
Основные параметры возбудимости: Возбудимость, параметры возбудимости. Биоэлектрические явления
Возбудимость, параметры возбудимости. Биоэлектрические явления
1. Тема: «Возбудимость, параметры возбудимости. Биоэлектрические явления».
План лекции:1. Физиологические свойства возбудимых тканей.
2. Возбудимость, её фазы.
3. Параметры возбудимости (порог возбуждения–реобаза,
полезное время, аккомодация, лабильность).
4. Электрические явления в возбудимых тканях.
2. Возбудимость – это свойство живых тканей отвечать на раздражение процессом возбуждения. Возбуждение – это сложный процесс,
характеризующийсяфизиологическими, биофизическими,
биохимическими и структурными
изменениями в тканях.
3. Параметры возбудимости
1. Порог возбуждения – это минимальная сила раздражителя, котораявызывает возбуждение, т.е. минимальную ответную реакцию.
2. Реобаза – это минимальная сила тока выраженная в Вольтах.
3. Полезное время – это наименьшее время, в течение которого должен
действовать раздражающий стимул в одну реобазу.

4. Хронаксия – это время, в течение которого сила тока, равная удвоенной
реобазе, вызывает возбуждение.
5. Аккомодация – это приспособление возбудимой ткани к медленно
6. Лабильность – это функциональная подвижность. Мерой лабильности
является максимальное количество импульсов, которое может
воспроизвести в единицу времени (в сек) возбудимая ткань в соответствии
с ритмом наносимых раздражений.
— нервная ткань: – 500–1000 имп/сек;
– аболютный рефрактерный период – 1–2 мсек.
— мышца: – 250–330 имп/сек;
– аболютный рефрактерный период – 4–5 мсек.
— синапс: – 100–125 имп/сек;
– аболютный рефрактерный период – 8–10 мсек.
4. Кривая «силы – времени» (Гоорвег, 1892 г.; Вейс, 1901г.; Лапик, 1909 г.)
1 – реобаза;2 – удвоенная реобаза.
а – полезное время
действия тока;
б – хронаксия.
5. Фазы изменения возбудимости при возбуждении
1. Абсолютный рефрактерный период.
В этот период ни на какие раздражения извне
ткань не отвечает.
Продолжительность этого периода, для:
— нервного волокна – 1-2 мсек;
— мышечного – 4-5 мсек;
— мионеврального синапса – 8-10 мсек.
2. Относительный рефрактерный период.
В этот период ответные реакции возникают
только при воздействии раздражителей выше
пороговой силы.
3. Супернормальный период.
В этот период ткань отвечает и на ниже
пороговые раздражения.
4.Субнормальный период.
В этот период происходит снижение
возбудимости и ткань отвечает на выше
пороговые раздражения.
6. Мембранный потенциал
Мембранный потенциал – эторазность потенциалов между
поверхностью клеточной
мембраны и её протоплазмой.
Снаружи мембрана заряжена – «+»;
Внутри мембрана заряжена – «-».
Величина мембранного потенциала
для мышечных волокон
– 60 — – 90 мв.
7. Потенциал действия
Потенциал действия– это колебание
мембранного
потенциала в
период
возбуждения
(происходит
перезарядка
мембраны).

8. Фазы потенциала действия
1. Локальный ответ.2. Деполяризация.
3. Реполяризация.
4. Отрицательный
5. Положительный
следовой потенциал.
9. Фазовые изменения возбудимости ткани во время её возбуждения
І – потенциал действия.ІІ – возбудимость ткани.
Физиология — 1 Тема Возбуждение Законы раздражения возбудимых
Физиология — 1 Тема: «Возбуждение. Законы раздражения возбудимых тканей» Лектор: доцент Евневич А. М.
Цель } по окончании лекции будете иметь представление о возбудимых тканях и их физиологических свойствах, параметрах возбудимости и законах раздражения возбудимых тканей, а также о применении этих знаний в клинической практике
План 1. 2. 3. 4. 5. Раздражимость как общебиологическое свойство живых тканей. Возбудимость, возбудимые ткани. Основные физиологические свойства возбудимой ткани. Параметры возбудимости.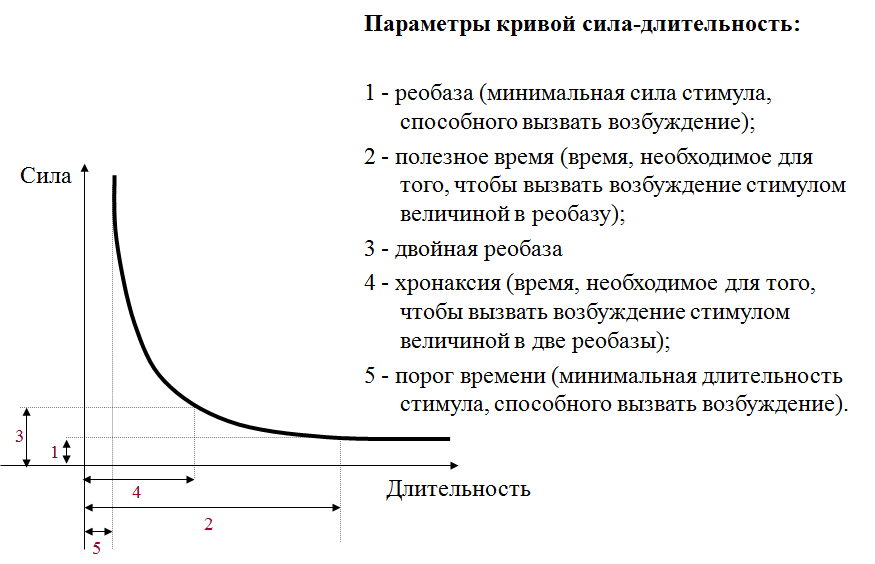
Основные понятия физиологии возбудимых тканей üраздражители и раздражение üраздражимость üвозбудимость и возбуждение üторможение
Возбудимые ткани
Возбудимость n способность живой ткани отвечать на действие достаточно сильного, быстрого и длительно действующего раздражителя изменением физиологических свойств и возникновением процесса возбуждения
Возбудимость n свойство возбудимых тканей отвечать на раздражение возбуждением, или генерацией потенциала действия (ПД)
Возбуждение волнообразный физиологический процесс, представляющий сложную биологическую реакцию на раздражение – «элементарные реакции» (Введенский) n неспецифические и специфические n
Параметры возбудимости n n n n порог раздражения реобаза полезное время хронаксия аккомодация лабильность рефрактерность
Классификация раздражителей n n n Физические – механические, температурные, электрические, световые, звуковые Физико-химические – изменения осмотического давления, р. Н, электролитного состава Химические – продукты обмена веществ, яды, лекарства, гормоны и т. д.
Н, электролитного состава Химические – продукты обмена веществ, яды, лекарства, гормоны и т. д.
Классификация раздражителей по физиологическому значению n Адекватные – раздражители, которые действуют на данную биологическую структуру в естественных условиях. n Неадекватные – раздражители, для восприятия которых данная клетка или орган не приспособлены.
Параметры возбудимости n n n n порог раздражения реобаза полезное время хронаксия аккомодация лабильность рефрактерность
Законы раздражения возбудимых тканей n n закон силы раздражения закон «все или ничего» закон «силы-времени» закон градиента силы
Порог раздражения n наименьшая сила раздражителя, которая способна вызвать минимальное возбуждение ткани
Закон силы раздражения n чем больше сила раздражителя, тем до определенного предела больше возбуждение, т. е. ответная реакция ткани
Закон силы
Мера возбудимости n n Порог раздражения (возбудимости) – та минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции. Подпороговое раздражение – раздражение меньшей интенсивности, не вызывающее ответные реакции.
Подпороговое раздражение – раздражение меньшей интенсивности, не вызывающее ответные реакции.
Порог раздражения
Закон «все или ничего» n подпороговые раздражения волокон не вызывают возбуждения ( «ничего» ), при пороговых стимулах возбуждение приобретает максимальную величину ( «все» ) (Боудич).
Формы электрических стимулов: прямоугольные n синусоидальные n линейно-нарастающие n экспоненциально-нарастающие n индукционные удары n конденсаторные разряды n
Применение электрического тока в клинике n наилучшее раздражающее действие вызывает постоянный ток прямоугольной формы
Применение электрического тока в клинике Электронные стимуляторы – импульсы любой формы, частоты и силы. n Диатермия – использование высокочастотного переменного тока от 0, 5 до 1 мгц для контролируемого местного прогревания ткани. n
n Диатермия – использование высокочастотного переменного тока от 0, 5 до 1 мгц для контролируемого местного прогревания ткани. n
Применение постоянного тока в медицине n Гальванизация – в лечебных целях используется ток низкого напряжения (30 -80 В) и небольшой силы (до 50 м. А) – способствует улучшению местного обмена веществ, регенерации эпителия.
Применение постоянного тока в медицине Лекарственный электрофорез – эффективное введение лекарственных веществ. n Электрообезболивание – в основе лежит явление электротона. n Используется для питания электродиагностических аппаратов. n
Параметры возбудимости Реобаза – минимальная сила постоянного тока, способная вызвать возбуждение (Лапик). n «Полезное время» – наименьшее время, в течение которого должен действовать ток, равный одной реобазе, чтобы вызвать возбуждение. n
Кривая «силы – времени» (Гоорвег-Вейс-Лапик) 1 – реобаза 2 – удвоенная реобаза а – полезное время действия тока б – хронаксия
Хронаксия n n n наименьшее время, в течение которого электрический ток, равный удвоенной реобазе, должен действовать на ткань, чтобы вызвать возбуждение характеризует скорость возникновения возбуждения при действии раздражителя чем больше скорость распространения возбуждения, тем меньше хронаксия
Хронаксия Ø Ø Субординационная (соподчиненная) – характерна для целого организма при сохраненной иннервации, при прохождении нервного импульса к мышце.
Кривая «силы – времени» (Гоорвег-Вейс- Лапик) n n n ОА – реобаза (пороговое напряжение электрического тока, в) ОС — полезное время, mc OF — хронаксия, mс.
Закон «силы — времени» n чем сильнее ток, тем меньше продолжительность его действия, необходимая для получения возбуждения
ЗАКОН «СИЛЫ — ВРЕМЕНИ»
Хронаксиметрия n n n метод определения возбудимости тканей или органов на основе выявления зависимости между пороговой силой раздражения, вызывающей процесс возбуждения и длительностью его действия. Фактически определяется субординационная хронаксия. При повреждении нерва или гибели мотонейронов спинного мозга выявляется конституционная хронаксия.
Применение хронаксиметрии n n n диагностика поражений центральной и периферической нервной систем, опорнодвигательного аппарата определение границ патологического очага и функционального состояния отдельных структур головного мозга в процессе проведения хирургических операций спортивная медицина, физиология труда, промышленная и санитарная гигиена
Параметры возбудимости n n Аккомодация – приспособление, адаптация к медленно нарастающему действию раздражителя, при которой возбудимость ткани снижается. Закон градиента силы – чем меньше крутизна нарастания раздражителя, тем выше порог возбудимости, а возбудимость ниже (Дюбуа — Реймон).
Закон градиента силы – чем меньше крутизна нарастания раздражителя, тем выше порог возбудимости, а возбудимость ниже (Дюбуа — Реймон).
Пороговый градиент аккомодации n n Мера скорости аккомодации – наименьшая крутизна нарастания силы раздражения, при которой ткань еще способна ответить возбуждением на данный раздражитель. Механизм аккомодации – инактивация натриевой и повышение калиевой проницаемости во время медленно нарастающей деполяризации мембраны.
Проводимость n способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани
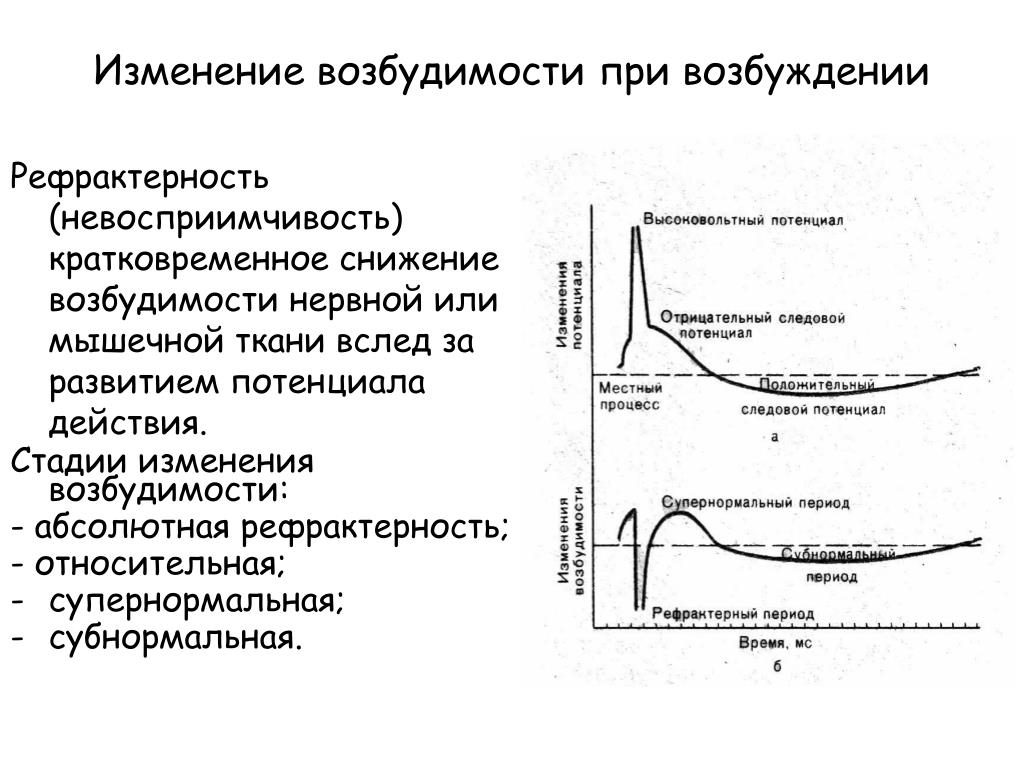
Рефрактерность n n n невозбудимость ткани при возбуждении время, за которое ткань способна вернуться в свое первоначальное состояние физиологического покоя временное снижение возбудимости одновременно с возникшим в ткани возбуждением
Рефрактерность n n Абсолютная – нет ответа ни на какой раздражитель. Относительная – ткань отвечает на сверхпороговый раздражитель.
Относительная – ткань отвечает на сверхпороговый раздражитель.
Изменение возбудимости при возбуждении
Лабильность (функциональная подвижность) (Н. Е. Введенский) n n способность возбудимой ткани реагировать на раздражение с определенной скоростью характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации
Лабильность (функциональная подвижность) (Н. Е. Введенский) максимальное число импульсов, которое может проводить ткань за 1 секунду n нервная ткань – 500 -1000 имп/с. n мышечная ткань – 150 -300 имп/с. n мионевральный синапс – 75 -150 имп/с. n
Лабильность отражает работоспособность возбудимых тканей n Снижение лабильности – утомление, холод, нагревание, сильный ток, механическое давление, наркотики, солевые растворы. n
Повышение лабильности способность воспроизводить более высокий ритм раздражения n Процесс усвоения ритма (Ухтомский) – опыт Г. Мевеса на изолированном нервном волокне лягушки – укорачивается время абсолютной рефрактерности. n
Мевеса на изолированном нервном волокне лягушки – укорачивается время абсолютной рефрактерности. n
Вывод чем меньше рефрактерность, тем ткань более лабильна, а значит более возбудима n имеет короткую хронаксию, низкий порог раздражения и обладает быстрой аккомодацией n
Свойства возбудимых тканей Возбудимость n Проводимость n Лабильность n Рефрактерность n Сократимость (для мышечной ткани) n
Особенности действия постоянного тока на ткани I. Полярный закон раздражения. II. Закон физиологического электротона. ІІІ. Сокращение мышцы в зависимости от силы постоянного тока и его направления.
I. Полярный закон раздражения Пфлюгера n Постоянный ток оказывает раздражающее действие на ткань только в момент замыкания под катодом и в момент размыкания под анодом.
I. Полярный закон раздражения Пфлюгера n В области приложения к поверхности ткани анода (+) – положительный потенциал на наружной стороне мембраны возрастает – происходит пассивная гиперполяризация, а прикладывании катода (-) положительный потенциал на наружной стороне мембраны снижается – возникает пассивная деполяризация.
I. Полярный закон раздражения Пфлюгера n n Увеличение мембранного потенциала под анодом (пассивная гиперполяризация) не сопровождается повышением ионной проницаемости мембраны даже при большой силе приложенного тока. Поэтому при замыкании постоянного тока возбуждение под анодом не возникает.
I. Полярный закон раздражения Пфлюгера n n Уменьшение мембранного потенциала под катодом (пассивная деполяризация) влечет за собой кратковременное повышение проницаемости мембраны для ионов натрия и медленно развивающееся стойкое повышение проницаемости ее для ионов калия. Поэтому при замыкании постоянного тока возбуждение возникает под катодом.
I. Полярный закон раздражения Пфлюгера n n Мгновенное замыкание приводит к постепенному возрастанию поляризации на аноде (гиперполяризации) и уменьшению поляризации на катоде (деполяризации). При размыкании происходит медленное возвращение мембранного потенциала к исходному уровню.
II. Закон физиологического электротона при длительном прохождении постоянного тока в ткани возбудимость меняется: Ø повышается на катоде – катэлектротон, порог раздражения понижается. Ø снижается на аноде – анэлектротон, порог раздражения повышается. n
II. Закон физиологического электротона n n при длительном действии постоянного тока под катодом начальное повышение возбудимости (катэлектротон) сменяется ее понижением – катодическая депрессия (Б. Ф. Вериго) под анодом сниженная возбудимость постепенно повышается – анодическая экзальтация
Физиологический электротон n n n Изменения мембранного потенциала и возбудимости ткани при действии постоянного тока а – изменения мембранного потенциала; б — изменения критического уровня деполяризации; в — изменения возбудимости. Примечание: стрелкой отмечено начало действия тока
ІІІ. Сокращение мышцы в зависимости от силы постоянного тока и его направления n n Если катод располагается дальше от мышцы, а анод ближе к мышце, то это восходящее направление тока. Если катод ближе к мышце, а анод дальше, то это направление – нисходящее. ↑ +а — к ↓ — к +а Восходящее Нисходящее По силе ток слабый – пороговый, средний – несколько выше порогового и сильный, вызывающий достаточно быстрое перемещение ионов к полюсам (физический электротон).
Если катод ближе к мышце, а анод дальше, то это направление – нисходящее. ↑ +а — к ↓ — к +а Восходящее Нисходящее По силе ток слабый – пороговый, средний – несколько выше порогового и сильный, вызывающий достаточно быстрое перемещение ионов к полюсам (физический электротон).
Действие постоянного тока различной силы и направления n n Слабый ток независимо от направления вызывает возбуждение нерва и сокращение мышцы только в момент замыкания. Средний ток независимо от направления вызывает возбуждение нерва и сокращение мышцы как при замыкании, так и при размыкании тока.
Действие постоянного тока различной силы и направления n n Сильный ток при нисходящем направлении, когда к мышце ближе катод вызывает сокращение мышцы при замыкании. Сильный ток при восходящем направлении, когда к мышце ближе анод вызывает сокращение мышцы при размыкании.
Контрольные вопросы (обратная связь) 1. Что такое пороговая сила? 2. Как зависит возбуждение от силы раздражителя? 3. Как соотносятся между собой сила раздражителя и время его действия? 4. Для каких тканей характерен закон «все или ничего» ? 5. Что происходит с возбудимыми тканями при медленном нарастании силы раздражителя?
Как зависит возбуждение от силы раздражителя? 3. Как соотносятся между собой сила раздражителя и время его действия? 4. Для каких тканей характерен закон «все или ничего» ? 5. Что происходит с возбудимыми тканями при медленном нарастании силы раздражителя?
Nerve Excitability Assessment in Chemotherapy-induced Neurotoxicity
1. Подготовка пациента
- Пациенты называют из отдела медицинской онкологии для базового возбудимости нервных тестирования до начала на химиотерапию.
- Пациент пригодности для возбудимость тестирования должны быть определены. Пациенты должны быть исключены, если они имеют в анамнезе или базовые нейрофизиологические доказательства периферическая невропатия, ранее получавших нейротоксическое химиотерапии или есть какие-то противопоказания для возбудимость тестирования.
2. Аксонального процедуры Возбудимость
- Проведение сенсорной и моторной возбудимостью протоколов на срединный нерв, с помощью полуавтоматической системы компьютеризированного QTracS (Институт неврологии, королева площади, Великобритания), изолированной линейной биполярной постоянного тока стимулятор (Digitimer, Welwyn Garden City, Великобритания) и усилителя ( Sapphire IIA, Medelec, Великобритания).

- Подготовка поверхности кожи на запястье и предплечье с абразивным гелем или колодки уменьшить кожисопротивления, с последующим применением спиртом.
- Подготовьте место для записей, двигатель с неполяризуемых электродами, расположенными на мышцы живота похитителя pollicis Brevis и 4 см дистального электрода для записи соединения двигателя потенциалы действия (CMAPs).
- Подготовьте место для сенсорной записи — с помощью кольца электродами, помещенными в проксимальных и дистальных межфаланговых суставов для записи и электродов, соответственно, для записи соединения сенсорных потенциалов действия (CSAPs).
- Поместите электрохирургического нейтральной пластины земли в ладони, с проводящим гелем.
- Электрические помехи в настройки записи следует удалить как можно больше, используя Humbug 50/60 Гц Шум выпрямитель (Quest Scientific Instruments, Северный Ванкувер, Канада).
- Стимулирование срединного нерва в запястье. Стимуляция сайт должен быть выбран в качестве места низкий порог, используя перемещаемый биполярного электрода, а затем не рolarizable электрода.
- Анодный электрод должен быть помещен 10 см проксимальнее стимулирующий электрод на кости.
- Мониторинг температуры в месте раздражения на протяжении всего испытания и убедиться, что температура превышает 32 ° C.


3. Аксонального протоколы Возбудимость
- Запись стимул-реакция кривой постепенного увеличения стимула до реакции максимальна и не увеличивает стимул, когда интенсивность еще более возрастает.
- Целевая амплитуда порог слежения автоматически устанавливается на 30% -40% от максимальной амплитудой, соответствующей области крутому склону на стимул-реакция кривой.
- В соответствии с методом Босток и соавт., 1998, изменения в порогового тока, необходимое для достижения целевого амплитуда отслеживаются онлайн.
- Запись нескольких параметров возбудимости, в том числе порог электротон (TE), восстановление цикла (RC) и текущего порога (I / V) отношениякак описано в Kiernan и соавт., 2000 и Kiernan и соавт., 2001.
- Порог электротон оценивается по 100 мс подпороговых поляризационные токи, с поляризационным текущий набор в пределах ± 40% порогового контроля (рис. 1). Изменение порогового тока, необходимые для поддержания амплитуды целевой ответа после как де-и гипер-поляризация записывается. Порог электротон дается оценка межузловых проводимости и мембранного потенциала, с ответами в гиперполяризующих направлении в конце поляризующего импульса (90-100 мс) тесно связан с мембранным потенциалом.
- Восстановление цикла оценивали с помощью парного импульса парадигмы (рис. 2), с начальным сверхпиковым стимул кондиционирования следуют через разные промежутки на тестовый стимул (от 2,5 мс до 200 мс). После сверхпиковым стимул, труднее для создания последующих ответов, называется «огнеупорность», что отражает инактивации напряжения гated Na +-каналов. После рефрактерный период, в течение содействие известный как superexcitability происходит.
- Текущий порог отношений оценивается с помощью поляризационного тока 200 мс, которые различаются по силе от +50% до 100% порога.


4. Оценка пациентов
- Используя эти методы, пациенты оценивали как остро и продольно по лечению химиотерапией. Для оценки острой нейротоксичности, пациенты возвращаются для последующей химиотерапией оценки в течение 48 часов с момента получения обращения.
- Для изучения хронической нейротоксичности, оценки, принятых до вливания химиотерапии по сравнению продольно по лечению циклов.
- В дополнение к аксонального тестирование возбудимости, обычной клинической классификации весы должны быть использованы для оценки вызванной химиотерапией нейротоксичности, в том числе Национальный институт рака — Common Criteria для неблагоприятных событий сенсорной невропатии подшкале, Общая оценка нейропатия и погладитьдиентом, сообщили результат оценки.
5. Анализ и интерпретация
- Для выявления острой нейротоксичности, сравнение результатов до и после химиотерапии. Для оценки развития продольных изменений в нервной функции, сравнение результатов по лечению. Основные параметры для оценки включают огнеупорность, superexcitability, протяженность порога изменение порога электротон (гиперполяризующих 90-100 мс), в дополнение к обычным параметрам, как максимальная амплитуда и время задержки.
- Для оценки общего изменения возбудимости параметры по лечению, композитный оценка возбудимости рассчитывается. Изменения в трех параметров — superexcitability, огнеупорность и порог электротон (гиперполяризующих 90-100 мс) — суммируется от начального до окончательного лечения, чтобы дать общий маркер изменения.
 Основные параметры для оценки включают огнеупорность, superexcitability, протяженность порога изменение порога электротон (гиперполяризующих 90-100 мс), в дополнение к обычным параметрам, как максимальная амплитуда и время задержки.
Основные параметры для оценки включают огнеупорность, superexcitability, протяженность порога изменение порога электротон (гиперполяризующих 90-100 мс), в дополнение к обычным параметрам, как максимальная амплитуда и время задержки.6. Представитель Результаты
Примеры возбудимость приводит к погладитьдиентом получавших оксалиплатин предоставляются. Сразу после инфузии оксалиплатина, резкие изменения как в сенсорной и моторной возбудимостью разработки, указывающие на развитие функциональных Na + channelopathy 10 -13. Тем не менее, существенные изменения в нескольких параметров возбудимость постепенно развивается через оксалиплатин лечение только в сенсорных аксонов, с аксонов двигателя не влияет (рис. 3), что отражает широкое сенсорных аксонального повреждения и изменения мембранного потенциала. Эта модель соответствует клинические проявления симптомов при хроническом оксалиплатин-индуцированной нейротоксичности. Возбудимость изменения в сенсорной аксонов предшествует снижение максимальной амплитудой, как оценивали с помощью традиционных методов нервной проводимости, и показывают, что аксонального методы возбудимость может служить чувствительным инструментом оценки рано оксалиплатин-индуцированной нейротоксичности.
3), что отражает широкое сенсорных аксонального повреждения и изменения мембранного потенциала. Эта модель соответствует клинические проявления симптомов при хроническом оксалиплатин-индуцированной нейротоксичности. Возбудимость изменения в сенсорной аксонов предшествует снижение максимальной амплитудой, как оценивали с помощью традиционных методов нервной проводимости, и показывают, что аксонального методы возбудимость может служить чувствительным инструментом оценки рано оксалиплатин-индуцированной нейротоксичности.
Рисунок 1. Ронг> Threshold электротон, изображающие сигналы в ответ на длительное подпороговых поляризующего тока (100 мс), с гиперполяризующих направлении построена в нижнем квадранте (синий) и деполяризующего направление укладывается в верхний квадрант (красный). Ниже стимул напряжения на генерацию порога срабатывания электротон.
Рисунок 2. Восстановление цикла возбудимости, демонстрируя характерные последовательности возбудимость изменения после проведения импульса, с периодом снижения возбудимости (огнеупорность) до 3 мс после стимула сверхпиковым, после периода повышенной возбудимости (superexcitability) с максимумом на 5 -7 мс, а затем уменьшить возбудимость (subexcitability). Парных импульсов парадигма стимул протокол вставкой.
Парных импульсов парадигма стимул протокол вставкой.
ИнжирЮр 3. возбудимость изменения в сенсорной аксонов оксалиплатин пациентов, с базовой записи черным цветом и последующей обработки записей белого цвета, после 4-6 месяцев лечения оксалиплатина. Эти изменения, как считается, отражают широкое распространение аксонального повреждения и изменения мембранного потенциала. Картина возбудимость настройка в условиях онкологии показан слева. Нажмите здесь, чтобы увеличить изображение .
Критические параметры для диагностических исследований при оказании неотложной помощи
Газы крови.pH — Потенциал водорода
Степень кислотности или щелочности любой жидкости (включая кровь) – это функция ее концентрации ионов водорода ([H+], а pH – это просто способ выражения активности ионов водорода. Отношение pH и концентрации ионов водорода описывается следующим образом [1]:
pH = -log aH+
где aH+ – активность ионов водорода.
Низкое значение pH связано с ацидозом, а высокое – с алкалозом.
Параметр pH имеется в следующих продуктах:
pCO2 – Парциальное давление углекислого газа
Углекислый газ (CO2) – это кислотный газ. Количество CO2в крови в большой степени контролируется частотой и глубиной дыхания или вентиляции. pCO2 – это парциальное давление CO2в крови. Это мера давления, создаваемого небольшой долей (~5%) общего CO2, которая остается в газообразном состоянии, растворенная в плазме крови [2]. pCO2 – это дыхательная составляющая кислотно-щелочного равновесия, отражающая адекватность вентиляции легких. Степень поражения, а также хронический характер можно оценить по сопутствующим изменениям кислотно-щелочного состояния
Параметр pCO2 имеется в следующих продуктах:
pO2 – Парциальное давление кислорода
Количество кислорода в крови контролируется множеством переменных, например вентиляцией/перфузией. pO2 – это парциальное давление кислорода в газовой фазе в равновесии с кровью. pO2отражает только малую долю (1–2%) общего кислорода в крови, растворенного в плазме крови [3]. Оставшиеся 98–99% кислорода, присутствующего в крови, связаны с гемоглобином в эритроцитах. pO2в первую очередь отражает поглощение кислорода легкими.
pO2 – это парциальное давление кислорода в газовой фазе в равновесии с кровью. pO2отражает только малую долю (1–2%) общего кислорода в крови, растворенного в плазме крови [3]. Оставшиеся 98–99% кислорода, присутствующего в крови, связаны с гемоглобином в эритроцитах. pO2в первую очередь отражает поглощение кислорода легкими.
Параметр pO2 имеется в следующих продуктах:
ЭлектролитыcNa+ – Натрий
Натрий (Na+) является доминирующим катионом во внеклеточной жидкости, где имеет концентрацию в 14 раз выше (∼140 ммоль/л), чем во внутриклеточной жидкости (∼10 ммоль/л). Na+вносит большой вклад в осмоляльность внеклеточной жидкости, и его основная функция в большой степени заключается в контроле и регуляции водного баланса, а также поддержании кровяного давления. Na+также важен для передачи нервных импульсов и активации сокращения мышц.
Параметр cNa+ имеется в следующих продуктах:
cK+ – Калий
Калий (K+) является основным катионом во внутриклеточной жидкости, где имеет концентрацию в 25–37 раз выше (∼150 ммоль/л в клетках тканей, ∼105 ммоль/л в эритроцитах), чем во внеклеточной жидкости (∼4 ммоль/л) [4, 5]. K+имеет несколько жизненно-важных функций в организме, например регуляция нервно-мышечной возбудимости, сердечного ритма, внутриклеточного и внеклеточного объема и кислотно-щелочного состояния.
Параметр cK+ имеется в следующих продуктах:
cCa2+ – Кальций
Ион кальция (Ca2+) – один их самых преобладающих катионов в организме, где приблизительно 1% присутствует во внеклеточной жидкости крови. Ca2+играет ключевую роль в костной минерализации и множестве клеточных процессов, например сократительной способности сердца и скелетной мускулатуры, нервно-мышечной передаче и гормональной секреции, а также действует в различных ферментных реакциях, таких как коагуляция крови.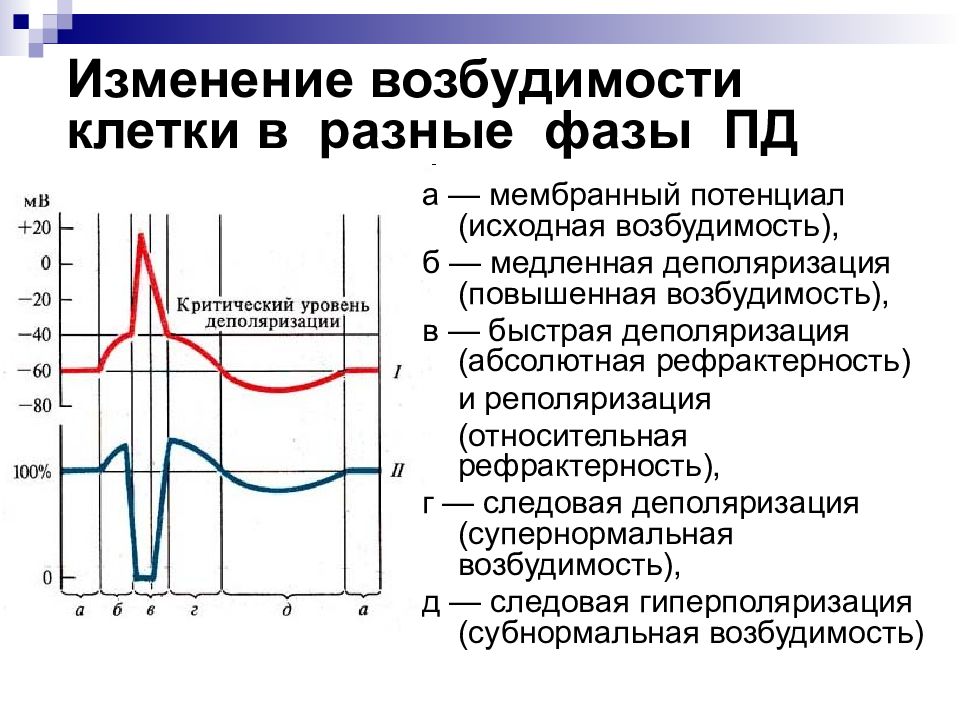
Параметр cCa2+ имеется в следующих продуктах:
cCl— – Хлорид
Хлорид (Cl—) является основным анионом во внеклеточной жидкости и одним из важнейших анионов в крови. Основная функция Cl—заключается в поддержании осмотического давления, жидкостного баланса, мышечной активности, ионной нейтральности в плазме. Он также помогает установить причину кислотно-щелочных нарушений.
Параметр cCl— имеется в следующих продуктах:
МетаболитыcGlu – Глюкоза
Глюкоза, самый обильный углевод в метаболизме человека, служит крупным источником внутриклеточной энергии (см. лактат). Глюкоза преимущественно производится из пищевых углеводов, но также – в основном в печени и почках – посредством анаболического процесса глюконеогенез и расщепления гликогена (гликогенолиз). Это эндогенная глюкоза помогает сохранять концентрацию глюкозы в крови в пределах нормы, когда пищевая глюкоза недоступна, например между приемами пищи или в период голодания.
Параметр cGlu имеется в следующих продуктах:
cLac – Лактат
Лактат, анион, являющийся результатом диссоциации молочной кислоты, это внутриклеточный метаболит глюкозы. Он производится клетками скелетной мускулатуры, красными кровяными тельцами (эритроцитами), головным мозгом и другими тканями в процессе анаэробного производства энергии (гликолиза). Лактат формируется во внутриклеточной жидкости из пирувата. Катализатором реакции является фермент лактатдегидрогеназ (LDH) [6].
Параметр cLac имеется в следующих продуктах:
cCrea – Креатинин
Креатинин – это эндогенный продукт мышечного метаболизма, производимый из креатина, крайне важной молекулы для производства энергии внутри клеток. Креатинин выводится из организма с мочой, и его концентрация в крови отражает клубочковую фильтрацию и, соответственно, функцию почек.
Параметр cCrea имеется в следующих продуктах:
ОксиметрияctHb – Общий гемоглобинКонцентрация общего гемоглобина (ctHb) в крови включает оксигемоглобин (cO2Hb), дезоксигемоглобин (cHHb), а также дисфункциональные виды гемоглобина, не способные связывать кислород:
карбоксигемоглобин (cCOHb) (см.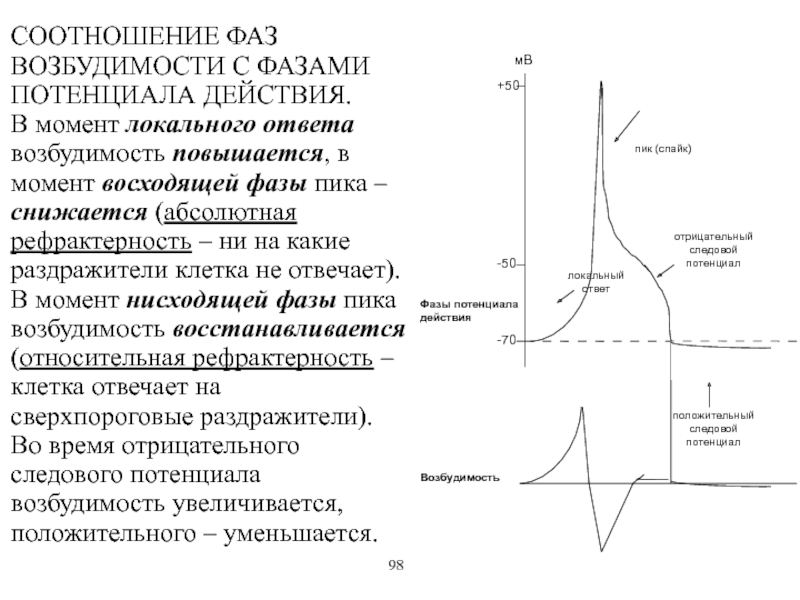 COHb), метгемоглобин (cMetHb) (см. MetHb) и сульфгемоглобин cSulfHb).
COHb), метгемоглобин (cMetHb) (см. MetHb) и сульфгемоглобин cSulfHb).
Таким образом:
ctHb = cO2Hb + cHHb + cCOHb + cMetHb + cSulfHb
Редкий sulfHb не включается в отчет по ctHb в большинстве оксиметров.
Параметр ctHb имеется в следующих продуктах:
MetHb – Метгемоглобин
FMetHb – этой доля общего гемоглобина (ctHb), присутствующего в виде метгемоглобина (MetHb). Как правило, эта доля выражается в виде процентов (%) [1].
В большинстве медицинских текстов MetHb(a) обозначается просто как MetHb.
Параметр MetHb имеется в следующих продуктах:
COHb – Карбоксигемоглобин
FCOHb – этой доля общего гемоглобина (ctHb), присутствующего в виде карбоксигемоглобина (COHb). Как правило, эта доля выражается в виде процентов (%). [1]
В диапазоне 0–60% COHb в артериальной (COHb(a)) и венозной крови (COHb(v)) схожи, т. е. можно делать анализ как венозной, так и артериальной крови [7]. В большинстве медицинских текстов FCOHb(a) обозначается просто как COHb.
е. можно делать анализ как венозной, так и артериальной крови [7]. В большинстве медицинских текстов FCOHb(a) обозначается просто как COHb.
Параметр COHb имеется в следующих продуктах:
sO2 – Сатурация кислорода
Сатурация кислорода (sO2) – это отношение концентрации оксигемоглобина к концентрации функционального гемоглобина (например оксигемоглобина (O2Hb) и дезоксигемоглобина (Hhb), способных переносить кислород [1].
Значение sO2отражает утилизацию способности транспортировать доступный в данный момент кислород.
В артериальной крови 98–99% кислорода транспортируется в эритроцитах, связанных с гемоглобином. Оставшиеся 1–2% кислорода, транспортируемые в крови, растворены в плазме крови. Эта часть указывается как парциальное давление кислорода (pO2) [8].
Параметр sO2 имеется в следующих продуктах:
FO2Hb – Доля оксигемоглобина
FO2Hb в общем гемоглобине крови.
Параметр FO2Hb имеется в следующих продуктах:
FHHb – Доля дезоксигемоглобина
FHHb в общем гемоглобине крови.
Параметр FHHb имеется в следующих продуктах:
FHbF – Доля фетального гемоглобина
FHHb в общем гемоглобине крови.
Параметр FHbF имеется в следующих продуктах:
ctBil – Билирубин
Билирубин – желтый продукт распада гем группы гемоглобина. Он транспортируется в крови от места производства, т. е. ретикулоэндотелиальной системы, в печень, где он биотрансформируется перед экскрецией в желчь. Желтуха, патологический желтый цвет кожи, вызывается нарушением аккумуляции билирубина в тканях в всегда ассоциируется с повышенной концентрации билирубина в крови (гипербилирубинемия).
Параметр ctBil имеется в следующих продуктах:
Гематокрит
Hct – Гематокрит
Гематокрит, отношение объема эритроцитов и объема всей крови.
Параметр Hct имеется в следующих продуктах:
Ссылки
- CLSI. Blood gas and pH analysis and related measurements; Approved Guidelines. CLSI document CA46-A2, 29, 8. Clinical and Laboratory Standards Institute, 940 West Valley Road, Suite 1400, Wayne, Pennsylvania 19087-1898 USA, 2009.
- Higgins C. Parameters that reflect the carbon dioxide content of blood. www.acutecaretesting.org Oct 2008.
- Wettstein R, Wilkins R. Interpretation of blood gases. In: Clinical assessment in respiratory care, 6th ed. St. Louis: Mosby, 2010.
- Burtis CA, Ashwood ER, Bruns DE. Tietz textbook of clinical chemistry and molecular diagnostics. 5th ed. St. Louis: Saunders Elsevier, 2012.
- Engquist A. Fluids/Electrolytes/Nutrition. 1st ed. Copenhagen: Munksgaard, 1985.
- Robergs RA, Ghiasvand F, Parker D. Biochemistry of exercise-induced metabolic acidosis. Am J Physiol Regul Integr Comp Physiol 2004; 287: R502-16.
- Lopez DM, Weingarten-Arams JS, Singer LP, Conway EE Jr. Relationship between arterial, mixed venous and internal jugular carboxyhemoglobin concentrations at low, medium and high concentrations in a piglet model of carbon monoxide toxicity. Crit Care Med 2000; 28: 1998-2001.
- Higgins C. Why measure blood gases? A three-part introduction for the novice. Part 1. www.acutecaretesting.org Jan 2012.

Поделиться этой страницей
Вопросы к кандидатским экзаменам 2
Вопросы к кандидатскому экзамену по дисциплине 03.03.01 «Физиология»
1. Предмет, цели и задачи физиологии.
2. Основные этапы развития физиологии.
3. Особенности современного периода развития физиологии.
4. Раздражимость, возбудимость как основа реакции ткани на раздражение.
5. Современные представления о строении и функции мембран. Активный и пассивный транспорт через мембраны.
6. Электрические явления в возбудимых тканях. История их открытия.
История их открытия.
7. Мембранный потенциал и его происхождение.
8. Потенциал действия и его фазы. Соотношение фаз возбудимости с фазами потенциала действия.
9. Одиночные сокращения и его виды. Тетанус. Факторы, влияющие на его величину.
10. Оптимум и пессимум раздражения.
11. Современная теория мышечного сокращения и расслабления.
12. Определение силы мышечного сокращения.
13. Распространение возбуждения по бeзмиелиновым и миелиновым волокнам.
14. Характеристика их возбудимости и лабильности.
15. Особенности строения и функционирования гладких мышц.
16. Строение и классификация синапсов. Механизм передачи возбуждения в синапсах.
17. 1Особенности строения и передачи возбуждения в нервно-мышечных синапсах.
18. Медиаторы, их синтез, секреция, взаимодействие с рецепторами.
19. Нейрон как структурная и функциональная единица ЦНС, его физиологические свойства и взаимосвязь с глиальными клетками.
20. Особенности передачи возбуждения в синапсах ЦНС. Возбуждающие синапсы и разнообразие медиаторов в ЦНС (ВПСП).
Особенности передачи возбуждения в синапсах ЦНС. Возбуждающие синапсы и разнообразие медиаторов в ЦНС (ВПСП).
21. Общие принципы координационной деятельности ЦНС.
22. Свойства нервных центров.
23. Структурно-функциональные особенности соматической и вегетативной нервной системы.
24. Торможение в ЦНС (И.М.Сeченов), его виды и роль. Современные представления о механизмах центрального торможения.
25. Основные принципы и особенности распространения возбуждения в ЦНС.
26. Конвергенция, дивергенция, одностороннее проведение.
27. Современные представления о функциональной организации мозга.
28. Роль спинного мозга в процессах регуляции деятельности опорно-двигательного аппарата и вегетативных функций организма. Характеристика спинальных животных. Спинальные рефлексы.
29. Продолговатый мозг и мост, их участие в процессах саморeгуляции функций.
30. Центры продолговатого мозга.
31. Физиология среднего мозга, его рефлекторная деятельность. Дeцеребрационная ригидность и механизм её возникновения. Роль среднего и продолговатого мозга в регуляции мышечного тонуса.
Дeцеребрационная ригидность и механизм её возникновения. Роль среднего и продолговатого мозга в регуляции мышечного тонуса.
32. Физиология мозжечка, его влияние на моторику и вегетативные функции организма.
Нарушение двигательной функции при поражении мозжечка.
33. Ретикулярная формация ствола мозга. Восходящие активирующие влияния на кору больших полушарий.
34. Гипоталамус. Характеристика основных ядерных групп. Участие гипоталамуса в рeгуляции вегетативных функций и в формировании эмоций и мотиваций.
35. Таламус. Функциональная характеристика основных ядерных групп.
36. Лимбичeская система и ее участие в формировании целостных поведенческих реакций организма.
37. Особенности строения различных отделов коры больших полушарий головного мозга.
38. Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы, синергизм и относительный антагонизм их влияния.
39. Роль ретикулярной формации, мозжечка и коры больших полушарий в регуляции деятельности вегетативной нервной системы.
40. Свойства вегетативных ганглиев.
41. Классификация рефлексов. Рефлекторный путь. Обратная aфферентация, её значение.
42. Физиологические механизмы образования условных рефлексов, их структурно-функциональная основа. Развитие представлений И.П.Пaвлова о механизмах формирования временных связей.
43. Условный рефлекс как форма приспособления животных и человека к изменяющимся условиям существования. Классификация условных рефлексов.
44. Химическая классификация гормонов. Современные представления о механизмах взаимодействия гормонов с клетками-мишенями. Центральные и периферические механизмы регуляции функций желез внутренней секреции.
45. Гормоны гипофиза, его функциональные связи с гипоталамусом и участие в регуляции деятельности эндокринных органов.
46. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функций организма.
47. Физиология щитовидной и околощитовидной желёз. Роль передней доли гипофиза в регуляции функций щитовидной железы.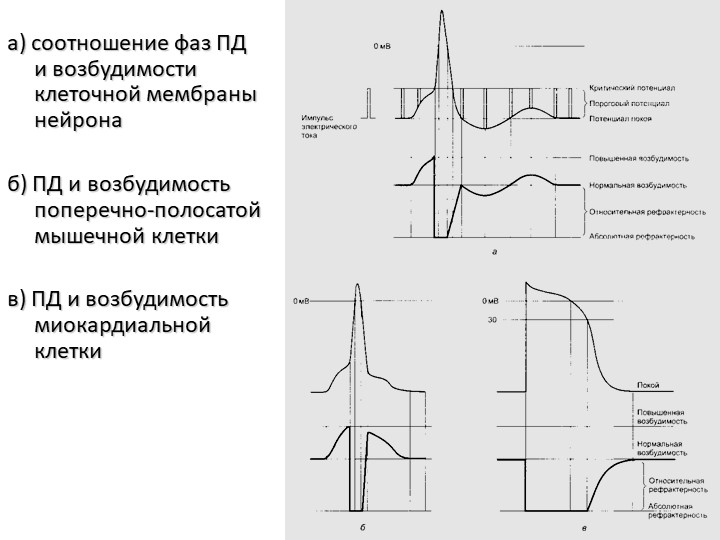
48. Поджелудочная железа и ее гормональная функция. Значение инсулина в углеводной обмене.
49. Половые гормоны.
50. Участие эндокринных желез в адаптации организма к нагрузкам, в том числе к экстремальным.
51. Понятие об обмене веществ в организме. Процессы ассимиляции и диссимиляции веществ. Пластическая и энергетическая роль питательных веществ.
52. Теплопродукция. Обмен веществ как источник образования тепла. Роль отдельных органов в теплопродукции.
53. Принципы составления рационов.
54. Определение дыхательного коэффициента, его значение для расчёта расхода энергии.
55. Основной обмен и значение его определения для клиники.
56. Значение минеральных веществ, микроэлементов и витаминов в организме.
57. Пищеварение в полости рта. Состав и физиологическая роль слюны.
58. Слюноотделение, его регуляция.
59. Методы исследования функций желудочно-кишечного тракта у животных и человека.
60. Пищеварение в желудке. Состав и свойства желудочного сока. Регуляция желудочной секреции. Фазы отделения желудочного сока.
Состав и свойства желудочного сока. Регуляция желудочной секреции. Фазы отделения желудочного сока.
61. Желчеобразования и желчевыделения.
62. Пищеварение в 12-ти перстной кишке. Внешнесекреторная деятельность поджелудочной железы. Регуляция и приспособительный характер панкреатической секреции к видам пищи и пищевым рационам.
63. Особенности пищеварения в толстой кишке.
64. Состав и свойства кишечного сока. Регуляция секреции кишечного сока.
65. Полостной и мембранный гидролиз пищевых веществ в различных отделах тонкой кишки.
66. Моторная и эвакуаторная деятельность желудка, её регуляция.
67. Всасывание веществ в различных отделах ЖКТ. Виды и механизм всасывания веществ через биологические мембраны.
68. Состав крови. Основные физиологические константы крови и механизм их поддержания. Свойства и функции крови.
69. Электролитный состав плазмы крови. Осмотическое давление крови.
70. Функциональная система, обеспечивающая постоянство осмотического давления крови.
71. Белки плазмы крови, их характеристика и функциональное значение. Онкотичeское давление крови и его роль.
72. Функциональная система, поддерживающая постоянство кислотно-щелочного равновесия.
73. Гeмопоэз. Гуморальная регуляция эритро- и лeйкопоэза.
74. Понятие о гемостазе. Процесс свёртывания крови и его фазы. Факторы, ускоряющие и замедляющие свёртывание крови.
75. Характеристика и роль форменных элементов крови: эритроциты, лейкоциты, тромбоциты.
76. Группы крови. Резус-фактор. Переливание крови. Кровезамещающие растворы.
77. Лейкоциты и их виды. Лейкоцитарная формула. Функции различных видов лейкоцитов.
78. Эритроциты, их функции. Виды гемоглобина, его соединения, их физиологическое значение.
79. Физиологические свойства и особенности миокарда. Автоматия сердца.
80. Сердце, значение его камер и клапанного аппарата, изменение давления и объёма крови в полостях сердца в различные фазы кардиоцикла. Систолический и минутный объём крови.
81. Электрокардиография.
82. Соотношение возбуждения, сокращения и возбудимости сердца в разные фазы сердечного цикла. Реакция сердечной мышцы на дополнительное раздражение.
83. Экстрасистолы.
84. Тоны сердца и их происхождение.
85. Регуляция сердечной деятельности (миогенная, гуморальная, нервная).
86. Гуморальная регуляция деятельности сердца.
87. Рефлекторная регуляция деятельности сердца. Характеристика влияний парасимпатических и симпатических нервных волокон на деятельность сердца.
88. Кровяное давление в различных отделах системы кровообращения. Факторы, определяющие его величину.
89. Рефлекторная регуляция системного артериального давления. Значение сосудистых рефлексогенных зон. Сосудодвигательный центр.
90. Основные законы гемодинамики. Факторы, обеспечивающие движение крови по сосудам.
91. Капиллярный кровоток и его особенности. Микроциркуляция и её роль в механизме обмена жидкости и различных веществ между кровью и тканями.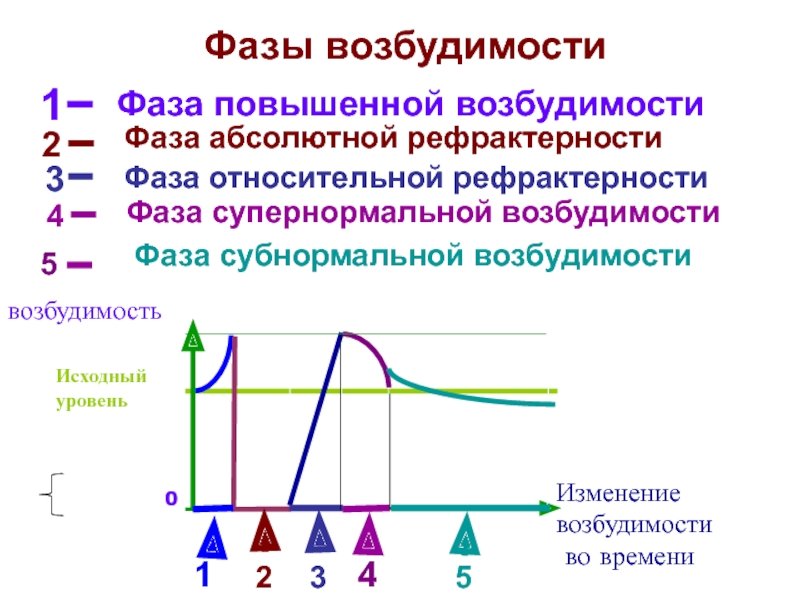
92. Рефлекторная регуляция тонуса сосудов, сосудодвигательный центр.
93. Бескровный метод определения кровяного давления (С.Ривe-Роччи, И.С.Коротков).
94. Дыхание, его основные этапы. Механизм внешнего дыхания. Биомеханика вдоха и выдоха.
95. Методы определения жизненной ёмкости лёгких. Спирометрия, спирография.
96. Давление в плевральной полости, его происхождение и значение в разные фазы дыхательного цикла. Механизм нарушения дыхания при пневмотораксе.
97. Газообмен в лёгких. Парциальное давление газов кислорода и углекислого газа в альвеолярном воздухе и напряжение газов в крови.
98. Определение минутной вентиляции лёгких в разных условиях.
99. Транспорт кислорода кровью. Кривая диссоциации оксигемоглобина, её характеристика.
100. Транспорт углекислого газа кровью. Значение карбоангидразы.
101. Дыхательный центр. Современные представления о его структуре и локализации. Автоматия дыхательного центра.
102. Рефлекторная саморeгуляция дыхания.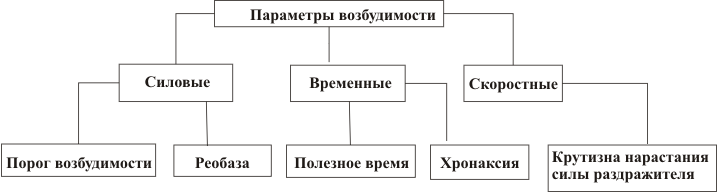 Механизм смены дыхательных фаз.
Механизм смены дыхательных фаз.
103. Регуляторное влияние на дыхательный центр со стороны высших отделов головного мозга (гипоталамуса коры больших полушарий).
104. Роль гуморальных факторов в регуляции дыхания. Роль углекислого газа.
105. Нефрон, строение, кровоснабжение. Механизм образования первичной мочи, её состав.
106. Образование конечной мочи, её состав и свойства. Рeабсорбция в канальцах, механизм её регуляции. Процессы секреции и экскреции в почечных канальцах.
107. Процесс мочеиспускания, его регуляция.
108. Регуляция деятельности почек. Роль нервных и гуморальных факторов.
109. Реобаза размыкательного удара 3 в. Нерв раздражается током в 10 в. Направление тока нисходящее. Что произойдет с мышцей при размыкании цепи раздражающего тока?
110. Нерв между раздражающими электродами перевязан. При действии тока мышца данного нервно-мышечного препарата сокращалась только в момент замыкания. Какой электрод находится ближе к мышце?
111. Какой электрод находится ближе к мышце, если при действии сильного тока сокращение возникает только при замыкании?
Какой электрод находится ближе к мышце, если при действии сильного тока сокращение возникает только при замыкании?
112. Мембранный потенциал нервного волокна равен 100 мв. Критический уровень деполяризации отличается от мембранного потенциала на 30%. Какова реобаза нерва, если 1 в. раздражающего тока сдвигает Ео на 5 мв.?
113. Параметры возбудимости ткани: реобаза, хронаксия, лабильность. Все ли параметры перечислены?
114. Что покажет гальванометр, если: а) микроэлектрод проколол мембрану; б) введен глубоко вглубь клетки?
115. Если бы клеточная мембрана была абсолютно непроницаема для ионов, как бы изменилась величина потенциала покоя?
116. Яд тетродотоксин блокирует электроуправляемые ионоселективные натриевые каналы мембраны возбудимой клетки. Как изменится мембранный потенциал (МП) нервного волокна, если подействовать на него тетродотоксином? Повлияет ли тетродотоксин на проведение возбуждения по данному волокну?
117. При нанесении алкалоида батрахотоксина на нервную клетку в эксперименте существенно увеличивается проницаемость плазматической мембраны для натрия. Как изменяется величина мембранного потенциала покоя (МП) нервной клетки при действии батрахотоксина?
Как изменяется величина мембранного потенциала покоя (МП) нервной клетки при действии батрахотоксина?
118. Гигантский __________аксон кальмара поместили в среду, которая по своему составу соответствовала межклеточной жидкости. При раздражении в аксоне возник ПД. Затем концентрацию ионов натрия в среде уравняли с их концентрацией в аксоне и повторили раздражение. Что обнаружили?
119. Что произойдет с нервной клеткой, если ее обработать цианидами?
120. Животному введена большая доза аминазина, который блокирует восходящую активирующую систему ретикулярной формации мозгового ствола. Как при этом меняется поведение животного и почему?
Физиология Flashcards | Quizlet
Физиология скелетных мышц. Известно три вида мышц:1) исчерченные (поперечно-полосатые) мышцы;
2) сердечная мышца;
3) неисчерченные (гладкие) мышцы.
Поперечно-полосатые скелетные мышцы и их значение, строение и физиологические свойства
Скелетные мышцы у человека занимают примерно 40-50% от массы тела.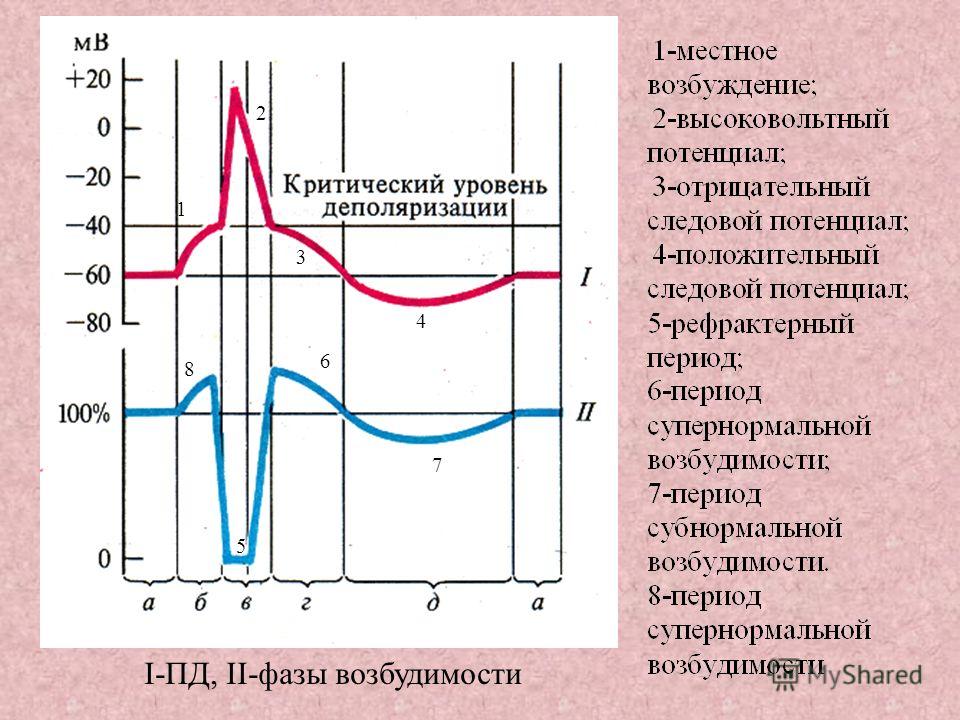 По разным данным, в нашем организме их насчитывается от 400 до 500. Это произвольные мышцы, т.е. они не могут сокращаться без «приказа» из ЦНС. Другими словами, они не обладают
По разным данным, в нашем организме их насчитывается от 400 до 500. Это произвольные мышцы, т.е. они не могут сокращаться без «приказа» из ЦНС. Другими словами, они не обладают
автоматизмом.
Значение скелетных мышц:
1) поддержание позы человека в пространстве;
2) перемещение тела в пространстве;
3) перемещение частей тела относительно друг друга;
4) обеспечение дыхательной функции;
5) выработка тепла;
6) помощь движению крови и лимфы;
7) участие в осуществлении половых функций;
8) механическая защита внутренних органов;
9) депонирование воды и соли;
10) участие в работе произвольных сфинктеров;
11) участие в осуществлении витальных рефлексов.
Физиологические свойства скелетных мышц:
1) возбудимость — способность генерировать потенциал действия;
2) проводимость — способность проводить волну возбуждения;
3) сократимость — способность укорачиваться или развивать напряжение;
4) эластичность — способность развивать напряжение при растягивании.
Свойства гладкой мышечной ткани
1. Возбудимость и проводимость ниже, чем у скелетных мышц;
2. 2. Удлинен латентный (скрытый) период до 1 сек.;
3. Сократимость миофибрил осуществляется с участием белков тропомиозина, тропонина и ионов Са++. Продолжительность до 100 сек.;
4. Пластический тонус — гладкие мышцы способны изменять линейные размеры (растягиваться) не изменяя своего тонуса.
5. Автоматизм — способность гладкой мышечной ткани сокращаться под воздействием импульсов, которые зарождаются в ней самой (собственная интрамуральная нервная система — нервные ганглии, которые самостоятельно генерируют потенциал действия).
По сравнению со скелетными мышцами
• менее возбудимы
• медленные движения
• длительные тонические сокращения
• низкая утомляемость
Типы гладкомышечной ткани: Унитарные ГМК — нервное волокно подходит к каждой клетке. Не обладают автоматией. Мышцы зрачка
Мультиунитарные ГМК — нервное волокно подходит к группе клеток. Обладают
автоматией. Мышцы внутренних органов. ГМК связаны щелевыми контактами — нексусами
Мышцы внутренних органов. ГМК связаны щелевыми контактами — нексусами
Возбудимость — обзор | Темы ScienceDirect
6.1.1 Краткое описание мышц млекопитающих
Чтобы воспроизвести аналогичные свойства биологических мышц в искусственных аналогах, мы должны полностью понимать механизмы, поведение и свойства естественных мышц. В этом разделе приводится краткое изложение анатомических и физиологических характеристик мышц человека или млекопитающих.
Мышцы обеспечивают подвижность человеческого тела и составляют 40–45% веса тела.Они обеспечивают средство прямого преобразования химической энергии в механическую (движение, работу и силу) энергию. Myology занимается научным изучением мышц.
Есть три различных типа мышечной ткани, которые различаются по микроскопической анатомии и нервному контролю. Это скелетные, сердечные и гладкие мышцы.
Ткани скелетных мышц в основном прикреплены к костям и обеспечивают движение скелета. Они полосатые; чередующиеся светлые и темные полосы видны при исследовании ткани под микроскопом. Они также классифицируются как произвольные мышечные ткани, поскольку находятся под сознательным контролем.
Они также классифицируются как произвольные мышечные ткани, поскольку находятся под сознательным контролем.
Ткань сердечной мышцы находится только в сердце. Он также имеет поперечно-полосатую форму, но непроизвольно, то есть его сокращение обычно не контролируется сознанием и использует кардиостимулятор, включенный в мышцу, чтобы произвести сердцебиение.
Гладкая мышечная ткань находится в стенках полых внутренних структур, таких как кровеносные сосуды, желудок, кишечник и большинство органов брюшной полости. Он беспорядочный и непроизвольный.
За счет сокращения или попеременного сокращения и расслабления мышечная ткань выполняет три основных функции: движение, стабилизация и выработка тепла.
Существует четыре следующих функциональных характеристики мышцы:
- 1.
Возбудимость , или способность реагировать на определенные стимулы путем создания потенциалов действия или импульсов электрического сигнала. Потенциалы действия, запускающие стимулы, представляют собой химические вещества, такие как нейротрансмиттеры, выделяемые нейронами, или гормоны, распространяемые кровью.
- 2.
Сократимость , или способность сокращаться и генерировать силу для выполнения работы. Сокращение происходит в ответ на один или несколько потенциалов мышечного действия.
- 3.
Растяжимость , или способность мышцы растягиваться (растягиваться) без повреждения ткани. Большинство скелетных мышц расположены в противоположных или антагонистических парах, в которых одна мышечная ткань сокращается, а другая расслабляется или даже растягивается.
- 4.
Эластичность , или способность мышечной ткани возвращаться к своей исходной форме после стимуляции [1].

Нервы и кровеносные сосуды в изобилии снабжают мышцы. Моторные нейроны отвечают за стимуляцию мышечных волокон. Чтобы мышечная ткань сокращалась, она использует большое количество аденозинтрифосфата (АТФ), молекулы, богатой энергией, и она должна производить АТФ за счет притока питательных веществ и кислорода через кровеносные капилляры (микроскопические кровеносные сосуды), которые в изобилии обнаруживаются в мышечной ткани. Каждое мышечное волокно (клетка) находится в тесном контакте с одним или несколькими капиллярами.
Каждое мышечное волокно (клетка) находится в тесном контакте с одним или несколькими капиллярами.
Соединительная ткань окружает и защищает мышечную ткань. Фасция — это лист широкополосной волокнистой соединительной ткани под кожей, вокруг мышц и других органов тела. Глубокая фасция, плотная соединительная ткань неправильной формы, выстилает стенки тела и конечности, скрепляет мышцы и разделяет их на функциональные группы. Глубокая фасция позволяет мышцам свободно двигаться; несет нервные, кровеносные и лимфатические сосуды; и заполняет промежутки между мышцами.За глубокой фасцией находятся три слоя плотных соединительных тканей неправильной формы, которые дополнительно защищают и укрепляют скелетные мышцы.
Самый внешний слой, окружающий всю мышцу, — это эпимизий . Перимизий затем окружает пучки ( faciculi или пучков ) из 10–100 или более отдельных мышечных волокон. Далее проникает внутрь каждого отдельного пучка и отделяет друг от друга мышечные волокна эндомизий . Заинтересованным читателям предлагается прочитать Приложение А к исх. 1.
Заинтересованным читателям предлагается прочитать Приложение А к исх. 1.
Моторный нейрон доставляет стимул, который в конечном итоге заставляет мышечное волокно сокращаться. Моторный нейрон вместе со всеми мышечными волокнами, которые он стимулирует, называется двигательной единицей .
Типичная скелетная мышца состоит из сотен или тысяч очень длинных цилиндрических клеток, называемых мышечных волокон . Мышечные волокна расположены параллельно друг другу и имеют диаметр от 10 до 100 мкм. Хотя типичная длина составляет 100 мкм, некоторые мышечные волокна достигают 30 см в длину.
Сарколемма — это плазматическая мембрана мышечного волокна, окружающая цитоплазму или саркоплазму мышечного волокна. Поскольку волокна скелетных мышц возникают в результате слияния множества более мелких клеток во время эмбрионального развития, каждое волокно имеет множество ядер, направляющих синтез новых белков. Ядра находятся на периферии клетки рядом с сарколеммой, удобно в стороне от сократительных элементов. Митохондрии (энергетические пакеты) лежат рядами по всему мышечному волокну, стратегически близко к мышечным белкам, которые используют АТФ для продолжения процесса сокращения.Внутри мышечных волокон находится миофибрилл , которые вытянуты вдоль саркоплазмы. Их яркие светлые и темные цвета полос, называемые поперечными полосами , заставляют все мышечные волокна казаться полосатыми или полосатыми.
Митохондрии (энергетические пакеты) лежат рядами по всему мышечному волокну, стратегически близко к мышечным белкам, которые используют АТФ для продолжения процесса сокращения.Внутри мышечных волокон находится миофибрилл , которые вытянуты вдоль саркоплазмы. Их яркие светлые и темные цвета полос, называемые поперечными полосами , заставляют все мышечные волокна казаться полосатыми или полосатыми.
Миофибриллы образуют сократительный элемент скелетной мышцы. Они имеют диаметр 1-2 мкм и содержат три типа более мелких филаментов, называемых миофиламентами . Это тонкие, толстые и эластичные нити соответственно.Тонкая нить составляет около 8 нм; толщина волокон составляет около 16 нм.
Переломные точки, вызванные дрейфом параметров в модели возбудимого океана
Хидэ Р. Некоторые эксперименты по тепловой конвекции во вращающейся жидкости. Q. J. R. Meteorol. Soc. 79 , 161 (1953).
ADS Статья Google Scholar

Стоммель, Х. Термохалинная конвекция с двумя устойчивыми режимами течения. Tellus 2 , 244–230 (1961).
Google Scholar
Лоренц, Э. Н. Детерминированный непериодический поток. J. Atmos. Sci. 20 , 130–141 (1963).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Веронис, Г. Анализ ветровой циркуляции океана с ограниченным числом компонентов Фурье. J. Atmos. Sci. 20 , 577–593 (1963).
ADS Статья Google Scholar
Гил М., Рид П. и Смит Л. Геофизические потоки как динамические системы: влияние экспериментов Хиде. Astron. Geophys. 51 , 4–28 (2010).
Артикул Google Scholar
Гил М. и Чайлдресс С. Темы геофизической гидродинамики: динамика атмосферы, теория динамо и динамика климата (Springer, 1987) ( переиздано в pdf, 2012 ).
MATH Книга Google Scholar
Дейкстра, Х. А. и Гил, М. Низкочастотная изменчивость крупномасштабной циркуляции океана: подход динамических систем. Rev. Geophys. 43 , RG3002 (2005).
ADS Статья Google Scholar
Гладуэлл, М. Переломный момент: как мелочи могут иметь большое значение (Little Brown, 2000).
Google Scholar
Lenton, T. M. et al. Опрокидывающие элементы в климатической системе Земли. Proc. Natl. Акад. Sci. США 105 , 1786–1793 (2008).
ADS CAS PubMed МАТЕМАТИКА Статья Google Scholar
Смейл С. Дифференцируемые динамические системы. Бык. Являюсь. Математика. Soc. 73 , 747–817 (1967).
MathSciNet МАТЕМАТИКА Статья Google Scholar

Гукенхаймер Дж. И Холмс П. Дж. Нелинейные колебания, динамические системы и бифуркации векторных полей (Springer, 1983).
MATH Книга Google Scholar
Крауэл Х. и Фландоли Ф. Аттракторы для случайных динамических систем. Probab. Теория Relat. Поля 100 , 365–393 (1994).
MathSciNet МАТЕМАТИКА Статья Google Scholar
Арнольд Л. Случайные динамические системы (Springer, 1998).
MATH Книга Google Scholar
Карвалью А., Ланга Дж. А. и Робинсон Дж. Аттракторы для бесконечномерных неавтономных динамических систем (Springer, 2012).
MATH Google Scholar
Карабалло, Т. и Хан, X. Прикладные неавтономные и случайные динамические системы: прикладные динамические системы (Springer, 2017).
MATH Google Scholar
Гил М. и Лукарини В. Физика изменчивости и изменения климата. Ред. Мод. Phys. 92 , 035002. https://doi.org/10.1103/RevModPhys.92.035002 (2020).
ADS MathSciNet CAS Статья Google Scholar
Куен, К. Математическая основа для критических переходов: бифуркации, быстрые и медленные системы и стохастическая динамика. Physica D 240 , 1020–1035. https://doi.org/10.1016/j.physd.2011.02.012 (2011).
ADS CAS Статья МАТЕМАТИКА Google Scholar
Эшвин, П., Вичорек, С., Витоло, Р. и Кокс, П. Переломные моменты в открытых системах: бифуркация, примеры, вызванные шумом и зависящие от скорости, в климатической системе. Philos. Пер. R. Soc. Математика. Phys. Англ. Sci. 370 , 1166–1184 (2012).
ADS Статья Google Scholar

Федель, У., Писарчик, А. Н. и Шоуолтер, К. Мультистабильность и склонность: от математики и физики до климата и мозга — мини-обзор и предисловие к основной теме. Хаос 28 , 033501 (2018).
ADS PubMed Статья Google Scholar
Гил М. Век нелинейности в науках о Земле. Earth Space Sci. 6 , 1007–1042 (2019).
ADS Статья Google Scholar
Perryman, C. & Wieczorek, S. Адаптация к изменяющейся среде: неочевидные пороги в многомасштабных системах. Proc. R. Soc. А 470 , 20140226 (2014).
ADS MathSciNet PubMed Статья Google Scholar
Эшвин, П., Перриман, К. и Вичорек, С. Сдвиги параметров для неавтономных систем в низкой размерности: опрокидывание, вызванное бифуркацией и скоростью. Нелинейность 30 , 2185 (2017).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Vanselow, A., Wieczorek, S. & Feudel, U. Когда очень медленно становится слишком быстро — коллапс системы хищник-жертва. J. Theor. Биол. 479 , 64–72 (2019).
MathSciNet PubMed Статья Google Scholar
О’Кифф, П. Э. и Вичорек, С. Опрокидывающие явления и точки невозврата в экосистемах: за пределами классических бифуркаций. SIAM J. Appl. Дин. Syst. 19 , 2371–2402 (2020).
MathSciNet МАТЕМАТИКА Статья Google Scholar
Дейкстра, Х.А. Нелинейная физическая океанография: подход динамических систем к крупномасштабной циркуляции океана и Эль-Ниньо 2-е изд.(Спрингер, 2005).
Google Scholar
Гил М., Чекроун М. Д. и Симоннет Э. Климатическая динамика и механика жидкости: естественная изменчивость и связанные с этим неопределенности. Physica D 237 , 2111–2126. https://doi.org/10.1016/j.physd.2008.03.036 (2008).
Д. и Симоннет Э. Климатическая динамика и механика жидкости: естественная изменчивость и связанные с этим неопределенности. Physica D 237 , 2111–2126. https://doi.org/10.1016/j.physd.2008.03.036 (2008).
ADS MathSciNet Статья МАТЕМАТИКА Google Scholar
Чекроун, М.Д., Симоннет, Э. и Гил, М. Стохастическая динамика климата: случайные аттракторы и инвариантные меры, зависящие от времени. Physica D 240 , 1685–1700 (2011).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Дротос, Г., Бодаи, Т. и Тел, Т. Вероятностные концепции в изменяющемся климате: снимок аттрактора. J. Clim. 28 , 3275–3288 (2015).
ADS Статья Google Scholar
Здесь, М., Дротос, Г., Хаспра, Т., Марфи, Дж. И Тел, Т. Теория параллельных реализаций климата как новая основа для анализа телекоммуникационных сетей. Sci. Отчет 7 , 44529 (2017).
Sci. Отчет 7 , 44529 (2017).
ADS PubMed PubMed Central Статья Google Scholar
Tél, T. et al. Теория параллельных реализаций климата. J. Stat. Phys. 179 , 1496–1530 (2020).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Вичорек, С., Эшвин, П., Люк, К. М. и Кокс, П. М. Возбудимость в наклонных системах: нестабильность компостной бомбы. Proc. R. Soc. А 467 , 1243–1269 (2011).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Kiers, C. & Jones, C.К. Р. Т. Об условиях опрокидывания, обусловленного скоростью, в многомерных динамических системах. J. Dyn. Отличаются. Equ. 32 , 483–503 (2020).
MathSciNet МАТЕМАТИКА Статья Google Scholar

Wieczorek, S., Xie, C. & Jones, C. K. R. T. Компактификация для асимптотически автономных динамических систем: теория, приложения и инвариантные многообразия. arXiv препринт arXiv: 2001.08733 (2020).
Гил М. Криотермодинамика: хаотическая динамика палеоклимата. Physica D 77 , 130–159 (1994).
ADS Статья Google Scholar
Хоффман, П. Ф., Кауфман, А. Дж., Халверсон, Г. П. и Шраг, Д. П. Неопротерозойский снежный ком Земля. Science 281 , 1342–1346 (1998).
ADS CAS PubMed Статья PubMed Central Google Scholar
Лукарини В., Фредрих К. и Лункейт Ф. Термодинамический анализ эксперимента по гистерезису земли снежного кома: эффективность, производство энтропии и необратимость. Q. J. R. Meteorol. Soc. 136 , 2–11 (2010).
ADS Статья Google Scholar

Kaszás, B., Feudel, U. & Tél, T. Явление опрокидывания в типичных динамических системах, подверженных дрейфу параметров. Sci. Отчетность 9 , 8654 (2019).
ADS PubMed PubMed Central Статья CAS Google Scholar
Ван дер Поль Б. О релаксационных колебаниях. Lond. Edinb. Dublin Philos. Mag. J. Sci. 2 , 978–992 (1926).
Артикул Google Scholar
Грасман Дж. Релаксационные колебания. В Энциклопедия сложности и системологии (изд.Мейерс, Р. А.) 1475–1488 (Springer, 2015).
Google Scholar
Линднер, Б., Гарсия-Ойалво, Дж., Нейман, А. и Шимански-Гейер, Л. Эффекты шума в возбудимых системах. Phys. Отчет 392 , 321–424 (2004).
ADS Статья Google Scholar

Пиковский А.С. и Куртс Дж. Когерентный резонанс в возбудимых системах, управляемых шумом. Phys. Rev. Lett. 78 , 775–778 (1997).
ADS MathSciNet CAS МАТЕМАТИКА Статья Google Scholar
Распятие, М. Осцилляторы и релаксационные явления в теории климата плейстоцена. Philos. Пер. R. Soc. А 370 , 1140–1165 (2012).
ADS Статья Google Scholar
Dansgaard, W. et al. Доказательства общей нестабильности климата в прошлом по данным ледового керна продолжительностью 250 тыс. Лет. Nature 364 , 218–220 (1993).
ADS Статья Google Scholar
Ганопольски А. и Рамсторф С. Резкие изменения ледникового климата из-за стохастического резонанса. Phys. Rev. Lett. 88 , 038501 (2002).
ADS PubMed Статья CAS Google Scholar

Дитлевсен, П. Д. и Йонсен, С. Дж. Переломные моменты: раннее предупреждение и принятие желаемого за действительное. Geophys. Res. Lett. 37 , L19703 (2010).
ADS Статья Google Scholar
Пельтье, У. Р. и Ветторетти, Дж. Дансгаард-Эшгер Осцилляции, предсказанные в комплексной модели ледникового климата: солевой осциллятор в Атлантике. Geophys. Res. Lett. 41 , 7306–7313 (2014).
ADS Статья Google Scholar
Ветторетти, Дж. И Пельтье, У. Р. Быстрая физика и медленная физика в нелинейных релаксационных колебаниях Дансгаарда – Эшгера. J. Clim. 31 , 3423–3449 (2018).
ADS Статья Google Scholar
Генрих Х. Происхождение и последствия циклического ледового сплава в северо-восточной части Атлантического океана за последние 130 000 лет. Quat. Res. 29 , 142–152 (1988).
Quat. Res. 29 , 142–152 (1988).
Артикул Google Scholar
MacAyeal, D.R.Binge / продувочные колебания лаурентидного ледникового покрова как причина событий Генриха в Северной Атлантике. Палеоокеанография 8 , 775–784 (1993).
ADS Статья Google Scholar
Делворт, Т. Л. и Грейтбэтч, Р.J. Многодесятилетняя изменчивость термохалинной циркуляции, вызванная воздействием атмосферных потоков на поверхности. J. Clim. 13 , 1481–1495 (2000).
ADS Статья Google Scholar
Jungclaus, J. H., Haak, H., Latif, M. & Mikolajewicz, U. Взаимодействие между Арктикой и Северной Атлантикой и многолетняя изменчивость меридиональной опрокидывающейся циркуляции. J. Clim. 18 , 4013–4031 (2005).
ADS Статья Google Scholar

Франккомб, Л. М., Дейкстра, Х. А. и фон дер Хейдт, А. Многолетняя изменчивость, вызванная шумом, в Северной Атлантике: возбуждение нормальных мод. J. Phys. Oceanogr. 39 , 220–233 (2009).
ADS Статья Google Scholar
Оттера, О. Х., Бентсен, М., Дрейндж, Х. и Суо, Л.Внешнее воздействие как метроном многомесячной изменчивости Атлантики. Nat. Geosci. 3 , 688–694 (2010).
ADS CAS Статья Google Scholar
Jiang, S., Jin, F.-F. И Гил, М. Множественные равновесия, периодические и апериодические решения в модели мелководья с двойным круговоротом и ветром. J. Phys. Oceanogr. 25 , 764–786 (1995).
ADS Статья Google Scholar
Пиерини, С. Исследование модели системы Kuroshio Extension: десятилетние хаотические автоколебания. J. Phys. Oceanogr. 36 , 1605–1625 (2006).
J. Phys. Oceanogr. 36 , 1605–1625 (2006).
ADS Статья Google Scholar
Пиерини, С. Когерентный резонанс в модели двойного круговорота Расширения Куросио. J. Phys. Oceanogr. 40 , 238–248 (2010).
ADS Статья Google Scholar
Пиерини, С. Куросио Бимодальность расширения и колебание в северной части Тихого океана: случай внутренней изменчивости, обусловленной внешним воздействием. J. Clim. 27 , 448–454 (2014).
ADS Статья Google Scholar
Taguchi, B. et al. Десятилетняя изменчивость расширения Куросио: наблюдения и ретроспективный прогноз модели с разрешением вихрей. J. Clim. 20 , 2357–2377 (2007).
ADS Статья Google Scholar

Пиерини С., Дейкстра Х. А. и Риччио А. Нелинейная теория бимодальности расширения Куросио. J. Phys. Oceanogr. 39 , 2212–2229 (2009).
ADS Статья Google Scholar
Пиерини С. и Дейкстра Х. А. Низкочастотная изменчивость расширения Куросио. Нелинейные процессы в геофизике. 16 , 665–675 (2009).
ADS Статья Google Scholar
Taguchi, B. et al. Десятилетняя изменчивость расширения Куросио: мезомасштабные водовороты и рециркуляции. Ocean Dyn. 60 , 673–691 (2010).
ADS Статья Google Scholar
Пиерини, С. Низкочастотная изменчивость, когерентный резонанс и фазовая селекция в модели низшего порядка ветровой циркуляции океана. J. Phys. Oceanogr. 41 , 1585–1604 (2011).
ADS Статья Google Scholar

Пиерини, С. Стохастические переломные моменты в динамике климата. Phys. Ред. E 85 , 027101 (2012).
ADS Статья CAS Google Scholar
Пиерини, С. Ансамблевое моделирование и обратные аттракторы периодически вынужденной системы двойного круговорота. J. Phys. Oceanogr. 44 , 3245–3254 (2014).
ADS Статья Google Scholar
Пиерини, С., Гил, М. и Чекроун, М. Д. Исследование обратных аттракторов квазигеострофической модели океана низкого порядка: детерминированный случай. J. Clim. 29 , 4185–4202 (2016).
ADS Статья Google Scholar
Пиерини, С., Чекроун, М. и Гил, М. Возникновение хаоса в неавтономных диссипативных динамических системах: исследование на примере модели океана низкого порядка. Нелинейные процессы в геофизике. 25 , 671–692 (2018).
25 , 671–692 (2018).
ADS Статья Google Scholar
Пиерини, С. Статистическая значимость небольших ансамблей моделирования и обнаружения внутренней изменчивости климата: тематическое исследование возбудимой океанической системы. J. Stat. Phys. 179 , 1475–1495 (2020).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Гил М. Циркуляция океана, приводимая в движение ветром: применение теории динамических систем к проблеме климата. Discrete Cont. Дин. Syst. А 37 , 189–228 (2017).
MathSciNet МАТЕМАТИКА Статья Google Scholar
Пиерини, С., Дейкстра, Х. А. и Му, М. Внутренняя низкочастотная изменчивость и предсказуемость течения Куросио и его распространения. Adv. Oceanogr. Лимнол. 5 , 79–122 (2014).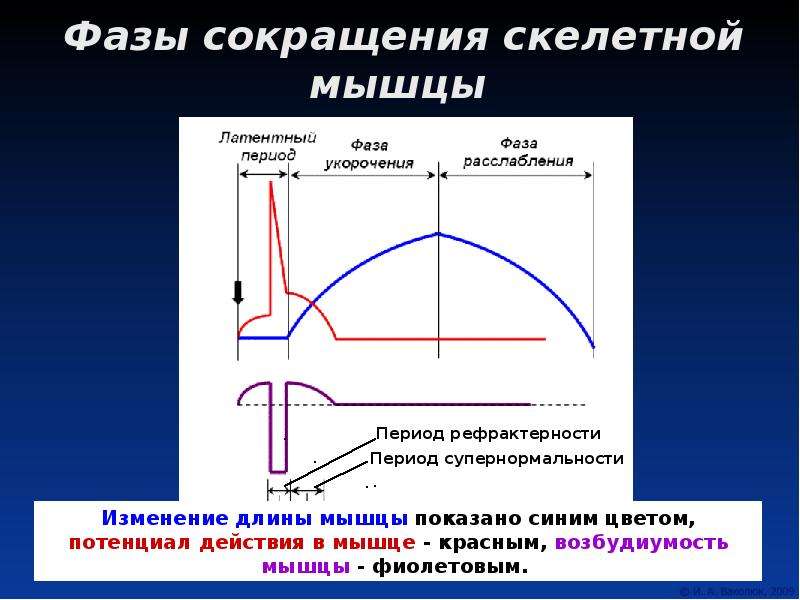
Артикул Google Scholar
Вексельбергер, М. Канардс. Scholarpedia 2 , 1356. https://doi.org/10.4249/scholarpedia.1356 (2007).
ADS Статья Google Scholar
Сутера, А. Стохастическое возмущение чисто конвективного движения. J. Atmos. Sci. 37 , 245–249 (1980).
ADS MathSciNet Статья Google Scholar
Гил, М. Проблемы Гильберта для наук о Земле в 21 веке. Нелинейные процессы в геофизике. 8 , 211–222 (2001).
ADS Статья Google Scholar
McWilliams, J. C. Непреодолимая неточность моделирования атмосферы и океана. Proc. Nat. Акад. Sci. 104 , 8709–8713 (2007).
ADS CAS PubMed Статья Google Scholar

Карри, Дж. А. и Вебстер, П. Дж. Климатология и монстр неопределенности. Бык. Являюсь. Meteorol. Soc. 92 , 1667–1682 (2011).
ADS Статья Google Scholar
De Saedeleer, B., Crucifix, M. & Wieczorek, S. Является ли астрономическая сила надежным и уникальным кардиостимулятором для климата? Концептуальная модель исследования. Клим. Дин. 40 , 273–294 (2013).
Артикул Google Scholar
Распятие, М. Почему ледниковые периоды могут быть непредсказуемыми ?. Клим. Прошлое 9 , 2253–2267 (2013).
Артикул Google Scholar
Боккалетти, С., Куртс, Дж., Осипов, Г., Валладарес, Д. Л., Чжоу, С. С. Синхронизация хаотических систем. Phys. Отчет 366 , 1–101 (2002).
ADS MathSciNet CAS МАТЕМАТИКА Статья Google Scholar

Пиковский А., Розенблюм М. и Куртс Дж. Синхронизация: универсальная концепция нелинейной науки (Cambridge University Press, 2003).
MATH Книга Google Scholar
Грот А. и Гил М. Анализ многомерного сингулярного спектра и путь к фазовой синхронизации. Phys. Ред. E 84 , 036206. https://doi.org/10.1103/PhysRevE.84.036206 (2011).
ADS CAS Статья Google Scholar
Feudel, U. & Grebogi, C. Мультистабильность и контроль сложности. Хаос междисциплинарный. J. Нелинейные науки. 7 , 597–604 (1997).
MathSciNet CAS МАТЕМАТИКА Статья Google Scholar
Чекроун, М. Д., Нилин, Дж. Д., Кондрашов, Д., Мак-Вильямс, Дж. К. и Гил, М. Грубая зависимость параметров в климатических моделях и роль резонансов Рюэля – Полликотта. Proc. Nat. Акад. Sci. 111 , 1684–1690. https://doi.org/10.1073/pnas.1321816111 (2014).
Nat. Акад. Sci. 111 , 1684–1690. https://doi.org/10.1073/pnas.1321816111 (2014).
ADS CAS Статья PubMed Google Scholar
Джин, Ф.-Ф., Нилин, Дж. Д. и Гил, М. Эл. Ниньо на лестнице дьявола: ежегодные субгармонические шаги к хаосу. Science 264 , 70–72 (1994).
ADS CAS PubMed Статья Google Scholar
Джин, Ф.-Ф., Нилин, Дж. Д. и Гил, М. Эл. Ниньо / Южное колебание и годовой цикл: субгармоническая синхронизация частоты и апериодичность. Physica D 98 , 442–465 (1996).
ADS МАТЕМАТИКА Статья Google Scholar
Collet, P. & Eckmann, J.-P. Концепции и результаты в хаотической динамике: краткий курс (Springer, 2007).
MATH Google Scholar

Lasota, A. & Mackey, M.C. Хаос, фракталы и шум: стохастические аспекты динамики Vol. 97 (Springer, 2013).
MATH Google Scholar
Гил М., Чекроун М. Д. и Симоннет Э. Климатическая динамика и механика жидкости: естественная изменчивость и связанные с этим неопределенности. Physica D 237 , 2111–2126 (2008).
ADS MathSciNet МАТЕМАТИКА Статья Google Scholar
Маранджо, Л., Седро, Дж., Галатоло, С., Гарбо, А. Д. и Гил, М. Карты Арнольда с шумом: дифференцируемость и немонотонность числа вращения. Дж.Стат. Phys. https://doi.org/10.1007/s10955-019-02421-1 (2019).
Артикул МАТЕМАТИКА Google Scholar
Бхаттачарья, К., Гил, М. и Вулис, И. Л. Внутренняя изменчивость модели энергетического баланса с отложенными эффектами альбедо. J. Atmos. Sci. 39 , 1747–1773 (1982).
Atmos. Sci. 39 , 1747–1773 (1982).
ADS Статья Google Scholar
Гил, М., Чекроун, М. Д. и Степан, Г. Сборник по динамике климата: множественные масштабы и эффекты памяти, введение. R. Soc. Proc. A 471 , 20150097. https://doi.org/10.1098/rspa.2015.0097 (2015).
ADS Статья Google Scholar
Кондрашов Д., Чекроун М. Д. и Гил М. Управляемые данными немарковские модели замыкания. Physica D 297 , 33–55. https://doi.org/10.1016 / j.physd.2014.12.005 (2015).
ADS MathSciNet Статья МАТЕМАТИКА Google Scholar
Мухин Д. и др. Прогнозирование критических переходов в моделях ЭНСО. Часть I: методология и простые модели с памятью. J. Clim. 28 , 1940–1961. https://doi.org/10.1175/JCLI-D-14-00239.1 (2015a).
ADS Статья Google Scholar
Мухин, Д. и др. Прогнозирование критических переходов в моделях ЭНСО. Часть II: пространственно-зависимые модели. J. Clim. 28 , 1962–1976. https://doi.org/10.1175/JCLI-D-14-00240.1 (2015b).
ADS Статья Google Scholar
Ливина В. Н., Квасниок Ф., Ломанн Г., Кантельхардт Дж. У. и Лентон Т. М. Изменение состояния и стабильности климата: от плиоцена до настоящего времени. Клим.Дин. 37 , 2437–2453 (2011).
Дейкстра, Х.А. Характеристика режима множественного равновесия в модели глобального океана. Tellus A 59 , 695–705 (2007).
ADS Статья Google Scholar
Является ли частота соматосенсорной электростимуляции ключевым параметром в модуляции кортикоспинальной возбудимости здоровых добровольцев и пациентов с инсультом со спастичностью?
Соматосенсорная электрическая стимуляция (СЭС) была предложена в качестве метода лечения пациентов с сенсорно-моторными нарушениями, такими как спастичность. Однако до сих пор нет единого мнения относительно того, какие параметры SES будут адекватными для лечения этих дефицитов. Таким образом, целью данного исследования было оценить эффекты применения SES на мышцы предплечья при четырех различных частотах стимуляции (3, 30, 150 и 300 Гц) и в двух интервалах времени (5 ‘и 30’) с помощью средства транскраниальной магнитной стимуляции и рефлекса Гофмана (H-рефлекс) у здоровых добровольцев (Эксперименты I и II). Группа пациентов с инсультом (эксперимент III) также была предварительно оценена, чтобы установить эффекты SES при низкой частоте (3 Гц), применяемой на 30 минут к мышцам спастических сгибателей предплечья, путем измерения пассивного крутящего момента лучезапястного сустава.Моторные вызванные потенциалы и H-рефлекс были собраны из различных мышц предплечья и кисти непосредственно до и после SES и до 5 ‘(Эксперимент I) и 10’ (Эксперименты I и II) позже. Ни одна из исследованных частот SES не могла работать как ключ при переключении модулирующих эффектов в центральной нервной системе здоровых добровольцев и пациентов с инсультом со спастичностью.
Однако до сих пор нет единого мнения относительно того, какие параметры SES будут адекватными для лечения этих дефицитов. Таким образом, целью данного исследования было оценить эффекты применения SES на мышцы предплечья при четырех различных частотах стимуляции (3, 30, 150 и 300 Гц) и в двух интервалах времени (5 ‘и 30’) с помощью средства транскраниальной магнитной стимуляции и рефлекса Гофмана (H-рефлекс) у здоровых добровольцев (Эксперименты I и II). Группа пациентов с инсультом (эксперимент III) также была предварительно оценена, чтобы установить эффекты SES при низкой частоте (3 Гц), применяемой на 30 минут к мышцам спастических сгибателей предплечья, путем измерения пассивного крутящего момента лучезапястного сустава.Моторные вызванные потенциалы и H-рефлекс были собраны из различных мышц предплечья и кисти непосредственно до и после SES и до 5 ‘(Эксперимент I) и 10’ (Эксперименты I и II) позже. Ни одна из исследованных частот SES не могла работать как ключ при переключении модулирующих эффектов в центральной нервной системе здоровых добровольцев и пациентов с инсультом со спастичностью.
1. Введение
В последние годы для лечения применялся подход, известный как соматосенсорная электрическая стимуляция (СЭС), заключающийся в применении периферической электростимуляции ниже или на уровне моторного порога (МП) [1–3]. лечат пациентов с сенсомоторными нарушениями, такими как спастичность [4–8].Несмотря на обнадеживающие клинические результаты, наблюдаемые у пациентов с инсультом [3, 9] и поражением спинного мозга [10, 11], некоторые методологические аспекты терапевтического использования СЭС остаются нерешенными [12]. Chipchase et al. [13] и Велдман и др. [14] провели систематические обзоры параметров СЭС на первичной моторной коре (M1) и предположили, что данных для определения их эффектов недостаточно. Вдобавок, похоже, также отсутствует консенсус относительно влияния SES на спинномозговые цепи, в основном из-за методологических ограничений, которые более подробно описаны в другом месте [15].
Среди механизмов, лежащих в основе СЭС, Уорд [16] предположил, что увеличение частоты стимуляции стимулируемого нерва позволит ионному току протекать более глубоко, что приведет к привлечению большего пула соматосенсорных рецепторов из разных тканей, прилегающих к сайт стимуляции СЭС.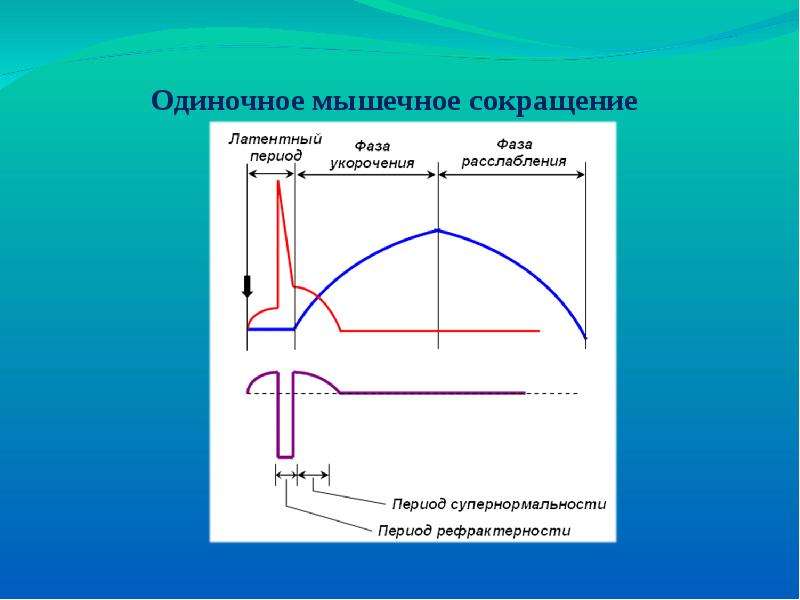 Таким образом, можно ожидать отчетливого воздействия на возбудимость в M1 и / или спинномозговой цепи в зависимости от среднего выхода задействованных сенсорных волокон. Однако, насколько нам известно, в предыдущих исследованиях сравнивалось влияние более чем двух частот SES на кортикоспинальную и спинномозговую возбудимость.
Таким образом, можно ожидать отчетливого воздействия на возбудимость в M1 и / или спинномозговой цепи в зависимости от среднего выхода задействованных сенсорных волокон. Однако, насколько нам известно, в предыдущих исследованиях сравнивалось влияние более чем двух частот SES на кортикоспинальную и спинномозговую возбудимость.
Таким образом, целью данного исследования было оценить с помощью транскраниальной магнитной стимуляции (ТМС) и рефлекса Гофмана (Н-рефлекс) у нормальных добровольцев эффект применения СЭС на мышцы кисти и предплечья при четырех различных частотах стимуляции, проведенных в двух опытах. Кроме того, группа пациентов, перенесших инсульт, также была предварительно оценена, чтобы установить эффекты SES с низкой частотой в мышцах спастических сгибателях предплечья на пассивный крутящий момент лучезапястного сустава.
2.Материалы и методы
2.1. Субъекты
Первые два эксперимента были выполнены с двумя группами добровольцев-правшей без неврологических расстройств. Первый состоял из четырнадцати добровольцев (семь мужчин и семь женщин; возраст: 19,0–39,0 лет, рост: 1,54–1,92 м, масса тела: 53,3–97,0 кг; Эдинбургская инвентаризация рук [17]: + 30,0– + 90,0) и во вторую — шесть добровольцев (пять мужчин и одна женщина; возраст: 20,0–37,0 лет, рост: 1,54–1,92 м, масса тела: 56 лет.0–97,0 кг; Эдинбургский опросный лист [17]: + 40,0– + 95,0). Критерии включения и исключения были основаны на Rossi et al. [18]. Первую и вторую группы подвергали 5 ‘и 30’ протоколам SES соответственно, далее именуемые экспериментами I и II. Оба времени подачи заявки были определены произвольно.
Первый состоял из четырнадцати добровольцев (семь мужчин и семь женщин; возраст: 19,0–39,0 лет, рост: 1,54–1,92 м, масса тела: 53,3–97,0 кг; Эдинбургская инвентаризация рук [17]: + 30,0– + 90,0) и во вторую — шесть добровольцев (пять мужчин и одна женщина; возраст: 20,0–37,0 лет, рост: 1,54–1,92 м, масса тела: 56 лет.0–97,0 кг; Эдинбургский опросный лист [17]: + 40,0– + 95,0). Критерии включения и исключения были основаны на Rossi et al. [18]. Первую и вторую группы подвергали 5 ‘и 30’ протоколам SES соответственно, далее именуемые экспериментами I и II. Оба времени подачи заявки были определены произвольно.
Третья группа, состоящая из пяти пациентов с хроническим инсультом (одна женщина и четыре мужчины; возраст: 45–70 лет, рост: 1,56–1,70 м, масса тела: 61,0–89,0 кг) со спастичностью, все правши (Edinburgh Handedness Опись [17]: до инсульта [+86.67– + 100,0]; после инсульта [-100,0– + 23,33]), также был привлечен к пилотному исследованию, касающемуся воздействия SES на спастические мышцы (Эксперимент III). Среди принятых критериев включения [18] основным было наличие спастичности в правом предплечье, ограничивающее движения запястья, с максимальной оценкой «2» («Более заметное повышение мышечного тонуса в большинстве диапазонов движений (ROM). , но затронутые части легко перемещаются ») на основе модифицированной шкалы Ашворта [19]. Кроме того, когнитивные способности и кожная чувствительность должны быть нормальными, что оценивалось с помощью тестов Mini Mental State Examination [20] и Pain and Light Touch Sensation [21], соответственно.Все пациенты с инсультом были независимо обследованы двумя опытными физиотерапевтами. Индивидуальные характеристики всех пациентов, перенесших инсульт, представлены в таблице 1.
Среди принятых критериев включения [18] основным было наличие спастичности в правом предплечье, ограничивающее движения запястья, с максимальной оценкой «2» («Более заметное повышение мышечного тонуса в большинстве диапазонов движений (ROM). , но затронутые части легко перемещаются ») на основе модифицированной шкалы Ашворта [19]. Кроме того, когнитивные способности и кожная чувствительность должны быть нормальными, что оценивалось с помощью тестов Mini Mental State Examination [20] и Pain and Light Touch Sensation [21], соответственно.Все пациенты с инсультом были независимо обследованы двумя опытными физиотерапевтами. Индивидуальные характеристики всех пациентов, перенесших инсульт, представлены в таблице 1.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Полные протоколы экспериментов были представлены в местный этический комитет (номер процесса: 082/08) и проводились после того, как каждый доброволец дал информированное согласие.
2.2. Планы экспериментов
Первую и вторую группы (эксперименты I и II) подвергали воздействию четырех различных частот SES: 3, 30, 150 и 300 Гц. Частоты были выбраны на основе предыдущих отчетов, в которых они приводили к улучшению моторного контроля и / или нейромодуляции [2, 22–24] за счет использования SES и других терапевтических методов электростимуляции.В обоих экспериментах частоты стимуляции применялись случайным образом. В эксперименте I влияние четырех частот SES на кортикоспинальную и спинномозговую возбудимость оценивали в одном и том же экспериментальном сеансе. Каждую частоту SES применяли в течение 5 минут, после чего следовали 10 минут отдыха. Моторные вызванные потенциалы (MEP) и H-рефлекс собирали непосредственно перед (исходный уровень) и после (0 ‘) применения SES, а затем через 5 минут (5’). Напротив, в эксперименте II добровольцы подвергались 30 ‘SES для каждой частоты, и протокол проводился в четыре разных дня, разделенных минимальным интервалом в 72 часа.Как и в эксперименте I, MEP и H-рефлекс собирали непосредственно перед (исходный уровень) и после (0 ‘) применения SES, а затем через 5’ (5 ‘) и 10’ (10 ‘) позже. Каждая экспериментальная сессия длилась примерно 4 часа.
В эксперименте III пациенты с инсультом проходили терапию SES только с частотой 3 Гц, которая также применялась в течение 30 минут за один сеанс, аналогично эксперименту II. Мы оправдываем такое решение из-за затрат времени (~ 4 ч) на эксперименты I и II, а также потому, что предыдущие авторы [14] сообщают об интересных модуляционных эффектах M1, когда SES применялась ниже 10 Гц.Кроме того, пассивное механическое сопротивление правого лучезапястного сустава при разгибании оценивалось с помощью изокинетической системы [25] только до (исходный уровень) и сразу после (0 ‘) терапии SES.
2.3. Приложение SES
Импульс SES представлял собой сигнал тока постоянной амплитуды (несимметричный асимметричный двухфазный импульс) длительностью 500 мкм с [26, 27]. СЭС проводилась с использованием специального электростимулятора (FES-PEB), созданного Веллозо и Соуза [27].Интенсивность пульса была установлена ниже МТ и определялась на основании отчетов добровольцев об ощущении покалывания в стимулируемой области (мышцы сгибателя предплечья) без какой-либо боли или видимого движения запястья и пальцев. Для SES использовали поверхностные самоклеящиеся электроды (5 × 5 см; модель: CF5050, Axelgaard Manufacturing Co., Ltd., Дания). Интенсивность SES была установлена в диапазоне 2–7 мА (эксперимент I), 3–8 мА (эксперимент II) и 3-5 мА (эксперимент III). Поверхностные электроды располагали над правым предплечьем, между суставами запястья (отрицательный электрод) и локтевым (положительный электрод) суставами, параллельно мышце FCR в продольном направлении (рис. 1).
Поскольку было высказано предположение, что вращение предплечья вокруг его продольной оси может модулировать спинномозговые рефлексы [28], добровольцы оставались сидящими в удобном кресле, а правое предплечье оставалось в положении лежа на протяжении всего эксперимента.
2.4. TMS и сбор данных H-Reflex
Система BIOPAC (модель: MP150; аналого-цифровой преобразователь: 16 бит; динамический диапазон: ± 10 В; частота дискретизации: 15 кГц; полосовой фильтр: 4-го порядка и 100–5000 Гц; прирост: 2000 г .; BIOPAC Systems, Inc., США) использовался для сбора МВП и H-рефлекса. Поверхностные многоразовые электроды BIOPAC (Ag / AgCl; диаметр: 8 мм) помещали в соответствии с рекомендациями SENIAM [29] на следующие мышцы живота: сгибатель (FCR) и разгибатель (ECR) carpi radialis и abductor pollicis brevis (APB). Мышцы ECR и APB были определены как контрольные в экспериментах I и II. В эксперименте III мы наблюдали еще за одной мышцей: контрлатеральным сгибателем лучевого сгибателя запястья (FCRc) непораженного левого предплечья.Поэтому в этом конкретном эксперименте мышцы FCRc, ECR и APB были определены как контрольные.
Катушка TMS Butterfly (модель: MagPro; MagVenture, Дания) была расположена над левым M1 в оптимальном положении кожи головы ( горячая точка ), чтобы вызвать двигательные реакции FCR в контралатеральном полушарии. Положение горячей точки было достигнуто с помощью колпачка с отметками (сетка: 1 × 1 см) в соответствии с международной системой 10-20 для электроэнцефалографии. Горячая точка FCR была определена как участок в M1, в котором одиночный магнитный импульс, установленный на минимальную интенсивность, вызывал ответ MEP (амплитуды более 50 мк В в трех из шести испытаний) в расслабленной мышце. [30–32].Затем интенсивность магнитного стимула была отрегулирована на 20% выше FCR покоя MT.
Как упоминалось ранее, MEP были собраны из всех мышц, исследованных непосредственно перед (исходный уровень) и сразу после (0 ‘) SES и до 5’ (Эксперимент I) и 10 ‘(Эксперименты II и III) позже после каждого сеанса SES и для каждой частоты. В каждом измерении подавалось не менее шести (эксперимент I) и десяти (эксперименты II и III) одиночных импульсов TMS с интервалом между импульсами 5–10 с.Сбор данных поверхностной электромиографии (пЭМГ) запускался аппаратным импульсом, подаваемым системой ТМС. Изменения в возбудимости позвоночника отслеживали посредством оценки среднего значения между максимальным и минимальным FCR H-рефлексными ответами, полученными с помощью кривой набора, собранного перед началом каждого сеанса SES. Чтобы вызвать H-рефлекс от FCR, прямоугольный монофазный импульс 800, 90 · 104 μ, 90 · 105 с подавался на пару поверхностных электродов, причем один из электродов помещался над срединным нервом (катод: Ag / AgCl; диаметр 4 мм. ), На ~ 4 см выше локтевого сустава, а второй на противоположной конечности (анод: Ag / AgCl; 47.5 см 2 ). Сбор сигнала пЭМГ запускался аппаратным импульсным триггером, обеспечиваемым стимулятором. После подачи электрического импульса было собрано и сохранено окно данных длительностью 100 мс. Межимпульсные интервалы варьировались от 5 до 10 с [33]. По крайней мере, четыре окна данных сигнала пЭМГ были получены для дальнейшего анализа из-за надежности собранных данных H-рефлекса. Что касается H-рефлекса FCR, эксперимент I предоставил подходящие данные только от семи добровольцев (один мужчина и шесть женщин; возраст: 19 лет.0–31,0 года; высота: 1,55–1,73 м; масса тела: 53,3–82,6 кг; Эдинбургская инвентаризация ручного труда [17]: + 60,0– + 90,0) из-за методологических ограничений при его регистрации. Точно так же, также из-за методологических ограничений, можно было собрать H-рефлекс FCR только у двух пациентов, перенесших инсульт, и поэтому мы решили не представлять эти данные в этом исследовании.
Визуальная обратная связь сигналов пЭМГ обеспечивалась на протяжении всех экспериментальных сессий, чтобы подтвердить, что все добровольцы были расслаблены.
2,5. Пассивное измерение крутящего момента запястья
Специальное устройство было создано для оценки пассивного сопротивления лучезапястного сустава при движении (рис. 2). Он состоял из шагового двигателя со статическим крутящим моментом 100 кгс · см, управляемого микрошаговым приводом, который делал небольшие шаги 0,036 °, которые сглаживали движение. Присоединенный к шаговому двигателю датчик нагрузки позволял измерять крутящий момент, прилагаемый к запястью. Данные были записаны с использованием системы Spider 8 (HBM, Hottinger Baldwin Messtechnik) с 16 битами, частотой дискретизации 4800 Гц и фильтром сглаживания, установленным на 960 Гц.
Среднеквадратичное значение силы, измеренной датчиком нагрузки, было рассчитано и использовано в качестве оценки крутящего момента (гс · см) в течение 5 циклов разгибания запястья на 35 градусов при угловой скорости 10 градусов в секунду, чтобы избежать рефлекса растяжения [34].
2.6. Данные и статистический анализ
Поверхностные ЭМГ-сигналы, собранные от каждой мышцы во время экспериментов I, II и III и в каждый интервал времени (исходный уровень, 0 ‘и 5’ или 10 ‘), анализировали с использованием алгоритма, встроенного в Matlab 6.5 (Mathworks, США). Этот алгоритм измерял значения MEP и H-рефлекса от пика до пика (P-P), которые представляют кортикоспинальную [35] и спинальную [36] возбудимость. Затем были получены средние значения MEP и H-рефлекса из каждых шести (Эксперимент I) и десяти (Эксперименты II и III) окон данных сигналов sEMG, полученных от мышц, ранее упомянутых для каждой группы, что дало одно значение для каждого интервала времени, что до (исходный уровень) и после (0 ‘) применения SES, а также через 5’ (Эксперимент I) и 10 ‘(Эксперименты II и III) позже для каждой частоты SES и для каждого добровольца.Затем медианы значений и H- были нормализованы путем вычисления отношения между данными для каждого интервала времени (0 ′, 5 ′ и 10 ′) и соответствующим базовым значением, достигнутым до SES, умноженным на 100 [2, 37–39 ].
Все нормализованные значения данных MEP были проанализированы с использованием двустороннего дисперсионного анализа с повторными измерениями (факторы: интервал времени × частота ) для каждой мышцы индивидуально. Для данных H-рефлекса использовали двухфакторный дисперсионный анализ с повторными измерениями (факторы: , интервал времени × , частота ), поскольку все измерения были взяты только для мышцы FCR.Для эксперимента III (TMS и H-рефлекс) анализ выполняли с использованием двухстороннего дисперсионного анализа с повторными измерениями (факторы: , интервал времени × мышцы ), в то время как изокинетические данные оценивали с помощью парного теста Стьюдента. Уровень значимости ( α ) был установлен на уровне 5%. Статистический анализ проводился с использованием R, версия 3.1. Результаты представлены в виде средних значений и стандартных отклонений.
3. Результаты
3.1. Эксперимент I
На рисунке 3 показаны результаты, полученные для нормализованных значений H и FCR H, соответственно, сразу после (0 ‘) SES и до 5’ позже для четырех частот SES, применяемых в течение 5 минут и для трех мышц (FCR , ECR и APB).Результаты, полученные после установки SES на всех исследованных частотах, не выявили какой-либо значимой разницы между частотами, () и интервалами времени () в данных по сравнению с исходным уровнем для всех мышц (Рисунки 3 (а), 3 (б) и 3 (в)).
Что касается нормализованных значений FCR H- (рис. 3 (d)), он также не показал какой-либо значимой разницы между частотами () и интервалами времени () от базовой линии.
3.2. Эксперимент II
На рисунке 4 показаны результаты, полученные для нормализованных значений H и FCR H, соответственно, сразу после (0 ‘) SES и до 5’ и 10 ‘позже для четырех частот SES, применяемых в течение 30 минут и для три мышцы (FCR, ECR и APB). Результаты, полученные после установки SES на всех исследованных частотах, не выявили существенной разницы между частотами для мышц FCR () и APB (). Хотя между частотами для ECR () была значительная разница, они не отличались статистически от исходного уровня ().Как и в эксперименте I, также не было значительной разницы между интервалами времени () в данных от базовой линии для всех мышц (рисунки 4 (a), 4 (b) и 4 (c)).
Наши результаты также не показали какой-либо значимой разницы между частотами (), интервалами времени () и взаимодействием интервала времени × частоты () относительно нормализованных значений FCR H- от базового уровня.
3.3. Эксперимент III
На рисунках 5 и 6 показаны результаты, полученные для нормализованных значений и задержки, соответственно, для SES, установленного на 3 Гц и в течение 30 минут.Не было значительных различий между интервалами времени в нормализованных относительных значениях от исходного уровня для всех трех мышц, то есть FCR (), FCRc (), ECR () и APB (). Измерения задержки также не показали какой-либо существенной разницы () относительно исходных значений.
Сопротивление пассивному разгибанию (Рисунок 7 (a);) и сгибанию (Рисунок 7 (b);) движениям лучезапястного сустава, оцененное с помощью изокинетической системы, также не показало какой-либо существенной разницы при сравнении измерения, собранные сразу после (0 ‘) (Post) SES и базовой линии (Pre).
4. Обсуждение
Было высказано предположение, что использование SES в клинической области может обеспечить улучшения, аналогичные тем, которые достигаются при интенсивном обучении [40]. Например, Conforto et al. [41] наблюдали повышение уровня силы щипковых движений у пациентов с инсультом после двух часов SES. Dos Santos-Fontes et al. [9] также предположили, что СЭС может привести к долгосрочному улучшению работы паретичной руки у пациентов с хроническим инсультом. Кроме того, SES был предложен в качестве альтернативного подхода к минимизации спастичности [6, 7, 42], хотя также нет ясности относительно его основных механизмов.Следовательно, несмотря на вышеупомянутые примеры, касающиеся положительных эффектов SES в клинических условиях, до сих пор нет единого мнения относительно того, как некоторые из параметров, используемых в SES (например, время применения, частота, интенсивность, форма волны и ширина импульса), являются учитывается центральной нервной системой (ЦНС) [14, 43]. Поэтому мы решили оценить влияние различных частот СЭС на спинномозговую и кортикоспинальную возбудимость здоровых добровольцев. Кроме того, мы также провели пилотное исследование для оценки эффекта SES, применяемого в течение 30 минут, в соответствии с клинической практикой, и выбрали частоту 3 Гц, применяемую к спастическим мышцам сгибателей предплечья у пяти пациентов, перенесших инсульт, с помощью TMS и пассивной терапии. оценка движений запястья.
Тем не менее, что говорит нам литература о различных частотах СЭС при кортикоспинальной модуляции и спастичности? Было показано, что возбуждающие, а также ингибирующие эффекты в M1 сохраняются от нескольких минут до часов после терапии SES [4, 13, 37, 44, 45]. Хотя некоторые авторы предположили, что увеличение частоты стимуляции может преимущественно приводить к увеличению возбудимости M1 [13, 22, 38, 46, 47], другие сообщили об обратном эффекте на уровне позвоночника [48].Мы предположили, что увеличение частоты позволит ионному току проходить более глубоко по всей протяженности предплечья и вокруг срединного нерва, который иннервирует кожу ладонной стороны большого пальца, указательного, среднего пальца и мышцы APB. [49]. В результате мы ожидали вызвать изменения кортикоспинальной возбудимости FCR и APB мышц. Тем не менее, мы также рассмотрели гипотезу о наблюдении некоторого эффекта в ECR-мышце, учитывая, что афферентные волокна типа II (неадаптирующиеся сенсорные волокна) в мышцах способны облегчать или подавлять мышцы-антагонисты [50], хотя Пьеро-Дезейлиньи и Берк [36] утверждают, что что влияние этого взаимодействия на верхнюю конечность до конца не изучено.
Тем не менее, в настоящем исследовании как короткая (5 ‘), так и большая (30’) длительность SES, применяемая с разной частотой, не приводила к изменениям спинномозговой и кортикоспинальной возбудимости мышцы при стимуляции, а также мышц, которые разделяют какие-либо кинезиологическое свойство со стимулированным. Несмотря на обескураживающие результаты, помимо частоты стимуляции необходимо учитывать некоторые вопросы, которые обсуждаются ниже.
4.1. Ширина импульса
Мы использовали более длительную длительность импульса (500 мкм с), чем у других авторов (от 100 до 300 мкм с) [2, 6, 22], которые более избирательны [26].Мы выбрали эту ширину импульса из-за необходимости задействовать широкий диапазон диаметров и модальностей сенсорных афферентных волокон. Поскольку нашей основной целью было оценить влияние различных частот SES, повышение избирательности рекрутирования могло замаскировать возможный источник модуляции кортикоспинальной возбудимости, полученный только на основании этого параметра.
Подобные отрицательные и расходящиеся с нашим результаты результаты также были описаны в литературе. Например, Tinazzi et al. [2] наблюдали увеличение и уменьшение кортикоспинальной возбудимости ECR и FCR, соответственно, после применения SES при 150 Гц над мышцами FCR в течение 30 минут.С другой стороны, Фернандес-дель-Ольмо и др. [49] выполнили аналогичный протокол и не наблюдали какого-либо облегчения или торможения обеих мышц. Любопытно, что в обоих исследованиях использовался более узкий импульс (100 90 · 104 мкм 90 · 105 с), который, как ожидается, будет более селективным для волокон большего диаметра [16].
Ожидается, что более короткая длительность импульса обеспечит более высокую избирательность при различении соматосенсорных, моторных и болевых сенсорных нервов. Ширина импульса 500 мкм с может привести к большему суммированию ответов от рецепторов, распределенных близко к месту SES, которые будут интегрированы и обработаны на разных уровнях ЦНС и, вероятно, переданы на M1 как «усредненный» сигнал. .Таким образом, в качестве первой гипотезы, объясняющей отсутствие модуляции SES, мы предполагаем, что этот усредненный входной сигнал может быть не в состоянии вызвать спинальную или кортикоспинальную модуляцию.
4.2. Интенсивность стимуляции SES
Другой важный вопрос касается интенсивности стимуляции. В некоторых исследованиях сообщается о различных эффектах SES, установленных на разных уровнях интенсивности на кортикоспинальную возбудимость, но также включая протоколы парной ассоциированной стимуляции (PAS). В этом контексте Pitcher et al.[22] применяли электрическую стимуляцию выше МТ и устанавливали на 3 Гц на первую спинную межкостную мышцу (FDI) в течение 30 минут, в то время как импульсы ТМС были синхронизированы по FDI hot — пятно в протоколе PAS. Они наблюдали уменьшение примерно на 40–50 ‘, предполагая, что периферическая электрическая стимуляция на такой низкой частоте опосредует задействование нервных цепей, которые вызывают долговременную депрессию (LTD). С другой стороны, Aimonetti и Nielsen [51] показали, что применение кондиционирующего электрического стимула чуть ниже MT над срединным нервом, который снабжает FCR, вызывает облегчение ECR на очень короткое время.Напротив, Bertolasi et al. [52] также показали, что применение кондиционирующего электрического стимула чуть выше МТ над срединным нервом вызывает ингибирование ECR. Veldman et al. [14] считают, что уровень кортикоспинальной возбудимости, по-видимому, модулируется тонкой настройкой интенсивности SES, которая может варьироваться от воспринимаемого до моторного порога и объясняется широким диапазоном и направлениями нейронных ответов, полученных от MEPs при TMS. эксперименты. Более того, их результаты предполагают, что интенсивность стимуляции, установленная на пороге восприятия, не способна вызвать какую-либо модуляцию кортикоспинальной возбудимости.Основываясь на этих замечаниях и на методологическом подходе, принятом в настоящем исследовании для настройки интенсивности SES (см. Подробности выше в разделе 2), в качестве второй гипотезы мы предполагаем, что обеспечивается на уровне или очень близко к порогу восприятия для большинства добровольцев. , SES не вызвала какого-либо измеримого физиологического эффекта для всех протестированных мышц, частот и интервалов времени.
4.3. Поддержание интенсивности SES
Другой важный аспект касается поддержания интенсивности стимуляции.Разные авторы регулируют интенсивность СЭС во время своих экспериментов, чтобы избежать привыкания [3, 41, 53, 54]. В отличие от большинства авторов, мы решили сохранить ту же интенсивность стимуляции, чтобы избежать смещения при оценке кортикоспинальной модуляции после SES. Кроме того, мы оценили два разных времени применения SES: очень короткое (5 минут) и долгое (30 минут). Следовательно, механизмы привыкания необходимо учитывать даже в кратковременном протоколе СЭС. В соответствии с этой третьей гипотезой работы Добкина [55] и Димитриевича и др.[56] поддержали идею о том, что регулярный и постоянный паттерн SES может очень быстро привести либо к привыканию / аккомодации, либо к неспособности произвести какое-либо изменение возбудимости M1, хотя этот процесс не кажется жестким. Таким образом, мы предлагаем в качестве третьей гипотезы, что эффект привыкания, вызванный поддержанием интенсивности стимуляции, может быть дополнительной переменной, не вызывающей какой-либо спинномозговой или кортикоспинальной модуляции от SES в настоящем исследовании.
4.4. H-рефлексы
P-P EvaluationНесмотря на то, что мы столкнулись с методологическими ограничениями при регистрации H-рефлекса у некоторых здоровых добровольцев, SES, похоже, не смог вызвать изменения возбудимости такого измерения FCR в экспериментах I и II.Несмотря на то, что H-рефлекс обычно считается моносинаптическим ответом, он также может модулироваться другими входами, которые сходятся к общим интернейронам и которые могут получать входные сигналы от спинномозговых и супраспинальных источников [7, 15]. Следовательно, SES, по-видимому, стимулирует различные специфические рецепторы, которые могут взаимно нейтрализовать себя и производить смесь возбуждения и торможения [7], что ранее предполагалось как одна из наших гипотез о том, что не наблюдается каких-либо существенных различий в кортикоспинальной возбудимости.
В отличие от показанного здесь отсутствия эффекта SES на H-рефлекс FCR, некоторые исследования [57] показали, что функциональная электрическая стимуляция (FES), которая позволяет задействовать сенсорные нейроны мышечного веретена Ia, способна индуцировать модуляцию H-рефлекса. -рефлекс у неврологических больных и здоровых людей. Таким образом, можно предположить, что афференты Ia, вряд ли рекрутируемые согласно протоколу SES, принятому в этом исследовании, могут быть способны вызывать модуляцию Н-рефлекса FCR. Следовательно, интенсивность стимуляции также может быть «ключевым моментом» в модуляции спинальных рефлексов.
4.5. Пациенты со спастическим инсультом
Целью этого предварительного исследования было первое, насколько нам известно, влияние SES, установленного на низкой частоте (3 Гц), на спастические мышцы-сгибатели предплечья на кортикоспинальном уровне у пациентов с хроническим инсультом. . Кроме того, мы намеревались оценить влияние SES на пассивное механическое сопротивление поврежденного лучезапястного сустава. Хотя мы не наблюдали какой-либо значительной спинальной и кортикоспинальной модуляции в экспериментах I и II, мы сначала предположили, что уменьшение кортикоспинального ответа, обеспечиваемого SES на низкой частоте (3 Гц) [22], могло бы способствовать временному уменьшению таких гиперактивность мышц.Однако, несмотря на отрицательные результаты, разные авторы привели некоторые интересные аргументы и результаты, которые предварительно подтверждают нашу предыдущую гипотезу. Как уже упоминалось, Pitcher et al. [22] наблюдали, что периферическая электрическая стимуляция с низкой частотой приводит к снижению кортикоспинальной возбудимости, хотя гипотезы о вероятных механизмах воздействия этой модели электростимуляции на спастичность все еще отсутствуют. Несмотря на это, Liepert et al.[58] предположили, что внутрикортикальное торможение может быть уменьшено у пациентов с инсультом из-за снижения ГАМКергической (ингибирующей) и / или повышенной глутаматергической (возбуждающей) активности, соответственно. Дисбаланс этих нейротрансмиттеров может способствовать проявлению спастичности супраспинального происхождения, хотя его физиопатология все еще исследуется [59]. Аналогичным образом, длительная (> 30 ‘) СЭС на низких частотах может способствовать увеличению набора ГАМКергических цепей и снижению кортикоспинальной возбудимости.Питчер и др. [22] также подтверждают, что модуляция этих нейронных цепей может быть частотно-зависимой и может быть оптимизирована с помощью стимуляции 3 Гц. Sonde et al. [60] также исследовали SES, установленную на низкой частоте (1,7 Гц) при лечении спастичности у лиц с инсультом, и, несмотря на улучшение двигательного паттерна, они также не наблюдали какого-либо значительного уменьшения спастичности, как оценивали с помощью Шкала Ашворта.
Несмотря на то, что данные о MEP и крутящем моменте запястья не достигли статистически значимых уровней, оценка механического сопротивления с помощью изокинетической системы, то есть более точного и разумного подхода, у четырех из шести пациентов было отмечено относительное снижение (5 .От 5 до 14,3%) при пассивном вращении запястья (сгибание и разгибание), что может рассматриваться как клинически значимое. Важно подчеркнуть, что шкала Эшворта не обеспечивает чувствительности к минимальным вариациям уровня спастичности.
Гипервозбудимость H-рефлекса рассматривается как показатель спастичности [48, 61]. Таким образом, отсутствие изменений в измерениях H-рефлекса не позволяет предполагать, что спинномозговая, а не кортикоспинальная возбудимость является вероятной частью ЦНС, подходящей для эффектов SES у пациентов с инсультом со спастичностью.
В итоге, поскольку мы ожидали наблюдать ковариацию между поведением MEP со снижением сопротивления пассивному разгибанию лучезапястного сустава, мы также можем предположить, что низкая частота SES, установленная на уровне 3 Гц, и порог восприятия могут быть не в состоянии вызвать какую-либо нейромодуляцию у этих пациентов.
5. Заключение
Основываясь на работе Ward [16], мы предположили, что увеличение частоты стимуляции SES позволит ионному току протекать более глубоко и, таким образом, большему пулу соматосенсорных рецепторов из разных тканей, прилегающих к SES будет привлечен.Однако, несмотря на то, что мы должны распознавать небольшие выборки участников в экспериментах II и III, результаты, полученные в наших экспериментах, предполагают, что ни одна из исследованных частот (3, 30, 150 и 300 Гц) SES вместе со всеми другими выбранными параметрами похоже, может действовать как ключ в переключении модулирующих эффектов в ЦНС здоровых добровольцев и пациентов с инсультом со спастичностью.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Выражение признательности
Эта работа была профинансирована Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento científico e Tecnológico (CNPq) и Fundação. Это исследование также проводилось в рамках деятельности Центра исследований, распространения и инноваций FAPESP для нейроматематики (грант 2013 / 07699-0, Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)).Сандро Сперандеи получил докторскую стипендию от FAPERJ и CAPES.
Потенциал сложного действия | Кривая «сила-продолжительность» | ||||||||||||||||||||||
Цель этой части лаборатории — изучить взаимозависимость между силой стимула и продолжительностью стимула в активации нерва, и построить кривую «сила-продолжительность». | |||||||||||||||||||||||
| |||||||||||||||||||||||
| Мы видели, как форма, амплитуда и продолжительность изменения ВП по мере увеличения силы стимула, потому что постепенно более сильная стимуляция активирует все больше и больше отдельных нервных волокон, индивидуальные потенциалы действия суммируются, чтобы получить ВП.Таким образом, когда стимул сильнее, большее количество волокон достигает порога. | |||||||||||||||||||||||
| Однако порог активации волокна зависит от не только от силы раздражителя, но и от продолжительность раздражителя. | |||||||||||||||||||||||
| Деполяризация возбудимой мембраны требует прохождения электрического заряда через мембрану.Из-за преобладающая электрическая емкость мембраны, соответствующий параметр для эффективной деполяризации мембраны — общее количество заряда переносится через мембрану. | |||||||||||||||||||||||
| Для кратковременного стимула, генерирующего установившийся трансмембранный ток, переносимый заряд ( Q ) равен пропорционально произведению тока I и времени T : | |||||||||||||||||||||||
Q = I x Т | |||||||||||||||||||||||
Следовательно, если сумма заряда, необходимая для активация волокна Q t , а длительность стимула D , текущая I t для активации потребуется: | |||||||||||||||||||||||
I т = Q т / D | |||||||||||||||||||||||
| Это говорит о том, что график порога сила стимула по сравнению с продолжительностью стимула должна снизиться до почти ноль при увеличении длительности стимула.Другими словами, стимул сила, необходимая для достижения порога, должна уменьшаться в течение более длительная стимуляция. Обратите внимание, что мы можем использовать напряжение (В) и ток (I) взаимозаменяемо как мера силы стимула. | |||||||||||||||||||||||
| Кривая «сила-продолжительность» для типичного нервная мембрана похожа, но отличается тем, что кривая четко выражена сглаживается с большой продолжительностью стимула, достигая асимптоты, называемой РЕБАЗА.Когда сила стимула ниже реобазы, стимуляция неэффективна даже при очень большой продолжительности стимула. | |||||||||||||||||||||||
Несоответствие наблюдаемой формы кривая «сила-продолжительность», предсказываемая приведенным выше уравнением, связана с тем, что что предсказанная взаимосвязь верна для идеальный конденсатор , без сопротивление утечке.При длительной стимуляции (большие значения t) уравнение не может предсказать передачу заряда через нервную мембрану, потому что под ними условиях фактическая передача заряда меньше, чем прогнозировалось, из-за утечки из-за заметное электрическое сопротивление мембраны . Из-за взаимодействия между резистивными и емкостными эффектами в мембране, перенос заряда (и мембрана потенциал) фактически возрастает экспоненциально до плато во время длительной стимуляции, вместо этого возрастает линейно со временем.Таким образом, если раздражитель слишком мал, мембрана потенциал никогда не достигает порога. | |||||||||||||||||||||||
| При исследовании силы-продолжительности связь в нервном стволе, содержащем тысячи нервных волокон, одно нужно быть осторожным, чтобы определить, какое из этих многих волокон является порогом в вопрос относится к. Пороговое напряжение стимула для CAP как целое на самом деле является порогом для самых быстрых и самых возбудимых волокон в нерве.Поскольку точно определить этот порог сложно, описанная ниже процедура использует в качестве опорного сигнала CAP, пик которого амплитуда составляет примерно одну пятую от максимальной. Таким образом, этот порог явно порог для другой менее возбудимой группы волокон. | |||||||||||||||||||||||
| Для начала устанавливается длительность стимула 1.0 мс с помощью ручки на стимуляторе. | |||||||||||||||||||||||
ШАГ A: напряжение стимула медленно увеличивается до появления ВП. В в этом примере мы отрегулировали напряжение пока амплитуда CAP не заполнит два интервала горизонтальной сетки на экране. Эта амплитуда CAP будет будет использоваться в качестве справочного материала для остальной части эксперимента. (Другой справочная линия также может быть выбрана при условии, что она сохраняется на протяжении всего упражнение) | |||||||||||||||||||||||
Считываем напряжение стимула со стимулятора, и соответствующую длительность стимула (в данном случае 1 мс) и введите эту пару значений в стол.Таким образом, длительность этого стимула и напряжение привели к небольшому пороговому значению. группа возбудимых волокон в нерве, хотя они не самые возбудимые. | |||||||||||||||||||||||
| ШАГ B: Теперь потихоньку убавляем длительность стимула до исчезновения ВП. (Останавливаемся, когда на дисплее отображается плоский линия, где раньше была CAP). | |||||||||||||||||||||||
| Затем повторяем ШАГ A; то есть, увеличивайте напряжение стимула до тех пор, пока CAP не достигнет эталонной амплитуды, указанной выше.(В этом случае заполнение двух интервалов сетки). Читаем новое напряжение стимула и длительность стимула от стимулятора и введите пару значений в таблицу. | |||||||||||||||||||||||
| Эта процедура повторяется, пока у нас не будет 10 разные пары значений. По нашим данным построена кривая сила-продолжительность: | |||||||||||||||||||||||
| |||||||||||||||||||||||
| Помимо реобазы, сила-продолжительность
Curve также предоставляет другую информацию — Chronaxie.Chronaxie — это
измерение длительности, соответствующее удвоенному значению Rheobase. Из приведенного выше графика, Rheobase составляет примерно 0,64 В, а Хронакси составляет около 0,16 мс. | |||||||||||||||||||||||
| Вопрос: Что такое
значение Реобазы? A: Обычно около 1 мс кривая сила-продолжительность, кривая выравнивается на реобазе, в точке, где постепенное увеличение длительности импульса больше не связано с прогрессирующим снижение напряжения.Другими словами, для более длительных стимулов минимальная напряжение, необходимое для доведения нерва до порога, будет реобазой. | |||||||||||||||||||||||
| Q: Каково значение
Хронакси? A: Учитывая, что у двух нервов одинаковые Реобаза, хронакси (длительность стимула, соответствующая удвоенной реобазе) может дают представление об их относительной возбудимости.На кривой «сила-продолжительность» справа нерв B более возбудимый. | |||||||||||||||||||||||
| Q: Каким образом
Кривая сила-продолжительность для набора медленных волокон (не очень возбудимых) по сравнению с
Кривая сила-продолжительность для набора быстрых волокон (очень возбудимая)? A: Кривая для более медленных волокна будет смещен вправо, указывая на то, что для данной силы стимула более длительный Длительность стимула потребуется, чтобы довести более медленные волокна до порогового значения. | |||||||||||||||||||||||
Нажмите здесь, чтобы продолжить тему Скорость проводимости (разностный и абсолютный методы) | |||||||||||||||||||||||
Проблема возбудимости | SpringerLink
‘) var cartStepActive = true var buybox = document.querySelector («[id-данных = id _» + отметка времени + «]»). parentNode ; []. slice.call (buybox.querySelectorAll («. покупка-опция»)). forEach (initCollapsibles) функция initCollapsibles (подписка, индекс) { var toggle = subscription.querySelector («. цена-опции-покупки») subscription.classList.remove («расширенный») var form = subscription.querySelector («. Purchase-option-form») if (form && cartStepActive) { var formAction = form.getAttribute («действие») form.setAttribute («действие», formAction.replace («/ checkout», «/ cart»)) document.querySelector («# сценариев электронной торговли»). addEventListener («загрузка», bindModal (форма, formAction, отметка времени, индекс), false) } var priceInfo = subscription.querySelector («. price-info») var buyOption = toggle.parentElement if (переключить && форму && priceInfo) { переключать.setAttribute («роль», «кнопка») toggle.setAttribute («tabindex», «0») toggle.addEventListener («клик», функция (событие) { var extended = toggle.getAttribute («aria-extended») === «true» || ложный toggle.setAttribute («расширенный ария»,! расширенный) form.hidden = расширенный если (! расширено) { покупка вариант.classList.add («расширенный») } еще { buyOption.classList.remove («расширенный») } priceInfo.hidden = расширенный }, ложный) } } function bindModal (form, formAction, timestamp, index) { var weHasBrowserSupport = window.fetch && Array.from return function () { var Buybox = EcommScripts? EcommScripts.Ящик для покупок: null var Modal = EcommScripts? EcommScripts.Modal: null if (weHasBrowserSupport && Buybox && Modal) { var modalID = «ecomm-modal_» + отметка времени + «_» + индекс var modal = новый модальный (modalID) modal.domEl.addEventListener («закрыть», закрыть) function close () { форма.querySelector («кнопка [тип = отправить]»). focus () } form.setAttribute ( «действие», formAction.replace («/ checkout», «/ cart? messageOnly = 1») ) form.addEventListener ( «Отправить», Buybox.interceptFormSubmit ( Buybox.fetchFormAction (window.fetch), Buybox.triggerModalAfterAddToCartSuccess (модальный), console.log, ), ложный ) document.body.appendChild (modal.domEl) } } } function initKeyControls () { документ.addEventListener («нажатие клавиши», функция (событие) { if (document.activeElement.classList.contains («покупка-опция-цена») && (event.code === «Space» || event.code === «Enter»)) { if (document.activeElement) { event.preventDefault () document.activeElement.click () } } }, ложный) } function initialStateOpen () { var buyboxWidth = buybox.offsetWidth ; []. slice.call (buybox.querySelectorAll («. покупка-опция»)). forEach (function (option, index) { var toggle = option.querySelector («. покупка-вариант-цена») var form = option.querySelector («. Purchase-option-form») var priceInfo = option.querySelector («. цена-информация») if (buyboxWidth> 480) { toggle.click () } еще { if (index === 0) { переключать.нажмите () } еще { toggle.setAttribute («расширенная ария», «ложь») form.hidden = «скрытый» priceInfo.hidden = «скрыто» } } }) } initialStateOpen () если (window.buyboxInitialised) вернуть window.buyboxInitialised = true initKeyControls () }) ()Транскраниальная стимуляция постоянным током в сочетании с вниманием повышает возбудимость коры головного мозга и улучшает двигательное обучение у здоровых добровольцев | Журнал нейроинжиниринга и реабилитации
Участники
В этом исследовании приняли участие тридцать три здоровых добровольца, девять (пять женщин) со средним возрастом 25 лет.6 лет (стандартное отклонение: SD, 2,7 года) в эксперименте 1 и 24 (12 женщин) со средним возрастом 24,8 года (SD, 2,4 года) в эксперименте 2. Размер выборки был определен на основе предыдущих исследований, посвященных изучению влияние внимания на моторную корковую пластичность [17, 18]. Доминирующая рука каждого участника была установлена с помощью теста доминантной руки Чепмена [19]. Все участники были правши. Ни один из участников не имел в анамнезе неврологических заболеваний и не принимал лекарства, влияющие на центральную нервную систему.Перед участием участники предоставили письменное информированное согласие. Исследование было одобрено институциональным наблюдательным советом реабилитационной больницы Токийского залива, Япония, и соответствовало стандартам, установленным последней редакцией Хельсинкской декларации. Исследование не было предварительно зарегистрировано, поскольку предварительная регистрация не была распространена в области нейрофизиологии человека в то время, когда проводилось исследование, то есть с 2012 по 2014 год.
Общая экспериментальная процедура
Два эксперимента были проведены в чтобы исследовать комбинированное влияние tDCS и внимания к целевой мышце на моторную кортикальную пластичность (эксперимент 1) и обучение двигательным навыкам (эксперимент 2).Методы каждого эксперимента подробно описаны ниже.
Эксперимент 1 (нейрофизиологический эксперимент)
tDCS
tDCS был доставлен с помощью DC-Stimulator-Plus (NeuroConn, Ильменау, Германия), подключенного к паре электродов с губчатой поверхностью, пропитанных 0,9% физиологическим раствором NaCl. Продолжительность стимуляции составляла 10 мин. Сила тока увеличивалась до 2 мА в течение 15-секундного периода, а в конце периода стимуляции использовалась линейная функция нисходящего тока. Анодный электрод (25 см 2 ) располагался над левым M1.Расположение руки M1 определяли на основании индукции самых больших MEP в правой первой спинной межкостной мышце (FDI), вызванной TMS. Электрод сравнения (50 см 2 ) помещали над ипсилатеральным плечом [20,21,22], чтобы свести к минимуму возможность того, что катодная стимуляция (электрод сравнения) вызывает нежелательные изменения возбудимости лобной коры головного мозга [20, 23]. Плотность тока составляла 0,08 мА / см 2 , а общая плотность поверхностного заряда составляла 0.048 Кл / см 2 , оба значения значительно ниже порога повреждения тканей [24].
Манипулирование вниманием
Участники удобно расположились перед столом в тихой комнате. Их руки при любых условиях были прикрыты коробкой, чтобы не обращать внимания на целевую мышцу. Участников попросили зафиксировать на маркере, расположенном перед ними, на протяжении всего задания.
Чтобы экспериментально манипулировать вниманием участников, их попросили выполнить задачу по обнаружению цели с помощью сенсорного стимула.Во время применения tDCS участникам были предъявлены стимулы двух сенсорных модальностей (то есть соматосенсорные и слуховые стимулы). В качестве соматосенсорного стимула слабый электрический импульс подавался на кожу чуть выше правой мышцы FDI. Длительность импульса составляла 1 мс, а интенсивность стимула в 1,1 раза превышала порог восприятия для каждого участника. В качестве слухового стимула через наушники подавался звуковой сигнал. Интенсивность слухового стимула в 1,1 раза превышала порог восприятия для каждого участника.Оба стимула предъявлялись 20 раз с полуслучайными интервалами каждые 30 с. Участников попросили устно сообщить об обнаружении сенсорного стимула, как только они его заметили. В состоянии соматосенсорного внимания участники улавливали только соматосенсорный раздражитель и должны были игнорировать звуковой раздражитель, тогда как в состоянии слухового внимания задача была обратной. Чтобы обнаружить сенсорный стимул, участникам необходимо было уделять избирательное внимание правой мышце FDI (состояние «Внимание к целевой мышце») или звуковому сигналу (условие «Внимание к звуку»), потому что стимулы были чуть выше их сенсорного порога. и трудно обнаружить без внимания.Отзывы участников не поступали. Ошибочные реакции определялись как пропущенные ответы (отсутствие реакции во время стимуляции) и неправильные ответы (реакция без стимуляции). Все условия стимула и данные ошибочной реакции представлены в дополнительных данных 1.
Электромиография
Перед присоединением электродов участки кожи натирали спиртом, и сопротивление кожи поддерживали ниже 5 кОм. Поверхностные электроды помещали на правый FDI, мышцу, отводящую большой палец (APB), и мышцу лучевого разгибателя запястья (ECR).Необработанный сигнал усиливался и фильтровался (полоса пропускания 5–3000 Гц) с использованием биоэлектрического усилителя (Neuropack MEB-2200; Nihon Kohden Corp., Токио, Япония), оцифровывался с частотой 4000 Гц и сохранялся для автономного анализа на лабораторном компьютере ( Система Power Lab; AD Instruments Pty Ltd., Новый Южный Уэльс, Австралия).
Транскраниальная магнитная стимуляция
ТМС вводили с помощью стимулятора Magstim 200, подключенного через модуль BiStim (Magstim Co., Dyfed, UK) к катушке в форме восьмерки с внутренним диаметром крыла 9 см.Магнитный стимулятор был способен создавать магнитное поле 2,2 Тл для импульса длительностью 100 мкс. Катушку помещали рукояткой назад, латерально под 45 ° от средней линии и примерно перпендикулярно центральной борозде.
Стимулирующую катушку поместили над участком, который был оптимальным для получения ответов в правильных FDI. Пороговое значение было определено в то время, когда ПИИ находились в состоянии покоя и во время добровольного сокращения. Порог определялся как минимальная интенсивность стимула, которая вызывала ответы 50 мкВ с аналогичной формой и задержкой во время пяти из 10 последовательных стимулов.Каждого участника попросили расслабиться во время измерения моторного порога покоя (rMT), в то время как тишина электромиограммы отслеживалась. Активный моторный порог (aMT) был определен как наименьшая интенсивность стимула, необходимая для получения MEP более 200 мкВ, по крайней мере, в пяти из 10 последовательных испытаний во время поддержания 100 мкВ произвольного изометрического сокращения FDI. Хотя параметры были скорректированы для правого FDI (целевой мышцы), APB и ECR были одновременно записаны, чтобы исследовать, наблюдались ли региональные эффекты на моторную кортикальную пластичность, когда участники обращали внимание на целевую мышцу.
Интенсивность стимуляции была установлена на уровне 120% rMT для оценки изменений возбудимости моторной коры. Испытания TMS проводились случайным образом 15 раз, и для каждой временной точки было записано 15 MEP. Размах амплитуды МВП усредняли, и ответы МВП выражали в процентах экспериментальных МВП относительно исходного уровня (% МВП).
Чтобы вызвать интракортикальное торможение с короткими интервалами (SICI) и интракортикальное облегчение (ICF), мы применили подпороговую кондиционирующую стимуляцию парными импульсами [25].Мы использовали 80% aMT для кондиционирующего стимула и 120% rMT для тестового стимула. На протяжении всего эксперимента тестовый стимул регулировался таким образом, чтобы амплитуда MEP оставалась равной амплитуде MEP FDI на исходном уровне. Межстимульные интервалы были установлены равными 2 мс (SICI 2 мс ) и 3 мс (SICI 3 мс ), а также 10 мс (ICF 10 мс ) и 15 мс (ICF 15 мс ), и 15 MEP были записаны из мышца FDI для каждой ISI и тестовой стимуляции. Условные амплитуды МВП выражали в процентах от средних тестовых амплитуд МВП.Время между импульсами стимула варьировалось от 5 до 7 с, чтобы избежать повторяющихся эффектов ТМС. Время стимула автоматически контролировалось с помощью LabVIEW (National Instruments, Остин, Техас, США).
Экспериментальная процедура
В настоящем исследовании использовался рандомизированный перекрестный дизайн с маской оценщика, и все участники выполняли следующие три условия в разные дни: 1) применялась анодная tDCS, в то время как участники обращали внимание на целевую мышцу FDI (анодальная tDCS + внимание к целевой мышце), 2) анодная tDCS применялась, пока участники обращали внимание на звук (анодная tDCS + внимание к звуку), и 3) анодная tDCS применялась без внимания участников к мышце FDI или звуку (анодная tDCS + Без внимания) (рис.1а). Порядок условий был сбалансирован для участников.
Рис. 1Экспериментальная парадигма для анодной tDCS в сочетании с вниманием. Динамика эксперимента 1 ( a ) и эксперимента 2 ( b )
Во всех условиях анодная tDCS применялась к моторной коре головного мозга FDI. В условиях анодной tDCS + «Внимание к целевой мышце» и анодной tDCS + «Внимание к звуку» участники выполняли задачу обнаружения соматосенсорной или слуховой цели, соответственно, во время tDCS.В анодном состоянии tDCS + No Attention участники не выполняли задачу по обнаружению цели, и их просили игнорировать целевую мышцу FDI или звук во время tDCS.
Изменения в MEP, SICI и ICF оценивались до и сразу после (0 мин), а также через 10 мин, 30 мин и 60 мин после задания. Чтобы предотвратить эффект переноса от предыдущих вмешательств, между сеансами были вставлены интервалы вымывания в 1 неделю или более. Основываясь на предыдущих выводах о том, что манипулирование вниманием в сочетании с PAS или сенсорным вводом увеличивает возбудимость коры и снижает внутрикортикальное торможение [21, 22], мы предположили, что анодный tDCS + внимание к целевой мышце увеличит MEP и снизит SICI только в целевой мышце FDI. , и что эффекты будут более заметными и сохраняться дольше, чем эффекты в других условиях.
Дополнительный эксперимент
Мы не включили фиктивное состояние tDCS в эксперимент 1. Оставалось неясным, было ли значительное увеличение MEP после tDCS в состоянии, когда участники обращали внимание на целевую мышцу FDI, было результатом взаимодействия tDCS и внимания. или это был только эффект внимания. Чтобы ответить на этот вопрос, еще восемь участников приняли участие в контрольном эксперименте с двумя интервенционными условиями в разные дни: 1) фиктивная tDCS + внимание к целевой мышце и 2) фиктивная tDCS + внимание к звуку.Эксперимент проводился по схеме двойного слепого фиктивного контроля. Изменения MEP мышцы FDI оценивали до и сразу после (0 мин), а также через 10 мин, 30 мин и 60 мин после задания. U-тесты Манна-Уитни с поправками Бонферрони были проведены для оценки различий в MEP между фиктивной tDCS + внимание к целевой мышце и анодной tDCS + вниманием к целевым мышцам, а также между фиктивной tDCS + Attention to Sound и анодной tDCS + Attention to Звуковые условия при каждом тестировании.
Эксперимент 2 (поведенческий эксперимент)
tDCS
Параметры tDCS, примененного к M1, были такими же, как и в эксперименте 1. Анодный электрод располагался над правым M1 APB, а электрод сравнения помещался над ипсилатеральным плечо. Для фиктивного состояния интенсивность была установлена на 2 мА, но ток подавался только в течение 30 с, чтобы имитировать ощущение нарастающего и снижающегося тока, приложенного в конце анодного состояния.
Манипуляция вниманием
Участники выполнили задачу соматосенсорного обнаружения с настройкой, идентичной настройке в эксперименте 1. В течение 10 минут реальной или фиктивной tDCS участникам предъявляли соматосенсорный стимул на APB недоминантной левой руки. через полуслучайные интервалы, примерно каждые 30 с, и их просили сообщить, когда они обнаружили стимул.
Двигательная задача
Баллистическая задача на сгибание использовалась в качестве практической, потому что неоднократно сообщалось, что обучение двигательным навыкам этой задачи модулируется повторяющимися TMS и tDCS [26,27,28,29].Следовательно, целевая мышца была изменена с мышцы FDI в эксперименте 1 на мышцу APB в эксперименте 2, потому что мышца APB играет важную роль в задаче движения баллистического сгибания. Предплечье фиксировали в нейтральном положении между пронацией и супинацией, большой палец был свободен для движения, в то время как пальцы фиксировали на месте жесткой связкой. Затем к подушечке большого пальца левой руки был прикреплен акселерометр. Пиковое ускорение баллистического движения большого пальца регистрировалось акселерометром с использованием встроенной электроники (модель 25A; Endevco, Сан-Хуан-Капистрано, Калифорния, США).Сигнал усиливался с помощью формирователя сигнала с низким уровнем шума (модель 4416B Isotron Signal Conditioner; Endevco) с питанием от батареи. Сигналы ускорения усиливались (в 10 раз) и оцифровывались с частотой 2000 Гц с помощью аналого-цифрового преобразователя и записывались на компьютере для автономного анализа. Специальная программа LabVIEW была создана для запуска движения с помощью слухового сигнала, обеспечения визуальной обратной связи и записи данных о двигательных характеристиках.
Участники сидели перед экраном компьютера.Их попросили как можно быстрее согнуть большой палец левой руки после звукового сигнала, а затем полностью расслабить левую руку до следующего удара. Сигналы ускорения измерялись через 1,5 с после звукового сигнала. Через 1,5 с после получения значения акселерометра участники получили визуальную обратную связь о пиковом ускорении их баллистического движения большого пальца через экран компьютера, который представлял цветной сигнал. Когда участники работали быстрее, чем медиана из предыдущих пяти значений ускорения, на экране компьютера отображался синий прямоугольник.Напротив, когда участники работали медленнее, чем медиана из предыдущих пяти значений ускорения, был представлен красный прямоугольник. Пиковое ускорение баллистического движения большого пальца было проанализировано как индикатор двигательной активности. Было рассчитано среднее значение пиковых ускорений в каждом блоке.
Методика эксперимента
Мы использовали двойной слепой эксперимент с фиктивным контролем. Участники были случайным образом разделены на одну из трех групп: 1) анодная tDCS + внимание к целевой мышце, 2) анодная tDCS + отсутствие внимания и 3) фиктивная tDCS + внимание к целевой мышце (рис.1б).
Перед вмешательством участники отработали 20 попыток баллистических движений большого пальца, чтобы привыкнуть к задаче. После этого участники выполнили один сеанс баллистической задачи (60 испытаний) в качестве исходного уровня. После вмешательства они выполнили пять сеансов баллистической задачи (всего 300 попыток). Последующие измерения (пять сеансов баллистической задачи) были проведены через 1 день, 7 дней и 30 дней после первой баллистической задачи, чтобы изучить долгосрочные различия в двигательных характеристиках между группами.Мы предположили, что усиление кортикальной пластичности, вызванное анодной tDCS + Внимание к целевой APB-мышце, улучшило бы моторное обучение баллистическому движению большого пальца и, таким образом, привело бы к более высокой долгосрочной производительности по сравнению с другими условиями [17, 18].
Статистический анализ
Тест Шапиро-Уилка использовался для определения нормального распределения амплитуды MEP,% MEP, SICI, ICF и данных производительности. Для эксперимента 1 был использован дисперсионный анализ смешанной модели с повторными измерениями (ANOVA) для оценки эффектов каждой задачи (анодная tDCS + внимание к целевой мышце, анодальная tDCS + внимание к звуку, анодная tDCS + отсутствие внимания) и каждого из них. время тестирования (Post0, Post10, Post30 и Post60) на% MEP, SICI и ICF при нормальном распределении данных.Парные t-тесты с поправками Бонферрони для множественных сравнений были выполнены для апостериорных сравнений. Для данных, которые не были нормально распределены, тест Краскала-Уоллиса использовался для оценки основного эффекта каждой задачи (анодная tDCS + внимание к целевой мышце, анодальная tDCS + внимание к звуку, анодальная tDCS + отсутствие внимания) в каждый момент времени. . U-тесты Манна-Уитни с поправками Бонферрони были проведены для оценки межгрупповых различий.
Для эксперимента 2 повторный анализ смешанной модели ANOVA с группой факторов (анодальный tDCS + внимание к целевой мышце, анодальный tDCS + NO Attention, фиктивный tDCS + внимание к целевой мышце) и сеанс (исходный уровень, 1 набор, 2 наборов, 3 подхода, 4 подхода и 5 подходов двигательной задачи) было выполнено, чтобы исследовать, могут ли эффекты анодной tDCS в сочетании с вниманием к APB-мышце улучшить приобретение баллистических движений большого пальца.ANOVA смешанной модели с повторными измерениями с группой факторов (анодная tDCS + внимание к целевой мышце, анодная tDCS + без внимания, фиктивная tDCS + внимание к целевой мышце) и временным курсом (исходный уровень, сразу после, 1 день после, 7 дней через 30 дней после двигательной задачи) также было выполнено, чтобы проверить, могут ли эффекты анодной tDCS в сочетании с анодной tDCS улучшить выполнение заученного движения. Множественные попарные сравнения с поправками Бонферрони были выполнены для апостериорных сравнений, когда значимый результат был получен в первичных анализах.Для данных, которые не были нормально распределены, были выполнены U-тесты Манна-Уитни с поправками Бонферрони для оценки различий внутри и между группами. Значения P <0,05 считались статистически значимыми для всех анализов. Статистический анализ выполнялся с использованием IBM SPSS 24.0 (IBM Corp., Нью-Йорк, Нью-Йорк, США) для Windows.
Данные одного участника отсутствовали из-за проблемы с устройством в фиктивном состоянии tDCS + «Внимание к целевой мышце» через 1 день после первой баллистической задачи.Также отсутствовали некоторые данные по анодному состоянию tDCS + внимание к целевой мышце (два участника), условному tDCS + внимание к целевой мышце (три участника) и анодному состоянию tDCS + без внимания (три участника) через 30 дней. после первой баллистической задачи по той же причине.
Результаты
Тест Шапиро-Уилка подтвердил, что все данные, кроме амплитуд MEP и% MEP, имели нормальное распределение.
Эксперимент 1
MEP
Средние необработанные значения (стандартное отклонение: SD) амплитуд MEP в мышце FDI на исходном уровне были равны 0.47 (0,18) мВ в анодной tDCS + внимание к целевой мышце, 0,59 (0,29) мВ в анодной tDCS + внимание к звуку и 0,57 (0,45) мВ в анодной tDCS + состояние отсутствия внимания. Эти исходные значения существенно не отличались друг от друга (критерий Краскела-Уоллиса, P = 0,314). Средние необработанные значения (SD) амплитуд MEP в APB на исходном уровне составляли 0,36 (0,42), 0,43 (0,37) и 0,28 (0,27) мВ, также существенно не отличались друг от друга (критерий Краскела-Уоллиса, P ). = 0.546). Средние необработанные значения (SD) амплитуд МВП в ЭКР на исходном уровне составляли 0,32 (0,30), 0,45 (0,28) и 0,31 (0,21) мВ, также существенно не отличающиеся друг от друга (критерий Краскела-Уоллиса, P ). = 0,447).
Временной ход% MEP показан на рис. 2. Чтобы подтвердить влияние анодного tDCS на амплитуды MEP между исходным уровнем и Post0 в каждой мышце (FDI, APB и ECR), были проведены односторонние знаковые ранговые тесты Вилкоксона. выполняется при каждом условии, исходя из предположения, что анодная tDCS увеличивает амплитуды МВП [7].По сравнению с исходным уровнем, амплитуды MEP значительно увеличились в Post0 в анодной tDCS + внимание к целевой мышце ( P = 0,002 для FDI-мышцы, P = 0,004 для APB-мышцы и P = 0,048 для ECR-мышцы). в анодном tDCS + внимание к звуку ( P = 0,049 для FDI-мышцы, P = 0,039 для APB-мышцы и P = 0,002 для ECR-мышцы), а в анодном tDCS + состояние отсутствия внимания ( P = 0,048 для мышцы FDI, P = 0.004 для мышцы APB и P = 0,004 для мышцы ECR). Эти результаты показывают, что анодная tDCS увеличивает возбудимость коры во всех мышцах сразу после стимуляции.
Рис. 2Эффекты анодной tDCS в сочетании с вниманием к моторным вызванным потенциалам (MEP). Амплитуды MEP в первой спинной межкостной мышце (FDI; a ), коротком отводящем большом пальце (APB; b ) и лучевом разгибателе запястья (ECR; c ) были нормализованы до базовой амплитуды (%) для каждого состояния. .Графики белого ящика обозначают анодную tDCS, примененную, когда участники обращали внимание на целевую мышцу FDI. Светло-серые прямоугольные диаграммы обозначают анодные tDCS, примененные, когда участники обращали внимание на звук. Темно-серые прямоугольные диаграммы обозначают анодную tDCS, примененную без внимания участников к целевой мышце FDI или звуку. Медианный и межквартильный диапазоны представлены горизонтальными линиями внутри прямоугольников и усов (представляющих минимальные и максимальные значения) соответственно. Звездочки указывают на существенные различия ( P <0.05) среди вмешательств
тестов Краскела-Уоллиса затем использовались для оценки эффектов каждой задачи в каждый момент времени. Были значительные основные эффекты задания на% MEP в мышце FDI в Post0 ( P = 0,002), Post10 ( P = 0,002), Post30 ( P = 0,004) и Post60 ( P = 0,003). ) (Рис. 2а). Не было обнаружено значительных основных эффектов на% MEP в мышце APB в Post0 ( P = 0,344), Post10 ( P = 0,448), Post30 ( P = 0.118) и Post60 ( P = 0,798) (рис. 2b), и нет значимых основных эффектов на% MEP в ECR-мышце в Post0 ( P = 0,615), Post10 ( P = 0,162), Post30. ( P = 0,927) и Post60 ( P = 0,395) (рис. 2c). Мы обнаружили, что анодная tDCS + внимание к целевой мышце значительно увеличивает% MEP в FDI-мышце по сравнению с анодной tDCS + Attention to Sound в Post0 ( P = 0,008), Post10 ( P = 0,007), Post30 ( P = 0.019) и Post60 ( P = 0,030) (рис. 2а). Кроме того, анодная tDCS + внимание к целевой мышце значительно увеличила% MEP в FDI-мышце по сравнению с анодной tDCS + No Attention at Post0 ( P = 0,045), Post30 ( P = 0,033) и Post60 ( P = 0,047) (рис. 2а). Эти результаты показывают, что внимание к целевой мышце усиливало индуцированную tDCS моторную кортикальную возбудимость, и региональные эффекты наблюдались в целевой мышце.
Результаты дополнительного эксперимента показывают, что анодный tDCS + внимание к целевой мышце значительно увеличивал% MEP в FDI на Post0 ( P <0.001), Post10 ( P <0,001), Post30 ( P = 0,004) и Post60 ( P = 0,008) по сравнению с фиктивным tDCS + внимание к целевой мышце (рис. 3). Не было значительных различий в основных эффектах задачи между анодной tDCS + Attention to Sound и фиктивной tDCS + Attention to Sound относительно% MEP в FDI в Post0 ( P = 0,321), Post10 ( P = 0,236), Post30. ( P = 0,963) и Post60 ( P = 0,423). Эти результаты показывают, что двигательная возбудимость коры головного мозга была увеличена только тогда, когда анодная tDCS была объединена с вниманием к целевой мышце.
Рис. 3Эффекты фиктивной tDCS в сочетании с вниманием к моторным вызванным потенциалам (MEP). Амплитуды MEP в первом дорсальном межкостном переходе (FDI) были нормализованы к базовой амплитуде (%) для каждого состояния. Графики белого ящика обозначают анодную tDCS, примененную, когда участники обращали внимание на целевую мышцу FDI. Светло-серые прямоугольные диаграммы обозначают фиктивную tDCS, примененную, когда участники обращали внимание на целевую мышцу FDI. Темно-серые прямоугольные диаграммы обозначают фиктивную tDCS, примененную, когда участники обращали внимание на звук.Медианный и межквартильный диапазоны представлены горизонтальными линиями внутри прямоугольников и усов (представляющих минимальные и максимальные значения) соответственно. Звездочки указывают на значительные различия ( P <0,05) между анодной tDCS в сочетании с вниманием к целевой мышце и фиктивной tDCS в сочетании с вниманием к целевой мышце. Не наблюдалось существенной разницы между ложными условиями tDCS ( P > 0,05)
SICI и ICF
. Значения SICI и ICF показаны в таблице 1.SICI снизился после анодной tDCS в сочетании с вниманием к целевой мышце FDI в течение 60 минут или дольше. Напротив, длительные эффекты наблюдались до 15 минут после анодной tDCS без внимания. Никаких явных изменений в SICI 2ms и SICI 3ms не наблюдалось в анодном состоянии tDCS + Attention to Sound. При сравнении SICI между условиями в каждой временной точке тестирования было обнаружено, что анодная tDCS + внимание к целевой мышце усиливает модулирующий эффект анодной tDCS на SICI 2 мс и SICI 3 мс .Для ICF никаких изменений не наблюдалось ни в одном из условий tDCS анода.
Таблица 1 Изменения в SICI и ICF до и после анодной tDCS в сочетании с вниманиемПриведенные выше результаты были подтверждены дисперсионным анализом, показывающим значительные взаимодействия между условиями и временем тестирования, которые были основными факторами, представляющими интерес для настоящего эксперимента. Значимые основные эффекты состояния (SICI 2 мс : F 2 , 16 = 12,28; SICI 3 мс : F 2 , 16 = 9.51) и время тестирования (SICI 2 мс : F 4 , 32 = 13,88; SICI 3 мс : F 4 , 32 = 20,86) были квалифицированы значительными взаимодействиями для SICI 2 мс ( F 8 , 64 = 2,45) и SICI 3 мс ( F 8 , 64 = 2,12). Взаимодействия показали, что время тестирования влияет на состояние, показывая, что влияние условия в основном связано с моментом времени тестирования на SICI 2 мс и SICI 3 мс .Чтобы прояснить этот эффект, для апостериорного анализа были выполнены множественные попарные сравнения.
По сравнению с исходными значениями, анодный tDCS + внимание к целевой мышце значительно снизил SICI 2 мс и SICI 3 мс в Post0, Post15 и Post60, в то время как анодальный tDCS + No Attention значительно снизил SICI 3 мс в Post0 и Post15 (см. Таблицу 1).
SICI 2 мс было значительно снижено анодной tDCS + внимание к целевой мышце по сравнению с анодной tDCS + Attention to Sound в Post0, Post15, Post30 и Post60 (см. Таблицу 1).По сравнению с анодным tDCS + No Attention, SICI 2 мс также значительно снизился на Post30 и Post60. SICI 3 мс был значительно снижен анодной tDCS + внимание к целевой мышце на Post30 и Post60 по сравнению с анодной tDCS + Attention to Sound. Анодный tDCS + No Attention значительно снизил SICI 2 мс и SICI 3 мс на Post0 по сравнению с анодным tDCS + Attention to Sound.
Не было обнаружено значимого взаимодействия для ICF 10 мс ( F 8 , 64 = 0.60) или ICF 15 мс ( F 8 , 64 = 1,12). Не было значительных основных эффектов протокола и времени тестирования для ICF 10 мс (условие: F 2 , 16 = 1,24; время тестирования: F 4 , 32 = 0,69) или ICF 15 мс (условие: F 2 , 16 = 2,29; время тестирования: F 4 , 32 = 1,70). Эти результаты показывают, что влияние условий не было связано с моментом времени тестирования на ICF 10 мс и ICF 15 мс .
Эксперимент 2
Средняя (SD) моторная производительность на исходном уровне, измеренная как пиковое ускорение, составила 3,35 (0,51) г в анодном tDCS + состояние целевых мышц, 3,54 (0,49) г в анодном tDCS + состояние без внимания и 3,22 (0,61) г в фиктивном состоянии tDCS + Attention to Target Muscle. Базовые двигательные характеристики не различались значимо среди трех условий (ANOVA, F 2 , 24 = 0,09, P = 0,914).
Немедленное влияние на моторное обучение
Динамика двигательной активности в каждом блоке после вмешательств показана на рис.4. Значительное взаимодействие было обнаружено для двигательной активности ( F 10 , 105 = 3,54, P <0,001). Был отмечен значительный основной эффект сеанса ( F 5 , 105 = 10,02, P <0,001), в то время как основной эффект не был обнаружен для группы ( F 2 , 21 = 1,46 , P = 0,254). Моторные характеристики улучшились после набора 5 в анодном состоянии tDCS + Внимание к целевой мышце по сравнению с исходным уровнем ( P = 0.013), и после первого набора задачи баллистического движения ( P = 0,039) (рис. 4). Производительность была значительно улучшена после набора 5 в состоянии анодного tDCS + Внимание к целевой мышце по сравнению с анодным tDCS + Без внимания ( P = 0,048) и фиктивным tDCS + Attention to Target Muscle ( P = 0,014). Это привело к тому, что анодная группа tDCS + «Внимание к целевой мышце» превзошла другие группы после первого набора задачи баллистического движения, что указывает на то, что анодная tDCS + «Внимание к целевой мышце», примененная перед задачей баллистического движения, улучшает приобретение двигательных навыков.
Рис. 4Непосредственные эффекты анодной tDCS в сочетании с вниманием к моторному обучению. Графики в белом поле обозначают анодную tDCS, примененную, когда участники обращали внимание на целевую мышцу APB. Светло-серые прямоугольные диаграммы обозначают анодную tDCS, примененную без внимания участников к целевой APB-мышце. Темно-серые прямоугольные диаграммы обозначают фиктивную tDCS, примененную, когда участники обращали внимание на целевую мышцу APB. Медиана и межквартильный размах представлены горизонтальными линиями внутри прямоугольников и усов (представляющих минимальные и максимальные значения), звездочки указывают на существенные различия ( P <0.05) между исходным уровнем и каждой временной точкой вмешательства или в рамках вмешательств
Изменения в производительности до 30 дней после вмешательств
Временной ход двигательной активности в каждый день после вмешательств показан на рис. 5. Существенное взаимодействие было обнаружено для моторных характеристик ( F 8 , 75 = 3,31, P = 0,003) и значимых основных эффектов группы ( F 2 , 21 = 3,56, P = 0.046) и хода времени ( F 4 , 75 = 13,09, P <0,001). По сравнению с исходными значениями, анодный tDCS + внимание к целевой мышце значительно улучшил двигательную активность через 1 день после ( P = 0,001), через 7 дней после ( P <0,001) и через 30 дней после двигательной задачи ( P = 0,012) (рис.5). Фальшивая tDCS + Внимание к задаче значительно улучшила моторные характеристики через 7 дней после моторной задачи ( P = 0,046), в то время как анодная tDCS + No Attention не улучшила производительность.Моторные характеристики были значительно увеличены за счет анодной tDCS + внимания к целевой мышце (по сравнению с анодной tDCS + без внимания и фиктивной tDCS + с вниманием к целевой мышце) через 1 день после (по сравнению с анодной tDCS + без внимания, P = 0,020), и через 7 дней после (по сравнению с анодной tDCS + без внимания, P = 0,024; по сравнению с фиктивной tDCS + с вниманием к целевой мышце, P = 0,039) вмешательства. Эти результаты показывают, что анодная tDCS в сочетании с вниманием к целевой мышце повышает эффективность выученного баллистического движения.
Рис. 5Изменения в характеристиках двигателя в течение 30 дней после анодной tDCS в сочетании с вниманием. Графики в белом поле обозначают анодную tDCS, примененную, когда участники обращали внимание на целевую мышцу APB. Светло-серые прямоугольные диаграммы обозначают анодную tDCS, примененную без внимания участников к целевой APB-мышце. Темно-серые прямоугольные диаграммы обозначают фиктивную tDCS, примененную, когда участники обращали внимание на целевую мышцу APB. Медиана и межквартильный размах представлены горизонтальными линиями внутри прямоугольников и усов (представляющих минимальные и максимальные значения), звездочки указывают на существенные различия ( P <0.05) между исходным уровнем и каждой временной точкой вмешательства или в рамках вмешательств
Повышенная возбудимость подкисленных скелетных мышц | Журнал общей физиологии
Чтобы определить, может ли влияние ацидоза на возбудимость мышц при 11 мМ K + (рис. 1) быть связано с изменением G Cl —, параметров кабеля от четырех групп мышц при 11 мМ K + (pH 7,4 или 6,8, с или без Cl — в буфере).Проникновение волокон от 4-недельных животных с двумя электродами одновременно приводило, однако, к значительной деполяризации, что делало использование больших удерживающих токов, необходимых для поддержания постоянного потенциала покоя. Чтобы избежать этой проблемы, исследование параметров кабеля проводилось на более крупных мышцах взрослых крыс. На рис. 2А показаны характерные ответы мембранного потенциала на гиперполяризационный ток -40 нА, зарегистрированные в четырех разных местах в волокне, инкубированном при pH 6,8 в буфере, содержащем Cl —.На основе этих записей были рассчитаны отношения ΔV м / I, которые соотносятся с межэлектродным расстоянием, как показано на рис. 2В, где показаны измерения отдельных волокон каждой из четырех групп мышц, отражающие среднее значение для каждой группы. В буфере, содержащем Cl —, низкий рН мышц приводил к более высоким отношениям ΔV m / I, чем нормальный рН, указывая на то, что проводимость мембраны была снижена из-за подкисления. Напротив, волокна в буфере, не содержащем Cl — , не подвергались воздействию подкисления, что указывает на то, что влияние pH только на проводимость мембраны было связано с изменением G Cl — .Для более качественного исследования влияния ацидоза на компонентную проводимость K + и Cl —, λ, R в и G m были определены в каждой из четырех групп в соответствии с методиками. Бойда и Мартина (1959). В таблице I показаны рассчитанные значения λ, R в и G м для волокон при 4 мМ K + (нормальный pH с Cl — ) и для четырех групп волокон при 11 мМ K +. . Когда pH снизился с 7.От 4 до 6,8 в мышцах, инкубированных в буфере, содержащем Cl — при 11 мМ K + , λ значительно увеличился на 18% (P <0,01) и R в на 40% (P <0,01). Следовательно, мышцы, инкубированные в буфере, содержащем Cl —, показали снижение проводимости мембраны с 2136 ± 150 мкСм / см 2 при pH 7,4 до 1393 ± 56 мкСм / см 2 при pH 6,8 (P <0,01). В мышцах, инкубированных в буфере, не содержащем Cl —, не было значительных изменений λ или R в с подкислением, что также отражалось небольшим и незначительным изменением G K + с 405 ± 20 мкСм. / см 2 при нормальном pH до 455 ± 30 мкСм / см 2 при низком pH (P <0.16). Основываясь на этих результатах, подкисление с 7,4 до 6,8 за счет увеличения CO 2 с 5 до 24% вызвало снижение G Cl — с 1731 ± 151 мкСм / см 2 при нормальном pH до 938 ± 64 мкСм / см 2 при низком pH, соответствующем снижению на 46% (P <0,01).
Чтобы проверить, повлияло ли на измерения G Cl — влияние ацидоза на диаметр мышечных волокон, внутриклеточное содержание воды было измерено в каждой из четырех групп мышц взрослых животных при 11 мМ К. + и сравнивается со средним диаметром волокна, рассчитанным на основе измерений параметров кабеля.Поскольку рассчитанный диаметр существенно не отличался между группами (таблица I), можно ожидать, что внутриклеточное содержание воды будет одинаковым во всех группах. В мышцах, инкубированных в буфере, содержащем Cl —, при pH 7,4, содержание внутриклеточной воды составляло 2,75 ± 0,05 г H 2 O / г сухого веса, что согласуется с ранее опубликованными значениями для взрослых крыс (Cieslar et al.