речь с нормальной артикуляцией, нарушение понимания синтаксиса предложений, а также нарушение именования предметов.lvPPA, вероятно, связан с патологией болезни Альцгеймера.
Первичная прогрессирующая афазия (PPA)
PPA вызывается дегенерацией частей мозга, контролирующих речь и язык (левая, или «доминирующая», часть мозга в лобных, височных и теменных областях, которые обычно контролируют речевую функцию). Этот тип афазии начинается постепенно, с речевых или языковых симптомов, которые отражают нормальную роль места первоначальной дегенерации. В конце концов, проблемы распространились по всей языковой сети.Подтипы PPA включают неглубокую первичную прогрессирующую афазию (nfvPPA), семантический вариант первичной прогрессирующей афазии (svPPA) и логопеническую первичную прогрессирующую афазию (lvPPA). Эти синдромы возникают в результате множества основных заболеваний, но чаще всего лобно-височной долевой дегенерации (ЛВП) (как подтипы тау-белка, так и подтипа TDP-43) или болезни Альцгеймера.
Афазия Вернике
Было показано, что повреждение задних верхних областей доминирующей височной доли языка (часто называемой областью Вернике) значительно влияет на понимание речи.Другими словами, информация слышится через неповрежденную слуховую кору в передней височной доле, однако, когда она достигает задних ассоциативных областей, информация не может быть в достаточной степени «переведена». В отличие от афазии Брока, человек с афазией Вернике разговаривает и свободно жестикулирует. Речь воспроизводится без усилий, а предложения имеют нормальную длину. Однако речь человека лишена смысла.
Этот образец рецептивной афазии обозначен:
- Беглая, грамматически правильная речь, мало значащая
- Плохое понимание
- Парафазные ошибки:
- называть ложку вилкой (смысловой)
- называя ложку «спудом» (букв.)
- Неологизмы (или бессмысленные слова)
Последствия неврологического заболевания
Болезнь Альцгеймера (AD)
При болезни Альцгеймера, наиболее частой причине слабоумия, речевое функционирование может быть относительно щадящим на ранних стадиях болезни, но, вероятно, существенно ухудшится на средних и поздних стадиях.Люди с БА часто испытывают трудности с языковым выражением, беглостью слов и именованием предметов. Синтаксис и понимание языка обычно сохраняются на ранних стадиях, однако на более поздних стадиях речь может прерываться из-за трудностей с поиском слов. Другими словами, пациентам очень трудно говорить полными предложениями из-за усилий, которые требуются, чтобы подобрать правильные слова. Письменные навыки часто могут быть скомпрометированы. Понимание речи может значительно ухудшиться в конечной стадии заболевания.
Кортикобазальный синдром (CBS)
CBS чаще всего вызывает трудности с языковым выражением, такие как трудности с поиском слов или проблемы с артикуляцией речи. Также могут быть нарушены чтение и письмо.
Лобно-височная деменция (FTD)
Поведенческий вариант ЛТД в первую очередь не влияет на язык. Часто люди с bvFTD молчат и меньше разговаривают, но это изменение происходит в большей степени из-за повышенной апатии и отсутствия инициации. Пациенты могут реагировать, когда к ним обращаются, но обычно не склонны говорить.С другой стороны, некоторые пациенты по мере прогрессирования болезни испытывают эйфорию и растормаживание, и эти люди могут быть склонны больше говорить.
Начальные симптомы семантического варианта PPA часто связаны с проблемами с поиском нужных слов во время разговора. Двустороннее ухудшение височных долей (особенно передних) приводит к:
- свободная речь,
- грамматически правильный язык,
- потеря смысла слова и предмета,
- относительно сохраненное понимание и
- дефицит в понимании эмоций других.
У людей с негибким вариантом PPA возникают трудности с речью. Они знают, что хотят сказать, но им очень трудно выговорить слова изо рта. Дегенерация коры головного мозга в доминирующих на языке областях лобной, теменной и височной долей (включая область Брока) приводит к:
- медленная, затрудненная нелегкая речь,
- аграмматизм (использование только ценных слов содержания без соединительных или описательных слов) и
- относительно сохраненное понимание слов.
Инсульт и травма
Лица, перенесшие неврологические травмы, такие как инсульт или черепно-мозговая травма, также могут испытывать речевой и языковой дефицит, особенно, но не исключительно, если поражена левая часть мозга. Афазия часто встречается у людей с левосторонними травмами головного мозга. Навыки разговорной речи, аудирования, чтения и письма могут быть затронуты в разной степени. Если удар влияет на части мозга, которые контролируют мышцы, задействованные в речи (мышцы языка, рта и губ), речь может стать невнятной или замедленной.
.