
Объединение нейронов в нервный центр: 1.2. Функциональная организация цнс . Центральная нервная система. Анатомия и физиология
1.2. Функциональная организация цнс . Центральная нервная система. Анатомия и физиология
Уровни функциональной организации
Единственное, с чем имеет дело мозг, – это информация. Все наши действия руководствуются информацией. Мозг функционирует, пока есть информация. Но что для мозга является информацией? Для мозга информация – это изменение среды, преобразованное в нервный код.
Элементарным обработчиком информации в мозге является нервная клетка, а единицей информации в ЦНС – потенциал действия. Потенциал действия – это электрический импульс, который генерирует нервная клетка и с помощью которого она воздействует на другие клетки. Нейрон кодирует передаваемую им информацию временной последовательностью потенциалов действия (рис. 15).
Рис. 15. Пример записи импульсной активности нервной клетки
Но нейрон – это лишь самый нижний уровень работы с информацией. Нервные клетки в ЦНС объединены в нервные центры. Нервный центр – это скопление нейронов, объединенных общей функцией.
Однако нервные центры – это не монолитные образования. Они подразделяются на модули. Модуль – это функциональное объединение нейронов для совместной обработки информации.
Вместе с тем нервные центры – это не обособленные, не независимые образования. Они объединяются в функциональные системы. Функциональная система – это набор нервных центров, участвующих в выполнении определенной функции.
Таким образом, мы можем выделить несколько уровней функциональной организации ЦНС:
1) нейрон – решающее устройство первого уровня, его задача – кодирование информации;
2) модуль – решающее устройство второго уровня, его задача – обработка информации;
3) нервный центр – командное устройство, его задача – формирование сообщений;
4) функциональная система, ее задача – реализация определенных функций.
Принципы обработки информации
Изучение мозга – это фактически изучение того, как он обрабатывает информацию. Современная наука представляет себе этот процесс в самом общем виде, и выглядит это следующим образом:
Современная наука представляет себе этот процесс в самом общем виде, и выглядит это следующим образом:
– обработка происходит в форме последовательных этапов, каждый этап реализуется определенным нервным центром;
– на каждом из этапов выполняется уникальная операция;
– система функционирует по принципу отрицательной обратной связи: решение нервным центром задачи прерывает его активность.
Такой алгоритм обработки информации возможен благодаря тому, что нервные центры находятся в иерархическом соподчинении. Иерархическая организация нервных центров выражается в следующем:
– нижележащий центр является информационным по отношению к вышележащему;
– вышележащий центр является управляющим по отношению к нижележащему.
В результате каждый нервный центр, входящий в определенную иерархическую систему, имеет два входа и два выхода (рис. 16).
Вход из нижележащего центра является информационным, вход из вышележащего центра – управляющим. Выход, направленный вверх является информационным, выход, направленный вниз, – управляющим.
Рис. 16. Входы и выходы нервного центра
Базовые процессы в нервной системе
Нервные центры взаимодействуют посредством двух процессов – возбуждения и торможения. Эти процессы характеризуются определенным набором свойств.
Свойства нервных процессов
1) Возбуждение и торможение могут иррадиировать.
Иррадиация – это распространение процесса из очага возникновения на окружающие клетки. Чем сильнее процесс в очаге возникновения, тем дальше он иррадиирует. Возбуждение иррадиирует быстрее торможения.
2) Иррадиация неизбежно сменяется концентрацией. Концентрация – это сосредоточение процесса в месте его первоначального возникновения. Концентрация происходит медленнее, чем иррадиация.
3) Концентрация сопровождается индукцией. Индукция – это свойство вызывать вокруг себя и после себя противоположный процесс. Индукция вытесняет процесс из той области, на которую он иррадиировал.
4) Возбуждение и торможение могут взаимодействовать. Это происходит при иррадиации процессов. Результат взаимодействия зависит от относительной силы конкурирующих процессов.
Распространение возбуждения и торможения
Возбуждение в нервной системе распространяется самостоятельно, торможение распространяется с помощью возбуждения (рис. 17).
Рис. 17. Схемы, иллюстрирующие процессы распространения возбуждения и торможения в нервной системе
Возбудительные связи между нервными центрами могут быть топическими (А), дивергентными (Б) и конвергентными (В) (рис. 18). Топическая связь обеспечивает передачу информации «точка в точку». Такой характер связей позволяет сохранять пространственные характеристики сигналов. Конвергентные и дивергентные связи обеспечивают обмен информацией между параллельными каналами обработки.
Рис. 18. Варианты возбудительных связей в нервной системе Круг – нейронный модуль, колонка кругов – нервный центр
Тормозные связи могут быть четырех типов. Прямое торможение (рис. 19, А) направлено на уменьшение количества каналов обработки.
Прямое торможение (рис. 19, А) направлено на уменьшение количества каналов обработки.
Рис. 19. Варианты тормозных связей Стрелками показаны возбудительные связи, треугольник обозначает тормозную связь. Белые кружки – возбудительные нейроны, черные кружки – тормозные нейроны
Реципрокное торможение (рис. 19, Б) обеспечивает согласованную работу систем-антагонистов: активация одной системы сопровождается подавлением другой. Возвратное торможение (рис. 19, В) позволяет ограничить длительность и силу реакции нервной клетки. Латеральное торможение (рис. 19, Г) призвано ограничить распространение возбуждения в нервном центре и подчеркнуть фокус реакции. Эфферентное торможение (рис. 19, Д) позволяет вышележащему центру регулировать активность нейронов нижележащего центра.
Торможение в ЦНС может быть двух видов: постсинаптическое и пресинаптическое (рис. 20). Первое направлено на дендрит или тело клетки, второе – на аксон. Первое изменяет заряд мембраны в сторону гиперполяризации и тем самым снижает возбудимость нейрона.
Рис. 20. Виды торможения
Базовые механизмы нервной деятельности
Одной из важных сторон нервной деятельности является формирование адаптивных реакций, вызванных изменениями среды. Эти реакции могут носить врожденный характер и обеспечиваться генетически закрепленными связями. Реакции могут быть и приобретенными. В этом случае они реализуются благодаря образованию новых функциональных связей. Кроме того, адаптивное поведение может носить произвольный характер и выражаться в форме деятельности, направленной на удовлетворение потребностей. В основе реализации адаптивного поведения лежит несколько базовых механизмов нервной деятельности.
Механизм реагирования посредством врожденных связей
Условия обитания закрепили за каждым видом определенный набор врожденных поведенческих реакций.
Элементарные безусловные рефлексы представлены простейшими реакциями, которые вызываются активацией локального поля рецепторов. Они отличаются высокой степенью автоматизма и стереотипности и выражаются в реализации простых движений или в приспособительных изменениях работы отдельных внутренних органов.
Координационные безусловные рефлексы представлены комплексными реакциями, которые запускаются активацией широкого спектра рецепторов. Это синхронные сокращения и расслабления разных мышечных групп, или согласованные изменения работы нескольких внутренних органов. В реализацию таких рефлексов включаются обратные связи. Этот уровень реагирования обеспечивает формирование целостных двигательных актов и гомеостатических реакций.
Интегративные безусловные рефлексы – это комплексные поведенческие акты, которые вызываются биологически значимыми стимулами. Они проявляются в виде сложноорганизованной двигательной активности, которая сопровождается адекватными вегетативными изменениями. На этом уровне реагирования происходит переход от безусловных рефлексов к поведенческим актам.
Сложнейшие безусловные рефлексы (инстинкты) – это комплексы безусловных рефлексов в виде фиксированных последовательностей действий. Инстинкты реализуются благодаря наследственной поведенческой программе, общей для всех представителей вида. Эти реакции отражают исторический опыт вида и запускаются ключевыми стимулами при условии наличия определенной биологической потребности. При реализации инстинкта завершение одной безусловной реакции является сигналом к началу следующей. Этим определяется жесткая последовательность инстинктивных реакций. Достижение данного уровня рефлекторного реагирования обеспечивает переход к сложным формам поведения.
Эти реакции отражают исторический опыт вида и запускаются ключевыми стимулами при условии наличия определенной биологической потребности. При реализации инстинкта завершение одной безусловной реакции является сигналом к началу следующей. Этим определяется жесткая последовательность инстинктивных реакций. Достижение данного уровня рефлекторного реагирования обеспечивает переход к сложным формам поведения.
Реагирование посредством врожденных связей – это механизм низшей нервной деятельности. На ранних этапах эволюции нервной системы безусловные реакции – единственная форма реагирования. Круг воспринимаемых раздражителей ограничен, ограничен и репертуар возможных ответных действий. Но безусловные рефлексы не в состоянии в полной мере обеспечить уравновешивание организма со средой. Среда изменчива, а набор врожденных реакций ограничен.
Механизм реагирования посредством приобретенных связей
Помимо врожденных стереотипных реакций нервная система обладает механизмом, позволяющим в течение жизни создавать новые рефлекторные реакции. Такое оперативное приспособление к условиям обитания возможно благодаря пластичности нервной системы. В основе образования новых рефлекторных реакций лежит механизм формирования временных связей между нервными центрами. Такие приобретенные реакции получили название условных рефлексов. Речь идет о формировании не анатомической (она существует изначально), а функциональной связи, которая возникает благодаря облегчению синаптической передачи. Такое «проторение пути» между нервными центрами возможно при условии их многократного одновременного возбуждения. Возбуждение из нервных центров иррадиирует навстречу друг другу, суммируется, и в области повышенного возбуждения происходит облегчение синаптической передачи. В результате формируется новая рефлекторная дуга, афферентным звеном которой является центр условного раздражителя, а эфферентным звеном – центр безусловного рефлекса. Благодаря образованию временнои связи условный сигнал начинает активировать центр безусловного рефлекса. Условный рефлекс – это индивидуально приобретенная реакция на ранее индифферентный раздражитель, воспроизводящая безусловную реакцию.
Такое оперативное приспособление к условиям обитания возможно благодаря пластичности нервной системы. В основе образования новых рефлекторных реакций лежит механизм формирования временных связей между нервными центрами. Такие приобретенные реакции получили название условных рефлексов. Речь идет о формировании не анатомической (она существует изначально), а функциональной связи, которая возникает благодаря облегчению синаптической передачи. Такое «проторение пути» между нервными центрами возможно при условии их многократного одновременного возбуждения. Возбуждение из нервных центров иррадиирует навстречу друг другу, суммируется, и в области повышенного возбуждения происходит облегчение синаптической передачи. В результате формируется новая рефлекторная дуга, афферентным звеном которой является центр условного раздражителя, а эфферентным звеном – центр безусловного рефлекса. Благодаря образованию временнои связи условный сигнал начинает активировать центр безусловного рефлекса. Условный рефлекс – это индивидуально приобретенная реакция на ранее индифферентный раздражитель, воспроизводящая безусловную реакцию. Адаптивное значение условного рефлекса состоит в том, что раздражитель, который прежде был не значимым, становится сигналом возможных изменений среды. Это позволяет организму подготовиться к предстоящим событиям. Образование временных связей – это механизм индивидуального приспособления: условные рефлексы изменчивы и бесконечно разнообразны, они образуются и исчезают.
Адаптивное значение условного рефлекса состоит в том, что раздражитель, который прежде был не значимым, становится сигналом возможных изменений среды. Это позволяет организму подготовиться к предстоящим событиям. Образование временных связей – это механизм индивидуального приспособления: условные рефлексы изменчивы и бесконечно разнообразны, они образуются и исчезают.
Реагирование посредством приобретенных связей – это механизм высшей нервной деятельности. Условные рефлексы встраиваются в качестве компонентов в сложные поведенческие акты. Однако условно-рефлекторное обучение является неосознаваемым. Никто из нас не в состоянии сказать, каким набором условных рефлексов он располагает.
Вместе с тем все разнообразие форм поведения невозможно объяснить только лишь рефлекторными реакциями, даже весьма сложными. В ЦНС реализуется еще один механизм, позволяющий сделать поведение бесконечно разнообразным и обеспечивающий наибольший адаптационный потенциал.
Механизм формирования доминанты
Когда достижение результата невозможно путем рефлекторного реагирования, в мозге формируется функциональная система, призванная удовлетворить возникшую потребность. Функциональная система – это временное объединение нервных центров. Оно возможно благодаря образованию доминанты.
Функциональная система – это временное объединение нервных центров. Оно возможно благодаря образованию доминанты.
Доминанта – это система нервных центров с устойчиво высокой возбудимостью. Это физиологическое проявление мотивации, определяющей направленность поведения. Она создает готовность организма к определенному виду деятельности. Доминанта характеризуется рядом свойств:
– повышенная возбудимость: даже слабые раздражители способны усилить возбуждение доминанты;
– стойкость, инерционность возбуждения: очаг возбуждения сохраняется длительное время, пока не будет удовлетворена потребность;
– способность к суммированию возбуждения: возбуждение центров, не входящих в доминанту, притягивается к доминанте и поддерживает ее возбуждение;
– сопряженное торможение центров-антагонистов: доминанта активно тормозит центры, которые используются для удовлетворения иных потребностей.
Формирование динамичных функциональных систем на основе образования доминант – это физиологический механизм психической деятельности.
Итак, именно благодаря нервной системе организм функционирует как единое целое, решая при этом комплекс важных задач, связанных с удовлетворением биологических, социальных и высших потребностей.
Контрольные вопросы
1. Что в НС является единицей информации?
2. Как по современным представлениям организован в мозге процесс обработки информации?
3. В чем выражается иерархическая организация нервных центров?
4. Что такое иррадиация?
5. Каков функциональный смысл реципрокного торможения?
6. В чем проявляется эволюция врожденных форм поведения?
7. Что является базовым механизмом низшей нервной деятельности?
8. Какой механизм лежит в основе образования новых рефлекторных реакций?
9. В чем состоит адаптивное значение условного рефлекса?
10. Что такое доминанта и каковы ее свойства?
Данный текст является ознакомительным фрагментом.
ЦНС Flashcards | Quizlet
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ1.Аксонный транспорт(БЕЛКИ КИНЕЗИН И ДИНЕИН) — непрерывное
движение белков, синтезированных в
клетке, медиаторов, а также органелл из тела нейронов по аксону в область нервных окончаний и обратно.
Различают быстрый и медленный, прямой и ретроградный(обратный), аксоток и дендроток.
2.Генераторная функция — способность к
генерации потенциалов
3.Трофическая функция — способность
регулировать метаболизм и геном клетки — мишени.
4.Суммация импульсов — способность суммировать импульсы, поступающие на нервную клетку и нервный центр:
Временная суммация — чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждения постсинаптических потенциалов (ВПСП). Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений.
Пространственная — когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД.
5. Трансформация ритма возбуждения —
способность изменять ритм проходящих
к нейронам нервного центра импульсов.
6.Последействие — рефлекторные акты
заканчиваются через некоторый период(заканчиваются не одновременно с
прекращением действия раздражителя)
7.Постетаническая потенциация — усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, выделяется большое количество нейромедиатора. Это приводит к нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
8.Интегративная функция — суммация ВПСП и ТПСП, возникающих в
различных участках мембраны, в
результате чего в нервной клетке
происходит генерация импульсов
различной модальности, регулирующих
деятельность других клеток и органов.
НЕРВНАЯ СЕТЬ-морфофункциональное объединение различных нейронов, расположенных на разных уровнях ЦНС и участвующих в формировании и регуляции функций организма (мотиваций, эмоций и т.д.)
Типы нервных сетей.
• — локальные
• — дивергентные
• — иерархические
Спонтанная активность нервного центра. Высшая нервная деятельность. Механизм передачи возбуждения в синапсе
Нервный центр – совокупность нервных клеток (нейронов), необходимая для регуляции деятельности других нервных центров или исполнительных органов. Простейший нервный центр состоит из нескольких нейронов, образующих узел (ганглий). У высших животных и человека нервный центр включает тысячи и даже миллионы нейронов. Большинство функций организма обеспечивается рядом нервных центров, расположенных на различных уровнях центральной нервной системы (напр., нервный центр находится в промежуточном, и в коре ). Нервный центр – сложное сочетание нейронов, согласованно включающихся:
- в регуляцию определенной функции;
- в осуществление рефлекторного акта.


Клетки нервного центра связаны между собой синаптическими контактами и отличаются огромным разнообразием и сложностью внешней и внутренней тектоники. В зависимости от выполняемой функции различают:
- чувствительные нервные центры;
- нервные центры вегетативных функций;
- двигательные нервные центры и др.
Понятие нервных центров
Нервный центр – центральный компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
Анатомическое понятие “нервный центр” – это совокупность нейронов, располагающихся в строго определенных отделах центральной нервной системы и осуществляющих один . Например: центр коленного – в передних рогах 2-4 поясничных сегментов ; центр глотания – на уровне : 5, 7, 9 пары черепно-мозговых нервов.
Физиологическое понятие “нервный центр” – это совокупность нейронов, расположенных на различных уровнях центральной нервной системы и регулирующих сложный рефлекторный процесс. Например: центр глотания входит в состав пищевого центра.
Например: центр глотания входит в состав пищевого центра.
Свойства нервных центров
Одностороннее проведение – возбуждение передается с афферентного на эфферентный . Причина: клапанное свойство синапса.
Задержка проведения возбуждения: скорость проведения возбуждения в нервном центре на много ниже таковой по остальным компонентам рефлекторной дуги. Чем сложнее нервный центр, тем дольше проходит по нему нервный импульс. Причина: синаптическая задержка. Время проведения возбуждения через нервный центр – центральное время рефлекса.
Суммация возбуждения – при действии одиночного подпорогового раздражителя ответной нет. При действии нескольких подпороговых раздражителей ответная реакция есть. Рецептивное поле рефлекса – зона расположения , возбуждение которых вызывает определенный рефлекторный акт.
Имеется 2 вида суммации: временная и пространственная.
Временная суммация – возникает ответная реакция при действии нескольких следующих друг за другом раздражителей. Механизм: суммируются возбуждающие постсинаптические потенциалы рецептивного поля одного рефлекса. Происходит суммация во времени потенциалов одних и тех же групп синапсов.
Механизм: суммируются возбуждающие постсинаптические потенциалы рецептивного поля одного рефлекса. Происходит суммация во времени потенциалов одних и тех же групп синапсов.
Пространственная суммация – возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей. Механизм: суммация возбуждающего постсинаптического потенциала от разных рецептивных полей. Суммируются потенциалы разных групп синапсов.
Центральное облегчение – объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный центр иннервирует определенное количество нервных клеток. Эти нейроны – нейронный пул. В каждом нервном центре много пулов. В каждом нейронном пуле – 2 зоны: центральная (здесь афферентное волокно над каждым нейроном образует достаточное для возбуждения количество синапсов), периферическая или краевая кайма (здесь количество синапсов недостаточно для возбуждения). При раздражении возбуждаются нейроны центральной зоны. Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны.
Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны.
Окклюзия – при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть меньше арифметической суммы раздражения каждого из них. Механизм: импульсы сходятся к одним и тем же нейронам центральной зоны. Возникновение окклюзии или центрального облегчения зависит от силы и частоты раздражения. При действии оптимального раздражителя, (максимального раздражителя (по силе и частоте) вызывающего максимальную ответную реакцию) – появляется центральное облегчение. При действии пессимального раздражителя (с силой и частотой вызывающих снижение ответной реакции) – возникает явление окклюзии.
Посттетаническая потенция – усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация возбуждения в синапсах;
Рефлекторное последействие – продолжение ответной реакции после прекращения действия раздражителя:
- кратковременное последействие – в течение нескольких долей секунды. Причина – следовая деполяризация нейронов;
- длительное последействие – в течение нескольких секунд. Причина: после прекращения действия раздражителя возбуждение продолжает циркулировать внутри нервного центра по замкнутым нейронным цепям.
 Причина – следовая деполяризация нейронов;
Причина – следовая деполяризация нейронов;Трансформация возбуждения – несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне происходит трансформация в сторону уменьшения из-за низкой лабильности синапса. На аксонах эфферентного нейрона, частота импульса больше частоты наносимых раздражений. Причина: внутри нервного центра образуются замкнутые нейронные цепи, в них циркулирует возбуждение и на выход из нервного центра импульсы подаются с большей частотой.
Высокая утомляемость нервных центров – связана с высокой утомляемостью синапсов.
Тонус нервного центра – умеренное возбуждение нейронов, которое регистрируется даже в состоянии относительного физиологического покоя. Причины: рефлекторное происхождение тонуса, гуморальное происхождение тонуса (действие метаболитов), влияние вышележащих отделов центральной нервной системы.
Высокий уровень обменных процессов и, как следствие, высокая в . Чем больше развиты нейроны, тем больше необходимо им кислорода. Нейроны проживут без кислорода 25-30 мин, нейроны ствола головного – 15-20 мин, нейроны коры головного мозга – 5-6 мин.
Все основные формы деятельности нервной системы в сложных многоклеточных организмах животных и человека связаны с функционированием определенных групп нервных клеток – нервных центров. Нервный центр является центральным компонентом рефлекторной дуги, в нем происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
Нервный центр – это совокупность нейронов, расположенных на различных уровнях ЦНС и регулирующих сложный рефлекторный процесс или функцию. В нервном центре выделяют следующие отделы: низший, или исполнительный, рабочий и высший, или регуляторный.
Низший (исполнительный ) отдел нервного центра локализован в спинном мозге и передает информацию от рабочего отдела к рабочим органам.
Рабочий отдел нервного центра – это отдел, ответственный за осуществление данной функции, как правило, находится в стволовых отделах головного мозга.
Высший (регуляторный ) отдел нервного центра расположен в коре больших полушарий мозга и регулирует активность рабочего отдела нервного центра, он вмешивается в регуляцию функций эпизодически, при необходимости корректировки автоматической деятельности рабочего отдела. Высшие отделы включаются в работу нервного центра по условно-рефлекторному механизму. Активность регуляторного (высшего) отдела нервного центра зависит от функционального состояния рабочего отдела.
Строение нервного центра можно рассмотреть на примере дыхательного центра. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам. Рабочий отдел представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту.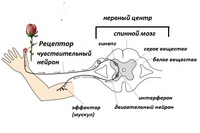 Нарушение этого отдела вызывает остановку дыхания. Регуляторный (высший) отдел дыхательного центра находится в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция ограничена, она зависит от функциональной активности рабочего отдела и афферентной импульсации, отражающей состояние внутренней среды (в данном случае pH крови, концентрации СО2 и О2 в крови).
Нарушение этого отдела вызывает остановку дыхания. Регуляторный (высший) отдел дыхательного центра находится в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция ограничена, она зависит от функциональной активности рабочего отдела и афферентной импульсации, отражающей состояние внутренней среды (в данном случае pH крови, концентрации СО2 и О2 в крови).
Свойства нервных центров обусловлены особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки:
- 1. Одностороннее проведение возбуждения – импульс проводится только в одном направлении, обратное проведение возбуждения через синапс невозможно.
- 2. Наличие латентного периода от начала действия сигнала до проявления рефлекторного акта, так называемая синаптическая задержка. Она обусловлена тем, что на выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5–2 мс. Соответственно, чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
- 3. Суммация возбуждений. В работе нервных центров происходят процессы пространственной и временной суммации подпороговых (недостаточных для передачи импульса через синапс) раздражений. Временная суммация наблюдается, если множество слабых импульсов приходит к нейрону по одному и тому же пути через один синапс с коротким интервалом времени. В результате их действие суммируется, приводя к генерации возбуждения. Пространственная суммация связана с суммированием подпороговых потенциалов, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика нейрона.
- 4. Более низкая скорость передачи импульса в синапсе по сравнению с передачей его по аксону (около 50–100 импульсов в секунду, что в 5-6 раз ниже скорости передачи в аксоне).
- 5. Утомление нервных центров – длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения. Этот процесс связан с деятельностью синапсов – в них наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, снижается реакция постсинаптического рецептора на медиатор. Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой.
- 6. Трансформация ритма – нервные клетки обладают свойством изменять частоту передающихся импульсов. Характер ответного разряда нейрона зависит от свойств раздражителя, а также и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях – чем сильнее раздражение, тем интенсивнее проявляется ответ.
- 7. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходимо постоянное поступление достаточного количества энергии и кислорода. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5-6 мин кислородного голодания они погибают, даже кратковременное ограничение мозгового кровообращения приводит у человека к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15–20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 мин отсутствия кровоснабжения.
- 8. Нервные центры всегда находятся в тонусе, который обеспечивается постоянно поступающими импульсами от разных структур мозга и исполнительных органов. В ответ центры посылают редкие импульсы к органам, поддерживая в них соответствующий тонус. Даже во время сна мышцы не расслабляются полностью и контролируются соответствующими центрами.
- 9. Нервные центры чувствительны к химическим веществам (в том числе лекарственным), проникающим через гематоэицефалический барьер, и обладают специфической реакцией на различные вещества. К примеру, стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов; хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров; апоморфин возбуждает рвотный центр; цититои и лобелии возбуждают дыхательный центр; морфин его угнетает, коразол возбуждает клетки двигательной зоны коры, вызывая судороги.
- 10. После окончания действия раздражителя активное состояние нервного центра продолжается еще некоторое время – так называемое последействие, или следовые процессы. Длительность следовых процессов различна: в спинном мозге – несколько секунд или минут, в подкорковых центрах мозга – десятки минут, часы и даже дни, в коре больших полушарий могут сохраняться до нескольких десятков лет. Следовые процессы имеют важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 ч связано с циркуляцией (реверберацией) импульсов в нервных цепях (Р. Лоренте де Но, 1934) и обеспечивает согласно ревербационной теории кратковременную память. Согласно биохимической теории памяти (X. Хиден, 1969) механизмы долговременной памяти обеспечиваются изменением структуры белков нейронов и глиальных клеток: при запоминании происходят структурные изменения в молекулах РНК, на основе которых строятся новые белки, несущие информацию о прежних раздражителях. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.
 Соответственно, чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
Соответственно, чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса. Этот процесс связан с деятельностью синапсов – в них наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, снижается реакция постсинаптического рецептора на медиатор. Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой.
Этот процесс связан с деятельностью синапсов – в них наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, снижается реакция постсинаптического рецептора на медиатор. Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5-6 мин кислородного голодания они погибают, даже кратковременное ограничение мозгового кровообращения приводит у человека к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15–20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 мин отсутствия кровоснабжения.
Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5-6 мин кислородного голодания они погибают, даже кратковременное ограничение мозгового кровообращения приводит у человека к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15–20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 мин отсутствия кровоснабжения. К примеру, стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов; хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров; апоморфин возбуждает рвотный центр; цититои и лобелии возбуждают дыхательный центр; морфин его угнетает, коразол возбуждает клетки двигательной зоны коры, вызывая судороги.
К примеру, стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов; хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров; апоморфин возбуждает рвотный центр; цититои и лобелии возбуждают дыхательный центр; морфин его угнетает, коразол возбуждает клетки двигательной зоны коры, вызывая судороги. Согласно биохимической теории памяти (X. Хиден, 1969) механизмы долговременной памяти обеспечиваются изменением структуры белков нейронов и глиальных клеток: при запоминании происходят структурные изменения в молекулах РНК, на основе которых строятся новые белки, несущие информацию о прежних раздражителях. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.
Согласно биохимической теории памяти (X. Хиден, 1969) механизмы долговременной памяти обеспечиваются изменением структуры белков нейронов и глиальных клеток: при запоминании происходят структурные изменения в молекулах РНК, на основе которых строятся новые белки, несущие информацию о прежних раздражителях. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.Нервные центры обладают большими возможностями, чем один нейрон. Это проявляется в их свойствах. Во многом эти свойства определяются наличием синапсов. Нервные центры обладают односторонностью проведения возбуждения.
Возбуждение в нервном центре проводитсяот афферентного нейрона к эфферентному. Нервный центр обладает трансформацией ритма; трансформация может быть повышающей (на один импульс нервный центр может ответить «автоматной очередью») или понижающей (в ответ на серию возбуждений нервный центр дает одиночный импульс или не отвечает вовсе). В нервном центре обнаружено последействие. Это свойство связано, вероятно, с наличием в нервных центрах нейронных цепей с положительной обратной связью и с таким свойством синапсов, как постсинаптическая потенциация. Последействие проявляется в продолжающемся рефлекторном процессе при отсутствии раздражителя, вызвавшего эту реакцию. Нервному центру характерна высокая чувствительность к недостатку кислорода. Это связано с интенсивными метаболическими процессами в нейронах. Нервный центр избирательно чувствителен к различным биологически активным веществам, ядам и другим химическим веществам. Это свойство называется хемотропностью. Хемотропность определяется специфическим набором медиаторов, ферментов, нейропептидов и других веществ, которые синтезируются системой нейронов, входящей в конкретный нервный центр. Нервные центры обладают низкой лабильностью и быстрой утомляемостью, а также замедлением проведения возбуждения через нервный центр. Эти свойства связаны с наличием в нервных центрах огромного числа синапсов.
Это свойство связано, вероятно, с наличием в нервных центрах нейронных цепей с положительной обратной связью и с таким свойством синапсов, как постсинаптическая потенциация. Последействие проявляется в продолжающемся рефлекторном процессе при отсутствии раздражителя, вызвавшего эту реакцию. Нервному центру характерна высокая чувствительность к недостатку кислорода. Это связано с интенсивными метаболическими процессами в нейронах. Нервный центр избирательно чувствителен к различным биологически активным веществам, ядам и другим химическим веществам. Это свойство называется хемотропностью. Хемотропность определяется специфическим набором медиаторов, ферментов, нейропептидов и других веществ, которые синтезируются системой нейронов, входящей в конкретный нервный центр. Нервные центры обладают низкой лабильностью и быстрой утомляемостью, а также замедлением проведения возбуждения через нервный центр. Эти свойства связаны с наличием в нервных центрах огромного числа синапсов. Нервные центры всегда находятся в тонусе.Тонус нервных центров (определенный уровень возбудимости) поддерживается притоком различной афферентации из периферии и поступлением возбуждения из нервных центров высших отделов ЦНС. Понижение или повышение тонуса нервных центров лежит в основе регуляции и организации функций.
Нервные центры всегда находятся в тонусе.Тонус нервных центров (определенный уровень возбудимости) поддерживается притоком различной афферентации из периферии и поступлением возбуждения из нервных центров высших отделов ЦНС. Понижение или повышение тонуса нервных центров лежит в основе регуляции и организации функций.
5. Принципы координационной деятельности
центральной нервной системы
Принцип рефлекса
Принцип рефлекса – это универсальная и своеобразная форма взаимодействия организма со средой, происходящая при участии нервной системы.
Понятие о рефлексе возникло в XVI веке в учении
Р. Декарта (1596-1650) о механической картине мира. Р. Декарт жил в эпоху расцвета механики, физики и математики. На его мировоззрение определяющее влияние оказало открытие У. Гарвеем механизма кровообращения и новаторские идеи А. Везалия о том, что носителями психики являются «животные духи», которые вырабатываются в желудочках мозга и передаются по нервам к соответствующим органам. Р. Декарт представлял нервные процессы по образцу системы кровообращения, используя существовавшие в то время принципы оптики и механики.
Р. Декарт представлял нервные процессы по образцу системы кровообращения, используя существовавшие в то время принципы оптики и механики.
Под рефлексом Р. Декарт понимал движение «животных духов» от мозга к мышцам по типу отражения светового луча. Согласно его схеме внешние предметы действуют на периферические окончания расположенных внутри нервных «трубок» нервных «нитей», которые, натягиваясь, открывают клапаны отверстий, ведущих из мозга в нервы. По каналам этих нервов «животные духи» перемещаются в соответствующие мышцы, которые в результате раздуваются, и, таким образом, происходит движение. Однако, находясь под влиянием социально-исторических противоречий своей эпохи, Р. Декарт сделал серьезные уступки идеализму: сознание человека он рассматривал в виде субстанционного начала, способного воздействовать посредством мозговой шишковидной железы на подчиненные рефлекторным законам телесные процессы. Согласно Р. Декарту, тело и душа – самостоятельные субстанции. Поведение и сознание разводились, превращались в два независимых ряда явлений. В этом проявился дуализм Р. Декарта.
В этом проявился дуализм Р. Декарта.
Биологическая концепция рефлекса была сформирована чешским анатомом и физиологом Йиржи Прохазкой (1749-1820). Свои представления о рефлексе Й. Прохазка выразил следующим образом: внешние впечатления, возникающие в чувствительных нервах, быстро распространяются по всей их длине до самого начала. Там они отражаются по определенному закону, переходят на соответствующие им двигательные нервы и по ним очень быстро направляются к мышцам, которые затем производят точные и строго ограниченные движения. Впервые термин «рефлекс» был введен в научный язык Й. Прохазкой. В отличие от Р. Декарта, Й. Прохазка утверждает монистическое представление о нервной системе, в целом относящейся к композиции «общего чувствилища», телесная часть которого локализуется в спинном мозге, а душевная – в головном.
Большой вклад в развитие рефлекторной теории сделали английский анатом и врач Чарльз Белл (1774-1842) и французский физиолог Ф. Мажанди (1783-1855). Они экспериментально показали наличие анатомической основы рефлекса. Так, Ч. Белл в 1811 году в трактате «О новой анатомии мозга» писал, что можно произвести перерезку заднего пучка нервов, исходящего из задней части спинного мозга, без конвульсивных сокращений мышц спины. Однако это становится невозможным даже при одном прикосновении кончиком ножа к переднему корешку. Переход нервного возбуждения по афферентным нервам через спинной мозг на эфферентные нервы получил название закона Белла-Мажанди. Ч. Белл создал теорию о «мышечной чувствительности» и сформулировал физиологическое обоснование циклической функции нервной системы . Между мозгом и мышцей имеется замкнутый нервный круг: один нерв передает влияние от мозга к мышце, а другой передает в мозг чувство состояния мышцы. Если круг будет разомкнут перерезкой моторного нерва, то исчезнет движение. Если же он размыкается перерезкой чувствительного нерва, пропадает ощущение самой мышцы, а вместе с тем исчезает и регуляция ее деятельности.
Они экспериментально показали наличие анатомической основы рефлекса. Так, Ч. Белл в 1811 году в трактате «О новой анатомии мозга» писал, что можно произвести перерезку заднего пучка нервов, исходящего из задней части спинного мозга, без конвульсивных сокращений мышц спины. Однако это становится невозможным даже при одном прикосновении кончиком ножа к переднему корешку. Переход нервного возбуждения по афферентным нервам через спинной мозг на эфферентные нервы получил название закона Белла-Мажанди. Ч. Белл создал теорию о «мышечной чувствительности» и сформулировал физиологическое обоснование циклической функции нервной системы . Между мозгом и мышцей имеется замкнутый нервный круг: один нерв передает влияние от мозга к мышце, а другой передает в мозг чувство состояния мышцы. Если круг будет разомкнут перерезкой моторного нерва, то исчезнет движение. Если же он размыкается перерезкой чувствительного нерва, пропадает ощущение самой мышцы, а вместе с тем исчезает и регуляция ее деятельности.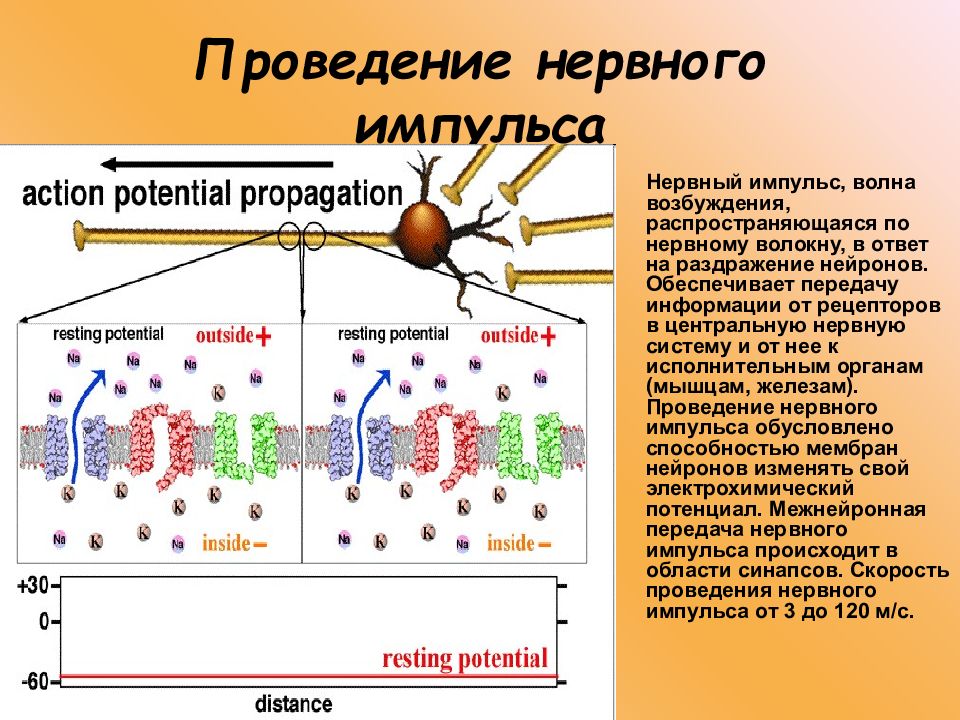 Так, например, у женщины на одной руке была потеряна чувствительность, а на другой – способность к движению. Она могла держать ребенка на руке, потерявшей только чувствительность, до тех пор, пока на него смотрела. Как только женщина отводила взор от ребенка, сразу же возникала опасность его падения на пол.
Так, например, у женщины на одной руке была потеряна чувствительность, а на другой – способность к движению. Она могла держать ребенка на руке, потерявшей только чувствительность, до тех пор, пока на него смотрела. Как только женщина отводила взор от ребенка, сразу же возникала опасность его падения на пол.
Рефлексы спинного мозга широко использовались клиницистами, среди которых наиболее значительными фигурами были английский врач Маршал Холл и немецкий физиолог Иоганес Мюллер. Именно М. Холлу принадлежит термин «рефлекторная дуга». Согласно его учению, рефлекторная дуга состоит из афферентного нерва, спинного мозга и эфферентного нерва. М. Холл и И. Мюллер настаивали на принципиальном отличии работы спинного мозга от головного. По их мнению, рефлекторный механизм свойственен только спинному мозгу. Головной мозг оказывался все дальше от сферы влияния физиологии.
Следующий этап развития рефлекторной теории связан с работами великого русского физиолога И.М. Сеченова (1829-1905).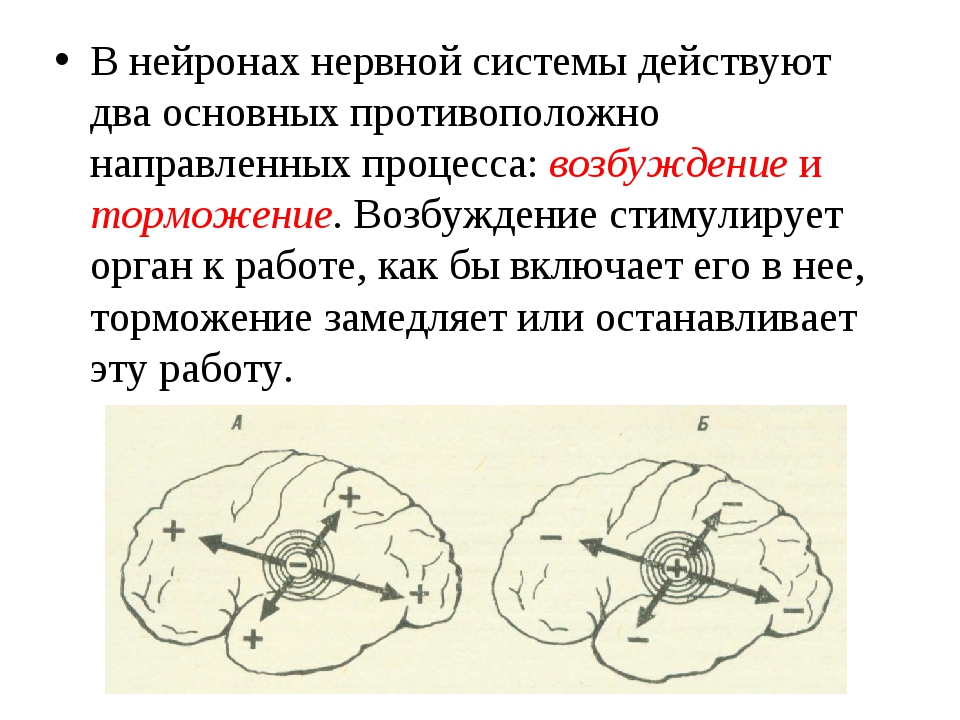 Само понятие о рефлекторном характере нервной деятельности у И.М. Сеченова претерпело существенные изменения. Основные положения его теории заключаются в следующем.
Само понятие о рефлекторном характере нервной деятельности у И.М. Сеченова претерпело существенные изменения. Основные положения его теории заключаются в следующем.
1. Рефлекс им понимался как универсальная и своеобразная форма взаимодействия организма со средой, опирающаяся на эволюционную биологию. И.М. Сеченов поставил вопрос о существовании двух родов рефлексов. Он выделил, во-первых, рефлексы постоянные врожденные, осуществляемые низшими отделами нервной системы, которые назвал «чистыми» рефлексами; во-вторых, рефлексы головного мозга изменчивые, приобретенные в индивидуальной жизни. Последние И.М. Сеченов представлял себе одновременно и физиологическим, и психическим явлением.
Таким образом, была впервые показана неотделимость психических процессов от мозга и одновременно обусловленность психики внешним миром. Важнейшим для И.М. Сеченова было положение о единстве организма и условий внешней среды. Большое значение в становлении рефлексов он придавал факторам эволюции.
2. Физиологический субстрат рефлекторных актов характеризуется как нейродинамика, отличная от динамики других систем. Открытие центрального торможения И.М. Сеченовым в 1862 году было первым шагом к созданию им новой физиологии головного мозга. Деятельность нервных центров мыслится ученым как непрерывная динамика процессов возбуждения и торможения.
3. На первый план ставятся межцентральные координационные отношения. Физиологическому анализу начинают подвергаться высшие мозговые центры. Если до И.М. Сеченова усиление или угнетение рефлекторных реакций трактовалось усилием воли, сознания, разума, то И.М. Сеченов переводит все это на строгий физиологический язык и показывает, как центры головного мозга могут задерживать или усиливать спинальные рефлексы.
4. Функции мозговых центров трактуются в широком плане биологической адаптации. Центры влияют усиливающим или тормозным образом на движения не потому, что высвобождается заложенная в них «психическая сила», и не потому, что укорачивается или удлиняется путь прохождения нервного импульса.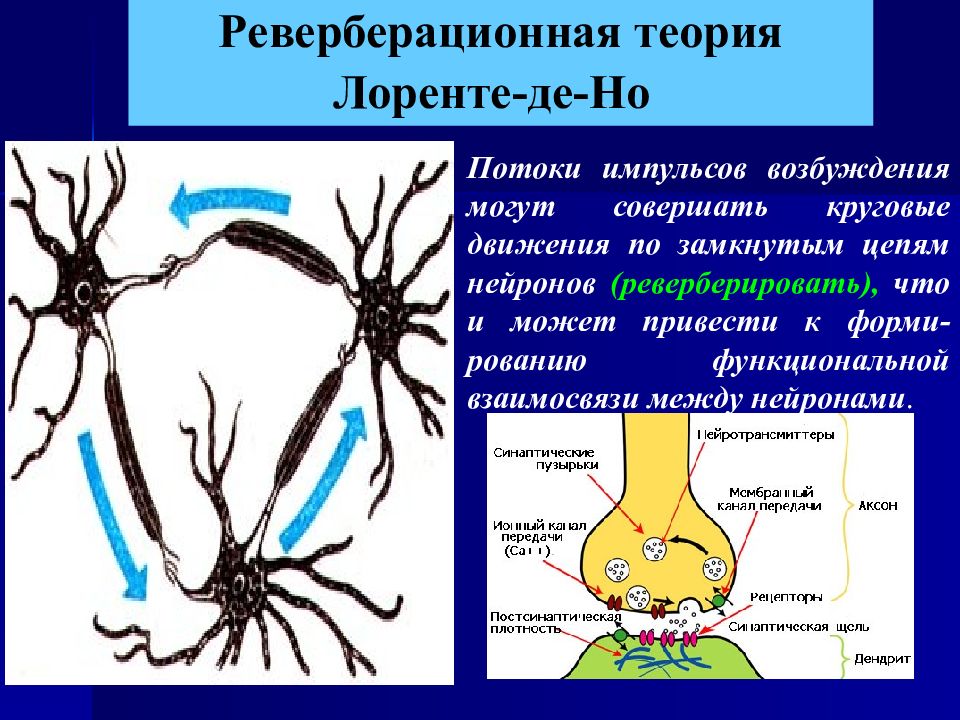 И.М. Сеченов вводит понятие «физиологического состояния центра», которое непосредственно связано с биологическими потребностями. Само же состояние центра, отражая характер отношений со средой, представляет собой нервный субстрат потребности. Таким образом, в учение о рефлексах вносится существенное дополнение. Реакция становится в непосредственную зависимость не только от наличных раздражений, но и от всей суммы прежних воздействий, оставивших длительно сохраняющиеся следы в нервных центрах.
И.М. Сеченов вводит понятие «физиологического состояния центра», которое непосредственно связано с биологическими потребностями. Само же состояние центра, отражая характер отношений со средой, представляет собой нервный субстрат потребности. Таким образом, в учение о рефлексах вносится существенное дополнение. Реакция становится в непосредственную зависимость не только от наличных раздражений, но и от всей суммы прежних воздействий, оставивших длительно сохраняющиеся следы в нервных центрах.
5. И.М. Сеченов считает, что мышечное чувство при выполнении одного движения становится в порядке ассоциации рефлексов сигналом для другого движения. Принцип ассоциации рефлексов лежит в основе обучения человека сложным формам трудовой деятельности. Устанавливается общий характер для движений и для психической деятельности – это наличие мышечной чувствительности.
По вопросу соотношения физиологического и психического И.М. Сеченов занял совершенно определенную позицию, которая отражена в его словах: «Для нас, как физиологов, достаточно и того, что мозг есть орган души, то есть такой живой механизм, который, будучи приведен какими ни на есть причинами в движение, дает в окончательном результате тот же ряд внешних явлений, которыми характеризуется психическая деятельность».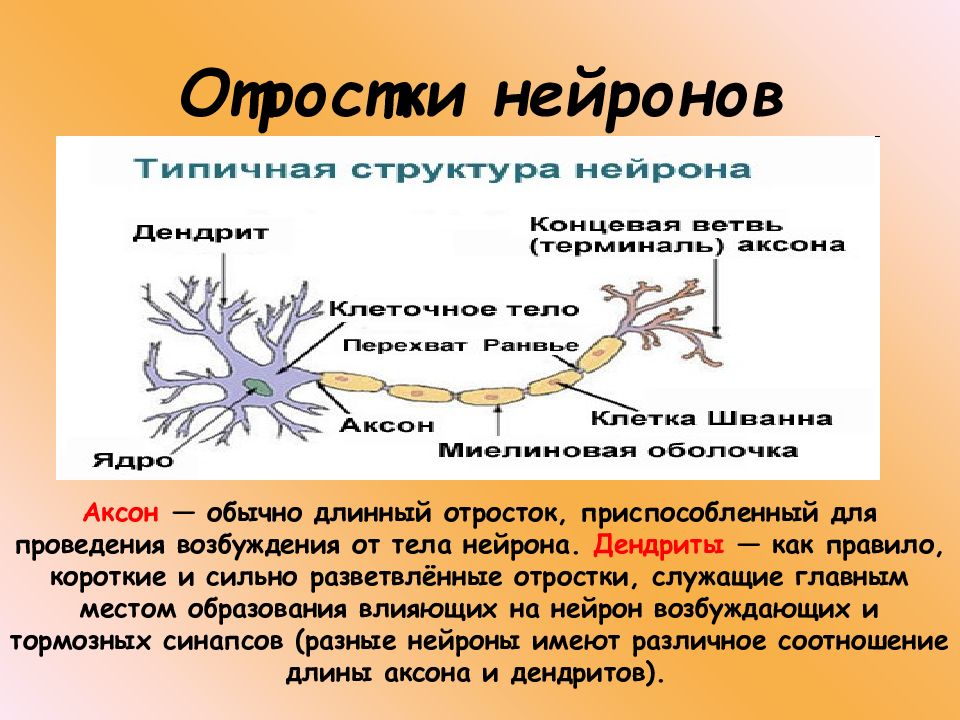
Для большей убедительности своих взглядов И.М. Сеченову не хватало экспериментального подтверждения.
Блестящие догадки, предвидения и мысли И.М. Сеченова экспериментально подкрепил И.П. Павлов. Он создал научную концепцию условного рефлекса, которая благодаря его блестящим экспериментам была введена в строгие рамки лабораторного опыта. Вот основные элементы рефлекторной теории И.П. Павлова.
1. Прежде всего был создан лабораторный метод объективного изучения приспособительной деятельности человека и животных – метод условных рефлексов.
2. Изучая условные рефлексы на целостном организме, И.П. Павлов подчеркивал их приспособительно-эволюционный смысл для животного мира.
3. И.П. Павлов предпринял попытку локализовать сам нервный процесс замыкания нервных связей в коре головного мозга у высших животных и человека. При этом он не исключал специфического участия других отделов мозга в этом процессе.
4. И.П. Павлов констатировал наличие в коре головного мозга процесса торможения, чем подкрепил сеченовские идеи о тормозящем влиянии головного мозга.
5. Было четко сформулировано учение о физиологии анализаторов, под которыми И.П. Павлов, как и И.М. Сеченов, мыслил конструкции, состоящие из периферических рецепторов, проводящих путей и мозговых центров, вплоть до коры больших полушарий.
6. И.П. Павлов сформировал представление о коре головного мозга как мозаике из возбуждений и торможений.
7. В конце творческой жизни И.П. Павлов выдвинул принцип системности в работе коры головного мозга, способной формировать динамический стереотип деятельности, уже в какой-то мере независимый от качества внешних раздражений.
Дальнейшее развитие рефлекторного принципа происходит в работах А.А. Ухтомского и П.К. Анохина.
А.А. Ухтомский подчеркивал: «… рефлекс – это такая реакция, которая мотивируется вполне отчетливо текущей обстановкой или средой. Этим, однако, не уничтожается спонтанное действие субстрата, оно лишь ставится в определенные границы в его противоположении факторам среды, и от этого оно становится более определенным по поддержанию и значению.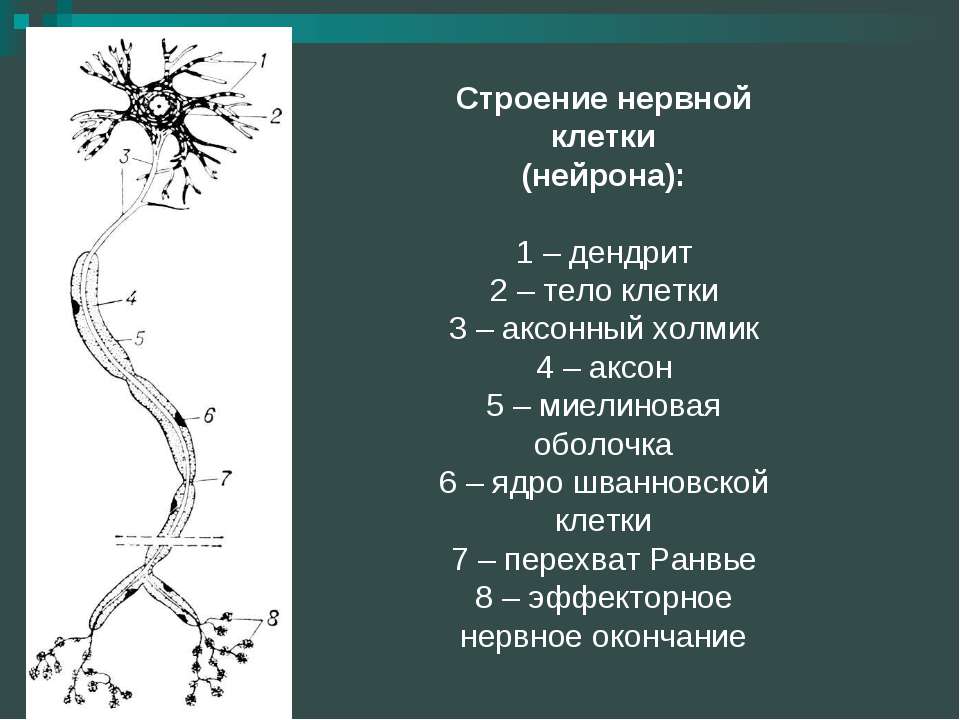 Рефлекс рисуется не чисто пассивным передвижением шарика под влиянием полученного им удара извне, так рефлекс мог изображаться, пока надо было подчеркнуть в особенности его мотивировку из среды. Но в полноте своей он представляется встречею во времени двух условий: с одной стороны, деятельности заготовленной, или сложившейся в самом субстрате (клетке) за предыдущую его историю, и, с другой стороны, внешних импульсов текущего момента». Принципы историзма и соотношения реактивности и активности в целостном поведении как способе разрешения противоречий, принесенные в физиологию А.А. Ухтомским, существенно обогатили рефлекторную теорию, которая окончательно избавилась от дуализма и механицизма картезианского толка, встав на твердые диалектические позиции.
Рефлекс рисуется не чисто пассивным передвижением шарика под влиянием полученного им удара извне, так рефлекс мог изображаться, пока надо было подчеркнуть в особенности его мотивировку из среды. Но в полноте своей он представляется встречею во времени двух условий: с одной стороны, деятельности заготовленной, или сложившейся в самом субстрате (клетке) за предыдущую его историю, и, с другой стороны, внешних импульсов текущего момента». Принципы историзма и соотношения реактивности и активности в целостном поведении как способе разрешения противоречий, принесенные в физиологию А.А. Ухтомским, существенно обогатили рефлекторную теорию, которая окончательно избавилась от дуализма и механицизма картезианского толка, встав на твердые диалектические позиции.
П.К. Анохин рассматривал поведение как системный процесс. В основе организации поведения, согласно П.К. Анохину, лежат функциональные системы. Созданием теории функциональных систем П.К. Анохин определил дальнейшее развитие рефлекторного принципа.
Принадлежит ведущая роль в обеспечении целостности организма, а также в его регуляции. Эти процессы осуществляются анатомо-физиологическим комплексом, включающим отделы ЦНС (центральной нервной системы). Он имеет свое название — нервный центр. Свойства, которыми он характеризуется: окклюзия, центральное облегчение, трансформация ритма. Они и некоторые другие будут изучены в данной статье.
Понятие нервного центра и его свойства
Ранее мы обозначили главную функцию нервной системы — интегрирующую. Она возможна благодаря структурам головного и спинного мозга. Например, дыхательный нервный центр, свойства которого — иннервация дыхательных движений (вдоха и выдоха). Он находится в четвертом желудочке, в области ретикулярной формации (продолговатый мозг). Согласно исследованиям Н. А. Миславского, он состоит из симметрично размещенных частей, ответственных за вдох и выдох.
В верхней зоне варолиевого моста находится пневмотаксический отдел, который регулирует вышеназванные части и структуры головного мозга, ответственные за дыхательные движения. Таким образом, общие свойства нервных центров обеспечивают регуляцию физиологических функций организма: сердечно-сосудистой деятельности, выделения, дыхания и пищеварения.
Таким образом, общие свойства нервных центров обеспечивают регуляцию физиологических функций организма: сердечно-сосудистой деятельности, выделения, дыхания и пищеварения.
Теория динамической локализации функций И. П. Павлова
Согласно воззрениям ученого, достаточно простые рефлекторные действия имеют в коре головного мозга, а также в спинном мозге стационарные зоны. Сложные процессы, такие как память, речь, мышление, связаны с определенными участками головного мозга и являются интегративным результатом функций многих его участков. Физиологические свойства нервных центров и обуславливают формирование основных процессов высшей нервной деятельности. В нейрологии, с анатомической точки зрения, участки центральной нервной системы, состоящие из афферентной и эфферентной частей нейронов, стали называть нервными центрами. Они, как считал российский ученый П. К. Анохин, образуют (объединение нейронов, выполняющие сходные функции и могущие находится в различных участках ЦНС).
Иррадиация возбуждения
Продолжая изучать основные свойства нервных центров, остановимся на форме распространения двух главных процессов, происходящих в нервной ткани — возбуждения и торможения. Он называется иррадиацией. Если сила раздражителя и время его действия велики, нервные импульсы расходятся по отросткам нейроцитов, а также по вставочным нейронам. Они объединяют афферентные и эфферентные нейроциты, обуславливая непрерывность рефлекторных дуг.
Он называется иррадиацией. Если сила раздражителя и время его действия велики, нервные импульсы расходятся по отросткам нейроцитов, а также по вставочным нейронам. Они объединяют афферентные и эфферентные нейроциты, обуславливая непрерывность рефлекторных дуг.
Рассмотрим торможение (как свойство нервных центров) более подробно. головного мозга обеспечивает как иррадиацию, так и другие свойства нервных центров. Физиология объясняет причины, ограничивающие или препятствующие распространению возбуждения. Например, наличие тормозных синапсов и нейроцитов. Эти структуры выполняют важные защитные функции, вследствие чего снижается риск перевозбуждения скелетной мускулатуры, способной перейти в судорожное состояние.
Рассмотрев иррадиацию возбуждения, нужно вспомнить следующую особенность нервного импульса. Он движется только от центростремительного нейрона к центробежному (для двухнейронной, рефлекторной дуги). Если рефлекс более сложный, то в головном или спинном мозге формируются интернейроны — вставочные нервные клетки.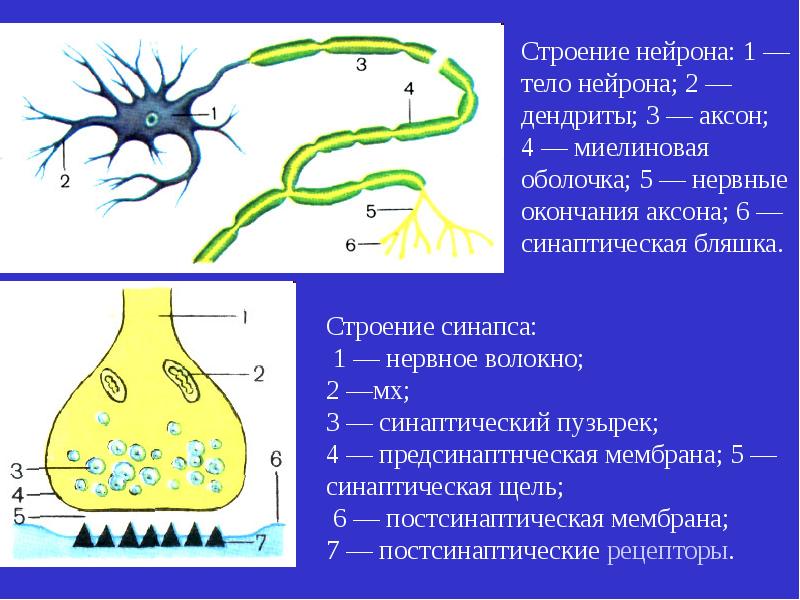 Они принимают возбуждение от афферентного нейроцита и далее передают его на двигательные нервные клетки. В синапсах биоэлектрические импульсы также однонаправленные: они движутся от пресинаптической мембраны первой нервной клетки, далее в синаптическую щель, а из неё — в постсинаптическую мембрану другого нейроцита.
Они принимают возбуждение от афферентного нейроцита и далее передают его на двигательные нервные клетки. В синапсах биоэлектрические импульсы также однонаправленные: они движутся от пресинаптической мембраны первой нервной клетки, далее в синаптическую щель, а из неё — в постсинаптическую мембрану другого нейроцита.
Суммация нервных импульсов
Продолжим изучать свойства нервных центров. Физиология главных отделов головного и спинного мозга, являясь наиболее важной и сложной отраслью медицины, изучает проведение возбуждения через совокупность нейронов, выполняющих общие функции. Их свойства — суммация, может быть временной или пространственной. В обоих случаях слабые нервные импульсы, вызванные подпороговыми раздражителями, складываются (суммируются). Это приводит к обильному выделению молекул ацетилхолина или другого нейромедиатора, что генерирует потенциал действия в нейроцитах.
Трансформация ритма
Этим термином обозначают изменение частоты возбуждения, которое проходит через комплексы нейронов ЦНС. Среди процессов, характеризующих свойства нервных центров — трансформация ритма импульсов, которая может возникать вследствие распределения возбуждения на несколько нейронов, длинные отростки которых формируют места контактов на одной нервной клетке (повышающая трансформация). Если же в нейроците появляется единичный потенциал действия, как результат суммации возбуждения постсинаптического потенциала — говорят о понижающей трансформации ритма.
Среди процессов, характеризующих свойства нервных центров — трансформация ритма импульсов, которая может возникать вследствие распределения возбуждения на несколько нейронов, длинные отростки которых формируют места контактов на одной нервной клетке (повышающая трансформация). Если же в нейроците появляется единичный потенциал действия, как результат суммации возбуждения постсинаптического потенциала — говорят о понижающей трансформации ритма.
Дивергенция и конвергенция возбуждения
Они являются взаимосвязанными процессами, характеризующими свойства нервных центров. Координация рефлекторной деятельности происходит благодаря тому, что в нейроцит одномоментно поступают импульсы от рецепторов различных анализаторов: зрительного, обонятельного и кожно-мышечной чувствительности. В нервной клетке они анализируются и суммируются в биоэлектрические потенциалы. Те, в свою очередь, передаются к другим участкам ретикулярной формации головного мозга. Этот важный процесс носит название конвергенции.
Однако каждый нейрон не только принимает импульсы от других клеток, но и сам образует синапсы с соседними нейроцитами. Это явление дивергенции. Оба свойства обеспечивают распространение возбуждения в ЦНС. Таким образом, совокупность нервных клеток головного и спинного мозга, выполняющих общие функции — это нервный центр, свойства которого мы рассматриваем. Он обеспечивает регуляцию работы всех органов и систем человеческого организма.
Фоновая активность
Физиологические свойства нервных центров, к одному из которых относится спонтанное, то есть фоновое образование электрических импульсов нейронами, например, дыхательного или пищеварительного центра, объясняются особенностями строения самой нервной ткани. Она способна к самогенерации биоэлектрических процессов возбуждения даже в период отсутствия адекватных раздражителей. Именно за счет дивергенции и конвергенции возбуждения, рассмотренных нами ранее, нейроциты получают импульсы от возбужденных нервных центров по постсинаптическим связям той же ретикулярной формации головного мозга.
Спонтанная активность может быть вызвана микродозами ацетилхолина, попадающего в нейроцит из синаптической щели. Конвергенция, дивергенция, фоновая активность, а также другие свойства нервного центра и их характеристика напрямую зависят от уровня обмена веществ как в нейроцитах, так и в нейроглии.
Виды суммации возбуждения
Они были рассмотрены в работах И. М. Сеченова, который доказал, что рефлекс можно вызвать несколькими слабыми (подпороговыми) раздражителями, которые достаточно часто действуют на нервный центр. Свойства его клеток, а именно: центральное облегчение и окклюзия, и будут рассмотрены нами далее.
При одновременном раздражении центростремительных отростков ответная реакция оказывается больше, чем арифметическая сумма силы раздражителей, действующих на каждое из этих волокон. Это свойство носит название центрального облегчения. Если же действие пессимальных раздражителей, независимо от их силы и частоты, вызывает снижение ответной реакции — это окклюзия. Она является обратным свойством суммации возбуждения и приводит к уменьшению силы нервных импульсов. Таким образом, свойства нервных центров — центральное облегчение, окклюзия — зависят от строения синаптического аппарата, состоящего из пороговой (центральной) зоны и подпороговой (периферической) каймы.
Она является обратным свойством суммации возбуждения и приводит к уменьшению силы нервных импульсов. Таким образом, свойства нервных центров — центральное облегчение, окклюзия — зависят от строения синаптического аппарата, состоящего из пороговой (центральной) зоны и подпороговой (периферической) каймы.
Утомляемость нервной ткани её роль
Физиология нервных центров, определение, виды и свойства, уже изученные нами ранее и присущие комплексам нейронов, будут неполными, если мы не рассмотрим такое явление, как утомляемость. Нервные центры вынуждены проводить через себя непрерывные серии импульсов, обеспечивая рефлекторные свойства центральных отделов нервной системы. В результате напряженных обменных процессов, осуществляемых как в самом теле нейрона, так и в глии, происходит накопление токсичных метаболических шлаков. Ухудшение кровоснабжения нервных комплексов также вызывает снижение их активности по причине дефицита кислорода и глюкозы. Свою лепту в развитие утомляемости нервных центров вносят также и места контактов нейронов — синапсы, которые быстро снижают выделение нейромедиаторов в синаптическую щель.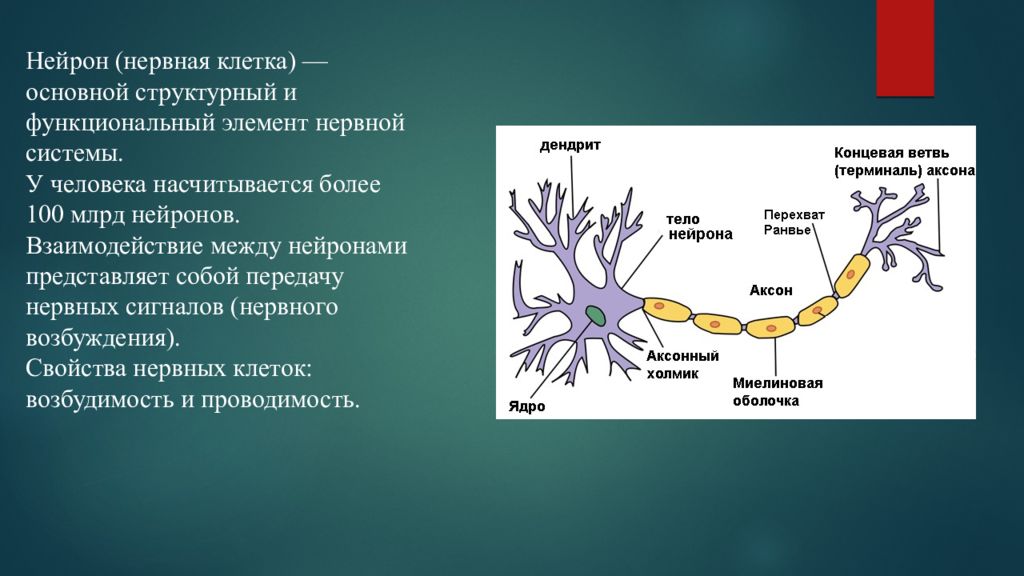
Генезис нервных центров
Комплексы нейроцитов, расположенные в и выполняющие координирующую роль в деятельности организма, претерпевают анатомо-физиологические изменения. Они объясняются усложнением физиологических и психологических функций, возникающих в течение жизни человека. Наиболее важные изменения, затрагивающие возрастные особенности свойств нервных центров, мы наблюдаем в становлении таких важных процессов, как прямохождение, речь и мышление, отличающие Homo sapiens от остальных представителей класса млекопитающих. Например, становление речи происходит в первые три года жизни ребенка. Являясь сложным конгломератом условных рефлексов, она формируется на базе раздражений, воспринимаемых проприорецепторами мышц языка, губ, голосовых связок гортани и дыхательной мускулатуры. К концу третьего года жизни ребенка все они объединяются в функциональную систему, в которую входит участок коры, лежащий в основании нижней лобной извилины. Он был назван центром Брока.
В формировании принимает участие и зона верхней височной извилины (центр Вернике).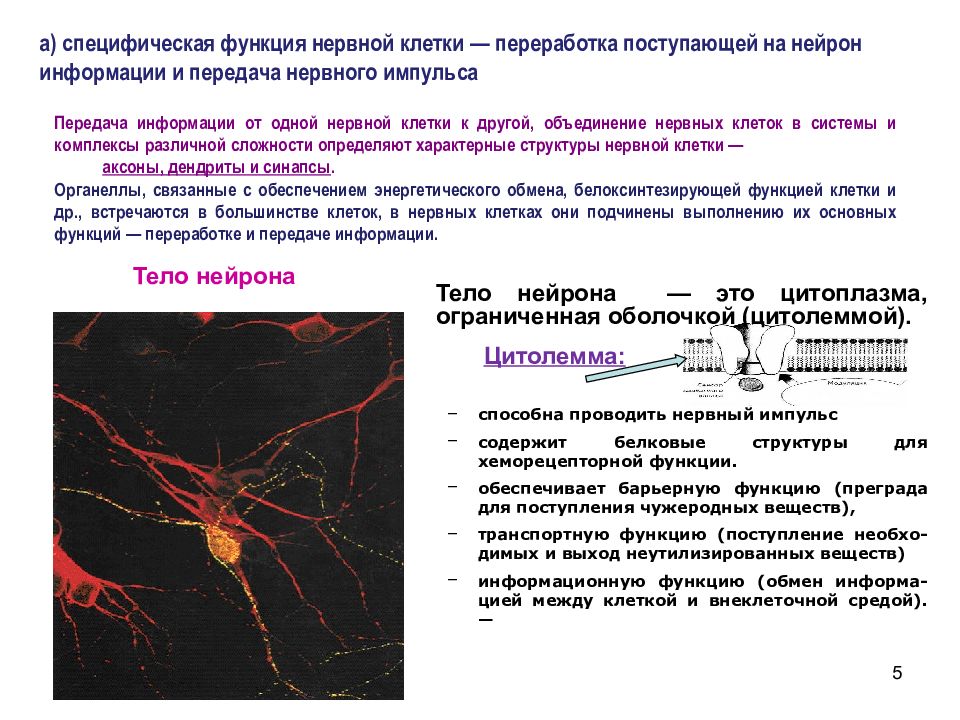 Возбуждение от нервных окончаний речевого аппарата поступает в двигательный, зрительный и слуховой центры коры головного мозга, где и формируются центры речи.
Возбуждение от нервных окончаний речевого аппарата поступает в двигательный, зрительный и слуховой центры коры головного мозга, где и формируются центры речи.
Одностороннее проведение возбуждения. В нервных центрах прохождение импульсов возбуждения по цепи нейронов осуществляется только в одном направлении: от чувствительного нерва через промежуточные к двигательному, а от двигательного — к органу-исполнителю. Это обусловлено односторонней синаптической передачей импульсов от одной нервной клетки к другой с помощью медиатора, который выделяется концевым аппаратом аксона и содержится лишь в пресинаптической щели. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление. Это свойство обеспечивает координирующую роль центральной нервной системы и способствует замыкательной функции условно-рефлекторных связей.
Замедление проведения возбуждения. Это свойство называется центральной задержкой, или латентным (скрытым) периодом рефлекса. Центральная задержка обусловлена более медленным проведением нервных импульсов через синапсы.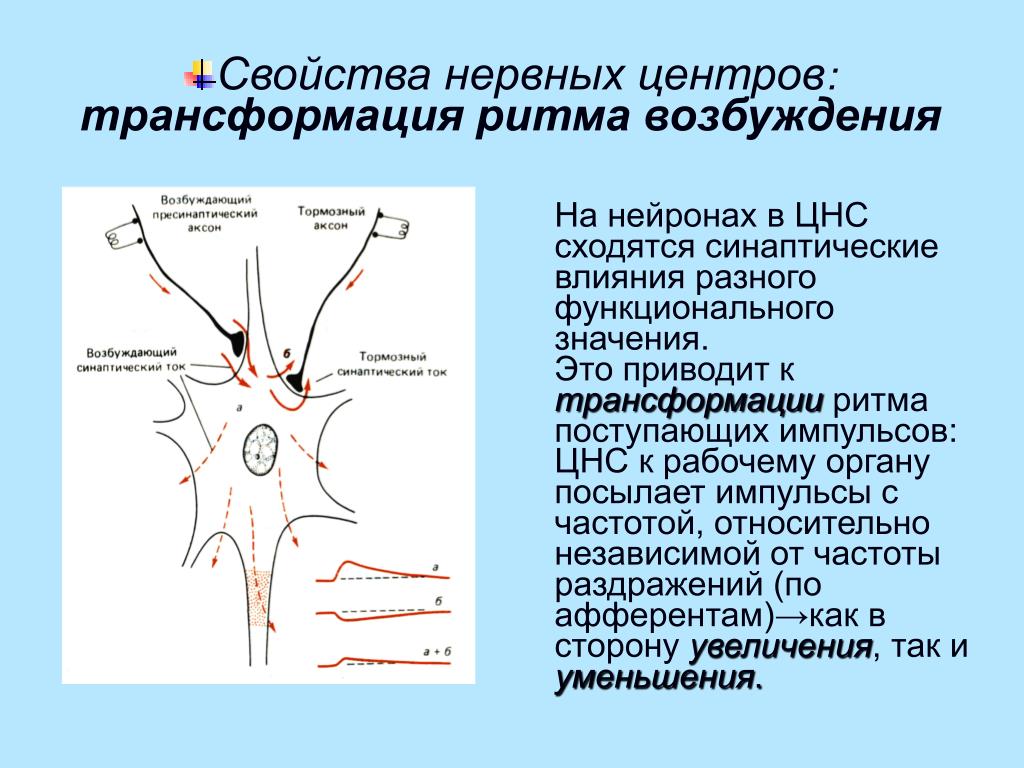
Интервал от начала раздражения рецептора до появления ответной реакции — 0,2–0,5 секунды. Чем сложнее рефлекс, тем длительнее центральная задержка. Дрессировщику необходимо учитывать скрытый период при выработке условного рефлекса и подкрепляющий раздражитель надо применять не раньше 0,5 секунды после воздействия сигнального раздражителя. Скрытый период рефлекса увеличивается при утомлении и заболевании животного.
Последействие. Присуще всем нервным центрам и характеризуется тем, что на короткое раздражение проявляется длительная ответная реакция, т. е. рефлекторный акт длится еще некоторое время после того, как раздражение рецептора прекращено. Такое последействие объясняется тем, что нервные импульсы от рецепторов поступают к двигательным центрам по различным нервным путям неодновременно: по коротким быстрее, чем по длинным. Запаздывающие импульсы поддерживают возбужденное состояние соответствующего нервного центра. Остаточное возбуждение в нервных центрах может сохраняться до 2 секунд и тем самым способствовать лучшей замыкательной функции при образовании условных рефлексов.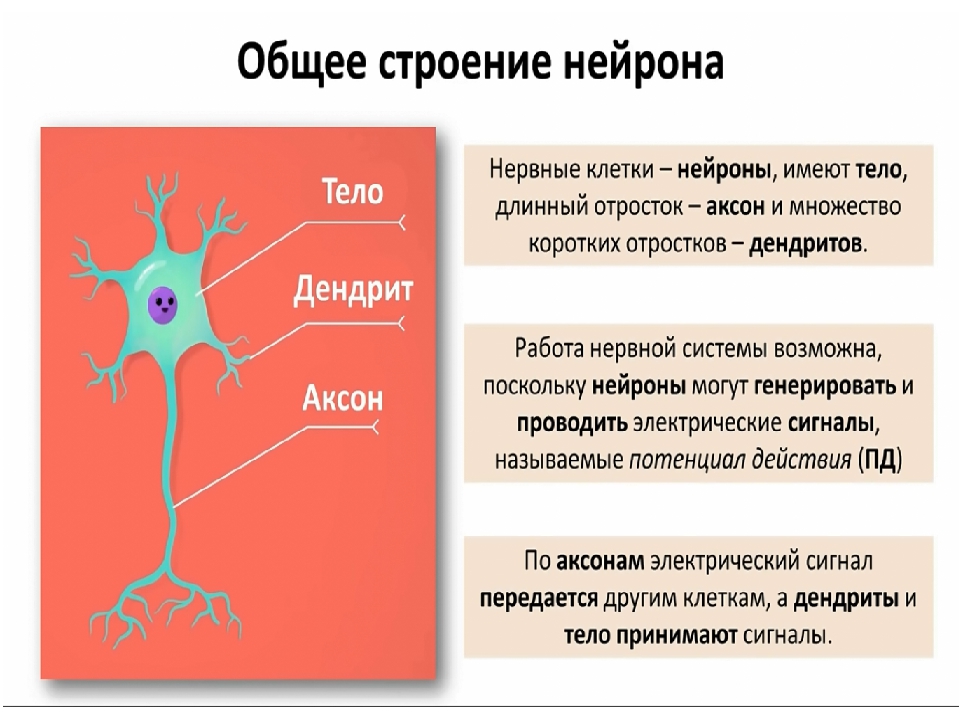 Кроме того, в практике дрессировки нужно помнить, что после окончания действия любого раздражителя требуется некоторое время для спада остаточного возбуждения нервного центра и освобождения рефлекторных путей для новой рефлекторной деятельности.
Кроме того, в практике дрессировки нужно помнить, что после окончания действия любого раздражителя требуется некоторое время для спада остаточного возбуждения нервного центра и освобождения рефлекторных путей для новой рефлекторной деятельности.
Суммация. Происходит путем накопления слабых допороговых раздражений до критического потенциала, способного вызвать возбуждение нервного центра. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного воздействия раздражителей допороговой силы на несколько чувствительных нейронов. Поступившие импульсы слабой силы суммируются в нервном центре и вызывают возбуждение. Суммация во времени происходит при активизации одного и того же чувствительного нейрона серией последовательных раздражений допороговой силы. Слабые импульсы от предыдущих раздражений накладываются друг на друга, суммируются и вызывают рефлекс. Суммация в пространстве и во времени в нервных центрах происходит одновременно. Они дополняют и усиливают друг друга и, например, увеличивают обонятельную и слуховую чувствительность у собаки, что так необходимо при работе по чутью, когда исключительно слабые раздражения запаховых частиц (1 молекула в одном литре воздуха) вызывают запаховые ощущения и соответствующие ответные реакции.
Трансформация. Свойство нервных центров изменять частоту и силу передающихся импульсов, т. е. трансформировать. Проявляется в активизации и перестройке нейронов на более высокий или более низкий ритм и изменении их лабильности, обеспечивает взаимодействие и установление связи между различными нервными центрами и другими отделами нервной системы, что имеет важное значение в замыкании условно-рефлекторных связей при дрессировке собак.
Облегчение. Свойство нервных центров обеспечивать высокую возбудимость и эффективность рефлекторной деятельности нервной системы. Сущность облегчения заключается в том, что после каждого раздражения в нервном центре повышается возбудимость к повторным раздражениям, следующим через небольшие промежутки времени. Один поток импульсов как бы облегчает действие другого и способствует образованию условного рефлекса.
Проторение. Способность одних нервных центров повышать возбудимость других и вступать во взаимодействие через обмен и передачу импульсов возбуждения между центрами по нейронным путям.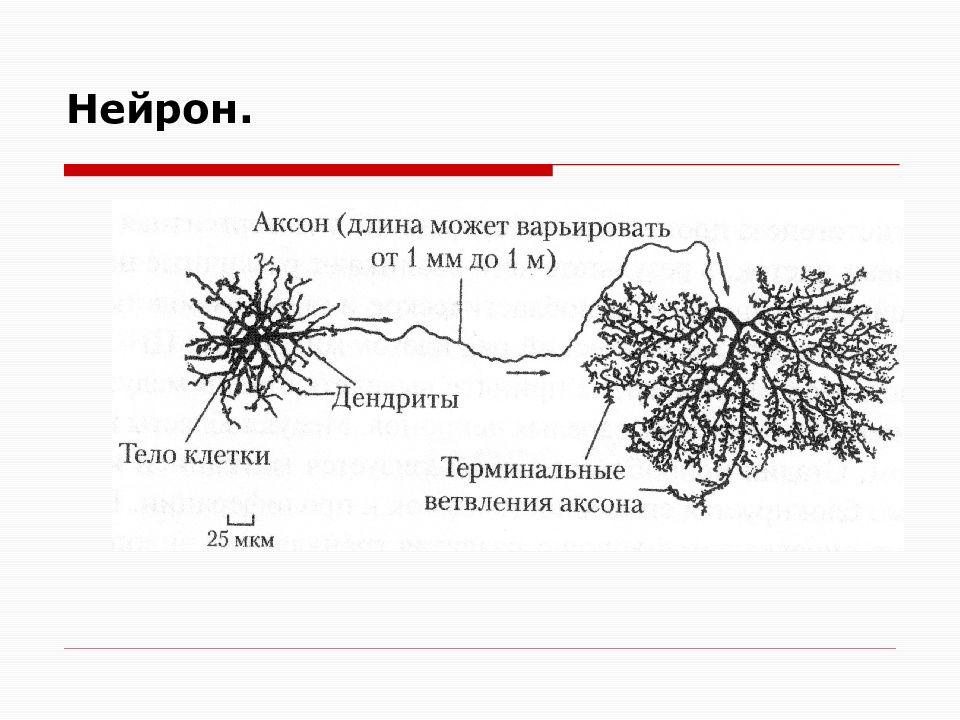 Процесс образования временной связи рассматривается как результат взаимодействия двух очагов возбуждения в коре, следствием чего является проторение пути между этими корковыми пунктами. Проходимость пути обусловлена функциональными и морфологическими изменениями в синапсах.
Процесс образования временной связи рассматривается как результат взаимодействия двух очагов возбуждения в коре, следствием чего является проторение пути между этими корковыми пунктами. Проходимость пути обусловлена функциональными и морфологическими изменениями в синапсах.
Иррадиация возбуждения. При сильном и длительном раздражении импульсы, поступившие в нервную систему, не ограничиваются возбуждением одного рефлекторного центра, а распространяются на другие центры. Чем сильнее раздражение, тем больше нервных центров охватывает иррадиация. Возбуждение большого количества различных нервных центров позволяет отобрать из них наиболее нужные и установить между ними новые функциональные связи — условные рефлексы. Большинство двигательных условных рефлексов формируется благодаря иррадиации возбуждения. Излишняя иррадиация возбуждения центральной нервной системы нарушает уравновешенность поведенческих реакций, приводит к расстройству рефлекторной деятельности. Иррадиация возбуждения ограничивается и уравновешивается торможением.
Торможение. Тормозной процесс — необходимое условие в координации нервной деятельности. Торможение возникает в определенных нервных структурах под влиянием волны возбуждения, которая подавляет другое возбуждение. При торможении выключается деятельность ненужных в данный момент органов и предохраняется от чрезмерного перенапряжения работа нервных центров. В нервной системе вырабатываются тормозные условные рефлексы: прекращения нежелательных действий, выдержки, дифференцировки и др.
Доминанта. Преобладание активности одних нервных центров над активностью других.
Доминирующий (господствующий) очаг возникает при определенном функциональном состоянии нервных центров. Одно из условий его образования — повышенная возбудимость нервных клеток одного или нескольких нервных центров под влиянием определенных нервных и гуморальных факторов. Возбудимость доминирующего очага усиливается суммированием импульсов, поступающих из других нервных центров, рефлекторная деятельность которых подавляется, а рефлекторная деятельность доминирующего очага заметно усиливается. Установившееся доминирование при инстинктах может быть продолжительным состоянием, которое определяет поведение животного на тот или иной срок. Доминанту можно наблюдать и в условно-рефлекторной деятельности. Доминирующий очаг возбуждения в коре головного мозга притягивает к себе импульсы нервных центров других очагов возбуждения и тем самым способствует суммации, облегчению, проторению и замыканию условно-рефлекторных связей на сигналы дрессировщика. Доминанта по теории академика И. П. Павлова имеет главенствующее значение в механизме образования и проявления условных рефлексов. У собаки всегда сильно проявляются стойко выработанные навыки, доминирующие над другими рефлексами и обеспечивающие безотказное управление собакой при отвлекающих раздражителям. Дрессировщику необходимо учитывать свойство доминанты при подготовке собак.
Установившееся доминирование при инстинктах может быть продолжительным состоянием, которое определяет поведение животного на тот или иной срок. Доминанту можно наблюдать и в условно-рефлекторной деятельности. Доминирующий очаг возбуждения в коре головного мозга притягивает к себе импульсы нервных центров других очагов возбуждения и тем самым способствует суммации, облегчению, проторению и замыканию условно-рефлекторных связей на сигналы дрессировщика. Доминанта по теории академика И. П. Павлова имеет главенствующее значение в механизме образования и проявления условных рефлексов. У собаки всегда сильно проявляются стойко выработанные навыки, доминирующие над другими рефлексами и обеспечивающие безотказное управление собакой при отвлекающих раздражителям. Дрессировщику необходимо учитывать свойство доминанты при подготовке собак.
Конвергенция (сближение). Схождение поступающих по чувствительным путям импульсов возбуждения в одном промежуточном или двигательном центре. В центральной нервной системе в 4–5 раз больше чувствительных путей, чем двигательных. Поэтому к одному и тому же двигательному центру могут подходить импульсы возбуждения по многим путям. Эта особенность прохождения возбуждения по нервным центрам противоположна иррадиации и является основой для концентрации возбуждения в отдельных пунктах коры головного мозга. Конвергенция обеспечивает специализацию условного рефлекса и формирование навыка на сложные и комплексные раздражители.
Поэтому к одному и тому же двигательному центру могут подходить импульсы возбуждения по многим путям. Эта особенность прохождения возбуждения по нервным центрам противоположна иррадиации и является основой для концентрации возбуждения в отдельных пунктах коры головного мозга. Конвергенция обеспечивает специализацию условного рефлекса и формирование навыка на сложные и комплексные раздражители.
Окклюзия (закупорка). Проявляется при сочетании особенно сильных раздражителей, дающих эффект меньше суммы величин этих реакций на каждый раздражитель в отдельности. Это свойство противоположно суммации. Окклюзия и пространственная суммация постоянно взаимодействуют между собой. При слабом возбуждении проявляется суммация, при сильных раздражениях — окклюзия. В дрессировке собак ошибочно применение сильных раздражителей для ускорения первоначального условного рефлекса. На громкие команды и сильные подкрепления вырабатываются, как правило, слабые условные рефлексы.
Пластичность (податливость). Способность перестройки функций нервных центров. Сформировавшиеся в процессе эволюции рефлекторные акты, реакции поведения и инстинкты могут перестраиваться, а нервные центры изменять свои функции. Опытами установлено, что перестройка функций нервных центров у животных происходит под регулирующим влиянием коры полушарий головного мозга. Благодаря этому свойству при изменении условий жизни меняется поведение животных, их привычки и навыки: собака и кошка могут мирно уживаться, инстинкт преследования собакой диких животных по следам заменяется реакцией поиска человека по его запаховому следу, вредные привычки, нежелательные связи у собаки могут быть исправлены систематической дрессировкой. Пластичность нервных центров позволяет при дрессировке вырабатывать сложные навыки в виде динамических стереотипов.
Способность перестройки функций нервных центров. Сформировавшиеся в процессе эволюции рефлекторные акты, реакции поведения и инстинкты могут перестраиваться, а нервные центры изменять свои функции. Опытами установлено, что перестройка функций нервных центров у животных происходит под регулирующим влиянием коры полушарий головного мозга. Благодаря этому свойству при изменении условий жизни меняется поведение животных, их привычки и навыки: собака и кошка могут мирно уживаться, инстинкт преследования собакой диких животных по следам заменяется реакцией поиска человека по его запаховому следу, вредные привычки, нежелательные связи у собаки могут быть исправлены систематической дрессировкой. Пластичность нервных центров позволяет при дрессировке вырабатывать сложные навыки в виде динамических стереотипов.
Инертность. Нервные центры обладают свойством переходить в состояние возбуждения только при относительно длительном раздражении. Возбудившись, они сохраняют это состояние возбуждения в течение определенного времени. Такое явление И. П. Павлов назвал инертностью. Способность нервных центров длительно сохранять в себе следы возбуждения и торможения, сильно выражена в нервных клетках коры полушарий головного мозга. И. П. Павлов говорил, что если бы у нервных клеток не было инертности, у нас не было бы никакой памяти, никакой выучки, не существовало бы никаких привычек. У животных существует два вида памяти: кратковременная и долговременная. Оба вида памяти обеспечивают возможность выработки у животных условных рефлексов и формирования стойких навыков. Кратковременная память у собаки проявляется в течение нескольких минут, долговременная, или долгосрочная, память — через много дней, месяцев и даже лет. Оба вида памяти у животных выражаются в репродукции образа воспринимаемых отдельных раздражителей или целостного объекта.
Такое явление И. П. Павлов назвал инертностью. Способность нервных центров длительно сохранять в себе следы возбуждения и торможения, сильно выражена в нервных клетках коры полушарий головного мозга. И. П. Павлов говорил, что если бы у нервных клеток не было инертности, у нас не было бы никакой памяти, никакой выучки, не существовало бы никаких привычек. У животных существует два вида памяти: кратковременная и долговременная. Оба вида памяти обеспечивают возможность выработки у животных условных рефлексов и формирования стойких навыков. Кратковременная память у собаки проявляется в течение нескольких минут, долговременная, или долгосрочная, память — через много дней, месяцев и даже лет. Оба вида памяти у животных выражаются в репродукции образа воспринимаемых отдельных раздражителей или целостного объекта.
Тонус. Нервные центры обладают свойством постоянно находиться в состоянии незначительного возбуждения при относительном рефлекторном покое. Они постоянно посылают импульсы, обеспечивающие тоническое сокращение скелетной мускулатуры. Тонус нервных центров поддерживается действием гуморальных веществ и непрерывным потоком импульсов, поступающих от рецепторов. Огромное значение в поддержании мышечного тонуса имеют нервные центры продолговатого, среднего и промежуточного мозга. Тоническое состояние нервных центров и мышечной системы обеспечивает устойчивую выработку условных рефлексов и хорошую работоспособность собаки. Состояние рабочего тонуса поддерживается правильно организованной дрессировкой, систематической тренировкой и регулярным использованием собаки на службе.
Тонус нервных центров поддерживается действием гуморальных веществ и непрерывным потоком импульсов, поступающих от рецепторов. Огромное значение в поддержании мышечного тонуса имеют нервные центры продолговатого, среднего и промежуточного мозга. Тоническое состояние нервных центров и мышечной системы обеспечивает устойчивую выработку условных рефлексов и хорошую работоспособность собаки. Состояние рабочего тонуса поддерживается правильно организованной дрессировкой, систематической тренировкой и регулярным использованием собаки на службе.
Перечисленные свойства нервных центров обеспечивают их функциональное назначение в нервной системе и организме.
Из книги Арасланов Филимон, Алексеев Алексей, Шигорин Валерий «Дрессировка собак»
Общая физиология нервной системы Лекция 2
Общая физиология нервной системы Лекция 2
• Нейроны устанавливают связи между собой через синапсы. • Синапсы бывают возбуждающие и тормозные. • Нейрон возбуждается, если возбуждающие влияния преобладают. • Нейрон тормозится, если преобладают тормозные влияния. • Нейрон находится в покое, если возбуждающие влияния уравновешены тормозными. • Покой — нейрон готов к принятию информации и ответу на неё. • Возбуждение – нейрон активен, генерирует электрические импульсы и передает их. • Торможение — нейрон угнетён, малочувствителен или нечувствителен к приходящей информации.
• Нейрон возбуждается, если возбуждающие влияния преобладают. • Нейрон тормозится, если преобладают тормозные влияния. • Нейрон находится в покое, если возбуждающие влияния уравновешены тормозными. • Покой — нейрон готов к принятию информации и ответу на неё. • Возбуждение – нейрон активен, генерирует электрические импульсы и передает их. • Торможение — нейрон угнетён, малочувствителен или нечувствителен к приходящей информации.
Кодирование информации нейроном Электронейронография
Нервная система – целостная морфологически и функционально совокупность структур организма, образованных нервной тканью и выполняющая следующие функции:
Основные функции нервной системы • Получение, хранение и переработка информации из внешней и внутренней среды. • Регуляция и координация деятельности всех органов и систем органов. • Обеспечение взаимодействия организма с внешней средой.
Нервная система Центральная üголовной мозг ü спинной мозг Периферическая üчувствительные и двигательные нервы; üчувствительные и вегетативные ганглии; üнервные сплетения.
Расхождение сигналов от одной клетки к нескольким
КОНВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ Схождение возбуждения к одной нервной клетке
Нервный центр – объединение нейронов, расположенных на различных уровнях центральной нервной системы и участвующих в регуляции определённой функции организма.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ 1. Одностороннее проведение 2. Центральная задержка рефлекса 3. Тонус нервного центра 4. Высокая утомляемость 5. Высокая химическая чувствительность
Классификация нервных центров • По общему функциональному значению: чувствительные, двигательные, вегетативные • По расположению: спинальные, бульбарные, мезенцефальные, таламические, гипоталамические, подкорковые, корковые • По регулируемой функции: зрительный, дыхательный, глотания, сгибания, разгибания, слюноотделения и т. д.
Основной принцип работы мозга рефлекторный Refleksus – отражённый Рефлекс – стереотипная ответная реакция организма на раздражение рецепторов, осуществляемая с участием ЦНС
Структурной основой рефлекса является рефлекторная дуга – путь, который проходит возбуждение при осуществлении рефлекса.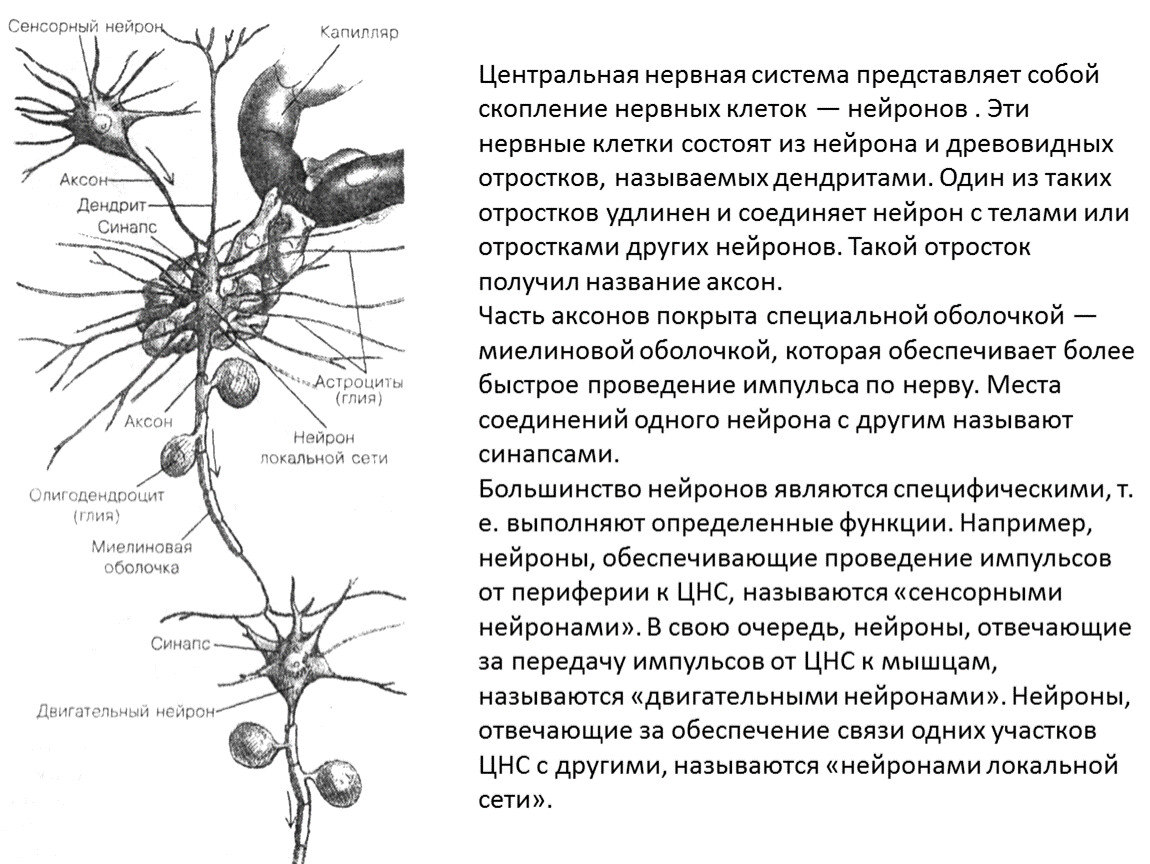 Рефлекторная дуга состоит из 5 звеньев: 1. Рецептор 2. Афферентный путь 3. Нервный центр 4. Эфферентный путь 5. Эффектор
Рефлекторная дуга состоит из 5 звеньев: 1. Рецептор 2. Афферентный путь 3. Нервный центр 4. Эфферентный путь 5. Эффектор
Простейшая рефлекторная дуга
Сложная рефлекторная дуга
Сложные рефлекторные дуги
• Для осуществления рефлекса необходимы анатомическая и функциональная целостность рефлекторной дуги • Время рефлекса (латентное) – время от начала раздражения до появления ответной реакции • Чем сильнее раздражитель и/или чем больше площадь раздражения, тем меньше время рефлекса.
Принципы классификации рефлексов 1. По происхождению – безусловные и условные. 2. По биологическому значению: пищевые, половые, защитные, локомоторные и т. д. 3. По виду рецепторов: зрительные, обонятельные и тд. 4. По месту расположения центра: спинальные, бульбарные, мезенцефальные, диенцефальные, корковые. 5. По характеру ответной реакции: двигательные и секреторные 6.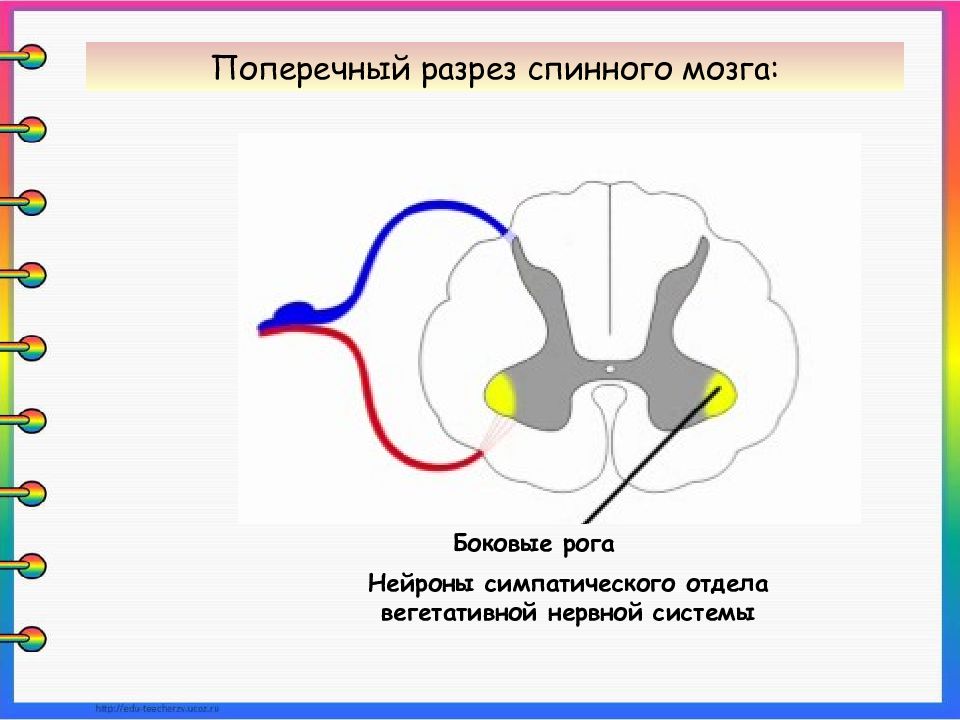 По принадлежности к системе органов: дыхательные, пищеварительные, сердечнососудистые и тд.
По принадлежности к системе органов: дыхательные, пищеварительные, сердечнососудистые и тд.
Рефлекторная деятельность мозга осуществляется по принципу ОБРАТНОЙ АФФЕРЕНТАЦИИ ОБРАТНАЯ АФФЕРЕНТАЦИЯ ДВИГАТЕЛЬНАЯ КОМАНДА
Впервые доказал, что в ЦНС наряду с процессами возбуждения протекают процессы торможения Сеченов И. М. (1829 -1905)
ФИЗИОЛОГИЯ ТОРМОЖЕНИЯ Торможение – это активный процесс, который препятствует возникновению и распространению возбуждения
Опыт Сеченова
В Зрительных буграх есть нервные центры, которые тормозят рефлексы спинного мозга
Торможение • Является результатом возбуждения специальных тормозных нейронов • Тормозные нейроны образуют тормозные синапсы, в которых выделяются медиаторы, оказывающие тормозное влияние на другие нейроны (ТПСП)
Чарльз Шеррингтон (1857 -1952): • В любом рефлекторном акте участвуют как процессы возбуждения, так и процессы торможения • Рефлекторный акт возможен только при сопряжённом (реципрокном) торможении мышц-антагонистов
Сопряжённое торможение центровантагонистов
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А. А. Ухтомскому, 1931) Д о м и н а н т а — временно господствующий рефлекс или поведенческий акт, который в данное время трансформирует и направляет работу прочих рефлекторных дуг, рефлекторного аппарата и поведения в целом
А. Ухтомскому, 1931) Д о м и н а н т а — временно господствующий рефлекс или поведенческий акт, который в данное время трансформирует и направляет работу прочих рефлекторных дуг, рефлекторного аппарата и поведения в целом
Принцип общего конечного пути в спинном мозге Множество разнообразных раздражении возбуждает одни и те же мотонейроны спинного мозга
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
Общая физиология ЦНС. Свойства нервных центров
1. Общая физиология ЦНС. Свойства нервных центров
Кафедра специальной психологии КГПУк.м.н., доц. Бардецкая Я.В.
2. НЕРВНЫЙ ЦЕНТР
• Нервный центр → функционально связаннаясовокупность нейронных ансамблей разных
этажей нервной системы, обеспечивающих
регуляцию определенных функций организма.
• Такое объединение (нервный центр в широком
смысле слова) позволяет осуществлять
наиболее адекватный для конкретных условий
принцип рефлекторной деятельности.
• Нервный центр → совокупность нейронов,
обеспечивающих реализацию определенного
рефлекса (мигания, глотания, кашля и т.д.).
3. ТИПЫ НЕЙРОНОВ НЕРВНОГО ЦЕНТРА
СенсорныеИнтернейроны
Моторные
ВОЗБУЖДАЮЩИЕ
ДВИГАТЕЛЬНЫЕ
МОНОМОДАЛЬНЫЕ
ТОРМОЗНЫЕ
СЕКРЕТОРНЫЕ
ПОЛИМОДАЛЬНЫЕ
МОДУЛИРУЮЩИЕ
ТРОФИЧЕСКИЕ
1. МОНОСЕНСОРНЫЕ
2. БИСЕНСОРНЫЕ
3. ПОЛИСЕНСОРНЫЕ
• Моносенсорные нейроны. Располагаются чаще в
первичных проекционных зонах коры и реагируют
только на сигналы своей сенсорности. Например,
значительная часть нейронов первичной зоны
зрительной области коры большого мозга реагирует
только на световое раздражение сетчатки глаза.
• Моносенсорные нейроны подразделяют
функционально по их чувствительности к разным
качествам одного раздражителя.
• Так, отдельные нейроны слуховой зоны коры
большого мозга могут реагировать на предъявления
тона 1000 Гц и не реагировать на тоны другой частоты.

Они называются мономодальными.
• Нейроны, реагирующие на два разных тона,
называются бимодальными, на три и более —
полимодальными.
• Бисенсорные нейроны. Чаще располагаются во
вторичных зонах коры какого-либо
анализатора и могут реагировать на сигналы
как своей, так и другой сенсорности.
• Например, нейроны вторичной зоны
зрительной области коры большого мозга
реагируют на зрительные и слуховые
раздражения.
• Полисенсорные нейроны. Это чаще всего
нейроны ассоциативных зон мозга;
• Способны реагировать на раздражение
слуховой, зрительной, кожной и других
рецептивных систем.
ПРИНЦИП СУБОРДИНАЦИИ
НЕРВНЫХ ЦЕНТРОВ
• Чем позже созревает отдел ЦНС в
процессе филогенетического
развития или в процессе онтогенеза,
тем сложнее и многообразнее его
строение и функции, и тем
отчетливее проявляется по
отношению к другим отделам НС его
руководящая, субординирующая
роль.

Нервные центры обладают рядом
характерных свойств,
определяемых:
• свойствами составляющих его
нейронов;
• особенностями синаптической
передачи нервных импульсов;
• структурой нейронных цепей,
образующих этот центр.
8. Основные свойства
1. Возбуждение в нервных центрахраспространяется односторонне — от рецептора
к эффектору, что обусловливается свойством
химических синапсов односторонне проводить
возбуждение от пресинаптической мембраны к
постсинаптической.
2. Возбуждение в нервных центрах проводится
медленнее, чем по нервному волокну. Это
обусловлено замедленным проведением
возбуждения через синапс (синаптическая
задержка).
3. В нервных центрах осуществляется суммация
возбуждений.
9. Свойства нервных центров
• Одностороннее проведение внервных центрах можно доказать при
раздражении передних корешков и
отведении потенциалов от задних. В
этом случае осциллограф не
зарегистрирует импульсов.
 Если
Еслипоменять электроды — импульсы будут
поступать нормально.
10. Одностороннее проведение в нервной цепи
• Дивергенция – это контактирование одногонейрона или нервного центра с множеством других.
• Так, существует разделение аксона чувствительного
нейрона в спинном мозге на множество коллатералей.
Получившиеся разветвления направляются к разным
сегментам спинного мозга и в головной мозг.
• Дивергенция пути сигнала наблюдается у многих
вставочных нейронов. Благодаря дивергенции могут
формироваться процессы параллельных вычислений,
что обеспечивает высокий уровень быстродействия
ЦНС.
• Дивергенция пути обеспечивает расширение области
распространения сигнала. Тем самым формируется
распространение процесса возбуждения на другие
нервные центры, то есть иррадиация возбуждения или
торможения.
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
Рецептор
• Конвергенция – это схождение нескольких нервных
путей к одним и тем же нейронам или нервным
центрам.

• Конвергенция многих нервных путей к одному
нейрону или центру делает его интегратором
соответствующих сигналов.
• Его состояние (импульсация или торможение) в
каждый момент времени определяется
алгебраическим сложением массы возбуждающих и
тормозных входов. Иными словами, суммой всех его
ВПСП и ТПСП, поступающих на данный центр или
нейрон.
• Мощная конвергенция обнаруживается на
мотонейронах спинного мозга и нейронах
ретикулярной формации ствола мозга.
• Благодаря конвергенции происходит «сжатие»,
уменьшение объема информации, поступающей от
рецепторов к ЦНС.
14. КОНВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ
Реверберация• Итальянский физиолог Лоренто де Но обнаружил
наличие в ЦНС замкнутых цепочек нейронов. Попав
в такую цепочку, импульс может минутами или
часами пробегать небольшой отрезок нейронной
цепи, модифицируя при этом синапсы.
• Сигнал может реверберировать до тех пор, пока или
какой-то внешний тормоз не выключит одно из
звеньев цепи, или в ней не наступит утомление.

• Выход из такой цепочки осуществляется по
коллатералям аксонов нейронов – участников цепи.
• Таким образом, при реверберации в цепочке
нейронов «хранится» тот сигнал, который включил в
ней циркуляцию (реверберацию) импульсов.
Считается, что реверберация может
служить основой кратковременной памяти.
16. РЕВЕРБЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СЕТИ по Лоренто-де-Но
17. Виды связей между нейронами
Задержка проведения в синапсах• По рефлекторной дуге проведение
возбуждения происходит медленнее, чем по
нервному волокну → в одном синапсе
переход медиатора к постсинаптической
мембране происходит за 0,3-0,5 мсек. (т.н.
синаптическая задержка).
• Чем больше синапсов в рефлекторной дуге,
тем больше время рефлекса, т.е. интервал от
начала раздражения до начала деятельности.
• С учетом синаптической задержки
проведение раздражения через один синапс
требует около 1,5-2 мсек.
• У человека наименьшую
продолжительность имеет время
сухожильных рефлексов (оно равно 20-24
мсек).
 У мигательного рефлекса оно больше 50-200 мсек.
У мигательного рефлекса оно больше 50-200 мсек.Суммация возбуждений
• В целостном организме рефлекторный акт может
осуществляться при действии подпороговых
стимулов, если они действуют на рецепторное поле
достаточно часто.
• Такое явление получило название временной
(последовательной) суммации. Пример — рефлекс
чесания у собаки можно вызвать, если подать в одну
точку подпороговые стимулы с частотой 18 гц.
• Суммация подпороговых стимулов можно получить и
тогда, когда они прикладываются на разные точки
кожи, но одновременно — это пространственная
суммация.
• В основе этих явлений лежит процесс суммации
возбуждающих постсинаптических потенциалов на
теле и дендритах нейронов → происходит накопление
медиатора в синаптической щели. В естественных
условиях оба вида суммации сосуществуют.
20. Схема временной (а) и пространственной (б) суммации возбуждений в ЦНС
21. Суммация в ЦНС
Последовательнаясуммация
Пространственная
суммация
23.
 Трансформация ритма возбуждения • изменение количества импульсов
Трансформация ритма возбуждения • изменение количества импульсоввозбуждения, выходящих из нервного центра,
по сравнению с числом импульсов,
приходящих к нему.
Различают два вида трансформации:
• понижающая трансформация, в основе
которой, в основном, лежит явление суммации
возбуждений (пространственной и временной),
когда в ответ на несколько возбуждений,
пришедших к нервной клетке, в последней
возникает только одно возбуждение;
• повышающая трансформация, в ее основе
лежат механизмы умножения
(мультипликации), способные резко увеличить
количество импульсов возбуждения
24. Схема умножения (мультипликации) возбуждения в центральной нервной
Трансформация ритма возбуждений• Частота и ритм импульсов, поступающих к нервным
центрам, и посылаемых ими на периферию, могут
не совпадать.
• В ряде случаев на одиночный импульс,
приложенный к афферентному волокну,
мотонейрон отвечает серией импульсов (в ответ
на одиночный выстрел нервная клетка отвечает
очередью).
 Чаще это бывает при длительном
Чаще это бывает при длительномпостсинаптическом потенциале и зависит от
триггерных свойств аксонного холмика.
• Другой механизм трансформации связан с
эффектами сложения фаз двух или более волн
возбуждения на нейроне → возможны эффекты как
увеличения, так и снижения частоты выходящих
из центра стимулов.
26. Трансформация ритма
Триггерные свойства аксонного холмикаПорог 30 мв
Порог 10 мв
Тело нейрона
Ек
Ео
Аксонный холмик
Ек
Ео
«На ружейный выстрел
нейрон отвечает
пулеметной очередью»
Последействие
Рефлекторные акты, в отличие от потенциалов
действия, заканчиваются не одновременно с
прекращением вызвавшего их раздражения, а
через некоторый период времени.
Продолжительность последействия может во
много раз превышать продолжительность
раздражения.
Основные механизмы последействия:
1. первый связан с суммацией следовой
деполяризации мембраны при частых раздражениях
(посттетаническая потенциация), когда нервная
клетка продолжает давать разряды импульсов,
несмотря на то, что кончилась серия раздражений.
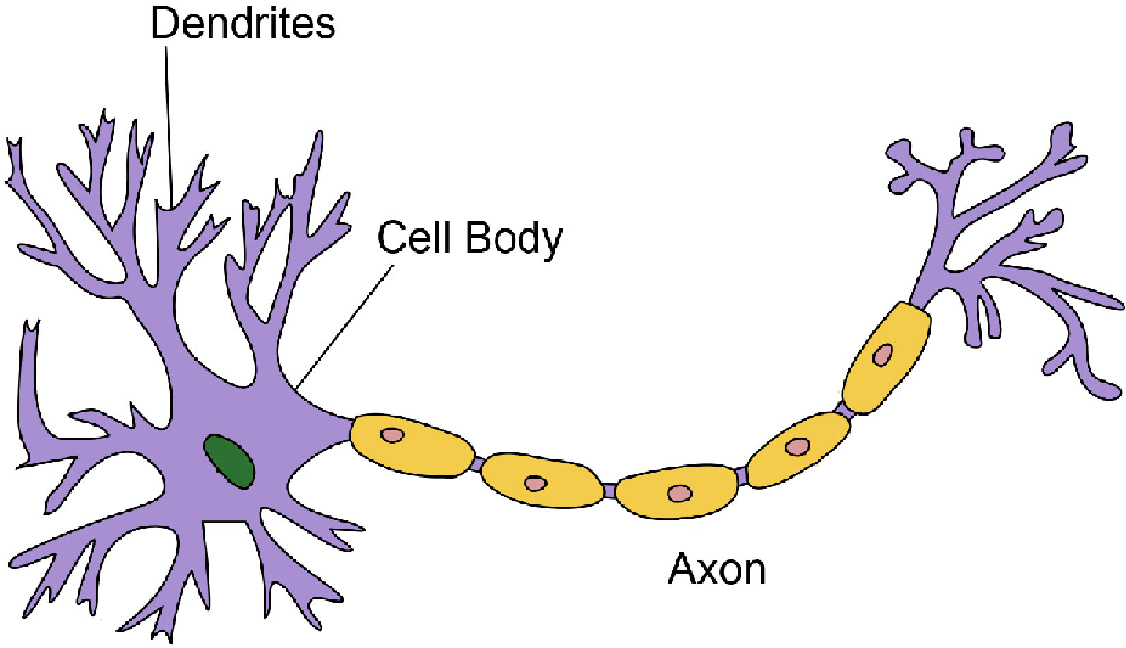
2. второй механизм связывает последействие с
циркуляцией нервных импульсов по замкнутым
нейронным сетям рефлекторного центра.
28. Схема пролонгирования возбуждения (нейронной ловушки) в центральной нервной системе
Утомление нервных центров• Проявляется в постепенном снижении и в конечном
итоге полном прекращении рефлекторного ответа при
продолжительном раздражении афферентных нервных
волокон.
• Утомление в нервных центрах связано прежде всего с
нарушением передачи возбуждения в межнейронных
синапсах.
Утомление зависит от:
— уменьшения запасов синтезированного медиатора;
— уменьшения чувствительности к медиатору
постсинаптической мембраны;
— уменьшение энергетических ресурсов нервной
клетки.
• Мало утомляемы проприоцептивные
тонические рефлексы
Рефлекторный тонус нервных
центров
• Нервные центры обладают тонусом, который
выражается в том, что даже при отсутствии
специальных раздражений, они постоянно
посылают импульсы к рабочим органам.

• В его поддержании участвуют как
афферентные импульсы, поступающие
непрерывно от периферических рецепторов в
ЦНС, так и различные гуморальные
раздражители (гормоны, углекислота и др.)
Высокая чувствительность к гипоксии
• Показано, что 100 г нервной ткани в единицу
времени потребляет кислорода в 22 раза
больше, чем 100 г мышечной ткани. Поэтому
нервные центры очень чувствительны к его
недостатку.
• Чем выше центр, тем более страдает он от
гипоксии.
• Для коры мозга 5-6 минут достаточно, чтобы
без кислорода произошли необратимые
изменения, клетки ствола мозга выдерживают
15-20 минут полного прекращения
кровообращения, а клетки спинного мозга — 2030 минут.
• При гипотермии, когда снижается обмен
веществ, ЦНС дольше переносит гипоксию.
Избирательная чувствительность к
химическим веществам
• Нервные центры, как и синапсы, обладают
высокой чувствительностью к действию
различных химических веществ, особенно
ядов.
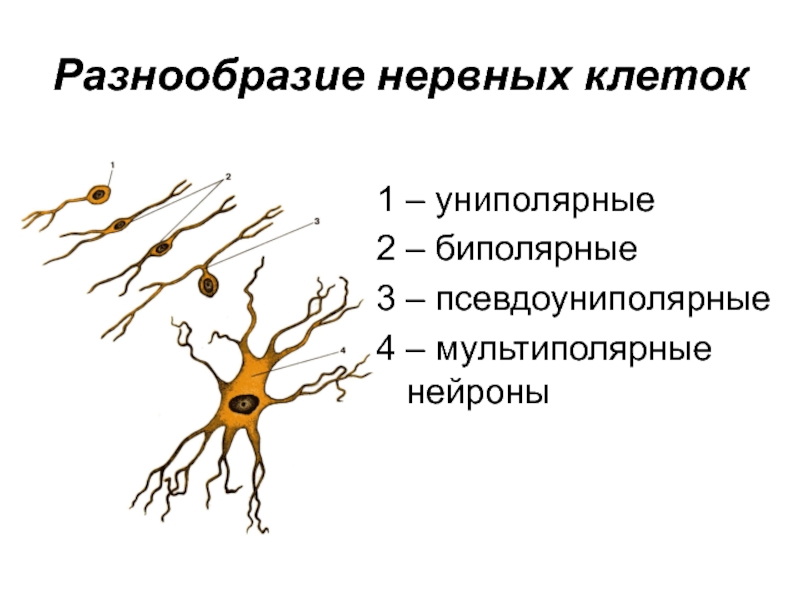 На одном нейроне могут располагаться
На одном нейроне могут располагатьсясинапсы, обладающие различной
чувствительностью к различным химическим
веществам.
• Поэтому можно подобрать такие химические
вещества, которые избирательно будут
блокировать одни синапсы, оставляя другие в
рабочем состоянии. Это делает возможным
корректировать состояния и реакции как
здорового, так и больного организма.
СПАСИБО
ЗА ВНИМАНИЕ!
Слияние аксонов: альтернативный и эффективный механизм восстановления нервов
Обзор
DOI: 10.1016 / j.pneurobio.2018.11.004. Epub 2018 27 ноя.Принадлежности Расширять
Принадлежности
- 1 Программа неврологии, Институт открытия биомедицины Монаша и факультет анатомии и биологии развития, Университет Монаша, Мельбурн, Виктория, 3800, Австралия. Электронный адрес: [email protected].
- 2 Центр Клема Джонса по исследованию старческой деменции, Квинслендский институт мозга, Квинслендский университет, Брисбен, QLD 4072, Австралия.
- 3 Центр исследования деменции Клема Джонса, Квинслендский институт мозга, Квинслендский университет, Брисбен, QLD 4072, Австралия. Электронный адрес: [email protected].
 Электронный адрес:
Электронный адрес: Элемент в буфере обмена
Обзор
Брент Нойман и др.Prog Neurobiol. 2019 фев.
Бесплатная статья Показать детали Показать вариантыПоказать варианты
Формат АннотацияPubMedPMID
DOI: 10.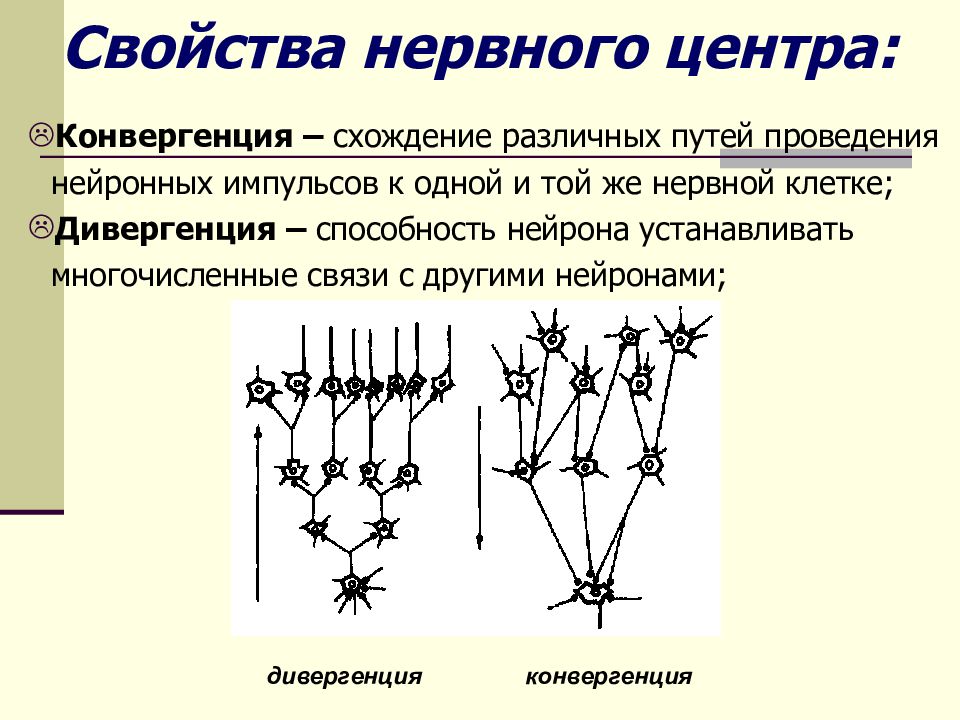 1016 / j.pneurobio.2018.11.004.
Epub 2018 27 ноя.
1016 / j.pneurobio.2018.11.004.
Epub 2018 27 ноя.Принадлежности
- 1 Программа неврологии, Институт открытия биомедицины Монаша и факультет анатомии и биологии развития, Университет Монаша, Мельбурн, Виктория, 3800, Австралия.Электронный адрес: [email protected].
- 2 Центр Клема Джонса по исследованию старческой деменции, Квинслендский институт мозга, Квинслендский университет, Брисбен, QLD 4072, Австралия.
- 3 Центр исследования деменции Клема Джонса, Квинслендский институт мозга, Квинслендский университет, Брисбен, QLD 4072, Австралия. Электронный адрес: [email protected].
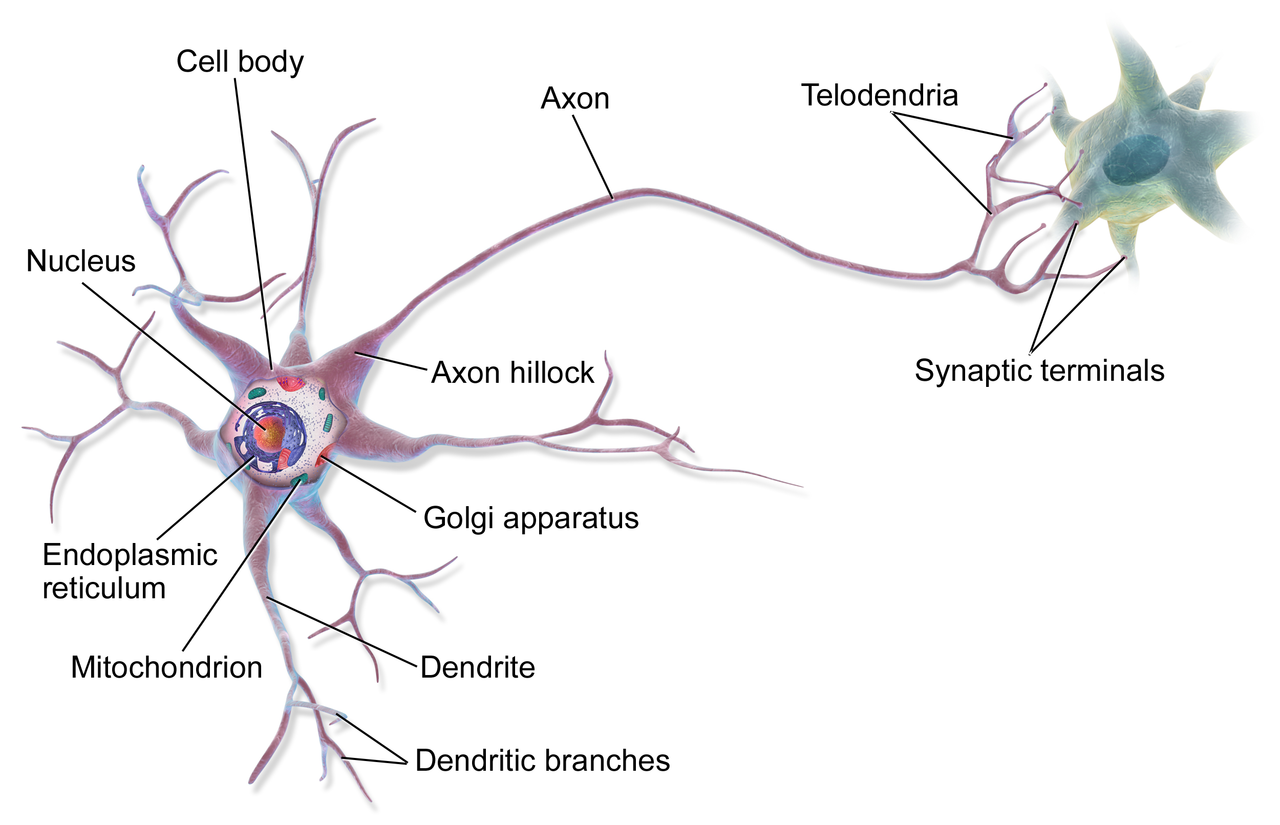 Электронный адрес:
Электронный адрес: Элемент в буфере обмена
Полнотекстовые ссылки Опции CiteDisplayПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Повреждения нервной системы могут вызвать пожизненные заболевания из-за разрыва связи между нервными клетками и их клеточными мишенями.Восстановление этих потерянных связей является конечной целью эндогенных регенеративных механизмов, а также тех, которые вызваны экзогенными манипуляциями в лабораторных или клинических условиях. Повторное соединение между разорванными нейронными волокнами происходит спонтанно у некоторых видов беспозвоночных и может быть индуцировано в системах млекопитающих.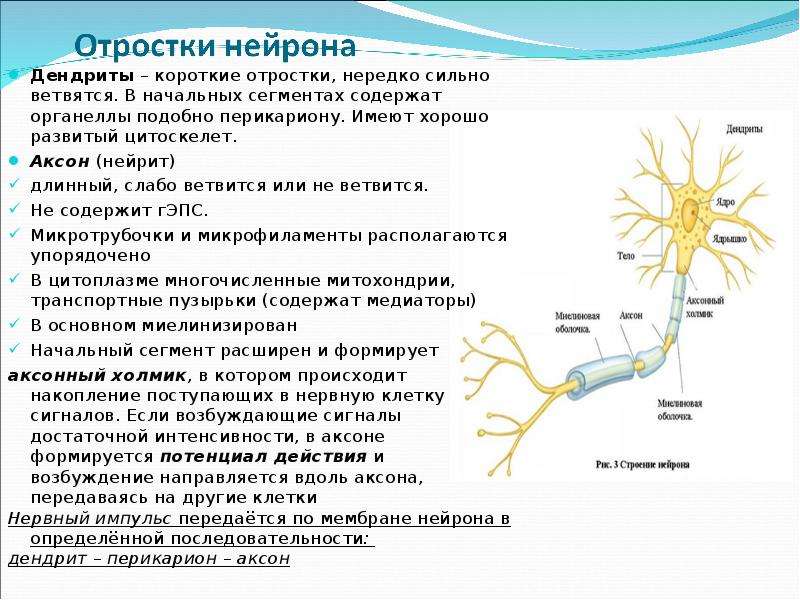 Этот процесс, известный как слияние аксонов, представляет собой высокоэффективное средство восстановления после травмы. Недавний прогресс значительно расширил наше понимание молекулярного контроля слияния аксонов, демонстрируя, что механизм, необходимый для поглощения апоптотических клеток, перепрофилируется, чтобы опосредовать воссоединение между оторванными фрагментами аксонов, которые впоследствии сливаются с помощью белков слияния.Здесь мы рассмотрим наше текущее понимание естественных событий слияния аксонов, а также тех, которые производятся эктопически, с целью достижения лучших клинических результатов.
Этот процесс, известный как слияние аксонов, представляет собой высокоэффективное средство восстановления после травмы. Недавний прогресс значительно расширил наше понимание молекулярного контроля слияния аксонов, демонстрируя, что механизм, необходимый для поглощения апоптотических клеток, перепрофилируется, чтобы опосредовать воссоединение между оторванными фрагментами аксонов, которые впоследствии сливаются с помощью белков слияния.Здесь мы рассмотрим наше текущее понимание естественных событий слияния аксонов, а также тех, которые производятся эктопически, с целью достижения лучших клинических результатов.
Ключевые слова: Распознавание апоптоза; Аксональное слияние; Функциональное восстановление; Фузогены; Ремонт нервов; Фосфатидилсерин; Регенерация.
Copyright © 2018 Авторы. Опубликовано Elsevier Ltd. Все права защищены.
Опубликовано Elsevier Ltd. Все права защищены.
Похожие статьи
- Фосфатидилсериновые спасательные сигналы управляют функциональным восстановлением оторванных аксонов у Caenorhabditis elegans .
Abay ZC, Wong MY, Teoh JS, Vijayaraghavan T., Hilliard MA, Neumann B. Абай З.С. и др.Proc Natl Acad Sci U S. A. 2017, 21 ноября; 114 (47): E10196-E10205. DOI: 10.1073 / pnas.1703807114. Epub 2017 6 ноя. Proc Natl Acad Sci U S A. 2017. PMID: 263 Бесплатная статья PMC.
- Нарушение RAB-5 увеличивает доступность фузогена EFF-1 на поверхности клетки и способствует регенеративной способности нейрона к слиянию аксонов.
Линтон К., Эр-Рияд, Массачусетс, Хо XY, Нойман Б., Джордано-Сантини Р., Хиллиард Массачусетс.
Линтон С. и др.
J Neurosci. 2019 10 апреля; 39 (15): 2823-2836. DOI: 10.1523 / JNEUROSCI.1952-18.2019. Epub 2019 8 февраля.
J Neurosci. 2019.
PMID: 30737314
Бесплатная статья PMC. - Регенерация аксонов происходит посредством специфического слияния аксонов в перерезанных нейронах C. elegans.
Нойман Б., Нгуен К.С., Холл Д.Х., Бен-Якар А., Хиллиард Массачусетс. Neumann B, et al. Dev Dyn.2011 июн; 240 (6): 1365-72. DOI: 10.1002 / dvdy.22606. Epub 2011 17 марта. Dev Dyn. 2011 г. PMID: 21416556 Бесплатная статья PMC.
- Преодоление разрыва: слияние аксонов способствует быстрому функциональному восстановлению нервной системы.
Теох Дж.С., Вонг М.Ю., Виджаярагхаван Т., Нойманн Б. Теох Дж. С. и др. Neural Regen Res.
2018 Апрель; 13 (4): 591-594. DOI: 10.4103 / 1673-5374.230271.
Neural Regen Res.2018.
PMID: 29722300
Бесплатная статья PMC.
Обзор. - Применение протеомики к исследованиям регенерации нервов.
Массинг М.В., Робинсон Г.А., Маркс К.Э., Альзате О., Мэдисон Р.Д. Массинг MW и др. В: Альзате О., редактор. Нейропротеомика. Бока-Ратон (Флорида): CRC Press / Taylor & Francis; 2010. Глава 15. В: Альзате О., редактор. Нейропротеомика. Бока-Ратон (Флорида): CRC Press / Taylor & Francis; 2010 г.Глава 15. PMID: 21882439 Бесплатные книги и документы. Обзор.
 Линтон С. и др.
J Neurosci. 2019 10 апреля; 39 (15): 2823-2836. DOI: 10.1523 / JNEUROSCI.1952-18.2019. Epub 2019 8 февраля.
J Neurosci. 2019.
PMID: 30737314
Бесплатная статья PMC.
Линтон С. и др.
J Neurosci. 2019 10 апреля; 39 (15): 2823-2836. DOI: 10.1523 / JNEUROSCI.1952-18.2019. Epub 2019 8 февраля.
J Neurosci. 2019.
PMID: 30737314
Бесплатная статья PMC.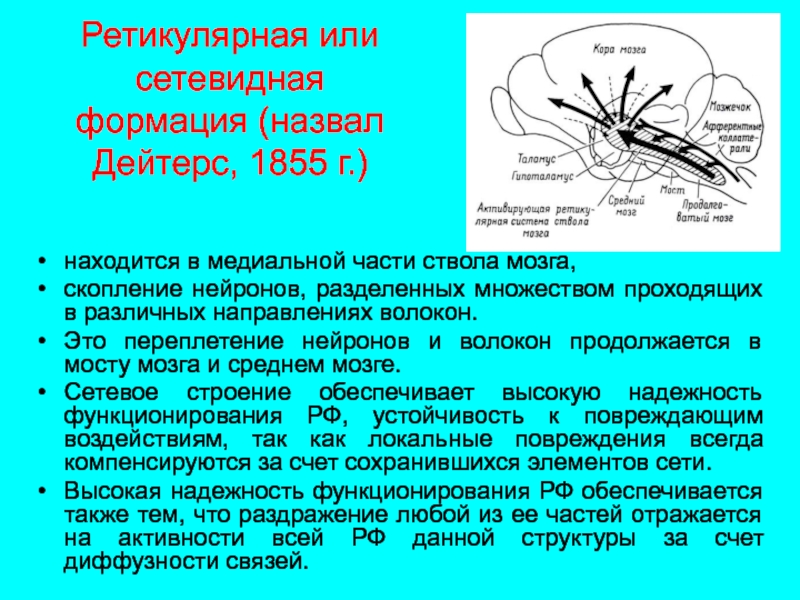 2018 Апрель; 13 (4): 591-594. DOI: 10.4103 / 1673-5374.230271.
Neural Regen Res.2018.
PMID: 29722300
Бесплатная статья PMC.
Обзор.
2018 Апрель; 13 (4): 591-594. DOI: 10.4103 / 1673-5374.230271.
Neural Regen Res.2018.
PMID: 29722300
Бесплатная статья PMC.
Обзор.Процитировано
8 статей- Обзор механизма Tuina, способствующего восстановлению после травмы периферических нервов.
Лю З, Ван Х, Ю Т, Цзяо Ю, Чжан Ю, Лю Д, Сюй Ю, Гуань Цюй, Лу М.Лю З. и др. Evid Based Complement Alternat Med. 1 июля 2021 г .; 1 июля 2021 г .: 6652099. DOI: 10.1155 / 2021/6652099. Электронная коллекция 2021 г. Evid Based Complement Alternat Med. 2021 г. PMID: 34285705 Бесплатная статья PMC. Обзор.
- Отмеченное слияние: передача сигналов фосфатидилсерина при слиянии клеток.
Уитлок Дж. М., Черномордик Л. В.. Whitlock JM и др. J Biol Chem. 2021, январь-июнь; 296: 100411.DOI: 10.1016 / j.jbc.2021.100411. Epub 2021 11 февраля. J Biol Chem. 2021 г. PMID: 33581114 Бесплатная статья PMC. Обзор.
- Пластичность мозга после лечения повреждений периферических нервов массажной терапией на основе функциональной магнитно-резонансной томографии в состоянии покоя.
Xing XX, Zheng MX, Hua XY, Ma SJ, Ma ZZ, Xu JG. Xing XX и др. Neural Regen Res. 2021 Февраль; 16 (2): 388-393.DOI: 10.4103 / 1673-5374.2. Neural Regen Res. 2021 г. PMID: 32859803 Бесплатная статья PMC.
- Устранение разрыва между повреждением периферических нервов у млекопитающих и беспозвоночных: протокол нового восстановления нерва.
Жилет М, Гуида А, Коломбини С, Кордес К, Пена Д, Маки М, Брионес М, Антонио С, Холлифилд С, Тиан Э, Джеймс Л, Борашан С, Вудсон Дж, Ровиг Дж, Шихадех Х, Карабачев А, Бросиус Дж. , Писторио А.Вест М и др. JMIR Res Protoc. 2020 27 августа; 9 (8): e18706. DOI: 10,2196 / 18706. JMIR Res Protoc. 2020. PMID: 32851981 Бесплатная статья PMC.
- Применение гидрогелей на основе фибрина для защиты нервов и регенерации после травм спинного мозга.
Ю З, Ли Х, Ся П, Конг В., Чанг И, Фу Ц, Ван К., Ян Х, Ци З. Ю. З. и др. J Biol Eng. 2020 3 августа; 14:22.DOI: 10.1186 / s13036-020-00244-3. Электронная коллекция 2020. J Biol Eng. 2020. PMID: 32774454 Бесплатная статья PMC. Обзор.
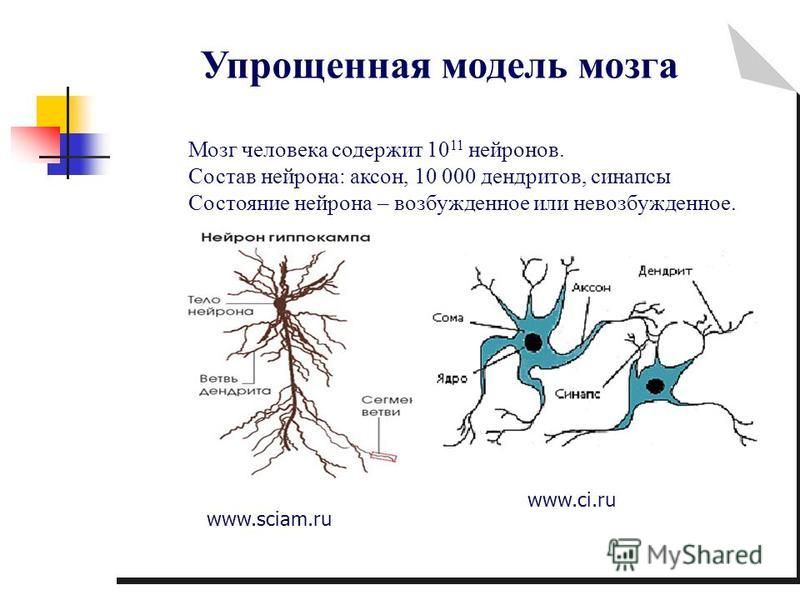
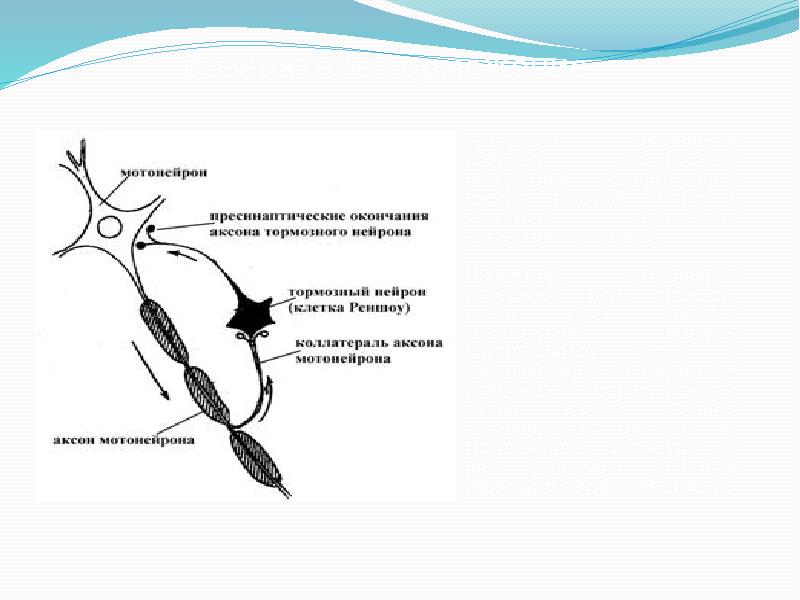
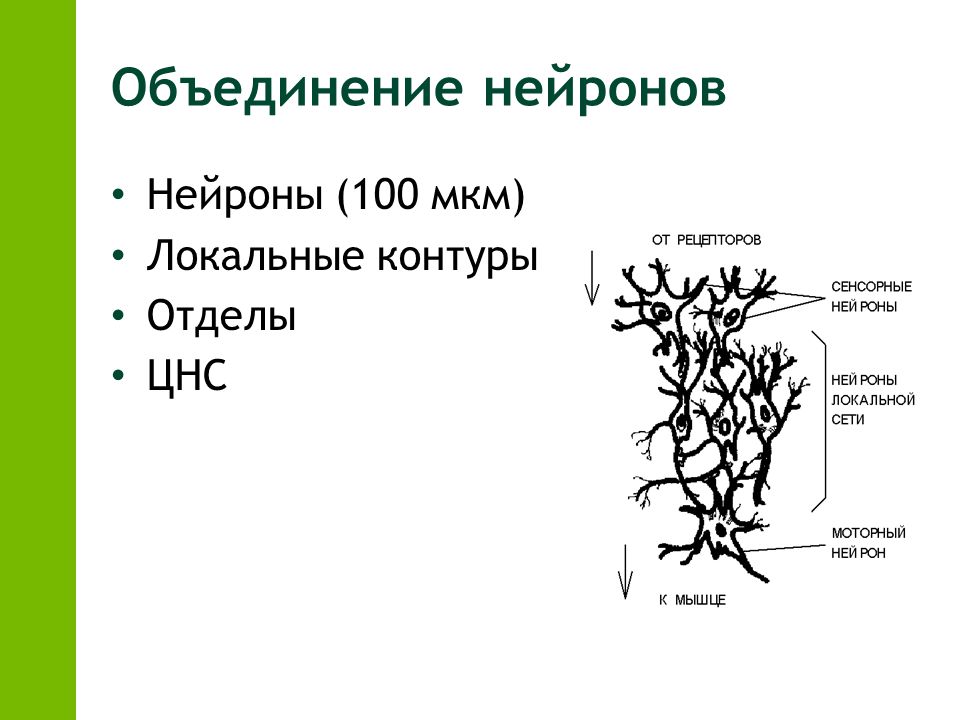
Типы публикаций
- Поддержка исследований, за пределами США. Правительство
Условия MeSH
- Клеточная коммуникация / физиология
- Регенерация нервов / физиология *
- Восстановление функций / физиологии *
цитировать
КопироватьФормат: AMA APA ГНД NLM
Нервная система | ЛОР 425 — Общая энтомология
Центральная нервная система
Вид тела сбоку, показывающий взаимное расположение кровеносной (желтый), пищеварительной (зеленый) и нервной (синий) систем.
Подобно большинству других членистоногих, насекомые имеют относительно простую центральную нервную систему с дорсальным мозгом, связанным с вентральным нервным канатиком, который состоит из парных сегментарных ганглиев , проходящих вдоль вентральной средней линии грудной клетки и брюшной полости. Ганглии в каждом сегменте связаны друг с другом коротким медиальным нервом (комиссурами), а также соединены межсегментарными связками с ганглиями в соседних сегментах тела. В целом центральная нервная система имеет лестничный вид: комиссур — ступеньки лестницы, а межсегментарные связки — рельсы.В более «продвинутых» отрядах насекомых отдельные ганглии имеют тенденцию объединяться (как латерально, так и продольно) в более крупные ганглии, которые обслуживают несколько сегментов тела.
Мозг насекомого представляет собой комплекс из шести сросшихся ганглиев (трех пар), расположенных дорсально внутри головной капсулы. Каждая часть мозга контролирует (иннервирует) ограниченный спектр действий в теле насекомого:
- Protocerebrum: Первая пара ганглиев в значительной степени связана со зрением; они иннервируют сложные глаза и глазки.
- Deutocerebrum: Вторая пара ганглиев обрабатывает сенсорную информацию, собранную антеннами.
- Tritocerebrum: Третья пара ганглиев иннервирует верхнюю губу и интегрирует сенсорные сигналы от прото- и дейтоцеребрумов. Они также связывают мозг с остальной частью брюшного нервного канатика и стоматологической нервной системой (см. Ниже), которая контролирует внутренние органы. Комиссура для тритоцеребрума обвивает пищеварительную систему, предполагая, что эти ганглии изначально располагались на за ртом и мигрировали вперед (вокруг пищевода) в ходе эволюции.

Расположен вентрально в головной капсуле (сразу под мозгом и пищеводом) — это еще один комплекс слитых ганглиев (вместе называемых подэзофагеальным ганглием). Эмбриологи полагают, что эта структура содержит нервные элементы из трех примитивных сегментов тела, которые слились с головой и образовали ротовой аппарат. У современных насекомых подэзофагеальный ганглий иннервирует не только нижнюю челюсть, верхнюю челюсть и губу, но также гортань, слюнные железы и мышцы шеи. Пара околопищеводных соединительных элементов петляет вокруг пищеварительной системы, чтобы связать вместе мозг и субэзофагеальный комплекс.
Пара околопищеводных соединительных элементов петляет вокруг пищеварительной системы, чтобы связать вместе мозг и субэзофагеальный комплекс.
В грудной клетке три пары грудных ганглиев (иногда слитых) контролируют движение, иннервируя ноги и крылья. Грудные мышцы и сенсорные рецепторы также связаны с этими ганглиями. Точно так же брюшные ганглии контролируют движения брюшных мышц. Дыхальца как в грудной клетке, так и в брюшной полости контролируются парой боковых нервов, которые отходят от каждого сегментарного ганглия (или срединным вентральным нервом, который разветвляется в каждую сторону). Пара терминальных брюшных ганглиев (обычно сливающихся с образованием большого хвостового ганглия ) иннервирует задний проход, внутренние и внешние гениталии и сенсорные рецепторы (например, церки), расположенные на заднем конце насекомого.
Паравертебральные ганглии — обзор
Расположение паравертебральных ганглиев
Паравертебральные ганглии связаны между собой и образуют симпатические цепи, идущие от шейного до крестцового уровней (рис. 6.4). Хотя они изначально развиваются сегментарно, может происходить значительное слияние соседних ганглиозных зачатков, так что общее количество макроскопических ганглиев варьирует.
6.4). Хотя они изначально развиваются сегментарно, может происходить значительное слияние соседних ганглиозных зачатков, так что общее количество макроскопических ганглиев варьирует.
В шейном отделе имеются три или четыре больших ганглия.Самым большим из них является верхний шейный ганглий, длина которого составляет около 2,5–3,0 см (рис. 6.4; Becker, Grunt, 1957; Mitchell, 1953a) и который обычно содержит более 1 миллиона нейронов (Ebbesson, 1963; Pearson and Pytel, 1978a). ). Ганглии обычно лежат непосредственно дорсальнее начала внутренней сонной артерии. Он имеет анатомические связи с черепными нервами (за исключением первого, второго и восьмого) либо напрямую, либо через внутренний сонный нерв и сплетение, а также с первым, четвертым или пятым шейными спинномозговыми нервами (Hoffman, 1957; Mitchell, 1953a, c, 1954; Parkinson, 1988; Parkinson et al., 1978; Пик и Шихан, 1946). Эти пути распределяют аксоны конечных мотонейронов к большинству их головных мишеней. Другие цели могут быть достигнуты с помощью волокон, следующих за головными артериями (Mitchell, 1954). Вдоль проксимального отдела внутренних сонных нервов есть небольшие микроганглии в дополнение к тем, которые описаны в кавернозном сплетении (Mitchell, 1953c).
Вдоль проксимального отдела внутренних сонных нервов есть небольшие микроганглии в дополнение к тем, которые описаны в кавернозном сплетении (Mitchell, 1953c).
Примерно у 60% людей имеется средний шейный ганглий длиной 0,7–0,8 см, расположенный примерно на уровне шестого шейного позвонка, чуть выше нижней щитовидной артерии (рис.6.4). Он имеет анатомические связи с пятым и шестым шейными спинномозговыми нервами и способствует развитию сплетений пищевода, трахеи и аорты. В некоторых случаях средний шейный ганглий сливается с нижним полюсом верхнего шейного ганглия с образованием комбинированного «средневерхнего» шейного ганглия длиной до 4,5 см (Becker and Grunt, 1957; Hoffman, 1957; Mitchell, 1953a; Wrete, 1959). ).
Ниже среднего шейного ганглия обычно находится небольшой позвоночный ганглий, лежащий непосредственно вентральнее позвоночной артерии.От этого ганглия возникают позвоночные нервы. Иногда он срастается со средним шейным ганглием (Becker, Grunt, 1957; Mitchell, 1953a; Wrete, 1959).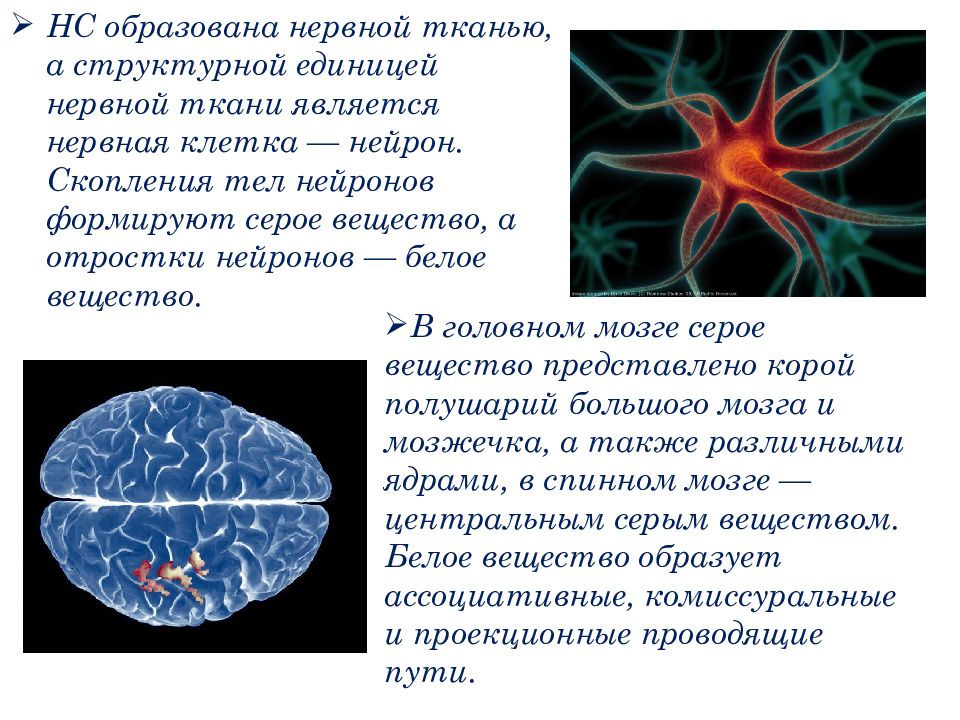
Самый нижний узел шеи — это нижний шейный узел, который обычно сливается с первым грудным узлом, образуя звездчатый узел (рис. 6.4; Becker and Grunt, 1957; Harman, 1900; Hoffman, 1957; Mitchell, 1953a; Пик и Шихан, 1946; Рет, 1959). Комбинированный узел может достигать 2,8 см. Иногда он и все грудные ганглии вплоть до четвертого сегмента сливаются (Wrete, 1959).
Оставшиеся грудные симпатические ганглии обычно связаны с каждым спинномозговым нервом, кроме самых нижних ганглиев, которые могут сливаться с соседними поясничными ганглиями (Pick and Sheehan, 1946; Wrete, 1959). Ганглии относительно малы, каждый содержит около 90 000–100 000 тел нервных клеток.
Грудные ганглии распределяют волокна по соответствующим или соседним спинномозговым нервам, а более верхние ганглии вносят непосредственный вклад в аортальное, сердечное, легочное и пищеводное сплетения (рис.6.4). Звездчатый ганглий также поставляет симпатические волокна к верхней конечности (Kuntz, 1945; Kuntz et al. , 1956; Mitchell, 1953a). Внутренние нервы отходят от грудных ганглиев и обеспечивают симпатические пути к внутренним органам брюшной полости. Вегетативные компоненты нервов включают не только аксоны конечных мотонейронов грудных ганглиев, но также преганглионарные аксоны, предназначенные для симпатических нейронов превертебральных ганглиев. Верхний грудной (или больший) внутренностный нерв является самым большим внутренним нервом и берет начало от пятого, шестого, девятого или десятого грудных ганглиев (рис.6.4; Пик и Шихан, 1946). Он имеет анатомические связи с пищеводным и аортальным сплетениями и соединяется дистально с чревными, верхними брыжеечными и аортекоренальными ганглиями. Там, где нерв пересекает аорту, обычно находится небольшой узел (Kuntz, 1945; Mitchell, 1953a). Средний грудной (или меньший) внутренностный нерв отходит от девятого, десятого, десятого и одиннадцатого грудных сегментов и сообщается с сердечным сплетением и ганглиями аорты (см. Ниже). Нижний грудной (или, по крайней мере,) внутренностный нерв, если присутствует, берет начало от нижних грудных ганглиев и в основном питается почечным сплетением и ганглиями (рис.
, 1956; Mitchell, 1953a). Внутренние нервы отходят от грудных ганглиев и обеспечивают симпатические пути к внутренним органам брюшной полости. Вегетативные компоненты нервов включают не только аксоны конечных мотонейронов грудных ганглиев, но также преганглионарные аксоны, предназначенные для симпатических нейронов превертебральных ганглиев. Верхний грудной (или больший) внутренностный нерв является самым большим внутренним нервом и берет начало от пятого, шестого, девятого или десятого грудных ганглиев (рис.6.4; Пик и Шихан, 1946). Он имеет анатомические связи с пищеводным и аортальным сплетениями и соединяется дистально с чревными, верхними брыжеечными и аортекоренальными ганглиями. Там, где нерв пересекает аорту, обычно находится небольшой узел (Kuntz, 1945; Mitchell, 1953a). Средний грудной (или меньший) внутренностный нерв отходит от девятого, десятого, десятого и одиннадцатого грудных сегментов и сообщается с сердечным сплетением и ганглиями аорты (см. Ниже). Нижний грудной (или, по крайней мере,) внутренностный нерв, если присутствует, берет начало от нижних грудных ганглиев и в основном питается почечным сплетением и ганглиями (рис.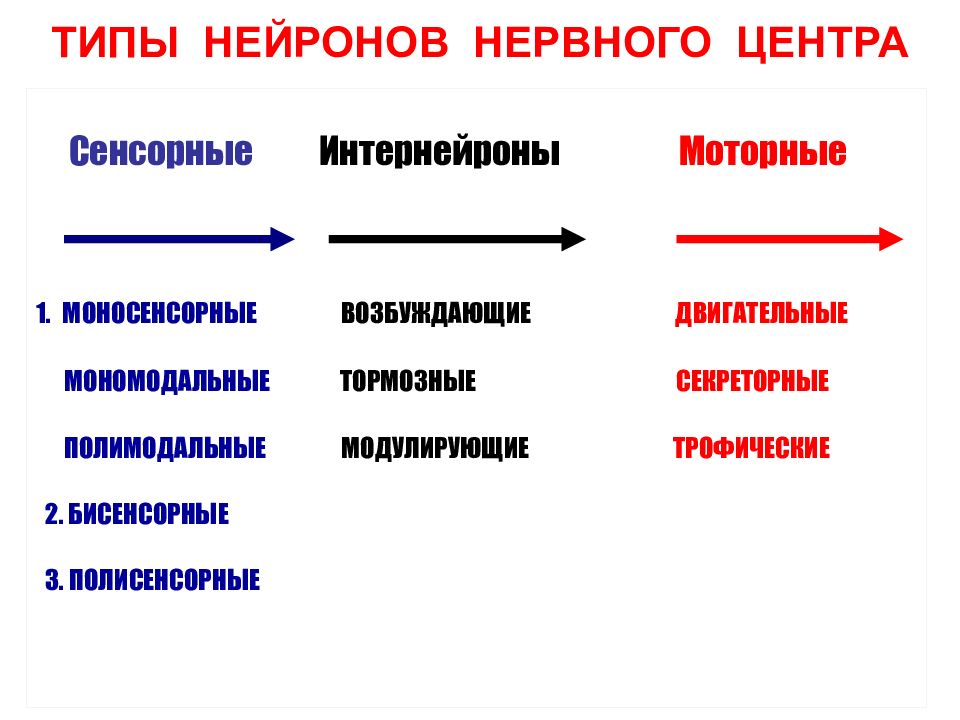 6.4).
6.4).
Поясничные симпатические ганглии чрезвычайно разнообразны по своему количеству и расположению вдоль поясничных симпатических цепей, и в их расположении нет постоянной закономерности (Cowley and Yeager, 1949; Kuntz and Alexander, 1950; Mitchell, 1953a; Webber, 1955, 1958). В среднем неслитые поясничные ганглии содержат от 60 000 до 85 000 тел нервных клеток (Webber, 1958). Поясничные ганглии имеют различные анатомические связи с поясничными спинномозговыми нервами и распределяют волокна с поясничными внутренними нервами к нижнему брыжеечному и гипогастральному сплетениям и сплетению аорты (рис.6.4). Многие нейроны в поясничных ганглиях проецируются на мишени в нижних конечностях (Kuntz and Alexander, 1950; Kuntz et al., 1956; Mitchell, 1953a; Pick and Sheehan, 1946; Webber, 1955, 1958). Иногда в поясничной области могут быть прямые перекрестные связи между двумя симпатическими цепями, что, вероятно, является единственной областью, где это происходит (Cowley and Yeager, 1949; Pick and Sheehan, 1946; Webber, 1955).
На уровне крестца симпатические ганглии маленькие и обычно расположены регулярно.Может присутствовать до шести ганглиев с каждой стороны (Wrete, 1959). Обе симпатические цепи встречаются и сливаются на копчиковых уровнях, где может быть один маленький ганглий («ganglion impar»; Mitchell, 1953a; Pick and Sheehan, 1946). Нейроны в этих ганглиях проходят через крестцовые и копчиковые нервы в тазовое сплетение (рис. 6.4) и, более дистально, в крупные кровеносные сосуды в области таза.
С симпатической цепью на всех уровнях связано большое количество «дополнительных» микроганглиев.Эти ганглии находятся в брюшных корешках, особенно на поясничном уровне (Alexander et al., , 1949; Boyd, 1957), и в симпатических корешках (серые сообщающиеся ветви) спинномозговых нервов (Boyd, 1957; Kuntz and Alexander , 1950; Kuntz et al., 1956; Pick and Sheehan, 1946; Skoog, 1947; Webber, 1955). Микроганглии также обнаруживаются в поясничных чревных нервах (Webber, 1955, 1958) и вдоль внутреннего сонного нерва и позвоночных нервов, отходящих от шейных ганглиев (Becker and Grunt, 1957; Harman, 1900; Hoffman, 1957; Kuntz et al.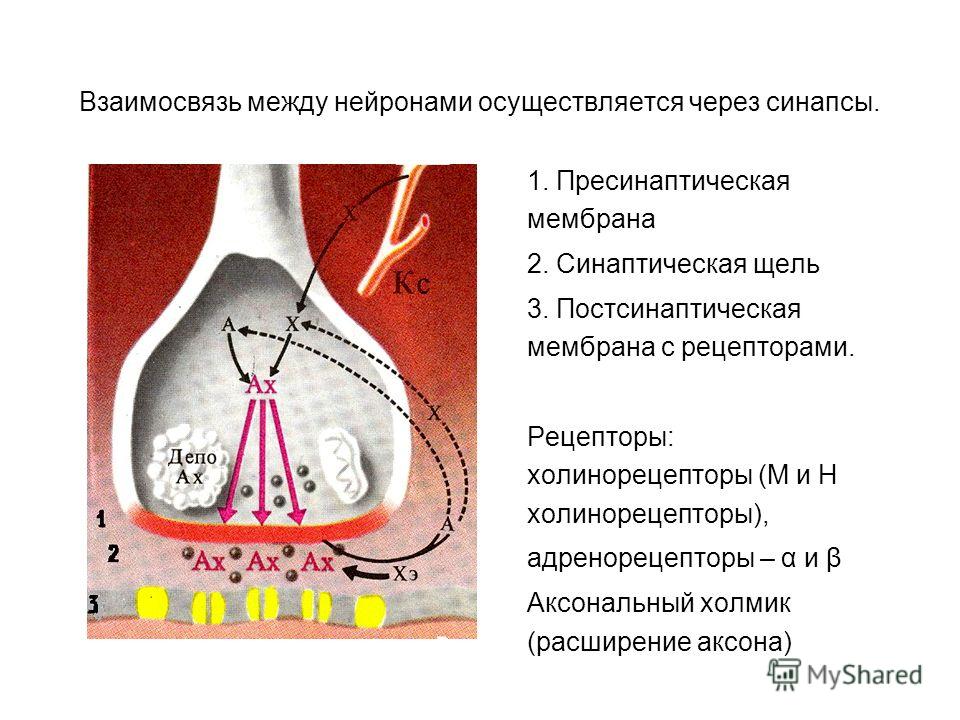 , 1957; Митчелл, 1953c). Микроганглии обычно содержат от нескольких сотен до нескольких тысяч тел нервных клеток (Kuntz et al., , 1956, 1957; Webber, 1955, 1958), хотя некоторые добавочные ганглии могут содержать от 10 000 до 20 000 нейронов (Alexander et al. , 1949; Kuntz et al., 1957; Webber, 1958).
, 1957; Митчелл, 1953c). Микроганглии обычно содержат от нескольких сотен до нескольких тысяч тел нервных клеток (Kuntz et al., , 1956, 1957; Webber, 1955, 1958), хотя некоторые добавочные ганглии могут содержать от 10 000 до 20 000 нейронов (Alexander et al. , 1949; Kuntz et al., 1957; Webber, 1958).
| Первичные пузырьки | Вторичные пузырьки | Строения для взрослых |
Пузырь переднего мозга ( prosencephalon ) | Telencephalon | Полушария головного мозга, состоящие из коры и медуллярный центр, базальные ганглии, терминальная пластинка, гиппокамп, полосатое тело и обонятельная система |
Промежуточный мозг | Таламус, эпиталамус, гипоталамус, субталамус, нейрогипофиз, шишковидная железа, сетчатка, зрительный нерв, мамиллярные тела | |
Пузырь среднего мозга ( mesencephalon ) | Средний мозг | Средний мозг |
Пузырь заднего мозга ( rhombencephalon ) | Metencephalon | Мост и мозжечок |
продолговатый мозг | Медулла |
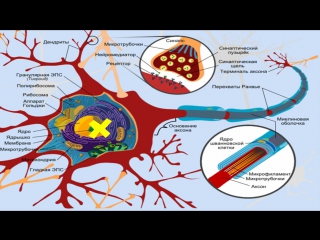 Передние нейропоры закрываются на 90-21 день включительно.
26 , а хвостовой нейропор закрывается перед концом четвертой недели .
Передние нейропоры закрываются на 90-21 день включительно.
26 , а хвостовой нейропор закрывается перед концом четвертой недели .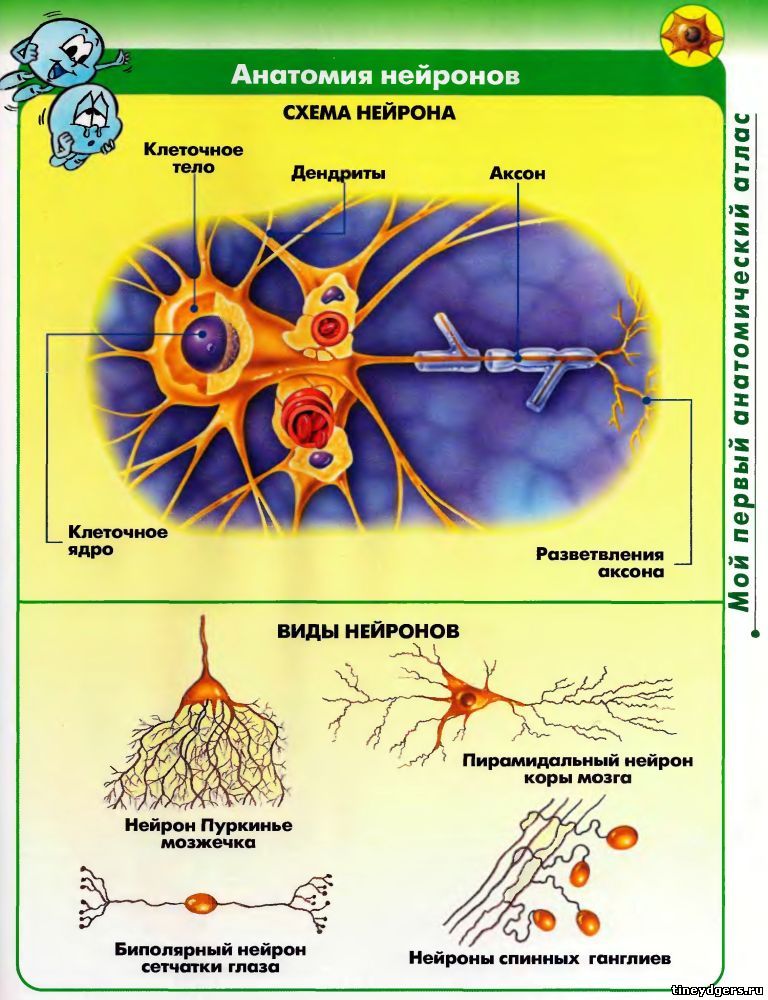 желудочковый
зона дает нейробластов (будущие нервные клетки) и глиобласты (будущие поддерживающие клетки), которые мигрируют в промежуточную зону образуют два набора ячеек ( alar
пластина и базальная пластина )
разделены бороздкой под названием борозда Лимиты . Ячейки в сигнале пластина становится афферентной ( сенсорной ) нейрон и образуют спинной
(задний) рог спинного мозга.Клетки
в базальной пластине становится эфферентным ( мотор ) нейрон и образуют вентральный
(передний) рог спинного мозга. В
два вентральных рога выпуклые вентрально, образуя вентральную срединную щель . В
спинные рога сливаются, образуя спинной
срединная перегородка . Просвет
нервная трубка становится центральным каналом спинного мозга.
желудочковый
зона дает нейробластов (будущие нервные клетки) и глиобласты (будущие поддерживающие клетки), которые мигрируют в промежуточную зону образуют два набора ячеек ( alar
пластина и базальная пластина )
разделены бороздкой под названием борозда Лимиты . Ячейки в сигнале пластина становится афферентной ( сенсорной ) нейрон и образуют спинной
(задний) рог спинного мозга.Клетки
в базальной пластине становится эфферентным ( мотор ) нейрон и образуют вентральный
(передний) рог спинного мозга. В
два вентральных рога выпуклые вентрально, образуя вентральную срединную щель . В
спинные рога сливаются, образуя спинной
срединная перегородка . Просвет
нервная трубка становится центральным каналом спинного мозга. У года рождения , мозговой конус
простирается до L3 позвонка.
У взрослого человека мозговой конус простирается до позвонка L1 . Спинальный
люмбальные проколы должны выполняться каудально к мозговому конусу во избежание
повреждение спинного мозга.
У года рождения , мозговой конус
простирается до L3 позвонка.
У взрослого человека мозговой конус простирается до позвонка L1 . Спинальный
люмбальные проколы должны выполняться каудально к мозговому конусу во избежание
повреждение спинного мозга.

 Он также вызывает микроцефалию и врожденный порок сердца .
Он также вызывает микроцефалию и врожденный порок сердца .Нейрорегенерация — Центр регенеративной медицины
Нейрорегенерация
Конусы роста спинномозговых нейронов
Флуоресцентные спинномозговые нейроны у развивающегося эмбриона Xenopus
Иммуноокрашивание нейронов гиппокампа для выявления зеленого цитоскелета микротрубочек
Совместное культивирование нервных мышц
Контактные спайки в конусе роста нервов (паксиллин — красным, микротрубочки — зеленым).
Адгезии субстрата в конусе роста, вызванные нейротрофическим фактором головного мозга

Сложные тонкие структуры, из которых состоит нервная система — головной мозг, спинной мозг и периферические нервы — подвержены различным типам травм, начиная от травм и заканчивая нейродегенеративными заболеваниями, вызывающими прогрессирующее ухудшение: болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз ( БАС, также известный как болезнь Лу Герига), рассеянный склероз и множественная системная атрофия.
К сожалению, из-за сложности головного и спинного мозга происходит небольшая спонтанная регенерация, восстановление или заживление. Следовательно, повреждение головного мозга, паралич в результате травмы спинного мозга и повреждение периферических нервов часто являются постоянными и приводят к потере трудоспособности.
Следовательно, повреждение головного мозга, паралич в результате травмы спинного мозга и повреждение периферических нервов часто являются постоянными и приводят к потере трудоспособности.
Пациентам с серьезными травмами нервной системы или инсультом часто требуется пожизненная помощь, что ложится огромным бременем на пациентов, их семьи и общество. Для улучшения лечения неврологических травм необходимы инновационные стратегии, меняющие парадигму.Исследования нейрорегенерации в клинике Мэйо находятся на переднем крае лечения нервной системы.
Более подробно о нейрорегенерации можно прочитать в буклете «Нейрорегенеративная медицина» в Mayo Clinic.
Основные направления
Клиницисты Mayo Clinic, ученые, инженеры и другие специалисты Центра регенеративной медицины применяют мультидисциплинарный интегративный подход к нейрорегенерации при ряде разрушительных неврологических состояний. Это многогранное исследование, начиная от фундаментальных научных открытий и заканчивая клиническим применением.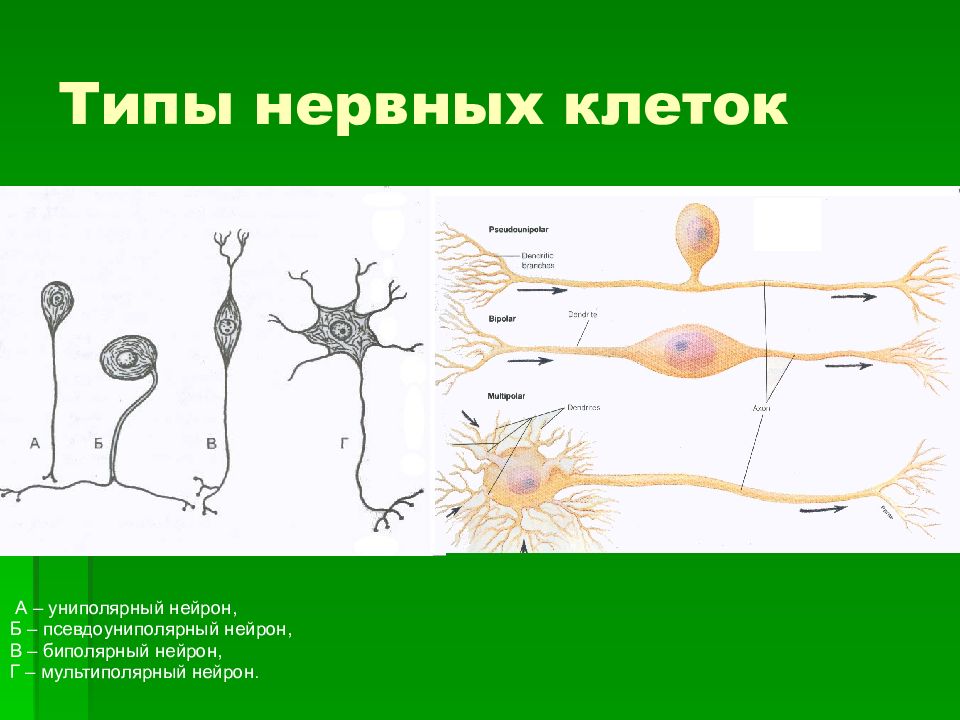
Исследования по конкретным заболеваниям
Болезнь Альцгеймера. Болезнь Альцгеймера является основной причиной деменции у пожилых людей с прогрессирующей потерей нейронов в областях мозга, ответственных за обучение и память. Усилия по исследованию болезни Альцгеймера сосредоточены на понимании того, почему нейроны в мозге дегенерируют с болезнью Альцгеймера и как замедлить этот процесс или заменить потерянные нейроны.
Исследователи Mayo изучают влияние восстановления цереброваскулярной функции посредством трансплантации клеток-предшественников сосудистых клеток-предшественников индуцированных плюрипотентных стволовых (iPS) клеток на амилоидную патологию и когнитивные функции у мышей, моделирующих амилоидную болезнь Альцгеймера.Клетки iPS , преобразованные из фибробластов кожи путем трансдукции четырех факторов транскрипции (Oct3 / 4, SOX2, Klf4, c-Myc), могут генерировать все ткани в организме, включая клетки сосудов.
Этот инновационный подход, вероятно, позволит разработать рациональные схемы регенеративной терапии сосудов против болезни Альцгеймера.
- Боковой амиотрофический склероз. Исследователи клиники Мэйо тестируют клеточную терапию бокового амиотрофического склероза (БАС).Это исследование все еще находится на ранней стадии, и в нем используются мезенхимальные стволовые клетки, полученные из жировой ткани, из собственного тела пациента. Эти клетки модифицируются в лаборатории и доставляются обратно в нервную систему пациента, чтобы способствовать регенерации нейронов.

Энтони Дж. Виндебанк, доктор медицины, и Натан П. Стафф, доктор медицины, доктор философии, неврологи и исследователи клиники Мэйо, обсуждают последние исследования в области лечения БАС.
Рассеянный склероз.
Хотя ученые много знают о повреждении нервов и их изолирующей оболочки (миелин) во время рассеянного склероза (РС) и о том, как иммунная система вызывает это повреждение, точные причины атаки иммунной системы очень плохо изучены. Отсутствие понимания точной причины MS является проблемой для разработки эффективных методов лечения, и лаборатории Mayo Clinic работают над тем, чтобы лучше понять это заболевание.Защита нервов и миелина от повреждения или восстановление миелина после того, как он был поврежден, также имеет потенциал для лечения MS .Повреждение нервов и миелина может быть серьезным в MS и является основной причиной функциональных нарушений. Однако спонтанное восстановление этого повреждения иногда наблюдается у людей с MS . Исследователи Центра регенеративной медицины активно занимаются разработкой методов лечения, призванных стимулировать это восстановление и тем самым способствовать восстановлению утраченных функций.
Были идентифицированы антитела, которые связываются с миелином и нервными клетками, защищают нервы от повреждений и стимулируют регенерацию миелина.Недавнее исследование также показало, что регенерацию миелиновой оболочки могут стимулировать небольшие свернутые молекулы ДНК (аптамеры).
- Болезнь Паркинсона. Исследователи изучают генетический вклад в предрасположенность к болезни Паркинсона путем создания банка кожи и линий клеток iPS от людей с болезнью Паркинсона. Наличие такой клеточной линии дает возможность генерировать клетки, которые умирают при нейродегенеративном заболевании, что позволяет исследователям лучше понять генетическую причину этого состояния и разработать новые методы лечения этих заболеваний в будущем.
Множественная системная атрофия. Множественная системная атрофия (МСА) — прогрессирующее нейродегенеративное заболевание со смертельным исходом.
Признак заболевания — глиальные цитоплазматические включения. Основным компонентом глиальных цитоплазматических включений является альфа-синуклеин. Агрегация микрофибрилл альфа-синуклеина приводит к цепочке событий, включая активацию микроглии, воспаление и дегенерацию глии и нейронов. Вероятные механизмы включают дефицит фактора роста (BDNF, GDNF), токсические цитокины и окислительное повреждение.Исследования сосредоточены на предотвращении агрегации альфа-синуклеина с помощью таких препаратов, как рифампицин или пароксетин; использование мезенхимальных стволовых клеток для обеспечения и доставки факторов роста; и противодействие активации микроглии и воспалительной реакции такими агентами, как внутривенный иммуноглобулин.
 Хотя ученые много знают о повреждении нервов и их изолирующей оболочки (миелин) во время рассеянного склероза (РС) и о том, как иммунная система вызывает это повреждение, точные причины атаки иммунной системы очень плохо изучены. Отсутствие понимания точной причины MS является проблемой для разработки эффективных методов лечения, и лаборатории Mayo Clinic работают над тем, чтобы лучше понять это заболевание.
Хотя ученые много знают о повреждении нервов и их изолирующей оболочки (миелин) во время рассеянного склероза (РС) и о том, как иммунная система вызывает это повреждение, точные причины атаки иммунной системы очень плохо изучены. Отсутствие понимания точной причины MS является проблемой для разработки эффективных методов лечения, и лаборатории Mayo Clinic работают над тем, чтобы лучше понять это заболевание.
 Признак заболевания — глиальные цитоплазматические включения. Основным компонентом глиальных цитоплазматических включений является альфа-синуклеин. Агрегация микрофибрилл альфа-синуклеина приводит к цепочке событий, включая активацию микроглии, воспаление и дегенерацию глии и нейронов. Вероятные механизмы включают дефицит фактора роста (BDNF, GDNF), токсические цитокины и окислительное повреждение.
Признак заболевания — глиальные цитоплазматические включения. Основным компонентом глиальных цитоплазматических включений является альфа-синуклеин. Агрегация микрофибрилл альфа-синуклеина приводит к цепочке событий, включая активацию микроглии, воспаление и дегенерацию глии и нейронов. Вероятные механизмы включают дефицит фактора роста (BDNF, GDNF), токсические цитокины и окислительное повреждение.Лечебные процедуры
Иммунный ответ и нейрорегенерация. Исследователи из Клинического центра регенеративной медицины Майо разрабатывают многочисленные подходы к ослаблению определенных типов иммунных клеток при воспалении центральной нервной системы (ЦНС) и применяют стратегии для лечения различных заболеваний, включая воспаление, развивающееся в ходе трансплантации стволовых клеток, генную терапию.
или управляемая факторами регенерация тканей ЦНС .Исследования продемонстрировали терапевтический эффект в снижении двигательной дисфункции и нарушения гематоэнцефалического барьера в модельных системах рассеянного склероза за счет устранения антиген-специфических ответов Т-лимфоцитов CD8. Оптимизируя визуализацию нейровоспаления с помощью конфокальной микроскопии высокого разрешения, МРТ мелких животных и профилирования инфильтрирующих ЦНС иммунных клеток с помощью проточной цитометрии, можно выделить и фенотипировать инфильтрующие ЦНС иммунные клетки in vivo и визуализировать в реальном времени ведущие события. воспалительному разрушению нервной ткани.
Ремонт спинного мозга. Рост нервных волокон (аксонов) необходим для восстановления и функционального восстановления спинного мозга. Деструкция тканей кистами и глиозом в месте травмы создает барьер для регенерации.
Текущие исследования используют тканевую инженерию с биоразлагаемыми полимерными каркасами (PLGA, PCLF, OPF), нагруженными различными клетками, способствующими росту (шванновские клетки, клетки-предшественники нейронов, мезенхимальные стволовые клетки) и различными факторами роста (GDNF, NT3, BDNF) для связывания промежуток, и способствовать регенерации аксонов и функциональному восстановлению в спинном мозге крыс и мышей, в конечном итоге для будущего использования у пациентов.
Кроме того, исследователи клиники Мэйо исследуют влияние физических упражнений и местной доставки стероидов на регенерацию аксонов и функциональное восстановление.
Регенерация и восстановление периферических нервов. Центр регенеративной медицины разрабатывает стратегии для расширения временного окна возможностей и улучшения функционального восстановления после повреждения и восстановления периферических нервов.
Одна из стратегий заключается в применении полимерных микросфер для доставки фактора роста эндотелия сосудов (VEGF) к участку восстановления нерва с контролируемым устойчивым высвобождением. VEGF способствует ангиогенезу и нейрогенезу и, таким образом, приводит к лучшему функциональному результату и большему окну возможностей для нерва, позволяющего продлить регенерацию.
Другая стратегия состоит в том, чтобы противодействовать нехватке здоровых шванновских клеток в месте восстановления нервов путем добавления функционирующих шванновских клеток, полученных из нервов, полученных в системе in vitro, или шванновских клеток, индуцированных из стволовых клеток жировой ткани.
Новые модели животных разрабатываются для определения природы и динамики денервационных мышечных изменений; определить ключевые показатели мышечной восприимчивости, включая электромиографические изменения, изменения типа мышечных волокон и изменения миогенных генов; и оценить влияние этих изменений на регенерацию нерва и потенциальный успех восстановления нерва.
- Рост нервных клеток: аксогенез. Исследователи Мэйо используют рыбок данио в качестве модельной системы на животных, чтобы исследовать, как особые сигналы в головном и спинном мозге могут побуждать или блокировать рост нервных клеток — эксперименты, которые помогают ученым понять, почему условия в месте повреждения нерва замедляют регенерацию. Эта работа дает новое понимание того, как нервные клетки растут в процессе развития нервной системы и как можно улучшить регенерацию нервов после травмы.
Нейрорегенерация инсульта. После инсульта нейроны около полутени уязвимы для отложенного, но прогрессирующего повреждения в результате ишемии.
Не существует эффективного лечения таких умирающих нейронов. Исследователи из Центра регенеративной медицины предположили, что мезенхимальные стволовые клетки (МСК) могут спасать поврежденные нейроны после воздействия стресса кислородно-глюкозной депривации (OGD).Исследования показали, что MSC может дифференцироваться в костную, хрящевую и жировую ткани.Эксперименты на животных моделях геморрагического инсульта показали, что терапия MSC улучшает функцию конечностей. Взятые вместе, эти данные лягут в основу использования MSC для лечения пациентов с недавно перенесенным геморрагическим инсультом.
Нейроонкология и нейрорегенеративные исследования. Исследования в настоящее время сосредоточены на инвазивных опухолях головного мозга (глиомах), по которым пациенты имеют очень плохой прогноз.
Однако есть и другие опухоли головного мозга — олигодендроглиома и астроцитома — с гораздо более благоприятным прогнозом.Исследователи клиники Майо интересуются мутациями, которые участвуют в развитии каждого из этих различных типов опухолей, и почему опухоли ведут себя по-разному.Идентифицирован целевой локус в бедной генами области, первоначально обнаруженный при сканировании генома. Исследовательские усилия сосредоточены на изучении функции этого изменения. Используя мышиные модели, нейральные стволовые клетки мыши и человека и индуцированные человеком плюрипотентные стволовые клетки, исследователи Мэйо изучают, как это изменение влияет на развитие глиальных клеток.
Нейрорегенерация и воспаление. Ограниченная способность нервной системы к восстановлению является серьезной медицинской проблемой. Центр регенеративной медицины разрабатывает новые инструменты для эффективного контроля процесса нервного повреждения и дегенерации и для создания микросреды, которая увеличивает способность к врожденному восстановлению и эффективность других стратегий регенерации, включая замену нервных клеток и нейрореабилитацию.
Исследовательские усилия сосредоточены на том, как можно нацелить протеазы с сильными лекарственными средствами (калликреины), чтобы предотвратить сложный каскад тканевого повреждения и аберрантной реорганизации, который является общепризнанным компонентом травмы CNS и который все чаще признается неотъемлемым фактором, лежащим в основе прогрессирование многих неврологических расстройств, включая те, которые классифицируются как нейродегенеративные или нейровоспалительные, а также те, которые имеют онкогенную основу.
Усилия направлены на понимание физиологических и патофизиологических последствий семейства рецепторов, связанных с G-белком (рецепторы, активируемые протеазой, или PAR), и на определение того, могут ли PAR или протеазы, которые их активируют, быть нацелены терапевтически для предотвращения патогенеза и способствовать пластичности и восстановлению CNS для улучшения функциональных результатов пациентов.
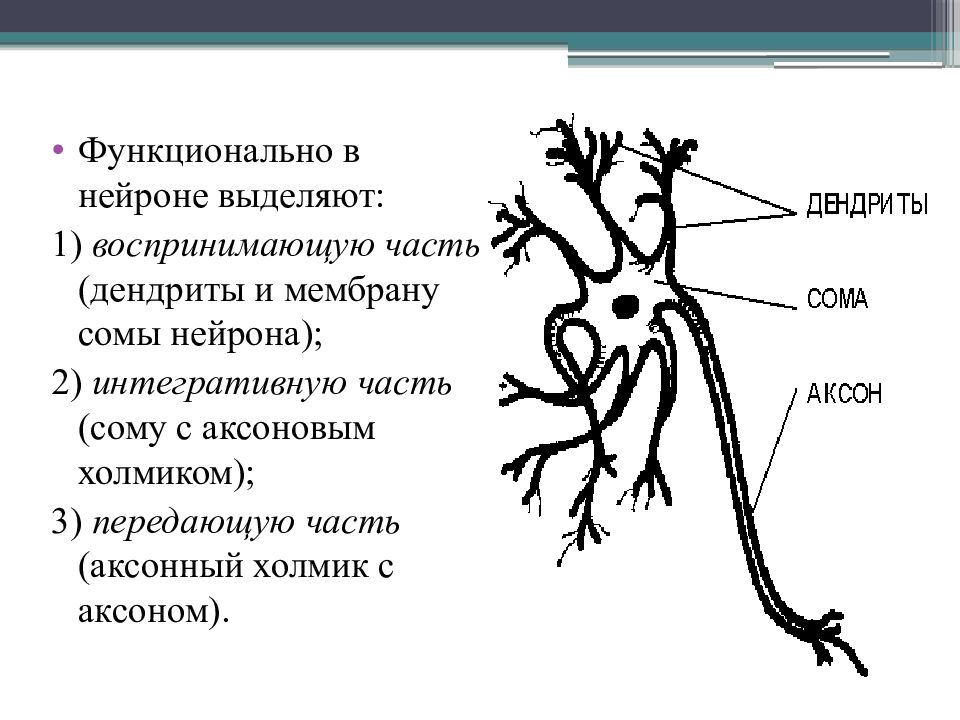 или управляемая факторами регенерация тканей ЦНС .
или управляемая факторами регенерация тканей ЦНС .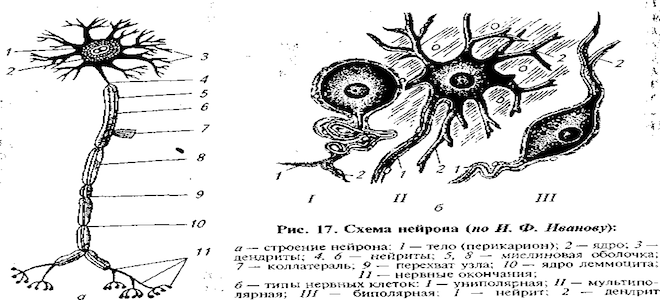

 Не существует эффективного лечения таких умирающих нейронов. Исследователи из Центра регенеративной медицины предположили, что мезенхимальные стволовые клетки (МСК) могут спасать поврежденные нейроны после воздействия стресса кислородно-глюкозной депривации (OGD).
Не существует эффективного лечения таких умирающих нейронов. Исследователи из Центра регенеративной медицины предположили, что мезенхимальные стволовые клетки (МСК) могут спасать поврежденные нейроны после воздействия стресса кислородно-глюкозной депривации (OGD).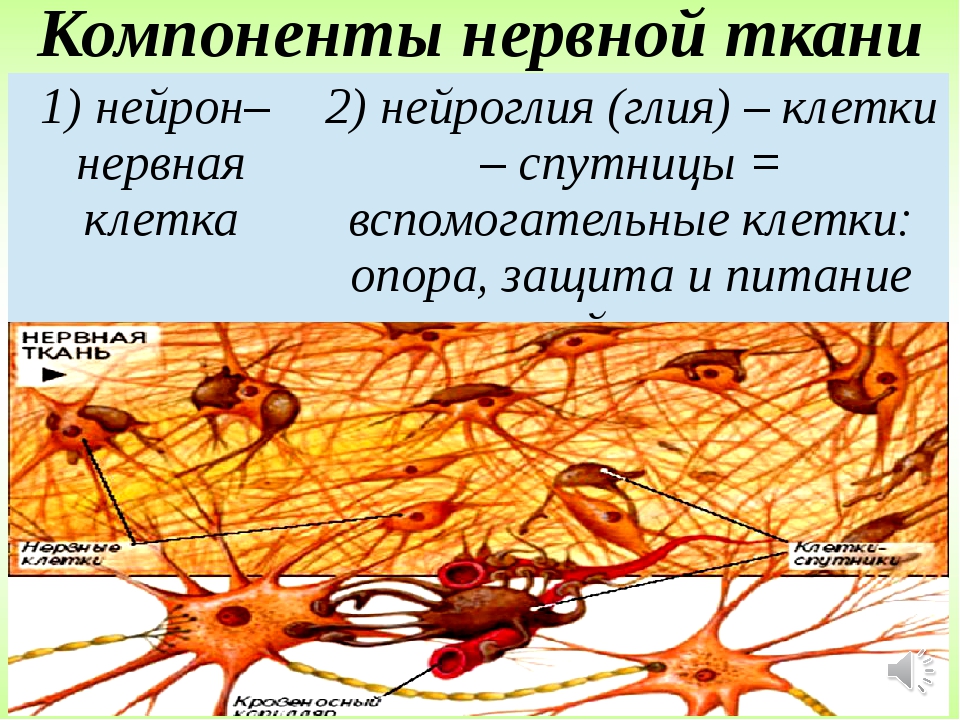 Однако есть и другие опухоли головного мозга — олигодендроглиома и астроцитома — с гораздо более благоприятным прогнозом.Исследователи клиники Майо интересуются мутациями, которые участвуют в развитии каждого из этих различных типов опухолей, и почему опухоли ведут себя по-разному.
Однако есть и другие опухоли головного мозга — олигодендроглиома и астроцитома — с гораздо более благоприятным прогнозом.Исследователи клиники Майо интересуются мутациями, которые участвуют в развитии каждого из этих различных типов опухолей, и почему опухоли ведут себя по-разному.
Методики
Глубокая стимуляция мозга при болезни Альцгеймера. Анекдотические и первоначальные отчеты об испытаниях, касающихся глубокой стимуляции мозга (DBS) сводов и гипоталамуса, были связаны с улучшением функции памяти и снижением ожидаемого снижения когнитивных функций у пациентов с ранней болезнью Альцгеймера. Свод представляет собой основной путь поступления и выхода из гиппокампа и медиальной височной доли.
Исследователи Мэйо начали новаторское пилотное исследование стимуляции двойного полушария субталамического ядра, свода и гипоталамуса, чтобы определить, может ли этот подход иметь положительный эффект в ослаблении когнитивного спада.Если это исследование предоставит положительные данные, тогда будет рассмотрен потенциал использования DBS свода в качестве лечения болезни Альцгеймера.
Детская анестезия, апоптоз и безопасность.
Воздействие нескольких анестетиков в молодом возрасте может быть связано с более поздними проблемами, такими как нарушение обучаемости и синдром дефицита внимания / гиперактивности. Исследователи из Центра регенеративной медицины работают над большим проектом, включающим подробное тестирование 1000 детей, чтобы попытаться лучше определить, какая травма (если таковая имеется) может быть связана с воздействием анестетика.Эта информация будет важна, чтобы увидеть, действительно ли это проблема в клинической практике, и если да, то для изменения практики, чтобы минимизировать проблемы.Исследователи проводят подробное тестирование нервной системы на выборке из когорты новорожденных детей, в том числе на батарее тестов, ранее использовавшихся на приматах, на которых было показано воздействие анестезии. Цель состоит в том, чтобы подтвердить (или опровергнуть) предыдущие результаты и впервые предоставить подробный фенотип связанного с анестезией повреждения (если таковой имеется).
Нейрогенез. Увеличивая понимание молекулярных мишеней, участвующих в регуляции нейрогенеза гиппокампа у взрослых (генерация нейронов) и связанных с ними поведенческих реакций, измененных в невропатологических условиях, ученые могут изучать лежащие в основе клеточные и молекулярные механизмы, которые регулируют производство, созревание и интеграцию новых нейронов в схемы, и как аберрантный нейрогенез играет роль в патогенезе болезни.Исследователи используют поведенческую нейробиологию для количественной оценки познавательных способностей, таких как обучение, память и тревожность.
Признавая терапевтический потенциал нейрогенеза у взрослых, исследователи из клиники Мэйо описывают системы лечения и клинически одобренные лекарства, которые позволяют диктовать нейрональное развитие в правильном направлении. Долгосрочная цель — использовать регенеративную способность нейрогенеза у взрослых для достижения оптимального клинического результата и улучшенных вариантов лечения заболеваний головного мозга.
Нейрореабилитация. Это исследование направлено на улучшение участия и качества жизни людей, функции мозга которых были нарушены в результате травмы или болезни. Основное внимание уделяется регенерации в том смысле, что улучшение поведенческих характеристик возможно только тогда, когда адаптивные анатомические и физиологические изменения происходят внутри и между системами мозга в ответ на терапевтическое вмешательство.
Разрабатывая подходы к лечению, которые приводят к улучшению функций и независимости, исследователи из Центра регенеративной медицины способствуют адаптивным регенеративным изменениям в функции мозга, которые делают возможным улучшение поведенческих характеристик.
Механизмы трансдукции, опосредующие рост двунаправленных нервов. Сигналы, высвобождаемые при распаде миелина после травм головного и спинного мозга, могут действовать как хемопелленты и ингибировать удлинение аксонов, что ограничивает функциональное восстановление. Напротив, положительные сигналы, такие как нейротрофины, могут способствовать удлинению аксонов и вызывать хемоаттракцию.
Это исследование направлено на определение того, как хемотропные сигналы в микросреде направляют рост нервов и как дисфункциональные механизмы управления могут вызывать заболевание.Понимание этих механизмов и открытие методов управления ими важны для разработки новых методов лечения, способствующих регенерации нервов после дегенеративного заболевания или травмы.
Исследователи определяют, как хемотропные сигналы в микросреде направляют рост нервов и как дисфункциональные механизмы управления могут вызывать заболевание. Это позволит ученым определить механизмы передачи пространственно-временного сигнала, с помощью которых конусы роста нервов обнаруживают внеклеточные ориентиры и динамически регулируют клеточные эффекторы, чтобы контролировать направление расширения аксона во время нормального эмбрионального развития и регенерацию нервов после травмы.
В более долгосрочной перспективе целью исследования является определение механизмов прайминга и направления регенерирующих аксонов к соответствующим синаптическим мишеням для завершения функциональных цепей.
 Воздействие нескольких анестетиков в молодом возрасте может быть связано с более поздними проблемами, такими как нарушение обучаемости и синдром дефицита внимания / гиперактивности. Исследователи из Центра регенеративной медицины работают над большим проектом, включающим подробное тестирование 1000 детей, чтобы попытаться лучше определить, какая травма (если таковая имеется) может быть связана с воздействием анестетика.Эта информация будет важна, чтобы увидеть, действительно ли это проблема в клинической практике, и если да, то для изменения практики, чтобы минимизировать проблемы.
Воздействие нескольких анестетиков в молодом возрасте может быть связано с более поздними проблемами, такими как нарушение обучаемости и синдром дефицита внимания / гиперактивности. Исследователи из Центра регенеративной медицины работают над большим проектом, включающим подробное тестирование 1000 детей, чтобы попытаться лучше определить, какая травма (если таковая имеется) может быть связана с воздействием анестетика.Эта информация будет важна, чтобы увидеть, действительно ли это проблема в клинической практике, и если да, то для изменения практики, чтобы минимизировать проблемы.Дополнительная информация
Нейрорегенеративная медицина в клинике Мэйо (PDF)
.
Дефицит кавеолина-1 нарушает синаптическую передачу в нейронах гиппокампа | Молекулярный мозг
Первичная культура нейронов
Области CA1-CA3 гиппокампа были выделены из неонатальных (0–1-дневных) крыс Sprague – Dawley (DBL, код штамма: NTac: SD) и помещены на покрытые полиорнитином покровные стекла.Нейроны трансфицировали через 8 дней после посева и дополнительно инкубировали в течение 14–21 дней в культуральной среде, как описано ранее [8]. Изображение нейронов производили через 14–21 день после посева. Все результаты получены как минимум из трех независимых первичных культур. Все процедуры на животных в этом исследовании выполнялись в соответствии с Руководством по уходу и использованию лабораторных животных с одобрения Институционального комитета по уходу и использованию животных Университета Кён Хи.
Плазмиды и трансфекция
Маленькая шпильковая (ингибирующая) РНК против Cav1 (shRNA-Cav1) с целевой последовательностью, 5′-GAT TGA TCT GGT CAA CCG C-3 ‘[9], была синтезирована, отожжена и лигирована в вектор pSuper в соответствии с инструкциями производителя.Тройной мутант Cav1 с удаленными сайтами пальмитоилирования, Cav1-C133A / C143A / C156A [10], был сконструирован с помощью набора для сайт-направленного мутагенеза (Stratagene) с использованием в качестве матрицы WT Cav1 (Cav1-mCh), меченного mCherry. Для оптической визуализации первичные культивированные нейроны гиппокампа трансфицировали указанными конструкциями с использованием метода осаждения фосфатом Ca 2+ , как описано ранее [8]. Вкратце, плазмиды инкубировали с 2X HEBS (273 мМ NaCl, 9,5 мМ KCl, 1,4 мМ Na 2 HPO 4 · 7H 2 O, 15 мМ D-глюкозы, 42 мМ HEPES pH 7.10), содержащий 2 мМ Ca 2+ , после чего смесь наносили на нейроны гиппокампа, культивированные в течение 8 дней in vitro (DIV8). Для вестерн-блоттинга первичные нейроны трансфицировали электропорацией с использованием системы NEPA21, как описано производителем (Bulldog Bio).
Иммунофлуоресценция
Для иммунофлуоресцентных анализов нейроны DIV14-18 фиксировали 4% параформальдегидом, проницаемостью 0,2% тритоном X-100 и блокировали 5% альбумином бычьей сыворотки (BSA).После инкубации с первичными антителами против синапсина1 (BD Bioscience) нейроны инкубировали с вторичными антителами, конъюгированными с Alexa 488 (Invitrogen).
Маркировка липидных рафтов
Нейрональные липидные рафты метили путем инкубации нейронов DIV14-18 с маркером липидных рафтов GM1 с использованием набора для маркировки липидных рафтов Vybrant Alexa Fluor 594 (Thermo Fisher) в соответствии с инструкциями производителя. Вкратце, нейроны инкубировали в течение 30 минут при 4 ° C с субъединицей B холерного токсина, конъюгированной с Alexa-594 (CT-B), которая специфически связывается с ганглиозидом плазматической мембраны, GM1, неотъемлемой частью липидных рафтов.После этого клетки инкубировали с антителом против CT-B в течение 30 мин при 4 ° C [11].
Оптическая визуализация
Для иммунофлуоресценции изображения фиксированных нейронов получали с помощью микроскопа Leica DMRBE с объективом PL Fluor 40x (1.0 NA.), Оснащенного камерой CoolSNAP HQ (фотометрической), управляемой программным обеспечением MetaMorph. Для тестов pHluorin в нервных окончаниях визуализацию живых клеток проводили на нейронах DIV14-21, трансфицированных Physin-pH, и указанными плазмидами через 8 дней после посева.Покровные стекла помещали в камеру для стимуляции с ламинарным потоком и перфузией на предметном столике изготовленного по индивидуальному заказу эпифлуоресцентного микроскопа с лазерной подсветкой (Zeiss Observer). Изображения живых клеток получали с помощью камеры EMCCD Andor iXon Ultra 897 (модель # DU-897U-CS0- # BV) с задней подсветкой. В качестве источника света использовался лазер OBIS 488 (когерентный) с диодной накачкой и затвор, который затворялся синхронизацией сигнала включения / выключения TTL от камеры EMCCD во время сбора данных. Возбуждение / испускание и сбор флуоресценции достигались с помощью объектива 40 × Fluar Zeiss (1.3 NA), эмиссионные 500–550 нм и дихроичные фильтры 498 нм (Chroma). Потенциалы действия (AP) вызывались пропусканием импульса тока длительностью 1 мс через платино-иридиевые электроды от изолированного стимулятора тока (World Precision Instruments). Нейроны перфузировали буфером Тирода, состоящим из 119 мМ NaCl, 2,5 мМ KCl, 2 мМ CaCl 2 , 2 мМ MgCl 2 , 25 мМ HEPES, 30 мМ глюкозы, 10 мкМ 6-циано-7-нитрохиноксалина-2, 3-дион (CNQX) и 50 мкМ D, L-2-амино-5-фосфоновалериановой кислоты (AP5) с доведением pH до 7.4. Все эксперименты проводились при 30 ° C; все изображения были получены с частотой 2 Гц с выдержкой 50 мс; и все химические вещества были приобретены у Sigma, если не указано иное.
Анализ изображений
Результаты анализов на основе pH-фторина были проанализированы, как описано ранее [12], с небольшими изменениями. Изображения были проанализированы с использованием Image J (http://rsb.info.nih.gov/ij) с плагином Time Series Analyzer, доступным по адресу https://imagej.nih.gov/ij/plugins/time-series.html. Различия в уровнях экспрессии Cav1 в присутствии и в отсутствие shRNA-Cav1 измеряли с помощью Image J.Синаптические бутоны нейронов были выбраны как представляющие интерес овальные области (диаметр, 10 пикселей), и интенсивность флуоресценции на основе pH-флюорина в синапсах была измерена и проанализирована с помощью Origin Pro 2020. Кинетика эндоцитоза и экзоцитоза была подогнана с использованием одного экспоненциального затухания. функция.
Вестерн-блоттинг
Области CA3-CA1 гиппокампа вырезали у однодневных крыс Sprague-Dawley, диссоциировали, высевали на покрытые полиорнитином 6-луночные чашки и инкубировали в течение 14-18 дней.На этапе DIV8 нейроны в трех лунках трансфицировали контрольными плазмидами, а нейроны в трех других лунках трансфецировали shRNA-Cav1. Клетки дополнительно инкубировали до DIV14-18. Все эксперименты проводились параллельно в одинаковых условиях. Клетки лизировали лизисным буфером, содержащим 10 мМ Трис (pH 7,4), 1% SDS, 10 мМ NaF и 1 мМ PMSF, с добавлением смеси ингибиторов протеаз (Complete Mini; Roche, Германия). Концентрацию белка в лизатах определяли с использованием анализа с бицинхониновой кислотой (BCA) (Thermo, IL).Образцы лизата, содержащие равные количества белка, подвергали электрофорезу в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и затем переносили на мембраны из поливинилидендифторида (PVDF). Мембраны блокировали инкубацией с 5% обезжиренным сухим молоком, а затем инкубировали с первичными антителами против Cav1 (BD Biosciences) и против β-актина (Санта-Крус, Калифорния). Интенсивность полос определяли количественно денситометрически, и соотношение [Cav1] / [β-актин] измеряли после вычитания фона.
Статистический анализ
Статистический анализ выполняли с использованием либо одностороннего дисперсионного анализа (ANOVA), либо критерия t Стьюдента, в зависимости от ситуации. Планки погрешностей указывают стандартные ошибки среднего (SEM).
Повторное исследование N-концевых доменов синтаксина-1 при слиянии везикул из центральных синапсов мыши
Существенных изменений (для авторов):
1. Авторы пытаются решить проблему снижения экспрессии белка, снижая уровни WT, используя только 1/12 вирусной нагрузки.Поразительно, но снижение уровней WT STX1A почти в 6 раз не повлияло на синаптическую передачу. Это поднимает важные вопросы. В какой момент STX1A становится ограничивающим скорость? И как эти уровни белка соотносятся с уровнями эндогенного STXA? Авторы должны показать уровни экспрессии своих мутантов относительно эндогенных уровней STX. Более того, в предыдущей статье той же группы сделан вывод, что титрование Synatxin1 действительно влияет на синаптическую передачу (Arancillo et al., J Neurosci 2013). Авторы должны обсудить, как их уровни экспрессии и результаты сравниваются с результатами их предыдущей статьи.
Целью наших экспериментов с понижающим титрованием было не определение того, какой уровень экзогенной экспрессии STX1A ограничивает высвобождение нейротрансмиттера, но мы скорее стремились проверить, сравнивается ли относительное снижение EPSC и других параметров высвобождения в нейронах STX1A LEOpen + ΔΝ с нейронами STX1A LEOpen было связано с относительным снижением их уровней STX1A. Поэтому мы уменьшили количество вирусных частиц, используемых для скважины 35 мм, с ~ 400.От 000 (1X) до ~ 35,000 для STX1A WT и до ~ 65,000 для STX1A LEOpen и успешно достигли уровней STX1A LEOpen + ΔΝ2-28 . На данный момент мы не наблюдали никакого влияния уровней экспрессии на параметры высвобождения ни при использовании STX1A WT , ни при использовании STX1A LEOpen . Таким образом, мы предположили, что снижение параметров высвобождения из-за делеций N-пептида на фоне STX1A LEOpen происходит не из-за его относительно более низких уровней экспрессии, а из-за функционального дефицита.
Тем не менее, мы благодарим рецензентов за их предложение сравнить уровни эндогенного STX1A с уровнями экзогенного STX1A, используемыми в этом исследовании, чтобы устранить возможное несоответствие между этим исследованием и нашим предыдущим исследованием (Arancillo et al., 2013). Важно отметить, что наша линия мышей имеет генотип STX1B FL / FL ; STX1A — / — . В настоящее время у нас нет линии мышей с генотипом STX1B FL / FL ; STX1A +/- для разведения. Следовательно, мы не можем сравнивать уровни эндогенной и экзогенной экспрессии STX1A с использованием однопометников.По этой причине мы проанализировали эндогенные уровни STX1A в нейронах WT по сравнению с лентивирусной экспрессией, полученной с помощью 1X объема лентивирусных частиц, кодирующих STX1A, в нейронах с нулевым значением STX1. Однако мы культивировали как WT, так и трансгенные нейроны в одной партии культур астроцитов и фиксировали их с помощью PFA в одном и том же DIV. Мы наблюдали, что 1X объем вирусных частиц приводил к ~ 3-кратному увеличению экспрессии STX1A по сравнению с эндогенными уровнями (Рисунок 4 — Дополнение 3).Это показывает, что даже самая низкая экспрессирующая конструкция STX1A LEOpen + ΔN2-28 достигает уровня экспрессии ~ 60% эндогенного STX1A WT . Это также говорит о том, что в нашем эксперименте с понижающим титрованием мы снова достигли 60% эндогенных уровней. Мы объяснили и обсудили наши новые данные в связи с нашим предыдущим исследованием (Arancillo et al., 2013) в строках 236-252.
Следует отметить, что нейроны WT также экспрессируют STX1B. Справедливо интерпретировать наши данные как таковые, что наши эксперименты не подпадают под исследования сверхэкспрессии, потому что нейроны, используемые в этом исследовании, не содержат STX1B.Следовательно, примерно в 3 раза более высокая экспрессия STX1A в отсутствие STX1B составляет общий уровень STX1, сравнимый с таковым в нейронах WT. Однако, поскольку STX1A LEOpen + ΔN2-28 действительно демонстрирует 60% экспрессию STX1A, мы удалили наше утверждение, что «высвобождение нейромедиатора предшествует нормальному, даже когда и STX1A, и Munc18-1 демонстрируют крайне низкую экспрессию». Теперь мы включили нашу новую интерпретацию, согласно которой наши эксперименты с понижающим титрованием недостаточны для достижения уровня ограничения скорости, определенного в нашем предыдущем исследовании, и что 60% STX1A в отсутствие STX1B неадекватны для восстановления уровней Munc18-1 (линии 242-249).
2. Хотя влияние N-пептида на большинство параметров высвобождения отсутствует / незначительно, наблюдалось устойчивое влияние на краткосрочную пластичность: до 70% больших EPSC в конце последовательности 10 Гц у мутантов N-пептида. Авторы недооценивают этот существенный эффект. Регулирование STP может быть важной функцией этого домена. Авторы лишь кратко упоминают этот эффект в аннотации, но должны обсудить его биологическое значение в Обсуждении. Во-вторых, типичные следы поездов на Рисунке 6 должны быть более высокого разрешения и увеличены, чтобы их можно было критически оценить.Например, постоянный (асинхронный) ток, который обычно возникает во время 10 Гц в аутапсах, кажется, отсутствует. Почему это так? Поскольку мутации N-пептида не изменяют PPR, наиболее логичным объяснением эффекта во время цуга 10 Гц является разница в накоплении общего Ca во время цуга. Это правдоподобно, учитывая (спорное) взаимодействие синтаксина с кальциевыми каналами. В идеале эксперименты следует повторить +/- EGTA-AM. В этом же эксперименте автор может изучить потенциальное влияние на Ca-зависимое восполнение или увеличение фузогенности (измерение размера RRP и фузогенности до и после 10 Гц).Как минимум, наблюдаемый эффект должен быть более четко обозначен в тексте.
Во-первых, мы благодарим рецензентов за то, что они указали на важность нашего открытия N-пептида STX1A в регуляции STP. Теперь мы обсудили этот вопрос также в нашем обсуждении с большим акцентом (строки 520-538).
Во-вторых, мы теперь включили увеличенные в масштабе примеры абсолютных значений EPSC для первых и последних 5 стимулов для всех групп (Рисунок 6 — Дополнение 2) для лучшей проверки, как предложено рецензентами.На этих увеличенных графиках также можно наблюдать, что стимуляция аутаптических нейронов частотой 10 Гц неадекватна для производства измеримого постоянного тока. Обычно токи, вызываемые 1 AP, достигают базовой линии менее чем за 100 мс, что является временным интервалом для стимуляции 10 Гц. Кроме того, время затухания EPSC влияет на время, необходимое для того, чтобы ток вернулся на базовый уровень, а в аутаптических нейронах более крупные EPSC могут привести к более длительному времени затухания и, таким образом, могут способствовать постоянному току (на основе внутренней информации в наших предыдущих экспериментах с использованием низких концентраций антагонистов AMPA для получения более мелких EPSC).Причина влияния размера EPSC на время распада и, следовательно, на постоянный ток — это перетекание глутамата, потому что аутаптические нейроны не являются полностью изолированной системой, в отличие от культур срезов. Мы понимаем, что составители обзора хотели бы знать влияние делеций N-пептида и мутации LE Open на асинхронное высвобождение. Однако потребуются более сложные эксперименты, такие как использование стронция вместо кальция во внеклеточном растворе (Friedmann and Regehr, 1999, Biophys J).
Наконец, мы хотели бы выразить некоторые из наших опасений относительно использования EGTA-AM для высокочастотной стимуляции. Прежде всего, эксперименты с EGTA-AM требуют предварительной инкубации нейронов в EGTA-AM перед электрофизиологической записью и, таким образом, приводят к плохо контролируемой концентрации этого ca 2+ -буфера в нейронах, что в конечном итоге может испортить результаты высоких частотная стимуляция. В то время как небольшие различия во внутриклеточной концентрации EGTA-AM среди отдельных нейронов одной группы могут быть незначительными, когда различия в чувствительности ca 2+ везикулярного высвобождения и пополнения RRP между двумя группами высоки, они могут сильно влиять на интерпретация данных, когда различия между двумя группами невелики, как в нашем исследовании.Кроме того, увеличение буферизации около 2+ приводит к сложным изменениям в переходных процессах около 2+ во время последовательностей точек доступа. Во время движения примерно 2+ -буфер насыщается, и удаление примерно 2+ с помощью плазмалеммальных насосов нарушается, что приводит к повышенным внутриклеточным уровням примерно 2+ из-за увеличенного времени удаления примерно 2+ . через насосы в конце поезда. Это еще больше усложняет интерпретацию данных 10 Гц с использованием EGTA-AM, особенно когда целью исследования является вскрытие механизмов, ведущих к небольшим изменениям в пополнении STP и RRP.Таким образом, мы предпочитаем не использовать EGTA-AM для изучения потенциального воздействия на зависящее от около 2+ восполнение или увеличение фузогенности. Мы хотели бы сфокусировать наше исследование на том, что N-пептид не является обязательным для высвобождения нейромедиаторов, а скорее регулирует STP и ca 2+ -чувствительность везикулярного высвобождения.
Однако мы обсудили роль N-пептидов в STP более с большим акцентом (строки 520-538), как упоминалось ранее.
3.Основная гипотеза, выдвинутая для снижения чувствительности к кальцию и нарушения восстановления RRP у мутантов N-пептида, заключается в нарушении образования комплекса SNARE. Как это согласуется с увеличением EPSC до 70% во время поездов 10 Гц? Прокомментируйте и обсудите.
Благодарим рецензентов за поднятие этого вопроса. Мы рассчитали чистую депрессию EPSC в течение 10 Гц для делеций N-пептида и увидели, что ответы, зарегистрированные от нейронов STX1A WT , уменьшаются до 60% от начального размера EPSC, а EPSC, записанные от мутантов N-пептида, уменьшаются до 80-90%.Мы также построили график абсолютных EPSC и наблюдали, что EPSC, зарегистрированные от нейронов STX1A ΔN2-19 и STΧ1A ΔN2-28 , не стали больше, а имели тенденцию оставаться меньшими по сравнению с теми, которые были записаны от нейронов STX1A WT (Рисунок 6 — Дополнение 1). Однако, поскольку у STX1A ΔN2-9 был начальный EPSC, сравнимый с таковым у STX1A WT , и поскольку эти нейроны показали кратковременную депрессию всего на 20%, абсолютные EPSC стали примерно на 30% больше по сравнению с таковым у STX1A WT .Мы обсудили этот вопрос в свете их способности образовывать комплекс SNARE и откорректировали нашу интерпретацию (строки 520-529).
4. Данные на рисунке 6 о взаимодействии между двумя способами связывания M18 Stx1 интригуют. Однако различные эффекты мутаций N-пептида на фоне WT и LE_Open трудно согласовать, и они не рассматриваются в дальнейшем в обсуждении. Поэтому трудно интерпретировать эти данные в свете функции N-пептида.Прокомментируйте и обсудите.
Мы уже обсуждали этот вопрос в нашем обсуждении (строки 426-441, особенно строки 437-441). Короче говоря, облегчение образования комплекса SNARE с помощью N-пептида и сужение связанной с Munc18-1 закрытой конформации за счет отсутствия N-пептида являются лишь незначительными (Burkhardt, 2008; Colbert, 2013). Более того, эффект N-пептида может стать незначительным в мембранной среде, когда присутствует TMR STX1 (Dawidowski and Cafiso, 2013).
Мы также предполагаем, что N-пептид регулирует ca 2+ -чувствительность везикулярного высвобождения, что является общим фенотипом делеций N-пептида на фоне WT и LE Open . Однако мы, скорее, остаемся консервативными в этом вопросе и только размышляем об этой «недавно открытой» функции N-пептида.
Поведение5. На рисунке 6F мутации N-пептида в Stx1 LE_Open замедляют депрессию во время первых нескольких стимуляций, но не влияют на STP в конце последовательности.Это противоположно эффекту, обнаруженному на фоне WT, где эти мутации влияют на STP в конце последовательности (рис. 6A). Как согласовать эти, казалось бы, разные результаты, следует обсудить в ходе обсуждения.
STP при высокочастотном стимуле зависит от исходного Pvr, фузогенности и чувствительности ca 2+ SV. В нашем исследовании мы показываем, что делеция N-пептида не влияет на слияние везикул ни на фоне STX1A WT , ни на фоне STX1A LEOpen (рис. 6).Однако мутация LE Open увеличивает степень слияния SV, которая остается высокой даже при удалении N-пептида (фиг. 6). Следовательно, мутанты STX1A LEOpen + ΔΝ демонстрируют высокую степень депрессии при стимуляции 10 Гц. С другой стороны, мы также показываем, что делеция N-пептида снижает чувствительность везикулярного высвобождения к ca 2+ как на фоне STX1A WT , так и на фоне STX1A LEOpen (Фиг.6). Однако снижение чувствительности ca 2+ на фоне STX1A WT относительно невелико по сравнению с таковым на фоне STX1A LEOpen .Это также очевидно в том, что снижения чувствительности ca 2+ на STX1A WT недостаточно для значительного снижения начального Pvr и EPSC. Следовательно, вероятно, что эффект глобального накопления ca 2+ в пресинапсе может наблюдаться только в поздней фазе стимула, когда N-пептид удален на фоне STX1A WT . Однако мутанты STX1A LEOpen + ΔΝ также демонстрируют высокую степень фузогенности, что может свести на нет эффект пониженной чувствительности к ca 2+ в конце последовательности AP.Наши интерпретации по этому поводу можно найти в строках 335-338, 348-350 и 473-491.
6. Учитывая, что амплитуда EPSC и Pvr для Stx1 LE_Open в два раза выше, этот мутант, скорее всего, в конечном итоге будет иметь такие же амплитуды EPSC в конце последовательности, как и для WT. Однако мутанты LE_Open Δ_N будут иметь более низкие амплитуды EPSC в конце последовательности. Это может привести к различным выводам о роли N-пептида, особенно в сочетании с аналогичными экспериментами с этими мутантами, как предложено в пункте 2.
Мы уже показали абсолютные значения EPSC во время стимула 10 Гц для STX1A WT и для всех мутантов (Рисунок 6 — Приложение 1). В то время как STX1A ∆N2-19 и STX1A ∆N2-28 , как правило, по-прежнему вызывали меньшие EPSC на протяжении высокочастотных стимулов по сравнению с таковыми из STX1A WT (Рисунок 6 — приложение 1), EPSC, записанные с STX1A Нейроны ∆N2-9 стали на ~ 30% больше в конце стимула по сравнению с нейронами STX1A WT (строки 290-294).С другой стороны, EPSC, зарегистрированные от нейронов STX1A LEOpen при стимуляции 10 Гц, как правило, изначально были больше, но они продолжали снижаться по сравнению с таковыми у нейронов STX1A WT (Рисунок 6 — приложение к рисунку 1). Мутант STX1A LEOpen + ∆N2-28 оставался производить меньшие EPSC на протяжении высокочастотных стимулов по сравнению с таковыми как у STX1A WT , так и у STX1A LEOpen (рисунок 6 — приложение к рисунку 1) (линии 328-333).
Эти данные подтверждают гипотезу о том, что делеции N-пептида как на фоне STX1A WT , так и STX1A LEOpen снижают эффективность образования комплекса SNARE и, следовательно, приводят к меньшим EPSC на протяжении всей последовательности.Исключением из этого наблюдения является то, что нейроны STX1A ΔΝ2-9 демонстрируют EPSC, изначально сопоставимые с таковыми у STX1A WT , что не согласуется с гипотезой о том, что на чувствительность ca 2+ SV и STP влияет N -пептидные делеции из-за нарушения образования комплекса SNARE. Мы обсудили этот вопрос более подробно в строках (520-535).
7. Интересно видеть на рис. 6J, что при низком уровне внешних Ca, N-пептидных мутаций на фоне LE_open восстанавливается отношение парных импульсов к уровням WT, тогда как при увеличении внешних уровней Ca это постепенно возвращается к фенотипу LE_open.Это должно быть рассмотрено в ходе обсуждения в свете различной чувствительности к кальцию и фузогенности для разных экспериментальных групп.
Мы уже обсуждали этот вопрос в строках 348-350 и предположили, что наблюдение, что делеция N-пептида приводит к измененному поведению только в начальной фазе стимулов 10 Гц — когда открытая конформация STX1A облегчается — согласуется с уменьшением ca 2+ -чувствительность (рис. 6I), но неизмененная фузогенность (рис. 6F) везикул.Рисунок, иллюстрирующий спекулятивную модель эффектов слияния, связывания около 2+ -канал-SV и около 2+ -чувствительности везикулярного высвобождения, можно найти на рисунке 8.
8. Если уменьшение притока Са объясняет пониженное сродство к Са у мутанта LE_open (рис. 6I), то как это согласуется с мутантом LE_open-deltaN, который имеет значительно меньший приток Са по сравнению с WT (рис. 7E), но не имеет существенной разницы в Ca чувствительность? Некоторые пояснения следует добавить в Обсуждение.
Мы хотели бы подчеркнуть, что наши результаты показывают, что мутант LE Open не снижает, а усиливает сродство ca 2+ везикулярного высвобождения, что согласуется с предыдущими сообщениями (Gerber, et al., 2008; Acuna et al., 2014). На рисунке 6I кривая дозовой реакции около 2+ для STX1A LEOpen отклоняется от кривой для STX1A WT со сдвигом влево, показывающим, что нейроны STX1A LEOpen высвобождают большую часть своей высвобождающей способности при более низком уровне около . 2+ -концентрация, поэтому они имеют более высокую чувствительность около 2+ .
ca 2+ -чувствительность везикулярного высвобождения является сложной функцией фузогенности SV и связи на расстоянии около 2+ -канал – SV. Наше применение субнасыщения сахарозы показывает, что STX1A LEOpen приводит к более высокой фузогенности, что способствует более высокой кажущейся чувствительности к ca 2+ . Однако мы не можем сделать вывод из наших экспериментов, что пузырьки состыкованы и праймированы на более близком расстоянии к примерно 2+ -каналов, что требует дальнейшего изучения.Таким образом, мы утверждаем, что даже несмотря на то, что концентрация ca 2+ -каналов может быть снижена в нейронах STX1A LEOpen , что должно быть дополнительно подтверждено дальнейшими экспериментами, ca 2+ -чувствительность может быть увеличена с адекватным ca 2+. Расстояние между каналом и SV и повышенная фузогенность SV.
С другой стороны, мутанты STX1A LEOpen + ∆N могут приводить к увеличению расстояния между пристыкованными / примированными везикулами и каналами ca 2+ , что, вероятно, связано с потерей N-пептида.Поскольку эти нейроны сохраняют высокую фузогенность SV из-за «открытия» STX1A, примерно 2+ -чувствительность слияния SV как запутанная функция фузогенности SV, и дистанционное сцепление примерно 2+ -канал – SV будет приближаться к WT-подобный уровень.
Мы уже обсуждали этот вопрос в нашей предыдущей версии рукописи (предыдущая версия 468-483). Тем не менее, мы включили более подробное обсуждение взаимосвязи между притоком около 2+ , фузогенностью и чувствительностью около 2+ (строки 509-519), и мы надеемся, что сможем решить вашу проблему.Кроме того, рисунок, иллюстрирующий спекулятивную модель эффектов слияния, связывания около 2+ -канал-SV, около 2+ -притока и около 2+ -чувствительности к везикулярному высвобождению, можно найти на рисунке 8.
9. Интерпретация авторов о том, что влияние притока кальция связано с уровнями экспрессии и выживаемостью, несколько оспаривается тем фактом, что мутант LE оказывает самое сильное влияние на приток кальция, хотя он далеко не мутант с самыми низкими уровнями экспрессии. , и при этом не испытывает снижения живучести.Прокомментируйте и обсудите.
Мы не связывали напрямую уровни экспрессии STX1A с уровнем глобального ca 2+ -influx, а скорее предложили это как возможную гипотезу (строки 359-363). Таким образом, мы измерили приток около 2+ от нейронов, экспрессирующих STX1A WT , но на таком же пониженном уровне, как STX1A LEOpen + ∆N2-28 , и не обнаружили никакой разницы. В этом отношении мы интерпретировали наши данные о том, что STX1A LEOpen снижает приток около 2+ из-за функционального нарушения, а не из-за низкого уровня экспрессии (строки 363-372).Приносим извинения, если возникла путаница.
10. На протяжении всей рукописи авторы определяют количество клеток как меру выживаемости. Некоторые мутации, например нокаут синтаксина и спасение мутанта DeltaHabc, сильно влияют на выживаемость клеток. Для интерпретации синаптической физиологии более значимым параметром будет число синапсов, особенно для оценки взаимосвязи между стыковкой (измеряется на профиль синапса) и RRP сахарозы (измеряется на клетку). Кажется возможным, если маловероятным, что синапсы теряются до того, как клетки умирают, и в этом случае корреляция RRP и стыковки может быть сильно затруднена.Авторы могут уже иметь данные в массовых культурах, и должна быть возможность количественно оценить количество синапсов (например, через плотность точек фагота) в этих существующих данных. Это должно сопровождаться обсуждением используемых систем (массовые культуры для морфологии и аутаптические культуры для электрофизиологии). Если номера синапсов изменяются, заряд RRP и количество стыкованных везикул не должны напрямую коррелироваться (как показано на рисунке 5E), потому что заряд RRP сильно зависит от количества синапсов, в то время как количество стыкованных везикул на синапс не коррелирует.Соответственно следует скорректировать обсуждение.
Мы благодарим рецензентов за то, что они подняли вопрос о возможном изменении числа синапсов в нейронах STX1A LEOpen и STX1A LEOpen + ∆N2-28 как возможную причину снижения их RRP. Чтобы проверить эту гипотезу, мы проанализировали наши иммуноцитохимические изображения VGlut1 аутаптических нейронов, используя изображения, которые были показаны ранее на Рисунке 4 — Дополнение 2. Мы не обнаружили разницы ни в количестве синапсов, ни в площади синапсов между STX1A WT , STX1A. LEOpen и STX1A LEOpen + ∆N2-28 нейронов.Мы включили наш результат в рисунок 5 — рисунок в приложении 1, а числовые значения можно найти в соответствующих исходных данных. Поскольку мы не обнаружили уменьшения числа синапсов, кроме RRP, мы сохранили наш первоначальный комментарий относительно корреляции между количеством пристыкованных пузырьков и зарядом RRP. С другой стороны, мы отметили, что стыковка везикул оценивается в массовых культурах, тогда как заряд RRP оценивается в аутаптических культурах. Наше объяснение того, что мы нашли число синапсов, можно найти в строках 265–272.
11. Авторы представляют двойную мутацию открывающего синтаксина (мутант LE) и делецию N-пептида как «прерывание обоих способов связывания Munc18-1» (строка заголовка 185, подписи к рисункам 4 и 5). Хотя мы понимаем, откуда взялась терминология для рассмотрения этого «нулевого значения для связывания Munc18», было бы гораздо лучше оставаться ближе к экспериментам в представлении данных и вместо этого использовать формулировку вроде «открытие синтаксина в сочетании с делецией N-пептида» . Как обсуждают авторы, взаимодействия Munc18-syntaxin сложны, и большинство данных указывает на то, что Munc18 также связывается с комплексами SNARE (и открытым синтаксином) во время их сборки.Открытие синтаксина и удаление N-пептида вряд ли полностью отменяет связывание Munc18, но смещает связывание синтаксина только с Munc18 в сторону связывания Munc18 с синтаксином, поскольку он включается в комплексы SNARE. Авторам следует отдать должное этому моменту в подписях к рисункам и обсудить его как таковой.
Приносим извинения за использование терминологии, которая может ввести читателей в заблуждение. Мы уже отмечали в нашем обсуждении, что Munc18-1, вероятно, все еще связан с комплексами STX1A и / или SNARE, даже когда N-пептид удален в открытой конформации STX1 (строки 416-425).Мы исправили строку заголовка 186 и подписи к рисункам 4 и 5, как предложили рецензенты.
12. Действие мутанта синтаксина LE было охарактеризовано несколькими биофизическими методами, которые показывают, что он сдвигает конформационное равновесие в сторону открытого состояния, но все же принимает закрытое состояние при связывании Munc18, как наблюдали SAXS (Colbert et al. 2013) и одиночная молекула FRET (Wang et al., EMBO J. 36, 816-, 2017). Более того, мутант LE лишь частично компенсирует делецию Munc13, т.е.е., спасение довольно ограничено, как это наблюдается в культурах аутаптических нейронов и в экспериментах с FRET одиночными молекулами (Lai et al., Neuron 95, 591-, 2017). Более того, результаты Wang et al., 2017 предполагают, что мутант LE мешает правильной функции Munc13 (через свои взаимодействия с синтаксином), что может быть связано с наблюдаемыми фенотипами этого мутанта. Пожалуйста, обсудите представленные здесь эксперименты в контексте этих биофизических открытий.
Мы уже обсуждали, что STX1A LEOpen связывается с Munc18-1 в закрытой конформации (строки 418-422).Теперь мы включили ссылки Wang, EMBO, 2017 и Lai, Neuron, 2017 в наше обсуждение этого пункта (строка 420). Мы также включили эти ссылки в наше обсуждение роли Munc18-1 и Munc13 как шаблона для образования комплекса SNARE (строки 444-445). Кроме того, мы обсудили возможное нарушение взаимодействия Munc13-STX1, когда STX1 включает мутацию LE Open (линии 448-454).
https://doi.org/10.7554/eLife.69498.sa2 .