
Клеточная мембрана строение: 1_1 Строение клеточной мембраны | Кинезиолог
1_1 Строение клеточной мембраны | Кинезиолог
Клеточная мембрана (плазм
алемма или плазмолемма)Определение понятия
Клеточная мембрана (синонимы: плазмалемма, плазмолемма, цитоплазматическая мембрана, биомембрана) — это тройная липопротеиновая (т.е. «жиро-белковая») оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен и связь между клеткой и окружающей её средой.
Главное в этом определении — не то, что мембрана отделяет клетку от среды, а как раз то, что она соединяет клетку с окружающей средой. Мембрана — это активная структура клетки, она постоянно работает.
Биологическая мембрана — это ультратонкая бимолекулярная пленка фосфолипидов, инкрустированная белками и полисахаридами. Эта клеточная структура лежит в основе барьерных, механических и матричных свойств живого организма (Антонов В.Ф., 1996).
Образное представление о мембране
Мне клеточная мембрана представляетсся в виде решетчатого забора с множеством дверей в нём, который окружает некую территорию. Всякая мелкая живность может через этот забор свободно перемещаться туда и обратно. Но более крупные посетители могут входить только через двери, да и то не всякие. У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей — транспортных механизмов мембраны!
Всякая мелкая живность может через этот забор свободно перемещаться туда и обратно. Но более крупные посетители могут входить только через двери, да и то не всякие. У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей — транспортных механизмов мембраны!
Свойства мембраны
1. Проницаемость.
2. Полупроницаемость (частичная проницаемость).
3. Избирательная (синоним: селективная) проницаемость.
4. Активная проницаемость (синоним: активный транспорт).
5. Управляемая проницаемость.
Как видим, основное свойство мембраны — это её проницаемость по отношению к различным веществам.
6. Фагоцитоц и пиноцитоз.
7. Экзоцитоз.
8. Наличие электрических и химических потенциалов, точнее разности потенциалов между внутренней и наружной сторонами мембраны. Образно можно сказать, что «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками». Подробности: смотреть тут.
9. Изменения электрического и химического потенциала.
10. Раздражимость. Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лигандов (управляющих веществ). Важно отметить, что сигнальное вещество воздействует на рецептор снаружи, а изменения продолжаются внутри клетки. Получается, что мембрана передала информацию из окружающей среды во внутреннюю среду клетки.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
12. Изменение формы поверхности и её площади. Это позволяет мембране образовывать выросты наружу или, наоборот, впячивания внутрь клетки.
13. Способность образовывать контакты с другими клеточными мембранами.
14. Адгезия — способность прилипать к твёрдым поверхностям.
Краткий список свойств мембраны
- Проницаемость.
- Эндоцитоз, экзоцитоз, трансцитоз.
- Потенциалы.
- Раздражимость.
- Ферментная активность.
- Контакты.
- Адгезия.
Функции мембраны
1. Неполная изоляция внутреннего содержимого от внешней среды.
2. Главное в работе клеточной мембраны — это обмен различными веществами между клеткой и межклеточной средой. Этому служит такое свойство мембраны как проницаемость. Кроме того, мембрана регулирует этот обмен за счёт того, что регулирует свою проницаемость.
3. Ещё одна важная функция мембраны — создание разности химических и электрических потенциалов между её внутренней и наружной сторонами. За счёт этого внутри клетка имеет отрицательный электрический потенциал — потенциал покоя.
За счёт этого внутри клетка имеет отрицательный электрический потенциал — потенциал покоя.
4. Через мембрану осуществляется также информационный обмен между клеткой и окружающей её средой. Специальные молекулярные рецепторы, расположенные на мембране, могут связываться с управляющими веществами (гормонами, медиаторами, модуляторами) и запускать в клетке биохимические реакции, приводящие к различным изменениям в работе клетки или в её структурах.
Видео: Строение мембраны клетки
Видеолекция: Подробно о строении мембраны и транспорте
Строение мембраны
Клеточная мембрана имеет универсальное трёхслойное строение. Её срединный жировой слой является сплошным, а верхний и нижний белковые слои покрывают его в виде мозаики из отдельных белковых участков. Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами — транспортёрами и ионными каналами. Зато важнейшие для всего живого газы — кислород и углекислый газ — легко перемещаются через мембрану как внутрь клетки, так и наружу.
Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами — транспортёрами и ионными каналами. Зато важнейшие для всего живого газы — кислород и углекислый газ — легко перемещаются через мембрану как внутрь клетки, так и наружу.
Ниже представлены микрофотографии реальных клеточных мембран контактирующих клеток, полученные с помощью электронного микроскопа, а также схематический рисунок, показывающий трёхслойность мембраны и мозаичность её белковых слоёв. Для увеличения изображения кликните на него.
Отдельное изображение внутреннего липидного (жирового) слоя клеточной мембраны, пронизанного интегральными встроенными белками. Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя
Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя
Рисунок выше: Неполное схематичное изображение клеточной мембраны (клеточной оболочки), приведённое в Википедии.
Учтите, что наружный и внутренний слои поверхностных белков здесь с мембраны сняты, чтобы нам лучше был виден центральный жировой двойной липидный слой. В реальной клеточной мембране сверху и снизу по жировой плёночке (мелкие шарики на рисунке) плавают большие белковые «острова», и мембрана получается более толстой, трёхслойной: белок-жир-белок. Так что она на самом деле похожа на сэндвич из двух белковых «кусков хлеба» с жирным слоем «масла» посередине, т.е. имеет трёхслойное строение, а не двухслойное.
На этом рисунке маленькие голубые и белые шарики соответствуют гидрофильным (смачиваемым) «головкам» липидов, а присоединённые к ним «ниточки» — гидрофобным (несмачиваемым) «хвостам». Из белков показаны только интегральные сквозные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс. Гликокаликс — это как бы углеводный («сахарный») «пушок» на мембране, образованный торчащими из неё длинными белково-углеводными молекулами.
Желтые овальные точки внутри мембраны — это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс. Гликокаликс — это как бы углеводный («сахарный») «пушок» на мембране, образованный торчащими из неё длинными белково-углеводными молекулами.
Модель цитоплазматической мембраны: Перейти для просмотра
Живая клетка — это маленький «белково-жировой мешочек», заполненный полужидким желеобразным содержимым, которое пронизано плёнками и трубочками.
Стенки этого мешочка образованы двойной жировой (липидной) плёночкой, облепленной изнутри и снаружи белками — клеточной мембраной. Поэтому говорят, что мембрана имеет трёхслойное строение: белки-жиры-белки. Внутри клетки также есть множество подобных жировых мембран, которые делят её внутреннее пространство на отсеки (=компартменты). Такими же мембранами окружены клеточные органеллы: ядро, митохондрии, хлоропласты. Так что мембрана — это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.
Так что мембрана — это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.
Слева — уже не реальная, а искусственная модель кусочка биологической мембраны: это мгновенный снимок жирового фосфолипидного бислоя (т.е. двойного слоя) в процессе его молекулярно-динамического моделирования. Показана расчётная ячейка модели — 96 молекул ФХ (фосфатидилхолина) и 2304 молекулы воды, всего 20544 атомов.
Справа — наглядная модель одиночной молекулы того самого липида, из которых как раз и собирается мембранный липидный бислой. Вверху у него гидрофильная (водолюбивая) головка, а снизу — два гидрофобных (боящихся воды) хвостика. У этого липида есть простое название: 1-стероил-2-докозагексаеноил-Sn-глицеро-3-фосфатидилхолин (18:0/22:6(n-3)cis ФХ), но вам нет нужды его запоминать, если вы только не планируете довести своего преподавателя до обморока глубиной своих познаний.
Можно дать и более точное научное определение клетке:
Клетка – это ограниченная активной мембраной, упорядоченная, структурированная неоднородная система биополимеров, участвующих в единой совокупности обменных, энергетических и информационных процессов, и также осуществляющих поддержание и воспроизведение всей системы в целом.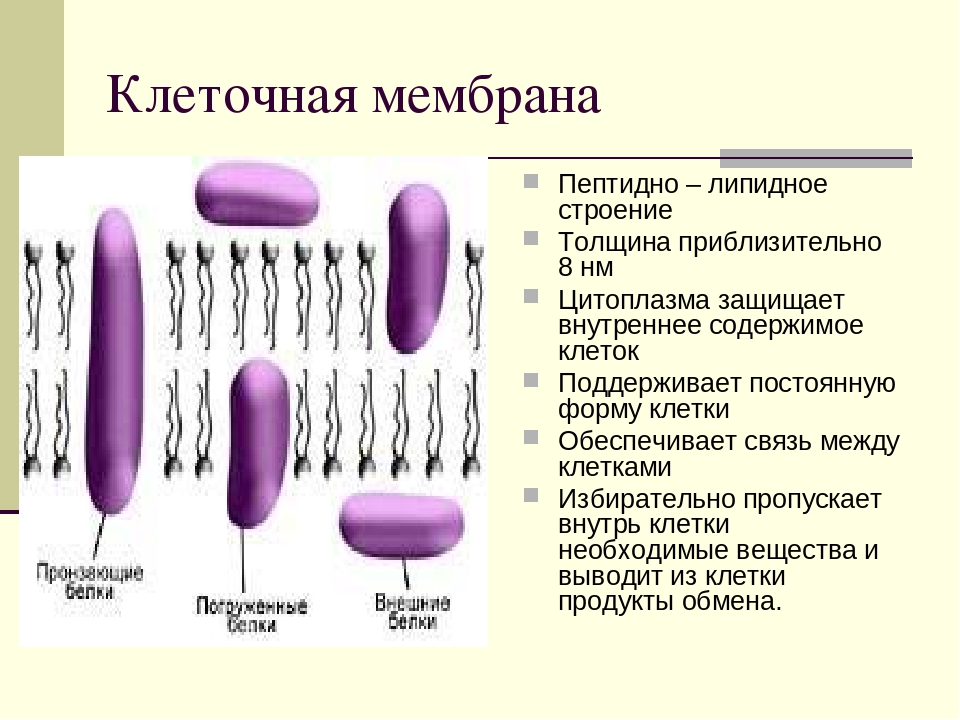
Внутри клетка также пронизана мембранами, а между мембранами находится не вода, а вязкий гель/золь изменяемой плотности. Поэтому взаимодействующие молекулы в клетке не плавают свободно, как в пробирке с водным раствором, а в основном сидят (иммобилизованы) на полимерных структурах цитоскелета или внутриклеточных мембранах. И химические реакции поэтому проходят внутри клетки почти как в твердом теле, а не в жидкости. Наружная мембрана, окружающая клетку, также облеплена ферментами и молекулярными рецепторами, что делает её очень активной частью клетки.
Клеточная мембрана (плазмалемма, плазмолемма) — это активная оболочка, отделяющая клетку от окружающей среды и связывающая её с окружающей средой. © Сазонов В.Ф., 2016.
Из этого определения мембраны следует, что она не просто ограничивает клетку, а активно работает, связывая её с окружающей её средой.
Мембранные липиды
Жир, из которого состоят мембраны, — особенный, поэтому его молекулы принято называть не просто жиром, а «липидами», «фосфолипидами», «сфинголипидами».
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин, а также в меньших количествах гликолипиды.
С химической точки зрения фосфолипид состоит из четырёх частей: глицерина, двух жирных кислот с длинной углеводородной цепью, фосфорной кислоты и особой для каждого фосфолипида группы, которую принято называть характеристической группой. Трёхатомный спирт глицерин связывает через сложно-эфирную связь две жирные кислоты и остаток фосфорной кислоты, к которой присоединена характеристическая группа (например, этаноламин).
Рис. ___. Структурная формула фосфатидилэтаноламина как пример амфифильной (гидрофобной/гидрофильной) молекулы фосфолипида. Кроме этаноламина характеристической группой фосфолипида может быть также холин, инозитол, серин и некоторые другие молекулы.
Рис. ___. Молекулярная структура фосфатидилхолина (=лецитина). Источник изображения: https://pandia.ru/text/80/650/73429-4.php
Мембранная плёночка является двойной, т. е. она состоит из двух липидных плёночек, слипшихся друг с другом с помощью своих липидных «хвостиков». Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из «бислоя«, т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая — не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами. Примерно так можно соединить две щётки, направив их щетиной друг к другу и слегка придавив.
е. она состоит из двух липидных плёночек, слипшихся друг с другом с помощью своих липидных «хвостиков». Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из «бислоя«, т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая — не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами. Примерно так можно соединить две щётки, направив их щетиной друг к другу и слегка придавив.
Мембранные белки
Белки мембраны включены в липидный двойной слой двумя способами:
- Гидрофильные радикалы аминокислот поверхностных мембранных белков связаны нековалентными связями с гидрофильной поверхностью липидного бислоя.
- Интегральные мембранные белки погружены в гидрофобную область бислоя.
Интегральные белки различаются по степени погруженности в гидрофобную часть бислоя. Они могут располагаться по обеим сторонам мембраны и при этом либо частично погружаются в мембрану, либо располагаются трансмембранно. Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связана с моносахаридными остатками или олигосахаридными цепями и представляет собой гликопротеины. В отличие от нерастворимых фибриллярных белков растворимые белки имеют почти сферическую (глобулярную) форму. Глобулярным белкам свойственна высокоупорядоченная пространственная структура (конформация), которая способствует выполнению специфических биологических функций (Албертс и соавт., 1994).
Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связана с моносахаридными остатками или олигосахаридными цепями и представляет собой гликопротеины. В отличие от нерастворимых фибриллярных белков растворимые белки имеют почти сферическую (глобулярную) форму. Глобулярным белкам свойственна высокоупорядоченная пространственная структура (конформация), которая способствует выполнению специфических биологических функций (Албертс и соавт., 1994).
Подвижными в мембране являются не только липиды, но и мембранные белки. Если белки не закреплены в мембране, они «плавают» в липидном бислое как в жидкости. Поэтому говорят, что биомембраны имеют жидкостно-мозаичную структуру. При этом «дрейф» белков в плоскости мембраны происходит достаточно легко, переход их с внешней стороны мембраны на внутреннюю («флип-флоп») невозможен, а переход липидов происходит крайне редко. Для «перескока» липидов необходимы специальные белки транслокаторы. Исключение составляет жир холестерин, который может легко переходить с одной стороны мембраны на другую. Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением. Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря» и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками. У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21–25 преимущественно гидрофобных аминокислот, которые образуют правую трансмембранную α-спираль с 6 или 7 витками (Фалер, Шилдс, 2004).
При этом «дрейф» белков в плоскости мембраны происходит достаточно легко, переход их с внешней стороны мембраны на внутреннюю («флип-флоп») невозможен, а переход липидов происходит крайне редко. Для «перескока» липидов необходимы специальные белки транслокаторы. Исключение составляет жир холестерин, который может легко переходить с одной стороны мембраны на другую. Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением. Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря» и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками. У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21–25 преимущественно гидрофобных аминокислот, которые образуют правую трансмембранную α-спираль с 6 или 7 витками (Фалер, Шилдс, 2004).
Мембрана бактерий
Оболочка прокариотической клетки грамотрицательных бактерий состоит из нескольких слоёв, показанных на рисунке ниже.
Слои оболочки грамотрицательных бактерий:
1. Внутренняя трёхслойная цитоплазматическая мембрана, которая соприкасается с цитоплазмой.
2. Клеточная стенка, которая состоит из муреина.
3. Наружная трёхслойная цитоплазматическая мембрана, которая имеет такую же систему липидов с белковыми комплексами, как и внутренняя мембрана.
Общение грамотрицательных бактериальных клеток с внешним миром через такую сложную трёхступенчатую структуру не даёт им преимущества в выживании в суровых условиях по сравнению с грамположительным бактериями, имеющими менее мощную оболочку. Они точно так же плохо переносят высокие температуры, повышенную кислотность и перепады давления.
Рис. Сложная тройная клеточная оболочка грамотрицательных бактерий. Источник изображения: https://probakterii.ru/prokaryotes/organelles/membrana-bakterij.html
Рис. Сравнение оболочек грамположительных и грамотрицательных бактерий. Источник изображения: https://myslide.ru/presentation/512325_skachat-stroenie-bakterialnoj-kletki
Рис . Рафтовые неоднородности в мембране различного масштаба. а — Нанокластеры холестерола, сфингомиелина, гликосфинголипидов и белков плазматической мембраны различаются по составу. Считается, что в эти кластеры входят ГФИ-заякоренные белки, трансмембранные (ТМ) белки, специфичные для рафтов, и цитоплазматические белки, связанные с актиновыми филаментами. «Обычные» ТМ-белки не входят в состав рафтов. б — В ответ на внешние сигналы нанокластеры могут сливаться с образованием рафтовой платформы, важной для ТМ передачи сигналов и мембранного транспорта. в — Рафтовая фаза, видимая в микроскоп (ø ≈1 мкм), наблюдается исключительно в равновесных мембранных системах, таких как гигантские синтетические или мембранные везикулы. В «нативных» мембранах постоянный обмен веществом и энергией «дробит» рафтовую фазу до субдифракционных размеров…. Читайте дальше на Биомолекуле: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni Источник изображения: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni
Рис. Domain-length scales and the biomembrane as a protein–lipid composite material. (a) Length scales of domains in biomembranes. Shells, complexes and nanoclusters range from 1–10 nm, whereas nanodomains such as caveolae can be as large as 100 nm. (b) A schematic representation of the biomembrane as a composite of lipids and proteins. Estimates of lateral protein concentration are about 30,000 per μm2 based on rhodopsin in the rod outer segment28,29 and transmembrane proteins in the baby hamster kidney (BHK) cell membrane27. Lipids were assumed to occupy a surface area of ∼0.68 nm2 (diameter ∼0.93 nm) and an α-helix ∼1 nm2 (diameter ∼1.1 nm). A 30 × 30 nm2 section of membrane is depicted with 32 lipids on a side, 35 transmembrane proteins with 15 single-span, 12 tetraspan and eight heptaspan α-helical proteins, having assumed crosssectional areas in the plane of the membrane of 1 nm2, 4.5 nm2 and 8 nm2, respectively. Taking into account the area excluded by the proteins, the numerical lipid : protein ratio is ∼50. For a single-span helix with a diameter of ∼1.1 nm, there are about seven lipids in the first boundary layer; for a tetraspan protein with a diameter of ∼2.4 nm, there are about 11 lipids in the first boundary layer; for a heptaspan protein (such as rhodopsin) with a diameter of ∼3.2 nm, there would be about 14 lipids in the first boundary layer. Such first-boundary layer lipids are shown in white, whereas the second layer is shown in red. All other lipids are shown in yellow. Lipid-binding proteins and adaptors linking transmembrane proteins to membrane proximate cytoskeletal filaments are also depicted as different coloured structures beneath the plane of the membrane, but ectodomains of the membrane proteins are omitted for clarity. Источник изображения: https://www.nature.com/articles/ncb0107-7
Видеолекция: Плазматическая мембрана. Е.В. Шеваль, к.б.н.
Видеолекция: Мембрана как клеточная граница. А. Иляскин
Важность ионных каналов мембраны
Легко понять, что через мембранную жировую плёнку могут проникать в клетку только жирорастворимые вещества. Это жиры, спирты, газы. Например, в эритроцитх прямо через мембрану легко проходят внутрь и наружу кислород и углекислый газ. А вот вода и водорастворимые вещества (например, ионы) просто так через мембрану не могут пройти внутрь любой клетки. Это значит, что для них нужны специальные отверстия. Но если просто сделать отверстие в жировой плёнке, то оно тут же затянется обратно. Что же делать? Выход в природе был найден: надо сделать специальные белковые транспортные структуры и протянуть их сквозь мембрану. Именно так и получаются каналы для пропускания не растворимых в жире веществ — ионные каналы мембраны клетки.
Итак, для придания своей мембране дополнительных свойства проницаемости для полярных молекул (ионов и воды) клетка синтезирует в цитоплазме специальные белки, которые затем встраиваются в мембрану. Они бывают двух типов: белки-транспортёры (например, транспортные АТФазы) и белки-каналоформеры (образователи каналов). Эти белки встраиваются в двойной жировой слой мембраны и формируют транспортные структуры в виде транспортёров или в виде ионных каналов. Через эти транспортные структуры теперь могут проходить различные водорастворимые вещества, которые по-другому проходить сквозь жировую мембранную плёнку не могут.
Вообще, встроенные в мембрану белки ещё называются интегральными, именно потому что они как бы включаются в состав мембраны и пронизывают её насквозь. Другие белки, не интегральные, образуют как бы острова, «плавающие» по поверхности мембраны: либо по её наружной поверхности, либо по внутренней. Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко!
Выводы
1. В целом, мембрана получается трёхслойной:
1) наружный слой из белковых «островов»,
2) жировое двухслойное «море» (липидный бислой), т.е. двойная липидная плёнка,
3) внутренний слой из белковых «островов».
Но есть ещё рыхлый наружный слой — гликокаликс, который образуют торчащие из мембраны гликопротеины. Они являются молекулярными рецепторами, с которыми связываются сигнальные управляющие вещества.
2. В мембрану встроены специальные белковые структуры, обеспечивающие её протицаемость для ионов или других веществ. Не надо забывать, что в некоторых местах жировое море пронизано интегральными белками насквозь. И именно интегральные белки образуют специальные транспортные структуры клеточной мембраны (смотрите раздел ). Через них вещества попадают внутрь клетки, а также выводятся из клетки наружу.
3. С любой стороны мембраны (наружной и внутренней), а также внутри мембраны могут располагаться белки-ферменты, которые влияют и на состояние самой мембраны и на жизнь всей клетки.
Так что мембрана клетки — это активная изменчивая структура, которая активно работает в интересах всей клетки и связывает её с окружающим миром, а не просто является «защитной оболочкой». Это — самое важное, что надо знать про клеточную мембрану.
В медицине мембранные белки зачастую используются как “мишени” для лекарственных средств. В качестве таких мишеней выступают рецепторы, ионные каналы, ферменты, транспортные системы. В последнее время кроме мембраны мишенью для лекарственных веществ становятся также гены, спрятанные в клеточном ядре.
Видео: Введение в биофизику клеточной мембраны: Структура мембран 1 (Владимиров Ю.А.)
Видео: История, строение и функции клеточной мембраны: Структура мембран 2 (Владимиров Ю.А.)
Дополнительно: Антонов В.Ф., 1996.
Подробности о биомембранах на сайте Биомолекула
Читать далее:
© 2010-2021 Сазонов В.Ф. © 2010-2016 kineziolog.bodhy.ru, © 2016-2021 kineziolog.su
Что такое клеточная мембрана? / Справочник :: Бингоскул
Каждая клеточка многоклеточного организма отделена от окружающей внутренней (или наружной) среды плазматической мембраной (цитолеммой, плазмалеммой) Это граница, которая «охраняет» внутреннее содержимое клетки от чужеродного вторжения. Но, как и на любой границе, она пропускает «своих» и выводит « чужих». Мембрана – это не панцирь, а подвижная, активная структура, благодаря которой осуществляется обмен веществ между клетками и окружающей средой.
Рис. 1. Клеточная мембранаВ переводе с латыни слово «мембрана» означает «пленка». Клетка завернута или упакована со всех сторон оболочкой, строение которой уникально. Она подвижна: тонкая, гибкая и эластичная диаметром 7,5-10 нм.
Клеточная мембрана похожа на гамбургер, состоящий из трех слоев: липиды – белки – липиды. Но это условное сравнение. На фото видно, что устроена она более сложно: два тонких липидных слоя пронизаны белковыми глобулами, которые встроены в оболочку. Вывод: мембрана подвижна и состоит из белков и липидов (жиров). Это динамичная структура, где липиды и белки способны перемещаться, меняя свое положение.
Строение
С 1925 года, когда впервые рассмотрели мембрану, гипотеза о ее строении претерпела ряд существенных изменений. С 1972 года общепризнанной считается жидкостно-мозаичная модель плазматической мембраны, которая представлена на схеме:
Рис. 2. Жидкостно-мозаичная модель мембраныФосфолипидный биослой устроен таким образом, что гидрофобные остатки жирных кислот обращены внутрь, а гидрофильные головки «смотрят» наружу. Двойной липидный слой – это барьер, благодаря которому содержимое клетки не растекается, и во внутреннее содержимое не попадают токсические вещества.
Рис. 3. Строение клеточной мембраныВ 2-хслойный липидный слой погружены подвижные молекулы белков, которые по выполняемым функциям делятся на 2 большие группы:
- Структурные, которые в прямом смысле поддерживают структуру клетки.
- Динамические, находящиеся на поверхности мембраны. Они участвуют в «барьерных» процессах. Среди динамических различают рецепторные, транспортные и ферментативные белковые глобулы.
На заметку: Соотношение компонентов в плазматической мембране следующее: белки составляют 65%, на фосфолипиды приходится 25%, холестерол – 13%, липиды другого строения – 4%, углеводы – 3%. Липиды (жиры) отталкивают воду, поэтому ее излишки не попадают в клетку.
Функции клеточной мембраны
Плазматическая мембрана ведет себя активно, как любой клеточный органоид. Это не просто барьер: «клеточная капсула» выполняет много задач.
Транспортная функция
Рис. 4. Механизмы прохождения веществ через клеточную мембрануОсуществляет выборочный транспорт веществ, поскольку липидные слои для большинства соединений непроницаемы. Через мембрану идет пассивный и активный транспорт:
1. Пассивный транспорт — это простая диффузия, которая идет с низкой скоростью. Через барьер проникают газы, вода, ряд органических веществ. Облегченная диффузия происходит с участием белков-переносчиков и идет с большей скоростью. На пассивный транспорт не требуется энергетических затрат. Белки-переносчики относятся к транспортным. Они «проносят» вещества через мембрану или через специальные каналы, предназначенные для проникновения различных ионов.
2. Перенос макромолекул или крупных частиц осуществляется путем эндоцитоза. При эндоцитозе на мембране возникают впячивания для захвата твердой частицы пищи. Вокруг нее образуется вакуоль, которая обволакивает частицу и переносит внутрь клетки.
Рис. 5. Эндоцитоз
На заметку: Эндоцитоз делится на фагоцитоз или захват твердой частицы и пиноцитоз, когда поглощается жидкий материал (коллоидный раствор, суспензии или просто раствор).
3. Вывод веществ из клетки через мембрану осуществляется путем экзоцитоза. Вещества, которые нужно переправить в межклеточную жидкость, «упаковываются» в мембранные пузырьки. Они подходят к цитолемме, встраиваются в нее и содержимое пузырьков выбрасывается из клетки. Это различные продукты метаболизма.
Рис. 6. Экзоцитоз4. Активный транспорт осуществляется против градиента концентрации и требует затрат энергии (АТФ). Вещества из области с меньшей концентрацией переходят в область с более концентрированным содержимым. Примером служит натрий-калиевый насос, когда из клетки выводятся ионы натрия и закачиваются ионы калия.
Проницаемость мембран разных клеток неодинакова. Чтобы попасть в клетку, вещество должно обладать определенным размером, химическими свойствами и электрическим зарядом. «Неподходящие» по своим параметрам молекулы просто не способны попасть во внутреннее содержимое клетки.
Другие функции клеточной мембраны
Плазматическая мембрана осуществляет:
- Структурную функцию, отделяя клетку от внешней среды. Так клетка работает как автономный организм. Она содержит набор органоидов, которые «плавают» во внутренней среде – цитоплазме.
- Рецепторную, реагируя на внешние раздражители. Подобную функцию осуществляют белки (гормоны, нейромедиаторы), которые получают сигналы-воздействия извне. Это служит отправной точкой для изменения хода обменных процессов внутри клетки.
- Ферментативную или метаболическую, когда мембранные белки-ферменты участвуют в разных химических процессах метаболизма. Большинство ферментов связаны с мембраной и в липидной оболочке созданы определенные условия для их работы, поэтому плазмалемма непосредственно и косвенно влияет на процессы метаболизма.
- Энергообразующую, которую осуществляют митохондрии – органоиды, которые относят к «маленьким силовым станциям клетки». Многие процессы, связанные с обменом между клеткой и межклеточным пространством нуждаются в дополнительных затратах энергии. Клетки также способны обмениваться энергией. Этот процесс осуществляется через белковые каналы.
- Матричную, когда цитолемма определяет местонахождение, фиксирует расположение органоидов внутри клетки и их позицию относительно друг друга. Оптимальное размещение позволяет органоидам легче взаимодействовать друг с другом.
- Маркировочную, так как каждая клетка снабжена «этикеткой» или маркировкой. Это антигены, состоящие из гликопротеинов (белков с разветвленными олигосахаридными цепями). Разветвления имеют разные конфигурации, поэтому каждый тип клеток снабжен уникальным «ярлыком», что позволяет идентифицировать или узнавать их. Например, иммунные клетки человека (макрофаги) распознают инородную клетку, проникшую в организм (бактерию или вирус). Они начинают работу по ее уничтожению. Таким же путем организм избавляется от старых, больных, мутировавших клеток.
Плазмалемма или мембрана – важнейшая составная часть живой клетки. Она выполняет не меньше функций, чем любой другой органоид. Имеет сложное строение и таит в себе много загадок, которые еще не раскрыты учеными. Этот «живой пропускной шлюз» продолжают изучать, чтобы использовать на благо человека.
Источники изображений:
- Рис. 1. — www.youtube.com/watch?v=uaC8KhzuH98&ab_channel=BogdanTuziak
- Рис. 3. — uchitel.pro/строение-и-функции-клетки/
- Рис. 4. — 900igr.net
|
Клеточная мембрана
2. История исследования
В 1925 году Гортер и Грендель с помощью осмотического «удара» получили так называемые «тени» эритроцитов — их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем с помощью ацетона выделили из оболочек липиды и определили количество липидов на единицу площади эритроцита — этого количества хватило на сплошной двойной слой. Хотя этот эксперимент привёл исследователей к правильному выводу, ими было допущено несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными плёнками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение — белки. В 1935 году Даниэлли и Доусон представили научному сообществу модель «сендвича», которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трёхслойное строение всех клеточных мембран.
Но постепенно накапливались аргументы против «бутербродной модели»:
- «бутербродная» модель термодинамически не выгодна — для поддержания такой структуры нужно затрачивать большое количество энергии, и протащить вещество через мембрану очень сложно;
- плазматическая мембрана может различаться по структуре даже в одной клетке, например в головке, шейке и хвосте сперматозоида;
- количество белков, связанных с мембраной электростатически, очень небольшое, в основном белки очень тяжело выделить из мембраны, так как они погружены в неё.
- оказалось, что структура мембраны при электронной микроскопии зависит от способа её фиксации;
- накапливались сведения о глобулярности плазматической мембраны;
Всё это привело к созданию в 1972 году С. Д. Сингером S. Jonathan Singer и Г. Л. Николсоном Garth L. Nicolson жидкостно-мозаичной модели строения мембраны. Согласно этой модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные, полуинтегральные и периферические. Периферические белки действительно находятся на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими взаимодействиями, но никогда не образуют сплошной слой. Доказательствами жидкостности мембраны служат методы FRAP, FLIP и соматическая гибридизация клеток, мозаичности — метод замораживания-скалывания, при котором на сколе мембраны видны бугорки и ямки, так как белки не расщепляются, а целиком отходят в один из слоёв мембраны.
Клеточная мембрана
Что такое клеточная мембрана
Поддержание жизнедеятельности клетки и контроль за ее целостностью осуществляет защитная пленка. Изучение мембран, их функционирования необходим для понимания причин возникновения заболеваний и способах лечения. Глубокое изучение клеточных мембран позволит создавать лекарства, снизить смертность и отыскать механизмы борьбы с болезнями внутри организма человека.
Каждая клетка в организме находится в специальной защитной пленке, которая и называется клеточной мембраной. Она выполняет много функций, благодаря которым поддерживается процесс жизнедеятельности клетки.
Название пленка и мембрана это одно и тоже, заимствованное из мертвого языка – латыни. Фактически это сложная структура, состоящая из двух пленок, которые соединены и имеют ряд свойств. Самое главное – это защита содержимого внутри оболочки и обеспечение возможности доставления внутрь клетки питательных веществ. Мембрана отвечает за полноценное обеспечение связи клеток друг с другом, и с окружающей средой. Бислой сформирован из липидных молекул, большая часть из которых это фосфолипиды или сложные липиды. Липидная молекула состоит из гидрофильной головки и гидрофобного хвоста. Структура мембраны похожа у многих организмов.
Мембрана включает и разные белки:
- интегральные (мембрана пронизана насквозь),
- полуинтегральные (в липидный слой опущен только один из концов),
- поверхностные (расположены вне клетки или внутри, но прилегают к мембране).
Функции клеточной мембраны
- Барьерная или защитная. Мембрана защищает содержимое клетки, создавая своеобразный барьер. Не позволяет проникать вредным веществам через стенки. Контролирует постоянство структуры клетки и оберегает от вредоносных молекул. При этом, в зависимости от ситуации, мембрана может вести себя активно или пассивно. Может проявлять активность в выборе или отторжении.
- Транспортная. Обеспечивает доставку полезных веществ внутрь клетки, происходит межклеточный обмен полезными веществами и поступает информация извне.
- Матричная. Мембрана строго разграничивает клетки,
- Механическая. Регулирует разграничение клеток между собой, поддерживает правильность их соединения. Здесь основная нагрузка ложится на стенки клетки. У животных активно принимает участие межклеточное вещество.
- Энергетическая. Через белок, содержащийся в клеточной мембране происходит процесс энергообмена.
- Рецепторная. Основную роль выполняют белки, которые выполняют роль рецепторов в клеточной мембране. Они отвечают за доставку сигналов в клетку от гормонов и нейромедиаторов. Это позволяет поддерживать стабильный гормональный фон и способствует беспрепятственному прохождению нервных импульсов.
- Ферментативная. Часть белков принимают участие в данной функции. Так, например, происходит синтез в эпителии кишечника.
- Маркировочная. Антиген. Присутствующий на мембране, действует как маркер-выделитель. Благодаря ему происходит распознавание клетки. Роль таких выделителей исполняют гликопротеины, играющие роль своеобразных антенн. У каждой клеточки свое обозначение, по которым происходит объединение в структуры или отторжение как чужеродных и вредных.
Клеточный обмен может происходить 3 способами
- Фагоцитоз. Обмен внутри клеток, главные участники которого – фагоциты. Они захватывают полезные вещества и перерабатывают их.
- Пиноцитоз. Здесь активной является сама мембранная клетка, которая специальными ловит капельку жидкости. Формируется небольшой пузырек, который постепенно втягивается в мембрану.
- Экзоцитоз. Прямо противоположный процесс, при котором из клетки уходит жидкость через стенки мембраны.
Свойства клеточной мембраны
Клеточные мембраны это не стабильные субстанции, а динамичные текучие образования. Молекулы липидов и белков не связаны между собой ковалентными связями, поэтому они способны быстро передвигаться внутри мембраны. Динамичность мембран проявляется в их способности легко расширяться, сужаться, восстанавливаться после повреждений.
Мембраны у разных биологических видов разные. Прежде всего отличаются по химическому составу. Также отличаются по количеству белков, липидов, по характеру имеющихся в них рецепторов. Каждый тип индивидуален, что определено гликопротеинами, участвующими в распознании факторов внешней среды и узнавании родственных клеток.
В мембранах находятся рецепторы, переносчики электронов, преобразователи энергии, ферментные белки.
Одно из главных свойств мембраны – выборочная или направленная проницаемость. Благодаря этой способности молекулы и ионы проникают через пленку стенок с разной скоростью. Чем крупнее молекула, тем медленнее ее скорость проникновения. Самая большая проникающая способность у воды и растворенных в ней газах. Ионы проходят через мембрану с меньшей скоростью.
Строение клеточной мембраны
Клеточная мембрана состоит из липидов
- Фосфолипиды (комбинация жиров и фосфора)
- Гликолипиды (комбинация жиров и углеводов)
- Холестерол
Фосфолипиды и гликолипиды состоят из гидрофильной головки и двух длинных гидрофобных хвостиков.
Холестерол же занимает пространство между этими хвостиками, не давая им изгибаться, все это в некоторых случаях делает мембрану определенных клеток весьма жесткой. Помимо всего этого молекулы холестерола упорядочивают структуру клеточной мембраны.
Внутри клеточной мембраны находятся разнообразные белки. Их окружают аннулярные липиды (структурированные жиры, оберегают белок и помогают ему функционировать).
Состав клеточной мембраны
В мембране имеется три слоя, главным из которых является однородный жидкий билипидный слой. Окружают его белки, благодаря которым обеспечивается проницаемость клеточных мембран. Фосфолипиды – фундамент мембраны, составляет до 90% от общего количества липидов.
Для проникновения сквозь мембрану таких веществ как калий и натрий, существуют специальные ионные каналы клеточных мембран.
Белки клеточной мембраны
Для клетки жизненно важно взаимодействовать как с соседними клетками, так и с окружающим миром. Некоторые микроскопические молекулы или потоки света беспрепятственно проникают сквозь мембрану, взаимодействуя с белками напрямую. При этом в клетке запускаются химические реакции выработки новых белков или появляется новая программа жизнедеятельности клетки. Пример ответных реакций это: деление клетки, выделять ферменты или гормоны. Клетка может запустить механизм самоуничтожения. Принцип у всех один – внутриклеточный запуск каскада превращений химических реакций.
Чтобы клетка могла функционировать продолжительное время, в нее должны поступать питательные вещества извне. Сигналы, достигающие внутриклеточного пространства, должны правильно обрабатываться и выдавать ответную реакцию. Для этого на поверхности мембраны есть специальные рецепторы: ионные каналы, порины, транспортеры, молекулярные моторы, структурные белки.
Появление гормонов или сигнальных молекул снаружи клетки вызывает в рецепторных белках сигнал. Самый яркий представитель – рецептор инсулина, который отвечает за снабжение клетки глюкозой. Транспорт ионов происходит через ионные каналы, которые поддерживают разницу в их концентрации между наружной средой и внутренней. Натриевые и калевые каналы отвечают за передачу нервного импульса. Порины и транспортеры отвечают за перенос воды и определенных молекул сквозь мембрану. Структурные белки поддерживают структуру мембраны и взаимодействуют с остальными белками.
Кроме всего прочего есть еще внутриклеточные пути передачи сигналов при помощи каскадов реакций.
Цитоплазма и клеточная мембрана
Цитоплазма – это часть клетки, которая находится между плазматической мембраной и ядром. Выделяют составляющие:
- гиалоплазму (основа цитоплазмы),
- органоиды (постоянные составляющие)
- включения (временные составляющие).
Химический состав цитоплазмы
До 90% занимает вода, остальное – всевозможные соединения органики и неорганических веществ. Цитоплазма имеет щелочную реакцию. Отличительная особенность цитоплазмы – циклоз или постоянное движение. Заметить это можно по перемещению внутри клетки хлоропластов. Жизнедеятельность клетки напрямую зависит от движения цитоплазмы. Прекращение движения ведет к гибели клетки, прекращению ее жизнедеятельности.
Гиалоплазма или цитозоль – коллоидный раствор, который не имеет цвета. По составу напоминает густую слизь. В этой жидкости протекают процессы, которые обеспечивают обменные процессы веществ. Благодаря цитозоли осуществляется связь между ядром и всеми органоидами
.
В свою очередь подразделяется на две формы, которые способны менять свое физическое состояние.
- золь – разжиженная,
- гель – тягучая.
Цитоплазма объединяет все внутренние составляющие клетки в единое целое. Ее среда – это место где протекают физиологические и биохимические клеточные процессы. Цитоплазма отвечает за жизнедеятельность и функционирование органоидов.
Проницаемость клеточных мембран
Проницаемость – это важнейшая функция защитного слоя клетки. Благодаря ей происходит движение внутрь и извне клетки многих метаболитов. Постоянно поддерживается форма клетки, баланс в ней веществ, осуществляется проведение нервного импульса, поддерживается жизнеспособность клетки.
Низкомолекулярные жирорастворимые вещества, такие как глицерин, спирты, мочевина могут беспрепятственно самостоятельно проникать через мембранную оболочку. Это лишь малая часть переносимых веществ, называется простая диффузия. Сложное перемещение называется транслокация и невозможно без дополнительных транспортных систем.
Есть предположение, что системы-переносчики состоят из белков или липопротеидов, а также ряд других компонентов. Переносчик или система сначала связывает переносимое вещество, а потом доставляет его через мембрану внутрь клетки. Выделяют также неподвижных переносчиков, которые не перемещаются внутри мембранной оболочки, а являются своеобразным туннелем или каналом. Выделяют также и вторичную транслокацию – переносчик осуществляет связь с переносимым веществом путем невалентных взаимодействий. Выделяют 3 вида:
- Облегченная диффузия (унипорт) – механизм переноса не зависит от переноса веществ в клетку или из нее. Этим способом переносится глюкоза в эритроциты.
- Котранспоорт (симпорт) – совместный транспорт двух или более веществ в одном направлении.
- Противотранспорт – доставка веществ в одном направлении соотносится с движением других частиц в противоположном направлении. Для этого вида транспорта требуется много энергии, которая образуется за счет сопряжения вторичной транслокации с ферментативными реакциями разрыва или образования химических связей.
Клеточная мембрана состоит из белков и липидов, основу которых составляют фосфолипиды. Фосфолипиды занимают значительную часть – 40-90% всех липидов в мембранной оболочке.
Липиды – это амфипатические молекулы, самостоятельно формирующие бислои.
Липиды имеют особенность: растворяются только в растворителях на органической основе и совсем не подвержены растворению в воде. Клеточная мембрана имеет несколько видов липидов: фосфолипиды, холестерол, гликолипиды.
Строение клеточной мембраны до конца не изучено. Происходит постоянное изучение и составление моделей состава мембраны. В одной – мембрана характеризуется как липидный двойной слой. В этом слое углеводородные хвосты липидов за счет гидрофобных взаимодействий удерживаются друг возле друга в вытянутом состоянии во внутренней полости, образуя двойной углеводородный слой. Полярные группы липидов находятся на внешней поверхности бислоя.
Изучение клеточных мембран перспективное направление в науке. Возможно, с полным пониманием механизмов, происходящих внутри клетки, позволит продлить жизнь. Может быть удастся найти ключик к долголетию.
Появление и эволюция клеточной мембраны
У всех современных организмов клеточная мембрана играет принципиальную роль в энергетическом обмене и других биохимических процессах. Новые исследования эволюции мембран позволяют ответить на многие каверзные вопросы: как мембрана появилась у нашего далекого предка LUCA, почему мембраны бактерий и архей так непохожи и каким образом эукариоты обзавелись мембранными органеллами.
Мембрана играет важнейшую роль в нормальном функционировании клетки: она обеспечивает отделение клетки от внешней среды и за счет компартментализации создает необходимую среду для протекания различных биохимических и энергетических процессов. Немало исследований посвящено изучению биохимии и биофизики биомембран [1], но не менее важное значение имеет и изучение их эволюции. Как и при каких обстоятельствах мембрана появилась в эволюции живого впервые? Когда появились первые эукариоты, и каким образом они обзавелись множеством внутренних мембран, которых нет у прокариот? Над этими вопросами ученые ломают головы уже долго, но до недавнего времени они могли оперировать только умозрительными гипотезами. Развитие масштабных методов анализа геномов (и прочих «омов» [2]), биоинформатики и математического моделирования в биологии [3] позволили если и не дать исчерпывающие ответов, то подобраться к ним вплотную.
Происхождение эукариот «наизнанку»
В недавно опубликованной в журнале BMC Biology статье [4] Дэвид и Базз Баумы, основываясь на большом количестве филогенетических данных, выдвинули новую гипотезу происхождения эукариотической клетки. Они называют эту гипотезу «моделью наизнанку» (inside-out, изнутри — наружу), в противовес господствовавшей до сих пор гипотезе «снаружи — внутрь» (outside-in). Согласно традиционной теории мембранные органеллы эукариот появились благодаря «впячиванию» своей наружной мембраны. Митохондрии, например, согласно этой гипотезе, были «проглочены» будущими эукариотами с помощью фагоцитоза. Однако со времени появления этой гипотезы накопилось немало данных, которые ей противоречат и указывают на то, что ситуация была противоположной. Вероятно, новые органеллы появились у будущих эукариот более дружелюбным способом — с помощью объятий. «Модель наизнанку» предполагает, что эукариотическое ядро образовалось из основной части предковой клетки, а цитоплазма с митохондриями и другими мембранными органеллами — из выростов этой клетки, которые по началу просто окружали клетки-симбионты (рис. 1). Новую гипотезу поддерживает множество важных фактов. Например, археи (они и были этими предковыми клетками) могут только выпячивать мембрану, а «впячивать» — нет. Несомненно, эта новая гипотеза требует дальнейшей проработки, но специалисты* оценивают ее позитивно: она действительно подтверждается известными данными о морфологии и биохимии прокариот и помогает сделать предсказания, которые можно проверить экспериментально (например, механизмы сборки ядерных пор и филогению белков фагоцитоза).
Рисунок 1. Схема того, как эукариотическая клетка могла возникнуть в соответствии с «моделью наизнанку». Выросты клетки-хозяина окружили клетки-симбионты, постепенно превратив их во внутренние мембранные органеллы. Рисунок из [4].
Математическое моделирование позволило другой группе ученых лучше разобраться с еще одним важным вопросом: какой была мембрана общего предка архей и бактерий, и как ее строение определило эволюцию этих двух групп прокариот. Об этом рассказывается в их недавней статье, вышедшей в журнале PLoS Biology [6].
Бактерии и археи: единство противоположностей
Все современные живые организмы относятся к одному из трех доменов жизни: бактерии, археи и эукариоты. По более-менее общепринятой гипотезе эукариоты происходят от своеобразного «слияния» двух других групп, которые являются гораздо более древними. Бактерии и археи происходят от общего предка — по-английски он называется LUCA (last universal common ancestor, последний универсальный общий предок). Бактерии и археи имеют много общих черт, включая одинаковый генетический код, механизмы транскрипции и рибосомной трансляции, но при этом отличаются в некоторых ключевых моментах. Они имеют разный химический состав клеточных мембран и стенок, по-разному устроенный гликолиз, ионные насосы и даже разные механизмы репликации ДНК.
Возможно, различия в устройстве клеточной мембраны являются ключевыми в этом списке различий (рис. 2) [7]. Мембраны современных бактерий состоят из фосфолипидов: сложных эфиров глицерина, двух остатков жирной кислоты и одного фосфатного остатка, к которому может быть присоединена дополнительная полярная группа. Гидрофобные хвосты жирных кислот образуют средний слой мембраны, а полярные остатки глицерина, фосфата и вспомогательных полярных групп — наружный и внутренний слои. Мембраны архей устроены в принципе похоже, но на другой химической основе. Вместо жирных кислот их липиды содержат терпеновые спирты, углеводородные цепочки которых несут метильные группы через каждые четыре атома. Моделирование молекулярной динамики мембран показало, что благодаря таким метильным «ответвлениям» мембраны становятся очень прочными, но при этом сохраняют гибкость [8, 9]. Терпеновые спирты простыми эфирными связями присоединяются к глицеринфосфату, фосфатный остаток может дополняться другими полярными головками, такими же, как у бактерий. Сам глицеринфосфат архей тоже отличается от бактериального — у архей используется другой его оптический изомер (глицерин-1-фосфат вместо глицерин-3-фосфата). Получается, что мембрана — важнейший элемент, обеспечивающей существование клетки как самостоятельной единицы, — появилась у бактерий и архей независимо. Из этого удивительного наблюдения некоторые ученые даже делают вывод о том, что у LUCA мембраны вообще не было [10]. Но это крайне маловероятно, учитывая, насколько важной для большинства биохимических процессов является мембрана. Сложно представить, что молекулярные механизмы, протекающие одинаково и у бактерий, и у архей, появились и могли функционировать еще до появления мембраны. Значит, какая-то мембрана у LUCA все-таки была. Группа ученых из Университетского Лондонского колледжа с помощью математического моделирования разработала модель, описывающую, как эта мембрана выглядела, и как из нее появились разные мембраны бактерий и архей [6].
Рисунок 2. Строение мембранных липидов бактерий (справа) и архей (слева) [7].
«Протекающая» мембрана
C различным строением мембраны бактерий и архей никак не вязалось то, что производство энергии в клетках обеих групп устроено очень похожим образом. Дело в том, что во всех современных клетках производство энергии (которая запасается в виде молекул АТФ) сопряжено с мембраной. Ключевыми стадиями этого процесса являются создание градиента протонов на мембране (избыток ионов Н+ с наружной стороны мембраны по сравнению с внутренней) и работа АТФ-синтазы за счет этого градиента. При этом протоны проходят через канал в АТФ-синтазе, вызывая тем самым механический поворот части АТФ-ситназного комплекса, который, в свою очередь, обеспечивает катализ синтеза АТФ. Согласно филогенетическим исследованиям, АТФ-синтазы всех организмов имеют общее эволюционное происхождение, и предковая молекула была уже у LUCA. У некоторых бактерий и архей вместо градиента протонов используется градиент ионов натрия, а у некоторых — и тот, и другой. Долгое время считалось, что Na+ выступает в качестве заменителя H+ у организмов, живущих в экстремальных условиях (термальных источниках или в сильнощелочной среде). Однако оказалось, что натрий-специфические ферменты занимают самые нижние ветви филогенетического древа в обоих доменах, что указывает на их древность. Модель функционирования древней мембраны, предложенная британскими учеными, успешно объясняет, как и зачем в процессе эволюции возникла способность АТФ-синтазы использовать ионы натрия. Но, прежде чем ответить на этот вопрос, они должны были разобраться с еще одной проблемой — несмотря на общее происхождение АТФ-синтаз, ионные насосы возникли у бактерий и архей независимо, т.е, вероятно, у LUCA их не было. Как же тогда древняя клетка могла избавляться от протонов, поступающих внутрь при работе АТФ-синтазы, и создавать градиент протонов?
По мнению авторов исследования, единственным объяснением могло быть то, что мембрана LUCA была «протекающей» (leaky), и клетка использовала естественные источники протонного градиента. На основе своих предположений ученые построили математическую модель древней клетки. В этой модели клетка находится на границе между двумя ламинарными потоками — кислотным (pH 5–7) и щелочным (pH 9–10), не смешивающимися за счет неорганического барьера (рис. 3). Подобные условия могли существовать в древнем океане рядом с подводными щелочными источниками (сама морская вода имела кислую реакцию). При этом мембрана клетки была полупроницаемой («протекающей») и свободно пропускала ионы H+с одной стороны клетки и ионы OH− с другой стороны. Эти ионы могут также свободно выходить через мембрану или взаимно нейтрализоваться внутри клетки с образованием воды. Молекула, способная к синтезу АТФ (древняя АТФ-синтаза), находится на «кислотной» стороне клетки и использует градиент протонов на этой мембране для своей работы. Согласно расчетам исследователей, разница pH в три единицы (т.е. тысячекратная разница в концентрации протонов) между щелочной и кислотной средами и молекулы АТФ-синтазы, занимающие 1% поверхности клетки, — это условия, необходимые и достаточные для того, чтобы клетка могла синтезировать необходимое количество АТФ для поддержания углеродного и энергетического метаболизма.
Рисунок 3. Условия с естественным градиентом протонов, в которых должна была обитать древняя клетка [6].
По мнению ученых, такая «протекающая» мембрана могла состоять из смеси амфифильных молекул, включая жирные кислоты и изопрены, но никак не могла содержать фосфолипиды, свойственные современным мембранам. Добавление фосфолипидов приводит к снижению проницаемости мембраны для ионов, так как полярные группы не могу проходить через неполярную внутреннюю часть мембраны. Такая мембрана не позволяла бы поддерживать градиент протонов, а значит, и работу АТФ-синтазы. Получается, что для клеток с «протекающей» мембраной не нужны ни фосфолипиды, ни ионные насосы (они никак не буду способствовать более эффективной работе АТФ-синтазы, т.к. все «накачанные» ионы будут утекать через мембрану). Чтобы понять, как произошел переход от «протекающей» мембраны к современным мембранам с ионными насосами, ученые обратились к уже упомянутому факту: некоторые АТФ-синтазы могут использовать не только протоны, но и ионы натрия.
Исследователи предположили, что необходимым шагом для перехода к современной мембране было появление способности использовать для создания энергии градиента ионов натрия. Создавать такой градиент могла бы молекула SPAP (sodium-proton antiporter, антипорт для ионов натрия и протонов), которая переносит один ион натрия в обмен на один протон. SPAP есть у многих представителей как архей, так и бактерий. Именно эта молекула могла бы использовать естественный градиент протонов для создания градиентов ионов натрия. Даже «протекающая» мембрана в шесть раз менее проницаема для ионов натри, чем для протонов, поэтому градиент ионов натрия гораздо более долговечен в таких условиях. Если АТФ-синтаза сможет использовать для производства АТФ и протоны, и ионы натрия, клетка, согласно подсчетам, сможет создавать на 60% больше энергии. Как уже было отмечено, некоторые современные АТФ-синтазы действительно способны использовать оба вида ионов. Другие используют только один тип ионов, но при этом все они отличаются только парой аминокислотных замен (вероятно, это связано со схожестью ионного радиуса и заряда ионов Na+ и H3O+ — форм, в которые этих ионы обычно транспортируются ионными каналами). Получившийся благодаря SPAP и смешанной работе АТФ-синтаз выигрыш в энергии клетки смогли бы использовать для того, чтобы начать занимать новые экологические ниши, в которых естественный градиент протонов был гораздо ниже (до 50 раз ниже) или был непостоянным. Кроме того, наличие SPAP делает выгодным наличие в клетке ионных насосов. Согласно расчетам модели, преимущество в использовании насосов возрастает со снижением проницаемости мембраны, вплоть до значений проницаемости, характерных для современных мембран.
Получатся, что SPAP — это та молекула, которая могла бы обеспечить переход от «протекающей» мембраны к почти непроницаемой современной, параллельно позволяя древним клеткам расширять ареал своего обитания. По мере расселения, в разных популяциях LUCA могли возникать различные типы насосов, поэтому в современном мире бактерий и архей мы наблюдаем такое разнообразие молекул, причем не все они имеют общее происхождение. Исследователи смогли ответить и на вопрос, связанный с принципиальным различием мембран бактерий и архей. Моделирование показало, что только после появления в эволюции ионных насосов клеткам стало выгодно снижать проницаемость мембраны за счет присоединения гидрофильных глицерол-фосфатных головок. Из-за того, что такой синтез фосфолипидов может происходить двумя путями, в зависимости от того, с какой стороны происходит нуклеофильная атака на карбонильный центр, появилось два разных хиральных варианта фосфолипидов у бактерий и архей. Получается, что разные популяции получили разные ионные насосы, а потом каждая из них пошла либо по «архейному» пути, либо по «бактериальному», в зависимости от реакции нуклеофильного замещения.
Рисунок 4. Эволюция архей и бактерий от общего предка LUCA. A—E — постепенный переход от «протекающей мембраны» к современной. F — дивергенция двух популяций, давших начало археям и бактериям, за счет эволюции мембраны. На рисунках обозначены АТФ-синтаза (ATPase), архейный и бактериальный ионные насосы (Archaeal pump, Bacterial pump) и SPAP, сыгравшие главные роkи в процессе расхождения архей и бактерий. Рисунки из [6, 11].
Заключение
Изучать появление и эволюцию жизни на самых ранних ее этапах — задача сложная и нетривиальная, требующая работы с большими объемами данных и особенных подходов. В последние годы у ученых в руках появляется все больше инструментов для таких исследований, позволяющих им проверять давно сформулированные гипотезы и выдвигать новые предположения. Иногда результаты удивляют и предполагают отказ от уже устоявшихся и давно вошедших в учебники теорий. Одно из новых исследований, например, показало, что стоит отказаться от теории происхождения мембранных органелл путем фагоцитоза, а обратить внимание на противоположную модель — модель расширения мембраны. Другое описанное в этой статье исследование предлагает еще одну достаточно революционную идею. Согласно математической модели британских ученых мембрана LUCA была «протекающей», а переход к современной мембране стал возможен благодаря антипорту протонов и ионов натрия. Эта модель подразумевает, что мембрана древних клеток состояла из жирных кислот и терпенов, хотя ранее такие мембраны считались неподходящими для производства энергии как раз из-за своей склонности к «протечкам».
Благодаря развитию информационных технологий и растущим объемам биологических баз данных ученые могут, хотя только в компьютерных моделях, заглянуть в далекое прошлое. Являются ли эти модели верными, покажут дальнейшие исследования, но уже сейчас они помогают понять многие критические точки в эволюции жизни на Земле.
- Липидный фундамент жизни;
- «Омики» — эпоха большой биологии;
- Вычислительное будущее биологии;
- Baum D.A., Baum B. (2014). An inside-out origin for the eukaryotic cell. BMC Biology 12, 76;
- ПостНаука: «Выдвинута новая гипотеза происхождения эукариотической клетки»;
- Sojo V., Pomiankowski A., Lane N. (2014) A Bioenergetic Basis for Membrane Divergence in Archaea and Bacteria. PLoS Biol 12, e1001926;
- Никитин М.А. (2013). Происхождение мембран и мембранной биоэнергетики. Химия и Жизнь № 9 (2013);
- Пресс-релиз на сайте ИБХ: «Прочные, но гибкие: молекулярная динамика объясняет уникальность биомембран архей»;
- Chugunov A.O., Volynsky P.E., Krylov N.A., Boldyrev I.A., Efremov R.G. (2014). Liquid but Durable: Molecular Dynamics Simulations Explain the Unique Properties of Archaeal-Like Membranes. Sci. Rep. 4, 7462;
- Martin W., Russell M.J. (2007). On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 1887–1925;;
- Robinson R. (2014). A Leaky Membrane and a Sodium Transporter at Life’s Great Divergence. PLoS Biol. 12, e1001927..
Урок по теме «Клеточная мембрана». 10-й класс
Тип урока: урок изучения нового материала (урок по модульной технологии)
Цель: продолжить формирование у учащихся знаний о строении эукариотической клетки.
Задачи
Образовательные
- Выявить сходство принципов построения клеток.
- Выяснить сущность принципа построения клеточной мембраны.
- Сформировать знания о функции клеточной мембраны.
- Сформировать познавательный интерес к процессам, происходящим в клетке.
- Углубить знания о связи строения и функции на примере клеточной мембраны.
- Показать связь молекулярного и субклеточного уровней организации живых существ.
- Выяснить, как осуществляется перенос веществ внутрь клетки и из клетки.
- Выяснить, чем пассивный транспорт веществ отличается от активного.
Развивающие
- Продолжить развитие познавательных способностей учащихся: логического мышления (анализа, синтеза, сравнения, умения выяснять причину и следствие, умения обобщать полученные результаты, конкретизировать, делать выводы).
- Способствовать развитию умственной деятельности (основных качеств ума – самостоятельности, критичности, гибкости, творческой инициативы, избирательности).
- Способствовать развитию активного умения слушать и воспринимать информацию для обеспечения дальнейшей деятельности.
- Продолжить формировать умения работы с текстом, схемами и рисунками учебника, находить сведения для составления схем, пользоваться рекомендациями к выполнению определённого вида деятельности.
- Развивать познавательный интерес к образовательной области “Биология”.
- Продолжить развитие навыков самостоятельной работы, анализа своей деятельности.
Воспитательные
- Продолжить формирование умения и навыков работать в режиме “само”, оценивать результаты своей работы и работы своих товарищей.
- Показать ученику, что каждый из них является перспективным подростком в принципиальной возможности развития его способностей, всей личности.
Планируемые результаты обучения
Предметные
Учащиеся должны: называть основные составные части клеточной мембраны, характеризовать строение и функции клеточной мембраны, устанавливать взаимосвязи между строением клеточной мембраны и её функциями.
Метапредметные
Познавательные. Учащиеся должны уметь: применять логические приемы анализа-синтеза, сравнения, систематизации для осмысления информации; выявлять причины происходящих в клетке процессов; выявлять взаимосвязь строения и функции; самостоятельно работать с учебной литературой; искать необходимую информацию для выполнения заданий; работать с учебно-тематической картой; владеть языком предмета; выявлять особенности строения и функции клеточной мембраны.
Коммуникативные. Учащиеся должны уметь: давать устные ответы, формулировать предположения в поиске ответа на проблемные вопросы.
Регулятивные. Учащиеся должны уметь: распределить рабочее время на выполнение работы с учебно-тематической картой, корректировать правильность выполнения заданий.
Личностная значимость: Я исследователь. Я открываю для себя тайны живой природы. Формирование научного мировоззрения о строении клеточной мембраны, убежденности в познаваемости мира, в важности биологических знаний.
Формируемая область понимания: субклеточный (надмолекулярный) уровень организации живых существ — клеточная мембрана
Контроль: индивидуальный, фронтальный, групповой, самоконтроль, контроль учителя.
Методы: наглядные, беседа, работа с книгой и учебно-тематической картой, волевые, социальные, познавательные, эмоциональные.
Оборудование: учебно-тематические карты для организации самостоятельного поиска, таблица “Клеточная мембрана”, схематическое изображение процессов фагоцитоза и пиноцитоза
Основные понятия урока: плазматическая (цитоплазматическая мембрана), билипидный слой, гидрофильная головка, гидрофобный хвост, мембранные белки, эндоцитоз, экзоцитоз, фагоцитоз, пиноцитоз, активный транспорт веществ, пассивный транспорт веществ, осмос, диффузия через мембранные каналы, простая диффузия, облегчённая диффузия, калий-натриевый насос.
Деятельность учащихся: самостоятельная работа с учебно-тематической картой с последующим обсуждением её результатов.
Ход урока
1. Оргмомент.
Проверить явку учащихся.
Проверить готовность класса к уроку.
2. Самостоятельная работа учащихся с учебно-тематическими картами и учебной литературой, поэтапное обсуждение выполнения заданий учебных элементов.
Рефлексия. Подведение итогов модульного урока, определение индивидуального домашнего задания.
Учебно-тематическая карта “Клеточная мембрана”
| Номер учебного элемента | Учебный материал с указанием учебного материала | Руководство по усвоению учебного материала |
| УЭ- 1 | Цель:
Устно ответьте на вопросы
(единство происхождения живых организмов, каждый организм в ходе эволюции шёл своим путём)? Какие клетки появились в ходе эволюции раньше: растительные или животные? Ответ поясните, исходя из строения этих клеток. | Проверка знаний и умений по теме
“Строение клетки. Типы клеточной организации” Устно ответьте на вопросы. Работайте с рис. 22 стр. 56, рис. 23 стр. 57 |
| УЭ-2 | Интегрирующая цель: в ходе работы с
учебно-тематической картой продолжить
формирование знаний о строении эукариотической
клетки. Задачи: выяснить сущность принципа построения клеточной мембраны; сформировать знания о функции клеточной мембраны; сформировать познавательный интерес к процессам, происходящим в клетке; углубить знания о связи строения и функции на примере клеточной мембраны; показать связь молекулярного и субклеточного уровней организации живых существ; выяснить, как осуществляется перенос веществ внутрь клетки и из клетки; выяснить, чем пассивный транспорт веществ отличается от активного. | Внимательно прочитайте цели и задачи урока. |
| УЭ-3 | Цель:
Устно ответьте на вопросы
Запомни! Молекулы белка и липидов подвижны, что обеспечивает динамичность плазматической мембраны. Запомни! Виды мембранных белков
Запомни! К некоторым белкам, находящимся на наружной поверхности, прикреплены углеводы (гликопротеины = белки + олигосахариды или полисахариды). Белки и углеводы у разных клеток на поверхности мембран неодинаковы и являются своеобразными указателями типа клеток. Например, с помощью этих указателей сперматозоиды узнают яйцеклетку. Благодаря мембранным полисахаридным “антеннам” клетки, принадлежащие к одному типу, объединяются вместе, образуя ткани. Запомни! Клеточная стенка (оболочка) = наружный слой + плазматическая мембрана
Наружный слой поверхности клеток животных тонкий, эластичный и состоит из разнообразных полисахаридов и белков (гликопротеины). Он получил название гликокаликс. 12. Запомни! Строение мембран всех других органоидов сходно с плазматической мембраной. Различаются они составом, соотношением липидов и белков, их расположением в структуре мембран. 13. Запомни! Билипидный слой представлен в основном фосфолипидами. 14. Запомни! На белки приходится в среднем 60% Периферические белки – располагаются на наружной или внутренней поверхности билипидного слоя. Полуинтегральные белки – белки, погружённые в билипидный слой на различную глубину. Интегральные белки или трансмембранные белки – пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой. | Изучите предложен-ную информацию и
устно ответьте на вопросы. Параграф 14 Стр. 55, р. 2 Рис. 24, стр. 58 |
| Выполните задания Чем представлена оболочка растительной клетки: А) плазматической мембраной Б) клеточной стенкой В) плазматической мембраной и наружным слоем из целлюлозы Г) наружным слоем, образованным из целлюлозы 2. Каково строение плазматической мембраны: А) образована бимолекулярным слоем липидов Б) ближе к цитоплазме бимолекулярный слой липидов, снаружи белковые молекулы В) ближе к цитоплазме – бимолекулярный слой липидов, снаружи – углеводные молекулы Г) бимолекулярный слой липидов, белки пронизывают всю ёё толщу и располагаются на её внешней и внутренней поверхности. 3. Какие вещества образуют основу клеточной мембраны: А) гликолипиды Б) фосфолипиды В) жиры Г) белки 4. Плотная оболочка отсутствует в клетках А) бактерий Б) млекопитающих В) земноводных Г) грибов Д) птиц Е) растений 5. Укажите компоненты биологических мембран А) ДНК Б) липиды В) т-РНК Г) углеводы Д) АТФ Е) белки | Максимальное количество баллов – 7 (1-3 задания по 1 баллу, 4-5 задания по 2 балла) | |
| УЭ- 4 | Цель:
Для переноса воды и различных ионов в клеточной мембране имеются поры, через которые в клетку пассивно поступает вода и некоторые ионы (калий, натрий, кальций и др.), которые имеют маленький диаметр. Существует активный перенос веществ с помощью специальных белков, входящих в состав клеточной мембраны. Перенос веществ осуществляется на основе процессов фагоцитоза и пиноцитоза (активный транспорт). Фагоцитоз – захват плазматической мембраной твёрдых частиц и впячивание (втягивание) их внутрь клетки. (Это явление можно наблюдать при захвате амёбой более мелких одноклеточных организмов, при захвате бактерий, проникающих в организм животного или человека, лейкоцитами крови). Так как клетки растений поверх наружной клеточной мембраны покрыты плотным слоем клетчатки, они не могут захватывать вещества при помощи фагоцитоза. Пиноцитоз – захват плазматической мембраной и впячивание капелек жидкости с растворёнными в ней веществами внутрь клетки. Он наиболее универсален, т.к. присущ клеткам растений, животных, грибов. Плазматическая мембрана образует впячивание в виде тонкого канальца, в который и попадает жидкость с растворёнными в ней веществами. Каково общее название фаго- и пиноцитоза? Что такое экзоцитоз? Каков механизм действия этого процесса? Запомни! Функции клеточной мембраны
Транспорт через мембраны жизненно важен для саморегуляции клетки, поддержания постоянства её состава, физико-химического гомеостаза, что необходимо для работы клеточных ферментов; он поставляет питательные вещества, которые служат источником энергии, а также “сырьё” для образования клеточных компонентов; от него зависят выведение токсичных отходов, а также секреция полезных веществ. Мембраны обладают свойством избирательной проницаемости, т. е. хорошо проницаемы для одних соединений и плохо или совсем непроницаемы для других соединений. Плазматическая мембрана хорошо проницаема для веществ, растворимых в липидах, т. к. мембраны состоят из липидов. Плазматическая мембрана более проницаема для незаряженных соединений, чем для ионов. Вода проходит через мембрану в 100000 раз быстрее, чем ионы калия и хлора. Пассивный транспорт веществ идёт без затрат энергии, связан с явлением диффузии, т.е. перемещением веществ по градиенту концентрации из области с более высокой концентрацией в область с более низкой. Например
Например *Натрий-калиевый насос, во время которого из клетки перекачиваются ионы Na+, а вкачиваются ионы К+. На работу натрий-калиевого насоса тратится почти 1/3 всей энергии, необходимой для жизнедеятельности клетки. В мышечных и нервных клетках натрий-калиевый насос обеспечивает возникновение в плазматической мембране разности потенциалов. *К активному транспорту веществ относятся эндо- и экзоцитоз. | Изучите предложен-ную информацию и
устно ответьте на вопросы.
Параграф 14 Стр. 55, р. 2 Рис. 24, стр. 58 |
| Выполните задания Каким образом вода попадает в клетку через клеточную оболочку: А) через гидрофильные каналы белковых молекул и через бимолекулярный слой липидов клеточной мембраны Б) за счёт активного транспорта В) за счёт фагоцитоза Г) за счёт пиноцитоза 2. Что такое фагоцитоз: А) работа калий-натриевого насоса Б) уничтожение микроорганизмов В) захват плазматической мембраной капель жидкости и втягивание их внутрь клетки Г) захват плазматической мембраной твёрдых частиц и втягивание их внутрь клетки 3. Что такое пиноцитоз: А) работа калий-натриевого насоса Б) уничтожение микроорганизмов В) захват плазматической мембраной капель жидкости и втягивание их внутрь клетки Г) захват плазматической мембраной твёрдых частиц и втягивание их внутрь клетки 4. Что такое активный транспорт: А) транспорт веществ через оболочку клетки, который идёт с затратой энергии АТФ Б) транспорт веществ через оболочку клетки, который идёт без затрат энергии АТФ В) Захват плазматической мембраной капель жидкости и втягивание их внутрь клетки Г) захват плазматической мембраной твёрдых частиц и втягивание их внутрь клетки 5. Какой вид транспорта наблюдается при поступлении воды в клетку: А) активный транспорт Б) диффузия В) осмос Г) все вышеперечисленные виды транспорта 6. Что такое осмос: А) движение молекул растворённого вещества через проницаемую мембрану Б) движение молекул растворителя через проницаемую мембрану В) транспорт веществ через оболочку клетки, который идёт с затратой энергии АТФ Г) захват плазматической мембраной капель жидкости и втягивание их внутрь клетки | Максималь-ное количество баллов – 6 (за каждое задание по 1 б) | |
| УЭ- 5 | Подведение итогов работы. Прочитайте цели урока. Достигли ли вы поставленной цели. Оцените свою работу (работа с ключом к заданиям).
Сдайте тетрадь на проверку учителю. Д/З а) выполнили всё на “5” — освобождаетесь от домашнего задания; выполнили на “4” — прочитайте параграфы более внимательно; выполнили на “3” — отработайте еще раз параграф14 по технологической карте “Клеточная мембрана” . Информацию, отсутствующую в учебнике занесите в тетрадь, воспользовавшись учебно-тематической картой “Клеточная мембрана”, и запомните. | Рефлексия
Максималь-ное количество баллов — 13 |
Литература
- Каменский А.А., Криксунов Е.А., Пасечник В.В. Общая биология. 10-11 класс: учеб. Для общеобразоват. учреждений. – М.: Дрофа, 2014. – 368 с.: ил.
- Колесников С.И.Биология. Большой справочник для подготовки к ЕГЭ: учебно-методическое пособие. – Ростов н/Д: Легион, 2015. – 544 с.
- Лемеза Н.А., Камлюк Л.В., Лисов Н.Д. Биология в экзаменационных вопросах и ответах. – М.: Рольф, Айрис-пресс, 1998. 496 с.: ил.
- Мустафин А.Г., Лагкуева Ф.К., Быстренина Н.Г. и др. под редакцией Ярыгина В.Н. Биология. Пособие для поступающих в вузы. – М.: Высшая школа, 2001. – 492 с.: ил.
- Общая биология. 9-11 классы: разноуровневые упражнения и тестовые задания/сост. М.В. Высоцкая. – Волгоград: Учитель, 2008. – 240 с.: ил.
Структура клеточной мембраны
Результаты обучения
- Описать строение клеточных мембран
- Идентифицировать компоненты клеточной мембраны, включая фосфолипиды, холестерин, белки и углеводы
Плазматическая мембрана клетки определяет клетку, очерчивает ее границы и определяет характер ее взаимодействия с окружающей средой. Клетки исключают одни вещества, поглощают другие и выделяют третьи в контролируемых количествах.Плазматическая мембрана должна быть очень гибкой, чтобы определенные клетки, такие как красные и белые кровяные тельца, могли изменять форму при прохождении через узкие капилляры. Это наиболее очевидные функции плазматической мембраны. Кроме того, на поверхности плазматической мембраны есть маркеры, которые позволяют клеткам узнавать друг друга, что жизненно важно для формирования тканей и органов на раннем этапе развития, а затем играет роль в различении иммунного ответа «я» и «чужое».
Одной из наиболее сложных функций плазматической мембраны является способность сложных интегральных белков рецепторов передавать сигналы.Эти белки действуют как приемники внеклеточного ввода, так и активаторы внутриклеточной обработки. Эти мембранные рецепторы обеспечивают внеклеточные сайты прикрепления для эффекторов, таких как гормоны и факторы роста, и они активируют каскады внутриклеточных ответов, когда их эффекторы связаны. Иногда вирусы захватывают рецепторы (ВИЧ, вирус иммунодефицита человека, является одним из примеров), которые используют их для проникновения в клетки, а иногда гены, кодирующие рецепторы, мутируют, вызывая сбой в процессе передачи сигнала с катастрофическими последствиями.
Жидкая мозаика Модель
Ученые идентифицировали плазматическую мембрану в 1890-х годах, а ее химические компоненты — в 1915 году. Основными компонентами, которые они идентифицировали, были липиды и белки. В 1935 году Хью Дэвсон и Джеймс Даниелли предложили структуру плазматической мембраны. Это была первая модель, которую широко приняли другие в научном сообществе. Это было основано на том, что плазматическая мембрана выглядела как «железнодорожный путь» на ранних электронных микрофотографиях. Дэвсон и Даниелли предположили, что структура плазматической мембраны напоминает сэндвич.Они провели аналогию белков с хлебом и липидов с начинкой. В 1950-х годах достижения в области микроскопии, особенно просвечивающей электронной микроскопии (ПЭМ), позволили исследователям увидеть, что ядро плазматической мембраны состоит из двойного, а не одинарного слоя. В 1972 году С.Дж. Сингер и Гарт Л. Николсон предложили новую модель, которая обеспечивает микроскопические наблюдения и лучше объясняет функцию плазматической мембраны.
Объяснение, модель жидкой мозаики , несколько эволюционировало с течением времени, но по-прежнему лучше всего объясняет структуру и функцию плазматической мембраны в том виде, в котором мы их теперь понимаем.Модель жидкой мозаики описывает структуру плазматической мембраны как мозаику компонентов, включая фосфолипиды, холестерин, белки и углеводы, что придает мембране жидкий характер. Плазменные мембраны имеют толщину от 5 до 10 нм. Для сравнения, красные кровяные тельца человека, видимые с помощью световой микроскопии, имеют ширину примерно 8 мкм, или примерно в 1000 раз шире плазматической мембраны. Мембрана действительно немного похожа на бутерброд (рис. 1).
Рис. 1. Жидкая мозаичная модель структуры плазматической мембраны описывает плазматическую мембрану как жидкую комбинацию фосфолипидов, холестерина, белков и углеводов.
Основными компонентами плазматической мембраны являются липиды (фосфолипиды и холестерин), белки и углеводы, присоединенные к некоторым липидам и белкам. Фосфолипид — это молекула, состоящая из глицерина, двух жирных кислот и головной группы, связанной с фосфатом. Холестерин, еще один липид, состоящий из четырех конденсированных углеродных колец, расположен рядом с фосфолипидами в ядре мембраны. Пропорции белков, липидов и углеводов в плазматической мембране зависят от типа клетки, но для типичной клетки человека белок составляет около 50 процентов композиции по массе, липиды (всех типов) составляют около 40 процентов, а углеводы. составляют оставшиеся 10 процентов.Однако концентрация белков и липидов зависит от клеточных мембран. Например, миелин, продукт мембраны специализированных клеток, изолирующий аксоны периферических нервов, содержит только 18 процентов белка и 76 процентов липидов. Внутренняя мембрана митохондрий содержит 76 процентов белка и только 24 процента липидов. Плазматическая мембрана эритроцитов человека на 30 процентов состоит из липидов. Углеводы присутствуют только на внешней поверхности плазматической мембраны и прикрепляются к белкам, образуя гликопротеинов , или присоединяются к липидам, образуя гликолипидов .
Как вирусы заражают определенные органы
Рис. 2. ВИЧ стыкуется и связывается с рецептором CD4, гликопротеином на поверхности Т-клеток, перед тем, как проникнуть в клетку или инфицировать ее. (кредит: модификация работы Национальных институтов здравоохранения США / Национального института аллергии и инфекционных заболеваний)
Гликопротеиновые и гликолипидные структуры на поверхности клеток дают многим вирусам возможность инфицирования. Вирусы ВИЧ и гепатита поражают только определенные органы или клетки в организме человека.ВИЧ способен проникать через плазматические мембраны подтипа лимфоцитов, называемых Т-хелперами, а также через некоторые моноциты и клетки центральной нервной системы. Вирус гепатита атакует клетки печени.
Эти вирусы способны проникать в эти клетки, потому что на поверхности клеток есть сайты связывания, которые специфичны и совместимы с определенными вирусами (рис. 2). Другие сайты узнавания на поверхности вируса взаимодействуют с иммунной системой человека, побуждая организм вырабатывать антитела.Антитела образуются в ответ на антигены или белки, связанные с инвазивными патогенами, или в ответ на чужеродные клетки, например, при трансплантации органов. Эти же сайты служат местами для прикрепления антител и либо уничтожения, либо подавления активности вируса. К сожалению, эти сайты узнавания на ВИЧ меняются очень быстро из-за мутаций, что делает эффективную вакцину против вируса очень сложной по мере того, как вирус эволюционирует и адаптируется. У человека, инфицированного ВИЧ, быстро разовьются разные популяции или варианты вируса, которые различаются по сайтам распознавания.Это быстрое изменение поверхностных маркеров снижает эффективность иммунной системы человека в атаке вируса, поскольку антитела не распознают новые вариации поверхностных структур. В случае ВИЧ проблема усугубляется тем, что вирус специфически инфицирует и разрушает клетки, участвующие в иммунном ответе, еще больше выводя из строя хозяина.
Вкратце: Структура клеточной мембраны
Современное понимание плазматической мембраны называется моделью жидкой мозаики.Плазматическая мембрана состоит из бислоя фосфолипидов с их гидрофобными хвостами жирных кислот, контактирующими друг с другом. Ландшафт мембраны усыпан белками, некоторые из которых покрывают мембрану. Некоторые из этих белков служат для транспортировки материалов в клетку или из клетки. Углеводы присоединяются к некоторым белкам и липидам на обращенной наружу поверхности мембраны. Они образуют комплексы, которые функционируют, чтобы идентифицировать клетку для других клеток. Жидкая природа мембраны обязана конфигурации хвостов жирных кислот, присутствию холестерина, встроенного в мембрану (в клетках животных), и мозаичному характеру белков и комплексов белок-углевод, которые не закреплены прочно. место.Плазменные мембраны окружают границы клеток, но они не статичны, а динамичны и постоянно изменяются.
Внесите свой вклад!
У вас была идея улучшить этот контент? Нам очень понравится ваш вклад.
Улучшить страницуПодробнее
Функция и структура клеточной мембраны
Клеточная мембрана (плазматическая мембрана) — это тонкая полупроницаемая мембрана, которая окружает цитоплазму клетки. Его функция заключается в защите целостности внутренней части клетки, позволяя одним веществам проникать в клетку, не допуская попадания других веществ.Он также служит основой для прикрепления цитоскелета у одних организмов и клеточной стенки у других. Таким образом, клеточная мембрана также помогает поддерживать клетку и помогает поддерживать ее форму.
Ключевые выводы
- Клеточная мембрана — это многогранная мембрана, которая окружает цитоплазму клетки. Он защищает целостность клетки, а также поддерживает клетку и помогает поддерживать форму клетки.
- Белки и липиды являются основными компонентами клеточной мембраны.Точная смесь или соотношение белков и липидов может варьироваться в зависимости от функции конкретной клетки.
- Фосфолипиды — важные компоненты клеточных мембран. Они спонтанно образуют липидный бислой, который является полупроницаемым, так что только определенные вещества могут диффундировать через мембрану внутрь клетки.
- Подобно клеточной мембране, некоторые клеточные органеллы окружены мембранами. Ядро и митохондрии — два примера.
Другая функция мембраны — регулировать рост клеток за счет баланса эндоцитоза и экзоцитоза.При эндоцитозе липиды и белки удаляются из клеточной мембраны по мере интернализации веществ. При экзоцитозе везикулы, содержащие липиды и белки, сливаются с клеточной мембраной, увеличивая размер клетки. Клетки животных, клетки растений, прокариотические клетки и клетки грибов имеют плазматические мембраны. Внутренние органеллы также покрыты мембранами.
Липиды клеточной мембраны
Вид фосфолипидов под микроскопом.Stocktrek Images / Getty Images
Фосфолипиды являются основным компонентом клеточных мембран.Фосфолипиды образуют липидный бислой, в котором их гидрофильные (привлеченные водой) области головы спонтанно располагаются так, чтобы быть обращенными к водному цитозолю и внеклеточной жидкости, в то время как их гидрофобные (отталкиваемые водой) участки хвоста обращены в сторону от цитозоля и внеклеточной жидкости. Липидный бислой является полупроницаемым, что позволяет только определенным молекулам диффундировать через мембрану.
Холестерин — еще один липидный компонент мембран клеток животных. Молекулы холестерина селективно распределены между фосфолипидами мембран.Это помогает удерживать клеточные мембраны от жесткости, предотвращая слишком плотную упаковку фосфолипидов. Холестерин не содержится в мембранах растительных клеток.
Гликолипиды расположены на поверхности клеточных мембран и имеют присоединенную к ним углеводную сахарную цепь. Они помогают клетке распознавать другие клетки тела.
Белки клеточной мембраны
Липопротеины и PCSK9 связываются с рецепторами.МАУРИЦИО ДЕ АНДЖЕЛИС / НАУЧНАЯ ФОТОБИБЛИОТЕКА / Getty Images
Клеточная мембрана содержит два типа связанных белков. Белки периферической мембраны находятся вне мембраны и связаны с ней посредством взаимодействия с другими белками. Интегральные мембранные белки вставлены в мембрану и в большинстве своем проходят через мембрану. Части этих трансмембранных белков открыты с обеих сторон мембраны. Белки клеточной мембраны выполняют ряд различных функций.
Структурные белки помогают придавать клеткам поддержку и форму.
Белки рецептора клеточной мембраны помогают клеткам общаться с внешней средой с помощью гормонов, нейротрансмиттеров и других сигнальных молекул.
Транспортные белки , такие как глобулярные белки, переносят молекулы через клеточные мембраны посредством облегченной диффузии.
Гликопротеины имеют прикрепленную к ним углеводную цепь. Они встроены в клеточную мембрану и помогают межклеточной коммуникации и транспорту молекул через мембрану.
Клеточная мембрана— полное руководство
Определение
Клеточная мембрана, также известная как плазматическая мембрана, представляет собой двойной слой липидов и белков, окружающий клетку.Он отделяет цитоплазму (содержимое клетки) от внешней среды. Это свойство всех клеток, как прокариотических, так и эукариотических.
Трехмерная диаграмма клеточной мембраны
Функция клеточной мембраны
Клеточная мембрана придает клетке ее структуру и регулирует материалы, которые входят в клетку и покидают ее. Это избирательно проницаемый барьер, то есть он позволяет одним веществам проникать, но не другим. Подобно разводному мосту, предназначенному для защиты замка и отпугивания врагов, клеточная мембрана позволяет входить или выходить только определенным молекулам.
Пересечение мембраны
Небольшие молекулы, такие как кислород, который нужен клеткам для выполнения метаболических функций, таких как клеточное дыхание, и углекислый газ, побочный продукт этих функций, могут легко входить и выходить через мембрану. Вода также может свободно проходить через мембрану, хотя и медленнее.
Однако высокозарядные молекулы, такие как ионы, не могут проходить напрямую, как и большие макромолекулы, такие как углеводы или аминокислоты.Вместо этого эти молекулы должны проходить через белки, встроенные в мембрану. Таким образом, клетка может контролировать скорость диффузии этих веществ.
Другой способ, которым клеточная мембрана может переносить молекулы в цитоплазму, — это эндоцитоз. Обратный процесс, когда клетка доставляет содержимое за пределы мембранного барьера, называется экзоцитозом.
Эндоцитоз включает фагоцитоз («поедание клеток») и пиноцитоз («питье клеток»). Во время этих процессов клеточная мембрана образует углубление, окружающее частицу, которую она поглощает.Затем он «отщипывается», образуя небольшую мембранную сферу, называемую пузырьком, которая содержит молекулу и транспортирует ее туда, где она будет использоваться в клетке.
Большие молекулы могут попадать в клетку в процессе эндоцитоза.
Клетки также могут доставлять вещества через клеточную мембрану во внешнюю среду посредством экзоцитоза, который противоположен эндоцитозу. Во время экзоцитоза везикулы образуются в цитоплазме и перемещаются на поверхность клеточной мембраны.Здесь они сливаются с мембраной и выпускают свое содержимое за пределы клетки. Экзоцитоз удаляет продукты жизнедеятельности клетки, которые являются частями молекул, которые клетка не использует, включая старые органеллы.
Передача сигналов на клеточной мембране
Клеточная мембрана также играет важную роль в передаче сигналов и коммуникации клеток. Мембрана содержит несколько встроенных белков, которые могут связывать молекулы, находящиеся вне клетки, и передавать сообщения внутрь клетки.
Важно отметить, что эти рецепторные белки на клеточной мембране могут связываться с веществами, вырабатываемыми другими частями тела, такими как гормоны. Когда молекула связывается со своим рецептором-мишенью на мембране, она инициирует путь передачи сигнала внутри клетки, который передает сигнал соответствующим молекулам.
В результате этих часто сложных сигнальных путей клетка может выполнять действие, определяемое сигнальной молекулой, например производство или остановку продукции определенного белка.
Как структура клеточной мембраны позволяет ей выполнять эти функции?
Структура клеточной мембраны
Двухслойный фосфолипид
Клеточная мембрана состоит из бислоя фосфолипидов. Фосфолипиды — это липидные молекулы, состоящие из головы фосфатной группы и двух хвостов жирных кислот. Важно отметить, что свойства молекул фосфолипидов позволяют им спонтанно образовывать двухслойную мембрану.
Головка фосфатной группы фосфолипида является гидрофильной, тогда как хвостовая часть фосфолипида гидрофобна.Это означает, что фосфатная группа притягивается к воде, тогда как хвост отталкивается водой.
Находясь в воде или водном растворе (в том числе внутри тела), гидрофобные головки фосфолипидов будут располагаться внутри, как можно дальше от воды. Напротив, гидрофильные головки будут снаружи, контактируя с водой. В результате образуется двойной слой фосфолипидов, причем гидрофобные головки сгруппированы вместе в центре, а гидрофильные хвосты образуют внешнюю часть структуры.Технический термин для этого двойного слоя фосфолипидов, который формирует клеточную мембрану, — это бислой фосфолипидов.
Структура клеточной мембраны и связанных с ней компонентов
Мембранные факторы
Помимо фосфолипидного бислоя клеточная мембрана также содержит липидные молекулы, особенно гликолипиды и стерины. Одним из важных стеринов является холестерин, который регулирует текучесть клеточной мембраны в клетках животных. Когда холестерина меньше, мембраны становятся более жидкими, но при этом более проницаемыми для молекул.Количество холестерина в мембране помогает поддерживать ее проницаемость, так что нужное количество молекул может проникать в клетку за раз.
Клеточная мембрана также содержит множество различных белков. Белки составляют около половины клеточной мембраны. Многие из этих белков являются трансмембранными белками, которые встроены в мембрану, но выступают с обеих сторон (т. Е. Охватывают весь липидный бислой).
Некоторые из этих белков являются рецепторами, которые связываются с сигнальными молекулами.Другие представляют собой ионные каналы, которые являются единственным средством, позволяющим ионам проникать в клетку или выходить из нее. Ученые используют модель жидкой мозаики для описания структуры клеточной мембраны. Клеточная мембрана имеет жидкую консистенцию из-за того, что большая часть состоит из фосфолипидов, и из-за этого белки свободно перемещаются по ее поверхности. Множество различных белков и липидов в клеточной мембране придают ей мозаичный вид.
Викторина
Библиография
Показать / Скрыть
Альбертс, Б., Johnson, A., Lewis, J., et al. Молекулярная биология клетки. 4-е издание. Нью-Йорк: наука о гирляндах; 2002. Липидный бислой. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK26871/
- Лодиш, Х., Берк, А., Зипурски, С.Л. и др. Молекулярная клеточная биология. 4-е издание. Нью-Йорк: У. Х. Фриман; 2000. Раздел 3.4, Мембранные белки. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK21570/
- Уотсон, Х. (2015). Биологические мембраны. Очерки биохимии , 59 , 43–69. https://doi.org/10.1042/bse05
Структура плазменной мембраны — MHCC Biology 112: Biology for Health Professions
Клетки тщательно контролируют обмен веществ внутри клетки и из нее. Некоторые вещества исключаются, другие принимаются внутрь, а третьи выводятся из организма — все в контролируемых количествах. Хотя плазматическая мембрана (клеточная мембрана) окружает границы клетки, она далеко не статический барьер; он динамичен и постоянно меняется.Плазматическая мембрана должна быть достаточно гибкой, чтобы определенные клетки, такие как эритроциты и лейкоциты, могли изменять форму при прохождении через узкие капилляры. В дополнение к этим более очевидным функциям поверхность плазматической мембраны несет маркеры, которые позволяют клеткам узнавать друг друга. Это жизненно важно, поскольку эти маркеры играют роль в различении иммунного ответа «я» и «не-я».
В 1972 г. С. Дж. Сингер и Гарт Л. Николсон предложили новую модель плазматической мембраны.Эта теория, по сравнению с более ранними теориями, лучше всего объясняет как микроскопические наблюдения, так и функцию плазматической мембраны. Эта теория получила название модели жидкой мозаики . Модель со временем несколько эволюционировала, но по-прежнему лучше всего объясняет структуру и функции плазматической мембраны в том виде, в котором мы их теперь понимаем. Модель жидкой мозаики описывает структуру плазматической мембраны как состоящую из различных компонентов, включая фосфолипиды, холестерин, белки и углеводы, которые способны течь и менять положение, сохраняя при этом основную целостность мембраны.Как молекулы фосфолипидов, так и встроенные белки могут перемещаться в мембране латерально. Текучесть плазматической мембраны необходима для активности определенных ферментов и транспортных молекул внутри мембраны.
Плазменные мембраны имеют толщину 5–10 нм. Для сравнения, красные кровяные тельца человека, видимые с помощью световой микроскопии, имеют толщину примерно 8 мкм, или примерно в 1000 раз толще плазматической мембраны.
Рисунок 1 Жидкая мозаичная модель структуры плазматической мембраны описывает плазматическую мембрану как жидкую комбинацию фосфолипидов, холестерина, белков и углеводов.Плазматическая мембрана состоит в основном из бислоя фосфолипидов со встроенными белками, углеводами, гликолипидами и гликопротеинами, а в клетках животных — холестерином (, рис. 1, ).
Фосфолипиды
Основная ткань мембраны состоит из двух слоев молекул фосфолипидов, и полярные концы этих молекул (которые выглядят как набор шариков в изображении модели художником) ( Рисунок 2 ) находятся в контакте с водной жидкость как внутри, так и снаружи клетки.Таким образом, обе поверхности плазматической мембраны являются гидрофильными и («любящими воду»). Напротив, внутренняя часть мембраны между двумя ее поверхностями представляет собой гидрофобную («водобоязненную») или неполярную область из-за хвостов жирных кислот. В этой области нет притяжения для воды или других полярных молекул.
Рисунок 2 Фосфолипидный бислой. «Внеклеточный» = вне клетки; «Внутриклеточный» = внутри клетки. Фотография предоставлена: Анатомия и физиология OpenStax.Молекула фосфолипида ( Рисунок 3 ) состоит из трехуглеродного глицеринового остова с двумя молекулами жирных кислот, присоединенными к атомам углерода 1 и 2, и фосфатсодержащей группой, присоединенной к третьему атому углерода.Такое расположение дает всей молекуле область, описываемую как ее голова (фосфатсодержащая группа), которая имеет полярный характер или отрицательный заряд, и область, называемую хвостом (жирные кислоты), которая не имеет заряда. Голова может образовывать водородные связи, а хвост — нет.
Рисунок 3 Эта молекула фосфолипида состоит из гидрофильной головки и двух гидрофобных хвостов. Гидрофильная головная группа состоит из фосфатной группы, присоединенной к молекуле глицерина. Гидрофобные хвосты, каждый из которых содержит насыщенную или ненасыщенную жирную кислоту, представляют собой длинные углеводородные цепи.Белки
Белки составляют второй главный химический компонент плазматических мембран (см. Рисунок 1 ). Белки встроены в плазматическую мембрану и могут полностью проходить через мембрану или обнаруживаться с одной или другой стороны ( Рисунок 1, ). Белки могут служить ферментами, структурными прикреплениями волокон цитоскелета или частью сайтов узнавания клетки. Белки, которые проходят через мембрану, могут служить каналами или насосами для перемещения материалов в клетку или из клетки.Белки также находятся либо на внешней, либо на внутренней поверхности мембран, где они могут быть прикреплены либо к другим белкам, либо к молекулам фосфолипидов ( Рисунок 1 ).
Ниже описаны пять различных типов белков, которые обычно связаны с клеточной мембраной.
Белки распознавания (идентификации)
Гликопротеины — это белки, к которым прикреплены молекулы сахара. Гликопротеины помогают клеткам узнавать друг друга.Уникальные структуры молекул сахара на поверхности клетки могут быть «прочитаны» соответствующими гликопротеинами на другой клетке. Однако этот процесс отличается от того, как мы читаем слова. Это больше похоже на вставку ключа в замок; гликопротеины часто связываются вместе, если они совпадают. Этот процесс привязки сообщает ячейке, что она нашла совпадение. Это то, что помогает нашей иммунной системе распознавать чужеродных захватчиков, а затем защищать наш организм от них. Верно и обратное. Вирусы используют гликопротеины для связывания и заражения клетки-хозяина.Распознавание гликопротеинов также помогает в репродукции.
Рецепторные белки
Рецепторы клеточной поверхности (мембранные рецепторы, трансмембранные рецепторы) — это рецепторы, встроенные в мембраны клеток. Рецептор представляет собой тип белка распознавания, который может избирательно связывать определенную молекулу вне клетки, и это связывание вызывает химическую реакцию внутри клетки. Лиганд — это специфическая молекула, которая связывается с рецептором и активирует его. Молекулы лиганда могут быть гормонами, нейротрансмиттерами, цитокинами, факторами роста, молекулами клеточной адгезии или питательными веществами; они реагируют с рецептором, вызывая изменения в метаболизме и активности клетки.
Каждый рецептор структурирован для связывания с определенным веществом. Связывание определенного вещества с его рецептором на плазматической мембране может активировать процессы внутри клетки, такие как активация ферментов, участвующих в метаболических путях. Эти метаболические пути могут иметь жизненно важное значение для обеспечения клетки энергией, производства веществ для клетки или разложения клеточных отходов или токсинов для утилизации. Точно так же внеклеточные гормоны и нейротрансмиттеры связываются с рецепторами плазматической мембраны, которые передают сигнал в клетку внутриклеточным молекулам.Некоторые сайты распознавания используются вирусами как точки прикрепления. Хотя они очень специфичны, патогены, такие как вирусы, могут развиваться, чтобы использовать рецепторы, чтобы проникнуть в клетку, имитируя конкретное вещество, которое рецептор должен связывать. Эта специфичность помогает объяснить, почему вирус иммунодефицита человека (ВИЧ) или любой из пяти типов вирусов гепатита проникает только в определенные клетки.
Белки движения
Канальные белки позволяют перемещать материалы с одной стороны мембраны на другую, не требуя энергии.Движущаяся молекула захватывается с одной стороны белка канала, и без использования энергии молекула попадает в клетку. Каналы используются для облегченной диффузии, которая является транспортом, особенно для больших полярных молекул и заряженных ионов, которые не могут свободно диффундировать через клеточные мембраны из-за гидрофобной природы жирнокислотных хвостов фосфолипидов, составляющих бислои.
Белки-носители используют энергию для перемещения молекул через мембрану.Транспортируемое химическое вещество должно сначала связываться в сайте связывания на белке-носителе. После связывания носитель примет и удержит перемещаемый материал. Затем белок-носитель меняет форму, так что отверстие в белке теперь обращено к другой стороне плазматической мембраны. Затем транспортируемый материал высвобождается с этой стороны мембраны.
Рисунок 4 Несколько мембранных белков. Канальный белок в центре содержит пору или канал, по которому может перемещаться материал.Из OpenStax Anatomy and PhysiologyAquaporins , также называемый водными каналами, представляет собой трансмембранный белок, который избирательно проводит молекулы воды в клетку и из нее, предотвращая прохождение ионов и других растворенных веществ. Молекулы воды движутся через поры канала одним файлом (рис. 5). Присутствие аквапоринов увеличивает проницаемость мембраны для воды. Молекулы воды проходят через поры канала единой цепью. Наличие водных каналов увеличивает проницаемость мембраны для воды.
Рисунок 5 Структура аквапорина или «белка водного канала».Муковисцидоз вызывается дефектом интегрального белка клеточной мембраны, который действует как канал. Белок CFTR перемещает ионы с одной стороны мембраны на другую. Когда он не функционирует должным образом, это вызывает скопление очень густой слизи в легких и пищеварительном тракте.
Когда белок канала CFTR функционирует правильно (1), ионы (маленькие шарики) могут проходить через мембрану.Когда он не функционирует должным образом (2), ионы не могут пересекать мембрану. Фото: LBudd14, май 2013 г. Викимедиа.Углеводы
Углеводы — третий важный компонент плазматических мембран. Они всегда находятся на внешней поверхности клеток и связаны либо с белками (образуя гликопротеинов ), либо с липидами (образуя гликолипидов ). Эти углеводные цепи могут состоять из 2-60 моносахаридных звеньев и могут быть прямыми или разветвленными.
Гликопротеины и гликолипиды образуют специализированные участки на поверхности клетки, которые позволяют клеткам узнавать друг друга. На этих участках есть уникальные узоры, которые позволяют распознавать клетку, во многом так же, как черты лица, уникальные для каждого человека, позволяют его или ее узнавать. Эта функция распознавания очень важна для клеток, поскольку она позволяет иммунной системе различать клетки тела (называемые «самими») и чужеродные клетки или ткани (так называемые «чужие»). Подобные типы гликопротеинов и гликолипидов находятся на поверхности вирусов и могут часто меняться, не позволяя иммунным клеткам распознавать их и атаковать их.
Углеводы, входящие в состав гликопротеинов, определяют группы крови человека A, B, O. Эти гликопротеины распознаются иммунной системой, что приводит к несовместимости по группам крови.
ABO Группы крови. На этом рисунке мембранный углевод представлен «леденцами». Их называют «антигенами». Фото: InvictaHOG, 2006. Викимедиа.Мозаичная характеристика мембраны, описанная в модели жидкой мозаики, помогает проиллюстрировать ее природу.Белки и другие компоненты, присутствующие в мембране, могут перемещаться относительно друг друга, как лодки, плавающие в озере. Однако мембрана не похожа на воздушный шар, который может расширяться и сжиматься; скорее, он довольно жесткий и может лопнуть, если в него проникнуть или если ячейка впитает слишком много воды. Однако из-за своей мозаичности очень тонкая игла может легко проникнуть в плазматическую мембрану, не вызывая ее разрыва, и мембрана будет течь и самоуплотняться при извлечении иглы.
Мозаичные характеристики мембраны частично объясняют ее текучесть. Есть два других фактора, которые помогают поддерживать эту характеристику жидкости. Одним из факторов является природа самих фосфолипидов. Структура хвостов жирных кислот в каждом фосфолипиде может сделать мембрану более плотной и жесткой или менее плотной и гибкой. Относительная текучесть мембраны особенно важна в холодных условиях. Холодная среда делает мембраны менее жидкими и более восприимчивыми к разрыву.Многие организмы (например, рыба) способны адаптироваться к холоду, изменяя пропорцию различных типов жирных кислот в своих мембранах в ответ на понижение температуры.
Животные имеют дополнительную мембранную составляющую, которая помогает поддерживать текучесть. Холестерин, , который находится рядом с фосфолипидами в мембране, имеет тенденцию ослаблять влияние температуры на мембрану. Таким образом, этот липид действует как буфер, предотвращая снижение текучести при низких температурах и предотвращая чрезмерное повышение текучести при повышенных температурах.Таким образом, холестерин расширяет в обоих направлениях диапазон температур, в котором мембрана является подходящей текучей и, следовательно, функциональной. Холестерин также выполняет другие функции, такие как организация кластеров белков в липидные рафты.
Если не указано иное, изображения на этой странице лицензированы OpenStax в соответствии с CC-BY 4.0.
Текст адаптирован из: OpenStax, Концепции биологии. OpenStax CNX. 18 мая 2016 г. http://cnx.org/contents/[email protected]
Молекулярные выражения Клеточная биология: плазменная мембрана
Плазменная мембрана
Все живые клетки, прокариотические и эукариотические, имеют плазматическую мембрану, которая закрывает их содержимое и служит полупористым барьером для внешней среды. Мембрана действует как граница, удерживая компоненты клетки вместе и препятствуя проникновению других веществ. Однако плазматическая мембрана проницаема для определенных молекул и позволяет питательным веществам и другим важным элементам проникать в клетку, а отходам — покидать клетку.Небольшие молекулы, такие как кислород, углекислый газ и вода, могут свободно проходить через мембрану, но прохождение более крупных молекул, таких как аминокислоты и сахара, тщательно регулируется.
Согласно принятой в настоящее время теории, известной как модель жидкой мозаики , плазматическая мембрана состоит из двойного слоя ( бислой ) липидов, маслянистых веществ, присутствующих во всех клетках (см. Рисунок 1). Большинство липидов в бислое можно более точно описать как фосфолипиды , то есть липиды, которые имеют фосфатную группу на одном конце каждой молекулы.Фосфолипиды обычно являются гидрофильными («водолюбивыми») на их фосфатных концах и гидрофобными («водобоязненными») вдоль их липидных хвостовых областей. В каждом слое плазматической мембраны гидрофобные липидные хвосты ориентированы внутрь, а гидрофильные фосфатные группы выровнены так, что они обращены наружу, либо к водному цитозолю клетки, либо к внешней среде. Фосфолипиды имеют тенденцию к спонтанной агрегации по этому механизму всякий раз, когда они подвергаются воздействию воды.
Внутри фосфолипидного бислоя плазматической мембраны встроено множество различных белков, тогда как другие белки просто прикрепляются к поверхностям бислоя. Некоторые из этих белков, в первую очередь те, которые, по крайней мере, частично открыты на внешней стороне мембраны, имеют углеводы, прикрепленные к их внешней поверхности, и поэтому называются гликопротеинами . Расположение белков вдоль плазматической мембраны частично связано с организацией филаментов, составляющих цитоскелет, которые помогают закрепить их на месте.Расположение белков также включает гидрофобные и гидрофильные области, обнаруженные на поверхности белков: гидрофобные области связаны с гидрофобной внутренней частью плазматической мембраны, а гидрофильные области простираются за поверхность мембраны либо во внутреннюю часть клетки, либо во внешнюю. среда.
Белки плазматической мембраны функционируют по-разному. Многие из белков играют роль в селективном транспорте определенных веществ через бислой фосфолипидов, действуя либо как каналы, либо как активные транспортные молекулы.Другие действуют как рецепторы, которые связывают молекулы, обеспечивающие информацию, такие как гормоны, и передают соответствующие сигналы на основе полученной информации внутрь клетки. Мембранные белки также могут проявлять ферментативную активность, катализируя различные реакции, связанные с плазматической мембраной.
С 1970-х годов плазматическую мембрану часто описывают как жидкую мозаику , что отражает открытие, что часто липидные молекулы в бислое могут перемещаться в плоскости мембраны.Однако в зависимости от ряда факторов, включая точный состав бислоя и температуру, плазменные мембраны могут претерпевать фазовых переходов , которые делают их молекулы менее динамичными и создают более гелеобразное или почти твердое состояние. Клетки способны регулировать текучесть своих плазматических мембран в соответствии со своими конкретными потребностями, синтезируя больше определенных типов молекул, например, с определенными типами связей, которые поддерживают их текучесть при более низких температурах.Присутствие холестерина и гликолипидов, которые содержатся в большинстве клеточных мембран, также может влиять на молекулярную динамику и ингибировать фазовые переходы.
У прокариот и растений плазматическая мембрана является внутренним слоем защиты, поскольку жесткая клеточная стенка образует внешнюю границу для их клеток. Стенка клетки имеет поры, которые позволяют материалам входить в клетку и покидать ее, но они не очень избирательны в отношении того, что проходит через нее. Плазматическая мембрана, выстилающая клеточную стенку, обеспечивает окончательный фильтр между внутренней частью клетки и окружающей средой.
Считается, что клетки эукариотических животных произошли от прокариот, утративших свои клеточные стенки. Оставив только гибкую плазматическую мембрану, окружающую их, эти первобытные существа смогли бы увеличиваться в размерах и становиться сложнее. Эукариотические клетки обычно в десять раз больше, чем прокариотические клетки, и имеют мембраны, содержащие внутренние компоненты, органеллы. Как и внешняя плазматическая мембрана, эти мембраны также регулируют поток материалов, позволяя клетке разделять свои химические функции на отдельные внутренние части.
НАЗАД К СТРУКТУРЕ ЖИВОТНОЙ КЛЕТКИ
НАЗАД К СТРУКТУРЕ КЛЕТКИ ЗАВОДА
Вопросы или комментарии? Отправить нам письмо.
© 1995-2021, автор — Майкл В. Дэвидсон и Государственный университет Флориды. Все права защищены. Никакие изображения, графика, программное обеспечение, сценарии или апплеты не могут быть воспроизведены или использованы каким-либо образом без разрешения правообладателей. Использование этого веб-сайта означает, что вы соглашаетесь со всеми юридическими положениями и условиями, изложенными владельцами.
Этот веб-сайт обслуживается нашим
Команда разработчиков графики и веб-программирования
в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля
.
Последнее изменение: пятница, 13 ноября 2015 г., 14:18
Счетчик доступа с 1 октября 2000 г .: 1429761
Микроскопы предоставленные:
Клеточная мембрана— обзор
II Бимолекулярная липидная мембрана
Термин плазматическая мембрана происходит от немецкого Plasmamembran, слова, придуманного Карлом Вильгельмом Нэгели (1817–1891) для описания твердой пленки, которая образуется, когда белковый сок поврежденной клетки контактирует с водой.Физиолог Л.В. Хейльбрунн назвал это и подобные явления «реакцией поверхностного осаждения», как описано в его книге The Dynamics of Living Protoplasm (1956). «Протоплазма» — это старый термин для обозначения вещества внутри клеток, который широко использовался до того, как методы электронной микроскопии и дифференциального центрифугирования помогли выяснить подробные структуры и специфические функции отдельных клеточных органелл. Участие того, что мы знаем сегодня как клеточная мембрана, и биохимия образования поверхностной пленки в ответ на повреждение клетки впоследствии не были объяснены.Таким образом, первоначальное использование термина «плазматическая мембрана» имеет неясное отношение к его нынешнему значению.
Многие знания о структуре и функциях мембран получены из исследований красных кровяных телец, как показано на микрофотографии, сделанной с помощью сканирующего электронного микроскопа, на рис. 3.1. Эритроциты высоко дифференцированы и специализируются на транспортировке кислорода и углекислого газа в крови. Они состоят в основном из плазматической мембраны, окружающей концентрированный раствор гемоглобина, и лишены ядра, митохондрий, эндоплазматического ретикулума, рибосом, аппарата Гольджи и лизосом.Более 100 лет назад обширные исследования осмотического давления и проницаемости эритроцитов Гамбургером, растительных клеток де Фриза и многих живых клеток Овертоном предоставили доказательства того, что липоидная мембрана окружает клетки. Жирорастворимые вещества, которые являются липофильными и легко растворяются в липидах, легко проникают в клетки, тогда как водорастворимые вещества поступают в клетки медленнее, если вообще. Овертон обнаружил корреляцию между коэффициентом разделения нефть-вода и проницаемостью мембраны; тем не менее, ни в одном из этих ранних исследований мембрана не постулировалась как отдельная структурная единица для объяснения результатов (см. Jacobs, 1962).
РИСУНОК 3.1. Сканирующая электронная микрофотография эритроцитов человека. Двояковогнутые дискоидные ячейки имеют диаметр 8 мкм, толщину 2,4 мкм по краю и 1,0 мкм в центре.
(от Bessis, M. 1974. Corpuscles. Атлас форм эритроцитов. Springer-Verlag, New York, рис. 1, с разрешения).В 1925 году Гортер и Грендель использовали ацетон для извлечения липидов из известного количества эритроцитов и, после испарения растворителя, измерили площадь, которую экстрагированные липиды занимали в виде мономолекулярной пленки на границе раздела воздух-вода, с помощью лотка Ленгмюра.По площади пленки экстрагированных липидов и площади поверхности эритроцитов, оцененной с помощью световой микроскопии, они пришли к выводу, что: «Очевидно, что все наши результаты хорошо согласуются с предположением о том, что хромоциты покрыты слоем. жирных веществ толщиной в две молекулы »(Гортер и Грендель, 1925, стр. 443). Однако примерно 40 лет спустя было отмечено, что площадь поверхности эритроцитов на самом деле на 50% больше; Кроме того, после экстракции ацетоном в призраках осталось около 30% липидов.К счастью, эти две ошибки, как правило, уравновешивали друг друга (Bar et al., 1966), показывая, что в редких случаях в науке вы можете быть правы по неправильным причинам. Бимолекулярный липидный листок толщиной 75–100 Å, впервые предложенный Гортером и Гренделем в качестве модели клеточной мембраны (рис. 3.2), до сих пор составляет основу современных представлений о структуре клеточных мембран. Суть этой модели состоит в том, что фосфолипиды мембраны расположены в параллельных слоях, образуя два полуслоя с их полярными головными группами, обращенными к водным внутриклеточным и внеклеточным растворам, а их неполярные цепи жирных кислот взаимодействуют латерально внутри гидрофобного ядра мембраны.
РИСУНОК 3.2. Модель бимолекулярной фосфолипидной мембраны, предложенная Гортером и Гренделем (1925).
Для чистых липидов ожидаемое поверхностное натяжение , измеренное в лотке Ленгмюра, составляет около 9 дин / см, но поверхностное натяжение морских яиц и других типов клеток примерно в 50–100 раз меньше всего за 0,1– 0,2 дин / см. Поверхностное натяжение можно представить как силу, необходимую для закрытия щели на поверхности мембраны. Даниелли и Харви обнаружили, что яичный белок может снизить поверхностное натяжение границы раздела масло-вода примерно до 0.6 дин / см, что привело Дэвсона и Даниелли (1943) к постулированию присутствия двух пленок белка, связанных с полярными головными группами на каждой стороне бимолекулярного липидного листка, модель, которая стала известна как пауцимолекулярная мембрана Дэвсона-Даниелли (Рис. 3.3). Предположительно, белок функционировал, чтобы укрепить и стабилизировать тонкую липидную пленку. Paucimolecular означает, что эта модель включала всего несколько молекул: бимолекулярный липидный листок с прилипшими белковыми пленками на внутренней и внешней поверхностях.
РИСУНОК 3.3. Пауцимолекулярная модель структуры мембраны.
(Из Даниелли, Дж. Ф. и Дэвсона, HA (1935). Вклад в теорию проницаемости тонких пленок. J Cell Comp Physiol. 5, 495–508, стр. 498, перепечатано с разрешения Wiley-Liss, Inc. , дочерняя компания John Wiley and Sons, Inc.)С помощью электрофизиологических методов было измерено электрическое сопротивление клеточных мембран, которое оказалось очень высоким, что также согласуется с предложением об изоляционной липидной мембране, окружающей клетки.В других исследованиях мембран было определено их двойное лучепреломление . Двулучепреломление — это оптическое свойство некоторых ориентированных материалов, которое можно определить, поместив образец между двумя скрещенными поляроидами на предметном столике микроскопа. Пленка Polaroid пропускает только свет, электрический вектор которого параллелен оси пленки Polaroid; две пленки Polaroid, пересеченные под прямым углом и поднятые к свету, кажутся черными. Но если кристалл или другое вещество, в котором ориентированы сами молекулы, поместить между скрещенными поляроидами, проходящий свет будет поляризован по кругу, и образец будет выглядеть ослепительно ярким.Образец, который кажется ярким при помещении между скрещенными поляроидами, называется двулучепреломляющим. Собственное двойное лучепреломление обусловлено ориентированной природой отдельных молекул, таких как нитчатые белки, тогда как формирует двойное лучепреломление обусловлено ориентированным расположением молекул в массиве, например, при параллельной упаковке актиновых и миозиновых нитей в мышцах. саркомеры. Когда мембраны эритроцитов просматривали с помощью поляризационного микроскопа, липиды вносили вклад в двойное лучепреломление, как и лежащий в основе цитоскелет, в соответствии с паучимолекулярной моделью структуры мембраны.
Электронные микрофотографии с высоким разрешением мембраны элемента также подтверждают паучимолекулярную модель Дэвсона – Даниелли. Термин элементарная мембрана относится к повсеместной триламинарной структуре толщиной 75–100 Å, наблюдаемой на электронных микрофотографиях тонких срезов клеток и органелл. Изображение выглядит как две темные линии, каждая толщиной около 25–30 ÅA, окружающие более светлую зону, и особенно хорошо разрешается в образцах, зафиксированных перманганатом калия. Электронно-микроскопический снимок тонкого среза плазматической мембраны эритроцитов человека показан на рис.3.4. Практически такая же триламинарная структура наблюдалась не только на поверхности эритроцитов, но также и в мышечных клетках, нервных клетках, эпителиальных клетках, растительных клетках, бактериальных клетках и практически во всех исследованных органеллах мембранных клеток. В ситуациях, когда две клетки плотно прилегали друг к другу, две триламинарные структуры составляли двойную мембрану . В миелиновой оболочке, окружающей нервные клетки, была видна серия триламинарных структур в спиральном расположении, согласующемся с оболочкой нервных аксонов мембраной шванновских клеток, как показано на рис.3.5A. Поскольку подробные химические реакции перманганата калия с тканью полностью не изучены, оставалась некоторая неопределенность в отношении основы изображения, которое наблюдалось в шлифах. Повсеместное распространение единичной мембраны, однако, было воспринято как веское свидетельство в поддержку паучимолекулярной модели Дэвсона-Даниелли. Дальнейшие исследования многослойной миелиновой оболочки нервных аксонов с помощью дифракции рентгеновских лучей центрифугированных незафиксированных и неокрашенных мембран дали профили электронной плотности, которые также согласуются с паучимолекулярной моделью Davson-Danielli (Worthington and McIntosh, 1973).Как видно на рис. 3.5B, электронная плотность низкая в гидрофобной сердцевине мембраны и высокая в полярных областях фосфатных групп. Более того, толщина мембраны, определенная на основе этих рассчитанных профилей электронной плотности неокрашенной миелиновой оболочки, количественно согласуется с толщиной мембраны в тонких срезах, фиксированных перманганатом калия — наблюдение, являющееся убедительным доказательством, подтверждающим предложение о бимолекулярном фосфолипиде. листочка как основа структуры клеточных мембран.Таким образом, доказательства для модели пауцимолекулярной мембраны состояли из исследований проницаемости, электрического сопротивления и микроскопических наблюдений двойного лучепреломления с помощью светового микроскопа, а также изображений с высоким разрешением как окрашенных, так и неокрашенных клеток с помощью электронного микроскопа. Эти убедительные аргументы были обобщены в классической монографии под названием The Permeability of Natural Membranes , написанной Дэвсоном и Даниелли (1943), книге, которая сильно повлияла на последующее развитие клеточной и мембранной физиологии.
РИСУНОК 3.4. Электронная микрофотография тонкого среза единичной мембраны эритроцита.
(микрофотография Дж. Д. Робертсона из Дайсона, Р. Д. (1974). Клеточная биология. Молекулярный подход. Аллин и Бэкон, Бостон, с разрешения).РИСУНОК 3.5. (A) Миелиновая оболочка аксона спинного мозга (любезно предоставлена доктором Седриком Рейном). (B) Профиль электронной плотности седалищного нерва лягушки.
(From Worthington, C.R. and McIntosh, T.J. (1973). Прямое определение профиля электронной плотности нервного миелина.Природа-новая биология. 245, 97–99, с. 99. Перепечатано с разрешения Nature , авторское право 1973 Macmillan Magazines Ltd.)16.2: Структура плазматической мембраны — Biology LibreTexts
В эукариотических клетках плазматическая мембрана окружает цитоплазму, заполненную рибосомами и органеллами. Органеллы — это структуры, которые сами заключены в мембраны. Некоторые органеллы (ядра, митохондрии, хлоропласты) даже окружены двойными мембранами. Все клеточные мембраны состоят из двух слоев фосфолипидов, залитых белками.Все они избирательно проницаемы (полупроницаемы), позволяя только определенным веществам проходить через мембрану. Уникальные функции клеточных мембран обусловлены их различным фосфолипидным и белковым составом. Десятилетия исследований выявили эти функции (например, см. Предыдущие обсуждения функции митохондрий и хлоропластов). Здесь мы опишем общие характеристики мембран на нашем примере плазматической мембраны.
A. Фосфолипидный бислой
Гортер и Грендель предсказали структуру двухслойной мембраны еще в 1925 году.Они знали, что красные кровяные тельца (эритроциты) не имеют ядра или других органелл и, следовательно, имеют только плазматическую мембрану. Они также знали, что основным химическим компонентом этих мембран являются фосфолипиды . Молекулярная модель, заполняющая пространство ниже, показывает основную структуру фосфолипидов, выделяя их гидрофильных (полярных) головок и гидрофобных хвостов.
Молекулы с гидрофильными и гидрофобными доменами — это амфипатических молекул .Гортер и Грендель измерили площадь поверхности красных кровяных телец. Затем они сделали «анализ крови» и разрушили известное количество красных кровяных телец. Затем они измерили количество фосфолипидов в экстрактах мембран. Исходя из этого, они подсчитали, что на клетку достаточно молекул липидов, чтобы дважды обернуть каждую клетку. На основании этих наблюдений они предсказали фосфолипидный бислой с жирными кислотами, взаимодействующими внутри бислоя. Любопытно, что Гортер и Грендель сделали две ошибки в расчетах при определении количества фосфолипидов на клетку.Тем не менее их ошибки компенсировали друг друга, так что их выводы, хотя и не были, строго говоря, правильными, оставались пророческими! Общие мембранные фосфолипиды показаны ниже.
Амфипатические молекулы, смешанные с водой, спонтанно объединяются, чтобы «скрыть» свои гидрофобные области от воды. В воде они образовали настоящие структуры, называемые липосомами, которые оседают при центрифугировании!
276 Мембранные липиды и бислой фосфолипидов
277 Эксперименты с липосомами и их использование
Структура липосомной мембраны соответствует предсказанному фосфолипидному бислою, при этом гидрофобные хвосты взаимодействуют друг с другом, а полярные головки обращены друг от друга, образуя фосфолипидный бислой .Это привело к картине мембранной архитектуры, основанной на взаимодействиях фосфолипидов. Характерная иллюстрация фосфолипидного бислоя с его внутренней гидрофобной жирной кислотой и гидрофильной внешней поверхностью представлена ниже.
Структура липосомной мембраны соответствует предсказанному фосфолипидному бислою, при этом гидрофобные хвосты взаимодействуют друг с другом, а полярные головки обращены друг от друга, образуя фосфолипидный бислой. Это привело к картине мембранной архитектуры, основанной на взаимодействиях фосфолипидов.Характерная иллюстрация фосфолипидного бислоя с его внутренней гидрофобной жирной кислотой и гидрофильной внешней поверхностью представлена ниже.
B. Модели структуры мембраны
В 1935 году Дэвсон и Даниелли предположили, что белки могут быть связаны с полярными головками фосфолипидов в плазматической мембране, создавая сэндвич белок / липид / белок. Десятилетия спустя Дж.Д. Робертсон наблюдал мембраны в просвечивающем электронном микроскопе при высоком увеличении, обнаружив, что все клеточные мембраны имеют трехламеллярную структуру .Классический вид трехламеллярной клеточной мембраны в электронном микроскопе проиллюстрирован ниже
.Триламеллярная структура согласуется с покрытыми белком гидрофильными поверхностями фосфолипидного бислоя в белково-липид-белковом сэндвиче Дэвсона и Даниелли. Заметив, что все клеточные мембраны имеют эту трехламеллярную структуру, Робертсон далее предложил свою модель Unit Membrane : все мембраны состоят из прозрачного фосфолипидного бислоя, покрытого электронно-плотными белками.
Статический вид трехламеллярных моделей мембранной структуры, подразумеваемый моделями Дэвсона-Даниелли или Робертсона, был заменен в 1972 году моделью Fluid Mosaic Зингера и Николсона (см. Модель жидкой мозаики мембран. Science 175: 720-731 ). Они предположили, что в дополнение к периферическим белкам , которые и связываются с поверхностями мембран, многие интегральные мембранные белки фактически охватывают мембрану. Интегральные мембранные белки представляли собой мозаику из белковых «плиток», заключенных в фосфолипидную среду.Но в отличие от мозаики из глазурованных плиток, уложенных в твердую цементоподобную структуру, белковые «плитки», по прогнозам, будут подвижными (, жидкость, ) в море фосфолипидов . В этой модели мембранные белки закреплены в мембранах одним или несколькими гидрофобными доменами; их гидрофильные домены будут сталкиваться с водной внешней и цитозольной средой. Таким образом, как и сами фосфолипиды, мембранные белки являются амфипатическими . Мы знаем, что клетки демонстрируют различные структурные (и функциональные) особенности поверхности водной среде на противоположных сторонах мембраны.Поэтому мы также говорим, что клеточные мембраны асимметричны . Типичная модель плазматической мембраны клетки проиллюстрирована ниже.
В этой модели периферические белки имеют гидрофобный домен, который не охватывает мембрану, но прикрепляет его к одной стороне мембраны. Другие периферические (или так называемые « поверхности ») белки связаны с мембраной посредством взаимодействий с полярными фосфатными группами фосфолипидов или с полярными доменами интегральных мембранных белков.
Благодаря своим собственным водным гидрофильным доменам мембранные белки являются естественным барьером для свободного прохождения заряженных молекул через мембрану. С другой стороны, мембранные белки отвечают за селективную проницаемость , мембран, облегчая перемещение определенных молекул внутрь и из клеток. Мембранные белки также отвечают за специфические и избирательные взаимодействия со своей внеклеточной средой. Эти взаимодействия включают в себя адгезию клеток друг к другу, их прикрепление к поверхностям, связь между клетками (как прямую, так и через гормоны и нейроны) и т. Д.«Сахарное покрытие» внеклеточных поверхностей плазматических мембран происходит из олигосахаридов , ковалентно связанных с мембранными белками (как гликопротеинов ) или с фосфолипидами (как гликолипиды ). Углеводные компоненты гликозилированных мембранных белков сообщают об их функции. Таким образом, гликопротеины позволяют клеткам специфически взаимодействовать друг с другом с образованием тканей. Они также позволяют взаимодействовать с внеклеточными поверхностями, к которым они должны прилипать.Кроме того, они играют важную роль как часть рецепторов многих гормонов и других биомолекул, связанных с химической связью. Белковые домены, открытые для цитоплазмы, хотя и не гликозилированы, часто соединяются с компонентами цитоскелета, придавая клеткам их форму и позволяя клеткам изменять форму при необходимости. Как мы увидим, многие мембранные белки обладают важными ферментативными свойствами. Учитывая решающую роль белков и гликопротеинов в функции мембраны, неудивительно, что белки составляют в среднем 40-50% массы мембраны.В некоторых случаях белки составляют до 70% массы мембраны (представьте кристаллические мембраны в митохондриях!).
278 Свойства белков, встроенных в бислой фосфолипидов
279 различных составов мембран
C. Доказательства структуры мембраны
Асимметрия мембраны относится к различным элементам мембраны, обращенным к противоположным сторонам мембраны. Это было напрямую продемонстрировано методом сканирующего электронного микроскопа замораживания-разрушения .Методика заключается в замораживании изолированных мембран в воде с последующим измельчением льда. Когда лед трескается, покрытые оболочкой мембраны разделяются по линии наименьшего сопротивления … которая оказывается между гидрофобными жирными кислотами, противоположными хвостами внутри мембраны. Затем сканирующая электронная микроскопия выявляет особенности внутренней и внешней поверхностей мембраны. Среди характерных особенностей сканирующей микрофотографии плазматических мембран с замораживанием — ямки и противоположные насыпи , обращенные друг к другу на противоположных створках мембраны, как показано ниже.
Другие показанные здесь особенности соответствуют структуре фосфолипидной мембраны.
280 Электронная микроскопия разрушения при замораживании клеточных мембран
Цитохимия подтвердила асимметрию плазматической мембраны, показав, что только внешние поверхности плазматических мембран покрыты сахаром. Проверьте ссылку ниже для более подробного описания экспериментов.
281 EM Цитохимическая демонстрация мембранной асимметрии
Наконец, асимметрия мембран была также продемонстрирована биохимически.В одном эксперименте целые клетки обрабатывали протеолитическими ферментами с последующей экстракцией мембран и затем выделением мембранных белков. Во втором эксперименте плазматические мембраны сначала выделяли из необработанных клеток, а затем , а затем обрабатывали ферментами. В третьем эксперименте белки экстрагировали из плазматических мембран, выделенных из необработанных клеток. Электрофоретическое разделение трех белковых экстрактов по размеру продемонстрировало, что разные компоненты интегральных мембранных белков присутствовали в двух экспериментах по гидролизу, подтверждая асимметрию плазматической мембраны.Опять же, для получения более подробной информации перейдите по ссылке ниже.
282 Электрофоретическая демонстрация мембранной асимметрии
Идея о том, что мембраны — это жидкость, также была проверена. В еще одном изящном эксперименте были созданы антитела к белкам клеточных мембран мыши и человека. Мембраны были изолированы и введены третьему животному (скорее всего, кролику). Кролик видел мембраны и связанные с ними белки как чужеродные и в ответ вырабатывал специфические молекулы антимембранных антител.Антитела против каждого мембранного источника были изолированы и отдельно помечены флуоресцентными метками разного цвета, чтобы они светились другим цветом при воздействии ультрафиолетового света. После того, как мышиные и человеческие клетки были смешаны в условиях, которые вызвали их слияние, в результате были получены гибридные клетки человека и мыши. При добавлении к слитым клеткам меченые антитела связывались с белками клеточной поверхности. Через короткое время было замечено смешивание различных флуоресцентных антител под флуоресцентным микроскопом в УФ-свете.Флуоресцентные метки, казалось, сдвинулись с их первоначального положения в слитых мембранах. Ясно, что встроенные в мембрану белки не статичны, но способны перемещаться в мембране латерально, фактически плавая и диффундируя в «море фосфолипидов». Мышиные антитела в гибридной клетке сразу после слияния показаны ниже.
283 Две демонстрации текучести мембраны: Жидкая мозаика
D. Регулируемая текучесть мембраны
1.Химические факторы, влияющие на текучесть мембраны
Как вы могли догадаться, текучесть мембраны зависит от ее химического состава и физических условий, окружающих ячейку, например, от температуры окружающей среды. Факторы, влияющие на текучесть мембран, кратко описаны ниже.
Подобно тому, как нагревание раствора заставляет растворенные молекулы и частицы двигаться быстрее, фосфолипидные и белковые компоненты мембран также становятся более текучими при более высоких температурах. Если жирные кислоты фосфолипидов имеют больше ненасыщенных (C = C) углеродных связей, эти гидрофобные хвосты будут иметь больше изгибов или изгибов.Изгибы имеют тенденцию раздвигать хвосты фосфолипидов. Благодаря большему пространству между хвостами жирных кислот компоненты мембраны могут двигаться более свободно. Таким образом, более полиненасыщенных жирных кислот в мембране делают ее более текучей. С другой стороны, молекулы холестерина имеют тенденцию заполнять пространство между жирными кислотами в гидрофобной внутренней части мембраны. Это снижает боковую подвижность фосфолипидных и белковых компонентов в мембране. Уменьшая текучесть, холестерин снижает проницаемость мембран для некоторых ионов.
2. Функциональные факторы, влияющие на текучесть мембраны
Evolution адаптировала клеточные мембраны к различным и меняющимся условиям, чтобы поддерживать текучесть, необходимую для правильного функционирования клеток. Пойкилотермные , или хладнокровные организмы, от прокариот до рыб и рептилий, не регулируют температуру своего тела. Таким образом, при воздействии более низких температур пойкилотермов реагируют увеличением содержания ненасыщенных жирных кислот в их клеточных мембранах.При более высоких температурах они увеличивают содержание в мембране насыщенных жирных кислот. Таким образом, клеточные мембраны рыб, живущих под арктическим льдом, поддерживают текучесть за счет высокого уровня как мононенасыщенных, так и полиненасыщенных жирных кислот. А как насчет видов рыб, которые составляют в более теплых и холодных средах (или которые живут в климате со сменой времен года). Для этих рыб состав мембран может изменяться, чтобы приспособиться к текучести в зависимости от окружающей среды.
Теплокровные ( гомеотермные, ) млекопитающие и птицы поддерживают более или менее постоянную температуру тела.В результате их мембранный состав также относительно постоянен. Но есть парадокс! Их клеточные мембраны очень жидкие, с более высоким соотношением полиненасыщенных жиров и мононенасыщенных жиров, чем, скажем, у рептилий. Однако очевидный парадокс разрешается, когда мы понимаем, что эта большая текучесть поддерживает более высокую скорость метаболизма теплокровных видов по сравнению с пойкилотермными. Просто сравните образ жизни почти любого млекопитающего с ленивым плавающим аллигатором или змеей, греющейся в тени скалы!
284 Факторы, влияющие на текучесть мембраны
E.Изготовление искусственных мембран и эксперименты с ними
Мембранные структуры могут образовываться спонтанно. Когда фосфолипиды взаимодействуют в водной среде, они объединяются, чтобы исключить свои гидрофобные жирные хвосты из воды, образуя мицеллы. Мицеллы представляют собой сферические фосфолипидные монослойные везикулы, которые самоорганизуются, естественная агрегация гидрофобных доменов жирных кислот этих амфипатических молекул.
Ниже изображена мицелла.
Мицеллы могут в дальнейшем самособираться в сферические фосфолипидные бислои, называемые липосомами (см. Ниже).
При создании в лаборатории эти структуры ведут себя как клетки, например, образуя осадок на дне пробирки при центрифугировании. Липосомы могут быть изготовлены по индивидуальному заказу из различных видов фосфолипидов и амфипатических белков, которые становятся неотъемлемой частью липосомных мембран. Когда липосомы могут быть получены в присутствии определенных белков или других молекул, которые не могут проникать через мембрану. Захваченные молекулы не могут выбраться из этой синтетической «органеллы».Именно такие исследования позволили идентифицировать комплексы дыхательной цепи митохондрий. Возможность манипулировать содержанием липосом и составом мембран также делает их кандидатами для доставки лекарств в определенные клетки и ткани (дополнительную информацию можно найти в липосомах Google).
F. Плазменная мембрана разделена на области с различными свойствами текучести и избирательной проницаемости
Как мы вскоре увидим, текучесть не приводит к равномерной диффузии всех компонентов мембраны вокруг поверхности клеточной мембраны.Вместо этого внеклеточные связи между клетками, а также внутриклеточные связи мембраны с дифференцированными областями цитоскелета эффективно разделяют мембрану на субрегионы. Чтобы понять это, представьте себе лист эпителия, как на рисунке ниже.
Лист клеток обнажает одну поверхность с уникальными функциями внутри органа, который они выстраивают. Он открывает противоположную поверхность, одна с совершенно другой функцией, другой стороне листа.Боковые поверхности ячеек представляют собой еще один мембранный отсек, который функционирует для соединения и связи между ячейками в листе. Компоненты, то есть мембранные белки, изображенные разными символическими формами и цветами, могут оставаться жидкими внутри отсека. Конечно, это макродифференцировка клеточных мембран, позволяющая взаимодействовать клетка-клетка и клетка-окружающая среда, имеет интуитивный смысл.
Недавнее наблюдение, что клеточные мембраны еще более разделены на части, было, возможно, менее ожидаемым.Фактически мембраны делятся на микрокомпоненты. Внутри этих отсеков компоненты текучие, но редко перемещаются между отсеками. Исследования показывают, что элементы цитоскелета создают и поддерживают эти микроноразрывы. Например, интегральные мембранные белки иммобилизуются в мембранах, если они прикреплены к цитоскелетным волокнам (например, актину) в цитоплазме. Более того, когда агрегаты этих белков выстраиваются в линию из-за сходных взаимодействий, они образуют своего рода барьер , препятствуя пересечению других компонентов мембраны.По аналогии, этот механизм микро-компартментализации называется моделью «Заборы и пикеты» ; белки, прикрепленные к цитоскелету, служат пикетами. Перемещение через ограждения (т. Е. Из одного отсека мембраны в другой) происходит нечасто. Предположительно, молекуле требуется дополнительная кинетическая энергия, чтобы «перепрыгнуть» через забор между отсеками. Следовательно, этот вид движения или hop diffusion отличает его от броуновского движения, подразумеваемого исходной жидкой мозаичной моделью.
285 Мембранные домены: региональная дифференциация плазматической мембраны
.