
На каком пути эволюционного развития находятся бактерии в настоящее время: На каком пути эволюционного развития находятся бактерии в настоящее время?
Революция в утилизации отходов: бактерия, пожирающая пластик
- Мэри Халтон
- Отдел науки, Би-би-си
Для просмотра этого контента вам надо включить JavaScript или использовать другой браузер
Подпись к видео,Британские ученые открыли «пожиратель пластика»
Ученым удалось усовершенствовать существующий в природе фермент, который способен разлагать некоторые из наиболее распространённых полимеров, загрязняющих окружающую среду.
Наиболее распространенный пластический материал — PET, или полиэтилен, — который используется при производстве бутылок, в течение сотен лет остается неизменным на свалках. Он крайне медленно распадается под воздействием природных факторов.
Модифицированный фермент, получивший обозначение PETase, начинает разлагать этот полимер в течение нескольких дней. Это может привести к революции в деле утилизации пластмассовых отбросов.
Только в Британии в течение года приобретается около 13 миллиардов пластмассовых бутылок, из которых более 3 миллиардов никогда не утилизируются и оказываются на свалках.
Находка на свалке
Первоначально этот фермент был обнаружен в Японии. Он является продуктом жизнедеятельности бактерии Ideonella sakaiensis, которая пожирает полиэтилен PET в качестве основного источника энергии.
Автор фото, David Jones
Японские ученые сообщили в 2016 году, что они обнаружили разновидность этой бактерии на заводе по переработке пластиковых бутылок в портовом городе Сакаи.
«Полимер PET стал появляться в огромных количествах только за последние 50 лет, и это не слишком длительный срок для развития бактерий, которые способны поглощать этот искусственный материал», — говорит профессор Джон Макгиан из Портсмутского университета, который участвовал в исследовании.
PET (полиэтилентерефталат) принадлежит к группе сложных полиэфиров, встречающихся в естественных условиях.
«Они присутствуют в листьях растений, — отмечает профессор. — В течение миллионов лет развились бактерии, которые питаются такими полиэфирами».
Автор фото, David Jones
Подпись к фото,Полимеры класса PET обычно используются в производстве пластиковых бутылок
Однако обнаружение бактерии, способной перерабатывать именно полиэтилены класса PET, было неожиданностью для биохимиков. Была сформирована международная группа ученых, которые поставили целью определение природы и путей эволюции фермента PETase.
Бактерия пожирает пластик
Биохимики создали трехмерную компьютерную модель фермента, применив мощный рентгеновский лазер.
Разобравшись в молекулярной структуре этого фермента, ученые отметили, что эффективность действия PETase можно улучшить, внеся изменения в его поверхностную структуру.
Автор фото, H Lee Woodcock
Подпись к фото,Модель молекулы фермента PETase позволила усовершенствовать ее эффективность
Это указывает на то, что встречающийся в природе фермент не оптимизирован, и что существует возможность его улучшения.
Фермент PETase испытывался также на полимерах класса PEF, основанных на биоматериалах растительного происхождения, но тоже очень медленно распадающихся в естественных условиях.
«Нас поразило то, что этот фермент еще лучше воздействует на полимеры PEF, чем на полимеры PET», — заявил профессор Макгиан.
Анализ — Дэвид Шукман, отдел науки Би-би-си
В состав группы исследователей в Портсмутском университете входят аспиранты и даже студенты, и когда я побывал в их лаборатории, то не мог не разделить их энтузиазма. Они знают, что изобретение полимера класса PET потребовало больших усилий химиков, и гордятся тем, что им удалось найти способ его ускоренного разложения. Этот полимер используется при производстве миллиардов пластиковых бутылок во всем мире. Нынешнее поколение молодых химиков осознаёт проблему пластикового загрязнения и прилагает все усилия для ее решения.
Однако на пути трансформации этого открытия в практически применимую технологию будет немало препятствий. Во-первых, предстоит разработать способы недорогого производства такого фермента в промышленных масштабах; во-вторых, необходимо получить надежные методы его применения и контроля над его действием.
Во-первых, предстоит разработать способы недорогого производства такого фермента в промышленных масштабах; во-вторых, необходимо получить надежные методы его применения и контроля над его действием.
Утилизация замкнутого цикла
Сложные полиэфиры, получаемые при переработке нефти, широко используются при производстве пластиковых бутылок и одежды. Существующие методы их утилизации основаны на снижении их качества на каждом этапе переработки. Например, пластиковые бутылки сначала превращаются в волокно, используемое в производстве одежды, затем в производстве ковров, после чего они часто заканчивают свой путь на свалке.
Автор фото, Dennis Schroeder/NREL
Подпись к фото,Измененный фермент PETase в течение нескольких дней разлагает пластиковые отходы — изображение с электронного микроскопа
Фермент PETase обращает этот процесс вспять, превращая сложные полиэфиры в более простые молекулы, которые можно использовать заново.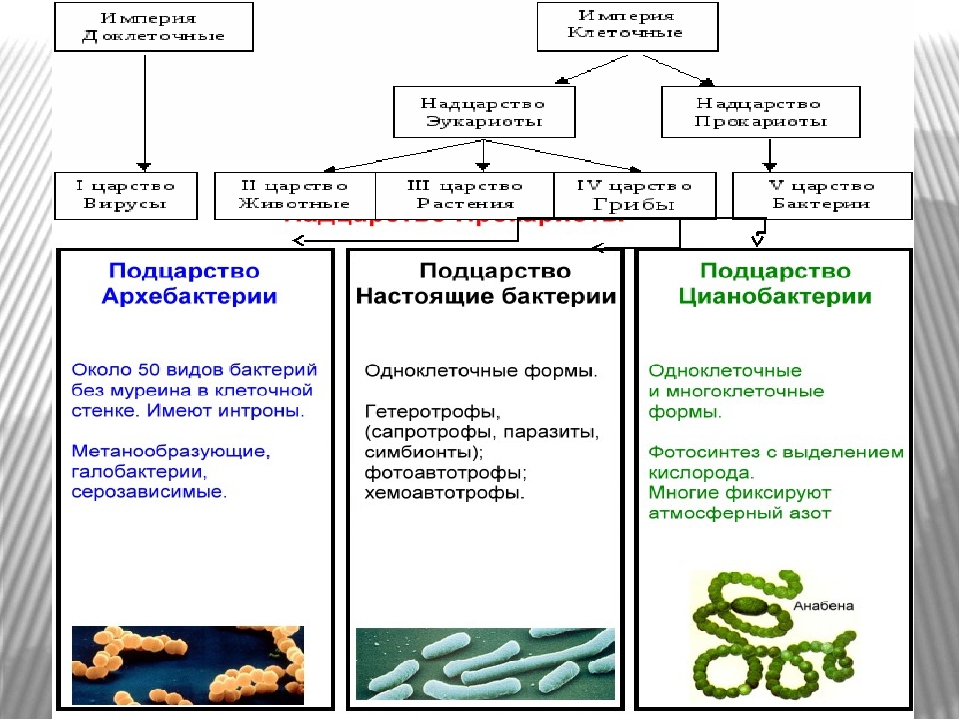
«Такие молекулы могут использоваться при производстве других полимеров, таким образом исключая из процесса нефть… В этом случае мы создаем замкнутый цикл производства и переработки, что необходимо при полной утилизации», — отмечает профессор Макгиан.
Этот фермент еще далек от промышленного использования. Необходимо ускорить его действие — в настоящее время он требует нескольких дней. В случае промышленного использования утилизация с его помощью должна занимать часы, а не дни.
Но профессор Макгиан надеется, что полученные результаты означают начало крупного сдвига в проблеме утилизации пластиковых отходов.
«В настоящее время остро ощущается потребность уменьшения объемов пластиковых отходов, которые заканчивают свой путь на свалках или попадают в окружающую среду, и если нам удастся применить новые методы, то мы получим решение этой проблемы в будущем», — говорит ученый.
Из опыта работы. Проверка знаний по теме «Прокариоты.
 Бактерии» в 10-м профильном классе
Бактерии» в 10-м профильном классеДвадцатипятилетний опыт работы учителем биологии привел меня к следующим выводам, которыми я руководствуюсь в своей повседневной работе.
- Только тщательно продуманный и спланированный урок, начиная с приветствия учителя и заканчивая тем, какое домашнее задание будет задано учащимся, позволит учителю достигнуть поставленных целей.
- Не давать ученикам все в готовом виде: прослушай, прочитай, запомни и отвечай. При таком учении мышление не развивается.
- Мало дать хорошую информацию, надо научить детей пользоваться полученными знаниями.
- Сколько времени учитель тратит на объяснение нового материала, столько же времени он должен потратить на освоение, закрепление и контроль знаний.
- Процесс накопления и закрепления знаний должен быть непрерывным. Закрепление нового материала должно проходить интенсивно, в сжатые сроки.
- Только творчество учителя порождает творчество учеников.

- Создавать на уроке положительную эмоциональную атмосферу. В такой атмосфере дети меньше устают.
Хочу поделиться опытом изучения темы “ Прокариоты. Бактерии.” в 10-м профильном классе.
Мои учащиеся обучаются в сельской школе, многие будут сдавать ЕГЭ по биологии. Поэтому свою работу я планирую с учетом этих особенностей.
Изучение темы “Прокариоты. Бактерии.” (да и других тем), я выстраиваю следующим образом:
– это урок теоретический. Лекция учителя, самостоятельная работа с учебником.Второй урок начинается с проверки знаний в игровой форме.
- Игра называется “Гонка за лидером”. Вызываются 2 ученика. Учитель
зачитывает подготовленные им короткие фразы, определения. Учащиеся должны их
закончить сразу через 2 секунды, не тратя время на раздумья.
 Вызываются 2 ученика. Учитель
зачитывает подготовленные им короткие фразы, определения. Учащиеся должны их
закончить сразу через 2 секунды, не тратя время на раздумья.
Вызываются 2 ученика. Учитель
зачитывает подготовленные им короткие фразы, определения. Учащиеся должны их
закончить сразу через 2 секунды, не тратя время на раздумья.- Самые древнейшие на земле организмы являются___________(доядерными).
- Доядерные организмы объединяются в царство____(Дробянки).
- Прокариоты не имеют оформленного____(ядра).
- Вместо ядра у прокариот в центре располагается___(кольцевая хромосома).
- Клеточная мембрана бактерий образует впячивания внутри клетки_____(мезосомы).
- Поверх плазматической мембраны у бактерий располагается оболочка из углевода____(муреина).
- В клетках прокариот встречаются следующие органоиды_____(только рибосомы).
- Бактерии, не использующие в энергетическом кислород называются____
- При неблагоприятных условиях бактерии образуют____(споры).
- Как переводится словосочетание “патогенные бактерии”_____(болезнетворные).

- Учащиеся всегда с удовольствием играют в эту игру. И хотя в ней участвуют только 2 уч-ся, остальные на местах проговаривают про себя ответы.
Учитель предлагает ученикам перейти к листам самоконтроля. Листы самоконтроля они получили от учителя на первом уроке после ознакомления с теоретическим материалом по прокариотам. Это было домашним заданием подготовиться отвечать по листам самоконтроля. Одно из главных требований к листам самоконтроля: ответы должны быть четкими, продуманными, аргументированными.
Назначаются два “консультанта” (из наиболее сильных учащихся, они помогут учителю “оценить” ответы одноклассников.
Лист самоконтроля.
- Какие организмы называют автотрофными, гетеротрофными, симбиотическими?
- В чем различие фототрофных и хемотрофных организмов по способу питания?
- Чем отличается питание сапротрофных организмов от паразитических?
- Какие организмы не имеют сформированного ядра в клетке?
- Как и насколько интенсивно размножаются бактерии?
- В чем заключается пастеризация и стерилизация как меря борьбы с бактериями? Что такое антибиотики?
- Чем отличается процесс аэробного дыхания от анаэробного?
- Какова роль гнилостных бактерий в природе и хозяйстве человека?
- При каких условиях бактерии гниения не наносят вреда?
- Для чего квасят и маринуют фрукты, овощи, готовят силос?
- Что такое симбиоз?
- Каким образом клубеньковые бактерии переводят азот воздуха в доступные для растений соединения?
- Учитель предлагает учащимся перейти к работе с обучающей картой.

Обучающая карта.
- Каково строение тела бактерии?
а) Многоклеточное; б) одноклеточное; в) колониальное. - Какую форму клетки имеет холерный вибрион?
а) шаровидную, б) палочковидную, в) извитую - Какие органеллы содержатся в клетках бактерий?
а) рибосомы; б) митохондрии; в) пластиды. - Какими по способу питания являются гнилостные бактерии?
а) автотрофными; б) симбиотическими; в) гетеротрофными. - Какие бактерии являются автотрофными?
а) паразитические; б) серобактерии; в) клубеньковые. - К какому типу размножения относится конъюгация у бактерий?
а) половое; б) бесполое; в) вегетативное. - Что образуют бактерии для перенесения неблагоприятных условий среды?
а) цисты; б) споры; в) клубеньки. - Чем по типу организации является бактериальная клетка?
а) прокариотической; б) зукариотической. - Что проводится для уничтожения патогенных бактерий?
а) консервирование; б) охлаждение; в) дезинфекция. - Какие бактерии используют кислород для дыхания?
а) анаэробные; б) аэробные.
Ответы к обучающей карте: 1- б, в; 2 – в; 3 – а; 4 – в; 5 – б; 6 – а; 7 – б; 8 – а; 9 – в; 10 – б.
“Консультанты” озвучивают правильные ответы к обучающей карте, ученики красными маркерами отмечают количество правильных ответов в своей обучающей карте.
- Последнее испытание для учащихся. Учитель предлагает им поработать с контрольными картами (тестовые задания более сложные).
Контрольная карта.
- Какое дыхание характерно для бактерий брожения?
а) кислородное; б) бескислородное. - Какие бактерии выполняют роль санитаров?
- Какие бактерии являются гетеротрофными?
а) нитрифицирующие; б) серобактерии; в) бактерии брожения; г) железобактерии. - Какие бактерии используются для жизнедеятельности энергию химических
связей неорганических веществ?
а) клубеньковые бактерии; б) паразитические; в) сапротрофные; г) хемотрофные; д) фототрофные. - Какие бактерии вызывают заболевания?
а) паразитические; б) бактерии гниения; в) бактерии молочнокислого брожения; г) патогенные.
а) бациллы; б) кокки; в) вибрионы; г) спирохеты.- Какие органеллы присутствуют в клетках бактерии?
а) митохондрии; б) центросомы; в) рибосомы; г) пластиды. - Отсутствие чего характерно для прокариотической клетки?
а) клеточной мембраны; б) обособленного ядра; в) комплекса Гольджи; г) клеточной стенки. - Что используют почвенные бактерии для передвижения?
а) реснички; б) жгутики; в) выделение слизи; г) газовые вакуоли. - На каком пути эволюционного развития находятся бактерии в настоящее
время?

Ответы к контрольной карте: 1) б; 2) б; 3) в; 4) г; 5) а, г; 6) б; 7) в; 8) б, в; 9) г; 10) а.
Учащиеся сами подсчитывают количество правильных ответов, выясняют не совсем понятные для них вопросы.
- Подведение итогов.
Выставление оценок.
Цель, поставленная учителем достигнута, весь класс активно работает, закрепляет полученные знания, умело использует их работе с тестами.
На следующем уроке учитель предложит познакомиться с контрольно-измерительными материалами ЕГЭ по изученной теме. Хорошее теоретическое усвоение темы позволит им успешно выполнить тестовые задания из методических пособий по ЕГЭ.
Учащиеся получают домашнее задание: выполнить тесты с выбором нескольких
правильных ответов.
- Генетический аппарат бактерий представлен:
а) белковыми молекулами;
б) кольцевой молекулой ДНК;
в) РНК;
г) липидными молекулами;
д) полисахаридами. - Нуклеоид – это:
а) мономер нуклеиновой кислоты;
б) генетический аппарат бактерий;
в) кольцевая молекула ДНК;
г) ДНК, связанная с белками-гистонами;
д) ДНК, не связанная с белками-гистонами. - Клетка бактерий имеет органоиды:
а) митохондрии;
б) комплекс Гольджи;
в) эндоплазматическая сеть;
г) рибосомы;
д) центросому. - Мезосомы бактерий выполняют функции:
а) органоидов движения;
б) митохондрий;
в) эндоплазматическая сеть;
г) комплекса Гольджи;
д) размножения. - Коньюгация бактерий – это:
а) половое размножение;
б) половой процесс;
в) обмен участками ДНК;
г) бесполое размножение;
д) множественное деление. - Болезнетворными являются бактерии:
а) холерные;
б) молочнокислые;
в) дифтерийные;
г) туберкулезные;
д) клубеньковые. - Способы борьбы с болезнетворными бактериями:
а) стерилизация хирургических инструментов;
б) дезинфекция помещений и одежды больных;
в) соблюдение гигиенических правил;
г) чистота продуктов питания;
д) контроль за качеством питьевой воды.
Правильные ответы: 1 – б; 2 – б, в, д; 3 – г; 4 – б, в, г; 5 – б, в; 6 – а, в, г; 7 – а, б, в, г, д.
Литература:
- Биология. Учебное пособие для учащихся средних школ, поступающие в высшие учебные заведения. 1995 г. Ижевск.
- Биология для абитуриентов. Р.Г. Заяц. И.В. Рачковская. Минск 2002 г.
- Биология. Пособие для поступающих в вузы. Москва. 1991 г.
- Общая биология. Готовимся к ЕГЭ. Дрофа. 2004 г.

Люди не смогут победить бактерий, с ними надо договариваться
Ежегодно Всемирная организация здравоохранения (ВОЗ) выбирает самую значимую для здоровья человечества проблему и обращается с международным призывом к совместным действиям, направленным на ее решение. Спустя 10 лет после публикации Глобальной стратегии ВОЗ по сдерживанию устойчивости к противомикробным препаратам эта проблема не только не решена, но стала настолько острой, что в 2011 году она объявлена темой Всемирного дня здоровья, а генеральный директор ВОЗ призвал мировое сообщество на борьбу с ее распространением.
В послании ВОЗ речь идет об одной большой группе микробов — надцарстве Bacteria. Сейчас охарактеризовано менее 9 тысяч видов, хотя всего их не меньше миллиарда. Надцарство — самый высокий ранг в систематике живых организмов. Если посмотреть на современное филогенетическое древо (оно показывает «родственные связи» разных видов, родов, семейств, классов), можно увидеть всего три главных ветви. Они делят все живое на надцарства — бактерии, археи (которых раньше причисляли к бактериям), и эукариоты (к нему принадлежат люди, животные, птицы, рыбы — те, у кого наследственная информация хранится в ядре клетки). Уже больше века ученые выясняют роль мира бактерий в жизни человека.
Они делят все живое на надцарства — бактерии, археи (которых раньше причисляли к бактериям), и эукариоты (к нему принадлежат люди, животные, птицы, рыбы — те, у кого наследственная информация хранится в ядре клетки). Уже больше века ученые выясняют роль мира бактерий в жизни человека.
«Родные» бактерии и «чувство кворума»
Сейчас известно, что жить без бактерий невозможно. Есть кефиры «с бифидо- и лактобактериями» и таблетки-пробиотики, которые нужно принимать после антибиотиков. Но также известно, что не бывает абсолютно безопасных бактерий. Они — всего лишь клетки с генетическим материалом, свернутым в кольцо (то есть в двойную спираль ДНК циркулярной формы), у которых немного целей в жизни: питаться и размножаться. А человек — всего лишь их инкубатор, поставляющий питательные вещества и обеспечивающий нормальную влажность и температурный режим. В ответ бактерии обеспечивают защиту от патогенов и токсинов, способствуют пищеварению и поддерживают в активном состоянии иммунитет. Если этот баланс смещается, какие-то представители «микро-биосферы», микробиоценоза, тоже могут изменить поведение и стать вирулентными, то есть вредными, несущими инфекции. Основные представители микрофлоры — это не широко разрекламированные бифидо- и лактобактерии, а грамотрицательные Bacteroides и грамположительные Firmicutes.
Если этот баланс смещается, какие-то представители «микро-биосферы», микробиоценоза, тоже могут изменить поведение и стать вирулентными, то есть вредными, несущими инфекции. Основные представители микрофлоры — это не широко разрекламированные бифидо- и лактобактерии, а грамотрицательные Bacteroides и грамположительные Firmicutes.
Каждое подразделение состоит из сотен видов бактерий и, что интересно, их набор уникален для каждого человека. Даже наличие разных видов бактерий и их количество строго индивидуально и, как было недавно показано, может быть связано с генотипом. Профессор Бухник из израильского Института технологий изучал различия в микробиоте мышей. Он обнаружил, что в желудках мышей одной из линий было намного больше бактерий штамма Lactobacillus johnsonii, причем все попытки заселить ими другую линию проваливались. Все потомки от скрещивания двух линий были способны нести в себе бактерии L.johnsonii. Эти данные говорят о том, что с изменением генотипа меняется и микробное сообщество в толстом кишечнике мышей. Вывод подтверждает генетик Эндрю Бенсон из университета Небраски. Проанализировав 645 «родственных» мышей, Бенсон с коллегами нашли 18 специфических участков генома (quantitative trait loci), которые были напрямую связаны с микробным составом мышиных желудков.
Вывод подтверждает генетик Эндрю Бенсон из университета Небраски. Проанализировав 645 «родственных» мышей, Бенсон с коллегами нашли 18 специфических участков генома (quantitative trait loci), которые были напрямую связаны с микробным составом мышиных желудков.
Интересно, что индивидуальность микрофлоры определяется еще и системой врожденного иммунитета человека, — ведь организм не должен реагировать на собственную микрофлору.
Внутри каждого человека для каждого присутствующего вида бактерии есть четко определенное место, функция и даже порядок колонизации (кто за кого «цепляется», и как быстро заселяет место в отсутствии соседа). Это сложная многоуровневая система, в которой необходимо поддерживать определенный баланс. Поэтому бактерии находятся в постоянном соревновании друг с другом, и такой антагонизм проявляется в выработке антимикробных веществ, подавляющих рост соседей. Для создания общего микромира, помимо противодействия, есть еще одно важное свойство системы — «чувство кворума» (quorum-sensing). На самом деле, это никакое не чувство, а огромное количество сигнальных молекул, которые синтезируют одни бактерии и химически чувствуют другие. Это одно-, двух-, и даже трехкомпонентные системы, причем каждая клетка может производить одновременно до нескольких десятков таких сигналов, сообщая о своем местоположении, взаимодействуя с соседями и организмом человека.
На самом деле, это никакое не чувство, а огромное количество сигнальных молекул, которые синтезируют одни бактерии и химически чувствуют другие. Это одно-, двух-, и даже трехкомпонентные системы, причем каждая клетка может производить одновременно до нескольких десятков таких сигналов, сообщая о своем местоположении, взаимодействуя с соседями и организмом человека.
Получается, что каждый человек с рождения окружен биологической «защитной пленкой», барьером из бактерий, которые находятся в таком тесном взаимодействии друг с другом и с иммунитетом хозяина, что просто не могут допустить к себе чужаков. От человека бактерии получают до 45 грамм углеводов из ежедневного рациона, но зато расщепляют полисахариды, липиды, а также до 30% неперевариваемых белков и 25% всей мочевины. Если баланс кворума сместится, изменятся концентрации специфических регуляторных пептидов, что может дать команду генетическому аппарату бактерий переключиться на вирулентный тип поведения, то есть стать инфекционными.
Поэтому когда врачи избавляются от «плохих» бактерий антибиотиками, они заодно убивают и своих, «родных», делают в естественном защитном барьере бреши, в которые беспрепятственно могут проникнуть патогены.
В настоящее время исследования пробиотиков, которые могут быть естественными врагами вирулентных бактерий, переживают подъем, но мы находимся только в начале пути к их повсеместному применению. До сих пор от «темной стороны» бактерий, их инфекционности, человечество спасали антибиотики, которые до недавнего времени, несмотря на то, что их действие сродни ковровым бомбардировкам, оставались эффективны.
Естественный отбор и устойчивость к антибиотикам
В общем случае действие антибиотика состоит в том, что он служит ингибитором, то есть тормозит или подавляет какую-то функцию, жизненно важную для микроба. Например, это может быть атака на аппарат белкового синтеза — и тогда бактерии теряют способность производить необходимые им белки. Некоторые антибиотики, тот же пенициллин, способствуют разрушению клеточной стенки бактерий.
Сейчас производят более эффективные антибиотики, нацеленные на старые мишени. Одновременно разрабатывают новые цели: нарушения в делении бактериальной клетки, биосинтез жирных кислот, ингибирование ферментов метаболизма. Но бактерии умудряются выживать даже в такой массированной атаке.
Подтверждены два пути того, как это может происходить. При первом, давно известном, в ответ на действие антибиотика быстро (хоть иногда и через несколько бактериальных поколений) появляются мутантные клетки, которые могут ему каким-либо образом противостоять. Например, у них идет более активный синтез питательных веществ, включаются «подстраховочные механизмы» белкового синтеза, или немного по-другому устроен белок, на который воздействует антибиотик. Может даже возникать спонтанная мутация, позволяющая им жить в среде, содержащей антибиотики. Соответственно, они выживают, растут, и постепенно заменяют собой всю популяцию. Для такой, новой, «усовершенствованной» популяции бактерий становятся не страшны даже высокие дозы антибиотика.
Второй путь, который подтвердили всего год назад, предполагает «благотворительность» со стороны бактериальных мутантов по отношению к своим соседям. В работе, опубликованной в Nature, описаны мутации, позволившие бактериям выживать под действием антибиотиков. Авторы-микробиологи нашли пять более или менее общих мутаций, которые позволяли активней выбрасывать антибиотик из клетки и синтезировать антиоксиданты. Но интересно другое — выживали и обычные, не мутировавшие бактерии. Оказалось, что антибиотик тормозил выработку индола — неспецифической защиты бактерий, в норме он синтезируется постоянно. Мутантные клетки быстро выбрасывали антибиотик, и, соответственно, могли синтезировать индол. Его количества было достаточно, чтобы защищать не только мутантов, но и их соседей.
Но не стоит забывать, что на выброс антибиотика из клетки затрачивается много «сил», и такие мутанты растут медленнее соседей. В итоге 1% приспособленных мутантов поддерживал 99% неприспособленных бактерий, которые, в свою очередь, обеспечивали высокие темпы роста всей колонии.
Процессы появления мутантов — не сложно спланированный механизм защиты бактерий от человеческого оружия, а всего лишь закономерное следствие теории эволюции. Один из ее важных принципов — существование различий между членами популяции, причем чем выше будет такая вариабельность, тем больше у вида (или, в данном случае, штамма) шансов на выживание в меняющихся условиях.
Люди сами создают вирулентные бактерии
Это верно и для бактерий, и для человека. Поэтому и происходит гонка вооружений между бактериями и фармацевтами. Ученые просто «выводят» новые виды опасных бактерий с помощью антибиотиков, а потом сами же с ними борются. «Помогают» им в этой селекции страны, в которых антибиотики дешевы, выписываются без рецепта, и, соответственно, у врача нет возможности проследить за выполнением всего цикла лечения. В итоге человек через пару дней приема антибиотиков чувствует себя значительно лучше, и забывает про них. А в это время в его организме остатки бактериальной популяции получают отличную возможность приспособиться к все понижающейся дозе антибиотика, и во время следующей болезни иметь целый арсенал защиты. Человек переходит на следующий антибиотик, потом на другой, но их не бесконечное количество. Вот о чем предупреждает ВОЗ — такими темпами через некоторое время человечество может оказаться в довоенной эре, полностью безоружным против им же созданных мутантных бактерий.
Человек переходит на следующий антибиотик, потом на другой, но их не бесконечное количество. Вот о чем предупреждает ВОЗ — такими темпами через некоторое время человечество может оказаться в довоенной эре, полностью безоружным против им же созданных мутантных бактерий.
Такая история уже произошла с туберкулезом, по поводу которого с 1944 года — первого применения стрептомицина, врачи были спокойны. Не так давно появились штаммы, устойчивые сразу к нескольким противотуберкулезным антибиотикам, так называемые MDR-формы (от англ. multidrug resistant). Сейчас MDR-штаммами туберкулезных бактерий инфицировано более 50 млн человек. «Горячие точки» — как раз страны с дешевыми легкодоступными антибиотиками: Восточная Европа, Россия, Китай. Но только от 5 до 10% инфицированных заболевает туберкулезом в течение жизни. Вероятно, это единственное, что спасает нас от эпидемии, и дает надежду найти оружие, которое использует иммунитет человека для защиты от микобактерии в тех самых 90-95% случаев.
Похожее развитие получил и золотистый стафилококк. Появились устойчивые к большинству антибиотиков штаммы — бактерии MRSA (methicillin resistant Staphylococcus aureus). Причем возникли они не где-нибудь, а в больницах и госпиталях. Микробиолог Марк Шпигельман из центра инфекционных болезней Лондонского University College считает, что избыточные меры гигиены уничтожают безвредных бактерий, оставляя нишу для MRSA, а также ведут к появлению устойчивых к антибиотикам штаммов и активизируют у бактерий защитные механизмы, которые обычно не работают. Естественно, что в операционной должно быть стерильно, а руки надо мыть с антибактериальным мылом. Однако последние данные заставляют всех, от мам маленьких детей до врачей, пересмотреть свои взгляды на гигиену.
К тому же, у бактерий есть свое «секретное оружие», кроме тех, к которым мы уже привыкли. У них есть горизонтальный перенос генетического материала, то есть патогенная бактерия может «поделиться» с безвредной своими «инфекционными» генами. Для этого есть много способов: фаги, плазмиды (короткие кольцевые ДНК), островки патогенности.
Для этого есть много способов: фаги, плазмиды (короткие кольцевые ДНК), островки патогенности.
Получается, что бактерия даже в своей «личной жизни» имеет большой шанс получить в геном патогенный участок, который позволит ей заражать человека. Например, благодаря горизонтальному переносу Bacillus cereus приобрел на каком-то этапе эволюции две плазмиды с генами патогенности, и стал возбудителем сибирской язвы. Аналогично возникла и чума. На настоящий момент внутри почти каждого рода есть несколько вредоносных штаммов, приводящим к тяжелым болезням. Какие есть новые подходы к борьбе с патогенными бактериями? И можно ли отказаться от использования антибиотиков в пользу новых безопасных лекарств?
Новые способы борьбы с патогенами
Накопленные знания про разнообразие форм бактерий позволяют считать, что схема «плохие — хорошие» бактерии устарела. Поэтому надо искать вещества, нацеленные только на вирулентные штаммы, а, еще лучше, на один конкретный штамм или даже вирулентный клон, имеющий тот же генетический материал, но с добавкой «островка патогенности». Например, если хорошо изучить механизм инфекционного процесса, можно с помощью специфических РНК блокировать синтез токсинов. Технологию создания таких искусственных РНК, нацеленных на контроль работы бактериальных генов, представила в этом году группа ученых из Китая под руководством профессора Венгонг Ю.
Например, если хорошо изучить механизм инфекционного процесса, можно с помощью специфических РНК блокировать синтез токсинов. Технологию создания таких искусственных РНК, нацеленных на контроль работы бактериальных генов, представила в этом году группа ученых из Китая под руководством профессора Венгонг Ю.
Другой вариант — влиять на сигналы «чувства кворума». Например, у стафилококков переключение на инвазивный тип существования, который приводит к инфекции, контролируется всего четырьмя генами. Они включаются и выключаются в ответ на концентрацию в среде специфических регуляторных пептидов. Поэтому, если ученые научатся использовать бактериоцины, продукция которых регулируется «чувством кворума», они смогут контролировать «вредность» некоторых штаммов бактерий, или даже избавляться от них. Например, бактериоцин низин уже сейчас успешно применяется в качестве консерванта в пищевой промышленности. Использование других биологических врагов бактерий — бактериофагов — затрудняется тем, что они могут вызывать аллергии, переносить патогенные участки ДНК/РНК между бактериями, к тому же, бактерии сами успешно с ними борются.
Сейчас в качестве вакцин будущего рассматривают введение в организм человека не ослабленных бактерий, а их специфических белковых последовательностей. Механика процесса поста — в кровь вводят антигены, и иммунная система человека делает всю «черную работу» — производит антитела, которые помогут ему бороться с настоящей, живой инфекцией. Но тут нужно выбирать, что использовать в качестве антигенов. Например, у основного белка клеточной стенки стрептококков и белка миокарда человека есть общие последовательности аминокислот, поэтому его нельзя использовать. Для борьбы с золотистым стафилококком используют в качестве мишени белок — токсин, который есть только у высоковирулентных штаммов. В итоге стафилококки-симбионты не повреждаются. Подход, который применяют в лаборатории Суворова, — использование смеси из нескольких антител с разными мишенями, — дает очень хорошие результаты. Видимо, в таком случае бактерия не может использовать запасные пути отступления (переключение пути метаболизма, выбрасывание антибиотиков, и т. д.) и вынуждена сдаться.
д.) и вынуждена сдаться.
Другой подход — не просто вакцинировать специфическими антигенами, но доставлять их с помощью безопасных, и даже полезных бактерий. Сейчас для этих целей часто используют штамм Lactococcus lactis. В работе Шуджи Лью с коллегами из Джиньянского университета (Китай) гены, контролирующие развитие фимбрий (отростков) «плохого» штамма Enterotoxigenic Escherichia coli искусственно вставляют в ДНК «хороших» лактококков. Лактококки сами по себе хорошо встраиваются в микро-сообщество человека, поэтому могут долго поддерживать специфический антигенный стимул. Кроме того, они сами стимулируют иммунитет. Аналогичная работа проводится в лаборатории профессора Суворова. В этом случае в качестве «корабля» используется широко распространенная кишечная палочка, а доставляет и синтезирует она гены, отвечающие за производство смеси поверхностных белков «плохих» штаммов стрептококков. В экспериментах на мышах такая система вызывала активный иммунный ответ на смертельную стрептококковую инфекцию. Это позволяет нам верить в перспективность именно такого способа борьбы с болезнетворными бактериями. С одним лишь условием — вакцина должна быть подобрана в соответствии с микробиотой конкретного человека и с учетом патогенных бактерий, циркулирующих в данном регионе.
Это позволяет нам верить в перспективность именно такого способа борьбы с болезнетворными бактериями. С одним лишь условием — вакцина должна быть подобрана в соответствии с микробиотой конкретного человека и с учетом патогенных бактерий, циркулирующих в данном регионе.
Со спора Пастера и Мечникова прошло уже больше века, и роль бактерий в жизни человека стала яснее. Теперь известно, что бактерии бывают необходимы и что их набор очень индивидуален, что они умеют приспосабливаться и что само человечество поневоле породило множество опасных штаммов. Но с развитием генетики и молекулярной биологии появляется надежда, что человечеству удастся договориться с микробным сообществом.
Мария Шутова
Страница не найдена |
Страница не найдена | 404. Страница не найдена
Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
3456789
10111213141516
17181920212223
24252627282930
31
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Фев
Мар
Апр
Май
Июн
Июл
Авг
Сен
Окт
Ноя
Дек
Метки
Настройки
для слабовидящих
Эволюционная биология развития (Evo-Devo): прошлое, настоящее и будущее | Эволюция: образование и пропаганда
Абухейф Э. , Рэй Г. Эволюция генной сети, лежащей в основе полифенизма крыльев у муравьев. Наука. 2002; 297: 249–52.
, Рэй Г. Эволюция генной сети, лежащей в основе полифенизма крыльев у муравьев. Наука. 2002; 297: 249–52.
CAS PubMed Google Scholar
Аллен Г. Науки о жизни в ХХ веке. Нью-Йорк: Уайли; 1975.
Google Scholar
Артур В.Теория эволюции развития. Чичестер: Уайли; 1988.
Google Scholar
Авероф М. Эволюция членистоногих: те же гены Hox , разные строения тела. Curr Biol. 1997; 7: R634–6.
CAS PubMed Google Scholar
Бальфур FM. Предварительный отчет о развитии пластиножаберных рыб. Q J Microsc Sci. 1874; 14: 323–64.
Google Scholar
Бейтсон В.Эволюционная вера и современные сомнения. Наука. 1922; 55: 53–61.
Google Scholar
Berrill NJ. Биология в действии: учебник для начинающих. Нью-Йорк: Додд, Мид и Ко .; 1966.
Биология в действии: учебник для начинающих. Нью-Йорк: Додд, Мид и Ко .; 1966.
Google Scholar
Бертосса RC. Морфология и поведение: функциональные звенья развития и эволюции. Филос Транс Соц Б. 2011; 366: 2056–68.
Google Scholar
Биемон К.От генотипа к фенотипу. Что нам говорят эпигенетика и эпигеномика? Наследственность. 2010; 105: 1–3.
PubMed Google Scholar
Bonner JT. Эволюция и развитие. Отчет Далемского семинара по эволюции и развитию, Берлин, 1981, 10–15 мая. Отчет об исследовании наук о жизни 22. Берлин: Springer; 1982.
Google Scholar
Bowler PJ. Прекрасная драма жизни.Эволюционная биология и реконструкция происхождения жизни 1860–1940 гг. Чикаго: Издательство Чикагского университета; 1996.
Google Scholar
Брыльский П, зал БК. Онтогенез макроэволюционного фенотипа: внешние защечные карманы геомиоидных грызунов. Эволюция. 1988a; 42: 391–5.
Онтогенез макроэволюционного фенотипа: внешние защечные карманы геомиоидных грызунов. Эволюция. 1988a; 42: 391–5.
Google Scholar
Брыльский П, зал БК. Эпителиальное поведение и пороговые эффекты в развитии внешних и внутренних защечных мешков у грызунов.Zool Syst Evolutionsforsch. 1988b; 26: 144–54.
Google Scholar
Калоу П. Жизненные циклы: эволюционный подход к физиологии воспроизводства, развития и старения. Лондон: Wiley; 1978.
Google Scholar
Калоу П. Принципы эволюции. Блэки: Глазго и Лондон; 1983.
Google Scholar
Кэрролл SB.Эво-дево и расширяющийся эволюционный синтез: генетическая теория морфологической эволюции. Клетка. 2008. 134: 25–36.
CAS PubMed Google Scholar
Carroll SB, Weatherbee SD, Langeland JA. Гомеотические гены, регуляция и эволюция числа крыльев насекомых. Природа. 1995; 375: 58–61.
Гомеотические гены, регуляция и эволюция числа крыльев насекомых. Природа. 1995; 375: 58–61.
CAS PubMed Google Scholar
Кэрролл С.Б., Гренье Дж.К., Уэтерби С.Д.От ДНК к разнообразию. Молекулярная генетика и эволюция дизайна животных. 2-е изд. Malden: Blackwell Science; 2005.
Google Scholar
Клементе Дж. К., Урселл Л. К., Палфри Л. В., Найт Б. Влияние микробиоты кишечника на здоровье человека: комплексный подход. Клетка. 2012; 148: 1258–70.
CAS PubMed Google Scholar
Collins JP, Gilbert S, Laubichler MD, Müller GB.Моделирование в EvoDevo: как интегрировать развитие, эволюцию и экологию. В: Лаубихлер MD, редактор. Корни теоретической биологии: 100-летие Prater Vivarium. Кембридж: MIT Press; 2007. с. 355–78.
Google Scholar
Даль Э. , Косеки Х., Баллинг Р. Гены Pax и органогенез. BioEssays. 1997; 19: 755–65.
, Косеки Х., Баллинг Р. Гены Pax и органогенез. BioEssays. 1997; 19: 755–65.
CAS PubMed Google Scholar
Дарвин CR.Происхождение видов путем естественного отбора. Лондон: Джон Мюррей; 1859.
Google Scholar
Дэвидсон Э. Регуляторный геном: регуляторные сети генов в развитии и эволюции. Сан-Диего: академический; 2006.
Google Scholar
Дэвидсон Э.Х., Эрвин Д.Х. Сети регуляции генов и эволюция строения тела животных. Наука. 2006; 311: 796–800.
CAS PubMed Google Scholar
de Beer GR. Эмбрионы и эволюция. Оксфорд: Кларендон; 1930.
Google Scholar
Дейчман У. Исследования начала 20 века на стыке генетики, развития и эволюции: размышления о прогрессе и тупиках. Dev Biol. 2011; 347: 3–12.
2011; 347: 3–12.
Google Scholar
Добжанский Т.Генетика и происхождение видов. Нью-Йорк: издательство Колумбийского университета; 1937.
Google Scholar
Эрвин DH, Wing SL. Глубокое время: взгляд палеобиологии. Палеобиология (Приложение). 2000. 26 (4): 1–371.
Google Scholar
Эрвин Д.Х., Лафламм М., Твидт С.М., Сперлинг Е.А., Пизани Д., Петерсон К.Дж. Кембрийская загадка: раннее расхождение и более поздний экологический успех в ранней истории животных.Наука. 2011; 334: 1091–7.
CAS PubMed Google Scholar
Фуско Г., Минелли А. Фенотипическая пластичность в развитии и эволюции: факты и концепции. Филос Транс Соц Б. 2010; 365: 547–56.
Google Scholar
Геринг WJ. Гомеобокс: ключ к пониманию развития? Клетка. 1985; 40: 3–5.
1985; 40: 3–5.
CAS PubMed Google Scholar
Геринг WJ.Управляйте генами управления в развитии и эволюции: история гомеобокса. Нью-Хейвен: издательство Йельского университета; 1998.
Google Scholar
Гилберт С.Ф. 2009 БИО. Evol Dev. 2009; 11: 331–2.
Google Scholar
Гилберт С.Ф., Апель Д. Экологическая биология развития: интеграция эпигенетики, медицины и эволюции. Сандерленд: Sinauer Associates; 2008.
Google Scholar
Гилберт С.Ф., Лоредо Г.А., Брукман А., Берк А.С.Морфогенез панциря черепахи: разработка новой структуры в эволюции четвероногих. Evol Dev. 2001; 3: 47–58.
CAS PubMed Google Scholar
Гиссис С.Б., Яблонька Э. Трансформации ламаркизма: от тонких флюидов к молекулярной биологии. Кембридж: MIT Press; 2011.
Кембридж: MIT Press; 2011.
Google Scholar
Глюкман П., Бидл А., Хэнсон М. Принципы эволюционной медицины.Оксфорд: издательство Оксфордского университета; 2009.
Google Scholar
Гольдшмидт Р. Предварительный отчет о некоторых генетических экспериментах, касающихся эволюции. Am Nat. 1918; 52: 28–50.
Google Scholar
Goodwin BC, Holder NJ, Wylie CC, редакторы. Развитие и эволюция. Кембридж: Издательство Кембриджского университета; 1983.
Google Scholar
Гулд С.Дж.Онтогенез и филогения. Кембридж: издательство Belknap Press Гарвардского университета; 1977.
Google Scholar
Грин Э. Полиморфизм развития гусеницы, вызванный диетой. Наука. 1989; 243: 643–66.
CAS PubMed Google Scholar
Грин Э. Фенотипическая изменчивость в развитии и эволюции личинок: полиморфизм, полифенизм и нормы реакции развития.В: Hall BK, Wake MH, редакторы. Происхождение и эволюция личиночной формы. Сан-Диего: академический; 1999. с. 379–410.
Фенотипическая изменчивость в развитии и эволюции личинок: полиморфизм, полифенизм и нормы реакции развития.В: Hall BK, Wake MH, редакторы. Происхождение и эволюция личиночной формы. Сан-Диего: академический; 1999. с. 379–410.
Google Scholar
Гренье Дж. К., Гарбер Т.Л., Уоррен Р., Уайтингтон П.М., Кэрролл С.Б. Эволюция всего набора генов членистоногих Hox предшествовала возникновению и радиации клады онихофоранов / членистоногих. Curr Biol. 1997; 7: 547–53.
CAS PubMed Google Scholar
Геккель Э.Generelle morphologie der organismen: Allgemeine grundzüge der organischen formen-wissenschaft, Mechanisch begin durch die von Charles Darwin, revision descendenz-theorie (2 тома). Берлин: Георг Реймер; 1866.
Google Scholar
Холл БК. Эволюционная биология развития. Лондон: Чепмен и Холл; 1992.
Google Scholar
Холл БК. Гомология: иерархическая основа сравнительной биологии.Сан-Диего: академический; 1994.
Google Scholar
Холл БК. Эволюционная биология развития. 2-е изд. Дордрехт: Kluwer Academic; 1999.
Google Scholar
Холл БК. Эво-дево или дево-эво — какое это имеет значение? Evol Dev. 2000; 2: 177–88.
CAS PubMed Google Scholar
Холл БК. Джон Сэмюэл Бюджетт (1872–1904): в погоне за Polypterus .Биология. 2001; 51: 399–407.
Google Scholar
Холл БК. Палеонтология и эволюционная биология развития: наука XIX и XXI веков. Палеонтология. 2002; 45: 647–69.
Google Scholar
Холл БК. Эволюция как контроль развития со стороны экологии. В: Холл Б.К., Пирсон Р., Мюллер Г.Б., редакторы. Окружающая среда, эволюция и развитие: к синтезу.Кембридж: MIT Press; 2003a. п. ix – xxiii.
В: Холл Б.К., Пирсон Р., Мюллер Г.Б., редакторы. Окружающая среда, эволюция и развитие: к синтезу.Кембридж: MIT Press; 2003a. п. ix – xxiii.
Google Scholar
Холл БК. Разблокировка черного ящика между генотипом и фенотипом: клетки и сгущения клеток как морфогенетические (модульные) единицы. Biol Philos. 2003b; 18: 219–27.
Google Scholar
Холл БК. Гомология, гомоплазия, новизна и поведение. Dev Psychobiol. 2012 (в печати).
Холл Б.К., Керни Р.Уровни биологической организации и происхождения новизны. J Exp Zool (Мол Дев Эвол). 2012; 314Б. DOI: 10.1002 / jez.b.21425.
Холл К., Олсон WM (ред.). Ключевые слова и понятия в эволюционной биологии развития . Кембридж, Массачусетс: Издательство Гарвардского университета; 2003.
Холл Б.К., Лаубихлер, доктор медицины, (ред.). Конрад Хэл Уоддингтон, теоретическая биология и evo – DevO. Биол. Теория. 2009. 3 (3): 185–289.
Биол. Теория. 2009. 3 (3): 185–289.
Халльгримссон Б., Холл Б.К., редакторы.Эпигенетика: связь генотипа и фенотипа в развитии и эволюции. Беркли: Калифорнийский университет Press; 2011.
Google Scholar
Hallgrímsson B, Hall BK (ред.). Вариация: центральное понятие в биологии . Нью-Йорк: Elsevier / Academic Press; 2005
Hoang T-H, McKay RI, Essam D, Hoai NX. О синергетическом взаимодействии между эволюцией, развитием и многоуровневым обучением. IEEE Trans Evol Comput.2011; 15: 287–311.
Google Scholar
Хаксли JS. Эволюция: современный синтез. Лондон: Аллен и Анвин; 1942.
Google Scholar
Косик К.С. МикроРНК рассказывают историю эволюции. Nat Rev Neurosci. 2009; 10: 754–9.
CAS PubMed Google Scholar
Laland KN, Odling-Smee J, Gilbert SF. EvoDevo и строительство ниш: наведение мостов. J Exp Zool (Мол Дев Эвол). 2008; 310B: 549–66.
EvoDevo и строительство ниш: наведение мостов. J Exp Zool (Мол Дев Эвол). 2008; 310B: 549–66.
Google Scholar
Лаубихлер MD. Эволюционная биология развития. В: Халл Д.Л., Русе М., ред. Кембриджский компаньон философии биологии. Кембридж: Издательство Кембриджского университета; 2007. с. 342–60.
Google Scholar
Лаубихлер MD. Эволюционная биология развития бросает серьезный вызов неодарвиновской парадигме.В: Аяла Ф.Дж., Арп Р., редакторы. Современные дискуссии по философии биологии. Молден: Вили-Блэквелл; 2010. с. 199–212.
Google Scholar
Laubichler MD, Maienschein J, редакторы. От эмбриологии до evo – DevO: история эволюции развития. Кембридж: MIT Press; 2007.
Google Scholar
Лаубихлер MD, Wagner GP. Введение в доклады симпозиума лауреата медали Ковалевского 2001 г. J Exp Zool (Мол Дев Эвол). 2004; 302B: 1–4.
J Exp Zool (Мол Дев Эвол). 2004; 302B: 1–4.
Google Scholar
Ledón-Rettig CC, Pfenning DW. Новые модельные системы в eco – evo – DevO: экологически чистая лопатоногая жаба. Evol Dev. 2011; 13: 391–400.
PubMed Google Scholar
Leliaert F, Verbruggen H, Zechman FW. В глубину: новые открытия в основе филогении зеленых растений. BioEssays.2011; 33: 683–92.
PubMed Google Scholar
Льюис Е.Б. Генный комплекс, контролирующий сегментацию Drosophila . Природа. 1978; 276: 565–70.
CAS PubMed Google Scholar
Ликлитер Р. Динамика развития и эволюции: выводы из поведенческой эмбриологии. Dev Psychobiol. 2007. 49: 749–57.
PubMed Google Scholar
MacDonald ME, Холл BK. Измененное время опосредованного внеклеточным матриксом эпителиально-мезенхимального взаимодействия, которое запускает скелетогенез нижней челюсти у трех инбредных линий мышей: развитие, гетерохрония и эволюционное изменение морфологии. J Exp Zool. 2001; 291: 258–73.
Измененное время опосредованного внеклеточным матриксом эпителиально-мезенхимального взаимодействия, которое запускает скелетогенез нижней челюсти у трех инбредных линий мышей: развитие, гетерохрония и эволюционное изменение морфологии. J Exp Zool. 2001; 291: 258–73.
CAS PubMed Google Scholar
Маклауд Р. Эмбриология и империя. В: Macleod R, Rehbock PF, редакторы. Лаборатория Дарвина. Эволюционная теория и естествознание в Тихом океане.Гонолулу: Гавайский университет Press; 1994. с. 140–65.
Google Scholar
Мейнард Смит Дж. Вейсманн и современная биология. В: Харви PH, Партридж Л., редакторы. Оксфордские обзоры по эволюционной биологии, т. 6. Оксфорд: Издательство Оксфордского университета; 1989. с. 1–12.
Google Scholar
Майр Э. Рост биологической мысли. Разнообразие, эволюция и наследование. Кембридж: издательство Belknap Press Гарвардского университета; 1982.
Кембридж: издательство Belknap Press Гарвардского университета; 1982.
Google Scholar
Майр Э. Становление эволюционной биологии как отдельной биологической дисциплины. BioEssays. 1997; 19: 263–6.
Google Scholar
McCain KW. Использование обмана для анализа цитируемого изображения: Конрад Хэл Уоддингтон и рост эволюционной биологии развития. J Am Soc Inf Sci Technol.2009; 60: 1301–19.
Google Scholar
McCain KW. Основная журнальная литература и постоянные темы исследований в развивающейся междисциплинарной области: изучение литературы по эволюционной биологии развития. J Informet. 2010; 4: 157–65.
Google Scholar
Моазед Д. Механизмы наследования состояний хроматина. Клетка. 2011; 146: 510–7.
CAS PubMed Google Scholar
Моларо А. , Ходжес Е., Фанг Ф, Сонг В., Маккомби В. Р., Хэннон Г. Дж..Профили метилирования сперматозоидов выявляют особенности эпигенетического наследования и эволюции у приматов. Клетка. 2011; 146: 1029–41.
, Ходжес Е., Фанг Ф, Сонг В., Маккомби В. Р., Хэннон Г. Дж..Профили метилирования сперматозоидов выявляют особенности эпигенетического наследования и эволюции у приматов. Клетка. 2011; 146: 1029–41.
CAS PubMed Google Scholar
Мюллер Ф. Фюр Дарвин. Лейпциг: Энгельман; 1864 (переведено У. С. Далласом как Фактов и аргументов для Дарвина , Лондон: Джон Мюррей, 1869).
Google Scholar
Müller GB. Шесть памяток для EvoDevo.В: Laubichler MD, Maienschein J, редакторы. От эмбриологии до evo – DevO: история эволюции развития. Кембридж: MIT Press; 2007a. п. 499–524.
Google Scholar
Müller GB. Evo – DevO », расширяющий эволюционный синтез. Nat Rev Genet. 2007b; 8: 943–9.
PubMed Google Scholar
Мюллер ГБ, Вагнер ГП. Новинка эволюции: перестройка концепции. Annu Rev Ecol Syst. 1991; 22: 229–56.
Annu Rev Ecol Syst. 1991; 22: 229–56.
Google Scholar
Nijhout HF. Гормональный контроль в развитии и эволюции личинок — насекомые. В: Hall BK, Wake MH, редакторы. Происхождение и эволюция личиночной формы. Сан-Диего: академический; 1999. с. 217–54.
Google Scholar
Никлас К.Дж. Эволюционная биология растений. Чикаго: Издательство Чикагского университета; 1997 г.
Google Scholar
Olsson L, Hoßfeld U, Breidbach O. Предисловие. Между Эрнстом Геккелем и гомеобоксом: роль биологии развития в эволюции. Теория Биоски. 2009; 128: 1–5.
PubMed Google Scholar
Петерсон К.Дж., Summons RE, Donoghue PCJ. Молекулярная палеобиология. Палеонтология. 2007; 50: 775–809.
Google Scholar
Петрусек А. , Толлиан Р., Швенк К., Хаас А., Лафорш К.«Терновый венец» — это индуцируемая защита, которая защищает Daphnia от древнего хищника. Proc Natl Acad Sci USA. 2009; 106: 2248–52.
, Толлиан Р., Швенк К., Хаас А., Лафорш К.«Терновый венец» — это индуцируемая защита, которая защищает Daphnia от древнего хищника. Proc Natl Acad Sci USA. 2009; 106: 2248–52.
CAS PubMed Google Scholar
Пфенниг DW. Полифенизм головастиков лопатоногих жаб как локально адаптированная эволюционно устойчивая стратегия. Эволюция. 1992; 46: 1408–20.
Google Scholar
Пиглюччи М., Мюллер ГБ, редакторы.Эволюция — расширенный синтез. Кембридж: MIT Press; 2010.
Google Scholar
Prum RO, Brush AH. Эволюционное происхождение разнообразия перьев. Q Rev Biol. 2002; 77: 261–95.
PubMed Google Scholar
Рафф РА. Прямые развивающиеся морские ежи и эволюционная перестройка раннего развития. BioEssays. 1992; 14: 211–8.
CAS PubMed Google Scholar
Рафф РА. Написано на камне: окаменелости, гены и evo – DevO. Nat Rev Genet. 2007; 8: 911–20.
Написано на камне: окаменелости, гены и evo – DevO. Nat Rev Genet. 2007; 8: 911–20.
CAS PubMed Google Scholar
Рафф Р.А., Лав AC. Ковалевский, сравнительная эволюционная эмбриология и интеллектуальное происхождение evo – DevO. J Exp Zool (Мол Дев Эвол). 2004; 302B: 19–34.
Google Scholar
Риппель О. Черепахи как обнадеживающие монстры. BioEssays. 2001; 23: 987–91.
CAS PubMed Google Scholar
Салазар-Сьюдад I. Ограничения развития против вариационных свойств: как формирование паттернов может помочь понять эволюцию и развитие. J Exp Zool (Мол Дев Эвол). 2006; 306B: 107–25.
Google Scholar
Сепкоски Д., Русе М. (ред.). Палеобиологическая революция: очерки развития современной палеонтологии.Чикаго: Издательство Чикагского университета; 2009.
Шубин Н., Табин С., Кэрролл С. Ископаемые останки, гены и эволюция конечностей животных. Природа. 1997. 388: 639–48.
CAS PubMed Google Scholar
Sommer RJ. Будущее evo – DevO: модельные системы и эволюционная теория. Nat Rev Genet. 2009; 10: 416–22.
CAS PubMed Google Scholar
Suga H, Tschopp P, Graziussi DF, Stierwald M, Schmid V, Gehring WJ.Гибкое использование генов Pax в развитии глаз на ранней стадии эволюции животных продемонстрировано исследованиями на гидрозойных медузах. Proc Natl Acad Sci USA. 2010; 107: 14263–8.
CAS PubMed Google Scholar
Томсон К.С. Морфогенез и эволюция. Оксфорд: издательство Оксфордского университета; 1988.
Google Scholar
Тиллс О, Рандл С.Д., Сэлинджер М. , Хаун Т., Пфеннингер М., Спайсер Дж.Генетическая основа внутривидовых различий в сроках развития? Evol Dev. 2011; 13: 542–8.
, Хаун Т., Пфеннингер М., Спайсер Дж.Генетическая основа внутривидовых различий в сроках развития? Evol Dev. 2011; 13: 542–8.
PubMed Google Scholar
Валентайн JW. О происхождении типов. Чикаго: Университет Чикаго Пресс; 2004.
Google Scholar
Waddington CH. Организаторы и гены. Кембридж: Издательство Кембриджского университета; 1940.
Google Scholar
Вагнера Г.П.Что обещает эволюция развития? Часть I. Почему биология развития необходима для объяснения эволюционных инноваций? J Exp Zool (Мол Дев Эвол). 2000. 288: 95–8.
CAS Google Scholar
Wagner GP, Larsson HCE. Что обещает эволюция развития? III. Тигель эволюционной эволюции. J Exp Zool (Мол Дев Эвол). 2003; 300B: 1–4.
Google Scholar
Wagner GP, Chiu C-H, Laubichler M.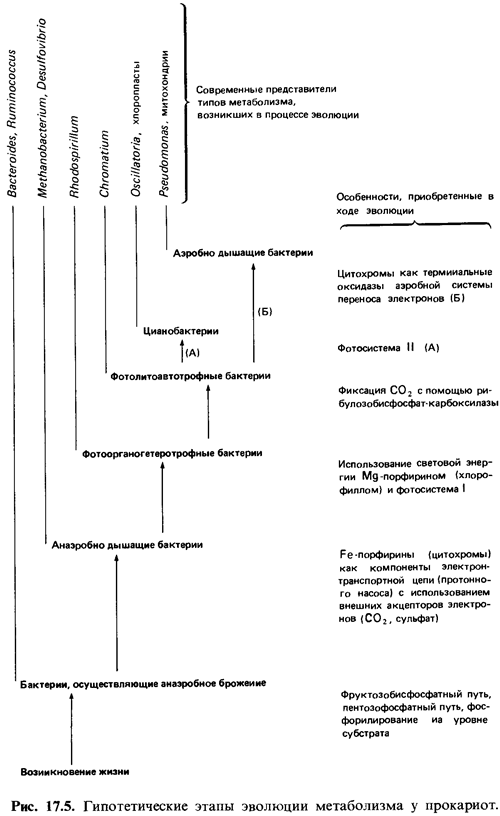 Эволюционная эволюция как механистическая наука: вывод от механизмов развития к эволюционным процессам. Am Zool. 2000; 40: 819–31.
Эволюционная эволюция как механистическая наука: вывод от механизмов развития к эволюционным процессам. Am Zool. 2000; 40: 819–31.
Google Scholar
Вагнер Г.П., Павлицев М., Чеверуд Ю.М. Путь к модульности. Nat Rev Genet. 2007; 8: 921–31.
CAS PubMed Google Scholar
Weiss KM. Томас Генри Хаксли (1825–1895) ставит нас на наше место.J Exp Zool (Мол Дев Эвол). 2004; 302B: 196–206.
Google Scholar
Вест-Эберхард MJ. Пластичность развития и эволюция. Оксфорд: издательство Оксфордского университета; 2003.
Google Scholar
Wilkins AS. Эволюция путей развития. Сандерленд: Sinauer Associates; 2002.
Google Scholar
Wilkins AS.Между «дизайном» и «бриколажем»: генетические сети, уровни отбора и адаптивная эволюция. Proc Natl Acad Sci USA. 2007. 104: 8590–6.
Proc Natl Acad Sci USA. 2007. 104: 8590–6.
CAS PubMed Google Scholar
Willmore KE. Развитие влияет на эволюцию. Am Sci. 2010; 98: 220–7.
Google Scholar
Xu X, You H, Du K, Han F. An Archeopteryx -подобный теропод из Китая и происхождение Avialae.Природа. 2011; 475: 465–70.
CAS PubMed Google Scholar
Зельдич М. За пределами гетерохронии: эволюция развития. Нью-Йорк: Уайли; 2001.
Google Scholar
Эволюция патогенов — эволюция микробов и совместная адаптация
ОБЗОР
Как заметил Ледерберг (2000), отношения хозяин-микроб представляют собой динамическое равновесие. Физиологические или генетические изменения у любого из партнеров могут побудить комменсальных микробов вторгнуться в ткань своего хозяина, тем самым вызывая иммунный ответ, который уничтожает захватчиков, но также может повредить или убить хозяина. Изучая этот процесс с точки зрения патогена и хозяина, докладчики семинара, представленные в этой главе, предложили различные возможные пути эволюции взаимоотношений хозяин-микроб, лежащих в основе инфекционных заболеваний.
Изучая этот процесс с точки зрения патогена и хозяина, докладчики семинара, представленные в этой главе, предложили различные возможные пути эволюции взаимоотношений хозяин-микроб, лежащих в основе инфекционных заболеваний.
Первая статья этой главы, написанная Стэнли Фалькоу из Стэнфордского университета, рассматривает природу бактериальной патогенности с точки зрения исторической точки зрения, выявленной его исследованиями и исследованиями его коллег из Стэнфордского университета. Он объясняет, как ключевые открытия, начиная с фундаментальной работы Ледерберга по бактериальной генетике, повлияли на развивающуюся область молекулярной биологии, а в частности, на почти 50-летние исследования Фалько генетической основы патогенности бактерий.
Используя инструменты молекулярной генетики для изучения Salmonella , Фалькоу и его коллеги наблюдали, как бактерии манипулируют функциями клетки-хозяина, как горизонтальный перенос генов формирует специализацию патогенов и как островки унаследованной патогенности превращают комменсальные бактерии в патогены. Проведя скрининг всего генома Salmonella на гены, которые связаны с разными стадиями заражения, с помощью стратегии отрицательной селекции на основе микрочипов, они идентифицировали многие гены патогенов, экспрессируемые в многоэтапном процессе инвазии хозяина.Используя мышиную модель, они также идентифицировали гены-хозяева и генные пути, экспрессируемые в ответ на инфекцию Salmonella .
Проведя скрининг всего генома Salmonella на гены, которые связаны с разными стадиями заражения, с помощью стратегии отрицательной селекции на основе микрочипов, они идентифицировали многие гены патогенов, экспрессируемые в многоэтапном процессе инвазии хозяина.Используя мышиную модель, они также идентифицировали гены-хозяева и генные пути, экспрессируемые в ответ на инфекцию Salmonella .
Фалькоу также учитывается важность микробов, которые он называет «комменсальными патогенами»: виды бактерий (например, Streptococcus pneumoniae , Neisseria meningitidis , Haemophilus influenzae типа b, Streptococcus pyogenes ), которые обычно населяют носоглотка человека протекает бессимптомно, но иногда вызывает заболевание.Их существование поднимает множество научных вопросов, касающихся взаимосвязи между микробной патогенностью, инфекционным заболеванием и иммунной функцией, — вопросы, к которым, по его мнению, следует подходить, изучая микробную патогенность как биологический феномен, а не только с точки зрения ее роли в вызывая болезнь.
Так же, как патогенность микробов — это больше, чем болезнь, инфекционное заболевание — это нечто большее, чем воздействие патогенов на клетки и системы хозяина.Вторая статья главы, написанная в соавторстве с Элисой Марголис и спикером семинара Брюсом Левином из Университета Эмори, рассматривает реакцию хозяина на микробную вирулентность, что, как отмечают авторы, не соответствует простым эволюционным моделям. Они исследуют, почему бактерии наносят вред (в основном людям) хозяевам, которые им необходимы для их выживания, предлагая доказательства того, что «большая часть вирулентности бактериальных инфекций может быть связана с кажущейся ошибочной реакцией иммунной защиты».
Эти иммунологические нарушения включают более энергичную реакцию, чем необходимо, как это происходит при бактериальном сепсисе; неправильная реакция на возбудителя, как при лепроматозной лепре; или реагирование на неправильные сигналы, как при синдроме токсического шока.Марголис и Левин исследуют эти и другие примеры «извращенности иммунной системы» и рассматривают эту точку зрения в свете различных текущих гипотез эволюции бактериальной вирулентности. Они предлагают возможные объяснения того, почему естественный отбор не смягчил чрезмерную реакцию иммунной системы на бактериальные инфекции, и обсуждают последствия своей точки зрения реакции хозяина на вирулентность для лечения бактериальных инфекций.
Они предлагают возможные объяснения того, почему естественный отбор не смягчил чрезмерную реакцию иммунной системы на бактериальные инфекции, и обсуждают последствия своей точки зрения реакции хозяина на вирулентность для лечения бактериальных инфекций.
Два дополнительных докладчика, Гордон Дуган и Джулиан Паркхилл из Института Wellcome Trust Sanger в Кембридже, Соединенное Королевство, внесли свой вклад в обсуждения на семинаре, касающиеся эволюции взаимоотношений хозяин-патоген.Каждый докладчик обсудил эволюционные пути, по которым серовары Salmonella стали разнообразными патогенами. К ним относятся Salmonella enterica серовар Typhimurium (далее S. typhimurium ), который инфицирует широкий круг хозяев и является основной причиной гастроэнтерита у людей, и S. enterica серовар Typhi (далее S. typhi ), специфический для человека агент системной инфекции брюшного тифа. У человека S.typhimurium обычно (но не всегда; см. ниже) локализуются в кишечном эпителии. S. typhi избегает разрушения иммунной системой и транспортируется через печень и селезенку в желчный пузырь и костный мозг, в которых бактерии могут сохраняться (рис. WO-9). Таким образом, значительное число людей, инфицированных брюшным тифом, в том числе бессимптомно инфицированных S. typhi , становятся хроническими носителями патогена и резервуарами заболевания, которое представляет значительную угрозу для здоровья населения.Однако с точки зрения S. typhi эта стратегия «скрытности» важна для его выживания.
ниже) локализуются в кишечном эпителии. S. typhi избегает разрушения иммунной системой и транспортируется через печень и селезенку в желчный пузырь и костный мозг, в которых бактерии могут сохраняться (рис. WO-9). Таким образом, значительное число людей, инфицированных брюшным тифом, в том числе бессимптомно инфицированных S. typhi , становятся хроническими носителями патогена и резервуарами заболевания, которое представляет значительную угрозу для здоровья населения.Однако с точки зрения S. typhi эта стратегия «скрытности» важна для его выживания.
Как и многие адаптированные к человеку патогены, такие как Yersinia pestis , Bacillus anthracis и Mycobacterium tuberculosis , S. typhi является монофилетическим; то есть, он ограничен с точки зрения геномной изменчивости, отметил Дуган. «Эти ограниченные человеком и недавно развившиеся патогены вошли в человеческую популяцию, как и многие другие патогены, не более 30–40 тысяч лет назад», и, таким образом, объяснил он, S. typhi эволюционировал одновременно с человеком и с аналогичной скоростью эволюции.
typhi эволюционировал одновременно с человеком и с аналогичной скоростью эволюции.
В своей презентации Паркхилл представил доказательства того, что помимо приобретения генов, которые придают инвазивность (острова патогенности, как описано Фалькоу), монофилетические патогены становятся вирулентными из-за потери функции генов, которые регулируют экспрессию факторов вирулентности (например, коклюшный токсин в Bordetella spp ., как подробно описано во вставке WO-2). Большая часть этих доказательств получена из определения идентичности нескольких различий между геномами монофилетических патогенов, выявленных с помощью геномики компаратора.
«Мы проводим компараторную геномику в надежде, что сравнение геномов расскажет нам что-то о сравнении фенотипов», — сказал Паркхилл. «Мы могли бы ожидать, что сможем пойти и поискать эти гены и найти [факторы вирулентности]», — продолжил он, но в случае Bordetella spp . Этого не произошло (вставка WO-2). Скорее, их сравнения показали, что Bordetella pertussis , основной возбудитель коклюша у людей, эволюционировал в сторону ограничения хозяина и большей вирулентности за счет потери функции генов, связанных с взаимодействием с хозяином (тем самым сужая диапазон хозяев), а также генов, регулирующих экспрессию факторов вирулентности, таких как токсин коклюша (Parkhill et al. , 2003).
, 2003).
Подобные события, по-видимому, повлияли на эволюцию различных патогенов человека, лошадей и растений, отметил Паркхилл. В случае S. typhi, большое количество псевдогенов (недавно инактивированных генов, на что указывает наличие точечных мутаций) имеют инактивированные белки клеточной поверхности и белки патогенности (McClelland et al., 2001). «Это признак организма, который сменил свою нишу», — сказал он. «Он перешел от фекально-орального патогена, который ограничен клетками, выстилающими кишечник, [] чтобы стать системным патогеном.Он потерял функцию. У него есть инактивированные гены, которые участвуют в патогенности, гены, которые были задействованы в его предыдущем образе жизни ».
«Почти наверняка некоторые из этих деактиваций являются выборочными», — продолжил он. «Они необходимы для этого изменения в нише, [например, для инактивации] секретируемых эффекторных генов типа III, которые, как мы знаем, важны для взаимодействия S. typhimurium с его хозяином. Мы видим, что известные нам гены участвуют в определении круга хозяев у S.typhimurium были инактивированы ». Однако, добавил он, «мы считаем, что многие из этих изменений, вероятно, являются сопутствующим ущербом. Происходит грандиозное событие, огромные изменения, которые организм не может контролировать ». Такие обстоятельства привели к возникновению узкого места в эволюции, во время которого огромное количество псевдогенов закрепилось по мере изменения диапазона хозяев и вирулентности патогена.
Мы видим, что известные нам гены участвуют в определении круга хозяев у S.typhimurium были инактивированы ». Однако, добавил он, «мы считаем, что многие из этих изменений, вероятно, являются сопутствующим ущербом. Происходит грандиозное событие, огромные изменения, которые организм не может контролировать ». Такие обстоятельства привели к возникновению узкого места в эволюции, во время которого огромное количество псевдогенов закрепилось по мере изменения диапазона хозяев и вирулентности патогена.
Сравнительное секвенирование S. typhi и второй независимой производной предка Salmonella enterica , S.enterica серовар Paratyphi A (далее S. paratyphi A), является дополнительным доказательством гипотезы Паркхилла (McClelland et al., 2004). Подобно S. typhi , S. paratyphi A стал системным патогеном, которым ограничено заражение людей. Каждый серовар содержит примерно 200 псевдогенов, но только около 30 из них являются общими для обоих. Эти общие псевдогены составляют «список генов, которые, по нашему мнению, были важными и которые, по нашему мнению, могут быть селективными для Salmonella , которая начинает становиться инвазивным патогеном, [таких как] секретируемые эффекторные белки, гены, участвующие в круге хозяев, и гены выделения, среди прочего другие, — заметил Паркхилл.Более того, он сказал: «Интересная особенность большинства этих распространенных псевдогенов в typhi и paratyphi A заключается в том, что они не имеют одинаковых инактивирующих мутаций. Они были приобретены самостоятельно. Это говорит о том, что они, вероятно, требуются выборочно ».
Эти общие псевдогены составляют «список генов, которые, по нашему мнению, были важными и которые, по нашему мнению, могут быть селективными для Salmonella , которая начинает становиться инвазивным патогеном, [таких как] секретируемые эффекторные белки, гены, участвующие в круге хозяев, и гены выделения, среди прочего другие, — заметил Паркхилл.Более того, он сказал: «Интересная особенность большинства этих распространенных псевдогенов в typhi и paratyphi A заключается в том, что они не имеют одинаковых инактивирующих мутаций. Они были приобретены самостоятельно. Это говорит о том, что они, вероятно, требуются выборочно ».
Те же самые эволюционные процессы привели к образованию менее вирулентных патогенов, сказал Паркхилл. Например, последовательность бактерии Streptococcus thermophilus , используемой для ферментации йогурта, обнаруживает свое происхождение от орального патогена человека, Streptococcus salivarius (Болотин и др., 2004). «Это предполагает — и это кажется вероятным — что люди начали ферментировать йогурт 10 000 лет назад в российских степях, плюясь в молоко, чтобы инициировать ферментацию», — пояснил он. «Через некоторое время они, вероятно, поняли, что это довольно отвратительно, или они нашли несколько действительно хороших сортов и размножили их, потому что из них получился хороший йогурт. По сути, то, что люди делали с йогуртом, — это микробиологический эксперимент, который длился 10 000 лет. Что произойдет, если вы возьмете патоген и приспособите его к новой нише — ферментирующему йогурту, которой раньше не было? Происходит то, что вы получаете огромное увеличение… псевдогенов, которые вырубили большинство генов, участвовавших в создании орального патогена.
«Это предполагает — и это кажется вероятным — что люди начали ферментировать йогурт 10 000 лет назад в российских степях, плюясь в молоко, чтобы инициировать ферментацию», — пояснил он. «Через некоторое время они, вероятно, поняли, что это довольно отвратительно, или они нашли несколько действительно хороших сортов и размножили их, потому что из них получился хороший йогурт. По сути, то, что люди делали с йогуртом, — это микробиологический эксперимент, который длился 10 000 лет. Что произойдет, если вы возьмете патоген и приспособите его к новой нише — ферментирующему йогурту, которой раньше не было? Происходит то, что вы получаете огромное увеличение… псевдогенов, которые вырубили большинство генов, участвовавших в создании орального патогена.
Таким образом, заключил он, присутствие многих псевдогенов в геноме организма свидетельствует о недавних и стремительных эволюционных изменениях, но не обязательно изменяет их в сторону патогенности. Псевдогены — это то, что остается в хромосоме организма, который быстро адаптировался к новой нише, заметил Паркхилл; потеря этих нефункциональных генов происходит гораздо медленнее. «Это говорит о том, что большая часть изменений, которые мы наблюдаем в этих организмах, на самом деле вызвана дрейфом», — сказал он. «Есть несколько выборочных изменений, но по большей части это случайный сдвиг.
«Это говорит о том, что большая часть изменений, которые мы наблюдаем в этих организмах, на самом деле вызвана дрейфом», — сказал он. «Есть несколько выборочных изменений, но по большей части это случайный сдвиг.
Обращаясь к недавним событиям в эволюции S. typhi , Дуган описал исследование секвенирования, которое он и его коллеги провели для сравнения 200 фрагментов генов примерно из 500 пар оснований, каждый из 105 глобально репрезентативных изолятов S. typhi . 1 У этого монофилетического патогена они идентифицировали только 88 однонуклеотидных полиморфизмов (SNP), которые включали по крайней мере 15 независимых мутаций одного и того же критического гена, кодирующего субъединицу ДНК-гиразы (Roumagnac et al., 2006). Эти мутации придают устойчивость к антибиотикам на основе фторхинолонов (налидиксовой кислоты), которые были введены в конце 1980-х годов для лечения мультиантибиотикорезистентных инфекций S. typhi .
Используя эту информацию, Дуган и его коллеги построили филогенетическое дерево S. typhi . На основании содержания SNP «любой новый изолят можно однозначно отнести к дереву», — сказал Дуган. Более того, «SNP на самом деле ассоциируются с разными типами мутаций в разных частях основной цепи белка, что приводит к появлению разных устойчивых к налидиксовой кислоте клонов.Таким образом, дерево можно использовать для различения изолятов, а также «стратифицировать приобретение вплоть до точечной мутации маркера устойчивости к лекарствам».
typhi . На основании содержания SNP «любой новый изолят можно однозначно отнести к дереву», — сказал Дуган. Более того, «SNP на самом деле ассоциируются с разными типами мутаций в разных частях основной цепи белка, что приводит к появлению разных устойчивых к налидиксовой кислоте клонов.Таким образом, дерево можно использовать для различения изолятов, а также «стратифицировать приобретение вплоть до точечной мутации маркера устойчивости к лекарствам».
Дуган предсказал, что этот метод, который он назвал сигнатурным типированием на основе ДНК, положит начало «новой эре» полевых и клинических исследований патогенеза микробов. По его словам, исследователи смогут связать фенотипы с конкретными маркерами SNP, присутствующими в бактериях, выделенных от пациентов; приложения могут включать усилия по выявлению генетической основы повышенной передачи или вирулентности в новых штаммах патогенов, выявлению носителей инфекционных заболеваний и проведению исследований эффективности вакцин для конкретных типов.
В дополнение к SNP, Дуган отметил еще один путь к устойчивости к антибиотикам, который, по-видимому, недавно был принят нетифоидными сероварами Salmonella , включая S. typhimurium . Эти инвазивные инфекции, вызываемые патогенами, которые обычно вызывают гастроэнтерит, стали основной причиной заболеваемости и смертности африканских детей (Gordon et al., 2008; Graham, 2002). «Большинство детей и людей в Африке, которые умирали от сальмонеллеза, инвазивной болезни, не умирали от S.typhi ; на самом деле они умирали от штаммов, которые обычно вызывают гастроэнтерит, таких как S. typhimurium и enteritidis », — заметил Дуган. Последовательности штаммов, вызывающих нетифоидный сальмонеллез (NTS), оказались генетически отличными от штаммов Salmonella (тех же сероваров), которые вызывают гастроэнтерит в западных популяциях: они несли плазмиды, содержащие два различных генетических элемента, которые придали устойчивость к нескольким антибиотикам, а также до четвертичного аммония (дезинфицирующее средство; Graham et al. , 2000). «Он почти создан природой, чтобы стать идеальным решением для попытки человека лечиться антибиотиками», — сказал Дуган, а также антибиотиками, такими как хлорамфеникол.
, 2000). «Он почти создан природой, чтобы стать идеальным решением для попытки человека лечиться антибиотиками», — сказал Дуган, а также антибиотиками, такими как хлорамфеникол.
Дуган предупредил, что эти гены устойчивости могут быстро распространяться путем горизонтального переноса на другие штаммы Salmonella после запланированного внедрения широкомасштабной антибиотикопрофилактики (триметоприм-сульфонамид) для африканских детей, инфицированных вирусом иммунодефицита человека (ВИЧ). «Мы говорили о взаимосвязи между комменсалами и патогенами: они не знают границ», — заметил он.«Я не могу думать, что пройдет очень много времени [после внедрения широкомасштабной антибиотикопрофилактики], прежде чем мы действительно начнем движение этого потенциального переносчика вокруг населения Африки. Меня это очень тревожит, и я думаю, что нам нужно немного подумать о том, как мы это делаем ».
Между тем, пытаясь понять генетическое происхождение NTS, Дуган и его коллеги обнаружили, что антитела защищают от болезни, которая непропорционально поражает детей в возрасте от четырех месяцев (до которого они защищены материнскими антителами) и до двух лет. (после чего их собственная иммунная система вырабатывает эффективную защиту от патогена; MacLennan et al., 2008). Это открытие предполагает, что вакцины против NTS могут быть эффективными в индукции защитных антител в уязвимой возрастной группе.
(после чего их собственная иммунная система вырабатывает эффективную защиту от патогена; MacLennan et al., 2008). Это открытие предполагает, что вакцины против NTS могут быть эффективными в индукции защитных антител в уязвимой возрастной группе.
БАКТЕРИАЛЬНАЯ ПАТОГЕННОСТЬ: ИСТОРИЧЕСКАЯ И ЭКСПЕРИМЕНТАЛЬНАЯ ПЕРСПЕКТИВА
Стэнли Фалькоу, Ph.D. 2
Стэнфордский университет
Джошуа Ледерберг отметил в своем эссе 1987 года, что «важность бактерий как возбудителей инфекционных заболеваний была четко установлена к 1876 году, но это не вызвало особого интереса к их фундаментальной биологии примерно до шестидесяти пяти лет спустя. »(Ледерберг, 1987).Его, как и меня, учили, что бактерии — это Schizomycetes — примитивные бесполые растения, поэтому трудно было думать о них как о патогенных по своей природе. Сальвадор Лурия сказал о тех временах, что микробиология была последним оплотом ламаркизма.
Ледерберг, будучи студентом, находился под влиянием нескольких важных открытий в середине 1940-х годов, которые проложили путь для его последующей работы по конъюгации бактерий, включая демонстрацию механизма бактериальной трансформации Эйвери, Маклаудом и Маккарти ( 1944) и бактериального мутагенеза и отбора Луриа и Дельбрюк (1943). Открытие Ледербергом бактериальной конъюгации позволило исследователям впервые изучить микробную генетику и биохимию. Это была мечта молодого Ледерберга; он вспомнил, что стер страницы книги по физиологической химии, которую он получил за свою бар-мицву. Джош также с самого начала осознавал, что разрабатываемые им методы могут иметь практическое применение для улучшения вакцин, а также для достижения «понимания вирулентности, современного расширения примитивных методов Пастера» (Lederberg, 1987).
Открытие Ледербергом бактериальной конъюгации позволило исследователям впервые изучить микробную генетику и биохимию. Это была мечта молодого Ледерберга; он вспомнил, что стер страницы книги по физиологической химии, которую он получил за свою бар-мицву. Джош также с самого начала осознавал, что разрабатываемые им методы могут иметь практическое применение для улучшения вакцин, а также для достижения «понимания вирулентности, современного расширения примитивных методов Пастера» (Lederberg, 1987).
Ледерберг со своим учеником Нортоном Зиндером и сотрудником Брюсом Стокером в начале 1950-х годов обнаружили, что любой фрагмент бактериальной ДНК может быть включен в геном бактериофага (Stocker et al., 1953). Из этого он понял, что рекомбинация генов, называемая генерализованной трансдукцией, вероятно, имеет место в природе, поскольку было показано, что фаги являются основой для нескольких различных видов известных серотипов Salmonella .
Фундаментальные исследования Ледерберга в области генетики бактерий явились важным фактором для открытий в последующие годы информационной РНК, генетического кода и работы Джейкоба и Моно (1961) по регуляции генов. Эта революционная работа стала основой современной молекулярной биологии, а также заложила основу для современных исследований патогенности бактерий. Поскольку меня попросили рассказать о патогенности бактерий как с исторической, так и с экспериментальной точки зрения, возможно, я буду прощен за использование некоторых моих собственных работ, а также некоторых работ моих коллег из Стэнфорда для обсуждения этой темы.
Эта революционная работа стала основой современной молекулярной биологии, а также заложила основу для современных исследований патогенности бактерий. Поскольку меня попросили рассказать о патогенности бактерий как с исторической, так и с экспериментальной точки зрения, возможно, я буду прощен за использование некоторых моих собственных работ, а также некоторых работ моих коллег из Стэнфорда для обсуждения этой темы.
Я начал свою работу над генетической основой патогенности в 1959 году, работая над тифоидной палочкой вместе с Луи С.Барон из Исследовательского института армии Уолтера Рида. Барон работал в лаборатории Ледерберга (Лу однажды сказал мне, что Ледерберг утверждал, что, если в эксперименте было больше шести планшетов и четырех пипеток, он был чрезмерно спроектирован). Моя цель на тот момент может быть просто сформулирована: я хотел узнать генетические различия между Salmonella и непатогенными обитателями кишечника, такими как Escherichia coli .
Я работал с медицинскими микробиологами, которые думали, как и многие до сих пор, что патоген — это любой организм, вызывающий болезнь. В то время микробиологи характеризовали патогенные бактерии как выродившиеся формы, которые заблудились и просто выросли за счет хозяина, вызывая тем самым ущерб (болезнь). Я подумал, как я сказал на семинаре в Колд-Спринг-Харбор в 1964 году, что патогены развили уникальные генетические черты, которые сделали их такими (на что очень известный ученый из аудитории ответил: «Фалькоу, никто не говорит как –– т. о брюшном тифе или патогенных микроорганизмах. Почему бы вам не заняться чем-то важным? »). Увы, в этой критике был смысл.Хотя я пытался показать, что существуют уникальные гены патогенности, мне не хватало необходимых генетических и молекулярных экспериментальных инструментов, чтобы доказать это.
В то время микробиологи характеризовали патогенные бактерии как выродившиеся формы, которые заблудились и просто выросли за счет хозяина, вызывая тем самым ущерб (болезнь). Я подумал, как я сказал на семинаре в Колд-Спринг-Харбор в 1964 году, что патогены развили уникальные генетические черты, которые сделали их такими (на что очень известный ученый из аудитории ответил: «Фалькоу, никто не говорит как –– т. о брюшном тифе или патогенных микроорганизмах. Почему бы вам не заняться чем-то важным? »). Увы, в этой критике был смысл.Хотя я пытался показать, что существуют уникальные гены патогенности, мне не хватало необходимых генетических и молекулярных экспериментальных инструментов, чтобы доказать это.
Вместо этого я обратился к изучению эписом, которые позже стали называть плазмидами (термин, придуманный Ледербергом). Джош и Эстер Ледерберг открыли первую плазмиду, фактор F, который оказался трансмиссивным генетическим элементом, определяющим фертильность E. coli K-12. Штаммы, несущие F, могут передавать свои гены другим бактериям. Работы Уильяма Хейса, а затем Джейкоба, Монода, Уоллмана и других впоследствии усовершенствовали биологию бактериальной фертильности. Вскоре появились сообщения о других примерах плазмид, включая передаваемую устойчивость к ряду антибиотиков. Инфекционная множественная лекарственная устойчивость была описана в западном мире примерно в 1960 году Цутому Ватанабэ и подтверждена работами Наоми Датта в Англии и Дэвида Смита и других в Соединенных Штатах. Эти R-плазмиды, как их называли, в середине 1960-х стали центром внимания большого числа ученых.Работа другого ученого по плазмидам — ветеринара по имени Х. Уильямс Смит — не получила должной оценки. Он продемонстрировал, что некоторые плазмиды могут передавать бактериальные токсины, адгезины и, в некоторой степени, специфичность к хозяину от одной бактериальной клетки к другой (Smith and Halls, 1967). Смит использовал классический подход Ледерберга, используя только пипетки, чашки Петри и простые генетические скрещивания, чтобы сделать эти важные открытия.
Работы Уильяма Хейса, а затем Джейкоба, Монода, Уоллмана и других впоследствии усовершенствовали биологию бактериальной фертильности. Вскоре появились сообщения о других примерах плазмид, включая передаваемую устойчивость к ряду антибиотиков. Инфекционная множественная лекарственная устойчивость была описана в западном мире примерно в 1960 году Цутому Ватанабэ и подтверждена работами Наоми Датта в Англии и Дэвида Смита и других в Соединенных Штатах. Эти R-плазмиды, как их называли, в середине 1960-х стали центром внимания большого числа ученых.Работа другого ученого по плазмидам — ветеринара по имени Х. Уильямс Смит — не получила должной оценки. Он продемонстрировал, что некоторые плазмиды могут передавать бактериальные токсины, адгезины и, в некоторой степени, специфичность к хозяину от одной бактериальной клетки к другой (Smith and Halls, 1967). Смит использовал классический подход Ледерберга, используя только пипетки, чашки Петри и простые генетические скрещивания, чтобы сделать эти важные открытия.
В 1972 году Стэнли Н. Коэн и Герберт Бойер обнаружили, что гены могут быть разрезаны и сплайсированы с использованием плазмид R и их производных, тем самым сигнализируя об открытии клонирования генов.Эта элегантная новая техника, а также развитие секвенирования ДНК позволили окончательно изучить гены патогенности.
Новое определение патогенности бактерий с помощью инструментов молекулярной генетики
Среди вещей, которые мы узнали о патогенности бактерий, есть следующие основные характеристики:
- Патогены — впечатляющие клеточные биологи. Накопленные за двадцать пять лет данные демонстрируют, что бактерии манипулируют нормальными функциями клетки-хозяина способами, которые приносят пользу бактериям ().
Горизонтальный перенос генов через мобильные генетические элементы стал чрезвычайно важным фактором в эволюции специализации бактерий, в том числе патогенов. Гены многих специализированных «бактериальных» продуктов, таких как токсины и адгезины, на самом деле находятся на транспозонах и фагах.
Наследование блоков генов, называемых островками патогенности, часто является ключом к выражению патогенности у бактерий.

РИСУНОК 3-1
Патогенные бактерии вмешиваются или манипулируют в своих интересах нормальной функцией (ями) клетки-хозяина.ИСТОЧНИК: Рисунок перепечатан из Wilson et al. (2002) с разрешения Cambridge University Press. Copyright 2002.
Когда мы начали использовать инструменты молекулярной генетики для изучения генов вирулентности и определения их функций, мы сначала попытались выделить определенные гены на токсины и другие вероятные продукты вирулентности. Сегодня мы используем совершенно другой подход, который отражает наше понимание поведения патогенов. Например, в отличие от комменсальных бактерий, Salmonella нарушает эпителиальный барьер хозяина, обычно в областях кишечного эпителия, известных как пятна Пейера, и поглощается фагоцитарными клетками.Вместо того, чтобы погибнуть, патоген размножается там, а затем распространяется в печень и селезенку. В конце концов, во многих случаях патоген будет распространяться хозяином, часто в течение длительного периода времени. Эти ключевые события в патогенезе инфекции, в дополнение к взаимодействию патогена с врожденной и адаптивной иммунной системой хозяина, проиллюстрированы на.
В конце концов, во многих случаях патоген будет распространяться хозяином, часто в течение длительного периода времени. Эти ключевые события в патогенезе инфекции, в дополнение к взаимодействию патогена с врожденной и адаптивной иммунной системой хозяина, проиллюстрированы на.
РИСУНОК 3-2
Инфекция Salmonella . IFN-γ = гамма-интерферон; MLN = брыжеечный лимфатический узел.ИСТОЧНИК: Monack et al. (2004b).
Различие между патогеном Salmonella и предковыми комменсальными организмами в кишечнике основано на наследовании островков патогенности, которые дают этим бактериям способность покидать пределы толстой кишки в места, где другие бактерии могут быть убиты. Здесь развивающийся патоген может действовать без конкуренции. Чтобы идентифицировать гены, которые делают возможными такие вторжения в хозяина, мы используем стратегию отрицательной селекции на основе микрочипов, как показано на рис. al., 2005).
РИСУНОК 3-3
Стратегия отрицательного отбора на основе микрочипов. hyb = гибридизация; IP = внутрибрюшинно. ИСТОЧНИК: Рисунок любезно предоставлен Каманом Чаном, доктором философии, Стэнфордский университет.
hyb = гибридизация; IP = внутрибрюшинно. ИСТОЧНИК: Рисунок любезно предоставлен Каманом Чаном, доктором философии, Стэнфордский университет.
Используя эту стратегию, можно обнаружить, что, хотя многие гены экспрессируются в течение первой недели болезни, существует группа генов, которые не экспрессируются до второй недели болезни, и даже другие, которые не экспрессируются до третьей или третьей недели болезни. четвертая неделя болезни (; Lawley et al., 2006).Эти результаты показывают, что определенные гены необходимы для различных стадий стойкой инфекции у мыши. Некоторые из генов участвуют в способности Salmonella выделять белки, убивающие макрофаги во время начальной инфекции, в то время как другие гены эволюционировали, чтобы позволить бактериям реплицироваться и сохраняться в вакуолях макрофагов.
РИСУНОК 3-4
Зависящий от времени отбор генов персистентности. Желтая рамка указывает на отсутствие гена персистентности.Синий или черный прямоугольник указывает на наличие гена персистентности. КОЕ = колониеобразующие единицы; ЛПС = липополисахарид; PMN = полиморфно-ядерный (подробнее …)
КОЕ = колониеобразующие единицы; ЛПС = липополисахарид; PMN = полиморфно-ядерный (подробнее …)
Таким образом, требуется много генов, чтобы способствовать способности Salmonella совершать свой путь от ротовой полости до пейеровского пятна в эпителии тонкой кишки и оттуда. в макрофаг, который распространяется на другие сайты. Благодаря доступной нам технологии мы можем легко идентифицировать все такие гены, но пройдет много лет, прежде чем мы сможем определить их точную биологическую функцию во взаимодействиях Salmonella -хост.
Хозяин как репортер ответа на инфекцию
Хозяин также может сказать нам, что происходит во время острых и стойких бактериальных инфекций. Когда Люсинда Томпсон, Дениз Монак и я исследовали экспрессию генов в периферической крови инфицированных мышей после заражения Salmonella , мы обнаружили, что 344 гена были индуцированы во время острой инфекции Salmonella у наивных мышей (). Эти гены связаны с передачей сигналов интерферона, инфильтрацией нейтрофилов и экстравазацией лейкоцитов.
РИСУНОК 3-5
Ответ на инфекцию Salmonella . Столбцы под синим прямоугольником соответствуют отдельным наивным животным до испытания. Каждый столбец под розовой рамкой соответствует активации гена у одного животного после заражения диким типом (дни 4–9 (подробнее …)
Мы изучили пути конкретных генов, которые были активированы или подавлены в ответ на Сальмонелла , и мы сравнили эти образцы ответа хозяина с теми, которые имели место у мышей, которые ранее были иммунизированы известным вакцинным штаммом Salmonella typhimurium , aroA .Подобные гены были индуцированы на ранних стадиях инфицирования у иммунизированных мышей, как и у наивных мышей, и поэтому составляют «сигнатуру иммунитета» (). Персистентно инфицированные мыши (через 10 месяцев после заражения) также экспрессировали характерный профиль индуцированных генов по сравнению с неинфицированными контрольными животными, который включал гены, участвующие в презентации антигена, защитных ответах и регуляции различных клеточных процессов. К шести месяцам большинство животных переходят к «нормальному» профилю экспрессии генов до инфицирования, но некоторые продолжают экспрессировать гены, связанные с инфекцией.
К шести месяцам большинство животных переходят к «нормальному» профилю экспрессии генов до инфицирования, но некоторые продолжают экспрессировать гены, связанные с инфекцией.
РИСУНОК 3-6
Ответ на инфекцию Salmonella у мышей 129sv после иммунизации. Транскрипционный ответ хозяина в клетках периферической крови измеряли у наивных мышей, зараженных оральной инфекцией Salmonella typhimurium (красный). Транскрипционный ответ (подробнее …)
Благодаря моделям мышей, разработанным Denise Monack et al. (2004a), теперь можно наблюдать за передачей Salmonella от постоянно инфицированных животных, которые протекают бессимптомно, но выделяют бактерии в течение года.Наивные мыши, помещенные в ту же клетку, что и постоянно инфицированные животные, заражаются. Некоторые, но не все, становятся «супершедерами», выделяя очень высокие уровни бактерий со стулом и передавая инфекцию своим неинфицированным товарищам по клетке (Lawley et al., 2008). Эпидемиологи выявили людей, которые также переносят некоторые инфекционные заболевания. Хотя они протекают бессимптомно, у супершеддеров Salmonella обнаружена выраженная воспалительная реакция по сравнению с другими постоянно инфицированными мышами (которые выделяют относительно низкие уровни бактерий).
Хотя они протекают бессимптомно, у супершеддеров Salmonella обнаружена выраженная воспалительная реакция по сравнению с другими постоянно инфицированными мышами (которые выделяют относительно низкие уровни бактерий).
Бактерии от супершедеров не более вирулентны или передаются, чем бактерии от других инфицированных мышей. Неясно, какие факторы способствовали появлению собственности супершеддера. Мыши являются генетически инбредными, поэтому, предположительно, превращение животных в супершеддер было обусловлено негенетическими факторами. У мышей с ослабленным иммунитетом из-за лечения стероидами больше не было шансов стать супершеддерами. Однако, если мышам вводят небольшую разовую дозу неомицина — достаточную для уменьшения комменсальной кишечной флоры менее чем на 48 часов — а затем через четыре дня они подвергаются воздействию Salmonella , почти все они становятся супершеддерами (Lawley et al., 2008). У этих животных уровни воспаления кишечника переходят от низкого к высокому, и в то же время они становятся очень эффективными переносчиками болезни. Одна из возможных интерпретаций этого наблюдения заключается в том, что тонкий баланс между комменсальной флорой и врожденной иммунной системой нарушается из-за устранения некоторых представителей нормальной флоры путем уничтожения антибиотиками и предоставляет окно возможностей для создания более крупных популяций сальмонелл . в кишечнике.
Одна из возможных интерпретаций этого наблюдения заключается в том, что тонкий баланс между комменсальной флорой и врожденной иммунной системой нарушается из-за устранения некоторых представителей нормальной флоры путем уничтожения антибиотиками и предоставляет окно возможностей для создания более крупных популяций сальмонелл . в кишечнике.
Патогены против комменсалов
Термин патоген происходит от греческого слова, означающего «рождение боли». Успешный патоген должен выполнять следующие действия:
Получать гены вирулентности
Оценивать окружающую среду
Включение и выключение генов вирулентности
Переход к месту заражения
Получение питательных веществ
Выживание в стрессе
Избегание иммунной системы хозяина
Подрыв цитоскелета хозяина и сигнальных путей
Распространение на другие сайты
- в некоторых случаях
- Передаться новому восприимчивому хозяину
Успешные комменсалы также делают многие из этих вещей, но, в отличие от патогена, комменсал не обладает внутренней способностью преодолевать анатомические барьеры или нарушать защитные механизмы хозяина, которые ограничивают выживание или репликацию других микробов . Как отмечалось ранее, патогены (но не комменсалы) предпочитают жить в опасных местах, чтобы избежать конкуренции и получить питательные вещества. Эти инвазивные свойства необходимы для выживания патогена в природе.
Как отмечалось ранее, патогены (но не комменсалы) предпочитают жить в опасных местах, чтобы избежать конкуренции и получить питательные вещества. Эти инвазивные свойства необходимы для выживания патогена в природе.
Тем не менее, некоторые представители бактериальной флоры человека, которые обычно беспрепятственно живут в носоглотке человека, включая Streptococcus pneumoniae , Neisseria meningitidis , Haemophilus influenzae типа b и Streptococcus pyogenes , иногда вызывают заболевания.Эти микробы, которых я называю «комменсальными патогенами», обладают детерминантами вирулентности, которые предполагают, что они регулярно вступают в тесный контакт с элементами врожденной и адаптивной иммунной системы (Falkow, 2006). Недавняя история показывает, что иммунизация против этих патогенов не только предотвращает заболевание человека, но также устраняет способность микробов эффективно колонизировать человека-хозяина. Такие комменсальные патогены сохраняются у значительной части человеческой популяции, подавляющее большинство которой являются бессимптомными носителями.
Это вызывает несколько вопросов: являются ли такие микробы патогенами или они комменсалы? Часто ли факторы «вирулентности» на самом деле являются подмножеством адаптивных факторов, которые позволяют микробам использовать определенную нишу, но не обязательно предназначены для того, чтобы вызывать заболевание? Являются ли они факторами вирулентности или лучше называть факторами колонизации ? Я полагаю, что внимание медицины к болезням может иногда отвлекать нас от понимания биологии патогенности. Конечно, важно, чтобы медицинское определение патогена основывалось на болезни.С другой стороны, я бы сказал, что болезнь не охватывает биологические аспекты патогенности и эволюции взаимоотношений паразит-хозяин. Многие патогены вызывают стойкие инфекции у людей (например, Mycobacterium tuberculosis , Treponema pallidum [причина сифилиса], Chlamydia , S. typhi и Helicobacter pylori ), и большинство таких инфекций протекает бессимптомно. Могут ли эти микробы считаться комменсальными патогенами?
Кроме того, как можно предположить, снижение числа инфекционных заболеваний в промышленно развитых странах сопровождалось увеличением иммунных расстройств, таких как рассеянный склероз, болезнь Крона и астма. Уничтожение Helicobacter было связано с увеличением числа случаев рака пищевода и, в последнее время, астмы (Blaser et al., 2008). Возможно, одна треть или более населения мира бессимптомно колонизирована микобактериями Mycobacterium tuberculosis . В течение прошлого века, когда западное общество устранило этот патоген с помощью основных мер общественного здравоохранения, кажется, что мы, возможно, действительно устранили микроб, с которым у нашего вида был диалог, микроб, который «разговаривал» с иммунной системой человека. и держал его готовым к оборонительным действиям.Возможно, как предположил карикатурист Pogo Уолт Келли, «мы встретили врага, и он — это мы».
Уничтожение Helicobacter было связано с увеличением числа случаев рака пищевода и, в последнее время, астмы (Blaser et al., 2008). Возможно, одна треть или более населения мира бессимптомно колонизирована микобактериями Mycobacterium tuberculosis . В течение прошлого века, когда западное общество устранило этот патоген с помощью основных мер общественного здравоохранения, кажется, что мы, возможно, действительно устранили микроб, с которым у нашего вида был диалог, микроб, который «разговаривал» с иммунной системой человека. и держал его готовым к оборонительным действиям.Возможно, как предположил карикатурист Pogo Уолт Келли, «мы встретили врага, и он — это мы».
РИСУНОК 3-7
Спад многих инфекционных заболеваний в промышленно развитых странах. (A) сопровождалось увеличением иммунных расстройств (B), США, 1950–2000 гг. ИСТОЧНИК: Bach (2002).
Если природа микробной патогенности является шизофренической — характеризуется несогласованными или противоречивыми элементами — тогда важно изучить каждый аспект его биологии и не отвлекаться на его роль в возникновении болезни. Вместо этого мы должны следовать инструкциям Томаса Хаксли, выраженным в цитате из письма Хаксли Чарльзу Кингсли от 23 сентября 1860 года, которое мы с Ледербергом очень дорожили: «Сядь перед фактом, как маленький ребенок, будь готов дать укрепляйте все предвзятые представления, смиренно следуйте, куда бы и в какие бы бездны ни приводила природа, иначе вы ничему не научитесь »(Huxley, 1901).
Вместо этого мы должны следовать инструкциям Томаса Хаксли, выраженным в цитате из письма Хаксли Чарльзу Кингсли от 23 сентября 1860 года, которое мы с Ледербергом очень дорожили: «Сядь перед фактом, как маленький ребенок, будь готов дать укрепляйте все предвзятые представления, смиренно следуйте, куда бы и в какие бы бездны ни приводила природа, иначе вы ничему не научитесь »(Huxley, 1901).
Благодарности
Джош Ледерберг был замечательным человеком с замечательным интеллектом. Его наследие еще предстоит полностью оценить.Он повлиял на меня с самого начала моей карьеры, и хотя я неоднократно говорил ему об этом лично, я считаю уместным выразить ему свою благодарность и здесь.
Мои замечания на этом симпозиуме были обобщены Эйлин Чоффнес и ее сотрудниками и легли в основу данной статьи. Я благодарен Люсинде Томпсон и Дениз Монак за разрешение представить здесь некоторые из их ранее неопубликованных результатов по реакции хозяина у наивных и иммунизированных мышей на инфекцию Salmonella . Я также благодарю Дениз Монак и Тревора Лоули за то, что они обсудили со мной их недавние захватывающие открытия относительно передачи Salmonella .
Я также благодарю Дениз Монак и Тревора Лоули за то, что они обсудили со мной их недавние захватывающие открытия относительно передачи Salmonella .
ЭВОЛЮЦИЯ ВЗАИМОДЕЙСТВИЙ БАКТЕРИАЛЬНО-ХОЗЯИН: ВИРУЛЕНТНОСТЬ И ИММУННАЯ ПЕРЕРЕАКЦИЯ
3Элиза Марголис 4
Университет Эмори
Брюс Р. Левин, доктор философии 4
Университет Эмори
Хотя многие люди могут не верить в эволюцию, для тех из нас, у кого отличный вкус и удача работать с бактериями, вирусами и одноклеточными грибами, эволюция — это не вопрос веры, и тем более веры.Эволюция — это то, что мы постоянно видим, хотим мы того или нет. Те, кто являются эволюционными биологами по образованию, склонности или стремлениям, обязаны быть чем-то большим, чем просто свидетелями и историками эволюции. Мы должны дать объяснения происхождению и поддержанию всех биологических явлений. Никаких исключений быть не может.
Придумать эти объяснения, а еще лучше — проверяемые эволюционные гипотезы, несложно для персонажей, которые обеспечивают очевидные преимущества приспособленности для организмов, которые их выражают. Рост устойчивости после введения антибиотиков не стал неожиданностью для биологов-эволюционистов. В присутствии антибиотиков бактерии, устойчивые к их действию, имеют очевидное селективное преимущество по сравнению со своими восприимчивыми предками. Сложнее объяснить ситуации, когда неясно, как рассматриваемый персонаж мог развиться в результате естественного отбора в пользу отдельных организмов. В то время как взаимодействия между паразитическими бактериями и их млекопитающими-хозяевами включают множество признаков, которые можно объяснить естественным отбором, действующим на уровне отдельных бактерий или отдельных хозяев (Burnet and White, 1972), многие не могут.Вирулентность — одна из этих черт, которую трудно объяснить простыми эволюционными моделями; почему бактерии причиняют вред хозяевам, необходимым им для их выживания?
Рост устойчивости после введения антибиотиков не стал неожиданностью для биологов-эволюционистов. В присутствии антибиотиков бактерии, устойчивые к их действию, имеют очевидное селективное преимущество по сравнению со своими восприимчивыми предками. Сложнее объяснить ситуации, когда неясно, как рассматриваемый персонаж мог развиться в результате естественного отбора в пользу отдельных организмов. В то время как взаимодействия между паразитическими бактериями и их млекопитающими-хозяевами включают множество признаков, которые можно объяснить естественным отбором, действующим на уровне отдельных бактерий или отдельных хозяев (Burnet and White, 1972), многие не могут.Вирулентность — одна из этих черт, которую трудно объяснить простыми эволюционными моделями; почему бактерии причиняют вред хозяевам, необходимым им для их выживания?
В этой главе (спекулятивная напыщенная речь, если хотите) мы сосредотачиваемся в первую очередь на аспектах эволюции взаимодействий бактерия-хозяин (в основном человека), которые не могут быть легко объяснены простыми эволюционными сценариями, ориентированными на пользу индивидуума. Мы постулируем и предоставляем доказательства того, что большая часть вирулентности бактериальных инфекций может быть связана с кажущейся ошибочной чрезмерной реакцией иммунной защиты, которую иногда называют «дружественным огнем» (Levin and Anita, 2001; Whitnack, 1993) или иммунопатологией ( Graham et al., 2005). Мы рассматриваем, как эта извращенность иммунной системы согласуется с текущими гипотезами об эволюции вирулентности, эволюции так называемых факторов вирулентности, и размышляем о причинах, по которым естественный отбор не смог или не смог подавить чрезмерную реакцию иммунной системы на бактериальные инфекции. . В заключение мы кратко обсудим значение этого взгляда на вирулентность для лечения бактериальных инфекций.
Мы постулируем и предоставляем доказательства того, что большая часть вирулентности бактериальных инфекций может быть связана с кажущейся ошибочной чрезмерной реакцией иммунной защиты, которую иногда называют «дружественным огнем» (Levin and Anita, 2001; Whitnack, 1993) или иммунопатологией ( Graham et al., 2005). Мы рассматриваем, как эта извращенность иммунной системы согласуется с текущими гипотезами об эволюции вирулентности, эволюции так называемых факторов вирулентности, и размышляем о причинах, по которым естественный отбор не смог или не смог подавить чрезмерную реакцию иммунной системы на бактериальные инфекции. . В заключение мы кратко обсудим значение этого взгляда на вирулентность для лечения бактериальных инфекций.
Вирулентность бактерий как иммунный ответ
Мы определяем вирулентность как величину заболеваемости и увеличение вероятности смерти в результате колонизации и размножения бактерий внутри или на хозяине.Чтобы облегчить рассмотрение этой вирулентности и ее эволюции, мы используем грубое упрощение, рисунок взаимодействия бактерии и хозяина, представленный на рис. Бактерии попадают в участок, синий ящик, где они размножаются, создают популяцию и колонизируют хозяина, но не вызывают заметных симптомов. Вирулентность требует их прохождения во второй участок, красный ящик, где присутствие бактерий (или их продуктов) может, но не обязательно, вызывать симптомы, например, для бактериемии Streptococcus pneumoniae синим участком является носоглотка, а красным — носоглотка. это кровоток.В этой модели красный участок не обязательно должен находиться в другом физическом месте. Это может быть другое состояние бактерий в месте их колонизации, например, для кожной инфекции Staphylococcus aureus синим участком будет кожа, а красным — фурункул.
Бактерии попадают в участок, синий ящик, где они размножаются, создают популяцию и колонизируют хозяина, но не вызывают заметных симптомов. Вирулентность требует их прохождения во второй участок, красный ящик, где присутствие бактерий (или их продуктов) может, но не обязательно, вызывать симптомы, например, для бактериемии Streptococcus pneumoniae синим участком является носоглотка, а красным — носоглотка. это кровоток.В этой модели красный участок не обязательно должен находиться в другом физическом месте. Это может быть другое состояние бактерий в месте их колонизации, например, для кожной инфекции Staphylococcus aureus синим участком будет кожа, а красным — фурункул.
РИСУНОК 3-8
Художественная концепция процесса заражения, иммунного ответа и избыточной реакции хозяина. Синий — место, где присутствие бактерий не вызывает симптомов — бессимптомно. Красный — участок / штат, где присутствует (подробнее…)
У млекопитающих, как и у млекопитающих, вирулентность проявляется двумя способами, оба из которых требуют, чтобы бактерии проникли в красный потенциально симптоматический участок или состояние: (i) прямое повреждение ткани хозяина вызвано репликацией бактерии и / или производство определенных продуктов (токсинов), или (ii) косвенное повреждение хозяина происходит из-за несоответствующей или чрезмерной реакции иммунной системы.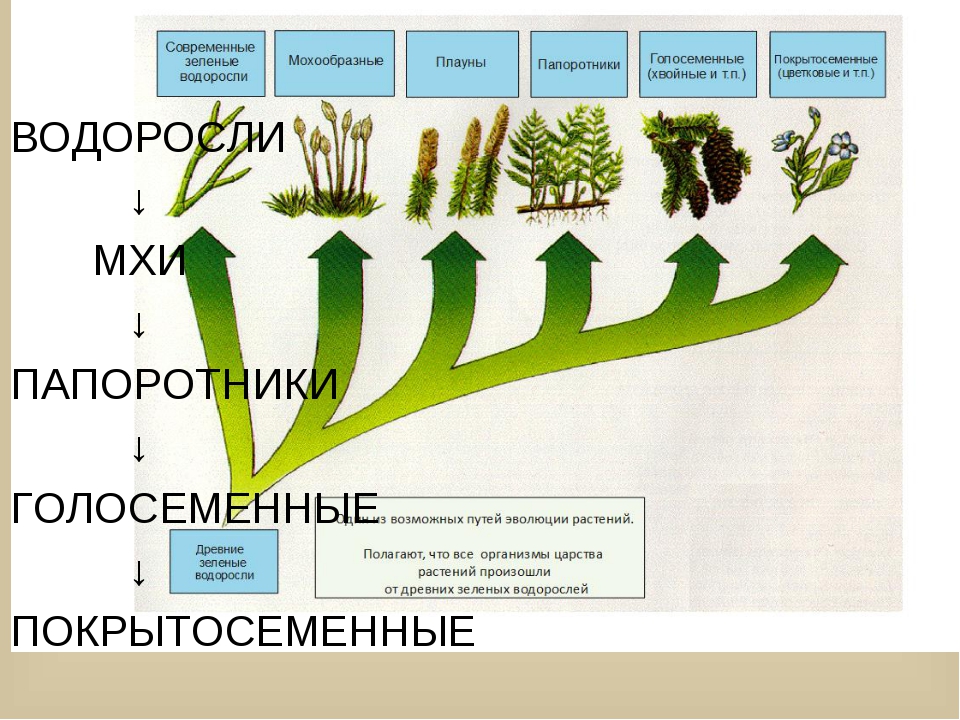 Оба типа повреждений обозначены знаком «!» внутри треугольника. В этой схеме иммунная защита может предотвращать вирулентность одним или несколькими из семи связанных способов:
Оба типа повреждений обозначены знаком «!» внутри треугольника. В этой схеме иммунная защита может предотвращать вирулентность одним или несколькими из семи связанных способов:
Ограничение проникновения бактерий в бессимптомный участок
Ограничение размножения бактерий в бессимптомном участке
Увеличение скорость выведения бактерий и их продуктов из бессимптомного участка
Предотвращение проникновения бактерий или их продуктов в потенциально симптоматический участок или состояние
Снижение скорости распространения бактерий в потенциально симптоматическом участке или состоянии
Увеличение скорости выведения бактерий и их продуктов из потенциально симптоматического места или состояния
Предотвращение иммунной избыточной реакции на бактерии или их продукты в потенциально симптоматическом месте или состоянии
Первые три из эти иммунные ответы поддерживают плотность бактерий и концентрация их продуктов в бессимптомном участке на уровне, при котором они вряд ли перейдут или иным образом проникнут на участок или состояние, где они могут вызвать симптомы.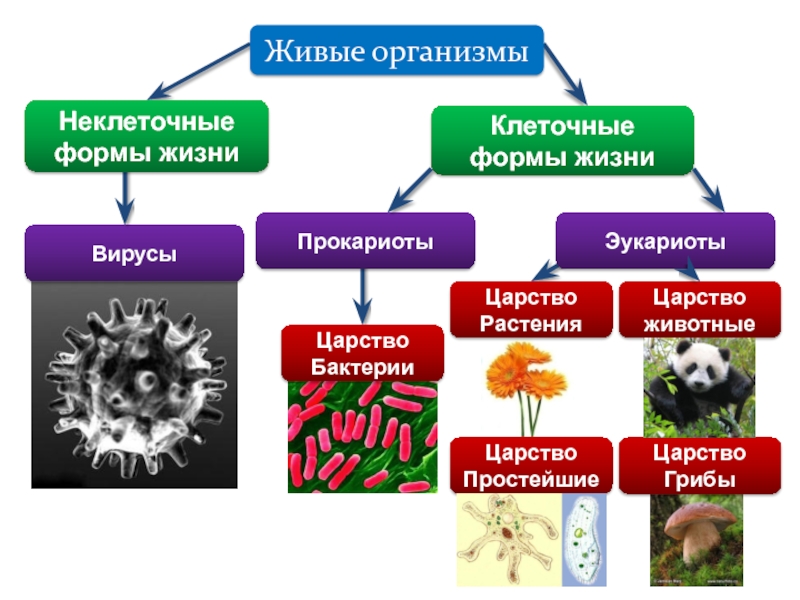 Действительно ли они вызывают симптомы, и величина этих симптомов при переходе в красный участок или состояние также зависит от того, насколько хорошо иммунная система ограничивает их плотность и концентрацию их продуктов. В цифре 8 обозначена инфекционная передача бактерий, способствующая возникновению симптомов, а цифра 9 — передача бактерий из бессимптомного участка. Все эти перечисленные этапы, на которых иммунная система ограничивает вирулентность бактерий, можно классифицировать как соответствующие реакции.Однако несоответствующие ответы, для которых мы используем термин , избыточный ответ , когда они приводят к повреждению хозяина, могут возникать из-за дефектов на одном из этих этапов или как вторичное следствие усиления иммунного ответа.
Действительно ли они вызывают симптомы, и величина этих симптомов при переходе в красный участок или состояние также зависит от того, насколько хорошо иммунная система ограничивает их плотность и концентрацию их продуктов. В цифре 8 обозначена инфекционная передача бактерий, способствующая возникновению симптомов, а цифра 9 — передача бактерий из бессимптомного участка. Все эти перечисленные этапы, на которых иммунная система ограничивает вирулентность бактерий, можно классифицировать как соответствующие реакции.Однако несоответствующие ответы, для которых мы используем термин , избыточный ответ , когда они приводят к повреждению хозяина, могут возникать из-за дефектов на одном из этих этапов или как вторичное следствие усиления иммунного ответа.
Далее мы сосредоточимся в первую очередь на вирулентности, возникающей в результате чрезмерной реакции иммунной системы. Однако есть примеры вирулентности, которые можно отнести к прямому повреждению ткани хозяина репликацией бактерий или секрецией их продуктов. К ним относятся (i) кариес зубов, возникающий в результате кислоты, вырабатываемой при метаболизме Lactobacillius acidophilus или Streptococcus mutans (Gibbons, 1964), (ii) паралич, вызванный нейротоксинами, секретируемыми Clostridium botulinum или Clostridium tetani , действующими на нервных и моторных замыкательных пластинках (Schiavo et al., 1992) и (iii) диарея, вызванная энтеротоксинами, которые ингибируют реабсорбцию хлорида натрия или способствуют его секреции. Примеры вирулентности, являющейся прямым продуктом взаимодействия между бактериями и клетками-хозяевами, по-видимому, редки по сравнению с теми, в которых заболеваемость и смертность можно отнести к косвенным повреждениям, вызванным избыточной иммунной реакцией.
К ним относятся (i) кариес зубов, возникающий в результате кислоты, вырабатываемой при метаболизме Lactobacillius acidophilus или Streptococcus mutans (Gibbons, 1964), (ii) паралич, вызванный нейротоксинами, секретируемыми Clostridium botulinum или Clostridium tetani , действующими на нервных и моторных замыкательных пластинках (Schiavo et al., 1992) и (iii) диарея, вызванная энтеротоксинами, которые ингибируют реабсорбцию хлорида натрия или способствуют его секреции. Примеры вирулентности, являющейся прямым продуктом взаимодействия между бактериями и клетками-хозяевами, по-видимому, редки по сравнению с теми, в которых заболеваемость и смертность можно отнести к косвенным повреждениям, вызванным избыточной иммунной реакцией.
Как показано на рисунке, заболеваемость и смертность от бактериальных инфекций может быть отнесена на счет иммунной системы хозяина, действующей одним из трех неподходящих способов: (i) быть более энергичным, чем необходимо, (ii) неподходящим для этого патогена, или (iii) ) реагируя на неправильные сигналы. Наиболее изученным примером слишком энергичной реакции иммунной системы является бактериальный сепсис, когда попадание цитокинов и бактерий в кровоток вызывает широко распространенное повреждение кровеносных сосудов и полиорганную недостаточность (нарушение функции легких, печени или почек).Здесь реакция на бактерии является приемлемой на одном уровне, поскольку высвобождаемые цитокины играют важную роль в привлечении нейтрофилов (иммунных клеток, фагоцитирующих бактерии) к локальному участку инфекции, но также являются чрезмерными (Kurahashi et al., 1999). Различие между несоответствующим и адекватным иммунным ответом можно увидеть в спектре заболеваний, связанных с Mycobacterium leprae (Modlin, 2002; Sieling et al., 1999). Хозяева, которые реагируют на инфекцию преимущественно антителами и очень небольшим количеством Т-лимфоцитов CD4, имеют инфекционные участки с крупными макрофагами, которые содержат множество микобактерий.Эти макрофаги ответственны за множественные поражения кожи и узелки, наблюдаемые при лепроматозной лепре, в то время как хозяин с ответом типа Т-хелперов 1 (высокая продукция интерферона-γ и низкий уровень интерлейкина-4 [IL-4]) имеет многочисленные хорошо сформированные гранулемы с очень мало микобактерий, образующих незначительные поражения кожи.
Наиболее изученным примером слишком энергичной реакции иммунной системы является бактериальный сепсис, когда попадание цитокинов и бактерий в кровоток вызывает широко распространенное повреждение кровеносных сосудов и полиорганную недостаточность (нарушение функции легких, печени или почек).Здесь реакция на бактерии является приемлемой на одном уровне, поскольку высвобождаемые цитокины играют важную роль в привлечении нейтрофилов (иммунных клеток, фагоцитирующих бактерии) к локальному участку инфекции, но также являются чрезмерными (Kurahashi et al., 1999). Различие между несоответствующим и адекватным иммунным ответом можно увидеть в спектре заболеваний, связанных с Mycobacterium leprae (Modlin, 2002; Sieling et al., 1999). Хозяева, которые реагируют на инфекцию преимущественно антителами и очень небольшим количеством Т-лимфоцитов CD4, имеют инфекционные участки с крупными макрофагами, которые содержат множество микобактерий.Эти макрофаги ответственны за множественные поражения кожи и узелки, наблюдаемые при лепроматозной лепре, в то время как хозяин с ответом типа Т-хелперов 1 (высокая продукция интерферона-γ и низкий уровень интерлейкина-4 [IL-4]) имеет многочисленные хорошо сформированные гранулемы с очень мало микобактерий, образующих незначительные поражения кожи.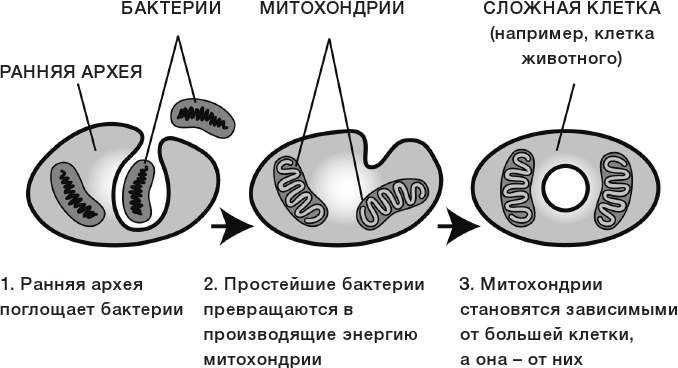 Суперантигены представляют собой пример того, как иммунная система реагирует на неверный сигнал. Суперантигены — это бактериальные продукты, которые стимулируют большое количество Т-клеток (от 1 до 40% Т-клеток будут реагировать) путем связывания с молекулами основного комплекса гистосовместимости класса II и рецепторами Т-клеток (бета-цепь) независимо от их специфичности в отношении антигенов (Rott and Fleischer , 1994). S. aureus , Streptococcus pyogenes , Mycoplasma arthritidis и Yersinia pseudotuberculosis относятся к числу бактерий, продуцирующих суперантигены. В случае синдрома токсического шока суперантигены, продуцируемые S. aureus , вызывают беспорядочную и подавляющую активацию Т-клеток, ведущую к выработке цитокинов, которые опосредуют шок и повреждение тканей. Во всех трех случаях заболеваемость и смертность хозяина могут быть связаны с явно ошибочной реакцией иммунной системы, которую мы называем чрезмерной реакцией.
Суперантигены представляют собой пример того, как иммунная система реагирует на неверный сигнал. Суперантигены — это бактериальные продукты, которые стимулируют большое количество Т-клеток (от 1 до 40% Т-клеток будут реагировать) путем связывания с молекулами основного комплекса гистосовместимости класса II и рецепторами Т-клеток (бета-цепь) независимо от их специфичности в отношении антигенов (Rott and Fleischer , 1994). S. aureus , Streptococcus pyogenes , Mycoplasma arthritidis и Yersinia pseudotuberculosis относятся к числу бактерий, продуцирующих суперантигены. В случае синдрома токсического шока суперантигены, продуцируемые S. aureus , вызывают беспорядочную и подавляющую активацию Т-клеток, ведущую к выработке цитокинов, которые опосредуют шок и повреждение тканей. Во всех трех случаях заболеваемость и смертность хозяина могут быть связаны с явно ошибочной реакцией иммунной системы, которую мы называем чрезмерной реакцией.
ТАБЛИЦА 3-1
Некоторые примеры вирулентности в результате иммунной избыточной реакции.
Эволюция бактериальной вирулентности как иммунный ответ
Как наблюдение, что большая часть заболеваемости и смертности может быть приписана чрезмерной реакции хозяина на бактерии, помогает понять эволюцию вирулентности? В перспективе, написанной десять лет назад, один из нас (Левин, 1996) перечислил четыре гипотезы, объясняющие эволюцию вирулентности: (i) общепринятое мнение, (ii) эпидемиологический отбор, (iii) случайная эволюция и (iv) близорукая эволюция внутри хозяина.С того времени, хотя был опубликован ряд теоретических, экспериментальных и спекулятивных статей об эволюции вирулентности (небольшую и, по общему признанию, предвзятую выборку см. Andre and Godelle, 2006; Bonhoeffer and Nowak, 1994; Brown et al., 2006). ; Bull, 1994; Ebert and Bull, 2003; Ebert and Herre, 1996; Frank, 1996; Grech et al., 2006; Lipsitch et al., 1995, 1996; Lipsitch, Moxon, 1997; Mackinnon and Read, 2004; Regoes et al., 2000), мы не знаем исследований, которые отвергли бы какую-либо из этих гипотез, и только одно добавило то, что может быть новой гипотезой: эволюция квазивидов (Pfeiffer and Kirkegaard, 2005). Эта пятая гипотеза может применяться только к вирусам с высокой частотой мутаций и, возможно, может быть отнесена к более широкой рубрике эволюции внутри хозяина. В этом разделе мы рассмотрим, как наблюдение о том, что заболеваемость и смертность от бактериальных инфекций могут быть приписаны чрезмерной иммунной реакции хозяев, соответствует каждой из этих гипотез эволюции вирулентности бактерий.
Эта пятая гипотеза может применяться только к вирусам с высокой частотой мутаций и, возможно, может быть отнесена к более широкой рубрике эволюции внутри хозяина. В этом разделе мы рассмотрим, как наблюдение о том, что заболеваемость и смертность от бактериальных инфекций могут быть приписаны чрезмерной иммунной реакции хозяев, соответствует каждой из этих гипотез эволюции вирулентности бактерий.
Общепринятая мудрость
Эту фразу, которую покойный Джон Кеннет Гэлбрейт придумал для описания идей и объяснений, которые широко признаны общественностью как истинные, Боб Мэй и Рой Андерсон (May and Anderson, 1983b) применили для описания тогдашний взгляд на эволюцию вирулентности.Согласно этой мудрости, вирулентность является артефактом относительной новизны ассоциации паразита с его хозяином. По мере развития отношений между паразитом и хозяином естественный отбор в популяции паразита или хозяина или в обоих приведет к исчезновению одного или другого вида или к эволюции симбиоза или мутуализма.
Хотя исходная теория, лежащая в основе этой гипотезы эволюции вирулентности инфекций, сводится к поговорке «не кусай руку, которая тебя кормит», доказательства в ее поддержку были и остаются убедительными.Многие из бактерий, ответственных за заболеваемость и смертность людей, были приобретены от других видов в недалеком прошлом (после появления сельского хозяйства), а некоторые постоянно приобретаются таким образом. К этим зоонозным (и протозоонозным) инфекциям относятся чума, туберкулез, болезнь легионеров, ботулизм, сибирская язва, бруцеллез, туляремия, пятнистая лихорадка Скалистых гор, холера и другие диарейные заболевания. Бактерии, вызывающие некоторые из этих инфекций, такие как Mycobacterium tuberculosis , передаются от человека к человеку и могут сохраняться без животного происхождения.Другие, такие как Legionella pneumophila , нет. Также с общепринятой точкой зрения согласуются коррелированные наблюдения о том, что лишь очень незначительное меньшинство из огромного числа видов бактерий, колонизирующих млекопитающих, вызывают болезни.
Может показаться, что предположение о том, что вирулентность бактериальных инфекций может быть объяснена чрезмерной реакцией иммунной системы хозяина, вполне согласуется с этим общепринятым мнением. То есть иммунная система еще не успела развиться, чтобы смягчить реакцию на эти новые бактерии и их продукты, и / или эти бактерии еще не превратились в хорошие.В конце концов, или, как это когда-то называли, в «день равновесия» (Левин и др., 2000) возобладает мутуализм, и иммунная избыточная реакция смягчается.
Эпидемиологический отбор
Принято считать, что это наблюдение, а не механизм, наблюдение, которое фокусируется на взаимодействиях между бактериями и отдельными хозяевами, которых они колонизируют. Однако, чтобы полностью понять эволюцию комменсальных и патогенных бактерий, необходимо рассмотреть их образ жизни вне организма-хозяина и, в частности, их передачу между хозяевами.Один из подходов к этой более полной картине эволюции паразитических микробов заключался в том, чтобы сделать выводы о природе и направлении отбора на основе эпидемиологических моделей (Levin et al. , 1982; Levin and Pimentel, 1981; May and Anderson, 1983a). В соответствии с этой точкой зрения, приспособленность конкретного штамма бактерий определяется его основным репродуктивным числом, R 0 , количеством вторичных инфекций, вызванных одним инфицированным индивидуумом в полностью восприимчивой популяции хозяев; чем выше значение R 0 , тем выше приспособленность бактерий.В этих традиционных эпидемиологических моделях вирулентность выражается только как смертность. Заболеваемость и другие более тонкие эффекты инфекций не рассматриваются напрямую в этих эпидемиологических моделях.
, 1982; Levin and Pimentel, 1981; May and Anderson, 1983a). В соответствии с этой точкой зрения, приспособленность конкретного штамма бактерий определяется его основным репродуктивным числом, R 0 , количеством вторичных инфекций, вызванных одним инфицированным индивидуумом в полностью восприимчивой популяции хозяев; чем выше значение R 0 , тем выше приспособленность бактерий.В этих традиционных эпидемиологических моделях вирулентность выражается только как смертность. Заболеваемость и другие более тонкие эффекты инфекций не рассматриваются напрямую в этих эпидемиологических моделях.
Пока для переноса к новым хозяевам требуются жизнеспособные хозяева, отбор будет отдавать предпочтение бактериям, которые не только инфекционно передаются с еще более высокой скоростью, но и дольше сохраняются в колонизированных хозяевах (т. Е. С меньшей вероятностью убивают хозяина). Другими словами, отбор будет отдавать предпочтение более доброкачественным, симбиотическим или, что еще лучше, мутуалистическим бактериям. Эволюция популяции хозяев также будет способствовать снижению вирулентности; предпочтение будет отдаваться хозяевам, которые менее подвержены инфекционным заболеваниям и смертности. Поскольку передача происходит из синего бессимптомного участка (8 дюймов), а не из красного участка (9 дюймов), эти эпидемиологические модели можно рассматривать как теоретическую основу общепринятого мнения (также см. Lenski and May, 1994). Однако, если передача и заболеваемость и смертность хозяина связаны так, что более вирулентные бактерии передаются с большей скоростью, чем более доброкачественные, существует компромисс между потерей хозяина и выгодой для бактерий; вирулентность будет благоприятствовать бактериальной популяции (Ebert and Bull, 2003).
Эволюция популяции хозяев также будет способствовать снижению вирулентности; предпочтение будет отдаваться хозяевам, которые менее подвержены инфекционным заболеваниям и смертности. Поскольку передача происходит из синего бессимптомного участка (8 дюймов), а не из красного участка (9 дюймов), эти эпидемиологические модели можно рассматривать как теоретическую основу общепринятого мнения (также см. Lenski and May, 1994). Однако, если передача и заболеваемость и смертность хозяина связаны так, что более вирулентные бактерии передаются с большей скоростью, чем более доброкачественные, существует компромисс между потерей хозяина и выгодой для бактерий; вирулентность будет благоприятствовать бактериальной популяции (Ebert and Bull, 2003).
На первый взгляд может показаться, что этот компромисс между передачей и вирулентностью несовместим с предположением о том, что заболеваемость и смертность от инфекции являются продуктом иммунной реакции хозяина. Мы полагаем, что это не всегда так. Чрезмерная реакция хозяина может быть побочным продуктом отбора, действующего на отдельные бактерии, чтобы способствовать их передаче. Хотя мы не знаем неопровержимых количественных и эмпирических доказательств того, что это справедливо для каких-либо патогенных бактерий (вирусы — другое дело; см. Fenner and Ratcliffe, 1965), эта интерпретация поддерживается аргументами разумного правдоподобия.Здесь мы рассмотрим два наиболее убедительных из этих примеров патогенных бактерий человека.
Хотя мы не знаем неопровержимых количественных и эмпирических доказательств того, что это справедливо для каких-либо патогенных бактерий (вирусы — другое дело; см. Fenner and Ratcliffe, 1965), эта интерпретация поддерживается аргументами разумного правдоподобия.Здесь мы рассмотрим два наиболее убедительных из этих примеров патогенных бактерий человека.
Первый — это диарейные заболевания, при которых человек играет важную роль в процессе передачи. Из-за массового размножения бактерий диарея, вероятно, увеличит плотность бактерий в воде и пищевых продуктах и, следовательно, скорость передачи этих бактерий. Таким образом, пока передаче способствует диарея, отбор в бактериальной популяции будет отдавать предпочтение механизмам, вызывающим диарею.В некоторых случаях индукция диареи объясняется тем, что можно рассматривать как чрезмерную реакцию иммунной системы. Дизентерийные бактерии Shigella flexneri индуцируют высвобождение цитокина интерлейкина 1 (ИЛ-1) в инфицированных макрофагах, что приводит к обширному повреждению слизистой оболочки толстой кишки, что, в свою очередь, приводит к потере жидкости и белка в просвет кишечника и, как следствие, диарея (Hilbi et al. , 1997). Эта гипотеза эволюции диареи с целью увеличения передачи требует, чтобы преимущество передачи более чем компенсировало потерю передачи из-за смертности хозяина.Насколько нам известно, нет количественных эмпирических исследований, демонстрирующих, что этот компромисс возможен при любом диарейном заболевании.
, 1997). Эта гипотеза эволюции диареи с целью увеличения передачи требует, чтобы преимущество передачи более чем компенсировало потерю передачи из-за смертности хозяина.Насколько нам известно, нет количественных эмпирических исследований, демонстрирующих, что этот компромисс возможен при любом диарейном заболевании.
Второй пример — чума. Хотя еще не так хорошо задокументирована, как часто рассказываемая мать всех историй о компромиссах, миксома и австралийские кролики (Fenner, 1965), новая история эволюции вирулентности бациллы чумы, Yersinia pestis , имеет параллели с той историей. Имеются убедительные доказательства того, что этот передающийся блохами патоген произошел от не очень вирулентного кишечного, орально-фекального происхождения Yersinia относительно недавно путем приобретения пары плазмид и нескольких хромосомных генов (Achtman et al., 1999; Карниэль, 2003). Поскольку блохи приобретают эти бактерии из крови грызунов, плотность бактерий в циркулирующей крови будет напрямую связана с вероятностью их передачи другим грызунам (или людям). Также с такой плотностью бактерий в крови напрямую связан сепсис, опасное проявление инфекции Y. pestis . Элизабет Карниэль (личное сообщение) предположила, что способность вызывать летальный сепсис — это не просто побочный продукт размножения бактерий в крови, но она может быть выбрана в бактериальной популяции.Хотя расчет рентабельности не производился, возможно, что скорость передачи бактерий увеличивается за счет уничтожения инфицированных грызунов, поскольку блохи перемещаются к новым хозяевам, когда их первоначальный хозяин умирает. И для диарейных заболеваний, и для чумы вирулентность, возникающая в результате чрезмерной реакции хозяина, связана с передачей инфекции. Очевидно, что потребуется дополнительная эмпирическая работа, чтобы подтвердить существование компромисса между бактериальной передачей и иммунной избыточной реакцией, а также постулируемое использование этой избыточной реакции для эпидемиологического преимущества паразита.
Также с такой плотностью бактерий в крови напрямую связан сепсис, опасное проявление инфекции Y. pestis . Элизабет Карниэль (личное сообщение) предположила, что способность вызывать летальный сепсис — это не просто побочный продукт размножения бактерий в крови, но она может быть выбрана в бактериальной популяции.Хотя расчет рентабельности не производился, возможно, что скорость передачи бактерий увеличивается за счет уничтожения инфицированных грызунов, поскольку блохи перемещаются к новым хозяевам, когда их первоначальный хозяин умирает. И для диарейных заболеваний, и для чумы вирулентность, возникающая в результате чрезмерной реакции хозяина, связана с передачей инфекции. Очевидно, что потребуется дополнительная эмпирическая работа, чтобы подтвердить существование компромисса между бактериальной передачей и иммунной избыточной реакцией, а также постулируемое использование этой избыточной реакции для эпидемиологического преимущества паразита.
Coincidental Evolution
В соответствии с этой гипотезой, у бактерий нет преимущества в том, чтобы вызвать заболевание хозяина, и, конечно же, нет преимущества для него, если он болен; вирулентность является следствием того, что бактерии находятся в неправильном хозяине или в неправильном месте у правильного хозяина (Levin and Svanborg Eden, 1990) (стрелка выше 7 дюймов). Бактериальные продукты, ответственные за заболеваемость и / или смертность хозяина, детерминанты вирулентности, эволюционировали в ответ на отбор по некоторой функции, отличной от вирулентности.
Бактериальные продукты, ответственные за заболеваемость и / или смертность хозяина, детерминанты вирулентности, эволюционировали в ответ на отбор по некоторой функции, отличной от вирулентности.
Разумными кандидатами на случайную вирулентность из-за избыточного иммунного ответа являются заболевания, связанные с Helicobacter pylori. Эти бактерии колонизируют и поддерживают популяции в желудках большинства людей на протяжении большей части их жизни, не вызывая симптомов, и, по-видимому, делали это с доисторических времен (Falush et al., 2003). Однако только после того, как Маршалл и Уоррен (1984) представили доказательства того, что изогнутая бактерия, которую мы теперь знаем как H. pylori , была этиологическим агентом желудочной и пептической язвы, эта, казалось бы, комменсальная бактерия была повышена до статуса патогена.Это различие было дополнительно усилено доказательствами того, что H. pylori также были связаны с раком желудка (Moss and Blaser, 2005; Tatematsu et al. , 2005). H. pylori. Колонизация может привести к хроническому воспалительному состоянию, которое возникает, когда реакции хозяина (такие как высвобождение IL-8 и других хемокинов, привлечение нейтрофилов, макрофагов и местная стимуляция Т-клеток) не в состоянии очистить от бактерий и лимфоидных агрегатов в собственной пластинке желудка и двенадцатиперстной кишки.Эта продолжающаяся стимуляция иммунных и воспалительных клеток (так называемый хронический атрофический гастрит) приводит к разрушению эпителия желудка, образованию пептических язв и повышенному риску рака желудка. Предположительно, но еще не продемонстрировано официально, индукция воспалительного ответа и последующих заболеваний не дает преимущества H. pylori в колонизированном хозяине или его передаче новым хозяевам. В этом смысле вирулентность H. pylori у колонизированных людей является случайной.
, 2005). H. pylori. Колонизация может привести к хроническому воспалительному состоянию, которое возникает, когда реакции хозяина (такие как высвобождение IL-8 и других хемокинов, привлечение нейтрофилов, макрофагов и местная стимуляция Т-клеток) не в состоянии очистить от бактерий и лимфоидных агрегатов в собственной пластинке желудка и двенадцатиперстной кишки.Эта продолжающаяся стимуляция иммунных и воспалительных клеток (так называемый хронический атрофический гастрит) приводит к разрушению эпителия желудка, образованию пептических язв и повышенному риску рака желудка. Предположительно, но еще не продемонстрировано официально, индукция воспалительного ответа и последующих заболеваний не дает преимущества H. pylori в колонизированном хозяине или его передаче новым хозяевам. В этом смысле вирулентность H. pylori у колонизированных людей является случайной.
Хотя они обычно описываются как патогены, особенно в заявках на гранты и людьми, страдающими от симптомов, которые они могут вызывать, ряд бактерий, ответственных за заболеваемость и смертность людей, также имеют хорошую репутацию в качестве комменсалов. Как и H. pylori , они переносятся многими бессимптомно и лишь у немногих вызывают заболевание. К наиболее известным из этих комменсальных патогенов для человека относятся S. aureus , Haemophilus influenzae , S.pneumoniae и Neisseria meningitidis . С эволюционной точки зрения, инвазивные заболевания, по-видимому, не подходят для этих бактерий — тупики. Места их вирулентности, кровь и мозговые оболочки, безусловно, не подходят для их передачи новым хозяевам обычным путем через респираторные капли. Редкая вирулентность этих комменсальных бактерий может быть объяснена иммунной реакцией на эти участки (Bergeron et al., 1998; Braun et al., 1999). Случайное перемещение бактерий в места, где они могут вызвать заболевание (красный цвет), может быть следствием случайной или случайной эволюции или, как мы утверждаем ниже, может быть следствием эволюции бактериальной популяции внутри хозяина.
Как и H. pylori , они переносятся многими бессимптомно и лишь у немногих вызывают заболевание. К наиболее известным из этих комменсальных патогенов для человека относятся S. aureus , Haemophilus influenzae , S.pneumoniae и Neisseria meningitidis . С эволюционной точки зрения, инвазивные заболевания, по-видимому, не подходят для этих бактерий — тупики. Места их вирулентности, кровь и мозговые оболочки, безусловно, не подходят для их передачи новым хозяевам обычным путем через респираторные капли. Редкая вирулентность этих комменсальных бактерий может быть объяснена иммунной реакцией на эти участки (Bergeron et al., 1998; Braun et al., 1999). Случайное перемещение бактерий в места, где они могут вызвать заболевание (красный цвет), может быть следствием случайной или случайной эволюции или, как мы утверждаем ниже, может быть следствием эволюции бактериальной популяции внутри хозяина.
Развитие внутри хозяина
В соответствии с этой гипотезой вирулентность бактерий является продуктом отбора в пользу более патогенных членов популяции, колонизирующей отдельного хозяина (Levin and Bull, 1994). Преимущество, полученное бактериями путем создания симптомов у колонизированного хозяина, ограничивается этим хозяином и может быть его недостатком при передаче новому хозяину; эта эволюция недальновидна. Мутантная комменсальная бактерия, способная создавать и поддерживать популяции в обычно стерильных участках, клетках или тканях, может быть предпочтительнее в колонизированном хозяине, потому что в этих участках меньше конкуренция за питательные вещества и / или эти мутантные бактерии в некоторой степени защищены от хозяина. иммунная защита.
Преимущество, полученное бактериями путем создания симптомов у колонизированного хозяина, ограничивается этим хозяином и может быть его недостатком при передаче новому хозяину; эта эволюция недальновидна. Мутантная комменсальная бактерия, способная создавать и поддерживать популяции в обычно стерильных участках, клетках или тканях, может быть предпочтительнее в колонизированном хозяине, потому что в этих участках меньше конкуренция за питательные вещества и / или эти мутантные бактерии в некоторой степени защищены от хозяина. иммунная защита.
Хотя мы можем привести веские доводы и даже привести доказательства вирулентности некоторых вирусов, таких как полиовирус и вирус Коксаки, являющихся продуктом эволюции внутри хозяина (Gay et al., 2006; Levin and Bull, 1994), для Лучшее, что мы можем сделать на этом этапе, — это представить аргументы, основанные на правдоподобности и согласованности с наблюдениями (см., например, Meyers et al., 2003). Центральным в этих аргументах являются результаты исследований на мышах и крысах, демонстрирующие, что бактерии, ответственные за инвазивность (заражение крови), обычно происходят из одной из очень немногих клеток (Meynell, 1957; Moxon and Murphy, 1978; Pluschke et al. , 1983; Рубин, 1987). Одно из возможных объяснений этих наблюдений состоит в том, что бактерии, ответственные за заражение крови, являются продуктами единичных мутантных клеток с повышенной способностью проникать в кровь и размножаться в ней.
, 1983; Рубин, 1987). Одно из возможных объяснений этих наблюдений состоит в том, что бактерии, ответственные за заражение крови, являются продуктами единичных мутантных клеток с повышенной способностью проникать в кровь и размножаться в ней.
Поддерживая гипотезу внутрихозяйственной эволюции вирулентности, эти наблюдения также согласуются с гипотезой случайной эволюции: только по чистой случайности только одна или несколько клеток вызывают инфицирование крови, что может быть связано с очень маленькими дырами в защите хозяина. через которую проходит только одна или несколько бактерий, указанная стрелкой выше 7 дюймов.Хотя гипотезы совпадения и гипотезы внутри хозяина можно отличить, продемонстрировав, что бактерии, вызывающие инфекцию крови, имеют унаследованную склонность к вторжению в кровь, насколько нам известно, нет опубликованных исследований, в которых проводился бы этот тест. Однако независимо от того, является ли инвазивность крови или других обычно стерильных участков случайной или обусловленной эволюцией внутри хозяина, вирулентность бактерий в этих участках может быть отнесена на счет избыточного иммунного ответа хозяина.
Эволюция детерминантов вирулентности
Не все бактерии или даже все представители одного и того же вида бактерий, способные колонизировать млекопитающих, несут ответственность за заболевание.Одним из объяснений того, почему одни бактерии вызывают заболевание, а другие нет, является то, что стало известно как факторы вирулентности или детерминанты вирулентности, выражение которых, по определению, необходимо для того, чтобы эти бактерии вызывали болезнь у (или на) колонизированных хозяев (Finlay и Фалькоу, 1989). Среди них есть признаки, которые способствуют адгезии к клеткам-хозяевам, уклоняются от конститутивной и индуцируемой иммунной защиты хозяина и продуцируют токсины. Соответственно, большая часть современной бактериологии посвящена пониманию молекулярной биологии, генетики, эволюционного происхождения и механизма действия детерминант вирулентности как способ понять бактериальные заболевания и в идеале предотвратить или лечить их.В то время как детерминанты (факторы) вирулентности почти наверняка являются продуктами адаптивной эволюции бактериальных популяций, не так однозначны факторы отбора, ответственные за их эволюцию и поддержание. Являются ли они благоприятными из-за вирулентности, то есть заболеваемости и смертности хозяина способствуют колонизации, сохранению и инфекционной передаче бактерий, которые выражают эти детерминанты? Являются ли факторы вирулентности побочными продуктами отбора для выполнения других функций, например, их экспрессия обеспечивает защиту от пасущихся простейших (Wildschutte et al., 2004) и / или способствует конкуренции с другими микробами? Или вирулентность, приписываемая этим факторам, является непреднамеренным побочным продуктом их нормальной функции в организме хозяина, примитивным характером, который будет утрачен в день равновесия или до него. Хотя эти гипотезы могут быть взаимоисключающими для любой конкретной бактерии-хозяина и фактора вирулентности, они явно не так коллективны. Независимо от того, развиваются ли они в ответ на отбор на вирулентность или нет, некоторые из этих факторов вирулентности ответственны за запуск избыточного иммунного ответа.
Являются ли они благоприятными из-за вирулентности, то есть заболеваемости и смертности хозяина способствуют колонизации, сохранению и инфекционной передаче бактерий, которые выражают эти детерминанты? Являются ли факторы вирулентности побочными продуктами отбора для выполнения других функций, например, их экспрессия обеспечивает защиту от пасущихся простейших (Wildschutte et al., 2004) и / или способствует конкуренции с другими микробами? Или вирулентность, приписываемая этим факторам, является непреднамеренным побочным продуктом их нормальной функции в организме хозяина, примитивным характером, который будет утрачен в день равновесия или до него. Хотя эти гипотезы могут быть взаимоисключающими для любой конкретной бактерии-хозяина и фактора вирулентности, они явно не так коллективны. Независимо от того, развиваются ли они в ответ на отбор на вирулентность или нет, некоторые из этих факторов вирулентности ответственны за запуск избыточного иммунного ответа.
Почему иммунная система слишком быстро реагирует?
Ранее мы изображали иммунную систему хозяина как ошибочную, чрезмерно реагирующую на нее, которая скорее вызывает, чем предотвращает заболеваемость и смертность от бактериальной инфекции. Однако с точки зрения эволюционной биологии «заблуждение» вряд ли может служить объяснением. Колонизация бактериями — не редкое явление, а скорее то, с чем млекопитающие сталкиваются все время, и чрезмерная реакция, приводящая к их заболеваемости и смертности, почти наверняка будет выбрана против.В своем обзоре «иммунопатологии» Грэм и его коллеги постулировали ряд причин такого нарушения иммунного ответа (Graham et al., 2005). Здесь мы предлагаем нашу точку зрения на этот вопрос.
Однако с точки зрения эволюционной биологии «заблуждение» вряд ли может служить объяснением. Колонизация бактериями — не редкое явление, а скорее то, с чем млекопитающие сталкиваются все время, и чрезмерная реакция, приводящая к их заболеваемости и смертности, почти наверняка будет выбрана против.В своем обзоре «иммунопатологии» Грэм и его коллеги постулировали ряд причин такого нарушения иммунного ответа (Graham et al., 2005). Здесь мы предлагаем нашу точку зрения на этот вопрос.
Как мы видим, существует два основных класса объяснений поддержания чрезмерной реакции иммунной системы. (i) Хотя инфекционное заболевание может быть основным источником заболеваемости и смертности (Haldane, 1949), отбор, опосредованный болезнью, может быть относительно слабым, и потребуется много времени для разработки механизмов, модулирующих иммунный ответ на определенные бактериальные инфекции. .(ii) Функциональные ограничения иммунной системы ограничивают способность естественного отбора полностью предотвращать и, возможно, даже частично снижать иммунный ответ на бактериальные инфекции.
(a) Даже если отбор повсеместно способствует сдерживанию чрезмерной реакции иммунной системы на инфекции и могут быть созданы предпочтительные генотипы (что мы ставим под вопрос ниже [b]), время, необходимое для развития умеренности, может быть значительным, особенно если чрезмерная реакция будет специфические для конкретных бактерий и / или их продуктов.Это связано с двумя факторами. (a) В максимальной степени интенсивность отбора для модулирования иммунного ответа на инфекцию равнялась бы доле популяции с этой инфекцией. Он был бы значительно ниже, если бы симптомы инфекции не проявлялись у всех колонизированных хозяев, редко были бы смертельными или стерилизующими, или проявлялись бы в первую очередь после репродуктивного возраста, или если бы величина снижения избыточной реакции предпочтительного генотипа была меньше абсолютной. . Большинство болезней, перечисленных в списке вирулентности, редко встречаются у колонизированных хозяев (менее 1%), и поэтому интенсивность отбора против иммунного избыточного ответа будет относительно слабой. (б) Редкому полезному мутанту может потребоваться значительное количество времени, чтобы подняться до значительных частот. Например, если отбор для снижения избыточной реакции действует на генотипы в одном локусе (лучший случай), начальная частота предпочтительного аллеля составляет 10 −3 , предпочтительный генотип имеет селективное преимущество 1%, и там не является доминированием, потребуется 1381 поколение (более 20000 лет для человека), чтобы этот ген достиг частоты 50%. Если предпочтительный генотип рецессивен, соответствующее количество поколений будет 100 491 (Crow and Kimura, 1971).
(б) Редкому полезному мутанту может потребоваться значительное количество времени, чтобы подняться до значительных частот. Например, если отбор для снижения избыточной реакции действует на генотипы в одном локусе (лучший случай), начальная частота предпочтительного аллеля составляет 10 −3 , предпочтительный генотип имеет селективное преимущество 1%, и там не является доминированием, потребуется 1381 поколение (более 20000 лет для человека), чтобы этот ген достиг частоты 50%. Если предпочтительный генотип рецессивен, соответствующее количество поколений будет 100 491 (Crow and Kimura, 1971).
Как насчет роли бактерий в эволюции более умеренной иммунной системы? Вследствие значительно более короткого времени генерации, гаплоидных геномов и склонности к получению генов и островков патогенности путем горизонтального переноса кажется разумным предположить, что бактерии будут иметь преимущество в эволюционной гонке вооружений со своими млекопитающими-хозяевами. Мы предполагаем, однако, что это преимущество почти не влияет на замедление темпа, с которым эволюция млекопитающих может модулировать избыточный иммунный ответ. Хотя могут быть ситуации, когда вирулентность положительно коррелирует с инфекционной передачей бактерий, в большинстве этих случаев заболеваемость и смертность, связанные с их передачей, не в пользу бактерий, а могут быть в их ущерб. Еще большая передача этих бактерий была бы возможна, если бы хозяева не были ослаблены или убиты в результате диареи или если бактериемии, необходимые для трансмиссивной передачи, не привели к сепсису. В этой интерпретации эволюция популяции бактерий не будет препятствовать эволюции более умеренной иммунной системы хозяина.Из всех примеров, рассмотренных в этой главе, единственный, в котором эволюция бактериальной популяции может способствовать чрезмерному иммунному ответу, — это предположение Карниэля о том, что, убивая своего хозяина, Y. pestis получает преимущество в передаче.
Хотя могут быть ситуации, когда вирулентность положительно коррелирует с инфекционной передачей бактерий, в большинстве этих случаев заболеваемость и смертность, связанные с их передачей, не в пользу бактерий, а могут быть в их ущерб. Еще большая передача этих бактерий была бы возможна, если бы хозяева не были ослаблены или убиты в результате диареи или если бактериемии, необходимые для трансмиссивной передачи, не привели к сепсису. В этой интерпретации эволюция популяции бактерий не будет препятствовать эволюции более умеренной иммунной системы хозяина.Из всех примеров, рассмотренных в этой главе, единственный, в котором эволюция бактериальной популяции может способствовать чрезмерному иммунному ответу, — это предположение Карниэля о том, что, убивая своего хозяина, Y. pestis получает преимущество в передаче.
(ii) Хотя приведенные выше реалии экологии и генетики естественного отбора могут быть частью ответа на вопрос, почему эволюция не устранила чрезмерную реакцию иммунной системы на бактериальные (и другие) инфекции, мы полагаем, что это не так.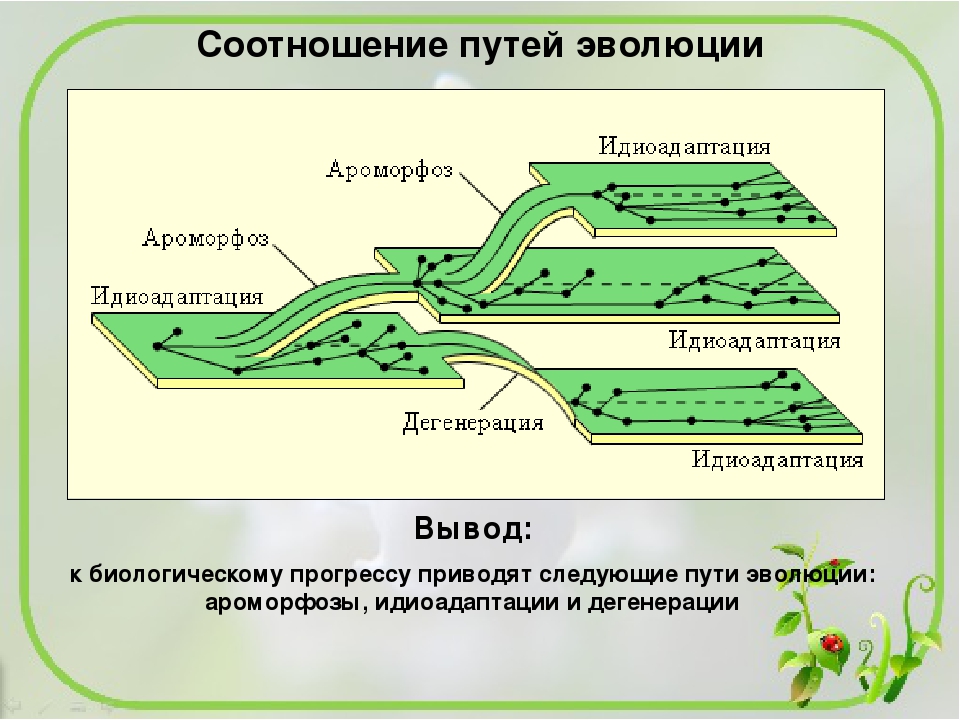 самая важная причина.Мы предполагаем, что основная причина, по которой эволюция млекопитающих не замедлилась и, возможно, не может сдержать чрезмерную реакцию иммунной системы на бактериальные и другие инфекции, — это функциональные ограничения, которые ограничивают степень, в которой иммунная система может быть изменена. Иммунная система не только устраняет бактериальные инфекции, но и выполняет другие функции. Было высказано предположение, что эти другие роли доминировали в эволюции иммунной системы млекопитающих (Burnet, 1970). Эти разные роли, а также необычайное разнообразие организмов, колонизирующих млекопитающих, бактерии, вирусы, грибы и червей разного рода, а также разнообразие мест колонизации предъявляют различные и потенциально противоречивые требования к иммунной защите, явление, называемое антагонистической плейотропией.Привлекательной гипотезой иммунопатологии, известной как аллергия, является чрезмерная реакция тех элементов иммунной системы, которые в менее древние времена в противном случае были бы заняты борьбой с гельминтными инфекциями (Wilson and Maizels, 2004).
самая важная причина.Мы предполагаем, что основная причина, по которой эволюция млекопитающих не замедлилась и, возможно, не может сдержать чрезмерную реакцию иммунной системы на бактериальные и другие инфекции, — это функциональные ограничения, которые ограничивают степень, в которой иммунная система может быть изменена. Иммунная система не только устраняет бактериальные инфекции, но и выполняет другие функции. Было высказано предположение, что эти другие роли доминировали в эволюции иммунной системы млекопитающих (Burnet, 1970). Эти разные роли, а также необычайное разнообразие организмов, колонизирующих млекопитающих, бактерии, вирусы, грибы и червей разного рода, а также разнообразие мест колонизации предъявляют различные и потенциально противоречивые требования к иммунной защите, явление, называемое антагонистической плейотропией.Привлекательной гипотезой иммунопатологии, известной как аллергия, является чрезмерная реакция тех элементов иммунной системы, которые в менее древние времена в противном случае были бы заняты борьбой с гельминтными инфекциями (Wilson and Maizels, 2004).
Существует тонкая грань между ответом (1–6 дюймов) и избыточным ответом (7 на этом рисунке), что может быть трудно для систем, регулирующих иммунный ответ, воспринимать, не говоря уже о том, чтобы избежать. Как предположил Франк (Андре и др., 2004), интенсивность иммунного ответа может определяться компромиссом между увеличением силы и скорости иммунной защиты и вирулентностью из-за чрезмерного отклика иммунной системы.
Есть ли доказательства в поддержку этих двух гипотез, почему эволюция не устранила вирулентность, возникающую в результате чрезмерного иммунного ответа? Немного — по крайней мере, пока. Однако мы предполагаем, что часть значительной наследственной изменчивости восприимчивости к инфекционным заболеваниям в популяциях людей (Bellamy and Hill, 1998; Bellamy et al., 2000; Segal and Hill, 2003; Sorensen et al., 1988) можно интерпретировать как поддержку этих гипотез. Безусловно, есть убедительные и даже неопровержимые доказательства того, что некоторые из этих вариаций поддерживаются опосредованным заболеванием балансированием или частотно-зависимым отбором, но это не относится ко всем или даже к большей части. Мы предполагаем, что большая часть устойчивых генетических вариаций восприимчивости к болезням в человеческих популяциях является отражением близорукости и ограничений естественного отбора: (i) относительной слабости отбора для модулирования иммунной избыточной реакции и (ii) даже в большей степени, бессилия естественный отбор из-за ограничений иммунной системы — антагонистическая плейотропия. Генетическая изменчивость, которая не воспринимается или плохо воспринимается естественным отбором, будет накапливаться и сохраняться (Crow and Kimura, 1971).
Мы предполагаем, что большая часть устойчивых генетических вариаций восприимчивости к болезням в человеческих популяциях является отражением близорукости и ограничений естественного отбора: (i) относительной слабости отбора для модулирования иммунной избыточной реакции и (ii) даже в большей степени, бессилия естественный отбор из-за ограничений иммунной системы — антагонистическая плейотропия. Генетическая изменчивость, которая не воспринимается или плохо воспринимается естественным отбором, будет накапливаться и сохраняться (Crow and Kimura, 1971).
Последствия
В то время как заболеваемость и смертность от большинства бактериальных инфекций можно объяснить чрезмерной реакцией иммунной системы, практически все наши усилия по лечению этих инфекций направлены на контроль распространения и уничтожение бактерий, в первую очередь с помощью антибиотиков.Этот подход был и остается эффективным, но не полностью. Лечение антибиотиками обычно не дает результатов, и пациенты умирают или остаются больными в течение длительного времени. Устойчивость патогена к антибиотикам, используемым для лечения, является лишь одной из причин этого неэффективности, а для некоторых инфекций не является основной, по крайней мере, пока (Levin and Rozen, 2006; Yu et al., 2003).
Устойчивость патогена к антибиотикам, используемым для лечения, является лишь одной из причин этого неэффективности, а для некоторых инфекций не является основной, по крайней мере, пока (Levin and Rozen, 2006; Yu et al., 2003).
Очевидным альтернативным подходом к лечению инфекций является снижение заболеваемости и предотвращение смертности за счет модуляции избыточной реакции иммунной системы.Это были попытки сделать именно это для лечения бактериального сепсиса. Клинические испытания оценивали использование глюкокортикоидов (Bone et al., 1987), препаратов, предназначенных для нейтрализации эндотоксинов (Ziegler et al., 1991), фактора некроза опухоли α (Fisher et al., 1996) и IL-1. β (Fisher et al., 1994), но ни один из этих методов лечения не был эффективным. На сегодняшний день наиболее успешные испытания на людях были с компонентом естественной антикоагулянтной системы, активированным протеином С, который обладает значительными противовоспалительными свойствами, а также является мощным антикоагулянтом (уменьшает образование сгустков, которые ответственны за органную недостаточность на поздних этапах). стадии сепсиса) (Fourrier, 2004).Кроме того, новые агенты перенаправляют иммунный ответ и обещают стать эффективным средством лечения сепсиса в будущем, такие как IL-12 (O’Suilleabhain et al., 1996) и антитела против комплемента (C5a) (Czermak et al., 1999). Однако понимание специфики иммунной сверхреакции и сложности механизмов обратной связи, которые контролируют иммунный ответ, необходимо для лечения, направленного на усиление или подавление иммунного ответа пациента.
стадии сепсиса) (Fourrier, 2004).Кроме того, новые агенты перенаправляют иммунный ответ и обещают стать эффективным средством лечения сепсиса в будущем, такие как IL-12 (O’Suilleabhain et al., 1996) и антитела против комплемента (C5a) (Czermak et al., 1999). Однако понимание специфики иммунной сверхреакции и сложности механизмов обратной связи, которые контролируют иммунный ответ, необходимо для лечения, направленного на усиление или подавление иммунного ответа пациента.
В настоящее время, в целом, успех этих иммуномодулирующих методов в предотвращении заболеваемости и смертности от бактериальных инфекций можно в лучшем случае охарактеризовать как скромный.Однако, поддерживая спекулятивный характер этой разглагольствования и желая получить оптимистичный вывод, мы предполагаем, что по мере того, как мы узнаем больше о регуляции иммунного ответа и разработаем процедуры для мониторинга, а также введения регуляторных иммунных молекул в режиме реального времени, эти методы будут становятся все более эффективными для лечения бактериальной инфекции.
Благодарности
Мы благодарим Элизабет Карниэль за то, что она поделилась своими идеями об эволюции вирулентности Y.pestis . Мы благодарны Джиму Буллу и Харрису Финбергу за ценные комментарии и предложения. Б. Р. Л. выражает свою постоянную благодарность Фернандо Бакеро за вдохновение, идеи, нескончаемые прихоти, поддержку и дружбу. Это начинание было поддержано грантом от NIH, AI40662 (B. R. L.), и грантом NIH на обучение (E. M.).
Эволюция бактерий: путь к сопротивлению
Вспомните, когда вы или ваш близкий в последний раз болели бактериальной инфекцией и нуждались в антибиотиках.Что, если антибиотики не подействовали? К сожалению, это становится все более распространенным во всем мире, вызывая не менее 700 000 смертей в год (O’Neill, 2016). Устойчивость к противомикробным препаратам, также известная как УПП, быстро развивается, часто в течение одной инфекции. Это происходит либо в результате обмена бактериями друг с другом генами, обеспечивающими устойчивость, либо в результате того, что отдельная бактерия становится устойчивой в результате мутаций в своих собственных генах. Устойчивые бактерии с большей вероятностью переживут лечение противомикробными препаратами и распространят инфекцию среди других людей.
Устойчивые бактерии с большей вероятностью переживут лечение противомикробными препаратами и распространят инфекцию среди других людей.
Во время инфекций бактерии имеют тенденцию расти на поверхностях в сообществах, называемых биопленками (Koo et al., 2017). Биопленки способствуют общению и сотрудничеству, а также физически защищают бактерии от антимикробных препаратов и иммунной системы хозяина. Эти свойства затрудняют лечение биопленочных инфекций даже при отсутствии УПП. Было высказано предположение, что бактерии, растущие в биопленках, могут быстрее развить УПП, что затрудняет прогнозирование результатов лечения. Однако мало исследований изучали, как различные способы роста влияют на способность бактерий развиваться (Steenackers et al., 2016).
Теперь в eLife Вон Купер и его коллеги из Университета Питтсбурга, в том числе Альфонсо Сантос-Лопес и Кристофер Маршалл в качестве первых авторов, сообщают, как «бактериальный образ жизни» влияет на эволюцию устойчивости к антибиотику ципрофлоксацину (CIP) в патоген Acinetobacter baumannii (Santos-Lopez et al. , 2019). Для этого они сравнили бактерии, которые плавали в жидкой культуре, когда они росли, с бактериями, которые росли в виде биопленок.Бактерии, растущие в жидкой культуре, были экспериментально эволюционированы путем ежедневного переноса небольших количеств культуры в новый контейнер со свежей питательной средой (рис. 1А). Биопленки были созданы путем выращивания A. baumannii на пластиковых шариках, погруженных в жидкую культуру, и ежедневного переноса этих покрытых биопленкой шариков в новый контейнер со свежей средой и чистым шариком (Рисунок 1B). В начале эксперимента клетки подвергались воздействию очень низкой дозы CIP, а затем доза CIP удваивалась каждые 72 часа по мере развития устойчивости в течение 12-дневного периода.
Экспериментальная эволюция устойчивости к ципрофлоксацину (CIP) у бактерий, выращенных в жидкости и на биопленках.
Чтобы понять, как образ жизни бактерий влияет на эволюцию устойчивости к противомикробным препаратам, штаммы бактерии A. baumannii были экспериментально выведены либо в жидкой культуре ( A ), либо в виде биолфимов на пластиковых шариках ( B ).Сантос-Лопес и др. обнаружили, что у штаммов A.baumannii , выращенных в жидкости и на биопленке, устойчивость к противомикробному CIP развивалась по-разному: бактерии, выращенные в жидкости, развили одну устойчивую линию, которая доминировала в популяции ( C ), тогда как бактерии, выращенные на биопленке развились множественные устойчивые линии, сосуществующие в популяции ( D ). Другие важные характеристики двух эволюционировавших штаммов (такие как устойчивость к CIP) обсуждаются в основном тексте.
baumannii были экспериментально выведены либо в жидкой культуре ( A ), либо в виде биолфимов на пластиковых шариках ( B ).Сантос-Лопес и др. обнаружили, что у штаммов A.baumannii , выращенных в жидкости и на биопленке, устойчивость к противомикробному CIP развивалась по-разному: бактерии, выращенные в жидкости, развили одну устойчивую линию, которая доминировала в популяции ( C ), тогда как бактерии, выращенные на биопленке развились множественные устойчивые линии, сосуществующие в популяции ( D ). Другие важные характеристики двух эволюционировавших штаммов (такие как устойчивость к CIP) обсуждаются в основном тексте.
Образцы бактерий были собраны из каждой культуры в разное время во время эксперимента, а также в конце эксперимента. Полногеномное секвенирование этих образцов показало, что выращенный в жидкой среде и выращенный на биопленке A. baumannii идет разными, но повторяемыми путями в ходе экспериментальной эволюции. Для бактерий, выращенных в жидкой культуре, Santos-Lopez et al. обнаружили, что наиболее устойчивая линия CIP превосходит другие бактериальные линии, что позволяет ее потомству быстро захватить популяцию (рис. 1C).Однако для бактерий, выращенных в биопленках, исследователи обнаружили, что конкуренция между клонами была меньше — предположительно потому, что они не так свободно перемещаются, как бактерии, выращенные в жидкости, — и что несколько сосуществующих линий смогли появиться (рис. 1D). ).
Для бактерий, выращенных в жидкой культуре, Santos-Lopez et al. обнаружили, что наиболее устойчивая линия CIP превосходит другие бактериальные линии, что позволяет ее потомству быстро захватить популяцию (рис. 1C).Однако для бактерий, выращенных в биопленках, исследователи обнаружили, что конкуренция между клонами была меньше — предположительно потому, что они не так свободно перемещаются, как бактерии, выращенные в жидкости, — и что несколько сосуществующих линий смогли появиться (рис. 1D). ).
Дальнейшие эксперименты показали, что бактерии, выращенные в жидкости и на биопленке, выработали уникальные стратегии противодействия CIP, что привело к разной степени устойчивости к CIP. Бактерии, выращенные в жидкости, развили устойчивость к CIP на низком уровне за счет мутаций, которые увеличили количество белков-помп, которые транспортируют CIP из клеток.Эти устойчивые клетки быстро взяли верх, а затем приобрели дополнительные мутации, которые препятствовали связыванию CIP с его белками-мишенями. Бактерии с обоими типами мутаций могли противостоять дозам CIP, которые были намного выше, чем те, которые использовались во время эволюционного эксперимента. Бактерии, выращенные на биопленке, также развили низкий уровень устойчивости к CIP за счет увеличения продукции помпового белка, но использовали другие насосы, чем их жидкие аналоги. Однако бактерии, выращенные на биопленке, не смогли развить высокий уровень устойчивости за счет дополнительных мутаций.Избирательное давление, создаваемое различными образами жизни, похоже, взаимодействует с давлением воздействия CIP и ограничивает эволюционные пути и, следовательно, результаты, доступные для каждого образа жизни.
Бактерии с обоими типами мутаций могли противостоять дозам CIP, которые были намного выше, чем те, которые использовались во время эволюционного эксперимента. Бактерии, выращенные на биопленке, также развили низкий уровень устойчивости к CIP за счет увеличения продукции помпового белка, но использовали другие насосы, чем их жидкие аналоги. Однако бактерии, выращенные на биопленке, не смогли развить высокий уровень устойчивости за счет дополнительных мутаций.Избирательное давление, создаваемое различными образами жизни, похоже, взаимодействует с давлением воздействия CIP и ограничивает эволюционные пути и, следовательно, результаты, доступные для каждого образа жизни.
Сантос-Лопес и др. обнаружили, что в отсутствие CIP устойчивые к CIP бактерии, образовавшиеся из жидкости, были менее приспособлены, чем штамм-предок, и их можно было легко победить. Примечательно, что о подобных компромиссах в фитнесе сообщалось и раньше, и считается, что снижение фитнеса замедляет распространение бактерий AMR. Вызывает тревогу то, что устойчивые к CIP штаммы, образовавшиеся из биопленок, были, по крайней мере, такими же пригодными, как и их предки, при выращивании без CIP. В то время как эволюционирующий из жидкости штамм часто становился устойчивым к дополнительным противомикробным препаратам, бактерии, выросшие на биопленке, обычно проявляли повышенную чувствительность по крайней мере к одному другому противомикробному лечению. Это означает, что, хотя штаммы, образовавшиеся из биопленок, могут быть более приспособленными и с большей вероятностью распространяться среди новых пациентов, их также может быть легче убить с помощью других противомикробных препаратов.
Вызывает тревогу то, что устойчивые к CIP штаммы, образовавшиеся из биопленок, были, по крайней мере, такими же пригодными, как и их предки, при выращивании без CIP. В то время как эволюционирующий из жидкости штамм часто становился устойчивым к дополнительным противомикробным препаратам, бактерии, выросшие на биопленке, обычно проявляли повышенную чувствительность по крайней мере к одному другому противомикробному лечению. Это означает, что, хотя штаммы, образовавшиеся из биопленок, могут быть более приспособленными и с большей вероятностью распространяться среди новых пациентов, их также может быть легче убить с помощью других противомикробных препаратов.
В этой работе Santos-Lopez et al.сосредоточились на том, как биопленки влияют на конкуренцию и давление отбора в процессе эволюции. Однако биопленки могут влиять на эволюцию другими способами, а бактериальный образ жизни — лишь один из многих факторов, определяющих эволюцию УПП «в дикой природе». Например, многие виды бактерий испытывают повышенное совместное использование генов и частоту мутаций при формировании биопленки, что также может способствовать развитию УПП (Steenackers et al. , 2016). Кроме того, развитие УПП может быть вызвано увеличением частоты мутаций, вызванных противомикробными препаратами (Cirz et al., 2005; Кохански и др., 2010; Gutierrez et al., 2013; Pribis et al., 2019) и другие факторы стресса, такие как голод, с которыми сталкиваются бактерии во время инфекции (Fitzgerald and Rosenberg, 2019).
, 2016). Кроме того, развитие УПП может быть вызвано увеличением частоты мутаций, вызванных противомикробными препаратами (Cirz et al., 2005; Кохански и др., 2010; Gutierrez et al., 2013; Pribis et al., 2019) и другие факторы стресса, такие как голод, с которыми сталкиваются бактерии во время инфекции (Fitzgerald and Rosenberg, 2019).
Эти результаты имеют важное значение для лечения A. baumannii, , который уже устойчив ко многим лекарственным препаратам и считается критической угрозой как Всемирной организацией здравоохранения, так и Центрами по контролю и профилактике заболеваний в США. Лучшее понимание переменных, формирующих эволюцию УПП, улучшит прогнозы эволюционных исходов, меры по контролю над противомикробными препаратами и клинические исходы для отдельных пациентов.
Эволюция и развитие (Стэнфордская энциклопедия философии)
1. Эволюция и развитие в историческом контексте
Первоначально концепции эволюции и развития были тесно связаны между собой. связанный. Фактически, с конца 17 -го века
понятие «эволюция» широко использовалось для описания
процессы индивидуального развития, и
гипотезы часто относились к тому, что сейчас называется эволюцией. В
кроме того, развитие (Entwicklung) часто считалось
не только для описания онтогенетических изменений организмов (Goethe 1790;
Debraw 1777), но также (что мы рассматриваем сегодня) филогенетические изменения.Например, Фридрих Шеллинг (1798) утверждает, что последовательность
стадий всех органических существ сформировались через постепенное
развитие («Entwicklung») той же организации. Этот
ситуация изменилась в начале 19 -го века, когда
эволюция использовалась некоторыми авторами для обозначения трансгенеративных
трансформации организмов, в том числе Чарльзом Лайелем и Георгом
W.F. Гегель. Однако в своем первом издании Происхождение
Вид (1859) Дарвин не использовал термин эволюция, скорее всего
потому что он хотел отделить свою теорию от более ранних
понимания слова.
связанный. Фактически, с конца 17 -го века
понятие «эволюция» широко использовалось для описания
процессы индивидуального развития, и
гипотезы часто относились к тому, что сейчас называется эволюцией. В
кроме того, развитие (Entwicklung) часто считалось
не только для описания онтогенетических изменений организмов (Goethe 1790;
Debraw 1777), но также (что мы рассматриваем сегодня) филогенетические изменения.Например, Фридрих Шеллинг (1798) утверждает, что последовательность
стадий всех органических существ сформировались через постепенное
развитие («Entwicklung») той же организации. Этот
ситуация изменилась в начале 19 -го века, когда
эволюция использовалась некоторыми авторами для обозначения трансгенеративных
трансформации организмов, в том числе Чарльзом Лайелем и Георгом
W.F. Гегель. Однако в своем первом издании Происхождение
Вид (1859) Дарвин не использовал термин эволюция, скорее всего
потому что он хотел отделить свою теорию от более ранних
понимания слова. Вместо этого он говорил о «происхождении с
модификации ». Но особенно благодаря работам Герберта
Спенсера, эволюция утвердилась как концепция, касающаяся
«Трансформация видов» и теория Дарвина
трансформация посредством отбора все чаще рассматривалась как образец
«теории эволюции» (Spencer 1852 [1891], 1).
Согласно этой теории, эволюция осуществляется через организмы.
«Борьба за существование» (Дарвин) и «выживание»
из сильнейших »(Спенсер).
Вместо этого он говорил о «происхождении с
модификации ». Но особенно благодаря работам Герберта
Спенсера, эволюция утвердилась как концепция, касающаяся
«Трансформация видов» и теория Дарвина
трансформация посредством отбора все чаще рассматривалась как образец
«теории эволюции» (Spencer 1852 [1891], 1).
Согласно этой теории, эволюция осуществляется через организмы.
«Борьба за существование» (Дарвин) и «выживание»
из сильнейших »(Спенсер).
Однако, помимо этого увеличивающегося концептуального расставания, эволюция и развитие рассматривались как тесно взаимосвязанные процессы многие ученые на протяжении 19 -х годов века.Основываясь на более ранние сравнительные исследования эмбриональных процессов в различных таксонах (среди других Карл Э. фон Бэр (1827) и Фриц Мюллер (1864)), Эрнст Геккель (1872) утверждал, что все многоклеточные животные имеют одинаковые ранние стадии развития (см. Gould 1977). Он понимал эти паттерны развития как модификации основного типа. В соответствии с этот биогенетический закон, развитие особей является резюме эволюционных изменений видов:
В ходе быстрого и непродолжительного индивидуального развития органическая особь […] повторяет важнейшие изменения форма, через которую прошли его предки в течение долгого и медленного ход палеонтологического развития, следуя законам наследственности и адаптация.
 (Геккель 1866 г., II 300)
(Геккель 1866 г., II 300) Эта взаимосвязь, при которой биоразнообразие постепенно возникло из
изменения в развитии, которые уже предвидел Иоганн Ф. Меккель
(1812) и Ричард Оуэн (1837), которые распараллелили
«Трансмутации» эмбриональных форм и трансмутации
разновидность. Также Дарвин (1837) описывает генезис индивидов как
«Укороченное повторение» морфологических изменений во время
филогенез. С конца 19 -го века этот вид на
развитие как резюме эволюции стало проблематичным (см.
Расмуссен 1991).Во-первых, за это время вид линейного
эволюция от беспозвоночных до позвоночных к человеку была
все чаще подвергаются сомнению. Во-вторых, на основе сравнительных исследований клеточного
В 1890-х годах ученые критиковали то, что
эволюционируют не только взрослые организмы, но и пути их развития.
Например, в 1898 году эмбриолог Эдмунд Б. Уилсон (1898) использовал
гомологичные паттерны деления эмбриональных клеток для объединения таксонов, в то время как
эмбриолог Франк Р. Лилли (1898) изучал различия в этих
шаблоны, чтобы показать истоки эволюционной адаптации.Эти
исследования (по крайней мере, до некоторой степени) противоречили теории Геккеля.
рекапитуляционизм, поскольку они видели онтогенез недостаточно объясненным
историческими причинами, а скорее нуждаются в независимой механистической
законы, влияющие на физические свойства онтогенетического материала (см.
Гуралник 2002). Кроме того, это исследование клеточного происхождения показало, что
что закон Геккеля о взаимосвязи между развитием и
эволюция должна быть инвертирована, поскольку клеточные клоны эволюционируют
эволюционное время. В более общем плане это означает, что разработка
не просто имитирует эволюцию.Скорее, это источник вариаций.
и, следовательно, причина эволюции и адаптации: «онтогенез не
резюмирует филогенез, он его создает »(Garstang 1922: 81). В
Короче говоря, эволюция происходит через модификации развития.
Лилли (1898) изучал различия в этих
шаблоны, чтобы показать истоки эволюционной адаптации.Эти
исследования (по крайней мере, до некоторой степени) противоречили теории Геккеля.
рекапитуляционизм, поскольку они видели онтогенез недостаточно объясненным
историческими причинами, а скорее нуждаются в независимой механистической
законы, влияющие на физические свойства онтогенетического материала (см.
Гуралник 2002). Кроме того, это исследование клеточного происхождения показало, что
что закон Геккеля о взаимосвязи между развитием и
эволюция должна быть инвертирована, поскольку клеточные клоны эволюционируют
эволюционное время. В более общем плане это означает, что разработка
не просто имитирует эволюцию.Скорее, это источник вариаций.
и, следовательно, причина эволюции и адаптации: «онтогенез не
резюмирует филогенез, он его создает »(Garstang 1922: 81). В
Короче говоря, эволюция происходит через модификации развития.
Еще одна характерная черта 19 -х годов века (и раннего
20 (-е гг. века) биология имела отсутствие четкой концептуальной
разрыв между процессами развития и размножением. Особенно
идея наследования приобретенных характеристик была частью
мейнстрим эволюционной мысли (Bowler 1983, 2017; Gissis
И Яблонька 2011).По словам Жана Батиста Ламарка (1809 г.),
изменения в структуре или функции черт, которые произошли во время
жизнь организма, в зависимости от их использования или неиспользования, может быть
по наследству. Дарвин принял эту идею в своей теории наследования
пангенезис (см. Holterhoff 2014). Он предположил, что все части тела, на
на каждой стадии развития содержат мелкие частицы, так называемые
«Геммулы», чувствительные к изменениям окружающей среды.
Эти частицы накапливаются в репродуктивных органах и
передается следующему поколению.Таким образом, геммулы модифицировались во время
развитие привело к изменениям в следующем поколении. Дарвин
заявил, что из-за этой тесной связи развития и
воспроизводства, «наследование следует рассматривать как просто форму
рост »(Дарвин 1868: II, 404). Однако эта связь между
развитие и наследование все чаще подвергались критике в первые
половина 20-го -го века. Вслед за Августом Вейсманном (1892 г.)
наследование вызванных окружающей средой изменчивости соматических клеток
(тело) организмов все чаще подвергалось сомнению.В
организмы, размножающиеся половым путем, для сперматозоидов используется только зародышевая плазма.
и яйца были признаны несущими информацию, которая передается
новое поколение. Согласно этой точке зрения, у зародышевой линии был иммунитет.
от изменений, происходящих в соматических клетках тела, и, таким образом,
наследование приобретенных характеристик или другие теории
плазматическое наследование были отвергнуты (но см. Harwood 1993, Gilbert
И Эпель 2015).
века) биология имела отсутствие четкой концептуальной
разрыв между процессами развития и размножением. Особенно
идея наследования приобретенных характеристик была частью
мейнстрим эволюционной мысли (Bowler 1983, 2017; Gissis
И Яблонька 2011).По словам Жана Батиста Ламарка (1809 г.),
изменения в структуре или функции черт, которые произошли во время
жизнь организма, в зависимости от их использования или неиспользования, может быть
по наследству. Дарвин принял эту идею в своей теории наследования
пангенезис (см. Holterhoff 2014). Он предположил, что все части тела, на
на каждой стадии развития содержат мелкие частицы, так называемые
«Геммулы», чувствительные к изменениям окружающей среды.
Эти частицы накапливаются в репродуктивных органах и
передается следующему поколению.Таким образом, геммулы модифицировались во время
развитие привело к изменениям в следующем поколении. Дарвин
заявил, что из-за этой тесной связи развития и
воспроизводства, «наследование следует рассматривать как просто форму
рост »(Дарвин 1868: II, 404). Однако эта связь между
развитие и наследование все чаще подвергались критике в первые
половина 20-го -го века. Вслед за Августом Вейсманном (1892 г.)
наследование вызванных окружающей средой изменчивости соматических клеток
(тело) организмов все чаще подвергалось сомнению.В
организмы, размножающиеся половым путем, для сперматозоидов используется только зародышевая плазма.
и яйца были признаны несущими информацию, которая передается
новое поколение. Согласно этой точке зрения, у зародышевой линии был иммунитет.
от изменений, происходящих в соматических клетках тела, и, таким образом,
наследование приобретенных характеристик или другие теории
плазматическое наследование были отвергнуты (но см. Harwood 1993, Gilbert
И Эпель 2015).
Это разделение между развитием, с одной стороны, и наследованием, и эволюции, с другой стороны, через теорию Вейсмана наследования, оказали долгосрочное влияние на рассуждения биологов о эволюция в последующие годы.Несмотря на продолжающиеся усилия неоламаркианцы вплоть до 1930-х годов (Bowler, 1983), развитие все больше исчезал с ландшафта эволюционной теории. От объединение законов Менделя и новых открытий в генетике в статистические основы популяционной генетики, современный синтез пришли к пониманию эволюции как изменения частот разные аллели в популяции (Фишер, 1930; Добжанский, 1951). Изменения, относящиеся к эволюции, производились только в генах зародышевая линия. Эти случайные изменения, мутации, были исключены из история развития личности.В 1940-х и 50-х годах это точка зрения все более и более проявлялась, не в последнюю очередь через так называемые «Центральная догма» молекулярной биологии (Crick 1958). Это утверждает (аналогично теории Вейсмана), что информационные потоки всегда от ДНК к белкам, и никогда наоборот. Таким образом, фенотипический изменения во время развития не могли повлиять на гены. Тенденция к связывать гены с популяциями в эволюционных объяснениях, а не развитие с эволюцией достигло своего апогея с появлением гена взгляд на эволюцию глазами (Williams 1966; Dawkins 1976).Сосредоточив внимание на вопрос, каковы могут быть определенные черты характера, такие как альтруистическое поведение полезный, он не только идентифицировал гены (а не организмы) как единственные единицы выбора. Кроме того, он описал развитие как ничего, кроме считывания генетической программы (Maienschein 2003; Пильуччи 2010). Гены контролируют развитие признаков и поведения организмов, и они копируются, чтобы защитить свои собственные дальнейшее размножение группами и популяциями.
В течение двадцатого века эти разрывы в развитии и эволюция привела к таким заявлениям, как: «Проблемы, связанные с упорядоченное развитие личности не связано с эволюция организмов во времени »(Wallace 1986: 149).Однако такие заявления также подверглись критике со стороны ряда критиков. Например, исключение биологии развития из современного синтез (Harrison 1937; De Beer 1954; Waddington 1957; Hamburger 1980), объяснительная автократия адаптационистской программы в эволюционной биологии (Gould & Lewontin 1979) и упущение удовлетворительная теория эволюционной новизны, в которой развитие сыграли бы роль (Goldschmidt 1940), подверглись критике. Не в последнюю очередь из-за этого отказа от перспектив развития эволюционная теория была обвинена в ограничении направления исследований в эволюционная биология (Provine 1989).Фактически, на протяжении двадцатого века был выдвинут ряд теорий, которые утверждают, что из-за частичной независимости генетической и фенотипической изменчивости, эволюционные исследования должны уделять больше внимания тому, как изменения в паттерны развития и поведения могут влиять на эволюционное изменять. Сюда входят случаи «канализованного» развития пути, которые сохраняются, даже если генотип или окружающая среда могла измениться до некоторой степени (Waddington 1957; см. также Nijhout 2002; Гилберт и Эпель, 2015 г .; Султан 2015).Кроме того, исследования фенотипической пластичности показали, что признак организма может по-разному реагировать на воздействие окружающей среды, и что геном кодирует широкий спектр потенциальных фенотипов (Waddington 1942; Nijhout 1990; Пильуччи 2001). Эволюционная значимость эти результаты были подчеркнуты исследованиями, изучающими, как диапазон вариации меняется от пластического признака к фиксированному или канал канализировал на протяжении поколений (Suzuki and Nijhout 2008; Вест-Эберхард 2003; Б.Baker et al. 2019). Более того, согласно так называемый «эффект Болдуина», изученный поведенческий паттерны (например, акклиматизация к новому стрессору), которые были изначально довольно пластичен, может повлиять на репродуктивный успех людей и, таким образом, из поколения в поколение и через естественные отбора, постепенно включаться в генетические и эпигенетические состав вида (Baldwin 1896; Simpson 1953; Piaget 1976 [1978]; Ньюман 2002). Хотя некоторые из этих перспектив развития эволюции соответствовали популяционно-генетической структуре эволюционной биологии и фактически постепенно вошли в эволюционной мысли, другие ставили перед теоретическая интеграция.
Основываясь на последнем, более проблематичном наборе подходов (особенно в последние годы) эволюционная теория столкнулась с звонки ориентированных на развитие биологов и философов биологии, чтобы расширить стандартные пояснительные и методологические подходы к эволюции (Bonner 1958; Alberch 1982; Bonner 1982; Raff & Кауфман 1983; Гилберт, Опиц и Рафф, 1996; Schlichting & Pigliucci 1998; Вест-Эберхард 2003; Г. Мюллер 2007; Пильуччи & Мюллер 2010; Бейтсон и Глюкман, 2011 г .; Яблонька и Баранина 2014; Лаланд и др.2014, 2015; Гилберт и Эпель, 2015 г .; Г. Мюллер 2017; для обсуждения см. Huneman И Уолш, 2017; Фабрегас-Техеда и Вергара-Силва 2018; Baedke et al. 2020b). Они утверждают, что эволюционные изменения не должны в первую очередь должны быть исследованы и объяснены как изменение генотипа частоты в популяциях, но (также) на уровне развивающихся и действующее лицо. Основная идея заключается в том, что фенотипический вариативность и гибкость реакции организмов на сигналы окружающей среды могут вносить неслучайные вариации и, таким образом, могут предвзятость и / или прямая морфологическая эволюция до некоторой степени.Этот включает не только экологические изменения в нормативных процессы, но также и физические ограничения развивающегося эмбриона. Сосредоточившись на этих явлениях, Кевин Лаланд и его коллеги (2014: 161) утверждают: «Альтернативное видение эволюции начинает проявляться. кристаллизуются, в которых процессы, с помощью которых организмы растут и развитие признаются причинами эволюции ». У других есть утверждал, что гены, вероятно, чаще следуют за эволюцией, чем лидеры (Вест-Эберхард 2003, 2005). Другими словами, экологически чуткий, развивающийся и действующий организм принимает Свинец.Он привносит неслучайным образом новые фенотипы в популяции, которые впоследствии стабилизируются генами.
Это «взгляд на разработку в первую очередь» (или иногда его называют «Взгляд на пластичность в первую очередь») эволюции в настоящее время принят особенно исследователями в трех областях исследований: эволюционных биология развития (эво-дево), эпигенетика и конструирование ниш теория. Evo-DevO изучает эволюцию путей развития и многое другое. важно для вышеприведенного взгляда, как ограничения и предубеждения развития может влиять на эволюционные траектории (Raff 1996, 2000; Gerhart & Киршнер 1997; Любовь 2003; Amundson 2005; Лаубихлер и Майеншайн 2007; Сансом и Брэндон 2007; Г.Мюллер 2007; Минелли 2009; Laubichler 2010; Пильуччи и Мюллер 2010; Гилберт и Эпель 2015; Moczek et al. 2015; Levis & Pfennig 2020). Это включает в себя спрашивая, как изменения в регуляторных сетях генов вызывают модификации процессы развития (связанные с основными сдвигами в морфологическом «Планы тела») и, таким образом, создают эволюционные новшества, или какими возможностями обладают организмы для создания наследуемых, адаптивных фенотипической изменчивости и, следовательно, эволюционировать в процессе эволюции (т. е. их эволюционируемость; см. Hendrikse et al.2007). Это фокус на том, как фенотипические создается вариация, а не то, как она выбирается, поддерживается исследования в области эпигенетики межпоколенческих и межпоколенческих эпигенетических наследование (см. запись системы наследования), как через передачу регуляторных факторов активности генов (Яблонка и Лэмб 1989, 2014, Янлонка и Раз 2009, Перес и Ленер 2019). Другие формы внегенетического наследования включают передача микробиоты (Gilbert et al.2012, 2014, Browne et al. 2017) и поведенчески опосредованные родительские эффекты (Kappeler & Meaney 2010; Rilling & Young 2014).Этот набор процессов содержит экологически чувствительные негенетические источники вариаций для организмы, которые могут передаваться из поколения в поколение, во многих случаях не связаны с передачей генетической информации. Наконец, ниша теория строительства подчеркивает самовоспроизводящийся и взаимный эффекты организмов как агентов, которые создают свою собственную нишу (и / или других видов) во время развития (Lewontin 1982; Стерельный 2001; Day et al. 2003; Odling-Smee et al. 2003; Лаланд и др. 2008, 2009, 2011; Odling-Smee 2010; Чиу и Гилберт 2015, 2020; Aaby & Ramsey готовится к печати).Эти недавние исследования эволюции развития будут обсуждаться в подробно в разделе 3.
Различные разработки в вышеуказанных областях привели к увеличению философский интерес к возникающим концептуальным и объяснительным вопросам с нового стыка развития и эволюции. К ним относятся выяснение, какие причинно-следственные связи и границы существуют между две области, в которых разные роли играют процессы развития. недавние эволюционные исследования, и как эти новые роли влияют на онтологические рамки организма.Кроме того, эти развития сопровождались дебатами о структуре объяснения эволюции развития. Наконец, они сообщили дискуссии по антропологическому пониманию и этическим вопросам относительно людей. Теперь мы обсудим дебаты по этим вопросам в деталь.
2. Концептуальные разделения и объединения эволюции и развития
Одна из центральных тем ранней философии биологии в 1960-е и 1970-е годы были попыткой разработать подходящую концептуальную фреймворк, который поддерживал бы разделение между разработкой и эволюция в соответствии с центральным предположением современного синтеза что эволюция — это изменение генетического состава популяций только (Добжанский 1951: 16; см. также Charlesworth et al.2017). Этот означает, как следствие, что развитие не существенные пути) причинно влияют на эволюцию. На протяжении десятилетий это предположение было поддержано исторически влиятельными концептуальное различие между непосредственными причинами и конечными причинами (Майр, 1961).
2.1 Ближайшее и высшее различие
Двойная структура «приблизительное против окончательного» обеспечивает качественное различие биологической причинности (для родственных различия, см. J. Baker 1938; Тинберген 1951, 1963).Он считает, что биологи, изучающие непосредственные причины, задают вопросы о индивидуальные процессы развития. Таким образом, функциональные биологи заинтересованные в таких ближайших причинах изучения, как работают системы. Вместо, эволюционные биологи, изучающие первопричины, задаются вопросом, почему например, почему филогенез произвел определенные эволюционные функции. Согласно этому различию, по крайней мере на поверхности, ближайший причины напоминают аристотелевские действенные причины, в то время как первопричины напоминают аристотелевские конечные причины.Чтобы проиллюстрировать это различие, Майр (1961) приводит пример миграции птиц. Миграция может быть изучили, задавая вопросы о том, как птицы мигрируют (т.е. как они развивают такие навыки, как навигации) или почему они мигрируют (т. е. из-за каких выборочных преимущество). Под этими двумя исследованиями понимаются оба важны и дополняют друг друга. Однако к ним следует относиться как к отличны друг от друга.
Ближайшее-окончательное различие может быть дано эпистемическим или онтологическое чтение. Во-первых, авторы интерпретировали это как различая разные виды объяснений (Amundson 2005; Calcott 2013; Scholl & Pigliucci 2015).Это эпистемическое прочтение включает что как вопросы не могут быть решены с помощью объяснений, цитирующих окончательные причины (то есть рассказ об адаптации) и вопросы, почему не могут быть рассмотрены с помощью объяснений с указанием ближайших причин (т. е. рассказывая историю развития черты характера). Во-вторых, авторы интерпретировали это различие как различие между разными онтологическими классы причин, работающих в онтогенетических и филогенетических процессах (Лаланд и др., 2013a). Это онтологическое прочтение подкреплено теоретически в соответствии с концепцией Вейсмана о отделении зародыша линия и сома, которая обеспечивает демаркационную линию между двумя различными классы причин.По сей день биологи и философы не достигли консенсуса относительно того, как именно разделение Должно быть «ближайший-окончательный» или «как-почему» понимается эпистемически или онтологически (Francis 1990; Dewsbury 1992, 1999; Стерельный 1992; Битти 1994; Ariew 2003). Несмотря на это отсутствие Соглашения, эта основа применялась в различных областях, начиная с эволюционная биология (E.O. Wilson 1975 [2000: 23]), эволюционная биология. психологии (Daly & Wilson 1978; Crawford 1998) и поведенческих от экологии (Morse 1980: 92–95) до гуманитарных наук, как в сотрудничество (Marchionni & Vromen 2009) и развивающее психология (Lickliter & Berry, 1990).Особенно в эволюционном биологии, он вносил свой вклад в основную причинно-следственную связь в течение долгого времени. время, даже среди биологов-эволюционистов, интересующихся вопросами развития процессы (см., например, Maynard Smith 1982: 6).
2.2 Объединение непосредственных и конечных причин
Постоянная критика ближайшего и окончательного различие (еще до Майра 1961 г.) и против лежащего в его основе идея принизить объяснительную или причинную значимость развития к эволюции.В последнее время обсуждение этого вопроса набирает обороты. благодаря новым открытиям в таких областях, как эпигенетика, эволюция и ниша теория строительства (Thierry 2005; Laland et al. 2011, 2013a, 2013b; Хейг 2011, 2013; Скотт-Филлипс и др. 2011; Дикинс и Рахман 2012; Герреро-Босанья 2012; Calcott 2013; Дикинс и Бартон, 2013 г .; Гарднер 2013; Mesoudi et al. 2013; Мартинес и Эспозито 2014; Scholl & Pigliucci 2015; Baedke 2018; Уллер и Лаланд 2019). В этом контексте некоторые ученые утверждают, что непосредственное-окончательное различие стоит «в центре некоторых из самые ожесточенные споры современной биологии »(Laland et al.2011: 1512) о роли пластичности развития, нише построение и инклюзивное наследование эволюционных траекторий. Участники этой дискуссии утверждали, что мы должны в силу различные эпистемологические или эвристические причины, придерживайтесь мнения Майра непосредственное-окончательное различие (Скотт-Филлипс и др., 2011; Дикинс & Barton 2013) или его пересмотренная или переосмысленная форма (Scholl И Пильуччи 2015; Otsuka 2015), расширите его на третье промежуточное звено. форму объяснения (Haig 2013) или заменить ее концепцией «Взаимная причинность» (Laland et al.2011, 2013a, 2013b, 2015; Mesoudi et al. 2013). В соответствии с более ранней философской работой (Oyama 1985; Keller 2010; Griffiths & Stotz 2013), последняя идея взаимной причинности должны позволять описывать процессы обратной связи между причинными факторами в развивающихся системах. Это включает в себя способность организмов к фенотипической пластичности или, более в частности, их деятельность по изменению давления отбора. Примеры парадигматической обратной связи — это нишевое конструктивное поведение организмы, которые изменяют свою среду обитания и, таким образом, формируют естественные давление отбора, работающее над ними.Другими словами, взаимное причинно-следственная связь утверждает, что организмы — это не только результат адаптивных процессы, но и причинные отправные точки эволюционного траектории. В этом смысле эта структура выступает против причинно-следственной связи. и / или объяснительное утверждение асимметрии ближайшего-окончательного различие. Он подчеркивает важную роль развития для эволюция.
Против этого нового подхода ученые утверждали, что взаимное причинно-следственная связь на самом деле не представляет никаких концептуальных проблем для эволюционная биология, так как она была включена с некоторого времени назад в поле (Svennson 2018).Однако настоящая проблема состоит в том, чтобы развить эту идею в методологически обоснованную основу, которая позволяет изучение и моделирование сложной нелинейной системы организм-среда связи. Другие ставят под сомнение центральную причинную роль единицы. организма, как предполагается, играет в рамках этой взаимности. (Baedke 2019a), или поставил под сомнение, может ли эта концептуализация в Фактически, охватить все представляющие интерес отношения причинной зависимости для эволюционная биология (Martínez & Esposito 2014; Scholl И Пильуччи 2015).Более того, некоторые утверждали, что и этот фреймворк полагается на дихотомию между развитием и эволюцией (Дикинс & Бартон 2013; Martínez & Esposito 2014) и что это не способствует успешная биологическая наука, так как она не приводит к фальсифицируемым вопросы (Dickins & Rahman 2012) и кровоточит непосредственно и окончательные объяснения друг друга, так что их различие становится бессмысленным (Gardner 2013; следует, однако, отметить, что это может быть самой целью этого подхода).В более общем плане было предложено, чтобы сторонники этого подхода предоставили больше концептуальные разъяснения того, что на самом деле является взаимной причинно-следственной связью должно означать (Buskell 2019).
Помимо различения развития и эволюции в качественном как непосредственные и окончательные причинные процессы, менее распространенный попытка состоит в том, чтобы количественно различить (или связать) их. Здесь, особенно различия, основанные на темпах или временных масштабах, на которых происходят различные процессы развития и биологические процессы (см. запись уровни организации в биологии).Например, Конрад Х. Уоддингтон (1957) разработал иерархическую модель временных шкал, включающая биохимические процессы на нижних молекулярные уровни организации с более высокой скоростью, средней скоростью процессы развития на среднем уровне и эволюционные процессы на более высоких уровнях с более медленным темпом. Согласно такой точке зрения, эволюционные процессы — это просто процессы, происходящие с разными темп и, следовательно, на другом уровне, чем уровни развития. Таким образом, они отличаются скорее постепенно, чем по сути.Рейтинговые различия имеют было описано, чтобы быть совместимым с окончательной ближайшей структурой (интерпретируя его как тот, который различает разные временные рамки фенотипического изменения; см. Francis 1990; Haig 2013) или как разные от ближайших до конечных различий (Baedke & Mc Manus 2018). В кроме того, концептуализация шкалы времени (или шкалы размеров) была применяется для разработки методологий и многомасштабного моделирования, интегрировать, среди прочего, процессы развития и эволюции (С.Левин 1992; Грин и Баттерман, 2017; Дакворт 2019).
3. Изменения в развитии как механизм эволюции
Идея о том, что эволюция представляет собой изменение развития отношения имеют центральное значение для области эволюционного развития биология (эво-дево). Эта область описывает себя как науку, которая изучает, как изменения в развитии создают вариации, которые природа может выбирать (Raff & Kaufman 1983; Gilbert, Opitz, & Raff 1996). Другими словами, естественный отбор не создавал вариаций; развитие создает вариации.Развитие — это художник; естественный выбор — куратор (Gilbert 2006, 2019). У обоих есть творческие агентство; но они работают на разных уровнях. Хотя это мнение были выражены такими учеными, как Томас Хаксли, Джулиан Хаксли, Конрад Х. Уоддингтон и Ричард Б. Гольдшмидт, он получил доверие благодаря более поздним открытиям, которые объяснили, как нормальное развитие могло произойти. Главным среди этих открытий было объяснение пути развития, которые связывают эмбриональную индукцию с геном выражение.Здесь паракринные факторы (белки, влияющие на поведения или паттерны экспрессии генов соседних клеток) секретируются одним набором клеток были получены рецепторы на мембранах другие клетки. Эти рецепторы затем активировали белки в цитоплазма, которая в конечном итоге активировала или подавляла поступающие белки ядро, регулирующее транскрипцию определенных генов.
Вторым крупным открытием, которое способствовало развитию evo-DevO, было открытие
модульные усилители. Вышеупомянутые факторы транскрипции будут
связываются с определенными участками ДНК, называемыми энхансерами.Большинство генов имеют
несколько энхансеров. Таким образом, у гена может быть «конечность».
энхансер, который активирует экспрессию гена в конечности, и
Энхансер «глаза», который обеспечивает экспрессию гена в
глаз. Более того, каждый энхансер обычно связывает несколько транскрипционных
факторы и могут быть активированы в логических
Эти различия в экспрессии генов можно разделить на четыре категории. категории (Артур 2004). Одна из этих категорий включает в себя место экспрессии генов, где разные популяции клетки экспрессируют определенный ген у разных видов. Например, ген gremlin в перепонках задних конечностей утки защищает эти клетки от клеточной гибели, что приводит к перепончатым лапам (Laufer et al, 1997; Merino et al.1999). Вторая категория включает изменения в время экспрессии гена, как в случае продолжающейся экспрессии ген fgf8 на кончике передних конечностей дельфина, таким образом позволяя выдвигать ласты (Richardson & Oelschläger 2002). Третья категория изменений включает изменения в величине экспрессии генов, как в различия в экспрессии гена Bmp4 , определяющие ширину клювов вьюрка (Абжанов и др., 2004). Четвертая категория сосредоточена на изменения фактической последовательности белка регуляторного белков, как в изменениях гена Antennapedia в насекомые, которые не позволяют насекомым образовывать более шести конечностей (Галант и Кэрролл, 2002).
Третьим открытием стало выяснение регуляторных сетей генов. (GRN). GRN основан на паракринных факторах, передаче сигнала каскады и факторы транскрипции. Игнорируя посттранскрипционная регуляция генов, эта концепция пытается объяснить как начальные условия (РНК и белки в ооците, положение эмбриона в матке и т. д.) могут создать условия при этом типы клеток различаются, хотя их геномы идентичны. Возглавляемый Эриком Дэвидсоном (2001, 2006), этот подход использует системы теоретические концепции, такие как модульность и диссоциативность, чтобы объяснить, как гены взаимодействуют иерархическим образом, производя различные типы клеток (Levine & Davidson 2005) и то, как типы клеток в родственные виды могут отличаться пополнением (кооптированием) конкретный GRN путем изменения связывания фактора транскрипции.Более в целом открытие GRN позволило интегрировать биология развития с палеонтологией (Jablonski 2017; Hinman et al. 2003; Hinman & Cheatle Jarvela 2014), а также может быть расширен на области симбиоза и конструирования ниш (Laubichler & Renn 2015).
Четвертым открытием эволюционной биологии развития было важность пластичности развития для эволюции (Nijhout 1990; Gilbert 2001; Пильуччи 2001). К концу ХХ века роль температуры, солнечного света, диеты, скученности, материнского поведения, и хищничество, как было замечено, играет важную роль в влиянии на фенотипы растения и животные.Таким образом, среда не только выбрана вариации, это помогло их произвести. Поскольку evo-DevO постулировал, что изменения в развитии вызывают эволюцию, и поскольку развитие пластичность сыграла роль в развитии, тогда возникла необходимость смотрите на изменения пластичности как на часть эволюции. Рано годы 21-го века, пластичность развития сыграла свою роль роли в эволюционных изменениях (West-Eberhard 2003; Abouheif et al. 2014; Suzuki & Nijhout 2006; Rajakumar et al. 2018; Левис и Pfennig 2020), и понятие «пластичность прежде всего эволюция »(или« эволюция в первую очередь ») интегрировали данные из многочисленных источников в программу, в которой физиологическая способность изменять фенотип из-за агенты окружающей среды могут стать канализованными и генетически закрепленными выбор.Механизмы, с помощью которых произошла такая эволюция, основанная на пластичности. осуществлено (разоблачение и отбор загадочных генетических вариантов, связанная со стрессом неспособность молекулярных шаперонов сворачивание мутантных белков и т. д.) стало новой исследовательской программой.
Пятое открытие заключалось в осознании того, что одним из основных агентами окружающей среды, влияющими на развитие, были симбиотические микробы (Макфолл-Нгай, 2002; Гилберт и др., 2012, 2015). Понятие холобионт (то есть интегрированный составной организм, состоящий из микробных и эукариотические виды-хозяева) организовали большую часть данных, чтобы посмотреть на роль микробов в нормальном развитии организм и варианты нормального развития (при болезни и эволюция) (Rosenberg & Zilber-Rosenberg 2016; см. запись биологические особи).Например, у мышей нормальное развитие иммунной система и капиллярная сеть кишечника зависят от конкретных бактерий полученный при рождении. Эти бактерии вызывают экспрессию определенные гены в эукариотических клетках и белки, производимые эти гены влияют на судьбу клеток (Hooper et al. 2001; McFall-Ngai et al. al. 2013). Другими словами, бактерии могут действовать как эмбриональная клетка. бы, регулируя экспрессию генов в соседних клетках. Здесь эукариотический организм нуждается в этих бактериях и ожидает их присутствия в течение нормальное развитие.
Как и в других случаях пластичности развития, следующим шагом было чтобы увидеть, могут ли изменения в развитии симбионтов вызвать изменения в эволюция (см. O’Malley 2015). Было показано, что изменения в симбионтах могут обеспечить выбираемые варианты для эволюции (Zhang et al., 2019), и он мог обеспечивают основу для репродуктивной изоляции через цитоплазматическую несовместимость или предпочтение спаривания (Brucker & Bordenstein 2013; Sharon et al. 2010). Одна из самых интересных возможностей, однако, исходит из точки зрения, что большинство, если не все, эукариотические организмы — холобионты, и симбионты открывают новые эволюционные траектории.Например, симбиотические микробы известны давно. отвечать за ферменты, переваривающие растения, в желудках жвачные животные. Без бактерий, переваривающих целлюлозу, коровы не переваривают трава или зерно. Более того, микробы помогают создавать рубец после они колонизируют пищеварительную систему при рождении. Таким образом, эти микробы также вызывают формирование органа, в котором они находятся, и позволяют им функция (Гилберт 2020; Чиу и Гилберт 2020). Развивающий симбиоз (симпоэзис), таким образом, открыл эволюционные траектории для некоторые млекопитающие.Это пример симбиоза развития и нишевое строительство. Конструкция ниши зависит от развития пластичность (Лаланд и др., 2008).
Эти пять новых (или обновленных) аспектов биологии развития имеют несколько философских выводов. Среди прочего, они касаются онтологические вопросы о том, что такое развивающиеся организмы и как они следует объяснять с комплексной точки зрения развития эволюция.
4. Онтологические проблемы эволюции развития
Многие онтологические споры о связи между развитием и эволюция фокусируется на организме как центральной единице, которая одновременно развивается и эволюционирует, в отличие, например, от генов или населения.Несколько теорий пытались прояснить природу этого организменная единица: Согласно одной влиятельной точке зрения, организм — это серия интегрированных процессов в течение жизненного цикла (Bonner 1965; Николсон и Дюпре, 2018; Fusco 2019a; DiFrisco 2019), с сложные и взаимные отношения между всем организмом и его части (Gilbert & Sarkar 2000; Esposito 2016; Peterson 2017). Центральные элементы этой точки зрения основаны на органицистской концепции. разработан Кантом, который утверждает, что в организмах «части, по отношению как к форме, так и к бытию, возможны только через их отношение к целому »и« что части связывают взаимно превращаются в единое целое таким образом, что они являются взаимно причиной и следствием друг друга »(Kant 1790/1793 [1902: 373]; см. также Lenoir 1982).Haraway (2008) подчеркивает последняя идея о взаимном взаимодействии частей организмов говоря: «Взаимная индукция — это название игры» (2008: 228), и это «взаимная сложность на всем пути. вниз »(2008: 42).
Эти идеи интеграции жизненного цикла и повсеместная взаимность предполагает более процессный и ориентированный на организм онтологический взгляд на организм. Эта точка зрения становится важен в исследованиях эволюции. Например, чтобы понять как развивается нервная система, нужно учитывать, что нервная система развивающегося организма имеет иные функции, чем нервная система взрослого человека и может использоваться для координации строения тела по мере развития (М.Левин 2019; Филдс и др. 2020). Это развивающееся системы демонстрируют истощение и конкуренцию, а развивающиеся системы показывают сотрудничество позволило Филдсу и Левину (готовится к печати) предположить, что процессы развития и эволюции могут быть интегрированы в безмасштабный уровень за счет языка обработки информации и коммуникация.
Второе онтологическое соображение гласит, что развитие и развитие организмы — это интегрированные коллективные особи, так называемые холобионты. или «мета-организмы» (Зильбер-Розенберг и Розенберг 2008; Bosch & McFall-Ngai 2011; Гилберт, Сапп и Таубер 2012; см. также O’Malley 2017; Baedke et al.2020a). Поэтому наши дискуссии об эволюции должны учитывать, что каждый организм — это консорциум, имеющий множество геномов, а не один, как традиционно предполагается. Математическое моделирование эволюции холобионтов, учитывающих это разнообразие, просто начало (Roughgarden et al. 2018; Osmanovic et al. 2018; Roughgarden 2020). Эта новая онтологическая основа утверждает, что симбиоз — это норма; это не периферийный. Эти симбионты могут действовать по-разному. этапы жизненного цикла и рассматриваются до развития каркаса (Гриземер 2014; Чиу и Гилберт 2015; Минелли 2016).В «Строительные леса», каждая стадия развития становится возможной объектами и процессами, которые катализируют эту деятельность, что позволяет новые и эволюционно релевантные процессы, которые будут происходить на более низком уровне трудности и затраты.
Третий онтологический момент касается точной природы ссылки. между развитием и эволюцией. Были предложены два подхода вперёд: Один опирается на идею, что биологическая сущность, которая причинно-следственная связь в обеих сферах может быть найдена только на уровне интегрированные коллективы симбиотических взаимодействий.Следуя в традиции представления Лейбница о возможности, а также Маргулис (1999) утверждает, что мы живем за счет «симбиотической планеты », эта точка зрения утверждает, что симбиотические коллективы не только существенные единицы развития, но также и эволюции. Это приводит с точки зрения эволюции, которая не сосредоточена на межвидовых конфликтах и соревнование между людьми (Huxley 1888; Williams 1966; Докинз 1976 г.). Вместо этого сущности, которые развиваются, сотрудничают соразвивающиеся коллективы (Rosenberg & Zilber-Rosenberg, 2016; Roughgarden et al.2018; для обсуждения см. Peacock 2011; Суарес 2018).
Ссылки на другую онтологическую структуру развитие и эволюция через сущность действующего организма (Nicholson 2014, 2018; Walsh 2015; обсуждение см. В Pradeu 2016; Baedke 2019a, 2019b). Эти подходы обычно меньше связаны с исследованиями симбиоза, чем исследованиями конструирования ниш или материнского последствия. Здесь организм (например, бобер, строящий плотину или дождевой червь, который обрабатывает почву) построен как самоопределение агент, который своим поведением модулирует давление отбора воздействуя на него (Odling-Smee et al.2003; Уллер и Хелантера 2019). Таким образом, согласно этому аргументу, он может оказывать влияние на население динамика. Перспектива холобионта и конструкция ниши перспективы объединяются, когда каждый понимает, что микробы и хозяева образуют среду друг друга. Здесь хозяин и микробы поддерживают развитие и эволюцию друг друга (Чиу И Гилберт 2015, 2020).
Независимо от того, рассматриваем ли мы коллективные или организменные индивидуальные агенты как основные субъекты, участвующие как в развитии, так и в эволюции, попытки интегрировать эти две области должны в каждом случае показать, что, Фактически, это то же самое подразделение, которое разрабатывает и эволюционирует .В другими словами, если мы хотим объединить развитие и эволюцию через единица биологического индивида (являющаяся единственной сущностью, которая принимает в обоих) эта единица должна соответствовать критериям как физиологических (например, метаболический) и эволюционной индивидуальности (см. биологические особи). Эволюционные личности традиционно рассматривались как репродуктивные единицы с дифференциальной приспособленностью и общими линиями (так называемые «дарвинисты»; см. Годфри-Смит 2009) или как единицы выбора («взаимодействующие»; см. Корпус 1980).К сожалению, обе эти единицы не всегда совпадают. (Годфри-Смит 2013; Pradeu 2016). Например, некоторые организмы (голобионты) образуют развивающиеся но без репродуктивных единиц, так как они включают множество линий передачи (например, микробные). Другие возможные единицы отбора (например, гены или популяции) не идентичны физиологическим индивидуумам. Таким образом, физиологический человек не обязательно может быть эволюционная единица или наоборот.
Это подводит нас к четвертому онтологическому пункту: развитие пластичность, которая, как считается, смещает или даже направляет эволюционные траектории (West-Eberhard 2003; Radersma et al.предстоящий). В концепция пластичности гласит, что развитие можно регулировать в важные пути окружающей среды. Это исключает генетический детерминизм (но не обязательно экологический детерминизм; см. Wagoner and Uller 2015). В первоначальной концепции производства фенотипа (т. Е. развития), Вильгельм Йоханнсен (1909) указал, что фенотип является продуктом как генотипа, так и условий окружающей среды, и Вольтерек (1909; см. Также Саркар 1999) утверждал, что то, что было унаследовано это « Reaktionsnorm », потенциал генерировать фенотипические вариации в ответ на факторы окружающей среды.В соответствии с этой точкой зрения многие тексты по эмбриологии в конце 1800-х годов (например, Hertwig 1894) провозгласили точка зрения, что развитие потребовало как взаимодействия между эмбриональные клетки и дальнейшее взаимодействие этих клеток с окружающая среда (см. Nyhart 1995).
Несмотря на эту историю, пластичность развития была маргинализована, поскольку генетические объяснения вышли на первый план в середине 20-го века (Саркар 1998; Келлер 2002). На этом фоне такие эмбриологи, как Льюис Вольперт (1994) мог спросить, является ли фенотип организма можно было бы вычислить, если бы у нас было полное описание яйца.Из-за вышеприведенные данные о взаимодействии организм-среда (см. 3) возникла иная точка зрения, более серьезно рассматривающая экологичность и пластичность развивающихся фенотип. Эта точка зрения включает переход от экстерналистов к интерналистам. или конструкционистское понимание организма-окружающей среды отношения (Годфри-Смит 1996). В то время как экстерналистский взгляд — ортодоксальный взгляд в эволюционной теории — концептуализирует свойства организмов в результате их окружающей среды (т.е. естественный отбор, нацеленный на генетические программы), интерналистский взгляд рассматривает «один набор органических свойств с точки зрения других внутренних или внутренние свойства органической системы »(Годфри-Смит 1996: 30). Согласно этим двум рассказам, организмы занимают окружающая среда, которая совпадает с ними или которая в значительной степени не зависит от их вариация. Вышеупомянутое исследование эволюции развития предлагает перейти от экстерналиста к конструкционисту перспектива, в которой организм активно формирует свои внутренние состояния и реагирует на внешнюю среду и изменяет ее (см. Laland et al.2014, 2015). Кроме того, в этой структуре также причинная роль окружающая среда становится более сложной. Теперь он включает идею, что окружающая среда имеет активный агент, который может определять фенотип. Вместо того, чтобы рассматривать окружающую среду как не что иное, как пассивное фильтра для эволюции, с этой точки зрения окружающая среда играет роль в активация фенотипа в дополнение к его выбору (Moczek 2015; Гилберт и Эпель 2015).
5. Объяснение эволюции развития
Помимо этих дискуссий об онтологии разработки и эволюционирующих организмов, другие центральные философские дискуссии о взаимодействие между развитием и эволюцией нацелены на тему научное объяснение.Имеется в виду вопросы о том, какие исследования эволюции развития (должны) объяснять и как они объясняют.
5.1 Механистические объяснения эволюции развития
Философы науки давно выступают за объяснительную автономию биологических объяснений. Особенно критиковали понимание биологического объяснения как аналогичного законным счетам объяснений в физике (см. Lange 2007). Напротив, ученые утверждали, что объяснение в бионауках часто включает описание механизма, который вызывает определенные биологические феномен (Bechtel & Richardson 1993; Craver 2007; Bechtel & Abrahamsen 2010; см. также запись механизмы в науке).Эво-дево было описано как парадигматический механистический наука, которая — вопреки крайнему и непосредственному различению — стремится определить механизмы развития, которые могут объяснить эволюционное изменение фенотипов (Gilbert 2003; Hall 2012). Этот механистический подход часто сопровождается математическими моделями различные паттерны развития, от изменений в регуляции генов сетей к моделям роста организмов и историческими повествованиями о том, как развиваются организмы и виды (Jaeger & Sharpe 2014; Winther 2015).Однако, помимо признанной центральности механистического объяснение эволюции развития, широко обсуждаемая тема что такое механизм развития и как он функционирует в эволюционное объяснение по сравнению со стандартными объяснениями, цитирующими естественный отбор.
Философы биологии (в так называемой новой механистической философии) концептуализировали механистические объяснения в биологии как построение моделей механизмов, соединяющих части систем, расположен на одном уровне организации, с поведением всего системы, обычно расположенной на более высоком уровне организации (Machamer et al. al.2000; Craver 2007; Иллари и Уильямсон 2012). В этом рамки, механистические модели связывают различные композиционные уровни организация, как гены и фенотипы или клетки и ткани. Эти межуровневые отношения существуют между причинными способностями частей системы и их организации, а также возможностей системы как все. Такие отношения устанавливаются в порядке декомпозиция и локализация (Bechtel & Richardson 1993; Craver 2007; Menzies 2012). Эта концептуальная основа для описания биологических механизмы и механистическое объяснение было разработано на основе тематические исследования в области молекулярной и клеточной биологии.Однако ученые поставить под сомнение полезность описания механистических объяснения в исследованиях развития и развития эволюция.
Что касается развития, то, во-первых, утверждалось, что организация играет иную роль в механистическом развитии объяснения (Mc Manus 2012). В отличие от вышеупомянутой структуры, что обычно предполагает, что уровни организации просто там, и, следовательно, не нужно разъяснять, как уровни организации на самом деле происходят, происхождение уровней и других форм организация (e.g., пространственные оси) специально рассматриваются в объяснения механистического развития. Во-вторых, философы утверждал, что отношения между уровнями прослеживаются с точки зрения развития механизмы не исчерпываются синхронными, определяющими отношениями между частями и целым, как предлагают некоторые новые механики (Craver & Bechtel 2007). Напротив, объяснения развития отслеживают изменение диахронические отношения между причинными способностями системы при разные уровни организации в разные промежутки времени (Ylikoski 2013; Бэдке и Мак Манус, 2018; Baedke 2020; см. также Love & Hüttemann 2011).В-третьих, утверждалось, что объяснения в evo-DevO с использованием механизмов развития сталкиваются с проблемой из-за неоднородность этих механизмов (Love 2018). При попытке объединить два типа объяснений механизмов развития — объяснения высококонсервативных молекулярно-генетических механизмов, как сети регуляции генов, и объяснения клеточно-физических механизмы, такие как миграция клеток — иногда возникает компромисс. Вместо того, чтобы давать более полное объяснение, объединение двух механизмы могут привести к менее общему объяснению, поскольку нефилогенетически консервативные клеточно-физические механизмы дают меньше общность объяснений.Этот компромисс может привести к объяснению предвзятость к проектам, которые стремятся объединить развитие и эволюцию. Это может побудить исследователей склоняться к общности объяснений, которые ссылаются на высококонсервативные молекулярно-генетические механизмы и не клеточно-физические механизмы, более комплексные объяснения, цитирующие оба виды механизмов.
Что касается концепции механизма эволюции развития, Бригандт (2015) подчеркивает, что некоторые механистические объяснения в evo-DevO — как те, что о том, как развитие влияет на эволюцию (Radersma et al.предстоящий), как новые вариации возникают благодаря пластичность развития (Pigliucci 2001, Gilbert 2006) и как организмы порождают наследственные адаптивные фенотипические вариации (эволюционируемость; см. Brown 2014) — значительно расширить стандартный анализ декомпозиции и локализации с помощью динамических модели (см. также Bechtel & Abrahamsen 2010; Brigandt 2013; Baedke 2020). Эти модели позволяют прогнозировать динамику развития системы. После разложения системы и выявления причинных вклады частей или подсистем, динамические модели помогают продемонстрировать, как эти вклады работают вместе, чтобы вызвать поведение всей системы.Таким образом они отвечают, как механизмы развития создают эволюционно релевантные качественные изменения фенотипических свойств, таких как устойчивость, фенотипические пластичность и модульность через лежащие в основе количественные изменения в их составные части и виды деятельности. Один классический пример этого являются математическими моделями устойчивости пространственного рассеяния и сегментация у Drosophila . Они обеспечивают количественные информация о взаимодействии лежащей в основе генной сети компоненты, включая, например, скорость транскрипции и распад генов уровни генных продуктов (von Dassow et al.2000).
Другие обсуждения на стыке развития и эволюции сосредоточьтесь на структуре объяснений, которые касаются того, как механизмы развиваются. Калкотт (2009, 2013) утверждает, что Майр различие характеризует два вида объяснения: объяснение, которое отвечает: «Как работают человека, человек за ? »И эволюционное объяснение, которое отвечает «Как популяции меняют с течением времени?» Однако существует третий вид, называемый «родословная». объяснение’.Объяснение происхождения отличается от приведенного выше на ответив «Как человека меняют сверх время ? »Поэтому они предлагают ряд механистических модели, в которых прослеживаются различия между механизмами развития особей, которые производят соответствующие морфологические структуры на разные времена. В течение эволюционного времени эти отношения претерпевают небольшие изменения. модификации, которые в конечном итоге приводят к нововведениям, таким как глаза и перья во всей системе. Таким образом, объяснение происхождения расширяет философские рамки стандартов из единственного описания механизм в серию механистических моделей.
5.2 Объяснительная сила объяснений эволюции сторонниками развития
Помимо этих дебатов о структуре биологических механизмов и их роль в объяснении эволюции развития, философы биология и биологи обсуждали в более общем плане, какая добродетель объяснения развития (могли бы) иметь для обращения к эволюционным явления. С этим вопросом связан вопрос, сколько — в смысл того, что за факты — только естественный отбор может объясните (см. запись адаптационизм).Широко признано, что такие объяснения могут касаться общих динамика частот черт и выживаемость (см. Sober 1984). Тем не мение, верно ли это для более детального рассмотрения развития отдельных черт личности — нерешенный вопрос. Хотя некоторые авторы утверждают, что эволюционные объяснения естественным отбором могут объяснять, почему у конкретного человека есть определенная черта, а не другая черта (Neander 1995; Forber 2005), другие отрицают это (Sober 1984; Stegmann 2010).Более того, утверждалось, что интегрирующие объяснения феноменов развития, таких как предвзятость в развитии, фенотипическая пластичность, конструирование ниш и инклюзивное наследование к объяснительной структуре эволюционного теория приведет к «значительно расширенному объяснительному емкость »этой теории (Pigliucci & Müller 2010: 12). Однако, хотя в эволюционной биологии часто существует согласие относительно существование этих феноменов развития (Laland et al. 2014; Рэй и др. 2014), их объяснительная значимость подвергается сомнению.Против На этом фоне ученые начали анализировать, на основании каких критериев объяснительной силы, такой как точность, соразмерность, чувствительность, и идеализация, эволюционистские эволюционные объяснения лучше, чем объяснения селекционера. Это включает в себя определение, какие компромиссы между пояснительными стандартами (например, между точностью и чувствительность или идеализация) те аккаунты, с которыми сталкиваются объединить объяснения развития и эволюции (Baedke et al. 2020b; Uller et al.2020).
6. Антропологические и этические аспекты эволюции развития
Вышеупомянутые дебаты о взаимосвязи между эволюцией и развитие в первую очередь возникло в рамках философии науки, особенно по истории и философии биологии. Однако есть и другие, более общие дебаты по антропологическим и этическим вопросам, которые касаются эволюция развития. В первую очередь относятся к тому, как мы концептуализируем развивающиеся и развивающиеся люди, их жизнь, тело и здоровье, а также как мы распределяем обязанности по медицинскому вмешательству.
Во-первых, новые открытия в пластичности развивающегося организма, их взаимосвязанность и способы передачи из поколения в поколение информация повлияла на научное и общественное понимание того, что люди. Например, если люди концептуализируются как холобионты — как коллективы совместно развивающихся и совместно эволюционирующих организмов — это также означает, что развитие — это вопрос совместное построение взаимодействий между видами. Значит, как Харауэй (2016) сформулировал это, что мы — как люди — очень буквально «стать с другими».’ В контексте, симпоэзис (симбиоз развития) означает, что развитие совместная разработка. На этом фоне Джон Дюпре и Морин О’Мэлли (2009) рассматривает живые существа как интерактивные коллекции: «Жизнь, как мы утверждаем, обычно находится в совместных пересечения многих родословных, и мы даже предлагаем сотрудничество следует рассматривать как центральную характеристику жизни иметь значение».
Во-вторых, биологические и биофилософские дебаты о развитии эволюция и пластичность организмов и окружающая среда отзывчивость повлияла на дебаты о том, что такое человеческое тело.За Например, Jörg Niewöhner (2011) утверждает, что новая концепция человеческое тело в настоящее время появляется в современной биологии, так называемые «Встроенное тело». Согласно этой точке зрения, человеческое тело больше не машиноподобная единица, которая генетически запрограммирована, нервно-контролируемый и ограниченный кожей, но открытый и динамичный единица, которую нельзя понять в отрыве от ее материальных и социальных окружающая среда (см. также Baedke 2017). Дополнительно в корпус встроен в различные временные шкалы, начиная от его эволюционного и трансгенерационные в онтогенетическое прошлое, которые постоянно составляют его настоящее.Другие возражали против стандартного человеческого рождения. повествования и Аристотеля, определившего временные границы людей при рождении и смерти, то рождение у людей не является создание нового человека. Напротив, рождение следует понимать как зарождение нового многовидового коллектива (Gilbert 2014; Chiu & Гилберт 2015).
Еще одна антропологическая проблема, возникшая в результате недавнего исследования evo-DevO и эпигенетика, в-третьих, вопрос, как определить нормальность и здоровье человека (см. Baedke 2019b).Есть свидетельства того, что бактерии необходимы для нашего нормального когнитивного и социального развития. За Например, стерильные мыши асоциальны и ведут себя как аутисты. (Desbonnet et al., 2014), и это поведение может быть воспроизведено имплантация микробиома аутичным пациентам (но не контроль пациентов) в стерильных мышей (Sharon et al., 2019). Такие случаи предполагают что биологическая нормальность не является неотъемлемым свойством организмов но возникает через взаимосвязи с другими организмами и окружающая обстановка.Кроме того, наше понимание здоровья все больше возрастает. оспаривается. Особенно это относится к взгляду, описывающему человеческое здоровье как свобода и автономия от внешнего вмешательства. Это обычно считает бактерии отклонениями от нормы, а паразитизм — патологический, потому что он угрожает и загрязняет чистоту индивидуальный энергетический паттерн. Вместо этого в (более процедурном) холобионтический каркас, микробы необходимы для нормального развития и считается, что они предотвращают развитие определенных заболеваний (Bello et al. al.2018; Кирьявайнен и др. 2019). Кроме того, некоторые организации, бактерии или вирусы, которые ранее считались вредными, теперь все чаще считается «хорошим» или «Здоровые» сотрудники, а не «плохие злоумышленники» (Дюпре и Гуттингер, 2016). В более общем смысле это означает что, поскольку микробиота становится все более важной компоненты, которые стабилизируют нормальное развитие и эволюционируют вместе с люди, следовательно, они обладают чертами, критически важными для физической формы человека, то есть здоровье. Эта новая перспектива может привести к радикальным изменениям в персонализированное наблюдение и лечение заболеваний и многое другое как правило, к новым стратегиям в разработке политики, которые заменяют идею сохранения автономной личности от заражения идеей поддержания (равновесных состояний) коллективов соразвивающихся и совместно развивающиеся люди.
Наконец, в более этическом аспекте эти выводы о открытость людей своему окружению и друг другу привело, во-первых, к дискуссиям о том, кто берет на себя ответственность за состояния здоровья людей и вмешательства (например, на эпигеномы и микробиомов) во временных масштабах внутри и между поколениями (Dupras & Равицкий 2016). Если развитие пластика может влиять на эволюцию, кто отвечает за результаты развития и эволюционные траектории в людях? Должен ли человек, являющийся центральным медицинским учреждением, агент, который «освобожден от цепей своих генов» (Пикерсгилл и др.2013), взять на себя эту ответственность? Или должен коллективы, такие как национальные государства или международные организации, должны быть отвечает за уровень токсинов в окружающей среде, а также за пища, которую едят люди, и стресс, которому они подвергаются (Hedlund 2012)? Последний отчет об ответственности направлен на то, чтобы не переоценивать причинная роль матери как важнейшего звена общественного здравоохранения агентов, которые должны быть привлечены к ответственности (и виновны), если их дети или более поздние поколения заболевают (Richardson et al.2014).
7. Заключительные замечания
Связь между эволюцией и развитием была долгой. обсуждаемая тема в истории биологии и философии биологии. В этой записи отобрана небольшая часть работ, относящихся к концептуальные, эпистемологические, антропологические и этические размышления об этих отношениях. Помимо обсуждаемых здесь вопросов, философы биологов и биологов обсудили, как развитие и филогенетические подходы к гомологии могут быть интегрированы (Amundson 2004; Вагнер 2014; DiFrisco 2019; см. запись биология развития), какие модельные организмы (Love 2009; Lloyd et al.2012; Минелли и Бедке, 2014 г .; Zuk et al. 2014; см. запись биология развития) и репрезентативные инструменты, которые ученые (должны) использовать для изучения отношения между эволюцией и развитием (например, нормальные пластины, карты судьбы клеток, эпигенетические ландшафты) и то, что эпистемологические и эвристические роли этих инструментов в научной практике (см. Haraway 1976; Gilbert 1991; Hopwood 2007; Любовь 2010; Baedke 2013; Бедке и Schöttler 2017; Никоглу 2018). Кроме того, давние дебаты по телеологии в биологии (см. запись телеологические понятия в биологии) обратил внимание на связь между эволюцией и развитием, чтобы различать телеологическое измерение развития от нетелеологического эволюционных процессов (Mayr 1961), чтобы ввести новые концептуальные фреймворки, такие как телеономия, которые относятся только к очевидным целенаправленным систем (Pittendrigh 1958), или чтобы подчеркнуть целенаправленное действие организменный агент как отправная точка для рассуждений о телеологических природа эволюции (Russell 1950; Piaget 1976 [1978]; Walsh 2015).
В настоящее время ведутся философские дебаты о подходящем концептуальном и объяснительный подход к объединению или интеграции развития и эволюционные процессы не достигли консенсуса. Это не в последнюю очередь в связи с тем, что соответствующие дисциплинарные области, такие как ево-дево, эпигенетика и теория конструирования ниш, которые способствуют развитию таких интеграции, все еще относительно молоды. Их теоретические и концептуальные рамки весьма разнородны. Таким образом, важные цели для будущих философских исследований — это понять препятствия на пути стабилизация и укрепление этих каркасов, чтобы идентифицировать их объяснительные достоинства и ограничения, а также внимание к их влиянию на то, как мы понимаем людей и людей здоровье.
Микробиом расширяет эволюционный потенциал хозяина
Blaser, M. J. & Falkow, S. Каковы последствия исчезновения микробиоты человека? Nat. Rev. Microbiol. 7 , 887–894 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Friesen, M. L. et al. Функциональные признаки растений, опосредованные микробами. Annu. Rev. Ecol. Evol. Syst. 42 , 23–46 (2011).
Артикул Google Scholar
McFall-Ngai, M. et al. Животные в бактериальном мире — новый императив для наук о жизни. Proc. Natl Acad. Sci. 110 , 3229–3236 (2013).
ADS CAS PubMed Статья PubMed Central Google Scholar
Дуглас А. Э. Симбиоз как общий принцип эволюции эукариот. Cold Spring Harb. Перспектива. Биол . 6 , а016113 (2014).
Стаппенбек, Т. С. и Вирджин, Х. У. Учет взаимных взаимодействий хозяина и микробиома в экспериментальной науке. Природа 534 , 191–199 (2016).
ADS CAS PubMed Статья PubMed Central Google Scholar
Моран, Н. А. и Слоан, Д. Б. Концепция гологенома: полезный или пустой? PLoS Biol. 13 , e1002311 (2015).
PubMed Статья CAS PubMed Central Google Scholar
Дуглас, А. Э. и Веррен, Дж. Х. Дыры в гологеноме: почему симбиозы хозяин-микроб не являются холобионтами. МБио 7 , e02099 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Коскелла, Б., Холл, Л. Дж. И Меткалф, К. Дж. Э. Микробиом за пределами экологической и эволюционной теории. Nat. Ecol. Evol. 1 , 1606–1615 (2017).
PubMed Статья PubMed Central Google Scholar
Моримото, Дж. И Балтрус, Д. А. Расширенный генотип: до какой степени? Комментарий к Carthey et al. Trends Ecol. Evol . 34 , 186–187 (2019).
Scheuring, I. & Yu, D. W. Как собрать полезный микробиом за три простых шага. Ecol. Lett. 15 , 1300–1307 (2012).
PubMed Статья PubMed Central Google Scholar
Докинз Р. Расширенный фенотип: долгая досягаемость гена . (Издательство Оксфордского университета, США, 1982).
Whitham, T. G. et al. Основы генетики сообществ и экосистем: от генов до экосистем. Nat. Преподобный Жене. 7 , 510–523 (2006).
CAS PubMed Статья PubMed Central Google Scholar
Мюллер, У. Г. и Сакс, Дж. Л. Разработка микробиомов для улучшения здоровья растений и животных. Trends Microbiol 23 , 606–617 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Зильбер-Розенберг, И. и Розенберг, Э. Роль микроорганизмов в эволюции животных и растений: теория эволюции гологенома. FEMS Microbiol. Ред. 32 , 723–735 (2008).
CAS PubMed Статья PubMed Central Google Scholar
Борденштейн, С. Р. и Тайс, К. Р. Биология хозяина в свете микробиома: десять принципов холобионтов и гологеномов. PLoS Biol. 13 , e1002226 (2015).
PubMed Статья CAS PubMed Central Google Scholar
Альберди А., Айзпуруа О., Боманн, К., Зепеда-Мендоза, М. Л. и Гилберт, М. Т. П. Обладают ли метагеномы кишечника позвоночных быстрой экологической адаптацией? Trends Ecol. Evol. 31 , 689–699 (2016).
PubMed Статья PubMed Central Google Scholar
Шапира, М. Кишечные микробиоты и эволюция хозяина: расширение симбиоза. Trends Ecol. Evol. 31 , 539–549 (2016).
PubMed Статья PubMed Central Google Scholar
Карриер, Т. Дж. И Райцель, А. М. Гологеном в окружающей среде и последствия ассоциированного с хозяином микробного репертуара. Фронт. Microbiol. 8 , 802 (2017).
PubMed Статья PubMed Central Google Scholar
Херст, Г. Д. Д. Расширенные геномы: симбиоз и эволюция. Интерфейс Focus 7 , 20170001 (2017).
Судакаран С., Кост, К. и Кальтенпот, М. Приобретение и замена симбионта как источник экологических инноваций. Trends Microbiol 25 , 375–390 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Карти, А.Дж. Р., Гиллингс, М. Р. и Блюмштейн, Д. Т. Расширенный генотип: обонятельная коммуникация, опосредованная микробами. Trends Ecol. Evol. 33 , 885–894 (2018).
PubMed Статья PubMed Central Google Scholar
Розенберг Э. и Зильбер-Розенберг И. Концепция эволюции гологенома через 10 лет. Микробиом 6 , 78 (2018).
PubMed Статья PubMed Central Google Scholar
Футуйма Д. Дж. И Морено Г. Эволюция экологической специализации. Annu. Rev. Ecol. Syst. 19 , 207–233 (1988).
Артикул Google Scholar
Пирсма Т. и Дрент Дж. Фенотипическая гибкость и эволюция конструкции организма. Trends Ecol. Evol. 18 , 228–233 (2003).
Артикул Google Scholar
Ланде, Р. Естественный отбор и случайный генетический дрейф в фенотипе. Evolution. Эволюция 30 , 314–334 (1976).
PubMed Статья PubMed Central Google Scholar
Вест-Эберхард, М. Дж. Фенотипическая пластичность и истоки разнообразия. Annu. Rev. Ecol. Syst. 20 , 249–278 (1989).
Артикул Google Scholar
Галамбор, К. К., Маккей, Дж. К., Кэрролл, С. П. и Резник, Д. Н. Адаптивная и неадаптивная фенотипическая пластичность и потенциал для современной адаптации в новых условиях. Функц. Ecol. 21 , 394–407 (2007).
Артикул Google Scholar
Bolnick, D. I. et al. Почему внутривидовые вариации признаков имеют значение в экологии сообщества. Trends Ecol. Evol. 26 , 183–192 (2011).
PubMed Статья PubMed Central Google Scholar
Меткалф, К. Дж. Э. и Коскелла, Б. Защитные микробиомы могут ограничивать эволюцию защиты хозяина от патогенов. Evol. Lett. 3 , 534–543 (2019).
PubMed Статья PubMed Central Google Scholar
Розенберг, Э., Корен, О., Решеф, Л., Ефроний, Р.И Зильбер-Розенберг, И. Роль микроорганизмов в здоровье кораллов, болезнях и эволюции. Nat. Rev. Microbiol. 5 , 355–362 (2007).
CAS PubMed Статья Google Scholar
Theis, K. R. et al. Правильная концепция хологенома: экоэволюционная структура для хозяев и их микробиомов. м Системы 1 , e00028 (2016).
Funkhouser, L.Дж. И Борденштейн, С. Р. Мама знает лучше всех: универсальность передачи микробов от матери. PLoS Biol. 11 , e1001631 (2013).
CAS PubMed Статья PubMed Central Google Scholar
Салем, Х., Флорез, Л., Херардо, Н. и Кальтенпот, М. Вне тела: внеклеточное измерение передачи мутуалистических бактерий у насекомых. Proc. R. Soc. Лондон.Б. 282 , 20142957 (2015).
Vacher, C. et al. Филосфера: микробные джунгли на границе раздела растений и климата. Annu. Rev. Ecol. Evol. Syst. 47 , 1–24 (2016).
Артикул Google Scholar
Rothschild, D. et al. В формировании микробиоты кишечника человека окружающая среда преобладает над генетикой хозяина. Природа 555 , 210–215 (2018).
ADS CAS PubMed Статья Google Scholar
Grieneisen, L.E. et al. Гены, геология и микробы: микробиота кишечника в гибридной зоне приматов объясняется свойствами почвы участка, а не видами хозяев. Proc. R. Soc. В: Биол. Sci. 286 , 201
(2019).
Артикул Google Scholar
МакКенни, Э.А., Келле, К., Данн, Р. Р. и Йодер, А. Д. Экосистемные услуги микробиомов животных. Мол. Ecol. 27 , 2164–2172 (2018).
CAS PubMed Статья Google Scholar
Спрокетт, Д., Фуками, Т. и Релман, Д. А. Роль приоритетных эффектов в ранней сборке кишечной микробиоты. Nat. Преподобный Гастроэнтерол. Гепатол. 15 , 197–205 (2018).
PubMed Статья PubMed Central Google Scholar
Ур, Г. Т., Дохналова, Л.И Тайсс, К. А. Измерение времени во взаимодействиях хозяина и микробиома. mSystems 4 , e00216–18 (2019).
ван Влит, С. и Доебели, М. Роль многоуровневого отбора в эволюции микробиома хозяина. Proc. Natl Acad. Sci. США 116 , 20591–20597 (2019).
PubMed Статья CAS PubMed Central Google Scholar
Бенсон, А.К. Микробиом кишечника — новая сложная черта. Nat. Genet. 48 , 1301 (2016).
CAS PubMed Статья PubMed Central Google Scholar
ван Опсталь, Э. Дж. И Борденштейн, С. Р. Переосмысление наследуемости микробиома. Наука 349 , 1172–1173 (2015).
ADS PubMed Статья Google Scholar
Beilsmith, K. et al. Полногеномные исследования ассоциации микробиома филлосферы: охват сложности взаимодействия хозяина и микроба. Завод J . 97 , 164–181 (2019).
Goodrich, J. K. et al. Человеческая генетика формирует микробиом кишечника. Ячейка 159 , 789–799 (2014).
CAS PubMed Статья PubMed Central Google Scholar
Гудрич, Дж.K. et al. Генетические детерминанты микробиома кишечника британских близнецов. Клеточный микроб-хозяин 19 , 731–743 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Ранний, А. М., Шанмугараджа, Н., Бучон, Н. и Кларк, А. Г. Генотип дрозофилы влияет на уровни комменсальных бактерий. PLoS ONE 12 , e0170332 (2017).
PubMed Статья CAS PubMed Central Google Scholar
Walters, W. A. et al. Крупномасштабное тиражируемое полевое исследование ризосферы кукурузы позволяет выявить наследственные микробы. Proc. Natl Acad. Sci. США 115 , 7368–7373 (2018).
PubMed Статья PubMed Central Google Scholar
Camarinha-Silva, A. et al. Влияние генома хозяина на микробный состав кишечника и микробное прогнозирование сложных признаков у свиней. Генетика 206 , 1637–1644 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Difford, G. F. et al. Генетика хозяина и микробиом рубца совместно связаны с выбросами метана у дойных коров. PLoS Genet 14 , e1007580 (2018).
PubMed Статья CAS PubMed Central Google Scholar
Кога, Р., Мэн, X.-Y., Tsuchida, T. & Fukatsu, T. Клеточный механизм избирательной вертикальной передачи симбионта облигатного насекомого на границе раздела бактериоцит-эмбрион. Proc. Natl Acad. Sci. США 109 , E1230 – E1237 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Нихолм С. В. и Макфолл-Нгаи М. Провеивание: установление симбиоза кальмаров и вибрионов. Nat. Rev. Microbiol. 2 , 632–642 (2004).
CAS PubMed Статья PubMed Central Google Scholar
Кальтенпот, М., Геттлер, В., Герцнер, Г. и Стром, Э. Симбиотические бактерии защищают личинок ос от заражения грибами. Curr. Биол. 15 , 475–479 (2005).
CAS PubMed Статья PubMed Central Google Scholar
Clark, R. I. et al. Отчетливые изменения в составе микробиоты во время старения дрозофилы нарушают функцию кишечника и приводят к смертности. Cell Rep. 12 , 1656–1667 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Franzosa, E.A. et al. Идентификация личных микробиомов с помощью метагеномных кодов. Proc. Natl Acad. Sci. США 112 , E2930 – E2938 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Lokmer, A. et al. Пространственная и временная динамика микробиоты гемолимфы тихоокеанских устриц в различных масштабах. Фронт. Microbiol. 7 , 1367 (2016).
PubMed Статья PubMed Central Google Scholar
Елена С. Ф., Ленски Р. Э. Эволюционные эксперименты с микроорганизмами: динамика и генетические основы адаптации. Nat. Преподобный Жене. 4 , 457–469 (2003).
CAS PubMed Статья PubMed Central Google Scholar
Лапьер, П. и Гогартен, Дж. П. Оценка размера бактериального пангенома. Trends Genet 25 , 107–110 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Кунин Э. В. и Вольф Ю. И. Эволюция микробов и вирусов: смена парадигмы в эволюционной биологии? Фронт. Клетка. Заразить. Microbiol. 2 , 119 (2012).
PubMed Статья PubMed Central Google Scholar
Феррейро А., Крук Н., Гаспаррини А. Дж. И Дантас Г. Мультимасштабная эволюционная динамика микробиомов, связанных с хозяином. Ячейка 172 , 1216–1227 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Кикучи, Ю., Хосокава, Т. и Фукацу, Т. Специфическое окно развития для установления симбиоза кишечника насекомых и микробов. Заявл. Environ. Microbiol. 77 , 4075–4081 (2011).
ADS CAS PubMed Статья PubMed Central Google Scholar
Kikuchi, Y. et al. Симбионт-опосредованная устойчивость к инсектицидам. Proc. Natl Acad. Sci. США 109 , 8618–8622 (2012).
ADS CAS PubMed Статья PubMed Central Google Scholar
Itoh, H. et al. Динамика заражения разлагающих инсектициды симбионтов от почвы к насекомым в ответ на распыление инсектицидов. ISME J. 12 , 909–920 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Коль, К. Д. и Диринг, М. Д. Микробиота кишечника лесной крысы как экспериментальная система для понимания микробного метаболизма пищевых токсинов. Фронт. Microbiol. 7 , 1165 (2016).
PubMed PubMed Central Google Scholar
Kohl, K. D., Weiss, R. B., Cox, J., Dale, C. & Dearing, M. D. Кишечные микробы травоядных млекопитающих способствуют поступлению токсинов растений. Ecol. Lett. 17 , 1238–1246 (2014).
PubMed Статья PubMed Central Google Scholar
Миллер, А. В., Коль, К. Д. и Деринг, М. Д. Желудочно-кишечный тракт белозвратного вудрата (Neotoma albigula) является местом обитания различных консорциумов оксалат-деградирующих бактерий. Заявл. Environ. Microbiol. 80 , 1595–1601 (2014).
ADS PubMed Статья CAS PubMed Central Google Scholar
Миллер А. В., Оаксон К. Ф., Дейл К. и Деринг М. Д. Влияние пищевого оксалата на микробиоту кишечника травоядного млекопитающего Neotoma albigula. Заявл. Environ. Microbiol. 82 , 2669–2675 (2016).
ADS CAS PubMed Статья PubMed Central Google Scholar
Коль, К. Д., Стенгель, А. и Деринг, М. Д. Инокуляция танин-деградирующих бактерий новым хозяевам увеличивает продуктивность при рационе, богатом танином. Environ. Microbiol. 18 , 1720–1729 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Коль, К. Д. и Деринг, М. Д. Опыт имеет значение: предварительное воздействие токсинов растений увеличивает разнообразие кишечных микробов у травоядных животных. Ecol. Lett. 15 , 1008–1015 (2012).
PubMed Статья PubMed Central Google Scholar
Редман Р. С., Шихан, К. Б., Стаут, Р. Г., Родригес, Р. Дж. И Хенсон, Дж. М. Термостойкость, вызванная симбиозом растений и грибов. Наука 298 , 1581 (2002).
CAS PubMed Статья PubMed Central Google Scholar
Rodriguez, R.J. et al. Стрессоустойчивость растений через симбиоз, адаптированный к среде обитания. ISME J. 2 , 404–416 (2008).
PubMed Статья PubMed Central Google Scholar
Миллер, Э. Т., Сванбэк, Р. и Боханнан, Б. Дж. М. Микробиомы как метасообщества: понимание микробов, связанных с хозяином, через экологию метасообщества. Trends Ecol. Evol. 33 , 926–935 (2018).
PubMed Статья PubMed Central Google Scholar
Ньюэлл П. Д. и Дуглас А. Е. Межвидовые взаимодействия определяют влияние кишечной микробиоты на распределение питательных веществ у Drosophila melanogaster. Заявл. Environ. Microbiol. 80 , 788–796 (2014).
ADS CAS PubMed Статья PubMed Central Google Scholar
Кибо, Э. С., Ямада, Р., Обадиа, Б., Лудингтон, В. Б. и Джа, В. В. Количество микробов влияет на питание, развитие и продолжительность жизни дрозофилы. Наука 4 , 247–259 (2018).
CAS Google Scholar
Gould, A. L. et al. Взаимодействие микробиома формирует приспособленность хозяина. Proc. Natl Acad. Sci. США 115 , E11951 – E11960 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Мушегян А.А., Вальзер Дж.-К., Саллам К.Э. и Эберт Д. Микробиота диапаузы: как ассоциации хозяина-микроба образуются после покоя у водных ракообразных. J. Anim. Ecol. 87 , 400–413 (2017).
PubMed Статья PubMed Central Google Scholar
Панке-Биссе, К., Пул, А.С., Гудрич, Дж. К., Лей, Р. Э. и Као-Книффин, Дж. Селекция на почвенные микробиомы выявляет воспроизводимые воздействия на функции растений. ISME J. 9 , 980–989 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Rolig, A. S., Parthasarathy, R., Burns, A. R., Bohannan, B. J. M. и Guillemin, K. Отдельные представители микробиоты непропорционально модулируют врожденные иммунные ответы хозяина. Клеточный микроб-хозяин 18 , 613–620 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Вебстер, Н. С. и Ройш, Т. Б. Х. Вклад микробов в устойчивость коралловых рифов. ISME J. 11 , 2167–2174 (2017).
PubMed Статья PubMed Central Google Scholar
Борн, Д. Г., Морроу, К. М. и Вебстер, Н. С. Анализ микробиома кораллов: обеспечение здоровья и устойчивости рифовых экосистем. Annu. Rev. Microbiol. 70 , 317–340 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Эйнсворт, Т. Д., Тербер, Р. В. и Гейтс, Р. Д. Будущее коралловых рифов: микробная перспектива. Trends Ecol. Evol. 25 , 233–240 (2010).
PubMed Статья PubMed Central Google Scholar
Sommer, F. et al. Микробиота кишечника регулирует энергетический обмен у зимующего бурого медведя Ursus arctos. Cell Rep. 14 , 1655–1661 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Меткалф, К. Дж. Э., Генри, Л. П., Реболледа-Гомес, М. и Коскелла, Б. Зачем нужно полагаться на микробиом для определения времени онтогенеза? МБио 10 , e01496-19 (2019).
Гилберт, С. Ф., Бош, Т. К. Г. и Ледон-Реттиг, К. Эко-Эво-Дево: симбиоз развития и пластичность развития как факторы эволюции. Nat. Преподобный Жене. 16 , 611–622 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Филиппы Т. и Сегер Дж. Хеджирование эволюционных ставок. Trends Ecol. Evol. 4 , 41–44 (1989).
CAS PubMed Статья PubMed Central Google Scholar
Storelli, G. et al. Lactobacillus plantarum способствует системному росту дрозофилы, модулируя гормональные сигналы посредством TOR-зависимого восприятия питательных веществ. Cell Metab. 14 , 403–414 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Брюйнинг, М., Генри, Л. П., Форсберг, С. К. Г., Меткалф, К. Дж. Э. и Эйролес, Дж. Ф. Когда микробиом определяет фенотип хозяина: отбор по вертикальной передаче в различных средах. bioRxiv 2020.09.02.280040 (2020) https://doi.org/10.1101/2020.09.02.280040.
Boone, C. K. et al. Бактерии, связанные с насекомыми-убийцами деревьев, снижают концентрацию защитных соединений растений. J. Chem. Ecol. 39 , 1003–1006 (2013).
CAS PubMed Статья PubMed Central Google Scholar
Berasategui, A. et al. Кишечная микробиота соснового долгоносика разрушает дитерпены хвойных растений и увеличивает приспособленность насекомых. Мол. Ecol. 26 , 4099–4110 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Ceja-Navarro, J. A. et al. Микробиота кишечника опосредует детоксикацию кофеина у основных насекомых-вредителей кофе. Nat. Commun. 6 , 7618 (2015).
ADS CAS PubMed Статья PubMed Central Google Scholar
Berasategui, A. et al. Микробиота кишечника соснового долгоносика похожа по всей Европе и напоминает микробиоту других хвойных жуков. Мол. Ecol. 25 , 4014–4031 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Сакс, Дж. Л., Скопхаммер, Р. Г. и Регус, Дж. У. Эволюционные переходы в бактериальном симбиозе. Proc. Natl Acad. Sci. США 108 , 10800–10807 (2011).
ADS CAS PubMed Статья PubMed Central Google Scholar
Обаяси Т.и другие. Кишечный орган насекомых для сортировки симбионтов. Proc. Natl Acad. Sci. США 112 , E5179 – E5188 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Itoh, H. et al. Специфичность «хозяин-симбионт», определяемая микробно-микробной конкуренцией в кишечнике насекомого. Proc. Natl Acad. Sci. США 116 , 22673–22682 (2019).
CAS PubMed Статья PubMed Central Google Scholar
Беннетт Г. М. и Моран Н. А. Наследственный симбиоз: преимущества и опасности эволюционной кроличьей норы. Proc. Natl Acad. Sci. 112 , 10169–10176 (2015).
ADS CAS PubMed Статья PubMed Central Google Scholar
Knight, R. et al. Лучшие практики для анализа микробиомов. Nat. Rev. Microbiol. 16 , 410–422 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Классен, Дж. Л. Определение функции микробиома. Nat. Microbiol 3 , 864–869 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Лау, Дж. А. и Леннон, Дж. Т. Быстрые реакции почвенных микроорганизмов улучшают приспособленность растений к новым условиям окружающей среды. Proc. Natl Acad. Sci. США 109 , 14058–14062 (2012).
ADS CAS PubMed Статья PubMed Central Google Scholar
Xu, L. et al. Засуха задерживает развитие микробиома корней сорго и увеличивает количество монодермальных бактерий. Proc. Natl Acad. Sci. США 115 , E4284 – E4293 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Чо И. и Блазер М. Дж. Микробиом человека: на стыке здоровья и болезни. Nat. Преподобный Жене. 13 , 260–270 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Райманн, К., Бобай, Л.-М. И Моран, Н. А. Антибиотики уменьшают генетическое разнообразие основных видов в микробиоме кишечника медоносной пчелы. Мол. Ecol. 27 , 2057–2066 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Дэвид, Л.A. et al. Диета быстро и воспроизводимо изменяет микробиом кишечника человека. Природа 505 , 559–563 (2014).
ADS CAS PubMed Статья PubMed Central Google Scholar
Циглер, М., Сенека, Ф. О., Юм, Л. К., Палумби, С. Р. и Вулстра, С. Р. Динамика бактериального сообщества связана с паттернами термостойкости кораллов. Nat. Commun. 8 , 14213 (2017).
ADS CAS PubMed Статья PubMed Central Google Scholar
Поллок, Дж., Глендиннинг, Л., Визедханвет, Т. и Уотсон, М. Безумие микробиома: попытка найти консенсусную «лучшую практику» для исследований микробиома 16S. Заявл. Environ. Microbiol. 84 , e02627–17 (2018).
PubMed Статья PubMed Central Google Scholar
Roth-Schulze, A. J. et al. Функциональная биогеография и специфичность бактериальных сообществ, ассоциированных с морскими зелеными водорослями Ulva spp. Мол. Ecol. 27 , 1952–1965 (2018).
PubMed Статья PubMed Central Google Scholar
Lozupone, C. A., Stombaugh, J. I., Gordon, J. I., Jansson, J. K. & Knight, R. Разнообразие, стабильность и устойчивость микробиоты кишечника человека. Природа 489 , 220–230 (2012).
ADS CAS PubMed Статья PubMed Central Google Scholar
Меден, С., Меткалф, К. Дж. Э. и Коскелла, Б. Влияние возраста хозяина и пространственного положения на состав бактериального сообщества дуба английского (Quercus robur). Environ. Microbiol. Реп. 8 , 649–658 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Lambais, M. R., Barrera, S. E., Santos, E. C., Crowley, D. E., Jumpponen, A. Метапротеомы деревьев из бразильского атлантического леса демонстрируют высокий уровень функциональной избыточности. Microb. Ecol. 73 , 123–134 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Квонг, В. К. и Моран, Н. А. Сообщества кишечных микробов социальных пчел. Nat.Rev. Microbiol. 14 , 374–384 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Oh, J. et al. Временная стабильность микробиома кожи человека. Ячейка 165 , 854–866 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Гаруд, Н. Р., Гуд, Б.Х., Халлатчек, О. и Поллард, К. С. Эволюционная динамика бактерий в микробиоме кишечника внутри и между хозяевами. PLoS Biol. 17 , e3000102 (2019).
PubMed Статья PubMed Central Google Scholar
Asnicar, F. et al. Изучение вертикальной передачи микробиома от матери младенцу с помощью метагеномного профилирования на уровне штаммов. mSystems 2 , e00164–16 (2017).
Yassour, M. et al. Штаммовый анализ передачи бактерий от матери ребенку в течение первых нескольких месяцев жизни. Клеточный микроб-хозяин 24 , 146–154.e4 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Фирер, Н. Принятие неизвестного: раскрытие сложностей микробиома почвы. Nat. Rev. Microbiol. 15 , 579–590 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Gilbert, J. A. et al. Современное понимание микробиома человека. Nat. Med. 24 , 392–400 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Abubucker, S. et al. Метаболическая реконструкция метагеномных данных и ее применение в микробиоме человека. PLoS Comput. Биол. 8 , e1002358 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Jorth, P. et al. Метатранскриптомика микробиома ротовой полости человека во время здоровья и болезни. MBio 5 , e01012 – e01014 (2014).
PubMed Статья PubMed Central Google Scholar
Башиардес, С., Зильберман-Шапира, Г. и Элинав, Э. Использование метатранскриптомики в исследовании микробиома. Bioinform. Биол. Insights 10 , 19–25 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Franzosa, E.A. et al. Связь метатранскриптома и метагенома кишечника человека. Proc. Natl Acad. Sci. США 111 , E2329 – E2338 (2014).
CAS PubMed Статья PubMed Central Google Scholar
Abu-Ali, G. S. et al. Метатранскриптом фекальных микробных сообществ человека в когорте взрослых мужчин. Nat. Microbiol. 3 , 356–366 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Хоанг, К. Л., Морран, Л. Т. и Херардо, Н.М. Экспериментальная эволюция как малоиспользуемый инструмент для изучения полезных взаимодействий животных и микробов. Фронт. Микробиол . 7 , 1444 (2016).
Martino, M. E. et al. Бактериальная адаптация к рациону хозяина — ключевая эволюционная сила, формирующая симбиоз дрозофилы и лактобацилл. Клеточный микроб-хозяин 24 , 109–119.e6 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Шлёттерер, К., Кофлер, Р., Версаче, Э., Тоблер, Р. и Франссен, С. У. Сочетание экспериментальной эволюции с секвенированием следующего поколения: мощный инструмент для изучения адаптации к существующей генетической изменчивости. Наследственность 114 , 431–440 (2015).
PubMed Статья PubMed Central Google Scholar
Генри, Л. П. и Эйролес, Дж. Ф. Мета-анализ показывает, что микробиом реагирует на эксперименты Evolve и Resequence у Drosophila melanogaster. BMC Microbiol 21 , 108 (2021).
CAS PubMed Статья PubMed Central Google Scholar
Wagner, M. R. et al. Природные почвенные микробы изменяют фенологию цветения и интенсивность отбора во время цветения у дикого родственника Arabidopsis. Ecol. Lett. 17 , 717–726 (2014).
PubMed Статья PubMed Central Google Scholar
Хендри, А. П. Экоэволюционная динамика . (Издательство Принстонского университета, 2017).
Bang, C. et al. Метаорганизмы в экстремальных условиях окружающей среды: играют ли микробы роль в адаптации организма? Зоология 127 , 1–19 (2018).
PubMed Статья PubMed Central Google Scholar
Hoyt, J. R. et al. Бактерии, выделенные от летучих мышей, подавляют рост Pseudogymnoascus destructans, возбудителя синдрома белого носа. PLoS ONE 10 , e0121329 (2015).
PubMed Статья CAS PubMed Central Google Scholar
Cheng, T. L. et al. Эффективность пробиотической бактерии для лечения летучих мышей, пораженных синдромом белого носа. J. Appl. Ecol. 54 , 701–708 (2016).
Артикул Google Scholar
Вудхэмс, Д.К., Блетц, М., Куенеман, Дж. И Маккензи, В. Управление болезнями земноводных с помощью микробиоты кожи. Trends Microbiol. 24 , 161–164 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Вайс, Б. и Аксой, С. Влияние микробиома на компетентность насекомых-переносчиков. Trends Parasitol. 27 , 514–522 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Цитвогель, Л., Ма, Ю., Рауль, Д., Кремер, Г., Гаевски, Т. Ф. Микробиом в иммунотерапии рака: инструменты диагностики и терапевтические стратегии. Наука 359 , 1366–1370 (2018).
ADS CAS PubMed Статья PubMed Central Google Scholar
Toju, H. et al. Основные микробиомы для устойчивых агроэкосистем. Nat. Растения 4 , 247–257 (2018).
PubMed Статья PubMed Central Google Scholar
Костелло, Э. К., Стагаман, К., Детлефсен, Л., Боханнан, Б. Дж. М. и Релман, Д. А. Применение экологической теории к пониманию микробиома человека. Наука 336 , 1255–1262 (2012).
ADS CAS PubMed Статья PubMed Central Google Scholar
Кристиан Н., Уитакер Б. К. и Клэй К. Микробиомы: объединение систем животных и растений через призму теории экологии сообществ. Фронт. Microbiol. 6 , 869 (2015).
PubMed Статья PubMed Central Google Scholar
Trevelline, B. K., Fontaine, S. S., Hartup, B. K. & Kohl, K. D. Биология сохранения нуждается в микробном возрождении: призыв к рассмотрению связанной с хозяином микробиоты в практике управления дикой природой. Proc. Биол. Sci. 286 , 20182448 (2019).
PubMed PubMed Central Google Scholar
Мюллер, Э. А., Вишноски, Н. И., Перальта, А. Л., Леннон, Дж. Т. Эффекты спасения микробов: как микробиомы могут спасти хозяев от вымирания. Функц. Экол . 34 , 2055-2064 (2020).
Хирд, С. М. Эволюционная биология нуждается в диких микробиомах. Фронт.Microbiol. 8 , 725 (2017).
PubMed Статья PubMed Central Google Scholar
Уэйд, М. Дж. Коэволюционная генетика экологических сообществ. Nat. Преподобный Жене. 8 , 185–195 (2007).
CAS PubMed Статья PubMed Central Google Scholar
Хаммер, Т. Дж., Янзен, Д. Х., Hallwachs, W., Jaffe, S.P. & Fierer, N. Гусеницы не имеют постоянного микробиома кишечника. Proc. Natl Acad. Sci. 114 , 9641–9646 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Хаммер, Т. Дж., Сандерс, Дж. Г. и Фирер, Н. Не всем животным нужен микробиом. FEMS Microbiol. Lett . 366 , fnz117 (2019).
Хит, К.Д. и Стинчкомб, Дж. Р. Объяснение вариации мутуализма: новый эволюционный парадокс? Evolution 68 , 309–317 (2014).
PubMed Статья PubMed Central Google Scholar
Сандовал-Мотта, С., Алдана, М., Мартинес-Ромеро, Э. и Франк, А. Микробиом человека и проблема отсутствующей наследственности. Фронт. Genet. 8 , 80 (2017).
PubMed Статья CAS PubMed Central Google Scholar
Wallace, R.J. et al. Наследственная подгруппа основного микробиома рубца определяет продуктивность дойных коров и выбросы. Sci. Adv. 5 , eaav8391 (2019).
ADS CAS PubMed Статья PubMed Central Google Scholar
Vollmar, S. et al. Микробная архитектура кишечника характеристик продуктивности у модельного вида домашней птицы японский перепел (Coturnix japonica), оцененная с помощью смешанных линейных моделей. G3 10 , 2553–2562 (2020).
CAS PubMed Статья PubMed Central Google Scholar
Хоффманн, А. А., Сгро, К. М. и Кристенсен, Т. Н. Пересмотр адаптивного потенциала, размера популяции и сохранения. Trends Ecol. Evol. 32 , 506–517 (2017).
PubMed Статья PubMed Central Google Scholar
Bruijning, M., Metcalf, C.J.E., Jongejans, E. & Ayroles, J.F. Эволюция управления дисперсией. Trends Ecol. Evol. 35 , 22–33 (2020).
PubMed Статья PubMed Central Google Scholar
Дуглас, Г. М., Белявски, Дж. П. и Лангиль, М. Г. И. Переоценка взаимосвязи между отсутствующей наследуемостью и микробиомом. Микробиом 8 , 87 (2020).
PubMed Статья PubMed Central Google Scholar
Йоханнес, Ф., Колот, В. и Янсен, Р. С. Динамика эпигенома: перспектива количественной генетики. Nat. Преподобный Жене. 9 , 883–890 (2008).
CAS PubMed Статья PubMed Central Google Scholar
Слаткин М. Эпигенетическая наследственность и проблема отсутствующей наследственности. Генетика 182 , 845–850 (2009).
PubMed Статья PubMed Central Google Scholar
Эрнандо-Эрраес, И., Гарсия-Перес, Р., Шарп, А. Дж. И Маркес-Бонет, Т. Метилирование ДНК: понимание эволюции человека. PLoS Genet 11 , e1005661 (2015).
PubMed Статья CAS PubMed Central Google Scholar
Pujol, B. et al. Отсутствующий ответ на отбор в дикой природе. Trends Ecol. Evol. 33 , 337–346 (2018).
PubMed Статья PubMed Central Google Scholar
Шоу Р. Г. От прошлого к будущему: учитывая ценность и пределы эволюционного предсказания. г. Nat. 193 , 1–10 (2019).
PubMed Статья PubMed Central Google Scholar
Гаруд, Н. Р., Гуд, Б. Х., Халлатчек, О. и Поллард, К. С. Эволюционная динамика бактерий в микробиоме кишечника внутри и между хозяевами. PLoS Biol. 17, e3000102 (2019).
Truong, D. T., Tett, A., Pasolli, E., Huttenhower, C. & Segata, N. Структура популяции микробных штаммов на уровне штаммов и генетическое разнообразие из метагеномов. Genome Res. 27 , 626–638 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Спор А., Корен О. и Лей Р. Выявление влияния окружающей среды и генотипа хозяина на микробиом кишечника. Nat. Rev. Microbiol. 9 , 279–290 (2011).
CAS PubMed Статья Google Scholar
Guo, Y. et al. Сети, лежащие в основе симбиоза, выявлены посредством межвидового сопоставления eQTL. Генетика 206 , 2175–2184 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Kohl, K. D. Вводное практическое руководство по включению исследований микробиома в интегративную и сравнительную биологию. Integr. Комп. Биол. 57 , 674–681 (2017).
PubMed Статья Google Scholar
Марчези, Дж. Р. и Равель, Дж. Словарь исследования микробиома: предложение. Микробиом 3 , 31 (2015).
PubMed Статья PubMed Central Google Scholar
Взгляд с конференции Mountain Lake по развитию и эволюции на JSTOR
АбстрактныйОграничения развития (определяемые как предубеждения в отношении производства вариантных фенотипов или ограничения фенотипической изменчивости, вызванные структурой, характером, составом или динамикой системы развития), несомненно, играют важную роль в эволюции.Тем не менее, нет единого мнения об их важности по сравнению с отбором, дрейфом и другими подобными факторами в формировании эволюционной истории. В этом обзоре проводится различие между «универсальными» и «локальными» ограничениями; в первую очередь речь идет о последних, которые применимы к ограниченному кругу таксонов. Такие ограничения, как правило, могут быть нарушены даже в рамках тех таксонов, к которым они применяются, хотя и с разной степенью сложности. Обсуждается происхождение ограничений, в явном виде рассматриваются пять отличительных особенностей ограничений.Изложены три средства выявления ограничений, а также четыре средства различения ограничений, связанных с развитием, от ограничений избирательного характера. Ни одно из последних (использование априорных адаптивных прогнозов, прямое измерение отбора, прямое измерение наследственной изменчивости и использование сравнительного метода) не является надежным. В заключительном разделе обсуждаются три более крупных вопроса, касающихся роли ограничений развития в эволюции: степень, в которой эволюционный застой может быть объяснена с точки зрения развития, степень, в которой эволюционные тенденции и закономерности могут быть следствием ограничений развития, и степень, в которой различные генетические механизмы и механизмы развития эволюционировали в силу необходимости клонов проявлять эволюционную пластичность (или приспособляемость), если они хотят выжить.Хотя окончательных выводов по этим более широким вопросам не сделано, мы используем последние достижения в биологии развития, теории эволюции и (в ограниченной степени) молекулярной биологии.
Информация о журналеТекущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. Ежеквартальный обзор биологии (QRB) представляет проницательные исторические, философские и технические трактовки важных биологических тем с 1926 года.Как ведущий обзорный журнал по биологии, QRB публикует выдающиеся обзорные статьи большого объема, которые руководствуются обширным, всеобъемлющим и часто гуманистическим пониманием биологии. Помимо основных биологических наук, QRB также является важным обзорным журналом для ученых в смежных областях, включая политические исследования, историю и философию науки. Обширный раздел обзоров новых книг по биологии предоставляет преподавателям и исследователям информацию о последних публикациях из жизни
Информация об издателеС момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета, University of Chicago Press взяла на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, которые способствуют образованию, способствуют общественному пониманию. , и обогатить культурную жизнь.Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Исследователи отслеживают, как микробиомные бактерии адаптируются к человеку посредством передачи — ScienceDaily
Полезные бактерии в кишечном микробиоме передаются от одного человека к другому разными способами, что влияет на их численность в кишечнике и функции, которые они обеспечивают.
В этом новом исследовании, проведенном учеными из института Wellcome Sanger, использовалось генетическое секвенирование, чтобы пролить новый свет на эволюцию, колонизацию и передачу кишечных бактерий, которые играют большую роль в здоровье человека.
Исследование, опубликованное сегодня (5 августа 2021 года) в Genome Biology , обеспечивает более глубокое понимание эволюции бактерий в микробиоме человека. Это также может послужить источником информации для разработки терапевтических средств на основе микробиома, где ключевые бактерии могут быть выбраны для лечения различных заболеваний, связанных с кишечником.
Люди населены чрезвычайно большим количеством микроорганизмов, называемых микробиомом, которые включают бактерии, вирусы, археи и грибы. Полезные бактерии в организме примерно соответствуют человеческим клеткам один к одному *, а в кишечнике этот набор бактерий играет важную роль для здоровья человека. Состав микробиома кишечника влияет на иммунную систему, предотвращает инфекции и поддерживает организм, расщепляя некоторые углеводы, с которыми человеческие клетки не могут справиться в одиночку.
Чтобы виды бактерий в кишечнике выжили, они должны иметь возможность передаваться от одного человека к другому.Кишечные бактерии в основном анаэробны, то есть они не могут выжить в кислороде. Для распространения некоторые из них производят споры, которые иногда сравнивают с семенами и могут бездействовать, пока не попадут в подходящие условия для роста.
В ходе этого исследования, проведенного Институтом Велкома Сэнгера, была изучена группа бактерий, известных как Firmicutes, которые доминируют в микробиоме человека и продуцируют споры. Исследователи проанализировали геномы 1358 Firmicutes и проследили эволюцию определенных характеристик, включая образование спор.Они заметили, что кишечные бактерии, образующие споры, были обнаружены в более низком количестве в кишечнике и имели больший геном по сравнению с теми, которые утратили способность производить споры (спорулят). В их геномах также было больше генов, связанных с метаболизмом углеводов и биосинтезом витаминов, что говорит о том, что у них есть важные метаболические функции.
Бактерии Firmicutes, которые больше не могли образовывать споры, имели меньшие геномы, но в большей степени присутствовали в кишечнике и имели более специализированный метаболизм, основанный на анализе генома.Они также были менее распространены в общей популяции, что означает, что они были обнаружены у меньшего числа людей, что свидетельствует о том, что потеря споруляции ограничивает их способность к широкому распространению.
Меньшие геномы и более специализированный метаболизм указывают на то, что бактерии, утратившие способность образовывать споры, становятся более приспособленными к своему человеческому хозяину, что может позволить им колонизировать более высокие уровни в кишечнике. С другой стороны, бактерии, которые все еще производят споры, кажутся менее приспособленными для людей из-за их более крупных геномов, что может объяснить, почему их не так много в кишечнике.
Эти различия показывают, что передача является важным процессом, определяющим эволюцию кишечных бактерий, и теперь необходимы дальнейшие исследования, чтобы продолжить изучение связи между передачей кишечных бактерий и ролью, которую они играют в здоровье человека. Понимание этих процессов может помочь информировать терапевтов, например, выяснить, можно ли давать людям определенные бактерии в зависимости от их способности к колонизации и как различный метаболизм этих бактерий может повлиять на состояние здоровья и лечение.
Доктор Хилари Браун, первый автор и штатный научный сотрудник Института Велкома Сэнгера, сказала: «Несмотря на то, что передача кишечных бактерий между людьми важна для их выживания, генетические и биологические особенности бактерий, которые позволяют им это делать, все еще сохраняются. Это исследование начинает разгадывать эту загадку, анализируя геномы и обнаруживая, что способность бактерий производить споры терялась несколько раз, что влияло на их эволюцию и функции.Необходимо продолжить изучение генетических деталей микробиома, чтобы понять роль конкретных бактерий и то, как их отсутствие может повлиять на здоровье человека ».
Доктор Тревор Лоули, старший автор и старший руководитель группы в Wellcome Sanger Institute, сказал: «Микробиом играет важную роль в здоровье и развитии человека и влияет на широкий спектр физиологических функций в организме. Подробнее о бактериях, которые населяют нас, и то, как они адаптируются к жизни в людях благодаря своему метаболизму, будет иметь важное значение для разработки новых методов лечения и диагностики заболеваний, опосредованных микробиомом.