
Наземные экосистемы: Наземная экосистема — это… Что такое Наземная экосистема?
Наземная экосистема — это… Что такое Наземная экосистема?
| В этой статье не хватает ссылок на источники информации. Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена. Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники. Эта отметка установлена 14 мая 2011. |
| Проверить информацию. Необходимо проверить точность фактов и достоверность сведений, изложенных в этой статье. |
Наземные экосистемы играют особую роль в жизни человека, поскольку урожай в них можно получать на всех трофических уровнях в отличие от водных сообществ, где используется только верхняя часть экологической пирамиды. Следовательно, особенности двух типов экосистем необходимо учитывать при эксплуатации природных ресурсов. В экосистемах суши продуценты (автотрофный компонент) представляют собой крупные организмы, у которых от года к году происходит накопление биомассы.

В каждой наземной экосистеме есть абиотический компонент – биотоп, или экотоп – участок с одинаковыми ландшафтными, климатическими, почвенными условиями; и биотический компонент – сообщество, или биоценоз – совокупность всех живых организмов, населяющих данный биотоп. Биотоп является общим местообитанием для всех членов сообщества. Биоценозы состоят из представителей многих видов растений, животных и микроорганизмов. Практически каждый вид в биоценозе представлен многими особями разного пола и возраста.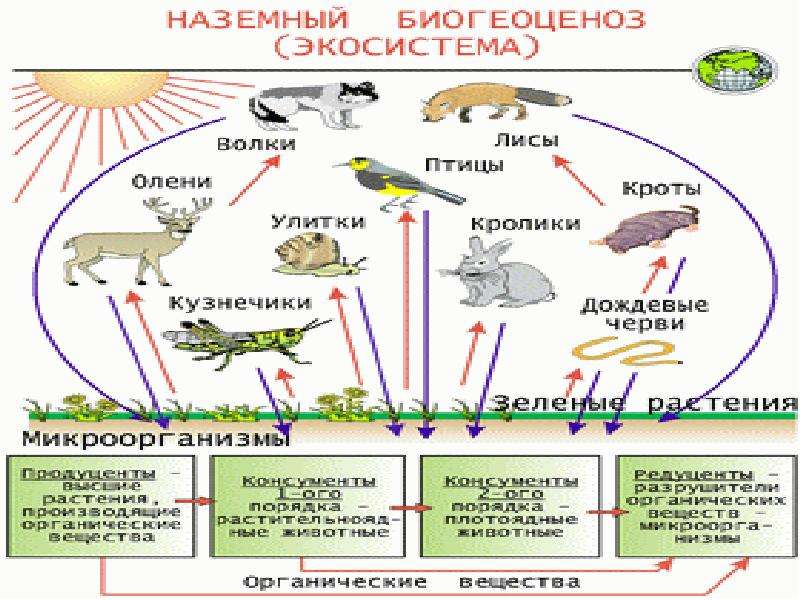
НАЗЕМНЫЕ ЭКОСИСТЕМЫ АРКТИКИ. Соловьев М. Ю.
Природные зоны России
Природные зоны России Арктические пустыни Природные условия Арктика расположена на островах Северного Ледовитого океана и на севере полуострова Таймыр. Арктика-страна льда и тьмы. Сурова природа этой
ПодробнееПрирода Красноярского края.

Природа Красноярского края. Географическое положение Красноярского края. Красноярский Край занимает центральное географическое положение в России и простирается от Северного Ледовитого океана до южных
ПодробнееИнструкция по выполнению работы
Часть 1 К каждому из заданий 1 12 даны четыре варианта ответа, из которых только один правильный. Инструкция по выполнению работы На выполнение теста по географии отводится 45 минут. Абитуриенту разрешается
ПодробнееТема: Природные зоны Евразии.
Тема: Природные зоны Евразии. Цель: Продолжить формирование у учащихся представлений и знаний об особенностях природы Евразии. Задачи: Ознакомить учащихся с особенностями природы и типичными представителями
Глава 1: Что такое экосистема?
Экосистемы БИОЛОГИЯ ЭКОСИСТЕМЫ ЭКОСИСТЕМЫ Глава 1: Что такое экосистема? Что такое экология? Экология это отрасль биологии, изучающая взаимодействие организмов друг с другом и с окружающей их средой.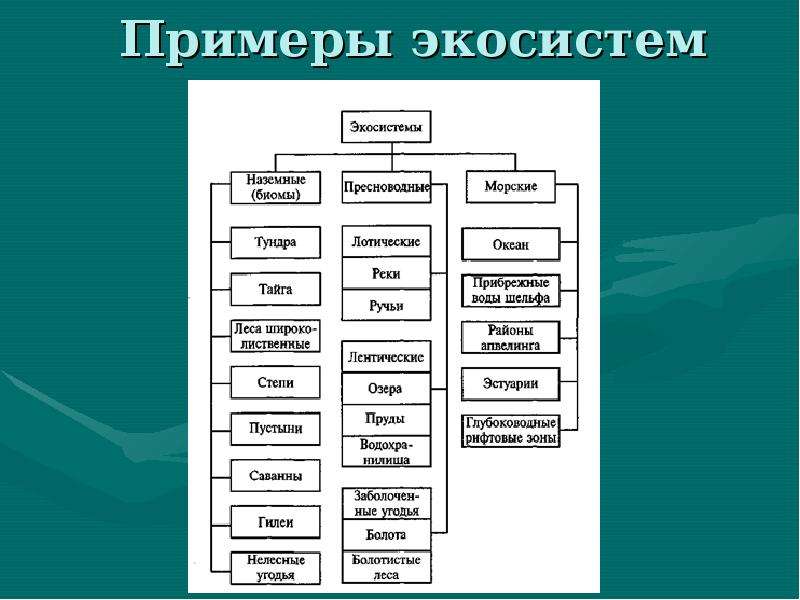 Экологи
Экологи
РИА Новости 28 октября 2015
Климатические изменения в Арктике: прогнозы и риски Г.В. Алексеев ГНЦ РФ «Арктический и антарктический научно-исследовательский институт» РИА Новости 28 октября 2015 СОДЕРЖАНИЕ Арктика в глобальной климатической
ПодробнееРис.1 Кольский полуостров
А.В. Семенов Мурманское управление по гидрометеорологии и мониторингу окружающей среды Г. Мурманск Россия Взаимосвязь изменения климата с гидрологическими характеристиками рек Кольского полуострова Климат
ПодробнееКонтрольная работа «Биосфера».
Разработчик: учитель географии МБОУ «Зимёнковская СОШ» Владимирской области, Муромского района, п. Зимёнки Чубарова Ю.Ю. Контрольная работа «Биосфера». 1 вариант. 1. Какого царства живых организмов не
ПодробнееББК 26.
 89(2Рос)я72 П99
89(2Рос)я72 П99ББК 26.89(2Рос)я72 П99 П99 Пятунин В.Б. География : 8 класс : рабочая тетрадь 2 к учебнику В.Б. Пятунина, Е.А. Таможней «География России. Природа. Население» для учащихся общеобразовательных организаций
Подробнее Контрольная работа по географии 8 класс
Муниципальное общеобразовательное учреждение Средняя общеобразовательная школа 57 Контрольная работа по географии 8 класс Составитель: учитель географии I категории Усольцева О.Г. Тюмень, 2008 Вариант
ПодробнееМОРСКИЕ ЭКОСИСТЕМЫ АРКТИКИ
МОРСКИЕ ЭКОСИСТЕМЫ АРКТИКИ Часть 1 проф. Л.В. Ильяш Границы Арктики Граница распространения деревьев Июльская изотерма 10 о С Арктика наземные и водные экосистемы. Водные экосистемы Северный Ледовитый
Подробнееотзыв на автореферат диссертации Федосова Владимира Эрнстовича «Основные закономерности дифференциации бриофлоры Гипоарктики на примере юго-восточного Таймыра» на соискание ученой степени доктора биологических
МИР РАСТЕНИЙ, 4 КЛАСС
ФАМИЛИЯ КЛАСС — ИМЯ Отметьте верный вариант ответа. Время выполнения работы — 90 минут. Часть I. За каждое верно выполненное задание начисляется 3 балла. 1. Определи, на какой картинке изображено растение,
Время выполнения работы — 90 минут. Часть I. За каждое верно выполненное задание начисляется 3 балла. 1. Определи, на какой картинке изображено растение,
Охрана растительности Тюменской области
Охрана растительности Тюменской области Арктическая тундра Типичная тундра Южная тундра Лесотундра Северная тайга Средняя тайга Южная тайга Лесостепь Типичные тундры дриада точечная осока мечелистная ива
ПодробнееОЛИМПИАДА ПО ГЕОЛОГИИ
Ответы на тесты Вы вносите в специальный бланк для ответов. У Вас есть 90 минут. Пользоваться шпаргалками, сотовыми телефонами, книгами, атласами не разрешается. Разговаривать с другими участниками запрещается.
ПодробнееУЧЕБНО — ТЕМАТИЧЕСКИЙ ПЛАН
УЧЕБНО — ТЕМАТИЧЕСКИЙ ПЛАН Класс: 8А, 8Б Плановых контрольных работ 5 Плановых зачётов Плановых практических работ 4 Лабораторных работ п/п Содержание Общее количество по разделу Лабораторные, практические
Тематическое планирование 5 класс.

п/п Тематическое планирование 5 класс. Раздел Тема урока Колво часов Практические работы Форма контроля 1. Введение 1. Что изучает география. 1 1 КЭС Дом. задание 2. Накопление знаний о Земле. 2. Познание
ПодробнееПояснительная записка
Пояснительная записка Итоговая контрольная работа по курсу географии материков и океанов 7 класс представлена в форме КИМов, составленых в формате ЕГЭ в двух вариантах и включают трех уровней сложности:
ПодробнееФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ Государственное образовательное учреждение высшего профессионального образования «Уральский государственный университет им. А.М. Горького» ИОНЦ «Экология и природопользование»
ПодробнееWhat is an Ecosystem? | Protocol (Translated to Russian)
27.
 1: Что такое экосистема?
1: Что такое экосистема?Обзор
Экосистема — это взаимодействие между всеми абиотическими и биотическими факторами окружающей среды, и их можно разделить на наземные или водные. Наземные экосистемы классифицируются в зависимости от климата, включая годовую температуру, количество осадков и сезонность. Водные экосистемы делятся на пресноводные и морские, а затем по глубине, что влияет на температуру воды и количество солнечного света, проникающего в воду.
Наземные экосистемы
Наземные экосистемы подразделяются на биомы, которые характеризуются как климатом окружающей среды, так и типами организмов, которые там обитают. Области около экватора Земли, которые демонстрируют высокие сезонные температуры и большое количество годовых осадков, являются тропическими лесами и являются одними из самых разнообразных мест обитания в мире. Экосистемы с высокими температурами, но с дефицитом годовых осадков — это пустыни. Хотя пустынные экосистемы обладают этими характеристиками, их биоразнообразие может сильно различаться.
С другой стороны, экосистема с очень низкими годовыми температурами и малым годовым количеством осадков — это арктический биом. У этой экосистемы самое низкое биоразнообразие. Между этими тремя основными биомами находятся экосистемы сезонной тундры, пастбищ умеренного пояса и саванны. Эти экосистемы демонстрируют колебательные годовые изменения как температуры, так и количества осадков, и служат убежищем для животных и растений, специально адаптированных к этим сезонным изменениям.
Пресноводные экосистемы
Водные экосистемы делятся на две категории: пресноводные и морские. Пресноводные экосистемы — самая редкая из экосистем, составляющих около 2% поверхности Земли, — это озера, реки, ручьи и водно-болотные угодья. Основными факторами, влияющими на биоразнообразие в этих экосистемах, являются приток воды, температура и глубина.
Мелководье у краев прудов и озер называется литоральной зоной. Прибрежная зона является самой теплой зоной водоема и включает в себя многих производителей, потребителей и животных, ведущих как наземный, так и водный образ жизни. Лимнетическая зона, или глубоководная поверхность, хорошо освещена и в ней преобладает планктон. Глубокая зона, или глубокая вода, в основном состоит из организмов, которые поедают мертвые организмы и не фотосинтезируют, поскольку в глубину проникает мало света.
Лимнетическая зона, или глубоководная поверхность, хорошо освещена и в ней преобладает планктон. Глубокая зона, или глубокая вода, в основном состоит из организмов, которые поедают мертвые организмы и не фотосинтезируют, поскольку в глубину проникает мало света.
В ручьях и реках вода течет быстро, рециркулируя кислород, а также сохраняя воду относительно прохладной. Разнообразие видов связано с шириной и глубиной реки, при этом более крупные участки реки более разнообразны, чем более узкие участки. Наконец, водно-болотные угодья — это участки неглубокой стоячей воды, такие как болота, топи и трясины, для которых характерна часто жаркая и влажная среда. Эти экосистемы обладают самым высоким биоразнообразием среди всех пресноводных экосистем, включая множество растений, животных, грибов и бактерий.
Морские экосистемы
Морские экосистемы — наиболее распространенные, составляющие почти 75% поверхности Земли, — отличаются от пресноводных экосистем из-за высокой солености воды. Они также разделены на основе глубины воды: мелководье, глубоководье и глубины океана. Самая разнообразная из этих экосистем — мелководье, на котором обитает большая часть коралловых рифов мира и связанных с ними морских обитателей. Помимо мелководья, в глубоководных областях океана также обитает множество морских обитателей — из-за количества проникающего солнечного света — в том числе планктон, который выполняет около 40% всего фотосинтеза на Земле.
Они также разделены на основе глубины воды: мелководье, глубоководье и глубины океана. Самая разнообразная из этих экосистем — мелководье, на котором обитает большая часть коралловых рифов мира и связанных с ними морских обитателей. Помимо мелководья, в глубоководных областях океана также обитает множество морских обитателей — из-за количества проникающего солнечного света — в том числе планктон, который выполняет около 40% всего фотосинтеза на Земле.
Из-за минимального количества света, проникающего в глубины воды, экосистема глубинного океана содержит менее разнообразный, но часто необычный набор организмов, адаптированных к темноте и высоким давлениям. Эта экосистема также наименее известна из всех экосистем из-за огромного пространства, которое она занимает. Однако теперь ученые могут исследовать эту границу благодаря новым технологиям, таким как управляемое человеком устройство Элвин. В настоящее время этот аппарат позволяет исследователям получать доступ к 2/3 дна океана на глубину до 4500 м.
Динамика экосистем
Независимо от характеристик конкретной экосистемы, все они имеют одинаковые динамические процессы , такие как преобразование энергии и материи через фотосинтез и пищевые цепочки между организмами. Экосистемы питаются от постоянного притока солнечной энергии, которая течет через экосистему в то время как материя перерабатывается.
Литература для дополнительного чтения
Murphy, Brett P., Alan N. Andersen, and Catherine L. Parr. “The Underestimated Biodiversity of Tropical Grassy Biomes.” Philosophical Transactions of the Royal Society B: Biological Sciences 371, no. 1703 (September 19, 2016). [Source]
Haddad, Nick M., Lars A. Brudvig, Jean Clobert, Kendi F. Davies, Andrew Gonzalez, Robert D. Holt, Thomas E. Lovejoy, et al. “Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems.” Science Advances 1, no. 2 (March 1, 2015): e1500052. [Source]
Техногенная трансформация наземных экосистем при эксплуатации ракетно-космической техники | Королева
1. Экологический мониторинг ракетно-космической деятельности. Принципы и методы / Под ред. Н.С. Касимова, О.А. Шпигуна. М., Рестарт, 2011. 472 с.
Экологический мониторинг ракетно-космической деятельности. Принципы и методы / Под ред. Н.С. Касимова, О.А. Шпигуна. М., Рестарт, 2011. 472 с.
2. Королева Т.В., Кондратьев А.Д., Кречетов П.П., Семенков И.Н., Шарапова А.В., Черницов О.В. Совершенствование экологических характеристик ракетно-космической техники и мониторинг ее воздействия на окружающую среду. Экология и промышленность России. 2015. Т. 19. № 6. С. 17—23.
3. Кондратьев А.Д., Королева Т.В. Жидкие ракетные топлива: контроль и оценка экологической опасности. Экология и промышленность России. 2017. Т. 21. № 2. С. 45—51.
4. Письмо Министерства охраны окружающей среды и природных ресурсов РФ от 27.12.1993г. №04-25-61-5678 «О порядке определения размеров ущерба от загрязнения земель химическими веществами».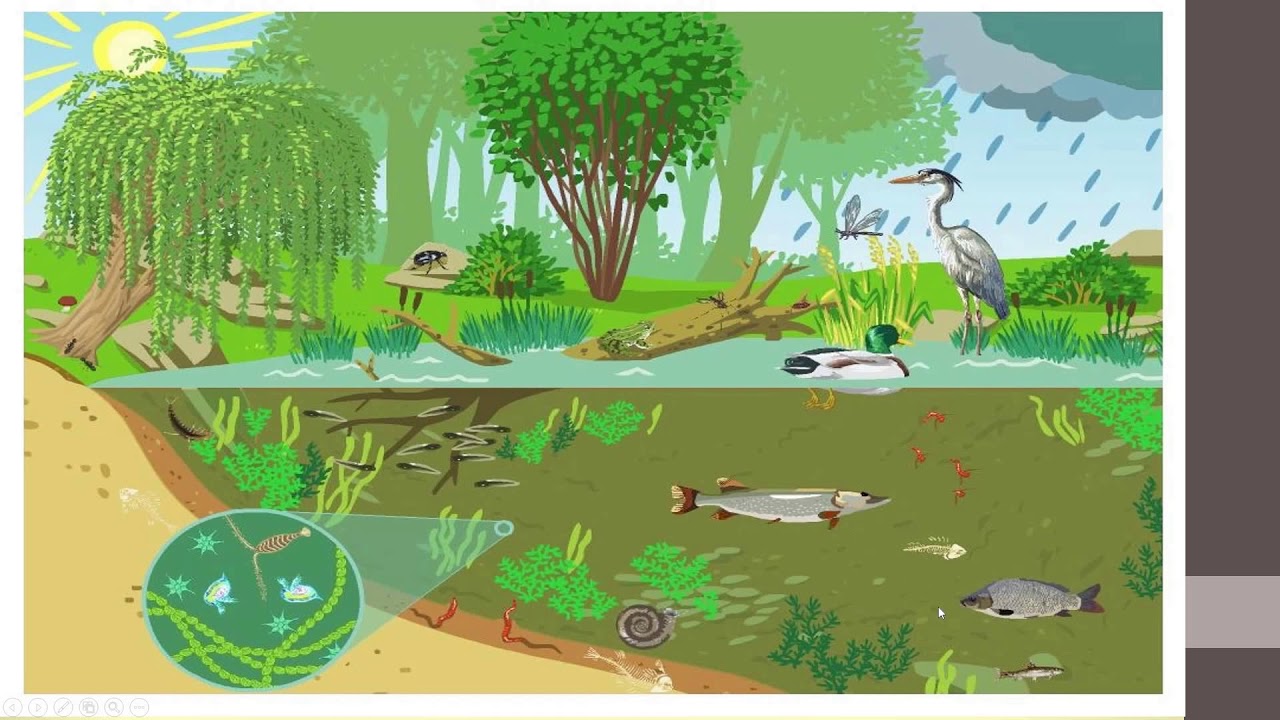 [Электронный ресурс]. URL: http:// legalacts.ru/doc/pismo-minprirody-rossii-ot-27121993-n-04-2561-5678 (дата обращения 20.06.2017 г.).
[Электронный ресурс]. URL: http:// legalacts.ru/doc/pismo-minprirody-rossii-ot-27121993-n-04-2561-5678 (дата обращения 20.06.2017 г.).
5. Экологическая безопасность ракетно-космической деятельности / Под ред. Н.С. Касимова. М., Спутник+ , 2015. 304 с.
6. Архипов В.А., Березиков А.П., Козлов Е.А., Третьяков Н.С., Шереметьева У.Н. Моделирование техногенных загрязнений при отделении ступеней ракет-носителей. Известия высших учебных заведений. Физика. 2005. Т. 48. № 11. С. 5—9.
7. Долотов А.Е., Кузнецов Г.В., Немова Т.Н. Моделирование процесса испарения несимметричного диметилгидразина в атмосфере земли. Известия Томского политехнического университета. 2008. Т. 313. № 4. С. 23—25.
8. Мороков Ю.Н. Моделирование падения в атмосфере остатков ракетного топлива. Вычислительные технологии. 2008. Т. 12. № 2. С. 52—59.
Мороков Ю.Н. Моделирование падения в атмосфере остатков ракетного топлива. Вычислительные технологии. 2008. Т. 12. № 2. С. 52—59.
9. Адам А.М., Архипов В.А., Бурков В.А., Плеханов И.Г., Ткаченко А.С. Влияние метеорологических условий на распространение аэрозольного облака жидких ракетных топлив. Оптика атмосферы и океана. 2008. Т. 21. № 6. С. 504—509.
10. Пузанов А.В., Горбачев И.Н., Архипов И.А. Оценка воздействия РКД на экосистемы Алтае-Саянской горной страны (1998—2010 годы). Мир науки, культуры, образования. 2010. № 5. С. 262—264.
11. Пузанов А.В., Балыкин С.Н., Балыкин Д.Н., Ефременков А.А., Горбачев И.В., Архипов И.А., Винокуров И.А. Результаты экологического мониторинга окружающей среды в районах падения отделяющихся частей ракет-носителей Алтае-Саянской горной страны. Приоритетные задачи экологической безопасности в районах падения Сибирского региона и пути их решения. М., Спутник+, 2016. С. 19—127.
Приоритетные задачи экологической безопасности в районах падения Сибирского региона и пути их решения. М., Спутник+, 2016. С. 19—127.
вопросов больше чем ответов • Алексей Гиляров • Новости науки на «Элементах» • Климат, Экология
Наземные экосистемы, поглощая или выделяя огромное количество парниковых газов (СО2, метана, оксидов азота), тем самым не только участвуют в глобальном круговороте углерода, но и оказывают заметное влияние на климат. Однако не меньшее значение имеет и обратное воздействие — влияние климата на процессы, протекающие в экосистемах. В статье, опубликованной в журнале Nature, Мартин Хайманн (Martin Heimann) и Маркус Райхштайн (Markus Reichstein) из Института биогеохимии Макса Планка (Йена, ФРГ) подчеркивают, что взаимосвязь климата и процессов, протекающих внутри экосистем, остается еще крайне малоизученной. Очевидно только, что эффекты глобального потепления могут экосистемами как ослабляться, так и усиливаться.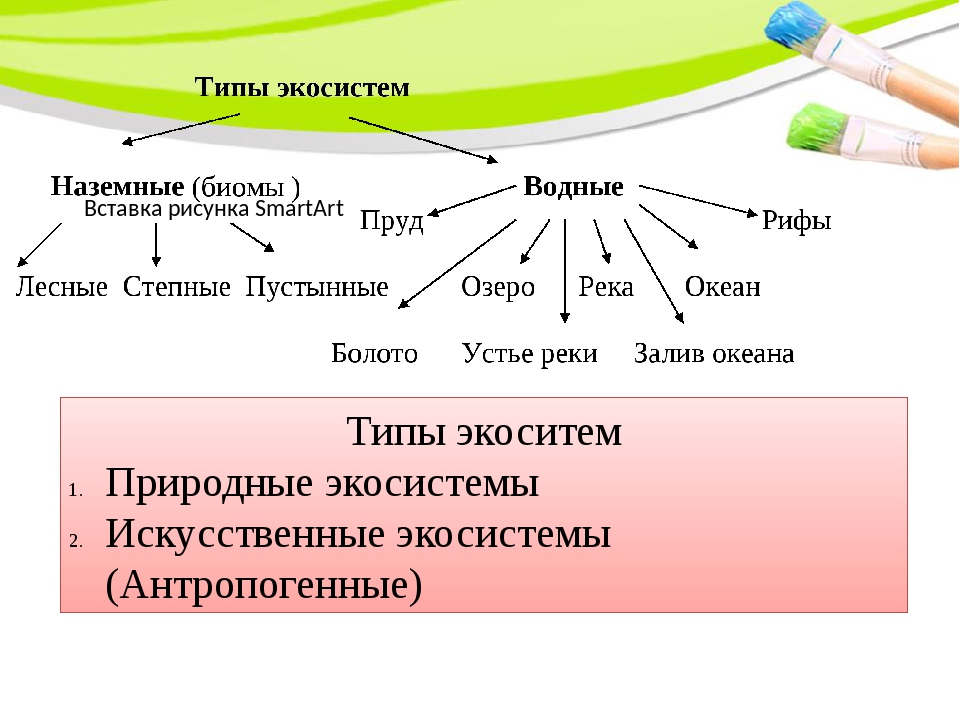
Если в какой-то экосистеме увеличивается масса растительности или запасы органического вещества в почве, то очевидно, что процессы потребления углекислого газа (СО2) преобладают над процессами выделения (экосистема работает как «сток» атмосферного углерода). Если же интенсивнее идут процессы разложения органического вещества, а количество СО2, выделяемого в ходе дыхания грибов и бактерий (а также самих растений), превышает количество СО2, фиксируемого в процессе фотосинтеза, то данная экосистема становится источником атмосферного углерода. Хотя принципы функционирования экосистем (в том числе трансформация ими соединений углерода) в основном понятны, детали протекания отдельных процессов, их количественная характеристика и то, как на них влияют внешние условия, остаются плохо изученными, что и оборачивается низким качеством прогнозов.
Так, например, судя по лабораторным экспериментам, зависимость интенсивности фотосинтеза от концентрации СО2 достаточно проста: по мере увеличения содержания СО2 фотосинтез растет, достигая порога насыщения при концентрации СО2 на уровне 800–1000 ppm (частей на миллион).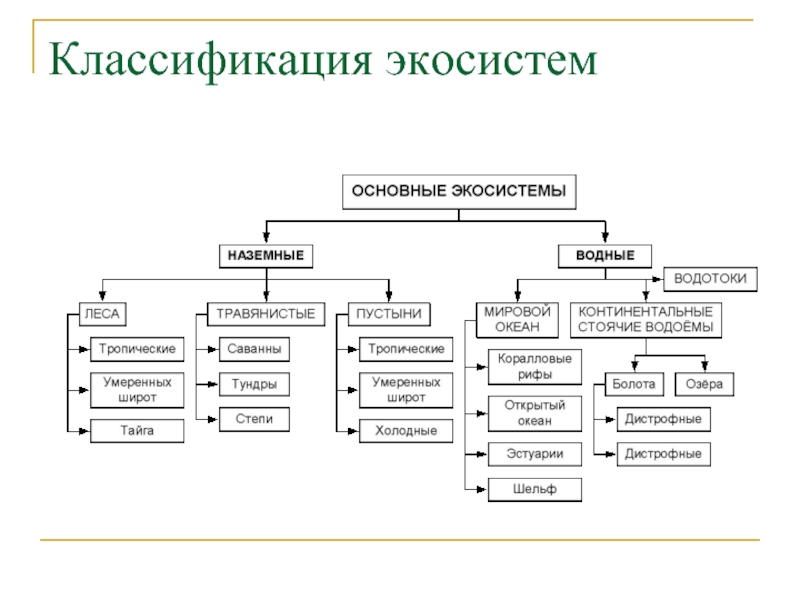 Поскольку современная концентрация составляет 380 ppm, то, казалось бы, существуют еще достаточные возможности противодействия росту СО2. Однако специалисты ожидают, что из-за нехватки элементов минерального питания насыщение углеродом произойдет, видимо, уже при 550–650 ppm СО2. Еще более существенные ограничения на фотосинтез накладывает дефицит влаги. В настоящее время развитие растительности лимитировано нехваткой воды более чем на половине территории суши, и с каждым годом эта доля только возрастает.
Поскольку современная концентрация составляет 380 ppm, то, казалось бы, существуют еще достаточные возможности противодействия росту СО2. Однако специалисты ожидают, что из-за нехватки элементов минерального питания насыщение углеродом произойдет, видимо, уже при 550–650 ppm СО2. Еще более существенные ограничения на фотосинтез накладывает дефицит влаги. В настоящее время развитие растительности лимитировано нехваткой воды более чем на половине территории суши, и с каждым годом эта доля только возрастает.
Параллельно росту содержания СО2 в атмосфере происходит и увеличение средней температуры (глобальное потепление есть уже твердо установленный факт, с которым нельзя не считаться). Поскольку интенсивность дыхания любых организмов при увеличении температуры растет экспоненциально (по закону Аррениуса), то неудивительно, что возрастает и суммарное дыхание экосистемы. В итоге леса тропического пояса (а это самые продуктивные экосистемы мира) становятся местами стока атмосферного углерода только в более влажные годы, а в годы более сухие оказываются местами его выделения. Отсюда — сильные межгодовые колебания содержания СО2 в атмосфере (см. рис. 1), коррелирующие с климатическими особенностями того или иного года, в частности с выраженностью явления Эль-Ниньо (точнее «Эль-Ниньо — Южная осцилляция»), которое сказывается на режиме выпадения осадков в Южной Америке и Юго-Восточной Азии. Однако переход к другим временным масштабам — к десятилетиям и столетиям — делает взаимодействие СО2 и климата куда менее понятным. Отсюда — крайняя неопределенность в оценках того, как уже менялось во времени связывание СО2 наземными экосистемами и каковы прогнозы на будущее (см. рис. 2).
Отсюда — сильные межгодовые колебания содержания СО2 в атмосфере (см. рис. 1), коррелирующие с климатическими особенностями того или иного года, в частности с выраженностью явления Эль-Ниньо (точнее «Эль-Ниньо — Южная осцилляция»), которое сказывается на режиме выпадения осадков в Южной Америке и Юго-Восточной Азии. Однако переход к другим временным масштабам — к десятилетиям и столетиям — делает взаимодействие СО2 и климата куда менее понятным. Отсюда — крайняя неопределенность в оценках того, как уже менялось во времени связывание СО2 наземными экосистемами и каковы прогнозы на будущее (см. рис. 2).
Оценить роль наземных экосистем в круговороте углерода непросто и потому, что в используемых для этого моделях процесс создания органического вещества, протекающий в надземных частях растений, рассматривается совершенно отдельно от процессов разложения органического вещества, происходящих в почве. На самом деле «надземные и «подземные» процессы, конечно, взаимодействуют, но как — не всегда ясно. Особенно плохо известно, что происходит в почве. Если, к примеру, органическое вещество сосредоточено преимущественно в самых верхних слоях, а корни растений располагаются гораздо глубже — там, где больше почвенной влаги или близок водоносный слой, то в случае быстрого наступления засухи разложение органического вещества может остановиться, а фотосинтез будет продолжаться (экосистема будет работать как «сток» углерода). Но если воды будет не хватать корням, то резко затормозится и фотосинтез.
Особенно плохо известно, что происходит в почве. Если, к примеру, органическое вещество сосредоточено преимущественно в самых верхних слоях, а корни растений располагаются гораздо глубже — там, где больше почвенной влаги или близок водоносный слой, то в случае быстрого наступления засухи разложение органического вещества может остановиться, а фотосинтез будет продолжаться (экосистема будет работать как «сток» углерода). Но если воды будет не хватать корням, то резко затормозится и фотосинтез.
Важно учитывать и то, что экосистемы суши реагируют не столько на средние показатели климата, сколько на конкретную обстановку, сложившуюся в определенном месте в определенное время. Так, сильнейшая жара в Европе летом 2003 года привела к столь резкому снижению первичной продукции (сумарного количества органического вещества, произведенного растительностью за определенное время; поскольку вещество это образовано в ходе фотосинтеза, то его можно выражать и в количестве связанного углерода; см. Primary production), что свела на нет результаты накопления органического вещества за предыдущие 4 года (см.: Ciais et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003). Кроме того, после таких сильных засух нередко наблюдается массовая гибель деревьев в последующие года. Жара, особенно если она сопровождается ветрами, резко повышает вероятность возникновения крупных лесных пожаров, в результате которых в атмосферу попадает огромное количество СО2.
Primary production), что свела на нет результаты накопления органического вещества за предыдущие 4 года (см.: Ciais et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003). Кроме того, после таких сильных засух нередко наблюдается массовая гибель деревьев в последующие года. Жара, особенно если она сопровождается ветрами, резко повышает вероятность возникновения крупных лесных пожаров, в результате которых в атмосферу попадает огромное количество СО2.
В своей статье Хайманн и Райхштайн приводят обобщенные схемы трех примеров, иллюстрирующих то, как происходящее внутри экосистем может сказываться на круговороте углерода, в частности на соотношении процессов выделения его в атмосферу и поглощения из атмосферы.
Первый пример (рис. 3a) касается районов вечной мерзлоты, где, по существующим оценкам, в замороженном состоянии находится по крайней мере 400 петаграмм (4 × 1011 тонн; «пета-» означает 1 × 1015) углерода органического вещества. Очевидно, что при повышении температуры воздуха возникает угроза поступления в атмосферу огромного количества метана и СО2. Важно при этом не только прямое нагревание замерзшей почвы солнечными лучами, но и то тепло, которое начинает выделяться изнутри, при разложении бактериями оттаявшего органического вещества (аналогичный разогрев происходит в любой куче гниющих растительных остатков). Таким образом тепловой эффект усиливается, что способствует выделению еще большего количества метана и углекислого газа.
Очевидно, что при повышении температуры воздуха возникает угроза поступления в атмосферу огромного количества метана и СО2. Важно при этом не только прямое нагревание замерзшей почвы солнечными лучами, но и то тепло, которое начинает выделяться изнутри, при разложении бактериями оттаявшего органического вещества (аналогичный разогрев происходит в любой куче гниющих растительных остатков). Таким образом тепловой эффект усиливается, что способствует выделению еще большего количества метана и углекислого газа.
Второй пример (рис. 3b) иллюстрирует ситуацию, когда добавление в систему нового, легко разлагаемого бактериями субстрата (например, глюкозы и даже целлюлозы) приводит к тому, что размножившиеся бактерии принимаются за разложение старого более устойчивого органического вещества. Такое происходит, в частности, когда в ответ на увеличение содержания в атмосфере СО2 стимулируется рост корней растений и выделение ими в среду легко разлагаемых органических веществ. Эти вещества дают пищу бактериям и активизируют тем самым их деятельность по разложению более глубоко лежащих органических веществ, ранее остававшихся стабильными. В результате также может достигаться значительное усиление процесса выделения в атмосферу углерода.
Эти вещества дают пищу бактериям и активизируют тем самым их деятельность по разложению более глубоко лежащих органических веществ, ранее остававшихся стабильными. В результате также может достигаться значительное усиление процесса выделения в атмосферу углерода.
Третий пример (рис. 3c) касается взаимодействия циклов азота и углерода. Во многих случаях рост первичной продукции, произошедшей в условиях более высокой концентрации атмосферного СО2,приводит к дефициту азота, причем не только для самих растений, но и для микроорганизмов, осуществляющих деструкцию органических веществ. В частности, грибы в таких условиях в качестве источника азота начинают активно использовать лигнин (очень стойкое вещество растительных клеточных стенок), который ранее оставался нетронутым. Таким образом в круговорот вовлекается дополнительное количество пребывавшего ранее в почве органического вещества. При этом, естественно, увеличивается и количество СО2, поступающее в атмосферу.
В заключение авторы подчеркивают, что, несмотря на значительную неопределенность в существующих оценках, очевидно, что наземные экосистемы могут работать как усилители воздействия повышенных температур на эмиссию парниковых газов. Однако для получения надежных количественных прогнозов нужны дополнительные данные. В частности, полезными могут оказаться результаты длительных многофакторных экспериментов в сочетании с наблюдениями, не нарушающими структуру экосистем (прежде всего — с оценкой потоков углерода).
Источник: Martin Heimann, Markus Reichstein. Terrestrial ecosystem carbon dynamics and climate feedbacks // Nature. 2008. V. 451. P. 289–292 (текст статьи будет находиться в открытом доступе в течение полугода после публикации).
См. также:
1) P. Ciais et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003 // Nature. 2005. V. 437. P. 529–533.
2) N. Gruber, J. N. Galloway. An Earth-system perspective of the global nitrogen cycle // Nature. 2008. V. 451. P. 293–296 (текст статьи будет находиться в открытом доступе в течение полугода после публикации).
2008. V. 451. P. 293–296 (текст статьи будет находиться в открытом доступе в течение полугода после публикации).
3) Биосфера уже не справляется с избытком СО2, «Элементы», 05.01.2008.
4) Таяние вечной мерзлоты ведет к выбросу в атмосферу миллионов тонн метана, «Элементы», 11.09.2006.
Алексей Гиляров
Урок 7. экосистемный уровень организации жизни. естественные и искусственные экологические системы — Экология — 10 класс
Экосистемный уровень организации жизни. Естественные и искусственные экосистемы
Необходимо запомнить
ВАЖНО!
Экосистема – это совокупность совместно обитающих живых организмов и неорганических компонентов, связанных потоком энергии и круговоротом веществ.
Биогеоценоз – экосистема в пределах фитоценоза (единицы растительного покрова). Понятия «экосистема» и «биогеоценоз» очень близкие, но не являются синонимами. Экосистема – понятие более общее. Каждый биогеоценоз – это экосистема. Но не любую экосистему можно назвать биогеоценозом. Экосистемы бывают естественные и искусственные. Крупные наземные экосистемы называют биомами. Это тундра, тайга, широколиственные и смешанные леса, степи, саванны, пустыни и др. Они соответствуют основным климатическим зонам Земли. Крупные экосистемы распадаются на экосистемы более низкого ранга.
Экосистема – понятие более общее. Каждый биогеоценоз – это экосистема. Но не любую экосистему можно назвать биогеоценозом. Экосистемы бывают естественные и искусственные. Крупные наземные экосистемы называют биомами. Это тундра, тайга, широколиственные и смешанные леса, степи, саванны, пустыни и др. Они соответствуют основным климатическим зонам Земли. Крупные экосистемы распадаются на экосистемы более низкого ранга.
Для осуществления биологического круговорота в структуре любой экосистемы должны присутствовать четыре необходимых компонента: биогенные элементы в окружающей среде и три разные по функциям группы организмов: продуценты, консументы и редуценты. Продуценты по типу питания относятся к автотрофам. Автотрофы используют для синтеза либо энергию солнечного света (фототрофы) или энергию химических связей неорганических веществ (хемотрофы). Гетеротрофы делятся на две группы: консументы и редуценты. Консументы – гетеротрофные организмы, потребляющие органическое вещество и перерабатывающие его в органические соединения собственных тел.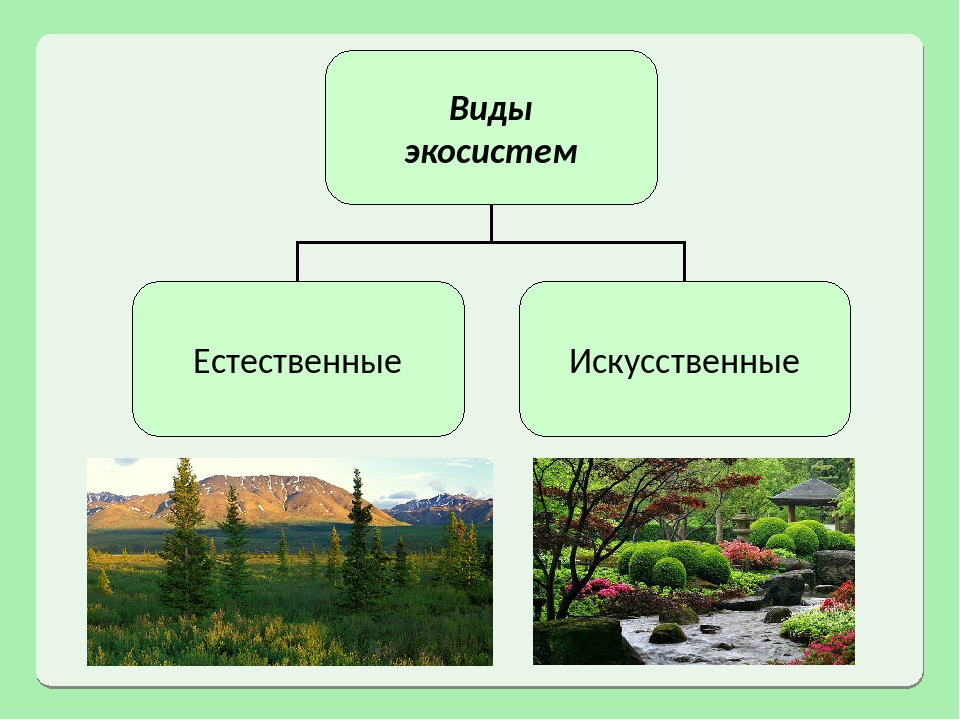 Редуценты – разлагают органические остатки до минеральных веществ. Устойчивые экосистемы – длительно существуют, например: дубрава, ковыльная степь, ельник-кисличник. Нестабильные экосистемы быстро изменяются. Последовательная, закономерная смена одних сообществ другими на определённом участке территории, обусловленная внутренними факторами развития экосистем, называется сукцессией. В результате сукцессионных изменений экосистема становится устойчивой и существует до тех пор, пока внешние силы не выведут её из равновесия. Завершающий, устойчивый этап сукцессии, называется климаксом.
Редуценты – разлагают органические остатки до минеральных веществ. Устойчивые экосистемы – длительно существуют, например: дубрава, ковыльная степь, ельник-кисличник. Нестабильные экосистемы быстро изменяются. Последовательная, закономерная смена одних сообществ другими на определённом участке территории, обусловленная внутренними факторами развития экосистем, называется сукцессией. В результате сукцессионных изменений экосистема становится устойчивой и существует до тех пор, пока внешние силы не выведут её из равновесия. Завершающий, устойчивый этап сукцессии, называется климаксом.
Интерактивная схема классификация экосистем
Различают водные и наземные экосистемы
В настоящее время концепция экосистемы играет весьма важную роль в экологии благодаря гибкости самого понятия: к экосистемам можно относить биотические сообщества любого масштаба с их средой обитания — от пруда до Мирового океана и от пня в лесу до обширного лесного массива — тайги и т. п. Экосистема – основная функциональная единица в экологии.
п. Экосистема – основная функциональная единица в экологии.
В каждой экосистеме существует две главные составляющие:
n биотический компонент – живой;
n абиотический – неживой. Абиотические компоненты (химические вещества, из которых состоят почва, воздух, вода) – преобладают и регулируют жизнедеятельность и само существование биотического компонента, т.е. живых организмов. Абиотические компоненты находятся как вне, так и внутри живых организмов.
С точки зрения трофической структуры, то есть структуры питания, экосистему можно разделить да два яруса:
1) верхний – автотрофный (самостоятельно питающийся) ярус, или «зеленый пояс», включающий растения или их части, содержащие хлорофилл, где преобладают фиксация энергии света, использование простых неорганических соединений и накопление сложных органических соединений;
2) нижний – гетеротрофный (питаемый другими) ярус, или «коричневый пояс» почв и осадков, разлагающихся веществ, корней и т. д., в котором преобладают трансформация и разложение сложных соединений.
С биологической точки зрения, в составе экосистемы удобно выделять следующие компоненты:
1) неорганические вещества (С, N, CO2, H2O и др.), включающиеся в круговороты;
2) органические соединения (белки, углеводы, липиды, гумусовые вещества и т.д.), связывающие биотическую и абиотическую части;
3) воздушную, водную и субстратную среду, включающую климатический режим и другие физические факторы;
4) продуцентов;
Продуценты — организмы, способные синтезировать органические вещества из неорганических с использованием внешних источников энергии. Так как продуценты сами производят органическое вещество, их называют автотрофами — самопитающимися, в отличие от всех остальных организмов, которые называют гетеротрофами — питаемыми другими.
В соответствии с источниками энергии, используемыми для синтеза органического вещества, автотрофы подразделяются на фототрофов (использующих энергию Солнца) и хемотрофов (использующих энергию химических связей, высвобождающуюся в процессе окисления минеральных веществ).
Основную массу фототрофов составляют зеленые растения, в клетках которых содержится хлорофилл и происходит процесс фотосинтеза. К этой категории также относятся цианобактерии и некоторые другие бактерии, проводящие фотосинтез не в хлорофилле, а в иных специализированных пигментах. К хемотрофам относятся только бактерии, окисляющие различные минеральные вещества (нитрофицирующие бактерии, железобактерии, серобактерии и др.).
5) консументов;
Консументы (от лат. konsumo — потребляю) — организмы, не способные строить свои организмы из неорганических веществ и нуждающиеся в готовой органической пище.
К консументам относятся все животные от мельчайших примитивных до самых совершенных, включая человека. Есть консументы и среди растений: это паразитирующие на других растениях. Существуют также растения со смешанным типом питания, например, росянки.
Среди консументов-животных выделяют растительноядных животных (консументы первого порядка), мелких и крупных хищников (консументов второго, третьего порядка и др. ).
).
Консументы также подразделяют на сапрофагов (питающихся мертвыми растительными остатками), фитофагов (поребителей живых растений), зоофагов (нуждающихся в живой пище) и некрофагов (трупоядных животных). Кроме того, организмы, питающиеся мертвыми остатками растений и животных — детритом, дополнительно выделяют в группу детритофагов.
6) редуцентов;
Редуценты (от лат. reducere — возвращать) — организмы, использующие в качестве пищи органическое вещество и подвергающие его минерализации. Поэтому данная категория организмов также называется деструкторами, ибо они окончательно разрушают органические вещества до относительно простых неорганических соединений, используемых консументами в качестве пищи. Тем самым осуществляется возврат вещества в начало природной цепи питания.
К редуцентам относятся многие виды бактерий и грибов, разлагающих в процессе метаболизма мертвое органическое вещество (трупы животных, гниющие растения, фекалии) до минеральных составляющих. Именно они (редуценты) завершают биологические циклы вещества в биосфере, возвращая в почву, воду и воздух биогены (С02, минеральные соли, воду, сероводород, азот и др.), которые вновь могут быть использованы растениями.
Именно они (редуценты) завершают биологические циклы вещества в биосфере, возвращая в почву, воду и воздух биогены (С02, минеральные соли, воду, сероводород, азот и др.), которые вновь могут быть использованы растениями.
Закономерности, характерные для экосистем:
n чем разнообразнее условия заселенного участка (биотопа) в пределах экосистемы, тем больше видов его населяет;
n чем больше видов содержит экосистема, тем меньше особей насчитывают соответствующие видовые популяции, например, в лесах видовое разнообразие животных значительное, а популяции относительно малочисленны;
n чем больше видов содержит экосистема, тем больше ее экологическая устойчивость;
n эксплуатируемые человеком системы, представленные одним или очень малым числом видов (например, поля для выращивания сельскохозяйственных культур) неустойчивы по своей природе и не могут самоподдерживаться.
n никакая часть экосистемы не может существовать без другой. Если, например, исчезает какая-либо группа организмов, вид, то по закону цепных реакций может разрушиться все сообщество.
n стабильность экосистемы — способность экосистемы сохранять свою структуру и функции при воздействии внешних и внутренних факторов; Гомеостаз экосистемы- способность биологических систем (организма, популяции и экосистем) противостоять изменениям и сохранять равновесие. Например, у пойкилотермных животных изменение температуры тела регулируется специальным мозговым центром, куда постоянно поступает сигнал обратной связи, содержащий данные об отклонении от нормы, а от центра поступает сигнал, возвращающий температуру к норме;
n Любая экосистема способна к саморегулированию. Это значит, что в экосистеме во времени и пространстве поддерживаются основные параметры и она находится в состоянии динамического равновесия.
В. Н. Сукачев в 1944 г. учение о «биогеоценозе».
Биогеоценоз (от греч. bios — жизнь, гео — Земля, ценоз — сообщество) – это совокупность на известном протяжении земной поверхности однородных природных явлений (атмосферы, горной породы, растительности, животного мира и мира микроорганизмов, почвы и гидрологических условий), имеющая специфику взаимодействий слагающих ее компонентов и определенный тип обмена веществом и энергией между собой и с другими явлениями природы.
Понятие «экосистема» и «биогеоценоз» близки друг к другу, но не являются синонимами. По определению А. Тэнсли, экосистемы – это безразмерные устойчивые системы живых и неживых компонентов, в которых совершается внешний и внутренний круговорот веществ и энергии. Таким образом, экосистема – это и капля воды с ее микробным населением, и горшок с цветком, и космический пилотируемый корабль, и индустриальный город. Под определение биогеоценоза они не подпадают, так как им не свойственны многие признаки этого определения.
Экосистема может включать несколько биогеоценозов.
Таким образом, понятие «экосистема» шире, чем «биогеоценоз», то есть любой биогеоценоз является экологической системой, но не всякая экосистема может считаться биогеоценозом, причем биогеоценозы – это сугубо наземные образования, имеющие свои четкие границы. Контур биогеоценоза устанавливается по границе растительного сообщества (фитоценоза).
Биогеоценоз включает две главные составляющие: совокупность на определенной территории абиотических факторов, то есть экотоп, и совокупность живых организмов – биоценоз.
Термин «биоценоз» был предложен немецким зоологом К. Мебиусом и обозначает организованную группу популяций растений, животных и микроорганизмов, приспособленных к совместному обитанию в пределах определенного объема пространства.
Биоценоз включает в себя сообщества животных (зооценоз), растений (фитоценоз) и микроорганизмов (микробиоценоз).
Любой биоценоз занимает определенный участок абиотической среды. Биотоп — пространство с более или менее однородными условиями, заселенное тем или иным сообществом организмов.
В свою очередь экотоп состоит из совокупности климатических факторов – климатопа и из совокупности почвенно-грунтовых факторов – эдафотопа.
ФИТОЦЕНОЗ + ЗООЦЕНОЗ + МИКРОБИОЦЕНОЗ = БИОЦЕНОЗ.
Одно из важнейших свойств биогеоценоза – взаимосвязь и взаимозависимость всех его компонентов.
Для наглядности представления взаимоотношений между организмами различных видов в различных системах принято использовать экологические пирамиды. Трофическую структуру системы можно изобразить графически, в виде так называемых экологических пирамид. Основанием пирамиды служит уровень продуцентов, а последующие уровни питания образуют этажи и вершину пирамиды.
Трофическую структуру системы можно изобразить графически, в виде так называемых экологических пирамид. Основанием пирамиды служит уровень продуцентов, а последующие уровни питания образуют этажи и вершину пирамиды.
Известны три основных типа экологических пирамид: 1) пирамидa численности, 2) пирамида биомассы, 3) пирамида продукции (или энергии).
Пирамида численности отображает количество особей, составляющих последовательный ряд звеньев от продуцентов к консументам, неуклонно уменьшается.
Пирамида биомасс четко указывает на количество живого вещества (биомассу — суммарную массу организмов) на каждом трофическом уровне. В наземных экосистемах действует следующее правило пирамиды биомасс: суммарная масса растений превышает массу всех травоядных, а их масса превышает всю биомассу хищников.
Самым фундаментальным способом отражения связей между организмами разных трофических уровней и функциональной организации биоценозов является пирамида энергий, в которой размер прямоугольников пропорционален энергетическому эквиваленту в единицу времени, т. е. количеству энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за принятый период.
е. количеству энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за принятый период.
В процессе жизнедеятельности системы создается и расходуется органическое вещество, т. е. соответствующая экосистема обладает определенной продуктивностью биомассы. Биомассу измеряют в единицах массы или выражают количеством энергии, заключенной в тканях.
Продуктивность — это скорость производства биомассы в единицу времени, которую нельзя взвесить, а можно только рассчитать в единицах энергии или накопления органических веществ.
Принято выделять четыре последовательные ступени (или стадии) процесса производства органического вещества:
1. валовая первичная продуктивность — общая скорость накопления органических веществ продуцентами (скорость фотосинтеза), включая те, что были израсходованы на дыхание и секреторные функции. Растения на процессы жизнедеятельности тратят примерно 20% производимой химической энергии;
2. чистая первичная продуктивность — скорость накопления органических веществ за вычетом тех, что были израсходованы при дыхании и секреции за изучаемый период. Эта энергия может быть использована организмами следующих трофических уровней;
чистая первичная продуктивность — скорость накопления органических веществ за вычетом тех, что были израсходованы при дыхании и секреции за изучаемый период. Эта энергия может быть использована организмами следующих трофических уровней;
3. чистая продуктивность сообщества — скорость общего накопления органических веществ, оставшихся после потребления гетеротрофами-консументами (чистая первичная продукция минус потребление гетеротрофа- ми). Она обычно измеряется за какой-то период; например, вегетационный период роста и развития растений или за год в целом;
4. вторичная продуктивность — скорость накопления энергии консументами. Ее не делят на «валовую» и «чистую», так как консументы потребляют лишь ранее созданные (готовые) питательные вещества, расходуя их на дыхание и секреторные нужды, а остальное превращая в собственные ткани.
Принцип Ле Шателье – Брауна:
n При любом внешнем воздействии, которое выводит систему из состояния равновесия, в системе усиливаются те процессы, которые ослабляют это воздействие, т.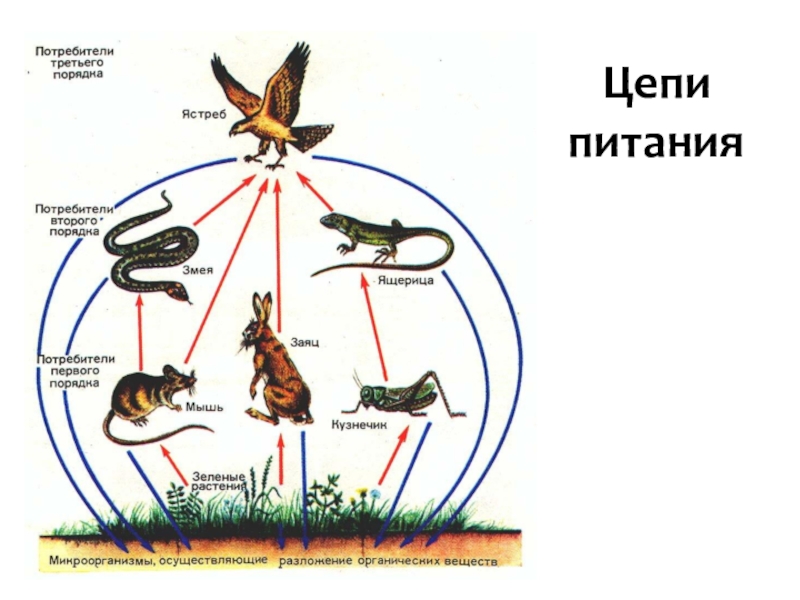 е. система стремиться вернуться в состояние равновесия.
е. система стремиться вернуться в состояние равновесия.
n Способность экосистемы возвращаться в исходное состояние после воздействия каких либо факторов, которые выводят ее из равновесия, называют устойчивостью экосистемы
Правило одного процента:
n Изменение энергии природной экосистемы в среднем на 1% (от 0,3 до 1%) выводит экосистему из состояния равновесия.
n Из этого правила следует, что относительно безопасный уровень потребления ресурсов биосферы не должен превышать 1%;
n Сегодня этот показатель составляет около 10%.
Наземные экосистемы — обзор
Введение
Наземные экосистемы занимают 144 150 000 км 2 (28%) поверхности Земли и состоят из сложной совокупности взаимодействующих сообществ с тысячами видов животных, растений, грибов и бактерий . Организмы в наземных экосистемах объединены в функциональную единицу особыми, динамическими отношениями из-за связанных процессов энергии и химического потока. В почве эта сложная сеть жизни отвечает за круговорот углерода, за воду и питательные вещества, за качество почвы, а также за питание и здоровье растений.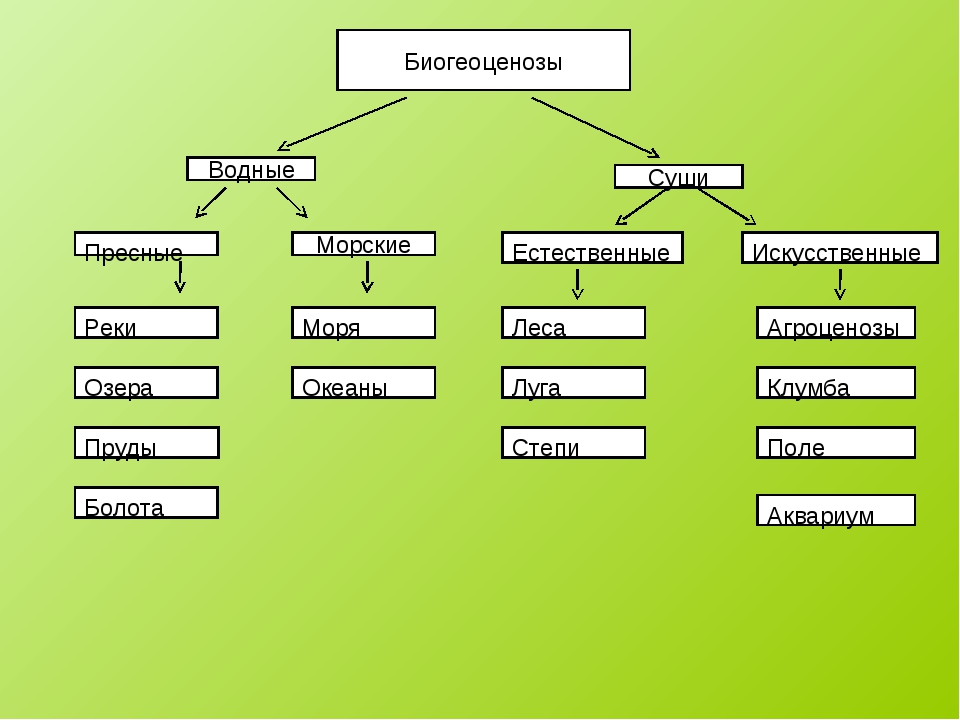 Чтобы полностью понять биологический и химический способ функционирования этих сложных взаимодействий, нужно рассматривать организмы не только как индивидуумы, но и как членов более крупного сообщества, учитывая взаимодействие и коммуникации между людьми внутри этих запутанных популяций, то есть их расширенный фенотип (Whitham et al. ., 2008). Одной из новых моделей таких исследований является взаимодействие между почвенными грибами и растительными сообществами (Martin et al., 2011).
Чтобы полностью понять биологический и химический способ функционирования этих сложных взаимодействий, нужно рассматривать организмы не только как индивидуумы, но и как членов более крупного сообщества, учитывая взаимодействие и коммуникации между людьми внутри этих запутанных популяций, то есть их расширенный фенотип (Whitham et al. ., 2008). Одной из новых моделей таких исследований является взаимодействие между почвенными грибами и растительными сообществами (Martin et al., 2011).
Грибы не только играют центральную роль в глобальном углеродном цикле, но и составляют основную группу патогенов растений в управляемых и естественных экосистемах и служат симбионтами как с гетеротрофными, так и с автотрофными организмами.Экологический успех и важность грибов для жизни на Земле, безусловно, можно объяснить большим разнообразием производимых ими ферментов и метаболитов, что дает им широкий спектр режимов питания и доступ к огромному разнообразию источников углерода и экологических ниш. Понимание того, как развивались и работают сапротрофные, симбиотические и патогенные грибы, является фундаментальным для понимания их экологических функций и их последующего влияния на судьбу растительных сообществ (Martin et al., 2011).
Понимание того, как развивались и работают сапротрофные, симбиотические и патогенные грибы, является фундаментальным для понимания их экологических функций и их последующего влияния на судьбу растительных сообществ (Martin et al., 2011).
Определение последовательности ДНК и РНК занимает центральное место во многих биологических исследованиях. ДНК была впервые секвенирована в 1977 году, с тех пор технологии позволили нам перейти к геномике — изучению полной последовательности генома организма. Первым свободноживущим организмом, геном которого был полностью секвенирован, была бактерия Haemophilus influenzae в 1995 году. В 1996 году Saccharomyces cerevisiae (пекарские дрожжи) были первой последовательностью эукариотического генома, а в 1998 году — первой последовательностью генома для многоклеточного эукариот, Caenorhabditis elegans , был выпущен.С тех пор были созданы сотни эукариотических геномов, включая геном человека. Из-за их относительно небольшого размера генома, роли патогенов человека / сельскохозяйственных культур и важности в области биотехнологии на сегодняшний день секвенировано более 200 геномов грибов. Однако в настоящее время наблюдается значительная предвзятость в отношении видов, являющихся патогенами человека. Тем не менее, многие из грибов, чьи геномы были секвенированы, являются обитателями почвы и растений (Martinez et al., 2004, 2009; Martin et al., 2008, 2010; Ом и др., 2010; Spanu et al., 2010; Stajich et al., 2010) и их геномные последовательности составят основу для изучения геномов и функций тысяч видов почвенных грибов, которые нельзя культивировать и секвенировать напрямую.
Однако в настоящее время наблюдается значительная предвзятость в отношении видов, являющихся патогенами человека. Тем не менее, многие из грибов, чьи геномы были секвенированы, являются обитателями почвы и растений (Martinez et al., 2004, 2009; Martin et al., 2008, 2010; Ом и др., 2010; Spanu et al., 2010; Stajich et al., 2010) и их геномные последовательности составят основу для изучения геномов и функций тысяч видов почвенных грибов, которые нельзя культивировать и секвенировать напрямую.
Набор экспрессируемых или транскрибируемых генов, известный как профиль экспрессии или транскриптом, возможно, является наиболее важным фактором в определении фенотипа и клеточных функций. Транскриптом представляет собой полный набор транскриптов в клетке и их количество для конкретной стадии развития или физиологического состояния.В отличие от генома, транскриптом очень динамичен и может очень быстро подвергаться радикальным модификациям в ответ на изменения окружающей среды или во время развития.
Несмотря на важные технологические достижения, достигнутые в последнее время, анализ паттернов экспрессии многих генов в ответ на изменение окружающей среды или процесс дифференциации является сложным. Были разработаны различные технологии для вывода и количественной оценки транскриптома, включая подходы на основе гибридизации или последовательностей.Такие технологии включают секвенирование тегов экспрессируемых последовательностей (EST), полиморфизмы длины амплифицированных фрагментов для анализа мРНК, серийный анализ экспрессии генов и очень широко используемые микроматрицы. Основное предположение, лежащее в основе стратегии, основанной на массовой идентификации мРНК, продуцируемых в определенных условиях, заключается в том, что гены со сходным поведением экспрессии (увеличивающиеся и уменьшающиеся при аналогичных обстоятельствах), вероятно, будут связаны функционально. Таким образом, генам без предшествующих функциональных назначений можно предварительно отвести роль в биологическом процессе на основе известных функций генов в том же кластере экспрессии.
В отличие от методов микроматрицы, подходы, основанные на последовательностях, непосредственно определяют последовательность кДНК. Первоначально использовалось секвенирование по Сэнгеру библиотек кДНК или EST, но этот подход является относительно малопроизводительным, дорогим и, как правило, не количественным. Относительно недавно разработка новых высокопроизводительных методов секвенирования ДНК предоставила новый метод как для картирования, так и для количественной оценки транскриптомов. Этот метод, получивший название RNA-Seq (секвенирование РНК), имеет явные преимущества по сравнению с существующими подходами и революционизирует способ анализа транскриптомов эукариот.
Непрерывное совершенствование технологий секвенирования генома и связанное с этим снижение затрат были настолько значительными, что позволили нам определить состав и динамику грибковых почвенных сообществ (Buée et al., 2009; Öpik et al., 2009; Jumpponen et al. др., 2010). Несколько 100 видов активны в почвах, и текущие исследования метагеномики и метатранскриптомики позволят выявить функции, закодированные в их геномах, а также в их экспрессируемых транскриптах (Martin and Martin, 2010).
Обилие геномных данных выдвинуло царство грибов на передний план эукариотической геномики.В то время как некоторые из секвенированных видов тесно связаны, другие разошлись 1 миллиард лет назад. Это позволяет нам использовать грибы для изучения эволюционных механизмов, связанных со структурой, организацией и содержанием эукариотического генома. Более того, прямое сравнение между двумя или более близкородственными видами, процесс, называемый сравнительной геномикой, позволяет нам определить метаболические и регуляторные пути и, следовательно, лучше понять образ жизни, связанный с сапротрофизмом, симбиозом, патогенезом.
Геномная энциклопедия грибов JGI FGP (http: // www.jgi.doe.gov/fungi) нацелены на грибковые геномы в трех областях: здоровье растений, биопереработка и разнообразие грибов. Как ясно указано Мартином и др. (2011), здоровье растений зависит от взаимодействия растения как с грибковыми мутуалистами, так и с паразитами. Находясь на поверхности, оба воздействия на хост ортогональны; они могут иметь общие характеристики, позволяющие избежать системы защиты растений и получить питательные вещества хозяина, жить внутри хозяина и поддерживать баланс между сапробным и паразитическим (или мутуалистическим) образом жизни (Martin et al.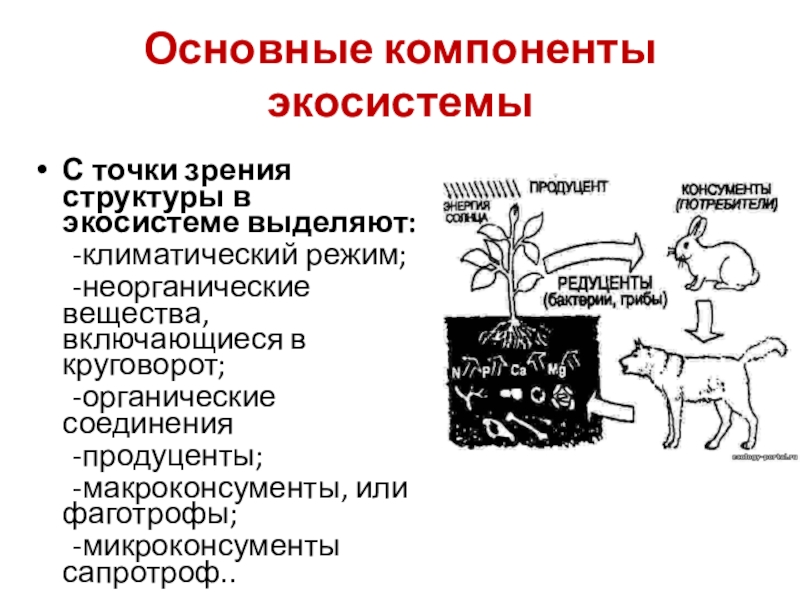 , 2011). Таким образом, наше понимание видов Trichoderma выиграет от достижений в области функционального геномного анализа патогенов и паразитов растений.
, 2011). Таким образом, наше понимание видов Trichoderma выиграет от достижений в области функционального геномного анализа патогенов и паразитов растений.
Являясь повсеместным и часто преобладающим компонентом микофлоры в многочисленных почвах (местных и сельскохозяйственных) во всех климатических зонах, виды Trichoderma играют важную роль в здоровье экосистемы (Klein and Eveleigh, 1998). Они тесно взаимодействуют с корнями растений, колонизируя внешние слои эпидермиса, и действуют как условно-патогенные, авирулентные симбионты растений, но также были обнаружены колонизирующие органы надземных растений (Shoresh et al., 2010). Однако способность этих грибов распознавать, вторгаться и уничтожать другие грибы была основной движущей силой их коммерческого успеха в качестве биопестицидов (Verma et al., 2007). В дополнение к защите, которую они обеспечивают растениям, убивая другие грибы и некоторые нематоды, эти грибы вызывают устойчивость к патогенам растений, вызывают устойчивость к абиотическому стрессу, улучшают приспособленность растений, солюбилизируют питательные вещества для растений и биовосстановление загрязнителей окружающей среды (Klein and Eveleigh, 1998; Lorito и другие. , 2010; Шореш и др., 2010; Mastouri et al., 2012).
, 2010; Шореш и др., 2010; Mastouri et al., 2012).
Каковы основные типы наземных экосистем?
Обновлено 22 ноября 2019 г.
Брюс Смит
Понятие экосистемы включает абиотическую (или неживую) и биотическую (или живую) части территории, а также взаимодействие между ними. Материя и энергия перемещаются между абиотическими и биотическими компонентами экосистемы. К абиотическим факторам, влияющим на экосистему, относятся температура, осадки, высота над уровнем моря и тип почвы.
Ученые делят экосистемы на наземные (наземные экосистемы) и неземные (неземные экосистемы). Экосистемы могут быть дополнительно классифицированы по их географическому региону и доминирующему типу растений. Водные, морские и водно-болотные угодья составляют внеземные экосистемы, а пятью основными наземными экосистемами являются пустыня, лес, луга, тайга и тундра.
Экосистемы пустыни
Количество осадков является основным абиотическим определяющим фактором экосистемы пустыни.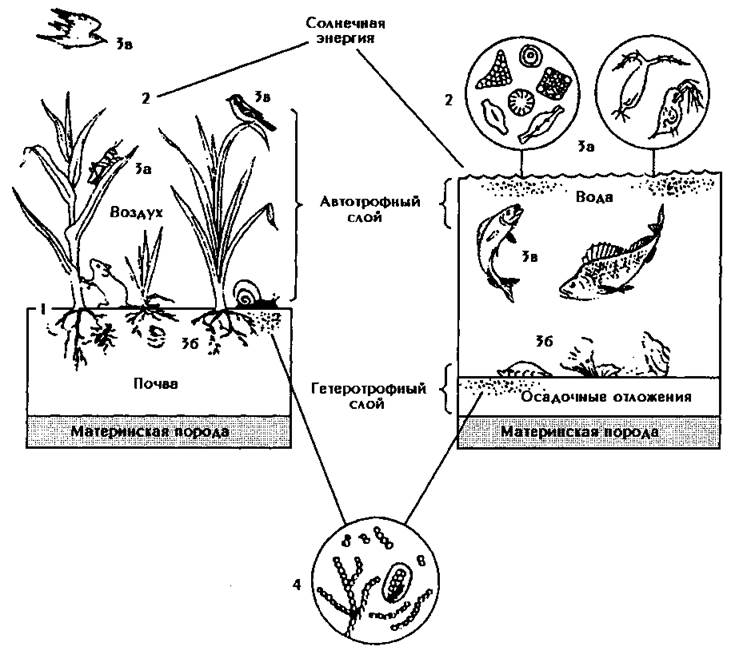 В пустынях выпадает менее 25 сантиметров (около 10 дюймов) дождя в год. Сильные колебания между дневной и ночной температурой характерны для наземной среды пустыни. Почвы содержат большое количество минералов и мало органических веществ.
В пустынях выпадает менее 25 сантиметров (около 10 дюймов) дождя в год. Сильные колебания между дневной и ночной температурой характерны для наземной среды пустыни. Почвы содержат большое количество минералов и мало органических веществ.
Растительность варьируется от несуществующей до большого количества высокоадаптированных растений. Экосистема пустыни Сонора содержит множество суккулентов и кактусов, а также деревья и кустарники. Они адаптировали структуру своих листьев, чтобы предотвратить потерю воды.Например, куст креозота имеет толстый слой, покрывающий его листья, чтобы предотвратить потерю воды из-за транспирации.
Одна из самых известных пустынных экосистем — пустыня Сахара, занимающая всю верхнюю часть африканского континента. По размеру сопоставима со всеми Соединенными Штатами и известна как самая большая жаркая пустыня в мире с температурой, превышающей 122 градуса по Фаренгейту.
Лесные экосистемы
Около одной трети суши Земли покрыто лесами. Основное растение в этой экосистеме — деревья. Лесные экосистемы подразделяются по типу деревьев, которые они содержат, и количеству получаемых осадков.
Основное растение в этой экосистеме — деревья. Лесные экосистемы подразделяются по типу деревьев, которые они содержат, и количеству получаемых осадков.
Некоторыми примерами лесов являются лиственные леса умеренного пояса, тропические леса умеренного пояса, влажные тропические леса, сухие тропические леса и северные хвойные леса. В сухих тропических лесах есть влажные и засушливые сезоны, а в тропических лесах дожди идут круглый год. Оба эти леса страдают от антропогенного давления, например, когда вырубают деревья, чтобы освободить место для ферм.Из-за обильных дождей и благоприятных температур тропические леса обладают высоким биоразнообразием.
Таежные экосистемы
Другой тип лесных экосистем — тайга, также известная как северные хвойные леса или бореальные леса. Он охватывает обширную территорию в северном полушарии. Ему не хватает биоразнообразия, есть только несколько видов. Экосистемы тайги характеризуются коротким вегетационным периодом, низкими температурами и бедной почвой.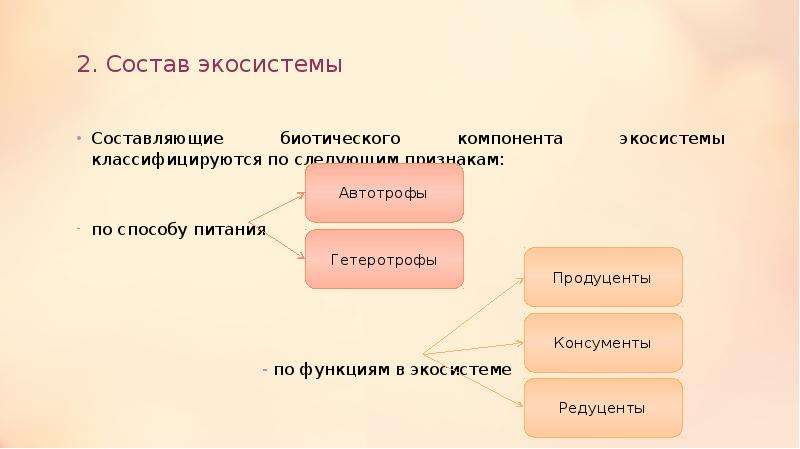
В этой земной среде длинные летние дни и очень короткие зимние дни.Среди животных, обитающих в тайге, — рыси, лоси, волки, медведи и роющие грызуны.
Экосистемы пастбищ
Пастбища умеренного пояса включают прерии и степи. У них есть сезонные изменения, но они не получают достаточного количества осадков, чтобы поддерживать большие леса.
Саваны — это тропические луга. Саванны имеют сезонную разницу в количестве осадков, но температуры остаются постоянными. Луга по всему миру были преобразованы в фермы, что уменьшило количество биоразнообразия в этих областях.Видными животными в экосистемах пастбищ являются травоядные, такие как газели и антилопы.
Тундра
Существует два типа тундры: арктическая и альпийская . Арктическая тундра расположена за Полярным кругом, к северу от бореальных лесов. Альпийские тундры встречаются на горных вершинах. Оба типа испытывают низкие температуры в течение всего года.
Из-за низких температур летом оттаивает только верхний слой почвы в этой земной среде; остальная часть остается замороженной круглый год — состояние, известное как вечная мерзлота.Растения в тундре — это в основном лишайники, кустарники и кустарники. В тундрах нет деревьев. Большинство животных, обитающих в тундре, на зиму мигрируют на юг или вниз с горы.
Ранняя жизнь на суше и первые наземные экосистемы | Экологические процессы
Абед РММ, Раметт А., Хюбнер В., Де Деккер П., де Бир Д.: Микробное разнообразие источников эоловой пыли из отложений соленых озер и биологических корок почвы в засушливой Южной Австралии. FEMS Microbiol Ecol 2011, 80 (2): 294–304.
Google Scholar
Адам Дж., Рашмер Т., О’Нил Дж., Фрэнсис Д.: Гадейские зеленые камни складчатого пояса Нуввуагиттук и происхождение ранней континентальной коры Земли. Геология 2012, 40: 363–366.
CAS Google Scholar
Аль-Бадер Д. , Элияс М., Райан Р., Радван С.: Переносимые воздухом ассоциации фототрофных и потребляющих углеводороды микроорганизмов: многообещающие консорциумы в биоремедиации летучих углеводородов. Environ Sci Pollut R 2012, 19 (9): 3997–4005.
, Элияс М., Райан Р., Радван С.: Переносимые воздухом ассоциации фототрофных и потребляющих углеводороды микроорганизмов: многообещающие консорциумы в биоремедиации летучих углеводородов. Environ Sci Pollut R 2012, 19 (9): 3997–4005.
CAS Google Scholar
Аллер Д.Ю., Аллер Р.К., Кемп П.Ф., Чистосердов А.Ю., Мадрид В.М.: Псевдоожиженные буровые растворы: новые условия для создания разнообразия биосферы в геологическом времени. Геобиология 2010, 8: 169–178.
CAS Google Scholar
Эллисон П.А., Боттьер Д.Д.: Тафономия: предвзятость и процесс во времени.В г. Тафономия: процесс и предвзятость во времени. Разделы геобиологии, т. 32 . Под редакцией: Allison PA, Bottjer DJ. Нью-Йорк: Спрингер; 2011: 1–17.
Google Scholar
Allwood AC, Walter MR, Kamber BS, Marshall CP, Burch IW: Строматолитовый риф из раннеархейской эры Австралии. Nature 2006, 441: 714–718.
Nature 2006, 441: 714–718.
CAS Google Scholar
Альтерманн В. Аккреция, улавливание и связывание отложений в архейских строматолитах — морфологическое выражение древности жизни.В Стратегии обнаружения жизни. Серия ISSI по космическим наукам, т. 25 . Под редакцией: Ботта О, Бада Дж., Гомес Э. Дж., Джаво Э., Селсис Ф., Саммонс Р. Нью-Йорк: Спрингер; 2008: 55–79.
Google Scholar
Альтерманн В., Шопф Дж. У.: микрофоссилии из неоархейской группы Кэмпбелл, грикваленд-западная секвенция Трансваальской супергруппы и их палеоэкологические и эволюционные последствия. Докембрийские исследования 1995, 75: 65–90.
CAS Google Scholar
Асплер Л. Б., Дональдсон Дж. А. Палеоклиматология бассейна Ноначо (ранний протерозой), Северо-Западные территории, Канада. Palaeogeogr Palaeoclimateol Palaeoecol 1986, 56: 17–34.
Google Scholar
Аврамик С: самые старые записи фотосинтеза. Photosynth Res 1992, 33: 75–89.
CAS Google Scholar
Awramik SM, Buchheim HP: гигантская система озер позднего архея: пачка Минтеена (формация Тумбиана; группа Фортескью), Западная Австралия. Докембрийские исследования 2009, 174: 215–240.
CAS Google Scholar
Аврамик С.М., Шопф Дж. В., Вальтер М.Р.: Нитчатые ископаемые бактерии из архея Западной Австралии. Докембрийские исследования 1983, 20: 357–374.
Google Scholar
Бамбах РК: Энергетика в мировой морской фауне: связь между земной диверсификацией и изменениями в морской биосфере. Geobios 1999, 32 (2): 131–144.
Google Scholar
Bamforth SS: Распространение микробов в пустынях и лесах Аризоны. Soil Biol Biochem 1984, 16 (2): 133–137.
Soil Biol Biochem 1984, 16 (2): 133–137.
Google Scholar
Bamforth SS: Водная пленочная фауна микробиотических корок теплой пустыни. J Arid Environ 2004, 56: 413–423.
Google Scholar
Bandopadhyay PC, Eriksson PG, Roberts RJ: Вертикальная палеопочва на стыке архея и протерозоя из кратона Сингхбхум-Орисса, восточная Индия. Докембрийские исследования 2010, 177 (3–4): 277–290.
CAS Google Scholar
Банерджи DM: палеопочва нижнего протерозоя на границе БГК-Аравалли в южно-центральном Раджастане. India J Geol Soc 1996, 48: 277–288.
CAS Google Scholar
Banerjee NR, Furnes H, Muehlenbachs K, Staudigel H, de Wit MJ: Сохранение ок. Микробные биомаркеры 3,4–3,5 млрд лет в подушечных лавах и гиалокластитах Зеленокаменного пояса Барбертона, Южная Африка. Earth Planet Sci Lett 2006, 241: 707–722.
Earth Planet Sci Lett 2006, 241: 707–722.
CAS Google Scholar
Баркер В.В., Велч С.А., Чу С., Банфилд Дж.Ф.: Экспериментальные наблюдения влияния бактерий на выветривание алюмосиликатов. Am Mineral 1998, 83: 1551–1563.
CAS Google Scholar
Баросс Дж. А., Хоффман С. Е.: Подводные гидротермальные источники и связанные с ними градиентные среды как места происхождения и эволюции жизни. Orig Life 1985, 15: 327–345.
CAS Google Scholar
Бартон HA, Northup DE: Геомикробиология в пещерных средах: прошлые, настоящие и будущие перспективы. J Cave Karst Stud 2007, 69: 163–178.
Google Scholar
Бейтс С.Т., Гарсия-Пичел Ф: независимое от культуры исследование свободноживущих грибов в биологических почвенных корках плато Колорадо: их разнообразие и относительный вклад в микробную биомассу.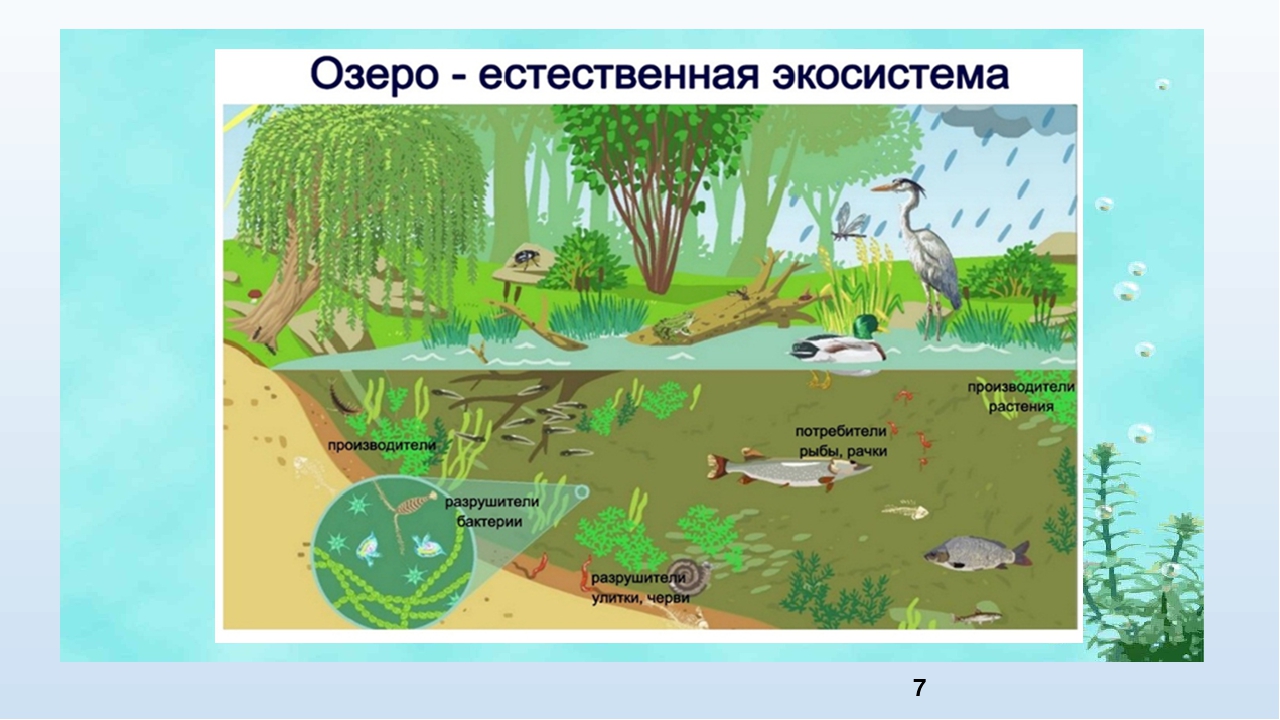 Environ Microbiol 2009, 11 (1): 56–67.
Environ Microbiol 2009, 11 (1): 56–67.
CAS Google Scholar
Бейтс С.Т., Клементе Дж. К., Флорес Г. Е., Уолтерс В. А., Вегенер-Парфри Л., Найт Р., Фирер Н.: Глобальная биогеография очень разнообразных протистанских сообществ в почве. ISME Jour 2013, 7: 652–659.
CAS Google Scholar
Battistuzzi FU, Hedges SB: крупная группа прокариот с древними приспособлениями к жизни на суше. Mol Biol Evol 2009, 26: 335–344.
CAS Google Scholar
Battistuzzi FU, Feijao A, Hedges SB: Геномная шкала времени эволюции прокариот: понимание происхождения метаногенеза, фототрофии и колонизации земли. BMC Evol Biol 2004, 4 (44): 1–14.
Google Scholar
Баулд Дж .: Геобиологическая роль цианобактериальных матов в осадочной среде: производство и сохранение органического вещества.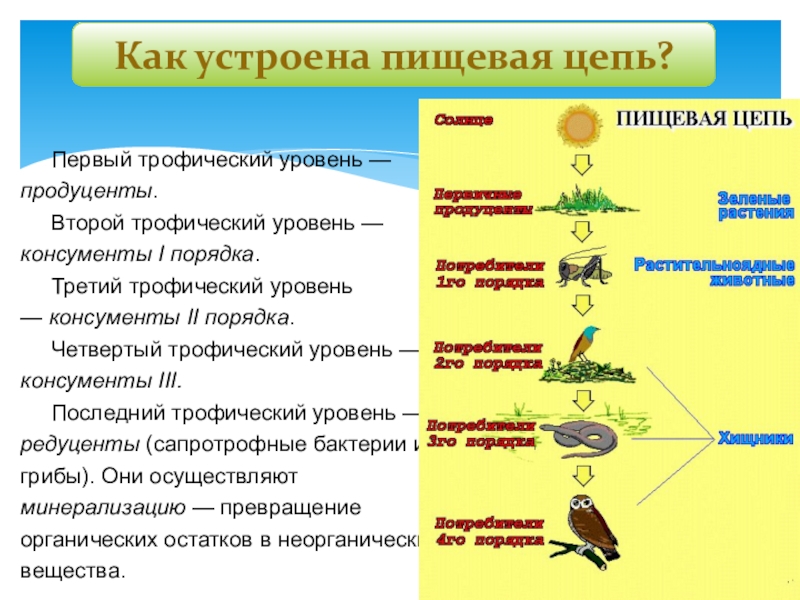 BMR J Aust Geol Geophys 1981, 6: 307–317.
BMR J Aust Geol Geophys 1981, 6: 307–317.
CAS Google Scholar
Баулд Дж., Д’Амелио Э., Фермер Дж. Д.: Современные микробные маты. В Протерозойская биосфера . Под редакцией: Schopf JW, Klein C. Нью-Йорк: Cambridge University Press; 1992: 261–269.
Google Scholar
Behrensmeyer AK, Damuth JD, DiMichele WA, Potts R: Наземные экосистемы во времени: эволюционная палеоэкология наземных растений и животных .Чикаго, Иллинойс: Издательство Чикагского университета; 1992.
Google Scholar
Белнап Дж., Джиллетт Д.А.: Уязвимость биологических почвенных корок пустынь к ветровой эрозии: влияние развития корки, текстуры почвы и нарушения. J Arid Environ 1998, 39: 133–142.
Google Scholar
Belnap J, Lange OL: Биологические корки почвы: структура, функции и управление. Серия экологических исследований, т. 150 . Берлин: Springer; 2001.
Серия экологических исследований, т. 150 . Берлин: Springer; 2001.
Google Scholar
Bennett PC, Rogers JR, Choi WJ: Силикаты, силикатное выветривание и микробная экология. Geomicrobiol J 2001, 18: 3–19.
CAS Google Scholar
Беральди-Кампеси Х., Хартнетт Х.Э., Анбар А., Гордон Г.В., Гарсия-Пичель Ф .: Влияние биологических почвенных корок на концентрации элементов в почве: последствия для биогеохимии и как прослеживаемые биосигнатуры древней жизни на суше. Геобиология 2009, 7: 348–359.
CAS Google Scholar
Беральди-Кампеси Х., Фермер Дж. Д., Гарика-Пичел Ф .: Доказательства мезопротерозойской жизни на суше и ее современного аналога в засушливых почвах . Материалы ежегодного собрания GSA, Миннеаполис, 9–12 октября 2011 г .; 2011.
Google Scholar
Бернар С. , Бензерара К., Бейссак О, Менгуи Н., Гайот Ф., Браун Г.Е. Младший, Гоффе Б.: Исключительная сохранность ископаемых спор растений в метаморфических породах высокого давления. Earth Planet Sc Lett 2007, 262: 257–272.
, Бензерара К., Бейссак О, Менгуи Н., Гайот Ф., Браун Г.Е. Младший, Гоффе Б.: Исключительная сохранность ископаемых спор растений в метаморфических породах высокого давления. Earth Planet Sc Lett 2007, 262: 257–272.
CAS Google Scholar
Бекес, штат Нью-Джерси, Дорланд Х, Гутцмер Дж, Недачи М., Омото Х: тропические латериты, жизнь на суше и история атмосферного кислорода в палеопротерозое. Геология 2002, 30: 491–494.
CAS Google Scholar
Блэквелл М: Наземная жизнь: изначально была грибком? Science 2000, 289 (5486): 1884–1885.
CAS Google Scholar
Бланк CE: Не такие старые археи — древность биогеохимических процессов в архейной сфере жизни. Геобиология 2009, 7 (5): 495–514.
CAS Google Scholar
Стенд WE: Водоросли как пионеры в сукцессии растений и их значение в борьбе с эрозией. Экология 1941, 22: 38–46.
Экология 1941, 22: 38–46.
Google Scholar
Бувье З .: Новое определение возраста Солнечной системы. Nat Geosci 2010, 3: 637–641.
CAS Google Scholar
Boyet M, Carlson RW: 142 Nd свидетельствует о ранней (> 4,53 млрд лет) глобальной дифференциации силикатной Земли. Наука 2005, 309: 576–581.
CAS Google Scholar
Brady NC, Weil RR: Природа и свойства почв . 14-е издание. Верхняя Сэдл-Ривер, Нью-Джерси: Пирсон-Прентис-холл; 2008 г.
Google Scholar
Брокс Дж. Дж.: Градиенты концентрации углеводородов в архейских сланцах в миллиметровом масштабе: утечка живой нефти или отпечаток загрязнения? Geochim Cosmochim Acta 2011, 75 (11): 3196–3213.
CAS Google Scholar
Büdel B, Weber B, KuhlM PH, Sultemeyer D, Wessels D: Изменение формы поверхности песчаника криптоэндолитическими цианобактериями: биощелачивание вызывает химическое выветривание в засушливых ландшафтах. Геобиология 2004, 2: 261–268.
Геобиология 2004, 2: 261–268.
Google Scholar
Buick R: Когда развился кислородный фотосинтез? Philos Trans R Soc Lond B Biol Sci 2008, 363 (1504): 2731–2743.
CAS Google Scholar
Buick R: Ранние годы: древние акритархи. Nature 2010, 463 (7283): 885–886.
CAS Google Scholar
Buick R, Thornett JR, McNaughton NJ, Smith JB, Barley ME, Savage M: запись формирующейся континентальной коры 3.5 миллиардов лет назад в кратоне Пилбара в Австралии. Nature 1995, 375: 574–577.
CAS Google Scholar
Campbell SE: Стабилизация почвы прокариотической корой пустыни: последствия для докембрийской наземной биоты. Orig Life 1979, 9: 335–348.
CAS Google Scholar
Кэмпбелл С.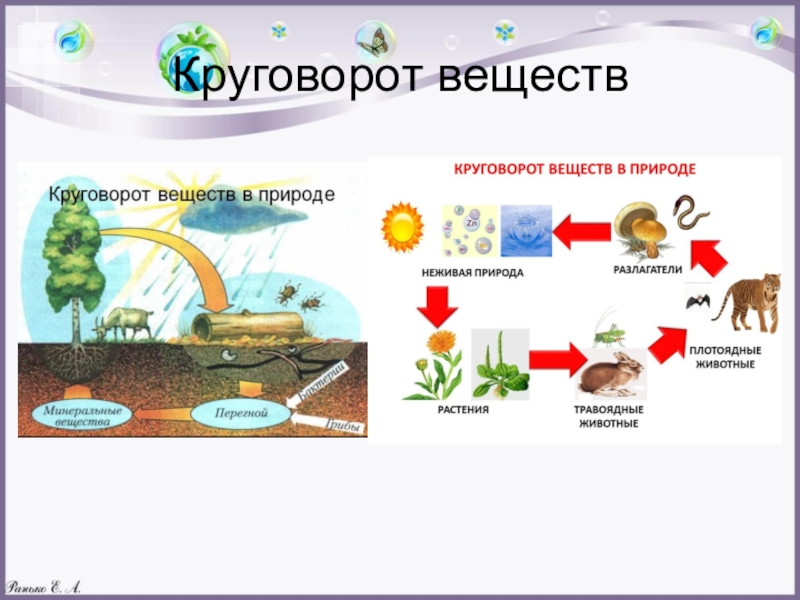 Е., Зеелер Дж., Голубич С. Формирование пустынной корки и стабилизация почвы.В: Дж. Скуджинс (ред.) Использование микробиологических процессов на засушливых землях для борьбы с опустыниванием и повышения производительности (ЮНЕП). Восстановление засушливых почв 1989, 3: 217–228.
Е., Зеелер Дж., Голубич С. Формирование пустынной корки и стабилизация почвы.В: Дж. Скуджинс (ред.) Использование микробиологических процессов на засушливых землях для борьбы с опустыниванием и повышения производительности (ЮНЕП). Восстановление засушливых почв 1989, 3: 217–228.
Google Scholar
Canup RM, Righter K (Eds): Происхождение Земли и Луны . Тусон, Аризона: Университет Аризоны Press; 2000.
Google Scholar
Catling DC, Zahnle KJ, McKay CP: Биогенный метан, утечка водорода и необратимое окисление ранней Земли. Наука 2001, 293: 839–843.
CAS Google Scholar
Cavosie AJ, Valley JW, Wilde S: Самая старая земная летопись минералов: обзор обломочных цирконов от 4400 до 3900 млн лет назад из Джек-Хиллз, Западная Австралия. В числе самых старых горных пород Земли. разработки в докембрийской геологии, серия 15 . Под редакцией: Ван Кранендонк MJ, Smithies RH, Беннет В. Амстердам: Elsevier; 2007: 91–111.
Под редакцией: Ван Кранендонк MJ, Smithies RH, Беннет В. Амстердам: Elsevier; 2007: 91–111.
Google Scholar
Чаналь А., Чапон В., Бензерара К., Баракат М., Кристен Р., Ачуак В., Баррас Ф., Хеулин Т.: Пустыня Татауин: экстремальная среда, в которой обитает большое разнообразие микроорганизмов и радиотолерантных бактерий. Environ Microbiol 2006, 8 (3): 514–525.
CAS Google Scholar
Чен И, Ву Л., Боден Р., Хиллебранд А, Кумаресан Д., Муссар Х., Басиу М., Лу И, Мюррелл Дж. К.: Жизнь без света: разнообразие микробов и свидетельства хемолитотрофии на основе серы и аммония в пещере Movile . ISME J 2009, 3: 1093–1104.
CAS Google Scholar
Chenu C, Stotzky G: Взаимодействие между микроорганизмами и частицами почвы: обзор.В Взаимодействие между частицами почвы и микроорганизмами: воздействие на экосистему суши . Под редакцией: Хуанг П.М., Боллаг Дж. М., Сенези Н. Чичестер: Джон Уайли и сыновья; 2002: 4–40.
Под редакцией: Хуанг П.М., Боллаг Дж. М., Сенези Н. Чичестер: Джон Уайли и сыновья; 2002: 4–40.
Google Scholar
Чоровер Дж., Кречмар Р., Гарсия-Пичель Ф., Спаркс Д.Л.: Биогеохимические процессы в почве в критической зоне. Элементы 2007, 3: 321–326.
CAS Google Scholar
Cloud P, микробы A: Новые допалеозойские наннофоссилии из формации Стоер (торридон), Северо-Западная Шотландия. Geol Soc Am Bull 1971, 82: 3469–3474.
Google Scholar
Condie KC: Докембрийские суперплюмы. В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 163–173.
Google Scholar
Corcoran PL, Mueller WU: Агрессивное архейское выветривание.В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 494–504.
Google Scholar
Dauphas N, van Zuilen M, Wadhwa M, Davis AM, Martey B, Janney PE: Ключ к разгадке изотопных вариаций Fe о происхождении ранних архейских BIF из Гренландии. Наука 2004, 302: 2077–2080.
Google Scholar
Дэвид Л.А., Альм Э.Дж.: Быстрые эволюционные инновации во время генетической экспансии архей. Nature 2011, 469 (7328): 93–96.
CAS Google Scholar
Дэвисон Дж .: Генетический обмен между бактериями в окружающей среде. Плазмида 1999, 42 (2): 73–91.
CAS Google Scholar
Деминг Д: Происхождение океана и континентов: единая теория Земли. Int Geol Rev 2002, 44: 137–152.
Google Scholar
DesMarais DJ: Когда на Земле зародился фотосинтез? Наука 2000, 289: 1703–1705.
Google Scholar
DiMichele WA, Hook RW: палеозойские наземные экосистемы. В земных экосистемах во времени . Под редакцией: Беренсмейер А.К., Дамут Дж.Д., ДиМишель В.А., Поттс Р., Зьюсс HD, Wing SL.Чикаго: Издательство Чикагского университета; 1992: 205–325.
Google Scholar
Dong GR, Li CZ, Jin T, Gao SY, Wu D: Некоторые результаты эксперимента с имитацией грунтовой аэродинамической трубы. Chinese Sci Bull 1987, 32: 297–301.
Google Scholar
Дотт Р. Х. Младший: Важность эоловой абразии в сверхзрелых кварцевых песчаниках и парадокс выветривания на свободных от растительности ландшафтах. J Geol 2003, 111: 387–405.
Google Scholar
Дризе С.Г., Гордон-Медарис Л. Младший: данные о биологическом и гидрологическом контроле над развитием палеопротерозойского палеопотеронного профиля в хребте Барабу, Висконсин, США. J Sediment Res 2008, 78: 443–457.
CAS Google Scholar
Дризе С.Г., Симпсон Э.Л., Эрикссон К.А.: Редоксиморфные палеопочвы в аллювиальных и озерных отложениях, 1.8 Формирование галактики, Маунт-Айза, Австралия: почвообразовательные процессы и последствия для палеоклимата. J Sediment Res 1995, A65: 675–689.
CAS Google Scholar
Эрлих HL: Геомикробиология . Нью-Йорк, Нью-Йорк: Марсель Деккер; 2002.
Google Scholar
Эль-Альбани А., Бенгтсон С., Кэнфилд Д.Е., Беккер А., Маккиарелли Р., Мазурье А., Хаммарлунд ЕС, Бульвэ П., Дюпюи Дж. Дж., Фонтен С., Фюршич Ф. Т., Готье-Лафай Ф. Крупные колониальные организмы со скоординированным ростом в насыщенная кислородом среда 2.1 млрд лет назад. Nature 2010, 466 (7302): 100–104.
CAS Google Scholar
Эльберт В., Вебер Б., Берроуз С., Стейнкамп Дж., Бюдель Б., Андреа М. О., Пёшль Ю.: Вклад криптогамных покрытий в глобальные циклы углерода и азота. Nat Geosci 2012, 5: 459–462.
CAS Google Scholar
Энгель А.С.: Микробное разнообразие пещерных экосистем.В Геомикробиология: молекулярные и экологические перспективы . Под редакцией: Бартон Л., Мандл М., Лой А. Нью-Йорк: Спрингер; 2010: 219–238.
Google Scholar
Эрикссон П.Г., Мартинс-Нето Массачусетс: Комментарий. В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Амстердам: Эльзевир; 2004: 677–680.
Google Scholar
Эрикссон П.Г., Симпсон Э.Л., Эрикссон К.А., Бамби А.Дж., Стейн Г.Л., Саркар С.: Мутные рулонные структуры в кремнисто-обломочных слоях ок.1,8 млрд лет Waterberg Group, Южная Африка. PALAIOS 2000, 15: 177–183.
Google Scholar
Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катунеану О: Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Амстердам: Эльзевир; 2004.
Google Scholar
Фермер Дж .: Тафономические режимы в окаменелости микробов.В Совет по космическим исследованиям (ред.) Пределы размеров очень мелких микроорганизмов . Вашингтон, округ Колумбия: Национальный исследовательский совет, Национальная академия прессы; 1999: 94–102.
Google Scholar
Фарроу К.Э., Моссман Д.Д.: Геология докембрийских палеопочв в основании гуронской супергруппы, озеро Эллиот, Онтарио, Канада. Докембрия Res 1988, 42: 107–139.
Google Scholar
Fedo CM, Whitehouse MJ: Метасоматическое происхождение кварц-пироксеновой породы, Акилия, Гренландия, и последствия для самой ранней жизни на Земле. Наука 2002, 296: 1448–1452.
CAS Google Scholar
Фирер Н., Немергут Д., Найт Р., Крейн Дж. М.: Изменения во времени: интеграция микроорганизмов в изучение сукцессии. Res Microbiol 2010, 161: 635–642.
Google Scholar
Флеминг Э.Д., Кастенхольц Р.В.: Влияние периодической сушки на синтез УФ-скринирующего соединения, сцитонемина, в цианобактериях. Environ Microbiol 2007, 9: 1448–1455.
CAS Google Scholar
Флетчер Дж. Э., Мартин В. П.: Некоторые эффекты водорослей и плесени в дождевой корке пустынных почв. Экология 1948, 29 (1): 95–100.
Google Scholar
Fliegel D, Wirth R, Simonetti A, Furnes H, Staudigel H, Hanski E, Muehlenbachs: Септатно-трубчатые текстуры в подушечных лавах 2,0 Ga из Печенгского зеленокаменного пояса: наноспектроскопический подход к исследованию их биогенности. Геобиология 2010, 8: 372–390.
CAS Google Scholar
Franz G, Mosbrugger V, Menge R: Карбо-пермские фрагменты листьев птеридофилла из фундамента амфиболитовой фации, Tauern Window, Австрия. Terra Nova 1991, 3: 137–141.
Google Scholar
Фридманн Э.И.: Эндолитическая микробная жизнь в жарких и холодных пустынях. Orig Life 1980, 10: 223–235.
CAS Google Scholar
Fritsch FE: Наземные водоросли. J Ecol 1922, 10 (2): 220–236.
Google Scholar
Furnes H, Banerjee NR, Muehlenbachs K, Staudigel H, de Wit M: Ранние годы жизни зарегистрированы в архейских подушечных лавах. Наука 2004, 304: 578–81.
CAS Google Scholar
Фурнес Х, де Вит М., Штаудигель Х, Розинг М., Мюленбах К.: остатки старейшего офиолита на Земле. Наука 2007, 315: 1704–1707.
CAS Google Scholar
Фурнес Х., Банерджи Н.Р., Стаудигель Х., Мюленбах К., Маклафлин Н., де Вит М., Ван Кранендонк М.: Сравнение петрографических сигнатур недавних биоизменений с мезоархейскими подушечными лавами: отслеживание подземной жизни в океанических магматических породах. Докембрия Res 2007, 158: 156–176.
CAS Google Scholar
Gadd GM (Ed): Грибы в биогеохимических циклах .Кембридж, Великобритания: Издательство Кембриджского университета; 2006.
Google Scholar
Gall Q: Протерозойские палеопочвы Телона, Северо-Западные территории, Канада. Докембрийское исследование 1994, 68: 115–137.
CAS Google Scholar
Gao Q, Garcia-Pichel F: солнцезащитные средства с микробным ультрафиолетом. Nat Rev Microbiol 2011, 9: 791–802.
CAS Google Scholar
Гарсия Руис Дж. М., Карнеруп А., Кристи А. Г., Велхэм Нью-Джерси, Хайд СТ: Морфология: неоднозначный индикатор биогенности. Астробиология 2002, 2 (2): 353–369.
Google Scholar
Гарсия-Пичель Ф .: Солнечный ультрафиолет и эволюционная история цианобактерий. Origins Life Evol. Биосфера 1998, 28: 321–347.
CAS Google Scholar
Гарсия-Пичель Ф., Кастенхольц Р.В.: Характеристика и биологические последствия ситонемина, пигмента оболочки цианобактерий. J Phycol 1991, 27: 395–409.
CAS Google Scholar
Гарсия-Пичель Ф., Принга О: Цианобактерии отслеживают воду в почвах пустынь. Nature 2001, 413: 380–381.
CAS Google Scholar
Гарсия-Пичель Ф., Войцеховски М.Ф .: Развитие способности строить надклеточные веревки позволило нитчатым цианобактериям колонизировать сильно разрушаемые субстраты. PLoS One 2009, 4 (11): e7801.
Google Scholar
Гарсия-Пичел Ф., Лопес-Кортес А., Нубель У.: Филогенетическое и морфологическое разнообразие цианобактерий в корках почвенной пустыни с плато Колорадо. Appl Environ Microbiol 2001, 67: 1902–1910.
CAS Google Scholar
Гарсия-Пичель Ф., Белнап Дж., Нойер С., Шанц Ф .: Оценка глобальной биомассы цианобактерий и ее распределения. Algol Stud 2003, 109: 213–227.
Google Scholar
Гарсия-Руис Дж. М., Хайд СТ, Карнеруп А. М., Кристи А. Г., Ван Кранендонк М. Дж., Велхэм Нью-Джерси: Самособирающиеся кремнеземно-карбонатные структуры и обнаружение древних микрофоссилий. Наука 2003, 302: 1194–1197.
CAS Google Scholar
Гоше К., Сиал А.Н., Халверсон Г.П., Фриммель HE: Неопротерозойско-кембрийская тектоника, глобальные изменения и эволюция: в центре внимания Юго-Западная Гондвана.События в геологии докембрия, т. 16 . Амстердам: Эльзевир; 2010.
Google Scholar
Гей А.Л., Грандстафф Д.Е.: Химия и минералогия докембрийских палеопочв на озере Эллиот, Онтарио, Канада. Докембрийское исследование 1980, 12: 349–373.
CAS Google Scholar
Geesey G, Jang L: внеклеточные полимеры для связывания металлов. В Микробное извлечение полезных ископаемых .Под редакцией: Эрлих Х.Л., Бриерли С.Л. Нью-Йорк: Макгроу-Хилл; 1990: 223–249.
Google Scholar
Gensel PG: Самые ранние наземные растения. Annu Rev Ecol Evol Syst 2008, 39: 459–477.
Google Scholar
Ghiorse W, Wilson J: Микробная экология земных недр. В Достижения прикладной микробиологии . Под редакцией: Ласкин А. Нью-Йорк: Академический; 1988: 107–172.
Google Scholar
Джорданино Ф., Штраух С.М., Вильяфанье В.Е., Хелблинг Е.В.: Влияние температуры и УФИ на фотосинтез и морфологию четырех видов цианобактерий. J Photochem Photobiol B: Biol 2011, 103: 68–77.
CAS Google Scholar
Гласс Дж. Б., Вульф-Саймон Ф., Анбар А. Д.: Коэволюция доступности металлов и ассимиляции азота у цианобактерий и водорослей. Геобиология 2009, 7: 100–123.
CAS Google Scholar
Гликсон A: Ранние архейские столкновения астероидов с Землей: стратиграфические и изотопные корреляции возраста и возможные геодинамические последствия. В числе самых старых горных пород Земли. События в геологии докембрия, серия 15 . Под редакцией: Ван Кранендонк MJ, Smithies RH, Беннет В. Амстердам: Elsevier; 2007: 1087–1103.
Google Scholar
Гловер Дж. Ф., Ках LC: Отложения спелеотем в протерозойском палеокарсте, группа мезопротерозойских мрачных озер, Арктическая Канада .2006. Рефераты Геологического общества Америки с программами 38 (3): 36 Рефераты Геологического общества Америки с программами 38 (3): 36
Google Scholar
Gogarten JP, Fournier G, Zhaxybayeva O: Перенос генов и реконструкция ранней истории жизни на основе геномных данных. В Стратегии обнаружения жизни. Серия космических наук ISSI, т. 25 . Отредактировано: Ботта О, Бада Дж. Л., Гомес-Эльвира Дж., Яво Э., Селсис Ф., Саммонс Р.Нью-Йорк: Спрингер; 2007: 115–131.
Google Scholar
Голубич С., Кэмпбелл С.Е.: Аналоги микробных форм в современных субаэральных местообитаниях и в кремнях докембрия: Gloethece coerulea Geitler и Eosynechococcus moorei Hoffmann. Докембрия Res 1979, 8: 201–217.
Google Scholar
Голубич С., Хофманн Х. Дж .: Сравнение современных и средних докембрийских Entophysalidaceae (Cyanophyta) в матах строматолитовых водорослей: деление и деградация клеток. J Paleont 1976, 50: 1074–1082.
Google Scholar
Голубич С., Сон-Джу Л.: Летопись окаменелостей ранних цианобактерий: сохранение, палеоокружение и идентификация. Eur J Phycol 1999, 34: 339–348.
Google Scholar
Гомес Р., Левисон Х. Ф., Циганис К., Морбиделли А. Происхождение катастрофического периода поздней тяжелой бомбардировки планет земной группы. Nature 2005, 435 (7041): 466–469.
CAS Google Scholar
Градзински М., Хмиэль М.Дж., Левандовска А., Михальска-Касперкевич Б. Силикокластические микростроматолиты в пещере из песчаника: роль улавливания и связывания частиц детрита в формировании пещерных отложений. Ann Soc Geol Polon 2010, 80 (3): 303–314.
Google Scholar
Грандстафф Д.Е., Эдельман М.Дж., Фостер Р.В., Збинден Э., Кимберли М.М.: Химия и минералогия палеопочв докембрия в основании групп Доминион и Понгола. Докембрия Res 1986, 32: 97–131.
CAS Google Scholar
Gray J, Boucot AJ: Остатки неморских животных раннесилурийского периода и природа раннеконтинентальной экосистемы. Acta Palaeontol Polon 1994, 38 (3–4): 303–328.
Google Scholar
Gray J, Shear WA: Ранняя жизнь на суше. Am Sci 1992, 80: 444–456.
Google Scholar
Gunatilaka A: Некоторые аспекты биологии и седиментологии слоистых водорослей из лагуны Маннар, северо-запад Цейлона. Sediment Geol 1975, 14: 275–300.
Google Scholar
Gutzmer J, Beukes NJ: самые ранние латериты и возможные свидетельства существования наземной растительности в раннем протерозое. Геология 1998, 26: 263–266.
CAS Google Scholar
Hallbauer DK: Углерод Витватерсранда — растительное происхождение. Minerals Sci Eng 1975, 7 (2): 111–131.
CAS Google Scholar
Hallbauer DK, van Warmelo KT: окаменелые растения в тухолите из докембрийских пород Витватерсранда, Южная Африка. Докембрийское исследование 1974, 1: 199–212.
Google Scholar
Хамблин В.К., Кристиансен Э.Х .: Динамические системы Земли . 10-е издание. Река Аппер Сэдл, штат Нью-Джерси: Prentice Hall; 2007.
Google Scholar
Хан TM, Руннегар B: Мегаскопические эукариотические водоросли из 2.Железная формация negaunee возрастом 1 миллиард лет, Мичиган. Science 1992, 257 (5067): 232–235.
CAS Google Scholar
Ханель М., Монтенари М., Кальт А: Определение возраста осадконакопления метаморфических гнейсов высокого содержания по их палинологическим данным: тематическое исследование в северной части Шварцвальда (Варисканский пояс, Германия). Int J Earth Sci 1999, 88: 49–59.
CAS Google Scholar
Харрисон С.П., Кофельд К.Э., Руландт К., Клакин Т.: Роль пыли в изменении климата сегодня, во время последнего ледникового максимума и в будущем. Earth Sci Rev. 2001, 54: 43–80.
CAS Google Scholar
Хартманн В.К., Райдер Дж., Донес Л., Гринспун Д.: Зависящая от времени интенсивная бомбардировка исконной системы Земля / Луна. В году Происхождение Земли и Луны . Под редакцией: Кануп Р.М., Райтер К. Тусон: Университет Аризоны Press; 2000: 493–512.
Google Scholar
Хазен Р.М., Сверженский Д.А.: Минеральные поверхности, геохимические сложности и происхождение жизни.В г. Истоки жизни, том 5 . 2-е издание. Отредактировано: Deamer D, Szostak JW. Колд-Спринг-Харбор: Лаборатория Колд-Спринг-Харбор; 2010: 1-21.
Google Scholar
Hazen RM, Golden J, Downs RT, Hystad G, Grew ES, Azzolini D, Sverjensky DA: Эволюция минералов ртути (Hg): минералогическая запись сборки суперконтинента, изменения геохимии океана и зарождающейся земной биосферы. Am Mineral 2012, 97: 1013–1042.
CAS Google Scholar
Хекман Д.С., Гейзер Д.М., Эйделл Б.Р., Штауфер Р.Л., Кардос Н.Л., Хеджес С.Б.: Молекулярные доказательства ранней колонизации земли грибами и растениями. Наука 2001, 293 (5532): 1129–1133.
CAS Google Scholar
Herrero A, Flores E (Eds): Цианобактерии: молекулярная биология, геномика и эволюция .Норфолк: Caister Academic Press; 2008.
Google Scholar
Хикман А.Х., Ван Кранендонк М.Дж .: Ранняя эволюция Земли: свидетельства геологической истории региона Пилбара в Западной Австралии за 3,5–1,8 млрд лет. Эпизоды 2012, 35 (1): 283–297.
Google Scholar
Hoashi M, Bevacqua DC, Otake T, Watanabe Y, Hickman AH, Utsunomiya S, Ohmoto H: образование первичного гематита в насыщенном кислородом море 3.46 миллиардов лет назад. Nat Geosci 2009, 2: 301–306.
CAS Google Scholar
Гофман П.Ф .: Древнейший наземный ландшафт. Nature 1995, 375 (6532): 537–538.
CAS Google Scholar
Хоффман П.Ф., Шраг Д.П. Гипотеза Земли как снежного кома: проверка пределов глобальных изменений. Terra Nova 2002, 14: 129–155.
CAS Google Scholar
Hofmann HJ: Докембрийская микрофлора, острова Белчер, Канада; значение и систематика. J Paleontol 1976, 50: 1040–1073.
Google Scholar
Голландия HD: Химическая эволюция атмосферы и океанов . Принстон: Издательство Принстонского университета; 1984.
Google Scholar
Голландия HD: Распространение и интерпретация палеосреды протерозойских палеопочв. В Протерозойская биосфера . Под редакцией: Schopf JW, Klein C.Кембридж: Издательство Кембриджского университета; 1992: 153–155.
Google Scholar
Холм Н.Г .: Морские гидротермальные системы и истоки жизни. Orig Life Evol Biosph 1992, 22: 181–242.
CAS Google Scholar
Хопперт М., Реймер Р., Кеммлинг А., Шредер А., Гюнцль Б., Хейнкен Т.: Структура и реакционная способность биологической корки почвы из ксерической песчаной почвы в Центральной Европе. Geomicrobiol J 2004, 21: 183–191.
Google Scholar
Городиски Р.Дж., Кнаут П.Л .: Жизнь на суше в докембрии. Science 1994, 263 (5146): 494–498.
CAS Google Scholar
Ху Ц., Лю И, Сун Л., Чжан Д.: Влияние водорослей почвы пустыни на стабилизацию мелкого песка. J Appl Phycol 2002, 14 (4): 281–292.
CAS Google Scholar
Hupe P: Sur des problematica du Precambrien III. Division des Mines et de la Géologie, Геологическая служба Марокко, Notes et Memoires 1952, 103: 297–383.
Google Scholar
Иидзука Т., Хорие К., Комия Т., Маруяма С., Хирата Т., Хидака Х, Виндли Б.Ф .: Ксенокристаллы циркона 4,2 млрд лет в гнейсе Акаста на северо-западе Канады: свидетельства ранней континентальной коры. Геология 2006, 34: 245–248.
CAS Google Scholar
Ishizuka H: протолиты комплекса Нэпиер на Земле Эндерби, Восточная Антарктида; обзор и значение для формирования земной коры архейских континентов. J Miner Petrol Sci 2008, 103: 218–225.
CAS Google Scholar
Джексон Т.А.: Ископаемые актиномицеты в ледниковых образованиях среднего докембрия. Science, New Series 1967, 155 (3765): 1003–1005.
CAS Google Scholar
Javaux EJ, Marshall CP, Bekker A: Органические окаменелости в мелководных морских силикокластических отложениях возрастом 3,2 миллиарда лет. Nature 2010, 463: 934–938.
CAS Google Scholar
Jickells TD, An ZS, Andersen KK, Baker AR, Bergametti G, Brooks N, Cao JJ, Boyd PW, Duce RA, Hunter KA, Kawahata H, Kubilay N, LaRoche J, Liss PS, Mahowald N, Prospero JM, Ridgwell AJ, Tegen I., Torres R: Глобальные связи железа между пылью пустыни, биогеохимией океана и климатом. Наука 2005, 308: 67–71.
CAS Google Scholar
Джонсон I, Ватанабе Y, Стюарт Б., Омото H: Самый старый на Земле (~ 3.4 млрд лет) латеритные палеопочвы в кратоне Пилбара, Западная Австралия, . Давос, Швейцария: Материалы конференции Гольдшмидта; 2009.
Google Scholar
Johnson I, Watanabe Y, Stewart B, Ohmoto H: Свидетельства земной жизни и богатой O2 атмосферы в старейших (~ 3,4 млрд лет) палеопочвах в восточной части кратона Пилбара, Западная Австралия . Лиг-Сити, Техас: Труды 6-й научной конференции по астробиологии; 2010 г.20–26 апреля 2010 г. 20–26 апреля 2010 г.
Google Scholar
Кандлер О. Раннее разнообразие жизни. В г. Ранняя жизнь на земле г. Под редакцией: Бенгстон С. Нью-Йорк: издательство Колумбийского университета; 1994: 152–161.
Google Scholar
Капплер А., Паскеро С., Конхаузер К.О., Ньюман Д.К.: Осаждение полосчатых железных образований аноксигенными фототрофными Fe (II) -окисляющими бактериями. Геология 2005, 33 (11): 865–868.
CAS Google Scholar
Кастинг J: Как найти обитаемую планету . Принстон: Издательство Принстонского университета; 2009.
Google Scholar
Като И., Сузуки К., Накамура К., Хикман А. Х., Недачи М., Кусакабе М., Беваква, округ Колумбия, Омото Х: образование гематита насыщенными кислородом грунтовыми водами более 2,76 миллиарда лет назад. Earth Planet Sci Lett 2009, 278: 40–49.
CAS Google Scholar
Келлер К.К., Вуд Б.Д .: Возможность химического выветривания до появления сосудистых наземных растений. Nature 1993, 364: 223–225.
CAS Google Scholar
Кендалл Б., Рейнхард, CT, Лион, TW, Кауфман, А.Дж., Поултон, юго-запад, Анбар, А.Д .: Повсеместная оксигенация вдоль окраин океана в поздний архей. Nat Geosci 2010, 3 (9): 647–652.
CAS Google Scholar
Кеннеди М.Дж., Вагнер Т .: Континентальный усилитель из глинистых минералов для связывания углерода в морской среде в тепличном океане. Proc Nat Acad Sci USA 2011, 108: 9776–9781.
CAS Google Scholar
Кеннеди М.Дж., Чедвик О.А., Витаусек П.М., Дерри Л.А., Хендрикс Д.М.: Замена выветривания атмосферными источниками основных катионов во время развития экосистемы, Гавайские острова. Геология 1998, 26: 1015–1018.
CAS Google Scholar
Кеннеди М., Дрозер М., Майер Л.М., Пивеар Д., Мрофка Д.: оксигенация в позднем докембрии; создание завода по производству глиняных минералов. Наука 2006, 311: 1446–1449.
CAS Google Scholar
Кимберли М.М., Грандстафф Д.Е.: Профили концентраций элементов в палеопочвах докембрия на базальтовых и гранитных материнских материалах. Докембрийское исследование 1986, 32: 133–154.
CAS Google Scholar
Киршвинк Дж. Л., Копп RE: Палеопротерозойские ледяные дома и эволюция ферментов, опосредующих кислород: случай позднего происхождения фотосистемы II. Фил Trans R Soc B 2008, 363: 2755–2765.
CAS Google Scholar
Кляйн С., Бекес, штат Нью-Джерси, Шопф Дж. У .: Нитчатые микрофоссилии в раннепротерозойской супергруппе Трансвааля: их морфология, значение и палеоэкологические условия. Докембрийская газета 1987, 36: 81–94.
Google Scholar
Knauth LP, Кеннеди MJ: Позднее докембрийское озеленение Земли. Nature 2009, 460: 728–732.
CAS Google Scholar
Knoll AH, Javaux EJ, Hewitt D, Cohen P: Эукариотические организмы в протерозойских океанах. Философские труды Королевского общества B 2006, 361 (1470): 1023–1038.
CAS Google Scholar
Knoll A, Canfield D, Konhauser K (Eds): Основы геобиологии . Чичестер: Вили-Блэквелл; 2012.
Google Scholar
Келер И., Конхаузер К.О., Капплер А: Роль микроорганизмов в полосчатых железных образованиях. В Геомикробиология: молекулярная и экологическая перспектива . Под редакцией: Бартон Л., Мандл М., Лой А.Нью-Йорк: Спрингер; 2010: 309–324.
Google Scholar
Комия Т., Маруяма С., Нода С., Масуда Т., Хаяси М., Окамото С.: тектоника плит 3,8–3,7 млрд лет; полевые данные из аккреционного комплекса Исуа, юг Западной Гренландии. J Geol 1999, 107: 515–554.
CAS Google Scholar
Konhauser K: Введение в геомикробиологию .Оксфорд: Блэквелл; 2007.
Google Scholar
Konhauser KO, Schultzelam S, Ferris FG, Fyfe WS, Longstaffe FJ, Beveridge TJ: осаждение минералов эпилитическими биопленками в Спид-Ривер, Онтарио, Канада. Appl Environ Microbiol 1994, 60: 549–553.
CAS Google Scholar
Конхаузер К.О., Ньюман Д.К., Капплер А: Потенциальное значение микробного восстановления Fe (III) во время отложения пластов докембрийского полосчатого железа. Геобиология 2005, 3: 167–177.
CAS Google Scholar
Косицин Н., Крапез Б.: SHRIMP Геохронология обломочного циркона U-Pb в позднеархейском бассейне Витватерсранд, связь между возрастными спектрами источника циркона и эволюцией бассейна. Докембрийские исследования 2004, 129: 141–168.
CAS Google Scholar
Лабандейра CC: Вторжение континентов: корки цианобактерий на древесных членистоногих. Trends Ecol Evol 2005, 20 (5): 253–262.
Google Scholar
Лалонд К., Муччи А., Уэлле А., Гелинас Y: Сохранение органического вещества в отложениях с помощью железа. Nature 2012, 483 (7388): 198–200.
CAS Google Scholar
Lannerbro R: Описание некоторых структур, возможно, окаменелостей, в песчанике йотнианцев из Монгсбодарны в Далекарлии. Geologiska Föreningens i Stockholm Förhandlingar 1954, 76: 46–50.
Google Scholar
Лау К.М., Ким М.К., Ким К.М.: Азиатские муссонные аномалии, вызванные прямым воздействием аэрозолей. Clim Dyn 2006, 26: 855–864.
Google Scholar
Ласкано А., Миллер С.Л.: Сколько времени потребовалось, чтобы жизнь зародилась и превратилась в цианобактерии? J Mol Evol 1994, 39: 546–554.
CAS Google Scholar
Li W, Johnson CM, Beard BL: данные изотопов U – Th – Pb указывают на фанерозойский возраст окисления базальта на вершине 3,4 млрд лет. Earth Planet Sci Lett 2012, 319–320: 197–206.
Google Scholar
Лю Ю.З., Донг Г.Р., Ли Ч.З.: исследование факторов, влияющих на эрозию почвы, посредством экспериментов в аэродинамической трубе. Chin J Arid Land Res 1994, 7: 359–367.
Google Scholar
Lopez-Garcia P, Moreira D, Douzery EJP, Forterre P, van Zuilen M: летопись древних окаменелостей и ранняя эволюция (примерно от 3,8 до 0,5 млрд лет). В г. От солнца к жизни: мультидисциплинарный подход к истории жизни на Земле. Земля, луна и планеты . Под редакцией: Гарго М. Нью-Йорк: Спрингер; 2006: 247–290.
Google Scholar
Lowe DR: Строматолиты возрастом 3400 млн лет из архея Западной Австралии. Nature 1980, 284: 441–443.
Google Scholar
Lozupone CA, Knight R: Глобальные закономерности в бактериальном разнообразии. Proc Natl Acad Sci USA 2007, 104: 11436–11440.
CAS Google Scholar
Macfarlane AW, Danielson A, Holland HD: Геология и химия основных и микроэлементов позднеархейских профилей выветривания в группе Фортескью, Западная Австралия: последствия для атмосферного pO 2 . Precam Res 1994, 65: 297–317.
CAS Google Scholar
Мэдиган М.Т., Маррс Б.Л .: Экстремофилы. Scientific American 1997, 276 (4): 66–71.
Google Scholar
Мэдиган М., Мартинко Дж., Паркер Дж .: Брок: биология микроорганизмов . 10-е издание. Верхняя Сэдл-Ривер, Нью-Джерси: Пирсон-Прентис-холл; 2003.
Google Scholar
Малам Исса О, Ле Биссонне И, Дефарж С., Трише Дж .: Роль цианобактериального покрова в структурной стабильности песчаных почв в Сахалинской части западного Нигера. Geoderma 2001, 101: 15–30.
Google Scholar
Martini JEJ: позднеархейско-палеопротерозойский (2,6 млрд лет) палеозоль на ультрамафитах в восточной части Трансвааля, Южная Африка. Докембрийское исследование 1994, 67: 159–180.
CAS Google Scholar
Mazor G, Kidron GJ, Vonshak A, Abeliovich A: Роль цианобактериальных экзополисахаридов в структурировании микробных корок пустыни. FEMS Microbiol Ecol 1996, 21: 121–130.
CAS Google Scholar
МакКоннелл Р.Л .: Предварительный отчет о микроструктурах вероятного биологического происхождения из формации мескаль (протерозой) в центральной Аризоне. Докембрийское исследование 1974, 1 (3): 227–234.
Google Scholar
МакКиган К.Д., Кудрявцев А.Б., Шопф Дж.В.: Рамановские и ионно-микроскопические изображения графитовых включений в апатите из супракрустальных пород Акилии старше 3830 млн лет в западной Гренландии. Геология 2007, 35: 591–594.
Google Scholar
McTainsh GH, Strong CL: Роль эоловой пыли в экосистемах. Геоморфология 2007, 89 (1–2): 39–54.
Google Scholar
Медоу Дж. Ф., Забинский, Калифорния: Пространственная неоднородность эукариотических микробных сообществ в неизученной геотермальной диатомовой биологической почвенной корке: Йеллоустонский национальный парк, Вайоминг, США. FEMS Microbiol Ecol 2012, 82 (1): 182–191.
CAS Google Scholar
Mloszewska AM, Pecoits E, Cates NL, Mojzsis SJ, O’Neil J, Robbins LJ, Konhauser KO: Состав старейших железных образований Земли: супракрустальный пояс Нуввуагиттук (Квебек, Канада). Earth Planet Sci Lett 2012, 317–318: 331–342.
Google Scholar
Мойзсис С.Дж., Харрисон М.Т., Пиджон Р.Т.: данные изотопного состава кислорода из древних цирконов для жидкой воды на поверхности Земли 4300 млн лет назад. Nature 2001, 409: 178–181.
CAS Google Scholar
Mossman DJ, Minter WEL, Dutkiewicz A, Hallbauer DK, George SC, Hennigh Q, Reimer TO, Horscroft FD: Местное происхождение Витватерсрандского «углерода». Докембрийское исследование 2008, 164: 173–186.
CAS Google Scholar
Мулкиджанян А.Ю., Бычковц А.Ю., Диброва Д.В., Гальперин М.Ю., Кунин Е.В.: Происхождение первых клеток на земных бескислородных геотермальных полях. Proc Natl Acad Sci USA 2012, 109: E821-E830.
CAS Google Scholar
Myers JS: Протолиты зеленокаменного пояса Исуа 3,8–3,7 млрд лет, Западная Гренландия. Докембрийские исследования 2001, 105: 129–141.
CAS Google Scholar
Myers JS: Загадки Исуа: иллюзорные тектонические, осадочные, вулканические и органические особенности зеленокаменного пояса Исуа> 3,7 млрд лет, юго-запад Гренландии.В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 66–73.
Google Scholar
Надь М., Перес А., Гарсиа-Пичел Ф: Прокариотическое разнообразие биологических почвенных корок в пустыне Сонора (Национальный памятник кактуса Органной трубы, Аризона). FEMS Microbiol Ecol 2005, 54: 233–245.
CAS Google Scholar
Неер Д.А., Левинс С.А., Вайхт Т.Р., Дарби Б.Дж.: Сообщества микроартропод, связанные с биологическими почвенными корками на плато Колорадо и в пустынях Чиуауа. J Arid Environ 2009, 73: 672–677.
Google Scholar
Нельсон Д.Р.: Образование Земли и первый миллиард лет. В г. «Докембрийская Земля: темпы и события», \.События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 3–27.
Google Scholar
Nemergut DR, Costello EK, Hamady M, Lozupone C, Jiang L, Schmidt SK, Fierer N, Townsend AR, Cleveland CC, Stanish L, Knight R: Глобальные закономерности в биогеографии бактериальных таксонов. Environ Microbiol 2011, 13 (1): 135–144.
Google Scholar
Несбитт Х.В., молодой GM: Агрессивное архейское выветривание.В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 482–493.
Google Scholar
Николас Дж., Бильдген П. Взаимосвязь между расположением карстовых бокситов в северном полушарии, глобальной тектоникой и климатическими изменениями в течение геологического времени. Palaeogeogr Palaeoclimateol Palaeoecol 1979, 28: 205–239.
Google Scholar
Noffke N: Геобиология: микробные маты в песчаных отложениях с архейской эры до наших дней . Берлин: Springer; 2010.
Google Scholar
Ноффке Н.: Современный взгляд на древнюю жизнь: микробные маты в песчаных морских условиях от архейской эры до наших дней. В г. Ранняя жизнь на Земле: среда обитания. Среды и методы обнаружения .Под редакцией: Голдинг С.Д., Гликсон М. Дордрехт: Спрингер; 2011: 171–182.
Google Scholar
Ноффке Н., Гердес Г., Кленке Т., Крамбейн В.Е.: перспективы. Осадочные образования, вызванные микробами — новая категория в классификации первичных осадочных структур. J Sediment Res 2001, 71 (5): 649–656.
Google Scholar
Noffke N, Eriksson KA, Hazen RM, Simpson EL: Новое окно в раннеархейскую жизнь: микробные маты в старейших силикокластических приливных отложениях Земли (3.2 Ga Moodies Group, Южная Африка). Геология 2006, 34 (4): 253–256.
CAS Google Scholar
Noffke N, Beukes N, Bower D, Hazen RM, Swift DJP: Актуалистический взгляд на архейские миры — (циано) бактериальные осадочные структуры в силикокластической секции Nhlazatse, супергруппа 2,9 Ga Pongola, Южная Африка. Геобиология 2008, 6: 5–20.
CAS Google Scholar
Ноффке Н., Кристиан Д. Р., Хазен Р. М.: (циано) бактериальная экосистема в архее 3.49 Ga Dresser Formation, Пилбара, Западная Австралия. Ежегодное собрание GSA в Миннеаполисе. Документ № 56–11. Рефераты Геологического общества Америки с программами 2011, 43 (5): 159.
Google Scholar
Nutman AP: Древность океанов и континентов. Элементы 2006, 2: 223–227.
CAS Google Scholar
Nutman AP, McGregor VR, Friend CRL, Bennett VC, Kinny PD: Комплекс Itsaq Gneiss на юге Западной Гренландии; самый обширный в мире рекорд ранней эволюции земной коры (3900–3600 млн лет назад). Докембрия Res 1996, 78: 1–39.
CAS Google Scholar
Nutman AP, Friend CRL, Bennett VC: свидетельства образования 3650–3600 млн лет назад северной оконечности комплекса Itsaq Gneiss, Гренландия: значение для ранней архейской тектоники. Тектоника 2002, 21 (1): 5–1–5–28.
Google Scholar
Nutman AP, Friend CRL, Bennett VC, Wright D, Norman MD: ≥3700 млн лет до-метаморфического доломита, образованного микробным посредником в супракрустальном поясе Исуа (W.Гренландия): простые доказательства ранней жизни? Докембрийский отчет 2010, 183: 725–737.
CAS Google Scholar
О’Нил Дж., Фрэнсис Д., Карлсон Р.В.: Влияние зеленокаменного пояса Нуввуагиттук на формирование ранней земной коры. J Petrol 2011, 52: 985–1009.
Google Scholar
Оч Л.М., Шилдс-Чжоу Г.А.: Неопротерозойское событие оксигенации.Нарушения окружающей среды и биогеохимический цикл. Earth Sci Rev. 2012, 110 (1–4): 26–57.
CAS Google Scholar
Омото H: свидетельства ранней эволюции атмосферного кислорода и наземной биоты в гапалеозолях до 2.2. Геология 1996, 24: 1135–1138.
CAS Google Scholar
Омото H: архейская атмосфера, гидросфера и биосфера.В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 361–368.
Google Scholar
Ohmoto H, Watanabe Y, Allwood A, Burch I, Knauth P, Yamaguchi K, Johnson I, Altinok E: формирование вероятных латеритных почв ~ 3,43 млрд лет в кратоне Пилбара, Западная Австралия. Geochimica et Cosmochimica Acta, Дополнение 2007, 71 (15): A733.
Google Scholar
Омелон С.Р., Поллард У.Х., Феррис Ф.Г .: Химическая и ультраструктурная характеристика криптоэндолитических местообитаний в высокой Арктике. Geomicrobiol J 2006, 23: 189–200.
CAS Google Scholar
О’Нил Дж., Карлсон Р.В., Фрэнсис Д., Стивенсон Р.К .: Ответ на комментарий к «Неодиму-142, свидетельствующему о наличии основной коры Гаде». Наука 2009, 325: 267b.
Google Scholar
Pace N: молекулярный взгляд на микробное разнообразие и биосферу. Наука 1997, 276: 734–740.
CAS Google Scholar
Палмер Дж. А., Филлипс Г. Н., Маккарти Т. С.: Палеопочвы и их отношение к составу докембрийской атмосферы. J Geol 1989, 97: 77–92.
CAS Google Scholar
Пандит М.К., де Валл Х., Чаухан Н.К .: Палеопочва на стыке архея и протерозоя на северо-западе Индии. J Earth Syst Sci 2008, 117 (3): 201–209.
CAS Google Scholar
Папино Д., ДеГрегорио Б.Т., Коди Г.Д., Фрис М.Д., Мойзсис С.Дж., Стил А., Страуд Р.М., Фогель М.Л.: Древний графит в эоархейских кварц-пироксеновых породах из Акилии на юго-западе Гренландии I: петрографические и спектроскопические характеристики. Geochim Cosmochim Acta 2010, 74: 5862–5883.
CAS Google Scholar
Papineau D, DeGregorio BT, Stroud RM, Steele A, Pecoits E, Konhauser K, Wang J, Fogel ML: Древний графит в эоархейской кварц-пироксеновой породе из Акилии на юге Западной Гренландии II: изотопный и химический составы и сравнение с палеопротерозойскими полосчатыми железными образованиями. Geochim Cosmochim Acta 2010, 74: 5884–5905.
CAS Google Scholar
Папино Д., Де Грегорио Б.Т., Коди Г.Д., О’Нил Дж., Стил А., Страуд Р.М., Фогель М.Л.: Молодой слабокристаллический графит в пластовой формации Нуввуагиттук возрастом 3,8 миллиарда лет. Nat Geosci 2011, 4 (6): 376–379.
CAS Google Scholar
Пол EA, Collins HP, Leavitt SW: Динамика устойчивого почвенного углерода сельскохозяйственных почв Среднего Запада, измеренная по естественному содержанию 14 C. Geoderma 2001, 104 (3–4): 239–256.
CAS Google Scholar
Портер М.Л., Энгель А.С., Кинкль Б., Кейн Т.К .: Взаимосвязь продуктивности и разнообразия в сульфидных карстовых системах на основе хемолитоавтотрофов. Int J Speleol 2009, 38: 27–40.
Google Scholar
Potts M: Механизмы устойчивости к высыханию у цианобактерий. Eur J Phycol 1999, 34: 319–328.
Google Scholar
Поттс М., Фридман Э.И.: Влияние водного стресса на криптоэндолитические цианобактерии из горячих пустынных пород. Arch Microbiol 1981, 130: 267–271.
CAS Google Scholar
Прасад Н., Роско С.М.: Свидетельства атмосферных изменений от бескислородного до оксидного в течение 2,45–2,22 млрд лет в нижних и верхних субгуронских палеопочвах, Канада. Катена 1996, 27: 105–121.
CAS Google Scholar
Праве AR: Жизнь на суше в протерозое: свидетельства из торридонских скал на северо-западе Шотландии. Геология 2002, 30 (9): 811–814.
Google Scholar
Пуфаль П.К., Хиатт Э.Э .: Оксигенация системы атмосфера-океан Земли: обзор физических и химических седиментологических реакций. J Mar Petrol Geol 2012, 32 (1): 1–20.
CAS Google Scholar
Расмуссен B: Нитчатые микрофоссилии в вулканогенных массивных сульфидных месторождениях возрастом 3 235 миллионов лет. Nature 2000, 405: 676–679.
CAS Google Scholar
Расмуссен Б., Флетчер И. Р., Брокс Дж. Дж., Килберн М. Р.: Переоценка первого появления эукариот и цианобактерий. Nature 2008, 455 (7216): 1101–1104.
CAS Google Scholar
Расмуссен Б., Блейк Т.С., Флетчер И.Р., Килберн М.Р.: Доказательства микробной жизни в синседиментарных полостях из земных сред 2,75 млрд лет. Геология 2009, 37: 423–426.
Google Scholar
Редди С.Г., Гарсия-Пичел Ф .: Сообщество и филогенетическое разнообразие биологических почвенных корок на плато Колорадо изучены с помощью молекулярного дактилоскопирования и интенсивного культивирования. Microb Ecol 2006, 52: 345–357.
Google Scholar
Реймер Т.О.: богатые глиноземом породы из раннего докембрия кратона Каапваал как индикаторы палеопочв и как продукты других реакций разложения. Докембрия Res 1986, 32: 155–179.
CAS Google Scholar
Retallack GJ: Летопись окаменелостей почв. Палеопочвы : их распознавание и интерпретация .Отредактировал: Райт П.В. Оксфорд: Блэквелл; 1986: 1–57.
Google Scholar
Retallack GJ: Переоценка палеопочвы возрастом 2200 млн лет около Waterval Onder, Южная Африка. Докембрийское исследование 1986, 32: 195–232.
CAS Google Scholar
Retallack GJ: Почвы прошлого . 2-е издание. Лондон: Blackwell Science; 2001.
Google Scholar
Retallack GJ: Кембрийско-ордовикские неморские окаменелости из Южной Австралии. Алчеринга 2009, 33: 355–391.
Google Scholar
Retallack: Критерии различения микробных матов и грунтов. В: Noffke N, Chafetz H (ed) Микробные маты в силикокластических осадочных системах с течением времени. Специальная публикация SEPM № 101. Общество осадочной геологии, Талса, ОК, стр. 2001, 139–152.
Google Scholar
Retallack GJ: Эдиакарская жизнь на суше. Nature 2013, 493 (7430): 89–92.
Google Scholar
Retallack GJ, Mindszenty A: Хорошо сохранившиеся палеопочвы позднего докембрия с северо-запада Шотландии. J Sediment Res 1994, A64: 264–281.
CAS Google Scholar
Reynolds R, Belnap J, Reheis M, Lamothe P, Luiszer F: Эоловая пыль в почвах плато Колорадо: поступление питательных веществ и недавнее изменение источника. Proc Natl Acad Sci USA 2001, 98: 7123–7127.
CAS Google Scholar
Рино С., Комия Т., Уиндли Б.Ф., Катаяма С., Мотоки А., Хирата Т.: значительное эпизодическое увеличение роста континентальной коры, определенное по возрасту циркона в речных песках; значение переворота мантии в раннем докембрии. Phys Earth Planet Inter 2004, 146: 369–394.
CAS Google Scholar
Римлянам Б.В., Грэхем С.А.: Глубокая перспектива связей суши и океана в осадочной летописи. Annu Rev Mar Sci 2013, 5: 69–94.
Google Scholar
Розинг М.Т., Фрей Р: богатые ураном отложения архейского морского дна из Гренландии: признаки кислородного фотосинтеза> 3700 млн лет. Earth Planet Sc Lett 2004, 217: 237–244.
CAS Google Scholar
Розинг М.Т., Берд Д.К., Сон Н.Х., Глассли В., Альбареде Ф .: Возникновение континентов — очерк геологических последствий фотосинтеза. Palaeogeogr Palaeoclimateol Palaeoecol 2006, 232: 99–113.
Google Scholar
Росси Ф., Потрафка Р. М., Гарсия-Пичель Ф., Де Филиппис Р: Роль экзополисахаридов в повышении гидравлической проводимости биологических почвенных корок. Soil Biol Biochem 2012, 46: 33–40.
CAS Google Scholar
Рожь Р., Голландия HD: Жизнь, связанная с 2.Эфемерный пруд 76 Га? Свидетельства из палеопочвы горы Роу №2. Геология 2000, 28: 483–486.
CAS Google Scholar
Сантош М.: Краткий обзор последних концептуальных моделей тектоники суперконтинента в связи с динамикой мантии, эволюцией жизни и окружающей средой на поверхности. J Geodyn 2010, 50 (3–4): 116–133.
Google Scholar
Sarbu SM, Kane TC, Kinkle BK: пещерная экосистема, основанная на химиоавтотрофах. Наука 1996, 272: 1953–1955.
CAS Google Scholar
Шау М., Хендерсон Дж. Б.: Архейское химическое выветривание в трех местах на канадском щите. Дев Докембрийская геология 1983, 7: 81–116.
Google Scholar
Шибер Дж., Бозе П.К., Эрикссон П.Г., Банерджи С., Саркар С., Альтерманн В., Катуно О. (ред.): Атлас микробных матов, сохранившихся в пласте кремнистой породы .Амстердам: Эльзевир; 2007.
Google Scholar
Schiffbauer JD, Xiao S: Новое применение сфокусированной ионно-лучевой электронной микроскопии (FIB-EM) для подготовки и анализа ультраструктур микрофоссилий: новый взгляд на сложность ранних эукариотических организмов. Palaios 2009, 24: 616–626.
Google Scholar
Schiffbauer JD, Yin L, Bodnar RJ, Kaufman AJ, Meng F, Hu J, Shen B, Yuan X, Bao H, Xiao S: Ультраструктурная и геохимическая характеристика архейско-палеопротерозойских частиц графита: последствия для распознавания следов жизнь в сильно метаморфизованных породах. Астробиология 2007, 7: 684–704.
CAS Google Scholar
Шиффбауэр Дж. Д., Уоллес А. Ф., Хантер Дж. Л. мл., Ковалевски М., Боднар Р. Дж., Сяо С.: Структурные и химические изменения, вызванные термическим воздействием на органические окаменелости: экспериментальный подход к пониманию сохранности окаменелостей в метаотложениях. Геобиология 2012, 10 (5): 402–423.
CAS Google Scholar
Schirrmeister BE, de Vosb JM, Antonelli A, Bagheri HC: Эволюция многоклеточности совпала с увеличением разнообразия цианобактерий и Великим событием окисления. PNAS-USA 2013. 10.1073 / pnas.12090
Google Scholar
Schmidt SK, Reed SC, Nemergut DR, Stuart-Grandy A, Cleveland CC, Weintraub MN, Hill AW, Costello EK, Meyer AF, Neff JC, Martin AM: самые ранние стадии сукцессии экосистемы на возвышенности ( 5000 метров над уровнем моря), недавно дегляцированные почвы. Proc Biol Sci 2008, 275 (1653): 2793–2802.
CAS Google Scholar
Schopf JW: Микрофлора образования горьких источников, поздний докембрий, центральная Австралия. J Paleontol 1968, 42: 651–688.
Google Scholar
Schopf JW (Ed): Самая ранняя биосфера Земли . Принстон, Нью-Джерси: Издательство Принстонского университета; 1983.
Google Scholar
Schopf JW: Палеобиология архея. В Протерозойская биосфера . Под редакцией: Schopf JW, Klein C. Нью-Йорк: Cambridge University Press; 1992.
Google Scholar
Schopf JW: Протерозойские прокариоты: родство, геологическое распределение и тенденции эволюции.В Протерозойская биосфера . Под редакцией: Schopf JW, Klein C. Нью-Йорк: Cambridge University Press; 1992.
Google Scholar
Schopf JW, Klein C (Eds): Протерозойская биосфера . Нью-Йорк: издательство Кембриджского университета; 1992.
Google Scholar
Шопф Дж. У., Вальтер М. Р., Руиджи К.: Самые ранние свидетельства существования жизни на Земле. Докембрийские исследования 2007, 158: 139–140.
CAS Google Scholar
Шварцман Д.В., Фольк Т.: Биотическое усиление выветривания и обитаемости земли. Nature 1989, 340: 457–460.
Google Scholar
Shear WA: Раннее развитие наземных экосистем. Nature 1991, 351: 283–289.
Google Scholar
Шелдон Н.Д .: Микробиологические осадочные структуры в ок.1100 млн лет назад — континентальный разлом Северной Америки. В Микробные маты в силикокластических осадочных системах с течением времени. Специальная публикация SEPM № 101 . Под редакцией: Ноффке Н., Чафец Х. Талса, ОК: Общество осадочной геологии; 2012: 153–162.
Google Scholar
Shen Y, Buick R: древность микробной сульфатредукции. Earth Sci Rev. 2004, 64: 243–272.
CAS Google Scholar
Шен Й, Фаркуар Дж., Мастерсон А., Кауфман А. Дж., Бьюик Р.: Оценка роли микробного восстановления сульфата в раннем архее с использованием четверной изотопной систематики. Earth Planet Sc Lett 2009, 279: 383–391.
CAS Google Scholar
Shephard KL: Испарение воды из слизи студенистого водорослевого сообщества. Br Phycol J 1987, 22: 181–185.
Google Scholar
Сиглер В.В., Кривий С., Зейер Дж .: Бактериальная сукцессия в ледниковых почвах переднего поля, характеризующаяся структурой сообщества, активностью и условно-динамической динамикой роста. Microb Ecol 2002, 44: 306–316.
CAS Google Scholar
Simpson WS, Simpson EL, Wizevich MC, Malenda HF, Hilbert-Wolf HL, Tindall SE: сохранившаяся биологическая почвенная корка позднего мелового периода в покрывающей пачке песчаника, формация Вахвип, национальный памятник Гранд-Лестница-Эскаланте, Юта, Палеоклиматический район подразумеваемое. Sediment Geol 2010, 230: 139–145.
Google Scholar
Сингх С.П., Кумари С., Растоги Р.П., Сингх К.Л., Рича С.Р.П .: Фотозащитные и биотехнологические возможности пигмента оболочки цианобактерий, скитонемина. Afr J Biotechnol 2010, 9: 580–588.
CAS Google Scholar
Sinha RP, Häder DP: УФ-индуцированное повреждение и восстановление ДНК: обзор. Photochem Photobiol Sci 2002, 1: 225–236.
CAS Google Scholar
Скотницки С.Дж., Кнаут Л.П.: Мескальный палеокарст среднего протерозоя, Центральная Аризона, США: развитие карста, окварцевание и пещерные отложения. J Sediment Res 2007, 77 (12): 1046–1062.
Google Scholar
Sleep NH, Bird DK: Ниши префотосинтетической биосферы и геологическое сохранение древнейшей экологии Земли. Геобиология 2007, 5: 101–117.
CAS Google Scholar
Smith SE, Читать D: Микоризный симбиоз . 3-е издание. Нью-Йорк: Эльзевир; 2008 г.
Google Scholar
Сквайр Р.Дж., Стюарт И.Р., Занг В.Л.: Акритархи в полидеформированных и сильно измененных кембрийских породах в западной Виктории. Aust J Earth Sci 2006, 53: 697–705.
Google Scholar
Stern RJ, Scholl DW: Инь и Ян создания и разрушения континентальной коры тектоническими процессами плит. Int Geol Rev 2010, 52: 1–31.
Google Scholar
Strother PK, Battison L, Brasier MD, Wellman CH: самые ранние неморские эукариоты на Земле. Nature 2011, 473: 505–509.
CAS Google Scholar
Stüeken EE, Catling DC, Buick R: Вклад в круговорот серы на суше в позднем архее. Nat Geosci 2012, 5: 722–725.
Google Scholar
Summons RE, Jahnke LL, Hope JM, Logan GA: 2-метилгопаноиды как биомаркеры кислородного фотосинтеза цианобактерий. Nature 1999, 400: 554–557.
CAS Google Scholar
Sun HJ, Friedmann EI: Рост в масштабе геологического времени в антарктическом криптоэндолитическом микробном сообществе. Geomicrobiol J 1999, 16: 193–202.
Google Scholar
Такеучи Н., Кохима С., Секо К.: Структура, образование, процесс потемнения материала, уменьшающего альбедо (криоконита) на гималайском леднике: гранулированный водорослевый мат, растущий на леднике. Arct Antarct Alp Res 2001, 33: 115–122.
Google Scholar
Тейлор Т.Н., Тейлор Э.Л., Крингс М: Палеоботаника: биология и эволюция ископаемых растений . 2-е издание. Амстердам: Эльзевир; 2009.
Google Scholar
Thiry M, Simon-Coincon R (Eds): Палеоповерхности и связанные с ними континентальные отложения. Специальная публикация 27 Международной ассоциации седиментологов .Оксфорд: Blackwell Science; 1999.
Google Scholar
Тайс М.М., Лоу Д.Р.: Фотосинтетические микробные маты в океане возрастом 3416 млн лет. Nature 2004, 431: 549–552.
CAS Google Scholar
Тирки Дж., Адхикари С.П.: Цианобактерии в биологических корках почвы Индии. Curr Sci 2005, 89: 515–521.
Google Scholar
Tomitani A, Knoll AH, Cavanaugh CM, Ohno T: Эволюционная диверсификация цианобактерий: молекулярно-филогенетические и палеонтологические перспективы. Proc Natl Acad Sci USA 2006, 103: 5442–5447.
CAS Google Scholar
Trendall AF, Blockley JG: Докембрийские железистые образования. В г. Докембрийская Земля: темпы и события. События в геологии докембрия, т. 12 . Под редакцией: Эрикссон П.Г., Альтерманн В., Нельсон Д.Р., Мюллер В.У., Катуняну О. Амстердам: Эльзевир; 2004: 403–420.
Google Scholar
Уэно Ю., Ямада К., Йошида Н., Маруяма С., Исодзаки Ю.: данные по флюидным включениям для микробного метаногенеза в раннюю архейскую эру. Nature 2006, 440: 516–519.
CAS Google Scholar
Ван Кранендонк MJ: Архейская тектоника 2004: обзор. Докембрийские исследования 2004, 131: 143–151.
CAS Google Scholar
Ван Кранендонк MJ: Морфология строматолита как индикатор биогенности древнейших окаменелостей Земли из кратона Пилбара 3,5–3,4 млрд лет в Западной Австралии.В г. Успехи в геобиологии строматолитов. Конспект лекций по наукам о Земле, т. 131 . Под редакцией: Рейтнер Дж., Кверик Н.В., Арп Г. Германия: Шпрингер; 2011.
Google Scholar
Кранендонк MJ Van, Smithies RH, Bennett V (Eds): самых старых горных пород Земли. Разработки в докембрийской геологии, серия 15 . Амстердам: Эльзевир; 2007a.
Google Scholar
Ван Кранендонк М.Дж., Смитис Р.Х., Хикман А.Х., Чемпион округ Колумбия: Обзор: вековая тектоническая эволюция архейской континентальной коры: взаимодействие горизонтальных и вертикальных процессов в формировании кратона Пилбара, Австралия. Terra Nova 2007, 19 (1): 1–38.
CAS Google Scholar
Ван Кранендонк М.Дж., Филиппот П., Лепот К., Бодоркос С., Пирайно Ф .: Геологические условия древнейших окаменелостей Земли в ок. Формация Дрессер, 3,5 млрд лет, кратон Пилбара, Западная Австралия. Докембрийские исследования 2008, 167: 93–124.
CAS Google Scholar
ван Зуилен М.А., Лепланд А, Аррениус Г.: Переоценка свидетельств самых ранних следов жизни. Nature 2002, 418: 627–630.
CAS Google Scholar
Voigt E: Tonrollen als Potentielle Pseudofossilien. Нат Мус 1972, 102 (11): 401–410.
Google Scholar
Wacey D, Kilburn MR, Saunders M, Cliff J, Brasier MD: микрофоссилии клеток, метаболизирующих серу, в скалах Западной Австралии возрастом 3,4 миллиарда лет. Nat Geosci 2011, 4: 698–702.
CAS Google Scholar
Уолш М.М.: микрофоссилий и возможные микрофоссилии из раннеархейской группы Onverwacht, Барбертон Маунтин Лэнд, Южная Африка. Докембрийское исследование 1992, 54: 271–292.
CAS Google Scholar
Уолш М.М., Лоу Д.Р.: Нитевидные микрофоссилии из группы Onverwacht Group 3500 млн лет назад, Барбертон Маунтин Лэнд, Южная Африка. Nature 1985, 314: 530–532.
Google Scholar
Уолш М.М., Лоу Д.Р.: Способы накопления углеродистого вещества в раннем архее: петрографические и геохимические исследования углеродистых кремней супергруппы Свазиленд. В г. Геологическая эволюция зеленокаменного пояса Барбертона, Южная Африка, Специальная статья Геологического общества Америки 329 Отредактировано: Lowe DR, Byerly GR. 1999, 115–132.
Google Scholar
Уолтер М.Р.: Архейские строматолиты: свидетельство самого раннего бентоса на Земле.В г. Самая ранняя биосфера Земли г. Под редакцией: Schopf JW. Принстон: Издательство Принстонского университета; 1983: 187–213.
Google Scholar
Walter MR, Buick R, Dunlop JSR: Строматолиты возрастом 3 400–3 500 млн лет из района Северного полюса, Западная Австралия. Nature 1980, 284: 443–445.
Google Scholar
Watanabe Y, Martini JEJ, Ohmoto H: Геохимические данные о наземных экосистемах 2.6 миллиардов лет назад. Nature 2000, 408: 574–578.
CAS Google Scholar
Велч С.А., Баркер В.В., Банфилд Дж.Ф.: Микробные внеклеточные полисахариды и растворение плагиоклаза. Geochim Cosmochim Acta 1999, 63: 1405–1419.
CAS Google Scholar
Westall F: Ранние годы жизни: природа, распространение и эволюция. В г. Истоки и эволюция жизни г.Под редакцией: Гарго М., Лопес-Гарсия П., Мартин Х. Кембридж: астробиологическая перспектива. Издательство Кембриджского университета; 2010: 391–413.
Google Scholar
Вестолл Ф., Де Вит М.Дж., Данн Дж., Ван Дер Гааст С., Де Ронд С., Гернеке Д. Ранние архейские ископаемые бактерии и биопленки в мелководных отложениях под гидротермальным влиянием, пояс Барбертона Гринстоуна, Южная Африка Докембрия Res 2001, 106: 91–112.
Google Scholar
Westall F, de Vries ST, Nijman W, Rouchon V, Orberger B, Pearson V, Watson J, Verchovsky A, Wright I, Rouzaud JN, Marchesini D, Anne S: The 3.466 Ga Kitty’s Gap Chert, раннеархейская микробная экосистема. В г. Процессы на ранней Земле. Специальная статья Геологического общества Америки 405 . Под редакцией: Реймольд У., Гибсон Р. Боулдер, Колорадо: Геологическое общество Америки; 2006a: 105–131.
Google Scholar
Westall F, de Ronde CEJ, Southam G, Grassineau N, Colas M, Cockell C, Lammer H: последствия субаэрального микробного мата возрастом 3,472–3,333 млрд лет из пояса Барбертон-Гринстоун, Южная Африка, для УФ-окружающей среды условия на ранней Земле. Phil Trans R Soc B 2006, 361: 1857–1875.
CAS Google Scholar
Westall F, Cavalazzi B, Lemelle L, Marrocchi Y, Rouzaud JN, Simionovici A, Salomé M, Mostefaoui S, Andreazza C, Foucher F, Toporski J, Jauss A, Thiel V, Southam G, MacLean L, Wirick С., Хофманн А., Мейбом А., Роберт Ф., Дефарж С. Влияние кальцификации in situ на фотосинтез в микробной биопленке возрастом 3,3 млрд лет из зеленокаменного пояса Барбертон, Южная Африка. Earth Planet Sci Lett 2011, 310 (3–4): 468–479.
CAS Google Scholar
Белый D: Физиология и биохимия прокариот . 2-е издание. Оксфорд: издательство Оксфордского университета; 2000.
Google Scholar
Whitton BA, Potts M: Экология цианобактерий: их разнообразие во времени и пространстве . Дордрехт: Клувер; 2000 г.
Google Scholar
Wilde SA, Valley JW, Peck WH, Graham CM: Обломочные цирконы свидетельствуют о существовании континентальной коры и океанов на Земле 4,4 млрд лет назад. Nature 2001, 409: 175–178.
CAS Google Scholar
Уильямс Дж. Д., Добровольск, JP, Западный Северо-Восток, Джиллетт Д.А.: Влияние микрофитной коры на ветровую эрозию. Trans ASAE 1995, 38: 131–137.
Google Scholar
Уильямс А.Дж., Бак Б.Дж., Бейен М.А.: Биологические корки почвы в пустыне Мохаве, США: микроморфология и почвообразование. Soil Sci Soc Am J 2012, 76 (5): 1685–1695.
CAS Google Scholar
Виндли Б. Обзор и история изучения ранних пород Земли. В числе самых старых горных пород Земли. Разработки в докембрийской геологии, серия 15 .Под редакцией: Ван Кранендонк MJ, Smithies RH, Беннет В. Амстердам: Elsevier; 2007: 3–7.
Google Scholar
Райт В. П.: Среда-предшественник для колонизации сосудистых растений. Phil Trans R Soc London B 1985, 309: 143–145.
Google Scholar
Xiao S, Kaufman AJ (Eds): Неопротерозойская геобиология и палеобиология. Разделы геобиологии, т.27 . Дордрехт: Спрингер; 2006.
Google Scholar
Сяо С., Кнаут LP: Палеонтология: окаменелости попадают на сушу. Nature 2013, 493 (7430): 28–29.
Google Scholar
Xiong J, Fischer WM, Inoue K, Nakahara M, Bauer CE: Молекулярные доказательства ранней эволюции фотосинтеза. Наука 2000, 289: 1724–1730.
CAS Google Scholar
Ян В., Голландия HD: палеопочва Хекпорта в пласте 1 в Гарбороне, Ботсвана: формирование почвы во время Великого окислительного события. Am J Sci 2003, 303: 187–220.
CAS Google Scholar
Ясуи А., Маккриди С.Дж.: Альтернативные пути восстановления повреждений ДНК, вызванных ультрафиолетом. Bioessays 1998, 20 (4): 291–297.
CAS Google Scholar
Ю.Г., Якобсен С.Б.: Быстрая аккреция Земли при позднем ударе гиганта, образующего Луну. PNAS USA 2011, 108 (43): 17604–9.
CAS Google Scholar
Занг В.Л.: Отложение и деформация отложений позднего архея и сохранение микрофоссилий в области Харрис Гринстоун, кратон Голер, Южная Австралия. Докембрийские исследования 2007, 156: 107–124.
CAS Google Scholar
Zbinden EA, Holland HD, Feakes CR, Dobos SK: Палеопочва Осетрового водопада и состав атмосферы 1.1 млрд лет назад. Докембрия Res 1988, 42: 141–163.
CAS Google Scholar
Чжан Дж .: Наблюдение за воздействием водорослей на субаэральное карстовое осаждение. Geogr Res 1992, 11 (2): 26–33.
Google Scholar
Чжао Б., Робб Л.Дж., Харрис С., Йордан Л.Дж .: Происхождение гидротермальных флюидов и золотого оруденения, связанного с контактным рифом Вентерсдорп, бассейн Витватерсранд, Южная Африка: ограничения изотопов S, O и H.В г. Процессы на ранней Земле. Специальная статья Геологического общества Америки 405 . Отредактировано: Reimold WU, Gibson RL. Боулдер, Колорадо: Геологическое общество Америки; 2006: 333–352.
Google Scholar
Журавлев А.Ю., Riding R (Eds): Экология кембрийской радиации . Нью-Йорк: издательство Колумбийского университета; 2001.
Google Scholar
Zonneveld KAF, Versteegh GJM, Kasten S, Eglinton TI, Emeis KC, Huguet C, Koch BP, De Lange GJ, De Leeuw JW, Middelburg JJ, Mollenhauer G, Prahl FG, Rethemeyer SG J, Sective Preservation: органического вещества в морской среде; процессы и влияние на осадочную запись. Биогеонауки 2010, 7: 483–511.
CAS Google Scholar
Дикая природа и наземные экосистемы — RMRS
Животный мир и наземные экосистемыПрограмма по дикой природе и наземным экосистемам хорошо согласуется с национальными целями в области дикой природы и рыб. Прогресс в достижении этих целей представляет собой вклад всех научных программ станций, а не только дикой природы и наземных местообитаний.Ниже приводится краткое описание целей программы.
Поддержание видов и экосистем, вызывающих озабоченность
В границах RMRS встречается больше экосистем, чем на любой другой станции, включая прерии, умеренные и тропические степи, пустыни, хвойные и прибрежные леса и тундру. Такое разнообразие экосистем выражается в разнообразии растений и животных, при этом в регионе RMRS обитает более 16 800 растений и животных. Более 3800 видов представляют интерес для сохранения, а 217 официально внесены в список находящихся под угрозой исчезновения.Поскольку более 90 процентов апелляций и судебных разбирательств по решениям по управлению земельными ресурсами основаны на проблемах дикой природы и рыб, исследования, которые предоставляют достоверную научную информацию, имеют решающее значение для выполнения юридических обязательств и сохранения находящихся под угрозой исчезновения и уязвимых видов и связанных с ними экосистем. Многоуровневые исследования выявляют множество факторов, которые влияют на сохранение видов, сообществ и экосистем, вызывающих озабоченность, таких как северные ястребы-тетеревятники в сосновых лесах на юго-западе пондероза и лесные хищники в Северных Скалистых горах.Контекст для большинства исследований включает экологические взаимодействия внутри и между сообществами растений, водных и наземных животных в самых разных экосистемах на всем Западном Межгорье и за его пределами.
Общие сведения об эффектах общественного пользования
Исследования информируют федеральные, государственные, племенные и местные агентства ресурсов о взаимодействиях между людьми и рыбами / дикой природой. Это обеспечивает лучшее понимание эффектов общественного использования, так что они могут быть смягчены соответствующими управленческими действиями.Хотя большая часть исследований сосредоточена на эффектах отдыха (OHV, туристы, орнитологи, хели-ски), аналогичные направления исследований оценивают эффекты коридоров передачи и косвенные эффекты шума, возникающие в результате землеустройства. В настоящее время проводятся многочисленные исследования для выяснения социальных и экономических ценностей, связанных с безвредным и непотребительским использованием рыбы / диких животных. Прогнозные модели среды обитания предоставляют ряд управленческих альтернатив для одновременного улучшения товарного и нетоварного использования рыбы / диких животных.
Управление наземными и водными средами обитания
Большинство оставшихся местообитаний для многих видов, вызывающих озабоченность, в настоящее время находятся в национальных лесах и пастбищах. Ученые РМРС отвечают на вопросы о количестве, виде, распределении и связности местообитаний, имеющих решающее значение для сохранения и численности этих видов. Эти исследователи также разрабатывают надежные методы мониторинга изменений видов и местообитаний и определения мест обитания, наиболее чувствительных к управлению или важных для сохранения видов.Ученые также исследуют пространственные и временные аспекты процессов, которые создают и поддерживают среды обитания, имеющие решающее значение для сохранения биоразнообразия, включая роль огня.
Оценка результатов использования земли / воды и природных нарушений
Широкий спектр антропогенных и природных нарушений влияет на наши луга, леса и водные пути. Природные и предписанные пожары, лесоводственные предписания, выпас скота, распространение и борьба с инвазивными видами, засуха, глобальное изменение климата, вспышки и фрагментация насекомых и болезней — все это влияет и потенциально угрожает средам обитания диких животных.Исследователи определяют непосредственные, долгосрочные и кумулятивные эффекты беспокойства у видов, вызывающих озабоченность и интерес, на Межгорном Западе.
Что такое экосистема суши? — Определение, примеры и типы — Видео и стенограмма урока
Типы и примеры наземных экосистем
Хотя с течением времени было разработано множество классификационных схем, в настоящее время принято считать, что существует шесть типов наземных экосистем.К ним относятся тайга, тундра, лиственные леса, луга, влажные тропические леса и пустыни.
Тайга — леса с холодным климатом, расположенные в северных широтах. Тайги — крупнейшая в мире наземная экосистема, на которую приходится около 29% лесов Земли. Самые крупные таежные экосистемы находятся в Канаде и России. Тайги известны своим субарктическим климатом с чрезвычайно холодной зимой и мягким летом. В основном они состоят из хвойных деревьев, таких как сосны, хотя есть и некоторые другие лиственные деревья, такие как ель и вяз, которые приспособились к жизни в этих областях, которые получают мало прямого солнечного света в течение большей части года.Тайги являются домом для крупных травоядных, таких как лоси, лоси и бизоны, а также для всеядных животных, таких как медведи.
тундра экосистем мира расположены в основном к северу от полярного круга. Они состоят из короткой растительности и практически без деревьев. Почва промерзает и покрыта вечной мерзлотой большую часть года. Карибу, белые медведи и овцебык — одни из самых известных видов, которые называют тундру своим домом.
Лиственные леса экосистемы составляют восточную половину Северной Америки и большую часть Европы.У них обычно средняя годовая температура 50 градусов по Фаренгейту, и в среднем в них выпадает около 30-60 дюймов дождя в год. В этих лесах обитает множество диких животных, в том числе олени, медведи, лисы, а также множество видов деревьев, кустарников и цветов. Если вы живете или когда-либо путешествовали на востоке США, вы бывали в лиственном лесу.
Луга также известны как равнины и прерии. Если вы представите «Дикий Запад» с перекати-поле, летящим по равнинам, и большими стадами оленей и буйволов, такими как прерии Небраски, то вы представляете луга.Эти экосистемы характеризуются 20-35 дюймовым количеством осадков в год и преобладающим покрытием различных видов трав. Они также известны своей очень плодородной почвой.
Влажные тропические леса экосистемы обладают наибольшим разнообразием растений и животных из всех экосистем. Они существуют в 28 градусах к северу или югу от экватора и известны своими высокими средними температурами, а также значительным количеством осадков. Пышные леса Амазонки в Южной Америке — классический пример тропических дождевых лесов.
Закройте глаза и представьте, что идете по горным песчаным дюнам, когда палящее солнце иссушает вам рот, пока вы ищете воду. Эта сцена дает вам возможность взглянуть на последнюю наземную экосистему, которой является пустыня . Эти экосистемы жаркие и засушливые, с очень небольшим количеством растительности и очень немногими видами животных, которые называют их своим домом, кроме рептилий, паукообразных и птиц. Большая пустыня Сахара занимает большую часть северной части африканского континента.
Краткое содержание урока
Как видите, все экосистем , которые полностью существуют на суше, являются наземными экосистемами .Они варьируются от самых холодных мест на Земле на крайних северных широтах до самых жарких пустынь , обнаруженных вокруг экватора. Каждая экосистема населена видами растений и животных, которые эволюционировали, чтобы процветать в них.
Результаты обучения
К концу этого урока вы должны уметь:
- Определять наземные экосистемы
- Назовите и опишите наземные экосистемы Земли
Наземные экосистемы
Возврат к экосистемам
Landbirds
Тихоокеанский гагара плавает в небольшом озере в дельте реки Колвилл.
(Источник: Райан Аскрен, Геологическая служба США. Общественное достояние)
Виды птиц, населяющие преимущественно наземные местообитания, известные под общим названием «наземные птицы», составляют самый крупный и наиболее экологически разнообразный компонент орнитофауны Аляски. Среда обитания наземных птиц варьируется от тропических лесов с умеренным климатом на юго-востоке Аляски до арктических тундр на большей части северной Аляски. Исследование USGS дает количественную оценку того, как наземные птицы являются сигналами качества и изменений наземной среды обитания.
гагары
Все три вида гагар на Аляске встречаются в Национальном нефтяном заповеднике Аляски (NPR-A) на северном побережье Аляски.Исследования, проведенные Геологической службой США, дают информацию о распространении и численности гагар на севере Аляски и о том, как они могут реагировать на экологические и антропогенные изменения северного ландшафта.
Императорские гуси на берегу в Кадьяке.
(Источник: Брайан Угер-Кох, Геологическая служба США. Общественное достояние.)
Shorebirds
Многочисленные виды составляют группу птиц, известных как кулики. Эти птицы — международные мигранты, прибывающие на Аляску из Океании, Азии и Южной Америки.Исследование USGS дает количественную оценку миграционных коридоров и движущих сил демографических тенденций.
водоплавающие птицы
Утки, гуси и лебеди составляют группу птиц, известных как водоплавающие птицы, и являются важным ресурсом для занятий спортом и вылова натурального урожая на Аляске и по всей Северной Америке из-за их миграционных схем. Исследование USGS дает количественную оценку движущих сил популяционных тенденций в популяциях водоплавающих птиц, таких как болезни, качество среды обитания и изменение окружающей среды.
Наземные млекопитающие
Три карибу стоят в тундре.Эти карибу являются частью стада карибу дикобраза.
(Источник: Эндрю Рэми, Геологическая служба США. Общественное достояние.)
Понимание динамики популяции, взаимоотношений хищник / жертва и экологии среды обитания крупных наземных млекопитающих имеет решающее значение для управления этими видами диких животных на Аляске. Исследования USGS предоставляют актуальную научную информацию по вопросам управления дикой природой, таким как влияние изменения климата, управление спортом и натуральным урожаем, а также последствия болезней и развития человека.
Арктика
Экосистема Арктики быстро меняется, и многочисленные виды диких животных и их среды обитания реагируют на эти изменения. Исследование USGS количественно оценивает реакцию (положительную, отрицательную и стабильную) видов диких животных и их местообитаний на изменение экосистемы в Арктике и предоставляет информацию об этих ответах для принятия управленческих решений, связанных с освоением нефтегазовых ресурсов, спортом и натуральным урожаем, и популяционные тенденции популяций диких животных северных широт.
Аналитика экосистем
Группа Ecosystems Analytics использует новейшие статистические, картографические и графические методы для проведения исследований по различным таксонам. Наша общая цель — помочь информировать о решениях по управлению дикой природой и земельными ресурсами, помогая партнерам в разработке программ мониторинга, анализе существующих данных и разработке новых исследовательских проектов. Наше внимание часто включает моделирование динамики популяций диких животных в пространстве и времени и прогнозирование того, как популяции могут реагировать на экологические и антропогенные изменения в будущем.
Наземные экосистемы | USGS.gov
Наземные экосистемы
Северные Скалистые горы наделены обилием видов растений и животных, которые остались относительно нетронутыми со времен экспедиции Льюиса и Кларка в начале 19 века. Расширяющееся развитие энергетики, растущее давление на население и постоянный спрос на глобальные водные ресурсы делают управление дикой природой более сложным, чем когда-либо.NOROCK работает совместно с партнерами над изучением основных жизненных циклов и потребностей этих видов в среде обитания.
Обзор науки о земных экосистемах:
Дата публикации: 1 июля 2020 г.
Статус: АктивныйХроническая болезнь истощения
За последние 20 лет хроническая болезнь истощения (CWD) в Вайоминге медленно распространилась из юго-восточного угла штата в сторону Большого Йеллоустоуна и кормовых угодий для лосей Вайоминга, где каждую зиму дополнительно кормят более 24000 лосей.
Дата публикации: 1 июля 2020 г.
Статус: АктивныйКоличественная экология болезней
Исследователи USGS работают над разработкой новых количественных методов для изучения динамики болезней в системах дикой природы, а также в системах на стыке дикой природы, домашнего хозяйства и человека. Большая часть нашей работы сосредоточена на том, как структура популяции хозяев влияет на инвазию, устойчивость и контроль болезней в системах болезней диких животных.Мы решаем эти проблемы, комбинируя моделирование и статистику …
Дата публикации: 21 июля 2019 г.
Статус: АктивныйЗападные шмели и местные опылители
Западный шмель встречается на западе США и в Канаде и был одним из самых распространенных шмелей в этом регионе. Однако наши и другие исследования показывают, что он снижается, а индекс U.Служба S. Fish and Wildlife Service рассматривает его для включения в список в соответствии с Законом об исчезающих видах. Данные необходимы для улучшения нашего понимания …
Дата публикации: 6 марта 2018 г.
Статус: АктивныйАльпийская дикая природа и динамика снежного покрова на северных каскадах
Ожидается, что горные экосистемы будут меняться в связи с продолжающимся сокращением годового снежного покрова, которое наблюдалось во всем мире за последние полвека.Недавние снежные засухи в Северной Америке были связаны с необычно высокими температурами, из-за которых зимние осадки выпадают в виде дождя, а не снега. Многие виды альпийской дикой природы зависят от снежного покрова для защиты от сильного холода и …
Дата публикации: 13 декабря 2017 г.
Статус: АктивныйМежведомственная группа по изучению медведей гризли
Межведомственная группа по изучению медведей гризли (IGBST) — это междисциплинарная группа ученых и биологов, ответственных за долгосрочный мониторинг и исследования медведей гризли в Большой Йеллоустонской экосистеме (GYE).Команда была сформирована Министерством внутренних дел (DOI) в 1973 году в результате разногласий …
Дата публикации: 16 ноября 2017 г.
Статус: АктивныйВоздействие болезни на волков в Йеллоустонском национальном парке
В 1995 и 1996 годах волки были повторно завезены в Северные Скалистые горы, где они с тех пор прижились и распространились. В Йеллоустонском национальном парке, одном из основных защищенных участков высвобождения, неуправляемая популяция постоянно увеличивалась до высокой плотности, создавая большую популяцию волков, восприимчивых к таким инфекциям, как парвовирус собак (CPV), вирус чумы собак (CDV) и саркоптоз…
Дата публикации: 16 ноября 2017 г.
Статус: АктивныйПневмония у снежного барана
Популяции снежных баранов часто поражаются вспышками пневмонии, которые предположительно исходят от домашних овец и коз.
Дата публикации: 16 ноября 2017 г.
Статус: АктивныйБруцеллез
Бруцеллез — это заболевание домашнего скота, регулируемое на национальном и международном уровнях, имеющее серьезные последствия для здоровья животных, здоровья населения и международной торговли.
Дата публикации: 19 сентября 2017 г.
Статус: АктивныйПрограмма исследования крупных хищников NOROCK
NOROCK обладает значительным опытом в исследованиях крупных хищников, в первую очередь с участием видов, перечисленных как находящиеся под угрозой исчезновения. Программа исследования крупных хищников NOROCK включает ученых из штаб-квартиры NOROCK, полевой станции West Glacier и полевой станции Южных Аппалачей.Исследования проводятся в самых разных ландшафтах США, а также за рубежом …
Дата публикации: 16 марта 2017 г.
Статус: АктивныйРазвитие энергетики и воздушное пространство
Сегодня альтернативная энергетика и телекоммуникации развивают воздушное пространство во многом так же, как рост мегаполисов и механизированное сельское хозяйство развивают ландшафт.