
Функции вставочного нейрона: Какую функцию выполняют вставочные и исполнительные нейроны. Моторный нейрон. Как они работают
Какую функцию выполняют вставочные и исполнительные нейроны. Моторный нейрон. Как они работают
Нейроны очень разнообразны по форме, величине, количеству и способу отхождения от тела отростков, химическому строению (имеется в виду, в первую очередь, синтез тех или иных нейромедиаторов) и т.д. (рис.9). Тела самых крупных нейронов достигают в диаметре 100-120 мкм (гигантские пирамиды Беца в коре больших полушарий), самых мелких – 4-5 мкм (зернистые клетки коры мозжечка). Приведем ниже основные способы классификации нервных клеток:
1.
Функционально нейроны подразделяются
на чувствительные (сенсорные), вставочные
(переключательные, интернейроны) и
исполнительные (двигательные или
мотонейроны и др.). Сенсорные нейроны – это нервные клетки, воспринимающие
раздражения из внешней или внутренней
среды организма. Отростки моторных
нейронов (мотонейронов )
подходят к мышечным волокнам и образуют
на них нервно-мышечный синапс. Кроме
того, некоторые нейроны иннервируют
железы (их отростки образуют синапс с
железой).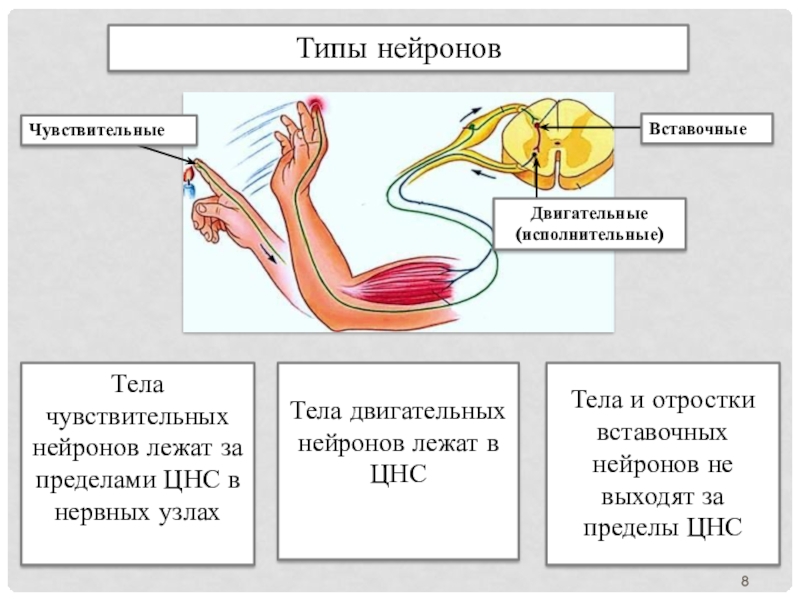
Исполнительные нейроны, управляющие сокращениями поперечно-полосатых мышечных волокон, называют двигательными (мотонейронами). Они образуют нервно-мышечные синапсы. Исполнительные нейроны, называемые вегетативными, управляют работой внутренних органов, включая гладкомышечные волокна, железистые клетки и др.
2.
По количеству отростков нейроны делятся
на униполярные, псевдоуниполярные,
биполярные и мультиполярные. Большинство
нейронов нервной системы (и почти все
нейроны в ЦНС) – это мультиполярные
нейроны (рис. 9, В-Д), они имеют один аксон и
несколько дендритов. Биполярные
нейроны (рис. 9, Б) имеют один аксон и один дендрит
и характерны для периферических отделов
анализаторных систем. Униполярных
нейронов, имеющих только один отросток,
у человека практически нет. Из тела псевдоуниполярного
нейрона (рис. 9, А) выходит один отросток, который
практически сразу делится на две ветви.
Одна из них выполняет функцию дендрита,
а другая – аксона. Такие нейроны
находятся в чувствительных спинномозговых
и черепных ганглиях. Их дендрит
морфологически (по строению) похож на
аксон: он гораздо длиннее аксона и часто
имеет миелиновую оболочку.
Униполярных
нейронов, имеющих только один отросток,
у человека практически нет. Из тела псевдоуниполярного
нейрона (рис. 9, А) выходит один отросток, который
практически сразу делится на две ветви.
Одна из них выполняет функцию дендрита,
а другая – аксона. Такие нейроны
находятся в чувствительных спинномозговых
и черепных ганглиях. Их дендрит
морфологически (по строению) похож на
аксон: он гораздо длиннее аксона и часто
имеет миелиновую оболочку.
3. По форме тела и характеру ветвления отростков выделяют звездчатые, пирамидные, веретеновидные, корзинчатые, зернистые и др. нейроны.
4. По длине аксона
нейроны делят на клетки типа Гольджи I
и типа Гольджи II (эта классификация
разработана итальянским ученым
К.Гольджи). Клетки Гольджи I имеют длинный
аксон, выходящий за пределы области, в
которой находится тело нейрона. Это,
например, пирамидные клетки коры больших
полушарий. У клеток Гольджи II короткий
и, как правило, очень разветвленный
аксон, не выходящий за пределы области,
в которой находится тело нейрона.
5. Каждый нейрон синтезирует только один основной нейромедиатор. Для того, чтобы определить нервную клетку с этой точки зрения к названию медиатора добавляют окончание «-ергический». Например, ацетилхолинергический нейрон образует ацетилхолин, глицинергический – глицин и т.д.
Нейрон
НЕЙРО́Н -а; м. [от греч. neuron — нерв] Спец. Нервная клетка со всеми отходящими от неё отростками.
нейро́н(от греч. néuron — нерв), нервная клетка, состоящая из тела и отходящих от него отростков — относительно коротких дендритов и длинного аксона; основная структурная и функциональная единица нервной системы. Нейроны проводят нервные импульсы от рецепторов в центральную нервную систему (чувствительный нейрон), от центральной нервной системы к исполнительным органам (двигательный нейрон), соединяют между собой несколько других нервных клеток (вставочные нейроны). Взаимодействуют нейроны между собой и с клетками исполнительных органов через синапсы.
НЕЙРО́Н (от греч. neuron — нерв), нервная клетка, состоящая из тела и отходящих от него отростков — относительно коротких дендритов (см. ДЕНДРИТЫ) и длинного аксона (см. АКСОН) ; основная структурная и функциональная единица нервной системы (см. схему). Нейроны проводят нервные импульсы от рецепторов в центральную нервную систему (чувствительный нейрон), от центральной нервной системы к исполнительным органам (двигательный нейрон), соединяют между собой несколько других нервных клеток (вставочные нейроны). Взаимодействуют нейроны между собой и с клетками исполнительных органов через синапсы. У коловратки (
Энциклопедический словарь . 2009 .
Синонимы :Смотреть что такое «нейрон» в других словарях:
Нейрон И9.66 советский IBM PC/XT совместимый персональный компьютер. Разработан в Киевском Научно Исследовательском Институте Радиоизмерительной Аппаратуры в середине 1980 х годов. Производством компьютера занималось ПО имени С.П. Королёва.… … Википедия
Производством компьютера занималось ПО имени С.П. Королёва.… … Википедия
нейрон — (от греч. n ё u гоп нерв) зрелая отросчатая клетка, являющаяся морфофункциональной единицей нервной ткани. В Н. выделяют тело и отростки. Существенной особенностью мембраны Н. является способность к проведению возбуждения своеобразного… … Большая психологическая энциклопедия
— (от греч. neuron нерв) нервная клетка, состоящая из тела и отходящих от него отростков относительно коротких дендритов и длинного аксона; основная структурная и функциональная единица нервной системы (см. схему). Нейроны проводят нервные импульсы … Большой Энциклопедический словарь
— (нервная клетка), основная структурная и функциональная единица НЕРВНОЙ СИСТЕМЫ, осуществляющая быструю передачу НЕРВНЫХ ИМПУЛЬСОВ между различными органами. Состоит из тела клетки, содержащей ядро, и нескольких ветвящихся пальцеобразных… … Научно-технический энциклопедический словарь
— (от греч. neuron жила, нерв), нервная клетка, нейроцит, осн. структурная и функциональная единица нервной системы, обладающая специфич. проявлениями возбудимости. Способен принимать сигналы, перерабатывать их в нервные импульсы и проводить к… … Биологический энциклопедический словарь
структурная и функциональная единица нервной системы, обладающая специфич. проявлениями возбудимости. Способен принимать сигналы, перерабатывать их в нервные импульсы и проводить к… … Биологический энциклопедический словарь
Неврон, нервная клетка, нейроцит Словарь русских синонимов. нейрон сущ., кол во синонимов: 5 клетка (126) … Словарь синонимов
— (от греческого neuron нерв), нервная клетка, состоящая из тела и отходящих от него отростков дендритов и аксона. Нейроны проводят нервные импульсы от рецепторов в центральную нервную систему, от центральной нервной системы к исполнительным… … Современная энциклопедия
НЕЙРОН, нейрона, муж. (греч. neuron волокно, нерв) (анат.). Нервная клетка. Толковый словарь Ушакова. Д.Н. Ушаков. 1935 1940 … Толковый словарь Ушакова
НЕЙРОН, а, муж. (спец.). Клетка 2, способная вырабатывать нервные импульсы и передавать их другим клеткам. Толковый словарь Ожегова. С.И. Ожегов, Н.Ю. Шведова. 1949 1992 … Толковый словарь Ожегова
Нейрон. См. нервная клетка. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
См. нервная клетка. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
нейрон — Обрабатывающий элемент нейронной сети. Тематики информационные технологии в целом … Справочник технического переводчика
Книги
- Модели волновой памяти , Кащенко С.А.. В настоящей книге рассматриваются модели нейронной среды, описываемой системой уравнений с запаздыванием. Каждый элемент среды (нейрон) является автогенератором, который в автономном режиме…
- Автономный искусственный интеллект , А. А. Жданов. Книга, посвященная моделированию нервных систем, дает возможные ответы на следующие вопросы: как должна быть устроена нервная система с логически-рациональной точки зрения? можно ли…
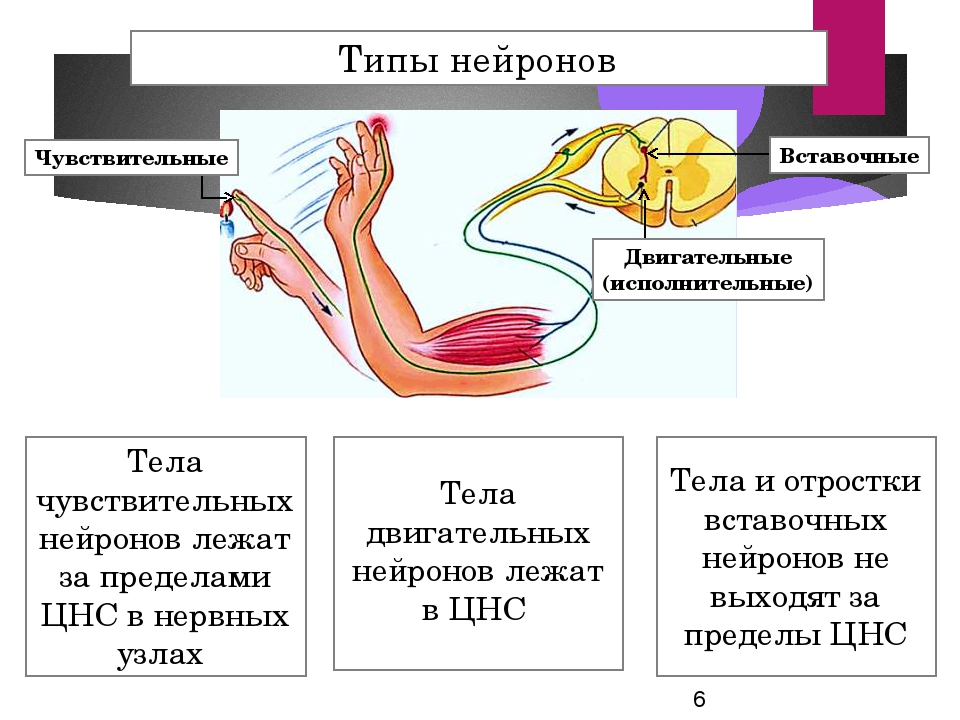
Для чего они нужны? Почему их так много? Что собой представляет чувствительный нейрон? Какую функцию выполняют вставочные и исполнительные нейроны? Давайте познакомимся поближе с этими потрясающими клетками.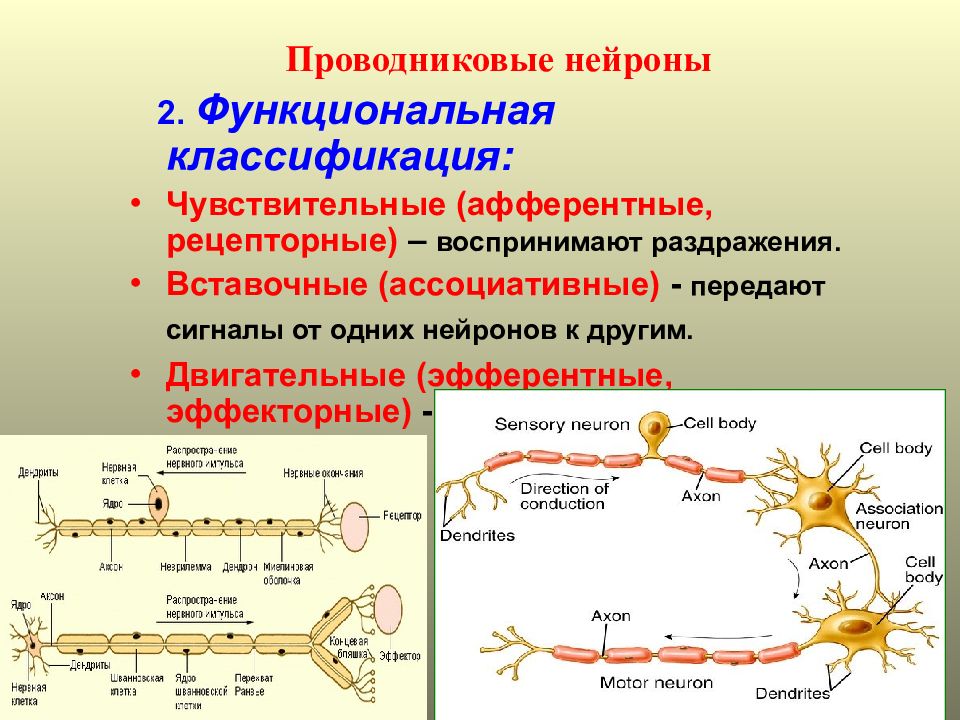
Функции
Ежесекундно через наш головной мозг проходит множество сигналов. Процесс не останавливается даже во сне. Организму нужно воспринимать окружающий мир, совершать движения, обеспечивать работу сердца, дыхательной, пищеварительной, мочеполовой системы и т.д. В организации всей этой деятельности участвуют две основные группы нейронов – чувствительные и двигательные.
Когда мы притрагиваемся к холодному или горячему и чувствуем температуру предмета – это заслуга именно чувствительных клеток. Они мгновенно передают полученную с периферии организма информацию. Так обеспечивается рефлекторная деятельность.
Нейроны формируют всю нашу ЦНС. Главные их задачи:
- получить информацию;
- передать ее по нервной системе.
Эти уникальные клетки способны мгновенно передавать электрические импульсы.
Чтобы обеспечить процесс жизнедеятельности, организм должен обрабатывать огромное количество информации, которая поступает к нему из окружающего мира, реагировать на любой признак изменения условий среды. Чтобы сделать этот процесс максимально эффективным, нейроны делятся по своим функциям на:
Чтобы сделать этот процесс максимально эффективным, нейроны делятся по своим функциям на:
- Чувствительные (афферентные) – это наши проводники в окружающий мир. Именно они воспринимают информацию извне, от органов чувств, и передают их в ЦНС. Особенность в том, что благодаря их контактной деятельности, мы чувствуем температуру, боль, давление, имеем другие чувства. Чувствительные клетки узкой специализации осуществляют передачу вкуса, запаха.
- Двигательные (моторные, эфферентные, мотонейроны). Двигательные нейроны передают информацию через электрические импульсы от ЦНС к мышечным группам, железам.
- Промежуточные (ассоциативные, интеркалярные, вставочные). Теперь подробнее разберемся, какую функцию выполняют вставочные нейроны, для чего они вообще нужны, в чем их отличие. Они располагаются между чувствительными и двигательными нейронами. Вставочные нейроны передают нервные импульсы от чувствительных волокон к двигательным. Они обеспечивают «общение» между эфферентными и афферентными нервными клетками.
 К ним нужно относиться, как к своеобразным природным «удлинителям», длинным полостям, которые помогают транслировать сигнал от сенсорного нейрона к двигательному. Без их участия это было бы невозможно сделать. В этом и заключается их функция.
К ним нужно относиться, как к своеобразным природным «удлинителям», длинным полостям, которые помогают транслировать сигнал от сенсорного нейрона к двигательному. Без их участия это было бы невозможно сделать. В этом и заключается их функция.
 К ним нужно относиться, как к своеобразным природным «удлинителям», длинным полостям, которые помогают транслировать сигнал от сенсорного нейрона к двигательному. Без их участия это было бы невозможно сделать. В этом и заключается их функция.
К ним нужно относиться, как к своеобразным природным «удлинителям», длинным полостям, которые помогают транслировать сигнал от сенсорного нейрона к двигательному. Без их участия это было бы невозможно сделать. В этом и заключается их функция.Сами рецепторы – это специально отведенные для данной функции клетки кожи, мышц, внутренних органов, суставов. Рецепторы могут начинаться еще в клетках эпидермиса, слизистой. Они умеют точно улавливать мельчайшие изменения, как снаружи организма, так и внутри него. Такие изменения могут быть физическими или химическими. Затем они молниеносно преображаются в специальные биоэлектрические импульсы и отправляются непосредственно к сенсорным нейронам. Так сигнал проходит путь от периферии к центру организма, где мозг расшифровывает его значение.
Импульсы от органа в мозг проводят все три группы нейронов – двигательные, чувствительные и промежуточные. Из этих групп клеток и состоит нервная система человека. Такое строение позволяет реагировать на сигналы из окружающего мира. Они обеспечивают рефлекторную деятельность организма.
Они обеспечивают рефлекторную деятельность организма.
Если человек перестает чувствовать вкус, запах, снижается слух, зрение, это может указывать на нарушения в ЦНС. В зависимости от того, какие органы чувств задеты, невропатолог может определить, в каком отделе мозга возникли проблемы.
1) Соматическая. Это сознательное управление мышцами скелета.
2) Вегетативная (автономная). Это неконтролируемое сознанием управление внутренними органами. Работа этой системы происходит, даже если человек находится в состоянии сна.
Сенсорные нейроны чаще всего униполярные. Это означает, что они снабжены лишь одним раздваивающимся отростком. Он выходит из тела клетки (сомы) и выполняет сразу функции и аксона, и дендрита. Аксон – это вход, а дендрит чувствительного нейрона – выход. После возбуждения чувствительных сенсорных клеток по аксону и дендриту проходит биоэлектрический сигнал.
Встречаются и биполярные нервные клетки, которые имеют соответственно два отростка. Их можно обнаружить, например, в сетчатке, структурах внутреннего уха.
Их можно обнаружить, например, в сетчатке, структурах внутреннего уха.
Тело чувствительной клетки по своей форме напоминает веретено. От тела отходит 1, а чаще 2 отростка (центральный и периферический).
Периферический по своей форме очень напоминает толстую длинную палочку. Он достигает поверхности слизистой или кожи. Такой отросток похож на дендрит нервных клеток.
Второй, противоположный отросток, отходит от противоположной части тела клетки и по форме напоминает тонкую нить, покрытую вздутиями (их называют варикозности). Это аналог нервного отростка нейрона. Данный отросток направлен в определенный отдел ЦНС и так разветвляется.
Чувствительные клетки еще называют периферическими. Их особенность в том, что они непосредственно находятся за периферической нервной системой и ЦНС, но без них работа данных систем немыслима. Например, обонятельные клетки размещены в эпителии слизистой носа.
Как они работают
Функция чувствительного нейрона состоит в приеме сигнала от специальных рецепторов, расположенных на периферии организма, определении его характеристик.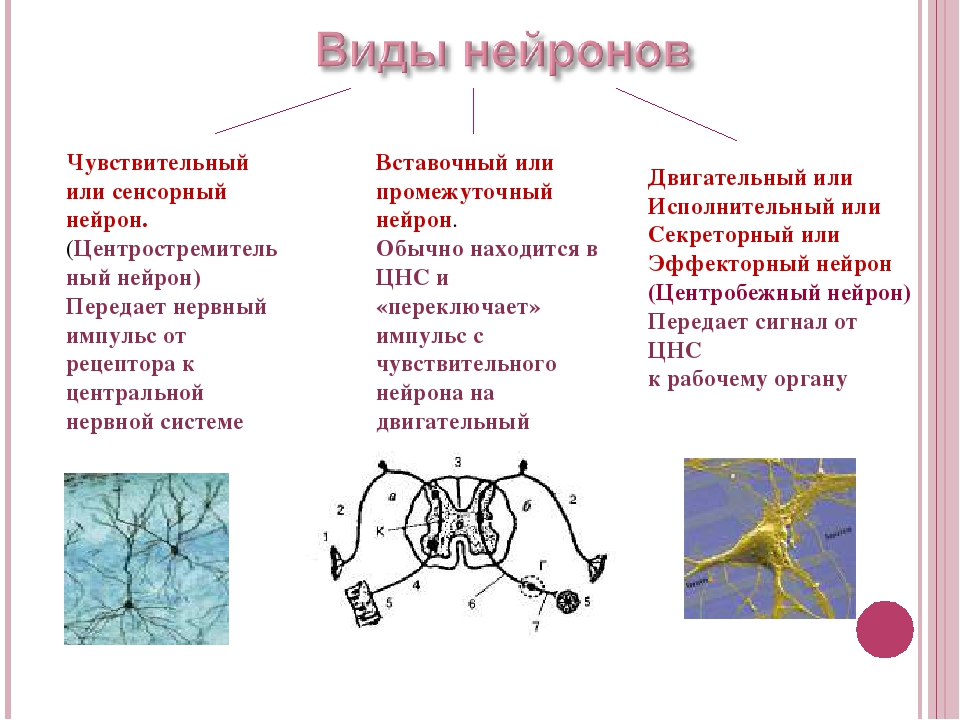 Импульсы воспринимаются периферическими отростками чувствительных нейронов, затем они передаются к их телу, а потом по центральным отросткам следуют непосредственно к ЦНС.
Импульсы воспринимаются периферическими отростками чувствительных нейронов, затем они передаются к их телу, а потом по центральным отросткам следуют непосредственно к ЦНС.
Дендриты сенсорных нейронов соединяются с различными рецепторами, а их аксоны – с остальными нейронами (вставочными). Для нервного импульса самым простым путем становится следующий – он должен пройти по трем нейронам: сенсорному, вставочному, моторному.
Самый типичный пример прохождения импульса – когда невропатолог стучит молоточком по коленному суставу. При этом моментально срабатывает простой рефлекс: коленное сухожилие после удара по нему приводит в движение мышцу, которая к нему прикреплена; чувствительные клетки от мышцы передают сигнал по чувствительным нейронам непосредственно в спинной мозг. Там сенсорные нейроны устанавливают контакт с двигательными, а те посылают импульсы обратно в мышцу, приводя ее в сокращение, нога при этом выпрямляется.
Кстати, в спинном мозге у каждого отдела (шейный, грудной, поясничный, крестцовый, копчиковый) находится сразу пара корешков: чувствительный задний, двигательный передний.
 Они образовывают единый ствол. Каждая из этих пар контролирует свою определенную часть тела и посылает центробежный сигнал, что делать дальше, как располагать конечность, туловище, что делать железе и т.д.
Они образовывают единый ствол. Каждая из этих пар контролирует свою определенную часть тела и посылает центробежный сигнал, что делать дальше, как располагать конечность, туловище, что делать железе и т.д.Чувствительные нейроны принимают участие в работе рефлекторной дуги. Она состоит из 5 элементов:
- Рецептор. Преобразует в нервный импульс раздражение.
- Импульс по нейрону следует от рецептора в ЦНС.
- Вставочный нейрон, который расположен в мозге, передает сигнал от нейрона чувствительного к исполнительному.
- По двигательному (исполнительному) нейрону основной импульс от мозга проводится к органу.
- Орган (исполнительный) – это мышца, железа и т.д. Он реагирует на полученный сигнал сокращением, выделением секрета и т.д.
Вывод
Биология человеческого организма очень продумана и совершенна. Благодаря деятельности множества чувствительных нейронов мы можем взаимодействовать с этим удивительным миром, реагировать на него. Наш организм очень восприимчивый, развитие его рецепторов и чувствительных нервных клеток достигло высочайшего уровня. Благодаря такой продуманной организации ЦНС наши органы чувств могут воспринимать и передавать мельчайшие оттенки вкуса, запаха, тактильных ощущений, звука, цвета.
Наш организм очень восприимчивый, развитие его рецепторов и чувствительных нервных клеток достигло высочайшего уровня. Благодаря такой продуманной организации ЦНС наши органы чувств могут воспринимать и передавать мельчайшие оттенки вкуса, запаха, тактильных ощущений, звука, цвета.
Нередко мы считаем, что главное в нашем сознании и деятельности организма – это кора и полушария мозга. При этом мы забываем, какие колоссальные возможности обеспечивает мозг спинной. Именно функционирование спинного мозга обеспечивает получение сигналов от всех рецепторов.
Трудно назвать предел этих возможностей. Наш организм очень пластичен. Чем больше человек развивается, тем больше возможностей предоставляется в его распоряжение. Такой простой принцип позволяет нам быстро приспособиться к изменениям окружающего мира.
Значение, строение и функционирование нервной системы
Нервная и эндокринная системы
В ходе этого урока мы ознакомимся со строением и функционированием нервной системы. А также поговорим о ее значении.
А также поговорим о ее значении.
Тема: Нервная и эндокринная система
Урок: Значение, строение и функционирование нервной системы
Нервная система — одна из главных систем, делающих наш организм не просто суммой миллиардов клеток, а уникальным единым организмом.
Нервная система регулирует и координирует работу всех систем и органов, поддерживает постоянство внутренней среды организма, и позволяет человеку успешно выживать в непростых, постоянно изменяющихся условиях.
Конечно, нервная система справляется с этим не в одиночку. Важнейшими системами, обеспечивающими целостность нашего организма, являются также эндокринная и иммунная. Тем не менее, говоря о регуляторных системах человеческого организма, в первую очередь имеют в виду нервную систему. Дело в том, что она первой успевает ответить на изменение ситуации, а ее реакция является самой быстрой и адресной. Для нервной системы характерна точная направленность нервных импульсов, большая скорость проведения информации.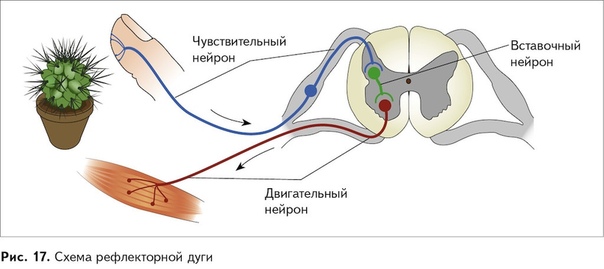 Именно работа этой системы служит основой для психической деятельности человека, его мышления, речи, сложных форм поведения.
Именно работа этой системы служит основой для психической деятельности человека, его мышления, речи, сложных форм поведения.
Основа нервной системы — нервная ткань . Нервная ткань состоит из нервных клеток — нейронов и вспомогательных нейроглиальных клеток, или клеток-спутниц. Вспомогательные клетки располагаются между нейронами и составляют межклеточное вещество нервной ткани. Выполняют опорную, защитную и питательную функции.
Рис. 1.
Нейрон — основная структурно-функциональная единица нервной ткани. Основные функции нейронов — генерация, проведение и передача нервного импульса — электрического сигнала, передающегося по нервным клеткам.
Рис. 2.
Нейрон состоит из тела и отростков. Отростки бывают короткими и длинными. Длинные отростки нервных клеток пронизывают организм и обеспечивают связь головного и спинного мозга с любым участком тела. У большинства нейронов длинный отросток имеет оболочку из особого жироподобного вещества миелина. Миелиновая оболочка способствует изоляции нервного волокна. Нервный импульс проводится по такому волокну быстрее, чем по лишенному миелина. По наличию или отсутствию оболочки все отростки делятся на миелинизированные и немиелинизированные.
Миелиновая оболочка способствует изоляции нервного волокна. Нервный импульс проводится по такому волокну быстрее, чем по лишенному миелина. По наличию или отсутствию оболочки все отростки делятся на миелинизированные и немиелинизированные.
Рис. 3.
Миелиновая оболочка имеет белый цвет, что позволило разделить вещество нервной системы на белое и серое. Тела нейронов и их короткие отростки образуют серое вещество мозга, а волокна — белое вещество.
Функциональное различие отростков нейронов связано с проведением нервного импульса.
Отросток, по которому импульс идет от тела нейрона, называется аксоном. У большинства нервных клеток аксон — это длинный отросток.
Отросток нейрона, по которому импульс идет к телу клетки, называется дендрит. Нейрон может иметь один или несколько дендритов. Дендриты, отходя от тела клетки, постепенно ветвятся под острым углом.
Рис. 4.
Передача сигнала от клетки к клетки осуществляется в особых образованиях — синапсах . Такое название им дал в 1897 г. Чарлз Шеррингтон. В них конечная веточка аксона утолщена и содержит пузырьки с раздражающим веществом — медиатором. Когда по аксону нервные импульсы дойдут до синапса, пузырьки лопаются и жидкость, содержащая медиаторы, попадает в синаптическую щель. В зависимости от ее состава клетка, регулируемая нейроном, может включиться в работу, то есть возбудиться, или выйти из работы (затормозиться).
Такое название им дал в 1897 г. Чарлз Шеррингтон. В них конечная веточка аксона утолщена и содержит пузырьки с раздражающим веществом — медиатором. Когда по аксону нервные импульсы дойдут до синапса, пузырьки лопаются и жидкость, содержащая медиаторы, попадает в синаптическую щель. В зависимости от ее состава клетка, регулируемая нейроном, может включиться в работу, то есть возбудиться, или выйти из работы (затормозиться).
Нейроны различаются по своим функциям и подразделяются на чувствительные, вставочные и двигательные.
Чувствительные нейроны — это нервные клетки, воспринимающие раздражения из внешней или внутренней среды организма.
Рис. 6.
Двигательные (исполнительные) нейроны — нейроны, иннервирующие мышечные волокна и железы.
Рис. 7.
Вставочные нейроны обеспечивают связь между чувствительными и двигательными нейронами.
Между чувствительным и двигательным нейроном может быть очень большое количество вставочных нейронов. Они собирают, анализируют информацию, полученную от чувствительных нейронов, и принимают решение о том, каким образом отреагировать на изменившиеся условия.
Они собирают, анализируют информацию, полученную от чувствительных нейронов, и принимают решение о том, каким образом отреагировать на изменившиеся условия.
Нервную систему (по месту расположения ) подразделяют на центральную и периферическую. К центральной нервной системе относят спинной и головной мозг, к периферической — нервы, нервные узлы и нервные окончания.
Рис. 8.
Нервы — пучки длинных отростков, покрытые общей оболочкой, выходящие за пределы головного и спинного мозга.
Если информация по нерву идет от рецепторов в головной или спинной мозг, то такие нервы называют чувствительными, центростремительными или афферентными. Эти нервы состоят из дендритов чувствительных нейронов.
Если информация по нерву идет из центральной нервной системы к исполнительным органам (мышцам или железам), то нерв называется двигательным или эфферентным. Двигательные нервы образованы аксонами двигательных нейронов.
В смешанных нервах проходят как чувствительные, так и двигательные волокна.
Нервные узлы — это скопления тел нейронов вне ЦНС.
Нервные окончания — разветвления отростков нейронов, служат для приема или передачи сигналов.
По функциям нервная система подразделяется на соматическую и вегетативную (автономную).
Рис. 9.
Соматическая нервная система (от греческого «сома» — «тело») регулирует работу скелетных мышц. Благодаря ей организм через органы чувств поддерживает связь с внешней средой. С ее помощью мы можем произвольно (по собственному желанию) управлять деятельностью скелетной мускулатуры.
Деятельностью внутренних органов, реакциями обмена веществ, поддержанием постоянства внутренней среды организма человека управляет автономная или вегетативная нервная система . Ее название происходит от греческого слова «автономия» — самоуправление. Работа этой системы не подчиняется воле человека. Нельзя, например, по желанию ускорить процесс пищеварения или сузить кровеносные сосуды.
Автономная система представлена двумя отделами — симпатическим и парасимпатическим. Симпатический отдел (система сложных ситуаций) включается во время интенсивной работы, требующей затраты энергии (что-то услышал неожиданное — расширяются зрачки, возрастает частота сокращений сердца, замедляется деятельность пищеварительной системы, учащается дыхание). Парасимпатический отдел можно назвать системой отбоя. Она возвращает организм в состояние покоя, создает условия для отдыха и восстановления организма.
Основной принцип работы нервной системы — рефлекторный. Любая ответная реакция организма на раздражитель, осуществляемая и контролируемая нервной системой, называется рефлексом. Основу рефлекторной реакции составляет рефлекторная дуга. В состав рефлекторной дуги входит рецептор, воспринимающий раздражение. По аксону чувствительного нейрона возбуждение попадает в центральную нервную систему и может распространиться непосредственно на двигательный нейрон или сначала на вставочные нейроны, а уже через них на эфферентный нейрон. По аксону эфферентного нейрона возбуждение достигает исполнительного органа, чаще всего мышцы. В результате возбуждения деятельность этого органа изменяется, например, мышца сокращается.
По аксону эфферентного нейрона возбуждение достигает исполнительного органа, чаще всего мышцы. В результате возбуждения деятельность этого органа изменяется, например, мышца сокращается.
Рис. 10.
Рефлексы подразделяются на соматические, заканчивающиеся сокращением скелетных мышц, и вегетативные, в результате которых меняется работа внутренних органов. Примером наиболее простого соматического рефлекса может служить дуга коленного рефлекса, состоящая всего из двух нейронов — чувствительного и двигательного.
1. Колесов Д.В., Маш Р.Д., Беляев И.Н. Биология 8 М.:Дрофа
2. Пасечник В.В., Каменский А.А., Швецов Г.Г. / Под ред. Пасечника В.В. Биология 8 М.:Дрофа.
3. Драгомилов А.Г., Маш Р.Д. Биология 8 М.: ВЕНТАНА-ГРАФ
1. Колесов Д.В., Маш Р.Д., Беляев И.Н. Биология 8 М.: Дрофа — с. 39, задания и вопрос 6,7,8,9.
2. Какие выделяют отделы нервной системы по месту расположения?
3. Опишите строение нейрона.
4. Подготовьте реферат о заболеваниях нервной системы.
функции и роль в формировании нейронных сетей. Типы нейронов
Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов.
Обзор
Сложность и многообразие функций нервной системы определяются взаимодействием между нейронами, которое, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
Строение
Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и . Нейрон имеет развитый и сложный цитоскелет, проникающий в его отростки.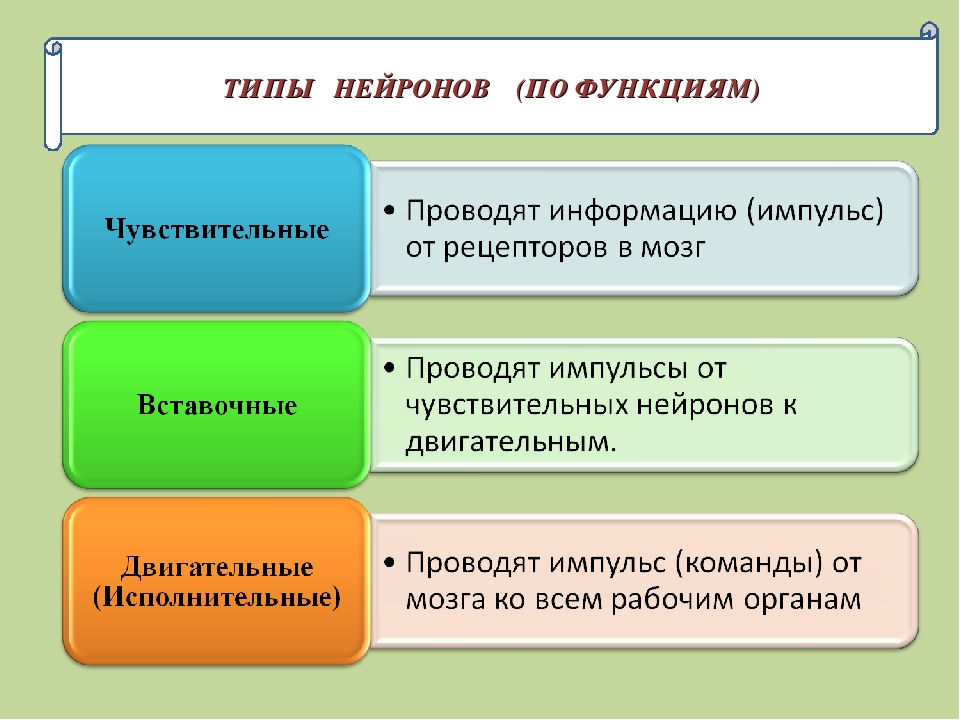 Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20-30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в . В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона.
Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20-30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в . В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Аксон — обычно длинный отросток, приспособленный для проведения от тела нейрона. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторнойклеткой. Служит для передачи между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсывызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Одни синапсывызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
Классификация
Структурная классификация
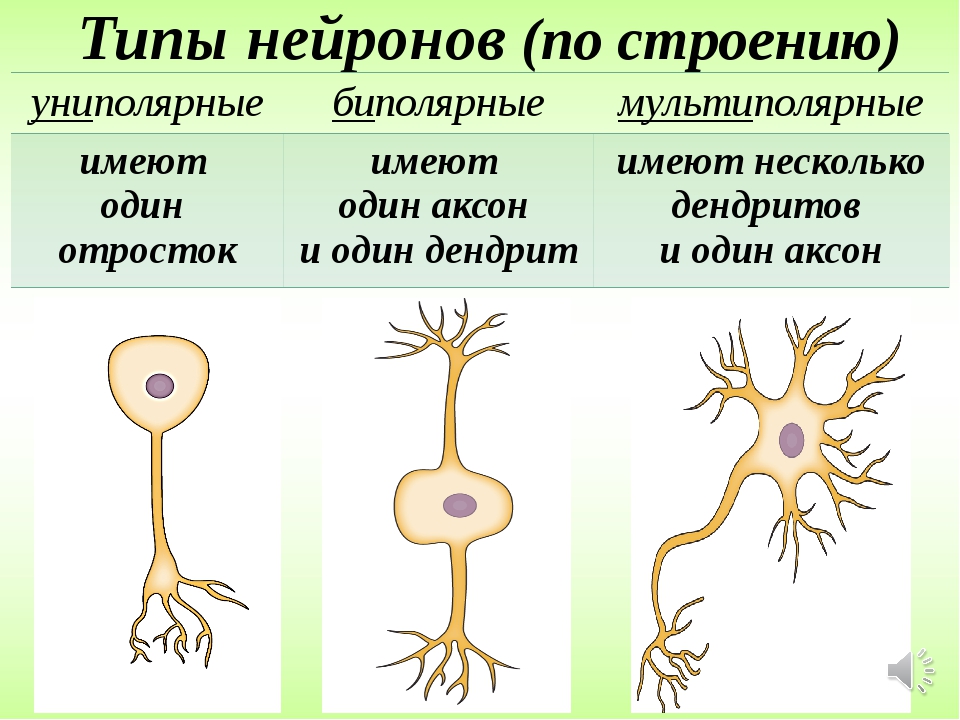
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в .
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в .
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация
Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона;
- количество и характер ветвления отростков;
- длину нейрона и наличие специализированных оболочек.

По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120-150 мкм у гигантских пирамидных нейронов. Длина нейрона у человека составляет от 150 мкм до 120 см.
По количеству отростков выделяют следующие морфологические типы нейронов:
- униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в ;
- псевдоуниполярные клетки, сгруппированные вблизи в межпозвоночных ганглиях;
- биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
- мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Развитие и рост нейрона
Нейрон развивается из небольшой клетки-предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
(Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Наш с вами спинной мозг — это наиболее древнее в эволюционном плане образование нервной системы. Появляясь впервые у ланцетника, в процессе эволюции спинной мозг с его эфферентными (двигательными) и афферентными (чувствительными) нейронами совершенствовался. Но при этом сохранял свои главные функции — проводящую и регуляторную. Именно благодаря чувствительным нейронам мы отдергиваем руку от горячей кастрюли еще до появления боли. О структуре этого органа центральной нервной системы и принципах его работы идет речь в данной статье.
Такой ранимый, но очень важный
Этот мягкий орган прячется внутри позвоночного столба. Спинной мозг человека весит всего до 40 граммов, имеет длину до 45 сантиметров, а толщина его сравнима с мизинцем — всего 8 миллиметров в диаметре. И, тем не менее, это управляющий центр сложной сети которая раскинулась по всему нашему телу. Без него не сможет выполнять свои аппарат и все жизненные органы нашего организма. Кроме позвонков спинной мозг защищают его оболочки. Наружная оболочка твердая, образована плотной соединительной тканью. В этой оболочке расположены кровеносные сосуды и нервы. А, кроме того, именно в ней наблюдается наивысшая концентрация болевых рецепторов в организме человека. А вот в самом мозге таких рецепторов нет. Вторая оболочка — паутинная, заполнена ликвором (спинномозговой жидкостью). Последняя оболочка — мягкая — плотно прилегает к мозгу, пронизана кровеносными и лимфатическими сосудами.
Без него не сможет выполнять свои аппарат и все жизненные органы нашего организма. Кроме позвонков спинной мозг защищают его оболочки. Наружная оболочка твердая, образована плотной соединительной тканью. В этой оболочке расположены кровеносные сосуды и нервы. А, кроме того, именно в ней наблюдается наивысшая концентрация болевых рецепторов в организме человека. А вот в самом мозге таких рецепторов нет. Вторая оболочка — паутинная, заполнена ликвором (спинномозговой жидкостью). Последняя оболочка — мягкая — плотно прилегает к мозгу, пронизана кровеносными и лимфатическими сосудами.
Несколько слов о нейронах
Структурной единицей нервной ткани являются нейроны. Совершенно особые клетки, главная функция которых образование и передача нервного импульса. Каждый нейрон имеет множество коротких отростков — дендритов, воспринимающих раздражение, и один длинный — аксон, который проводит нервный импульс только в одном направлении. В зависимости от задачи и бывают чувствительные и двигательные.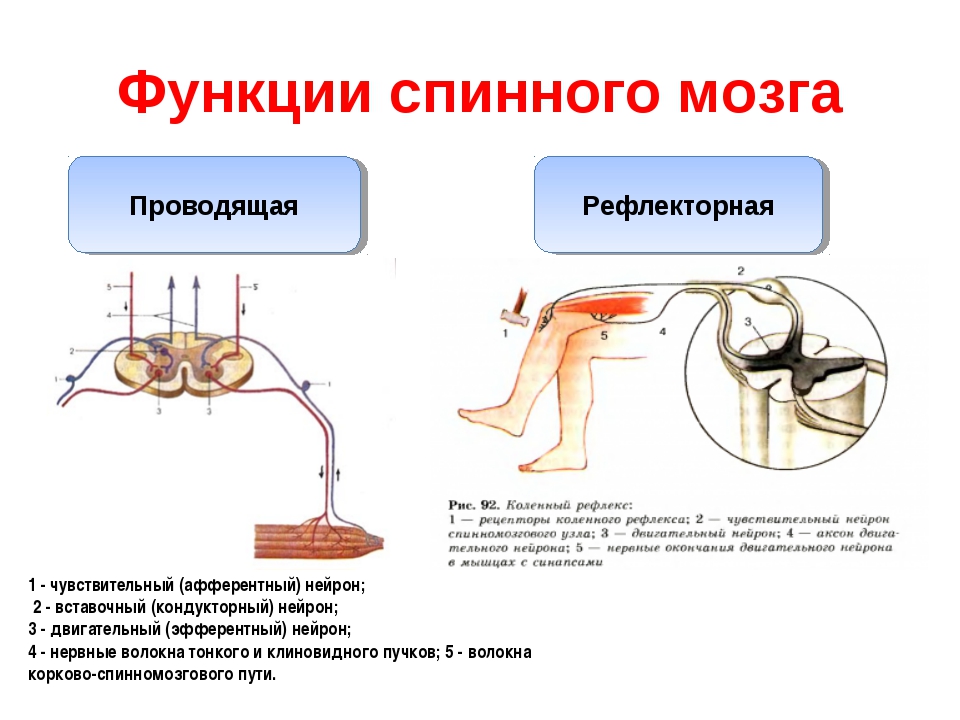 Нейроны промежуточные или вставочные — это своеобразные «удлинители», которые передают импульс между другими нейронами.
Нейроны промежуточные или вставочные — это своеобразные «удлинители», которые передают импульс между другими нейронами.
Строение спинного мозга
Начинается спинной мозг с затылочного отверстия черепа, заканчивается в поясничных позвонках. Он состоит из 31-33 сегментов, которые не отделены друг от друга: С1-С8 — шейные, Th2- Th22 — грудные, L1-L5 — поясничные, S1-S5 — крестцовые, Co1-Co3 — копчиковые. Ниже в канале позвоночника расположены продолжения нервов, собранные в пучок и именуемые конским хвостом (видимо за внешнее сходство), которые иннервируют нижние конечности и органы таза. Каждый сегмент имеет две пары корешков, которые соединяются в 31 пару спинномозговых нервов. Два задних (дорсальных) корешка образованы аксонами чувствительных нейронов и имеют утолщение — где находятся тела этих нейронов. Два передних (вентральных) корешка образованы аксонами двигательных нейронов.
Такие разные и важные
В спинном мозге человека находится порядка 13 миллионов нервных клеток. Функционально они делятся на 4 группы:
Функционально они делятся на 4 группы:
- Двигательные — образуют передние рога и передние корешки.
- Интернейроны — образуют задние рога. Здесь находятся чувствительные нейроны, в которых возникает на различные раздражения (болевые, тактильные, вибрационные, температурные).
- Симпатические и парасимпатические нейроны — находятся в боковых рогах и образуют передние корешки.
- Ассоциативные — это уже клетки головного мозга, которые устанавливают связь между сегментами спинного мозга.
Серая бабочка в окружении белого
В центре спинного мозга расположено серое вещество, образующее передние, задние и боковые рога. Это тела нейронов. В спинальных ганглиях расположены чувствительные нейроны, длинный отросток которых находится на периферии и заканчивается рецептором, а короткий — в нейронах задних рогов. Передние рога образованы аксоны которых идут к скелетным мышцам. В боковых рогах расположены нейроны вегетативной системы. Серое вещество окружено белым — это нервные волокна, образованные аксонами восходящих и нисходящих проводных путей. Первые чувствительные нейроны расположены в следующих сегментах: шейном С7, грудных Th2- Th22, поясничных L1-L3, крестцовых S2-S4. При этом спинномозговой нерв соединяет в один ствол задние (чувствительные) и передние (двигательные) корешки. При этом каждая пара спинномозговых нервов контролирует определенные части тела.
Первые чувствительные нейроны расположены в следующих сегментах: шейном С7, грудных Th2- Th22, поясничных L1-L3, крестцовых S2-S4. При этом спинномозговой нерв соединяет в один ствол задние (чувствительные) и передние (двигательные) корешки. При этом каждая пара спинномозговых нервов контролирует определенные части тела.
Как это работает
Разветвленные дендриты чувствительных нейронов спинальных центров вегетативной нервной системы заканчиваются рецепторами, которые представляют собой биологические структуры, в которых формируется нервный импульс при контакте с конкретным раздражителем. Рецепторы обеспечивают вегетовисцеральную чувствительность — воспринимают раздражение от таких частей нашего тела как кровеносные сосуды и сердце, желудочно-кишечный тракт, печень и поджелудочная железа, почки и другие. По дендриту импульс передается к телу нейрона. Далее по аксонам афферентных (чувствительных) нейронов поступает в спинной мозг, где образуют синоптические соединения с дендритами эфферентных (двигательных) нейронов.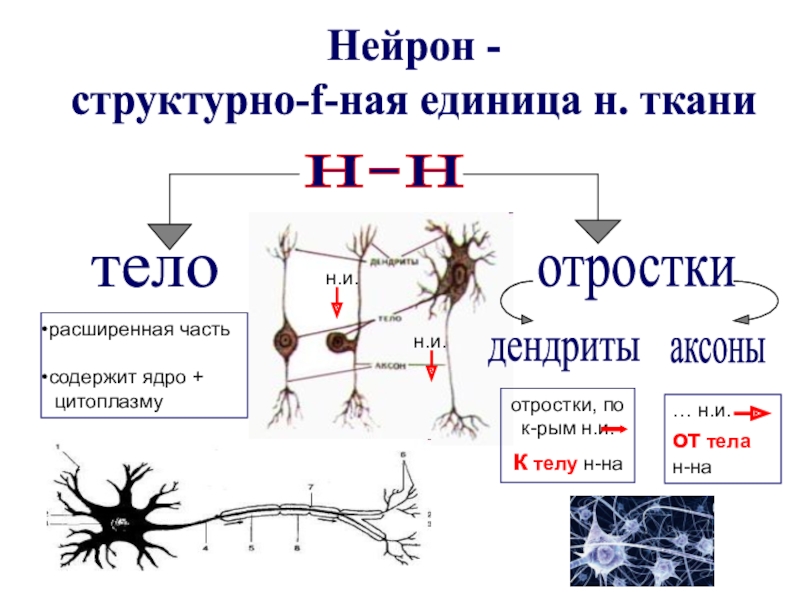 Именно благодаря такому прямому контакту мы отдергиваем руку от горячей кастрюли или утюга еще до того, как наш главный командир — головной мозг — проанализирует возникшие болевые ощущения.
Именно благодаря такому прямому контакту мы отдергиваем руку от горячей кастрюли или утюга еще до того, как наш главный командир — головной мозг — проанализирует возникшие болевые ощущения.
Подводим итог
Все наши автоматические и рефлекторные действия происходят под надзором именно спинного мозга. Исключение составляют лишь те, которые контролирует сам головной мозг. Например, воспринимая увиденное с использованием глазного нерва, который идет прямо в головной мозг, мы меняем угол зрения при помощи мышц глазного яблока, которые уже контролируются спинным мозгом. Плачем мы, кстати, тоже по приказу спинного мозга — слезными железами «командует» именно он. Сознательные наши действия начинаются в головном мозге, но как только они становятся автоматическим, их контроль переходит к спинному мозгу. Можно сказать, что нашему пытливому головному мозгу нравится учиться. А когда он уже научился, ему становится скучно и он отдает «бразды правления» своему более древнему в эволюционном плане собрату.
1) центральная — спинной и
2) периферическая — нервы и нервные узлы.
- Нервы — это пучки нервных волокон, окруженные соединительнотканной оболочкой.
- Нервные узлы — это скопления тел нейронов за пределами ЦНС, например, солнечное сплетение.
Нервная система по функциям делится на 2 части
1) соматическая — управляет скелетными мышцами, подчиняется сознанию.
2) вегетативная (автономная) — управляет внутренними органами, не подчиняется сознанию. Состоит из двух частей:
- симпатическая : управляет органами во время стресса и физической нагрузки
- повышает пульс, давление и концентрацию глюкозы в крови
- активизирует работу нервной системы и органов чувств
- расширяет бронхи и зрачок
- тормозит работу пищеварительной системы.
- парасимпатическая система работает в состоянии покоя, приводит работу органов в норму (функции противоположные).

Рефлекторная дуга
Это путь, по которому нервный импульс проходит при осуществлении . Состоит из 5 частей
1) Рецептор — чувствительное образование, способное реагировать на определенный вид раздражителя; преобразует раздражение в нервный импульс.
2) По чувствительному нейрону нервный импульс идет от рецептора в центральную нервную систему (спинной или головной мозг).
3) Вставочный нейрон находится в мозге, передает сигнал с чувствительного нейрона на исполнительный.
4) По исполнительному (двигательному) нейрону нервный импульс идет от мозга к рабочему органу.
5) Рабочий (исполнительный) орган — мышца (сокращается), железа (выделяет секрет) и т.п.
Анализатор
Это система нейронов, воспринимающих раздражение, проводящих нервные импульсы и обеспечивающих переработку информации. Состоит из 3 отделов:
1) периферического – это рецепторы, например, колбочки и палочки в сетчатке глаза
2) проводникового – это нервы и проводящие пути мозга
3) центрального , расположенного в коре – здесь происходит окончательный анализ информации.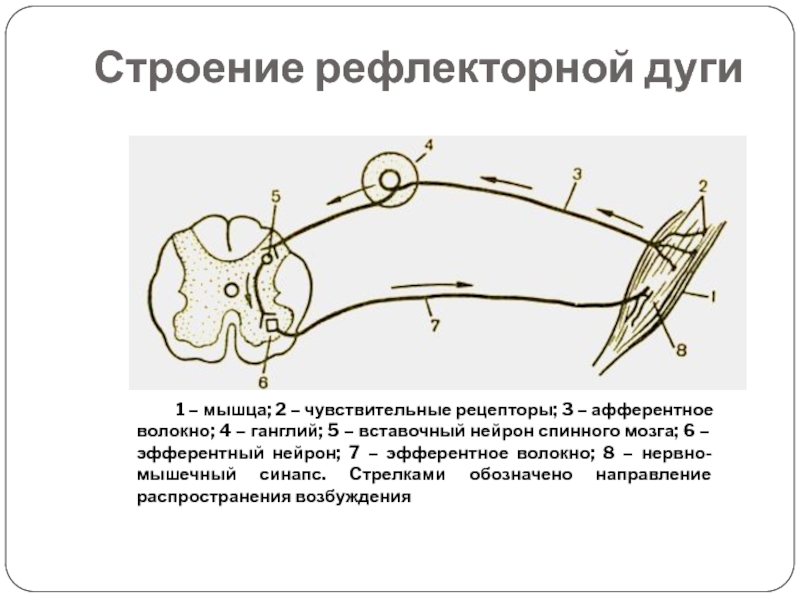
Выберите один, наиболее правильный вариант. Отдел слухового анализатора, передающий нервные импульсы в головной мозг человека, образован
1) слуховыми нервами
2) рецепторами, расположенными в улитке
3) барабанной перепонкой
4) слуховыми косточками
Ответ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Какие примеры иллюстрируют возбуждение симпатической нервной системы?
1) усиление сердечных сокращений
2) усиление перистальтики кишечника
3) понижение артериального давления
4) расширение зрачков глаз
5) увеличение сахара в крови
6) сужение бронхов и бронхиол
Ответ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Какое влияние оказывает парасимпатическая нервная система на организм человека?
1) увеличивает частоту сокращений сердца
2) активизирует слюнообразование
3) стимулирует выработку адреналина
4) усиливает образование желчи
5) увеличивает перистальтику кишечника
6) осуществляет мобилизацию функций органов при стрессе
Ответ
Выберите один, наиболее правильный вариант. Нервные импульсы от рецепторов в центральную нервную систему проводят
Нервные импульсы от рецепторов в центральную нервную систему проводят
1) чувствительные нейроны
2) двигательные нейроны
3) чувствительные и двигательные нейроны
4) вставочные и двигательные нейроны
Ответ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Рецепторы – это нервные окончания в организме человека, которые
1) воспринимают информацию из внешней среды
2) воспринимают импульсы из внутренней среды
3) воспринимают возбуждение, передающееся к ним по двигательным нейронам
4) располагаются в исполнительном органе
5) преобразуют воспринимаемые раздражения в нервные импульсы
6) реализуют ответную реакцию организма на раздражение из внешней и внутренней среды
Ответ
Выберите один, наиболее правильный вариант. Периферическая часть зрительного анализатора
1) зрительный нерв
2) зрительные рецепторы
3) зрачок и хрусталик
4) зрительная зона коры
Ответ
Выберите один, наиболее правильный вариант. Рефлексы, которые не могут быть усилены или заторможены по воле человека, осуществляются через нервную систему
Рефлексы, которые не могут быть усилены или заторможены по воле человека, осуществляются через нервную систему
1) центральную
2) вегетативную
3) соматическую
4) периферическую
Ответ
1. Установите соответствие между особенностью регуляции и отделом нервной системы, который ее осуществляет: 1) соматический, 2) вегетативный
А) регулирует работу скелетных мышц
Б) регулирует процессы обмена веществ
В) обеспечивает произвольные движения
Г) осуществляется автономно независимо от желания человека
Д) контролирует деятельность гладкой мускулатуры
Ответ
2. Установите соответствие между функцией периферической нервной системы человека и отделом, который эту функцию выполняет: 1) соматическая, 2) вегетативная
А) направляет команды к скелетным мышцам
Б) иннервирует гладкую мускулатуру внутренних органов
В) обеспечивает перемещение тела в пространстве
Г) регулирует работу сердца
Д) усиливает работу пищеварительных желёз
Ответ
3. Установите соответствие между характеристикой и отделом нервной системы человека: 1) соматическая, 2) вегетативная. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
Установите соответствие между характеристикой и отделом нервной системы человека: 1) соматическая, 2) вегетативная. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) направляет команды к скелетным мышцам
Б) изменяет деятельность различных желёз
В) образует только трёхнейронную рефлекторную дугу
Г) изменяет частоту сердечных сокращений
Д) обусловливает произвольные движения тела
Е) регулирует сокращение гладкой мускулатуры
Ответ
4. Установите соответствие между свойствами нервной системы и ее типами: 1) соматическая, 2) вегетативная. Запишите цифры 1 и 2 в правильном порядке.
А) иннервирует кожу и скелетные мышцы
Б) иннервирует все внутренние органы
В) действия неподвластны сознанию (автономны)
Г) действия подконтрольны сознанию (произвольны)
Д) способствует поддержанию связи организма с внешней средой
Е) регулирует обменные процессы, рост организма
Ответ
5. Установите соответствие между типами нервной системы и их характеристиками: 1) вегетативная, 2) соматическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) регулирует работу внутренних органов
Б) регулирует работу скелетных мышц
В) рефлексы осуществляются быстро и подчиняются сознанию человека
Г) рефлексы медленные и не подчиняются сознанию человека
Д) высший орган этой системы гипоталамус
Е) высший центр этой системы — кора больших полушарий
Ответ
6н. Установите соответствие между характеристикой и отделом нервной системы человека, к которому её относят: 1) соматическая, 2) вегетативная. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) регулирует диаметр кровеносных сосудов
Б) имеет двигательный путь рефлекторной дуги, состоящий из двух нейронов
В) обеспечивает разнообразные движения тела
Г) работает произвольно
Д) поддерживает деятельность внутренних органов
Ответ
Установите соответствие между органами и видами нервной системы, которые контролируют их деятельность: 1) соматическая, 2) вегетативная. Запишите цифры 1 и 2 в правильном порядке.
Запишите цифры 1 и 2 в правильном порядке.
А) мочевой пузырь
Б) печень
В) бицепсы
Г) межреберные мышцы
Д) кишечник
Е) глазодвигательные мышцы
Ответ
Выберите три варианта. Слуховой анализатор включает в себя
1) слуховые косточки
2) рецепторные клетки
3) слуховую трубу
4) чувствительный нерв
5) полукружные каналы
6) кору височной доли
Ответ
Выберите один, наиболее правильный вариант. Нервные импульсы передаются в мозг по нейронам
1) двигательным
2) вставочным
3) чувствительным
4) исполнительным
Ответ
Выберите три последствия раздражения симпатического отдела центральной нервной системы:
1) учащение и усиление сокращений сердца
2) замедление и ослабление сокращений сердца
3) замедление процессов образования желудочного сока
4) усиление интенсивности деятельности желёз желудка
5) ослабление волнообразных сокращений стенок кишечника
6) усиление волнообразных сокращений стенок кишечника
Ответ
1. Установите соответствие между функцией органов и отделом вегетативной нервной системы, который ее осуществляет: 1) симпатический, 2) парасимпатический
Установите соответствие между функцией органов и отделом вегетативной нервной системы, который ее осуществляет: 1) симпатический, 2) парасимпатический
А) усиление выделения пищеварительных соков
Б) замедление частоты сердечных сокращений
В) усиление вентиляции легких
Г) расширение зрачка
Д) усиление волнообразных движений кишечника
Ответ
2. Установите соответствие между функцией органов и отделом вегетативной нервной системы, который ее осуществляет: 1) симпатический, 2) парасимпатический
А) повышает частоту сердечных сокращений
Б) уменьшает частоту дыхания
В) стимулирует секрецию пищеварительных соков
Г) стимулирует выброс адреналина в кровь
Д) усиливает вентиляцию лёгких
Ответ
3. Установите соответствие между функцией вегетативной нервной системы и её отделом: 1) симпатический, 2) парасимпатический
А) повышает кровяное давление
Б) усиливает отделение пищеварительных соков
В) понижает частоту сердечных сокращений
Г) ослабляет перистальтику кишечника
Д) усиливает кровоток в мышцах
Ответ
4. Установите соответствие между функциями и отделами вегетативной нервной системы: 1) симпатическая, 2) парасимпатическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
Установите соответствие между функциями и отделами вегетативной нервной системы: 1) симпатическая, 2) парасимпатическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) расширяет просветы артерий
Б) учащает сердцебиение
В) усиливает перистальтику кишечника и стимулирует работу пищеварительных желез
Г) сужает бронхи и бронхиолы, уменьшает вентиляцию легких
Д) расширяет зрачки
Ответ
Выберите один, наиболее правильный вариант. Чем образованы нервы?
1) скоплением нервных клеток в головном мозге
2) скоплениями нервных клеток вне центральной нервной системы
3) нервными волокнами с соединительнотканной оболочкой
4) белым веществом, расположенным в центральной нервной системе
Ответ
Выберите три анатомические структуры, являющиеся начальным звеном анализаторов человека
1) веки с ресницами
2) палочки и колбочки сетчатки
3) ушная раковина
4) клетки вестибулярного аппарата
5) хрусталик глаза
6) вкусовые сосочки языка
Ответ
Выберите один, наиболее правильный вариант. Систему нейронов, воспринимающих раздражения, проводящих нервные импульсы и обеспечивающих переработку информации, называют
Систему нейронов, воспринимающих раздражения, проводящих нервные импульсы и обеспечивающих переработку информации, называют
1) нервным волокном
3) нервом
4) анализатором
Ответ
Выберите один, наиболее правильный вариант. Как называют систему нейронов, воспринимающих раздражения, проводящих нервные импульсы и обеспечивающих переработку информации
1) нервным волокном
2) центральной нервной системой
3) нервом
4) анализатором
Ответ
Выберите три варианта. Зрительный анализатор включает
1) белочную оболочку глаза
2) рецепторы сетчатки
3) стекловидное тело
4) чувствительный нерв
5) кору затылочной доли
6) хрусталик
Ответ
Выберите один, наиболее правильный вариант. Периферическую часть слухового анализатора человека образуют
1) слуховой проход и барабанная перепонка
2) косточки среднего уха
3) слуховые нервы
4) чувствительные клетки улитки
Ответ
При возбуждении симпатической нервной системы в отличие от возбуждения парасимпатической нервной системы
1) расширяются артерии
2) повышается артериальное давление
3) усиливается перистальтика кишечника
4) сужается зрачок
5) увеличивается количество сахара в крови
6) учащаются сокращения сердца
Ответ
1. Установите последовательность частей рефлекторной дуги при прохождении по ней нервного импульса. Запишите соответствующую последовательность цифр.
Установите последовательность частей рефлекторной дуги при прохождении по ней нервного импульса. Запишите соответствующую последовательность цифр.
1) чувствительный нейрон
2) рабочий орган
3) вставочный нейрон
4) отдел коры больших полушарий
5) рецептор
6) двигательный нейрон
Ответ
2. Установите последовательность звеньев рефлекторной дуги рефлекса потоотделения. Запишите соответствующую последовательность цифр.
1) возникновение в рецепторах нервных импульсов
2) потоотделение
3) возбуждение двигательных нейронов
4) раздражение рецепторов кожи, воспринимающих тепло
5) передача нервных импульсов к потовым железам
6) передача нервных импульсов по чувствительным нейронам в ЦНС
Ответ
3. Установите последовательность проведения нервного импульса в рефлекторной дуге, обеспечивающей один из механизмов терморегуляции в организме человека. Запишите соответствующую последовательность цифр.
1) передача нервного импульса по чувствительному нейрону в центральную нервную систему
2) передача нервного импульса на двигательные нейроны
3) возбуждение терморецепторов кожи при понижении температуры
4) передача нервного импульса на вставочные нейроны
5) уменьшение просвета кровеносных сосудов кожи
Ответ
Выберите три варианта.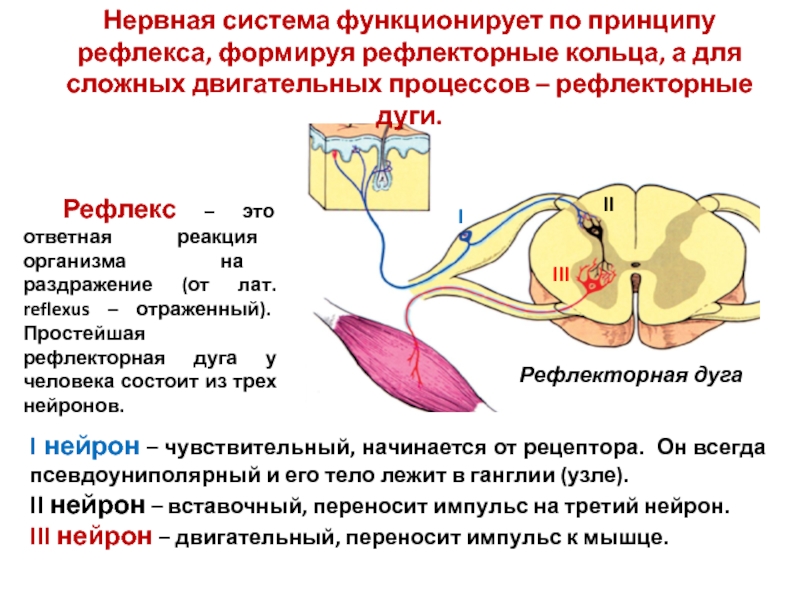 В нервной системе человека вставочные нейроны передают нервные импульсы
В нервной системе человека вставочные нейроны передают нервные импульсы
1) с двигательного нейрона в головной мозг
2) от рабочего органа в спинной мозг
3) от спинного мозга в головной мозг
4) от чувствительных нейронов к рабочим органам
5) от чувствительных нейронов к двигательным нейронам
6) из головного мозга к двигательным нейронам
Ответ
Расположите в правильном порядке элементы рефлекторной дуги коленного рефлекса человека. Запишите в ответе цифры в порядке, соответствующем буквам.
1) Двигательный нейрон
2) Чувствительный нейрон
3) Спиной мозг
4) Рецепторы сухожилия
5) Четырёхглавая мышца бедра
Ответ
Выберите три функции симпатической нервной системы. Запишите цифры, под которыми они указаны.
1) усиливает вентиляцию лёгких
2) уменьшает частоту сердечных сокращений
3) снижает кровяное давление
4) угнетает секрецию пищеварительных соков
5) усиливает перистальтику кишечника
6) расширяет зрачки
Ответ
Выберите один, наиболее правильный вариант. Чувствительные нейроны в трехнейронной рефлекторной дуге соединены с
Чувствительные нейроны в трехнейронной рефлекторной дуге соединены с
1) отростками вставочных нейронов
2) телами вставочных нейронов
3) двигательными нейронами
4) исполнительными нейронами
Ответ
Установите соответствие между функциями и типами нейронов: 1) чувствительные, 2) вставочные, 3) двигательные. Запишите цифры 1, 2, 3 в порядке, соответствующем буквам.
А) передача нервных импульсов от органов чувств в мозг
Б) передача нервных импульсов от внутренних органов в мозг
В) передача нервных импульсов к мышцам
Г) передача нервных импульсов к железам
Д) передача нервных импульсов от одного нейрона к другому
Ответ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Какими органами управляет вегетативная нервная система?
1) органы пищеварительного тракта
2) половые железы
3) мышцы конечностей
4) сердце и кровеносные сосуды
5) межреберные мышцы
6) жевательные мышцы
Ответ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. К центральной нервной системе относят
К центральной нервной системе относят
1) чувствительные нервы
2) спинной мозг
3) двигательные нервы
4) мозжечок
5) мост
6) нервные узлы
Ответ
Проанализируйте таблицу «Нейроны». Для каждой ячейки, обозначенной буквой, выберите соответствующий термин из приведенного списка. © Д.В.Поздняков, 2009-2019
Нейрон является специфической, электрически возбудимой клеткой в нервной системе человека и обладает уникальными особенностями. Его функции заключаются в обработке, хранении и передаче информации. Нейроны характеризуются сложным строением и узкой специализацией. Они также делятся на три вида. В этой статье подробно описывается вставочный нейрон и его роль в действии центральной нервной системы.
Классификация нейронов
Головной мозг человека насчитывает примерно 65 миллиардов нейронов, которые постоянно взаимодействуют между собой. Эти клетки подразделяются на несколько видов, каждый из которых выполняет свои особенные функции.
Чувствительный нейрон играет роль передатчика информации между органами чувств и центральными отделами человеческой нервной системы. Он воспринимает разнообразные раздражения, которые преобразовывает в нервные импульсы, а далее передает сигнал в головной мозг человека.
Двигательный — посылает импульсы в различные органы и ткани. В основном данный тип задействован в контроле над рефлексами спинного мозга.
За переработку и переключение импульсов отвечает вставочный нейрон. Функции данного типа клеток заключаются в получении и обработке информации от чувствительных и двигательных нейронов, между которыми они находятся. Более того, вставочные (или промежуточные) нейроны занимают 90 % центральной нервной системы человека, а также в больших количествах находятся во всех сферах головного и спинного мозга.
Строение промежуточных нейронов
Вставочный нейрон состоит из тела, аксона и дендритов. Каждая часть имеет свои специфические функции и отвечает за определенное действие. В его теле содержатся все компоненты, из которых созданы клеточные структуры. Важная роль этой части нейрона заключается в генерировании нервных импульсов и выполнении трофической функции. Продолговатый отросток, который несет сигнал от тела клетки, называется аксоном. Он делится на два типа: миелиновый и безмиелиновый. На конце аксона находятся различные синапсы. Третья составляющая нейронов — дендриты. Они являются короткими отростками, которые разветвляются в разные стороны. Их функция заключается в доставке импульсов к телу нейрона, что обеспечивает связь между различными видами нейронов центральной нервной системы.
Сфера воздействия
Что определяет область влияния вставочного нейрона? В первую очередь его собственное строение. В основном у клеток данного типа имеются аксоны, синапсы которых оканчиваются на нейронах этого же центра, что обеспечивает их объединение. Некоторые промежуточные нейроны активируются другими, из иных центров, а затем доставляют информацию в свой нейронный центр. Такие действия усиливают воздействие сигнала, который повторяется в параллельных путях, тем самым удлиняя срок хранения информационных данных в центре. В результате место, куда был доставлен сигнал, увеличивает надежность влияния на исполнительную структуру. Иные вставочные нейроны могут получать активацию от соединений двигательных «братьев» из своего центра. Потом они становятся передатчиками информации назад в свой центр, чем создают обратные связи. Таким образом, вставочный нейрон играет важную роль в образование особых замкнутых сетей, которые продлевают срок хранения информации в нервном центре.
Возбуждающий тип промежуточных нейронов
Вставочные нейроны делятся на два типа: возбуждающие и тормозные. При активации первых облегчается передача данных из одной нейронной группы в другую. Такую задачу выполняют именно «медленные» нейроны, которые имеют способность к длительной активации. Они передают сигналы на протяжении довольно длительного времени. Параллельно с этими действиями промежуточные нейроны активизируют и своих «быстрых» «коллег». Когда усиливается активность «медленных» нейронов, то уменьшается время реакции «быстрых». Одновременно с этим последние несколько замедляют работу «медленных».
Тормозной тип промежуточных нейронов
Вставочный нейрон тормозного типа приходит в активное состояние за счет прямых сигналов, которые поступают в их центр или исходят из него. Данное действие происходит путем обратных связей. Прямое возбуждение данного типа вставочных нейронов является характерным для промежуточных центров чувствительных путей спинного мозга. А в двигательных центрах коры головного мозга происходит активизация вставочных нейронов благодаря обратным связям.
Роль вставочных нейронов в работе спинного мозга
В работе спинного мозга человека важная роль отводится проводящим путям, которые расположены снаружи от пучков, исполняющих проводниковую функцию. Именно по этим дорожкам и передвигаются импульсы, которые посылает вставочный и чувствительный нейроны. Сигналы проходят вверх и вниз по этим путям, передавая различную информацию в соответствующие части мозга. Вставочные нейроны спинного мозга находятся в промежуточно-медиальном ядре, которое, в свою очередь, расположено в заднем роге. Промежуточные нейроны являются важной передней частью спинно-мозжечкового пути. На обратной стороне рога спинного мозга расположены волокна, состоящие из вставочных нейронов. Они образуют боковой спинно-таламический путь, который выполняет особую функцию. Он является проводником, то есть передает сигналы о болевых ощущениях и температурной чувствительности сначала в промежуточный мозг, а потом и в саму кору головного мозга.
Дополнительная информация о вставочных нейронах
В нервной системе человека вставочные нейроны выполняют особую и крайне важную функцию. Они связывают между собой различные группы нервных клеток, передают сигнал из головного мозга в спинной. Хотя именно этот тип является наиболее мелким по размерам. По форме вставочные нейроны напоминают звезду. Основное количество данных элементов располагается в сером веществе головного мозга, а их отростки не выступают за пределы центральной нервной системы человека.
Нейрон является специфической, электрически возбудимой клеткой в нервной системе человека и обладает уникальными особенностями. Его функции заключаются в обработке, хранении и передаче информации. Нейроны характеризуются сложным строением и узкой специализацией. Они также делятся на три вида. В этой статье подробно описывается вставочный нейрон и его роль в действии центральной нервной системы.
Классификация нейронов
Головной мозг человека насчитывает примерно 65 миллиардов нейронов, которые постоянно взаимодействуют между собой. Эти клетки подразделяются на несколько видов, каждый из которых выполняет свои особенные функции.
Чувствительный нейрон играет роль передатчика информации между органами чувств и центральными отделами человеческой нервной системы. Он воспринимает разнообразные раздражения, которые преобразовывает в нервные импульсы, а далее передает сигнал в головной мозг человека.
Двигательный — посылает импульсы в различные органы и ткани. В основном данный тип задействован в контроле над рефлексами спинного мозга.
За переработку и переключение импульсов отвечает вставочный нейрон. Функции данного типа клеток заключаются в получении и обработке информации от чувствительных и двигательных нейронов, между которыми они находятся. Более того, вставочные (или промежуточные) нейроны занимают 90 % центральной нервной системы человека, а также в больших количествах находятся во всех сферах головного и спинного мозга.
Строение промежуточных нейронов
Вставочный нейрон состоит из тела, аксона и дендритов. Каждая часть имеет свои специфические функции и отвечает за определенное действие. В его теле содержатся все компоненты, из которых созданы клеточные структуры. Важная роль этой части нейрона заключается в генерировании нервных импульсов и выполнении трофической функции. Продолговатый отросток, который несет сигнал от тела клетки, называется аксоном. Он делится на два типа: миелиновый и безмиелиновый. На конце аксона находятся различные синапсы. Третья составляющая нейронов — дендриты. Они являются короткими отростками, которые разветвляются в разные стороны. Их функция заключается в доставке импульсов к телу нейрона, что обеспечивает связь между различными видами нейронов центральной нервной системы.
Сфера воздействия
Что определяет область влияния вставочного нейрона? В первую очередь его собственное строение. В основном у клеток данного типа имеются аксоны, синапсы которых оканчиваются на нейронах этого же центра, что обеспечивает их объединение. Некоторые промежуточные нейроны активируются другими, из иных центров, а затем доставляют информацию в свой нейронный центр. Такие действия усиливают воздействие сигнала, который повторяется в параллельных путях, тем самым удлиняя срок хранения информационных данных в центре. В результате место, куда был доставлен сигнал, увеличивает надежность влияния на исполнительную структуру. Иные вставочные нейроны могут получать активацию от соединений двигательных «братьев» из своего центра. Потом они становятся передатчиками информации назад в свой центр, чем создают обратные связи. Таким образом, вставочный нейрон играет важную роль в образование особых замкнутых сетей, которые продлевают срок хранения информации в нервном центре.
Возбуждающий тип промежуточных нейронов
Вставочные нейроны делятся на два типа: возбуждающие и тормозные. При активации первых облегчается передача данных из одной нейронной группы в другую. Такую задачу выполняют именно «медленные» нейроны, которые имеют способность к длительной активации. Они передают сигналы на протяжении довольно длительного времени. Параллельно с этими действиями промежуточные нейроны активизируют и своих «быстрых» «коллег». Когда усиливается активность «медленных» нейронов, то уменьшается время реакции «быстрых». Одновременно с этим последние несколько замедляют работу «медленных».
Тормозной тип промежуточных нейронов
Вставочный нейрон тормозного типа приходит в активное состояние за счет прямых сигналов, которые поступают в их центр или исходят из него. Данное действие происходит путем обратных связей. Прямое возбуждение данного типа вставочных нейронов является характерным для промежуточных центров чувствительных путей спинного мозга. А в двигательных центрах коры головного мозга происходит активизация вставочных нейронов благодаря обратным связям.
Роль вставочных нейронов в работе спинного мозга
В работе спинного мозга человека важная роль отводится проводящим путям, которые расположены снаружи от пучков, исполняющих проводниковую функцию. Именно по этим дорожкам и передвигаются импульсы, которые посылает вставочный и чувствительный нейроны. Сигналы проходят вверх и вниз по этим путям, передавая различную информацию в соответствующие части мозга. Вставочные нейроны спинного мозга находятся в промежуточно-медиальном ядре, которое, в свою очередь, расположено в заднем роге. Промежуточные нейроны являются важной передней частью спинно-мозжечкового пути. На обратной стороне рога спинного мозга расположены волокна, состоящие из вставочных нейронов. Они образуют боковой спинно-таламический путь, который выполняет особую функцию. Он является проводником, то есть передает сигналы о болевых ощущениях и температурной чувствительности сначала в промежуточный мозг, а потом и в саму кору головного мозга.
Дополнительная информация о вставочных нейронах
В нервной системе человека вставочные нейроны выполняют особую и крайне важную функцию. Они связывают между собой различные группы нервных клеток, передают сигнал из головного мозга в спинной. Хотя именно этот тип является наиболее мелким по размерам. По форме вставочные нейроны напоминают звезду. Основное количество данных элементов располагается в сером веществе головного мозга, а их отростки не выступают за пределы центральной нервной системы человека.
Функции нейрона:
Воспринимающая – чувствительные окончания Ннов на дендритах и синапсы способны принимать Ин от внешних или внутренних раздражителей (трансформируя специфическую энергию раздражителя в неспецифический процесс нервного возбуждения – нервный импульс) и других Ннов для обработки и последующей передачи по каналам связи.
Интегративная – обработка одновременно или в течение короткого интервала времени поступающих нервных сигналов по механизму их алгебраической суммации, в результате которого на выходе нейрона формируется сигнал, несущий в себе информацию всех суммированных сигналов.
Мнестическая – существуют тонкие молекулярные биофизические процессы, сохраняющие след от всякого предыдущего воздействия и благодаря этому трансформирующие характер ответной реакции на всякое последующее. По существу, это элементарная форма памяти и научения.
Проводниковая – от тела нейрона по аксону к его окончанию в естественных условиях только в одном этом направлении распространяется, не затухая, нервный импульс. Скорость его распространения в зависимости от морфофункциональных особенностей проводника колеблется от нескольких сантиметров до 100-120 метров в секунду.
Эффекторная – передача Ин на др Кл (Нн или Кл исполнит органа) через химич. синапсы окончания аксона.
Конкретные функции тех или иных нейронов определяются их положением в нервной системе и заключаются в восприятии сигналов в виде электрической импульсации с периферии или от других нейронов, переработке и передаче ее на соседние нейроны или исполнительные органы, осуществляя тем самым регуляцию и координацию деятельности всех органов и систем организма.
Разным Ннам присущи разные функции:
1) Чувствительные Нны (афферентные, сенсорные) – передают Ин от органов чувств и рецепторов на др. Нны.
2) Вставочные – располагаются в ЦНС – передают Ин с Нна на Нн.
3) Двигательные (эфферентные, мотонейроны) – несут Ин от ЦНС к органам тела.
Кол-во Н-нов у человека значит превышает его потребности. Их образуется в эмбриогенезе с избытком и с течением времени те клетки которые постоянно задействованы специализируются, «приобретают опыт», а лишние Нны – постепенно отмирают. Не делятся, не восстанавливаются, т. к. включение нового «необученного» Нна в сложившийся нейронный ансамбль затруднило бы работу системы.
Деятельность нервной системы носит рефлекторный характер.
Если ущипнуть заднюю конечность децеребрированной (с интактным спинным мозгом) лягушки, она ее отдергивает. Если положить на спину такой лягушки смоченный кислотой кусочек фильтровальной бумаги, он вскоре будет сброшен точным движением ближайшей задней конечности. Подобного рода автоматическую, стереотипную, целенаправленную реакцию организма на стимул физиолог Унцер в 1771 г. назвал рефлексом.
Рефлекс – это простейшая ответная реакция организма на то или иное раздражение (внешнее или внутреннее воздействие), которая происходит при участи нервной системы. В осуществлении рефлекторной реакции принимают участие все типы нейронов по принципу так называемой рефлекторной дуги. Рефлекторная дуга -–это цепь нервных клеток от места возникновения раздражения (рецептор) к месту приложения (мышцы, железы). Первый нейрон дуги всегда афферентный, последний – всегда эфферентный. Простейшая рефлекторная дуга состоит из двух нейронов – чувствительного и двигательного. Тело первого (афферентного) нейрона находится в спинно-мозговом узле спинномозгового нерва или чувствительном узле черепного нерва. Дендрит этой клетки в составе соответствующего нерва направляется на периферию, где заканчивается рецепторным аппаратом. В рецепторе энергия внешнего или внутреннего раздражителя преобразуется в нервный импульс, который по нервному волокну передается к телу клетки. Асон через задний корешок спинно-мозгового или соответствующего корешка черепного нерва следует в спинной или головной мозг. В сером веществе спинного или ядрах головного мозга этот отросток чувствительной клетки образует синапс с телом второго (эфферентного) нейрона. Аксон двигательного нейрона выходит из спинного (головного) мозга в составе передних корешков спинно-мозгового или соответствующего черепного нерва и направляется к рабочему органу.
Чаще всего рефлекторная дуга состоит из многих нейронов. Тогда между афферентным и эфферентным клетками расположены вставочные нейроны. В такой рефлекторной дуге возбуждение от чувствительного нейрона передается по аксону к одному или нескольким вставочным нейронам. Аксоны одних вставочных нейронов направляются к двигательным клеткам передних рогов спинного мозга того же сегмента и образуют синапс с эффекторным нейроном, замыкая тем самым трехнейронную рефлекторную дугу. Аксоны других вставочных нейронов разделяются Т-образно, их ветви идут к эфферентным клеткам передних рогов спинного мозга ниже- или вышележащих сегментов (от 1 до 3-х в каждом направлении). Помимо этого, от каждого из описанных отростков нервных клеток отходят ответвления, направляющиеся к эфферентным клеткам тех сегментов, мимо которых они проходят. Поэтому раздражение одной точки тела может передаваться не только к соответствующему сегменту мозга, но и охватывать несколько соседних сегментов. В результате этого простой рефлекс перерастает в ответную реакцию, захватывающую несколько групп мышц. Возникает сложное, координированное, но все-таки рефлекторное движение.
Всем известно множество примеров врожденных рефлексов (безусловных по Павлову) — роговичный, кашлевой, глотательный, реакция отдергивания и т.д., а также рефлексов, приобретенных в течение жизни (условных). Однако в большинстве случаев мы их даже не осознаем. Это — рефлексы, обеспечивающие прохождение и переваривание пищи в желудке и кишечнике, непрерывно приспосабливающие кровообращение и дыхание к текущим потребностям организма, и т д.
В психологии представления о рефлекторном механизме движения положили начало различным теориям поведения, делающим упор на связи стимул – реакция (бихевиоризм). Однако имеется точка зрения, что поведение далеко не всегда имеет рефлекторный характер. Децеребрированные (с удаленным головным мозгом) собаки способны к ритмичным движениям, например к почесыванию спины задней лапой или локомоции, даже после отключения всех сенсорных входов спинного мозга путем перерезки дорсальных корешков. Следовательно, организация движений не всегда основана на рефлексах.
Ряд движений вообще регулируется нервной системой даже в отсутствие всяких внешних стимулов, например, дыхание. Такие последовательности движений, поддерживаемые ЦНС без внешней стимуляции, называются «запрограммированными», или автоматическими. После того как была обнаружена способность ЦНС к такой деятельности, быстро получила признание гипотеза, согласно которой движения регулируются в основном программами, а не рефлексами, и представление о «программной организации» ЦНС стало общепринятым. Дыхание, ходьба, чесание-все это примеры врожденных программ, к которым в течение жизни индивида добавляется множество приобретенных. Среди последних есть спортивные или профессиональные навыки (гимнастические движения, печатание на машинке и т. п.), становящиеся в результате соответствующей практики почти автоматическими.
Совокупность нейронов образует нервную систему
Нервные клетки образуют структуры нервной системы.
Функция нервной системы – регуляция процессов в организме адекватно информации, которая поступает по каналам сенсорной связи. Функционально нервную систему условно подразделяют на две части – соматическую (анимальную) и вегетативную (автономную), имеющие как центральный так и периферический отделы.
Соматическая нервная система иннервирует главным образом тело (сому), а именно скелетные мышцы, кожу и некоторые внутренние органы (язык, гортань, глотку, мышцы глазного яблока, среднего уха), обеспечивает связь организма с внешней средой.
Вегетативная (автономная) нервная система состоит из симпатической и парасимпатической частей, которые включают скопления клеток, расположенных в головном и спинном мозге, вегетативные нервы, узлы и сплетения. Она иннервирует гладкие мышцы внутренних органов и сосудов, сердце, железы, а также обеспечивает регуляцию обменных процессов во всех органах и тканях.
В соответствии с основными функциональными задачами нервная система состоит из 3-х функциональных 3-х звеньев
чувствительное – чувствительные волокна нервов и нервные центры и сенсорные участки коры конечного мозга, обрабатывающие сенсорную информацию – обеспечивают восприятие сигнала раздражителя и его обработку. И.П.Павлов ввел понятие анализатора – относительно автономная структура, обеспечивающая переработку специфической сенсорной Ин и прохождение ее на всех уровнях НС.
двигательное (моторное) – структуры центральной и периферической нервной системы, формирующие движения, вплоть до возникновения целостных поведенческих актов
интегративное (АССОЦИАТИВНОЕ) – ядра нервных центров и некоторые области коры конечного мозга, которые обеспечивают связь чувствительного звена с двигательным. Интегративные функции ЦНС не связаны непосредственно с обработкой сенсорных сигналов или управлением двигательными и вегетативными структурами, а связаны с их координацией.
Чувствительно, моторное и ассоциативное звенья имеют место во всех отделах нервной системы.
Структурно нервная система подразделяется на две части: центральную и периферическую. К периферической нервной системе относятся спинномозговые и черепномозговые узлы и периферические нервы (12 пар черепномозговых и 31 пара спинномозговых), их сплетения и узлы, лежащие в различных отделах тела. С помощью пучков нервных волокон, тела которых находятся в центральной части нервной системы, периферическая нервная система передает информацию от органов и тканей в ЦНС и обратно.
Центральная нервная система (ЦНС) представлена головным и спинным мозгом.
Она защищена костной тканью черепа, окружена оболочками, состоит из белого и серого вещества. Серое в-во – ядра нейронов. Белое в-во — отростки нейронов – образуют восходящие чувствительные пути и нисходящие двигательные пути.
Выделяют в ЦНС:
Спинной мозг — цилиндрический тяж (45 см у ♂, 42см у ♀), который располагается в канале позвоночного столба и имеет сегментное (метамерное) строение. Количество сегментов соответствует количеству позвонков. От каждого сегмента отходят 2 пары корешков (нервных пучков) – передние и задние, которые, соединяясь образуют периферические спинномозговые нервы.
Спинной мозг осуществляет 2 основные функции – рефлекторную и проводниковую. Большинство рефлексов связано с работой спинного мозга. С разрушением спинного мозга все рефлексы исчезают. Проводниковая функция заключается в осуществлении двусторонней связи головного мозга с различными системами организма.
СМ сверху переходит в продолговатый мозг, а внизу заканчивается конусом с концевой нитью.
Головной мозг состоит из ряда отделов:
1) задний мозг
В основании заднего мозга располагается продолговатый мозг, который является непосредственным продолжением спинного. Продолговатый мозг является важным звеном, контролирующим многие базовые биологические функции организма.
Во-первых, он регулирует деятельность сердечно-сосудистой, пищеварительной и дыхательной систем, каждую секунду определяя, как часто и насколько глубоко нам нужно дышать, с какой частотой должно биться наше сердце, сколько крови оно должно прокачивать.
Во-вторых, он позволяет осуществлять ряд важных рефлексов, направленных на восприятие переработку и проглатывание пищи (глотания, сосания, жевания, слюноотделение) и защиты (рефлексы рвоты, чихания, кашля, слезоотделения, замыкания век).
В-третьих, он помогает поддерживать равновесие тела, управляя ориентацией головы и конечностей, на которые действует гравитация.
Сразу за продолговатым мозгом задний мозг расширяется, и этот участок известен как мост. Мост содержит области, интегрирующие движения и ощущения, поступающие от лицевой мускулатуры, языка, глаз и ушей. Другие участки моста принимают участие в регуляции уровня бодрствования и инициируют процесс засыпания.
Наиболее зрительно выделяющейся частью заднего мозга является мозжечок —массивная структура, которая находится позади моста и закрывает продолговатый мозг. Мозжечок работает как сложнейший компьютер, более тридцати миллиардов нейронов которого суммируют информацию от гравитационных рецепторов, а также от мышц, связок и сухожилий всего тела. Эта своеобразная структура имеет ряд отдельных областей. Одна область полностью контролирует равновесие тела. Любое повреждение этой области, будь то травма, заболевание или алкогольная интоксикация, приводит к нарушениям координации движений и шатающейся походке. Другая область мозжечка контролирует выполнение точных, профессиональных движений, будь то игра в теннис или фортепьянное арпеджио. Повреждение этой области вызывает тремор (дрожание) конечностей при выполнении движений и невозможность быстрого выполнения чередующихся движений (например, поочередное постукивание пальцами).
2) Средний мозг
Средний мозг управляет формированием слуховых и зрительных стимулов. Одна из областей среднего мозга контролирует движения глаз. Другие области среднего мозга принимают участие в терморегуляции, болевой рецепции (восприятии боли), а также сотрудничают с мостом в регуляции цикла сон-бодрствование. Средний мозг осуществляет преобразование простых движений, порождаемых задним мозгом, в более сложные действия.
3) Передний мозг
Передний мозг включает в себя все, что располагается над средним. Передний мозг человека настолько велик, что он .окружает практически весь средний и часть заднего мозга. Наиболее заметной частью переднего мозга является большая морщинистая поверхность — кора головного мозга
Передний мозг обладает ярко выраженной билатеральной симметрией. Особенно заметной эта симметрия становится при рассмотрении коры, по обе стороны от глубокой продольной борозды, проходящей между полушариями. В каждом полушарии под корой находится остальная часть переднего мозга, включающая ряд подкорковых структур:
1) таламус — располагается глубоко в толще тканей над средним мозгом. В нем концентрируется множество центров, функционирующих как ретрансляционные (переключающие) станции почти для всей сенсорной информации, подающейся к коре.
2) гипоталамус — находится непосредственно под таламусом и управляет непроизвольным поведением, таким, как питание, поддержание адекватной температуры тела, реакция на угрозу и вовлечение в сексуальную активность, осуществляет регуляцию работы всех внутренних органов человека.
3) базальные ганглии — располагаются сразу над таламусом в каждом полушарии. Они принимают активное участие в регуляции мышечных сокращений (особенно плавных), а также предохраняют наши движения от судорог. Болезнь Паркинсона вызывается дегенерацией клеток в области базальных ганглиев. У страдающих этой болезнью обычно отмечается снижение мышечного тонуса, неподвижное маскообразное лицо, медленные движения и тремор конечностей в расслабленном состоянии. Базальные ганглии поражаются и при болезни Хантингтона (хорее), прогрессирующем наследственном заболевании, которое может вызывать судороги конечностей, лицевые тики и неконтролируемые судороги всего тела.
4) лимбическая система — группа взаимосвязанных подкорковых структур, окружающая таламус и базальные ганглии (от французского limbique — граничащий, ограничивающий). Лимбическая система включает в себя такие структуры, как миндалина и гиппокамп, имеет тесные анатомические связи со многими другими структурами мозга, в особенности таламусом, корой и центрами обоняния. Она вовлечена в регуляцию эмоций и мотиваций, а также в процессы памяти и научения.
Кора переднего мозга (Кора больших полушарий, КБП)– высший центр регуляции жизненных функций и поведения. Осуществляет анализ и синтез всей Ин, поступающей из внешней и внутренней сред организма.
Кора конечного мозга составляет 80 процентов всего мозга, однако она — лишь его участок толщиной 2-3 мм. Если разгладить все извилины коры человеческого мозга, то она заняла бы площадь порядка 2500 кв. см. Чтобы вместить эту структуру в развернутом виде, понадобилась бы голова нестандартных размеров (и, что важнее, понадобились бы гораздо более длинные нервы для присоединения всех ее областей). Но к счастью, эта гигантская структура в сжатом виде умещается в весьма ограниченном пространстве человеческого черепа.
Некоторые из извилин образуют очень глубокие борозды. Самой глубокой является продольная борозда, которая разделяет правое и левое полушария. Другие борозды отграничивают несколько больших участков в каждом полушарии, именуемых долями. Существует четыре таких доли, названных по имени близлежащей кости черепа В каждом полушарии лобная и теменная доли формируют самую верхнюю часть мозга; борозда, разделяющая их, именуется центральной. Височные доли, располагающиеся по бокам от лобных, отделены боковой (латеральной) бороздой. Последняя, затылочная доля плотно примыкает сзади к височным и теменной долям.
В первой половине XX в. ученые произвели подробное картирование коры головного мозга человека на основании тщательного анализа неврологических и психических нарушений, возникающих при ее местных повреждениях (вследствие огнестрельных ранений или заболеваний) и высказали предположение о жесткой приуроченности тех или иных психических функций к определенным участкам коры.
ЛОКАЛИЗАЦИОНИЗМ – каждая психическая функция локализована (привязана) в определенном участке КБП.
А.Р.Лурия выделил 3 блока:
1) поддержание активности – центральные области КБП,
2) познавательные процессы – получение, переработка и хранение Ин – задн. и височн. отделы КБП,
3) мышление, поведенческая регуляция, самоконтроль – передние отделы КБП.
Эксперименты показали наличие первичных двигательных зон, которые ответственны за формирование двигательных актов, и первичных сенсорных зон, связанных с обработкой специфической сенсорной информацией.
Например, первичная соматосенсорная зона расположена в теменной доле каждого полушария и является центром приема сенсорной информации от рецепторов кожи. Подобные первичные проекционные зоны существуют для зрения и слуха. Они располагаются в затылочной и височной долях соответственно. Пациенты, которым стимулировали визуальные проекционные зоны, говорили о своих зрительных ощущениях — довольно ярких, но весьма неопределенных: мерцающие огни, полосы цвета. Стимуляция же слуховых зон вызывала бессмысленные и невнятные слуховые ощущения: щелчки, шумы, шорохи, звонки.
2/3 площади (до 75%) занимают вторичные зоны (ассоциативные, интегральные) – обеспечивают синтез и интегрирование отдельных элементов информации в целостную картину. Они элементарные ощущения складывают в целостное восприятие, а из отдельных движений формируют целостные поведенческие акты.
На сегодняшний день к ассоциативной (неспецифичной, интегративной) коре в узком смысле слова относят: теменно-височно-затылочную (высшие сенсорные функций и речь), лобную, точнее, префронтальную (координирование сложных движений и поведения в целом, выработка стратегий поведения) и лимбическую (функции памяти и эмоций) ассоциативные зоны.
В последние годы в опытах на животных были получены данные, сильно поколебавшие классические представления о разделении коры на сенсорную, двигательную и ассоциативную. По крайней мере, в работах на крысах с использованием пероксидазы хрена показано, что все изученные области коры получают зрительную, слуховую и соматовисцеральную афферентацию от таламуса и в свою очередь посылают сигналы к этому отделу. Места для ассоциативных зон между сенсорными полями просто не обнаруживается. С этими анатомическими данными согласуются и результаты опытов, в которых животные после разрушения соответствующих первичных проекционных областей коры не становились ни слепыми, ни глухими; кроме того, оказалось, что агнозию (неспособность интерпретировать определенные сенсорные стимулы или соотносить их с окружающей обстановкой) невозможно вызвать путем разрушения только лишь ассоциативных полей. Представления о двигательной коре также нуждаются в пересмотре; как выяснилось, аксоны почти от всех зон коры подходят к передним рогам спинного мозга (как будто вся кора — «двигательная»), причем повреждения первичных моторных областей практически не приводят к двигательным расстройствам, если не считать нарушения движений пальцев.
В противовес теории локализации функций высказывалась и противоположная (холистическая – целостная) точка зрения, оспаривающая столь дискретную локализацию различных психических функций и утверждающая, что мозг во всех случаях работает как единое целое. Так, Лешли обнаружил, что нарушения, возникающие у крыс при удалении различных участков коры, в большей степени зависят от величины разрушаемой области, нежели от ее местоположения. На основании этих данных он сформулировал концепцию «эквипотенциальности» всех отделов головного мозга, в соответствии с которой любой из них может принимать на себя функции другого.
Что делает вставочный нейрон. Чувствительный или сенсорный нейрон. Интегративная деятельность нейрона
Вообще, в зависимости возложенных на нейроны задач и обязанностей, они делятся на три категории:
— Сенсорные (чувствительные) нейроны принимают и передают импульсы от рецепторов «в центр», т.е. центральную нервную систему. Причем сами рецепторы — это специально обученные клетки органов чувств, мышц, кожи и суставов умеющие обнаруживать физические или химические изменения внутри и снаружи нашего организма, преобразовывать их в импульсы и радостно передавать их сенсорным нейронам. Таким образом, сигналы идут от периферии к центру.
Следующий тип:
— Моторные (двигательные) нейроны, которые урча, фырча и бибикая, несут сигналы, выходящие из головного или спинного мозга, к исполнительным органам, коими являются мышцы, железы и т.д. Ага, значит, сигналы идут от центра к периферии.
Ну а промежуточные (вставочные) нейроны, попросту говоря, являются «удлинителями», т.е. получают сигналы от сенсорных нейронов и посылают эти импульсы дальше к другим промежуточным нейронам, ну или сразу к моторным нейронам.
В общем и целом вот что получается: у сенсорных нейронов дендриты соединены с рецепторами, а аксоны — с другими нейронами (вставочными). У двигательных нейронов наоборот, дендриты соединены с другими нейронами (вставочными), а аксоны — с каким-нибудь эффектором, т.е. стимулятором сокращения какой-нибудь мышцы или секреции железы. Ну а, соответственно, у вставочных нейронов и дендриты и аксоны соединяются с другими нейронами.
Получается что самый простой путь, по которому может идти нервный импульс, будет состоять из трех нейронов: одного сенсорного, одного вставочного и одного моторного.
Ага, а давайте теперь вспомним дядьку — очень «нервного патолога», с ехидной улыбкой стучащего своим «волшебным» молоточком по колену. Знакомо? Вот, это и есть простейший рефлекс: когда он ударяет по коленному сухожилию, прикрепленная к нему мышца растягивается и сигнал от находящихся в ней чувствительных клеток (рецепторов) передается по сенсорным нейронам в спинной мозг. А уже в нем сенсорные нейроны контактируют либо через вставочные, либо непосредственно с моторными нейронами, которые в ответ посылают импульсы назад в ту же самую мышцу, заставляя ее сокращаться, а ногу — распрямляться.
Сам же спинной мозг удобно примостился внутри нашего позвоночника. Он мягкий и ранимый, потому и прячется в позвонках. Спинной мозг всего 40-45 сантиметров в длину, с мизинец толщиной (около 8 мм) и весит каких-то 30 грамм! Но, несмотря на всю свою тщедушность, спинной мозг является управляющим центром сложной сети нервов, раскинутой по телу. Практически как центр управлениями полетами! 🙂 Без него ни опорно-двигательный аппарат, ни основные жизненные органы ну никак не могут действовать и работать.
Свое начало спинной мозг берет на уровне края затылочного отверстия черепа, а заканчивается на уровне первого-второго поясничных позвонков. А вот уже ниже спинного мозга в позвоночном канале находится такой густой пучок нервных корешков, прикольно именуемый конским хвостом, видимо за сходство с ним. Так вот, конский хвост – это продолжение нервов, выходящих из спинного мозга. Они отвечают за иннервацию нижних конечностей и органов таза, т.е. передают сигналы от спинного мозга к ним.
Спинной мозг окружен тремя оболочками: мягкой, паутинной и твердой. А пространство между мягкой и паутинной оболочками заполнено еще и большим количеством спинномозговой жидкости. Через межпозвоночные отверстия от спинного мозга отходят спинномозговые нервы: 8 пар шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 или 2 копчиковых. Почему пар? Да потому, что спинномозговой нерв выходит двумя корешками: задним (чувствительным) и передним (двигательным), соединенными в один ствол. Так вот, каждая такая пара контролирует определенную часть тела. Т.е., например, если вы нечаянно схватились за горячую кастрюлю (не дай бог! Тьфу-тьфу-тьфу!), то в окончаниях чувствительного нерва тут же возникает болевой сигнал, сразу же поступающий в спинной мозг, и уже оттуда — в парный двигательный нерв, который и передает приказ: «Ахтунг-ахтунг! Немедленно убрать руку!» Причем, поверьте, это происходит очень быстро — еще до того, как головной мозг зарегистрирует болевой импульс. В итоге, вы успеваете отдернуть руку от кастрюли еще до того, как почувствуете боль. Конечно же, такая реакция спасает нас от тяжелых ожогов или других повреждений.
Вообще, практически все наши автоматические и рефлекторные действия контролируются спинным мозгом, ну за исключением тех, за которыми следит сам головной мозг. Ну, вот, например: мы воспринимаем увиденное с помощью глазного нерва идущего в головной мозг, и в то же время обращаем свой взор в разные стороны при помощи глазных мышц, которые управляются уже спинным мозгом. Да и плачем мы то же по приказу спинного мозга, который «заведует» слезными железами.
Можно сказать, что наши сознательные действия идут от головного мозга, но как только эти действия мы начинаем выполнять уже автоматически и рефлекторно — они передаются в ведение спинного мозга. Так что, когда мы только учимся что-то делать, то, конечно же, сознательно обдумываем и продумываем и осмысливаем каждое движение, а значит, используем головной мозг, но со временем мы уже можем делать это автоматически, и это значит, что головной мозг передает «бразды правления» этим действием спинному, просто ему уже стало скучно и неинтересно….потому как, наш головной мозг очень пытливый, любознательный и любит учиться!
Ну вот, пришло и нам время полюбопытствовать……
Нейрон является специфической, электрически возбудимой клеткой в нервной системе человека и обладает уникальными особенностями. Его функции заключаются в обработке, хранении и передаче информации. Нейроны характеризуются сложным строением и узкой специализацией. Они также делятся на три вида. В этой статье подробно описывается вставочный нейрон и его роль в действии центральной нервной системы.
Классификация нейронов
Головной мозг человека насчитывает примерно 65 миллиардов нейронов, которые постоянно взаимодействуют между собой. Эти клетки подразделяются на несколько видов, каждый из которых выполняет свои особенные функции.
Чувствительный нейрон играет роль передатчика информации между органами чувств и центральными отделами человеческой нервной системы. Он воспринимает разнообразные раздражения, которые преобразовывает в нервные импульсы, а далее передает сигнал в головной мозг человека.
Двигательный — посылает импульсы в различные органы и ткани. В основном данный тип задействован в контроле над рефлексами спинного мозга.
За переработку и переключение импульсов отвечает вставочный нейрон. Функции данного типа клеток заключаются в получении и обработке информации от чувствительных и двигательных нейронов, между которыми они находятся. Более того, вставочные (или промежуточные) нейроны занимают 90 % центральной нервной системы человека, а также в больших количествах находятся во всех сферах головного и спинного мозга.
Строение промежуточных нейронов
Вставочный нейрон состоит из тела, аксона и дендритов. Каждая часть имеет свои специфические функции и отвечает за определенное действие. В его теле содержатся все компоненты, из которых созданы клеточные структуры. Важная роль этой части нейрона заключается в генерировании нервных импульсов и выполнении трофической функции. Продолговатый отросток, который несет сигнал от тела клетки, называется аксоном. Он делится на два типа: миелиновый и безмиелиновый. На конце аксона находятся различные синапсы. Третья составляющая нейронов — дендриты. Они являются короткими отростками, которые разветвляются в разные стороны. Их функция заключается в доставке импульсов к телу нейрона, что обеспечивает связь между различными видами нейронов центральной нервной системы.
Сфера воздействия
Что определяет область влияния вставочного нейрона? В первую очередь его собственное строение. В основном у клеток данного типа имеются аксоны, синапсы которых оканчиваются на нейронах этого же центра, что обеспечивает их объединение. Некоторые промежуточные нейроны активируются другими, из иных центров, а затем доставляют информацию в свой нейронный центр. Такие действия усиливают воздействие сигнала, который повторяется в параллельных путях, тем самым удлиняя срок хранения информационных данных в центре. В результате место, куда был доставлен сигнал, увеличивает надежность влияния на исполнительную структуру. Иные вставочные нейроны могут получать активацию от соединений двигательных «братьев» из своего центра. Потом они становятся передатчиками информации назад в свой центр, чем создают обратные связи. Таким образом, вставочный нейрон играет важную роль в образование особых замкнутых сетей, которые продлевают срок хранения информации в нервном центре.
Возбуждающий тип промежуточных нейронов
Вставочные нейроны делятся на два типа: возбуждающие и тормозные. При активации первых облегчается передача данных из одной нейронной группы в другую. Такую задачу выполняют именно «медленные» нейроны, которые имеют способность к длительной активации. Они передают сигналы на протяжении довольно длительного времени. Параллельно с этими действиями промежуточные нейроны активизируют и своих «быстрых» «коллег». Когда усиливается активность «медленных» нейронов, то уменьшается время реакции «быстрых». Одновременно с этим последние несколько замедляют работу «медленных».
Тормозной тип промежуточных нейронов
Вставочный нейрон тормозного типа приходит в активное состояние за счет прямых сигналов, которые поступают в их центр или исходят из него. Данное действие происходит путем обратных связей. Прямое возбуждение данного типа вставочных нейронов является характерным для промежуточных центров чувствительных путей спинного мозга. А в двигательных центрах коры головного мозга происходит активизация вставочных нейронов благодаря обратным связям.
Роль вставочных нейронов в работе спинного мозга
В работе спинного мозга человека важная роль отводится проводящим путям, которые расположены снаружи от пучков, исполняющих проводниковую функцию. Именно по этим дорожкам и передвигаются импульсы, которые посылает вставочный и чувствительный нейроны. Сигналы проходят вверх и вниз по этим путям, передавая различную информацию в соответствующие части мозга. Вставочные нейроны спинного мозга находятся в промежуточно-медиальном ядре, которое, в свою очередь, расположено в заднем роге. Промежуточные нейроны являются важной передней частью спинно-мозжечкового пути. На обратной стороне рога спинного мозга расположены волокна, состоящие из вставочных нейронов. Они образуют боковой спинно-таламический путь, который выполняет особую функцию. Он является проводником, то есть передает сигналы о болевых ощущениях и температурной чувствительности сначала в промежуточный мозг, а потом и в саму кору головного мозга.
Дополнительная информация о вставочных нейронах
В нервной системе человека вставочные нейроны выполняют особую и крайне важную функцию. Они связывают между собой различные группы нервных клеток, передают сигнал из головного мозга в спинной. Хотя именно этот тип является наиболее мелким по размерам. По форме вставочные нейроны напоминают звезду. Основное количество данных элементов располагается в сером веществе головного мозга, а их отростки не выступают за пределы центральной нервной системы человека.
Соединяющий нейрон, который находится между сенсорным (афферентным) и моторными (эфферентным) нейронами. Находится в центральной нервной системе. Также называется промежуточный нейрон, и в более старых текстах – ассоциативный нейрон.
Смотреть значение
Вставочный Нейрон в других словаряхВставочный Прил. — 1. Предназначенный для вставки, вставления.
Толковый словарь Ефремовой
Нейрон М. — 1. То же, что: неврон.
Толковый словарь Ефремовой
Вставочный — (шн), вставочная, вставочное. Прил. к вставка.
Толковый словарь Ушакова
Нейрон — нейрона, м. (греч. neuron — волокно, нерв) (анат.). Нервная клетка.
Толковый словарь Ушакова
Нейрон — -а; м. [от греч. neuron — нерв] Спец. Нервная клетка со всеми отходящими от неё отростками.
Толковый словарь Кузнецова
Вставочный Диск — (discus intercalatus, LNH) общее название микроскопических структур в месте контакта смежных мышечных клеток миокарда, обеспечивающих их соединение в мышечные комплексы и передачу……..
Большой медицинский словарь
Двигательный Нейрон — , нервная клетка, проводящая информацию на ЭФФЕКТОРЫ (обычно мышцы), от ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ (ЦНС), таким образом вызывая соответствующую реакцию. Аксоны (отростки,……..
Нейрон — (нервная клетка), основная структурная и функциональная единица НЕРВНОЙ СИСТЕМЫ, осуществляющая быструю передачу НЕРВНЫХ ИМПУЛЬСОВ между различными органами. Состоит……..
Научно-технический энциклопедический словарь
Сенсорный Нейрон — (чувствительный нейрон), нервная клетка, проводящая информацию от РЕЦЕПТОРОВ в любой части тела к ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ (ЦНС). Их нервные окончания находятся в……..
Научно-технический энциклопедический словарь
Нейрон — (neuronum, neurocytus, LNH; греч. neuron жила, нерв; син.: клетка нервная, невроцит, нейроцит) клетка, способная воспринимать раздражение, приходить в состояние возбуждения, вырабатывать……..
Большой медицинский словарь
Нейрон Амакриновый — (n. amacrinum, LNH) Н., расположенный во внутреннем зернистом слое сетчатки и обеспечивающий связь между нейронами этого слоя.
Большой медицинский словарь
Нейрон Ассоциативный — см. Нейрон вставочный.
Большой медицинский словарь
Нейрон Афферентный — (n. afferens, n. sensorium: син.: Н. рецепторный, Н. сенсорный, Н. чувствительный) Н., осуществляющий восприятие и передачу возбуждения от рецепторов к другим Н. центральной нервной системы.
Большой медицинский словарь
Нейрон Биполярный — (n. bipolare, LNH) Н., имеющий два отростка — аксон и дендрит.
Большой медицинский словарь
Нейрон Вегетативный — общее название Н., входящих в состав ганглиев, сплетений и нервов вегетативной нервной системы.
Большой медицинский словарь
Нейрон Веретеновидный — (n. fusiforme, LNH) мультиполярный вставочный Н. вытянутой формы, встречающийся в молекулярной пластинке коры полушарий большого мозга.
Большой медицинский словарь
Нейрон Веретеновидный Горизонтальный — (n. fusiforme horizontale, LNH) мультиполярный Н. вытянутой формы, встречающийся преимущественно между слоем грушевидных нейронов и зернистым слоем коры мозжечка.
Большой медицинский словарь
Нейрон Внутренний — (n. internum, LNH) Н. внутренних отделов переднего рога спинного мозга, аксон которого переходит через белую спайку на противоположную половину спинного мозга.
Большой медицинский словарь
Нейрон Вставочный — (n. intercalatum; син.: Н. ассоциативный, Н. промежуточный) Н., участвующий в передаче возбуждения от афферентных Н. к эфферентным.
Большой медицинский словарь
Нейрон Входной — формальный нейрон, выполняющий в конкретной системе нейронов (нейронной сети) функцию входа, т. е. воспринимающий сигналы только от внешней для данной системы среды.
Большой медицинский словарь
Нейрон Гигантопирамидальный — (n. gigantopyramidale, LNH; син.: Беца клетка, клетка пирамидная гигантская) крупный пирамидальный Н. внутренней пирамидальной пластинки коры большого мозга; аксоны Н. г. образуют……..
Большой медицинский словарь
Нейрон Горизонтальный — (n. horizomale, LNH) 1) Н. внутреннего зернистого слоя сетчатки глаза, отростки которого контактируют с центральными окончаниями фоторецепторных клеток, осуществляя перераспределение……..
Большой медицинский словарь
Нейрон Грушевидный — (n. piriforme, LNH; син. Пуркинье клетка) эфферентный Н. коры мозжечка, расположенный в ее ганглионарном слое и имеющий грушевидную форму.
Большой медицинский словарь
Нейрон Двигательный — см. Мотонейрон.
Большой медицинский словарь
Нейрон Длинноаксонный — (n. longiaxonicum, LNH; син. Догеля клетка I типа) мультиполярный вегетативный Н., аксон которого передает импульсы к гладкой или сердечной мышечной ткани.
Большой медицинский словарь
Нейрон Звездчатый — (n. stellatum, LNH) вставочный Н. звездчатой формы.
Большой медицинский словарь
Нейрон Звездчатый Длинноаксонный — (n. stellatum longiaxonicum, LNH) Н. з., расположенный в зернистом слое коры мозжечка, имеющий аксон, выходящий в белое вещество.
Большой медицинский словарь
Нейрон Звездчатый Короткоаксонный — (n. stellatum breviaxonicum, LNH) Н. з. зернистого слоя коры мозжечка, имеющий аксон, идущий к клубочкам мозжечка.
Большой медицинский словарь
Нейрон Зерновидный — (n. granulare, LNH) общее название мелких Н. округлой, угловатой и пирамидальной формы, расположенных в наружной зернистой пластинке коры большого мозга, дендриты которых поднимаются……..
Большой медицинский словарь
Нейрон Зерновидный Большой — (granoneurocytus magnus, LNH) общее название крупных Н., расположенных в молекулярном слое коры мозжечка, дендриты которых распространяются в молекулярном слое, а аксоны идут в зернистый……..
Большой медицинский словарь
Составляют 90% всех нейронов. Отростки не покидают пределов ЦНС, но обеспечивают многочисленные связи по горизонтали и вертикали.
Особенность: могут генерировать потенциал действия с частотой 1000 в сек. Причина — короткая фаза следовой гиперполяризации.
Вставочные нейроны осуществляют обработку информации; осуществляют связь между эфферентными и афферентными нейронами. Делятся на возбуждающие и тормозные.
Эфферентные нейроны .
Это нейроны, передающие информацию от нервного центра к исполнительным органам.
Пирамидные клетки двигательной зоны коры больших полушарий, посылающие импульсы к мотонейронам передних рогов спинного мозга.
Мотонейроны – аксоны выходят за пределы ЦНС и заканчиваются синапсом на эффекторных структурах.
Терминальная часть аксона ветвится, но есть ответвления и вначале аксона – аксонные коллатерали. Место перехода тела мотонейрона в аксон – аксонный холмик – наиболее возбудимый участок. Здесь генерируется ПД, затем распространяется по аксону.
На теле нейрона огромное количество синапсов. Если синапс образован аксоном возбуждающего интернейрона, то при действии медиатора на постсинаптической мембране возникает деполяризация или ВПСП (возбуждающий постсинаптический потенциал). Если синапс образован аксоном тормозной клетки, то при действии медиатора на постсинаптической мембране возникает гиперполяризация или ТПСП. Алгебраическая сумма ВПСП и ТПСП на теле нервной клетке проявляется в возникновении ПД в аксонном холмике.
Ритмическая активность мотонейронов в нормальных условиях 10 импульсов в секунду, но может возрастать в несколько раз.
Проведение возбуждения.
ПД распространяется за счет местных токов ионов, возникающих между возбужденным и невозбужденным участками мембраны. Так как ПД генерируется без затрат энергии, то нерв обладает самой низкой утомляемостью.
Объединения нейронов .
Существуют разные термины, обозначающие объединения нейронов.
Нервный центр – комплекс нейронов в одном или разных местах ЦНС (например, дыхательный центр).
Нейронные цепи – последовательно соединенные нейроны, выполняющие определенную задачу (с этой точки зрения рефлекторная дуга – тоже нейронные цепи).
Нейронные сети – более обширное понятие, т.к. помимо последовательных цепей имеются параллельные цепи нейронов, а также связи между ними. Нейронные сети – это структуры, выполняющие сложные задачи (например, задачи по обработке информации).
НЕРВНАЯ РЕГУЛЯЦИЯ
| | | следующая лекция ==> | |
Нейрон является специфической, электрически возбудимой клеткой в нервной системе человека и обладает уникальными особенностями. Его функции заключаются в обработке, хранении и передаче информации. Нейроны характеризуются сложным строением и узкой специализацией. Они также делятся на три вида. В этой статье подробно описывается вставочный нейрон и его роль в действии центральной нервной системы.
Классификация нейронов
Головной мозг человека насчитывает примерно 65 миллиардов нейронов, которые постоянно взаимодействуют между собой. Эти клетки подразделяются на несколько видов, каждый из которых выполняет свои особенные функции.
Чувствительный нейрон играет роль передатчика информации между органами чувств и центральными отделами человеческой нервной системы. Он воспринимает разнообразные раздражения, которые преобразовывает в нервные импульсы, а далее передает сигнал в головной мозг человека.
Двигательный — посылает импульсы в различные органы и ткани. В основном данный тип задействован в контроле над рефлексами спинного мозга.
За переработку и переключение импульсов отвечает вставочный нейрон. Функции данного типа клеток заключаются в получении и обработке информации от чувствительных и двигательных нейронов, между которыми они находятся. Более того, вставочные (или промежуточные) нейроны занимают 90 % центральной нервной системы человека, а также в больших количествах находятся во всех сферах головного и спинного мозга.
Строение промежуточных нейронов
Вставочный нейрон состоит из тела, аксона и дендритов. Каждая часть имеет свои специфические функции и отвечает за определенное действие. В его теле содержатся все компоненты, из которых созданы клеточные структуры. Важная роль этой части нейрона заключается в генерировании нервных импульсов и выполнении трофической функции. Продолговатый отросток, который несет сигнал от тела клетки, называется аксоном. Он делится на два типа: миелиновый и безмиелиновый. На конце аксона находятся различные синапсы. Третья составляющая нейронов — дендриты. Они являются короткими отростками, которые разветвляются в разные стороны. Их функция заключается в доставке импульсов к телу нейрона, что обеспечивает связь между различными видами нейронов центральной нервной системы.
Сфера воздействия
Что определяет область влияния вставочного нейрона? В первую очередь его собственное строение. В основном у клеток данного типа имеются аксоны, синапсы которых оканчиваются на нейронах этого же центра, что обеспечивает их объединение. Некоторые промежуточные нейроны активируются другими, из иных центров, а затем доставляют информацию в свой нейронный центр. Такие действия усиливают воздействие сигнала, который повторяется в параллельных путях, тем самым удлиняя срок хранения информационных данных в центре. В результате место, куда был доставлен сигнал, увеличивает надежность влияния на исполнительную структуру. Иные вставочные нейроны могут получать активацию от соединений двигательных «братьев» из своего центра. Потом они становятся передатчиками информации назад в свой центр, чем создают обратные связи. Таким образом, вставочный нейрон играет важную роль в образование особых замкнутых сетей, которые продлевают срок хранения информации в нервном центре.
Возбуждающий тип промежуточных нейронов
Вставочные нейроны делятся на два типа: возбуждающие и тормозные. При активации первых облегчается передача данных из одной нейронной группы в другую. Такую задачу выполняют именно «медленные» нейроны, которые имеют способность к длительной активации. Они передают сигналы на протяжении довольно длительного времени. Параллельно с этими действиями промежуточные нейроны активизируют и своих «быстрых» «коллег». Когда усиливается активность «медленных» нейронов, то уменьшается время реакции «быстрых». Одновременно с этим последние несколько замедляют работу «медленных».
Тормозной тип промежуточных нейронов
Вставочный нейрон тормозного типа приходит в активное состояние за счет прямых сигналов, которые поступают в их центр или исходят из него. Данное действие происходит путем обратных связей. Прямое возбуждение данного типа вставочных нейронов является характерным для промежуточных центров чувствительных путей спинного мозга. А в двигательных центрах коры головного мозга происходит активизация вставочных нейронов благодаря обратным связям.
Роль вставочных нейронов в работе спинного мозга
В работе спинного мозга человека важная роль отводится проводящим путям, которые расположены снаружи от пучков, исполняющих проводниковую функцию. Именно по этим дорожкам и передвигаются импульсы, которые посылает вставочный и чувствительный нейроны. Сигналы проходят вверх и вниз по этим путям, передавая различную информацию в соответствующие части мозга. Вставочные нейроны спинного мозга находятся в промежуточно-медиальном ядре, которое, в свою очередь, расположено в заднем роге. Промежуточные нейроны являются важной передней частью спинно-мозжечкового пути. На обратной стороне рога спинного мозга расположены волокна, состоящие из вставочных нейронов. Они образуют боковой спинно-таламический путь, который выполняет особую функцию. Он является проводником, то есть передает сигналы о болевых ощущениях и температурной чувствительности сначала в промежуточный мозг, а потом и в саму кору головного мозга.
Дополнительная информация о вставочных нейронах
В нервной системе человека вставочные нейроны выполняют особую и крайне важную функцию. Они связывают между собой различные группы нервных клеток, передают сигнал из головного мозга в спинной. Хотя именно этот тип является наиболее мелким по размерам. По форме вставочные нейроны напоминают звезду. Основное количество данных элементов располагается в сером веществе головного мозга, а их отростки не выступают за пределы центральной нервной системы человека.
Интернейрон — это… Что такое Интернейрон?
интернейрон — интернейрон … Орфографический словарь-справочник
Центральная нервная система — основная часть нервной системы животных и человека, состоящая из скопления нервных клеток (нейронов) и их отростков; представлена у беспозвоночных системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных и… … Большая советская энциклопедия
Клетка Реншоу — Клетки Реншоу (КР) (англ. Renshaw cells) тормозные вставочные нейроны, расположенные в передних рогах спинного мозга, несколько дорсальнее и медиальнее, чем мотонейроны (МН). Это небольшие клетки. Диаметр тела клетки Реншоу равен 10 20 … Википедия
Коленный рефлекс — … Википедия
Клетки реншоу — (КР) (англ. Renshaw cells) это тормозные вставочные нейроны, расположенные в передних рогах спинного мозга, несколько дорзальнее и медиальнее, чем мотонейроны (МН). Это небольшие клетки. Диаметр тела клетки Реншоу равен 10 20 мкм, дендриты имеют … Википедия
Нейроны — Не следует путать с нейтроном. Пирамидальные ячейки нейронов в коре головного мозга мыши Нейрон (нервная клетка) – это структурно функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко специализирована и по структуре… … Википедия
Синапс — Основные элементы синапса Синапс[1] (греч … Википедия
Аксон — 9 дневной мыши Аксон (греч. ἀξον ось) нейрит, осевой цилиндр, отросток нервной клетки, по кото … Википедия
Нейроглия — Нейроглия, или просто глия (от др. греч. νεῦρον «волокно, нерв» и γλία «клей») совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Термин ввёл в 1846 году Рудольф Вирхов[1]. Глиальные клетки … Википедия
Астроциты — Астроциты клетки нейроглии. Совокупность астроцитов называется астроглией. Содержание 1 Функции астроглии 2 Виды астроцитов … Википедия
Нейроны для «чайников» — vechnayamolodost.ru
Нейроны – особая группа клеток организма, распространяющих информацию по всему телу. Используя электрические и химические сигналы, они помогают мозгу координировать все жизненно необходимые функции.
Если упростить, задачи нервной системы – собрать сигналы, поступающие из окружающей среды или из организма, оценить ситуацию, принять решение, как на них отреагировать (например, изменить частоту сердечных сокращений), а также подумать о происходящем и запомнить это. Основной инструмент для выполнения этих задач – нейроны, сплетенные по всему организму сложной сетью.
По средним оценкам, количество нейронов в головном мозге составляет 86 миллиардов, каждый из них связан еще с 1000 нейронов. Это создает невероятную сеть взаимодействия. Нейрон – основная единица нервной системы.
Нейроны (нервные клетки) составляют около 10% мозга, остальное – глиальные клетки и астроциты, функция которых заключается в поддержании и питании нейронов.
Как выглядит нейрон?
В строении нейрона можно выделить три части:
· Тело нейрона (сома) – получает информацию. Содержит ядро клетки.
· Дендриты – короткие отростки, принимающие информацию от других нейронов.
· Аксон – длинный отросток, несет информацию от тела нейрона в другие клетки. Чаще всего аксон оканчивается синапсом (контактом) с дендритами других нейронов.
Схема строения нейрона (здесь и далее рисунки из Википедии).
Дендриты и аксоны называют нервными волокнами.
Аксоны сильно варьируют по длине, от нескольких миллиметров до метра и более. Самыми длинными являются аксоны спинномозговых ганглиев.
Типы нейронов
Классификацию нейронов можно провести по нескольким параметрам, например, по строению или выполняемой функции.
Типы нейронов в зависимости от функции:
· Эфферентные (двигательные) нейроны – несут информацию от центральной нервной системы (головного и спинного мозга) к клеткам других частей тела.
· Афферентные (чувствительные) нейроны – собирают информацию от всего организма и несут ее в центральную нервную систему.
· Вставочные нейроны – передают информацию между нейронами, чаще в пределах центральной нервной системы.
Как нейроны передают информацию?
Нейрон, получая информацию от других клеток, накапливает ее до тех пор, пока она не превысит определенный порог. После этого нейрон посылает по аксону электрический импульс – потенциал действия.
Потенциал действия формируется движением электрически заряженных частиц через мембрану аксона.
В состоянии покоя электрический заряд внутри нейрона отрицательный относительно окружающей его межклеточной жидкости. Эта разница называется мембранным потенциалом. Обычно он составляет 70 милливольт.
Когда тело нейрона получает достаточно заряда, и он «выстреливает», в соседнем участке аксона происходит деполяризация – мембранный потенциал быстро растет, а затем падает примерно за 1/1000 секунды. Этот процесс запускает деполяризацию соседнего участка аксона, и так далее, пока импульс не пройдет по всей длине аксона. После процесса деполяризации наступает гиперполяризация – кратковременное состояние отдыха, в этот момент передача импульса невозможна.
Потенциал действия чаще всего генерируют ионы калия (К+) и натрия (Na+), которые по ионным каналам перемещаются из межклеточной жидкости внутрь клетки и обратно, меняя заряд нейрона и делая его сначала положительным, а затем снижая его.
Потенциал действия обеспечивает работу клетки по принципу «все или ничего», то есть импульс или передается, или нет. Слабые сигналы будут накапливаться в теле нейрона до тех пор, пока их заряда не будет достаточно для передачи по отросткам.
Миелин
Миелин – это белое густое вещество, покрывающее большинство аксонов. Это покрытие обеспечивает электроизоляцию волокна и повышает скорость прохождения импульса по нему .
Миелинизированное волокно в сравнении с немиелинизированным.
Миелин вырабатывается шванновскими клетками на периферии и олигодендроцитами в центральной нервной системе. По ходу волокна миелиновая оболочка прерывается – это перехваты Ранвье. Потенциал действия перемещается от перехвата к перехвату, что обеспечивает быструю передачу импульса.
Такое распространенное и серьезное заболевание, как рассеянный склероз, вызвано разрушением миелиновой оболочки.
Как работают синапсы
Нейроны и ткани, которым они передают импульс, физически не соприкасаются, между клетками всегда существует пространство – синапс.
В зависимости от способа передачи информации, синапсы могут быть химическими и электрическими.
Химический синапсПосле того как сигнал, передвигаясь по отростку нейрона, достигает синапса, происходит высвобождение химических веществ – нейромедиаторов (нейротрансмиттеров) в пространство между двумя нейронами. Это пространство называют синаптической щелью.
Схема строения химического синапса.
Нейромедиатор из передающего (пресинаптического) нейрона, попадая в синаптическую щель, взаимодействует с рецепторами на мембране принимающего (постсинаптического) нейрона, запуская целую цепь процессов.
Виды химических синапсов:
· глютаматэргический – медиатором является глютаминовая кислота, обладает возбуждающим эффектом на синапс;
· ГАМК-эргический – медиатором является гамма-аминомасляная кислота (ГАМК), обладает тормозящим эффектом на синапс;
· холинергический – медиатором является ацетилхолин, осуществляет нервно-мышечную передачу информации;
· адренергический – медиатором является адреналин.
Электрические синапсы
Электрические синапсы встречаются реже, распространены в центральной нервной системе. Клетки сообщаются посредством особых белковых каналов. Пресинаптическая и постсинаптическая мембраны в электрических синапсах расположены близко друг к другу, поэтому импульс способен проходить непосредственно от клетки к клетке.
Скорость передачи импульса по электрическим синапсам гораздо выше, чем по химическим, поэтому они расположены преимущественно в тех отделах, где необходима быстрая реакция, например, отвечающих за защитные рефлексы.
Еще одно отличие двух типов синапсов в направлении передачи информации: если химические синапсы могут передавать импульс только в одном направлении, то электрические в этом смысле универсальны.
Заключение
Нейроны – это, пожалуй, самые необычные клетки организма. Каждое действие, которое осуществляет тело человека, обеспечивается работой нейронов. Сложная нейронная сеть формирует личность и сознание. Они отвечают как за самые примитивные рефлексы, так и за самые сложные процессы, связанные с мышлением.
Аминат Аджиева, портал «Вечная молодость» http://vechnayamolodost.ru по материалам Medical News Today: Neurons: The basics.
Типы нейронов. Сенсорные (чувствительные), моторные (двигательные), промежуточные (вставочные) нейроны
Вообще, в зависимости возложенных на нейроны задач и обязанностей, они делятся на три категории:
— Сенсорные (чувствительные) нейроны принимают и передают импульсы от рецепторов «в центр», т.е. центральную нервную систему. Причем сами рецепторы — это специально обученные клетки органов чувств, мышц, кожи и суставов умеющие обнаруживать физические или химические изменения внутри и снаружи нашего организма, преобразовывать их в импульсы и радостно передавать их сенсорным нейронам. Таким образом, сигналы идут от периферии к центру.
Следующий тип:
— Моторные (двигательные) нейроны, которые урча, фырча и бибикая, несут сигналы, выходящие из головного или спинного мозга, к исполнительным органам, коими являются мышцы, железы и т.д. Ага, значит, сигналы идут от центра к периферии.
Ну а промежуточные (вставочные) нейроны, попросту говоря, являются «удлинителями», т.е. получают сигналы от сенсорных нейронов и посылают эти импульсы дальше к другим промежуточным нейронам, ну или сразу к моторным нейронам.
В общем и целом вот что получается: у сенсорных нейронов дендриты соединены с рецепторами, а аксоны — с другими нейронами (вставочными). У двигательных нейронов наоборот, дендриты соединены с другими нейронами (вставочными), а аксоны — с каким-нибудь эффектором, т.е. стимулятором сокращения какой-нибудь мышцы или секреции железы. Ну а, соответственно, у вставочных нейронов и дендриты и аксоны соединяются с другими нейронами.
Получается что самый простой путь, по которому может идти нервный импульс, будет состоять из трех нейронов: одного сенсорного, одного вставочного и одного моторного.
Ага, а давайте теперь вспомним дядьку — очень «нервного патолога», с ехидной улыбкой стучащего своим «волшебным» молоточком по колену. Знакомо? Вот, это и есть простейший рефлекс: когда он ударяет по коленному сухожилию, прикрепленная к нему мышца растягивается и сигнал от находящихся в ней чувствительных клеток (рецепторов) передается по сенсорным нейронам в спинной мозг. А уже в нем сенсорные нейроны контактируют либо через вставочные, либо непосредственно с моторными нейронами, которые в ответ посылают импульсы назад в ту же самую мышцу, заставляя ее сокращаться, а ногу — распрямляться.
Сам же спинной мозг удобно примостился внутри нашего позвоночника. Он мягкий и ранимый, потому и прячется в позвонках. Спинной мозг всего 40-45 сантиметров в длину, с мизинец толщиной (около 8 мм) и весит каких-то 30 грамм! Но, несмотря на всю свою тщедушность, спинной мозг является управляющим центром сложной сети нервов, раскинутой по телу. Практически как центр управлениями полетами! 🙂 Без него ни опорно-двигательный аппарат, ни основные жизненные органы ну никак не могут действовать и работать.
Свое начало спинной мозг берет на уровне края затылочного отверстия черепа, а заканчивается на уровне первого-второго поясничных позвонков. А вот уже ниже спинного мозга в позвоночном канале находится такой густой пучок нервных корешков, прикольно именуемый конским хвостом, видимо за сходство с ним. Так вот, конский хвост – это продолжение нервов, выходящих из спинного мозга. Они отвечают за иннервацию нижних конечностей и органов таза, т.е. передают сигналы от спинного мозга к ним.
Спинной мозг окружен тремя оболочками: мягкой, паутинной и твердой. А пространство между мягкой и паутинной оболочками заполнено еще и большим количеством спинномозговой жидкости. Через межпозвоночные отверстия от спинного мозга отходят спинномозговые нервы: 8 пар шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 или 2 копчиковых. Почему пар? Да потому, что спинномозговой нерв выходит двумя корешками: задним (чувствительным) и передним (двигательным), соединенными в один ствол. Так вот, каждая такая пара контролирует определенную часть тела. Т.е., например, если вы нечаянно схватились за горячую кастрюлю (не дай бог! Тьфу-тьфу-тьфу!), то в окончаниях чувствительного нерва тут же возникает болевой сигнал, сразу же поступающий в спинной мозг, и уже оттуда — в парный двигательный нерв, который и передает приказ: «Ахтунг-ахтунг! Немедленно убрать руку!» Причем, поверьте, это происходит очень быстро — еще до того, как головной мозг зарегистрирует болевой импульс. В итоге, вы успеваете отдернуть руку от кастрюли еще до того, как почувствуете боль. Конечно же, такая реакция спасает нас от тяжелых ожогов или других повреждений.
Вообще, практически все наши автоматические и рефлекторные действия контролируются спинным мозгом, ну за исключением тех, за которыми следит сам головной мозг. Ну, вот, например: мы воспринимаем увиденное с помощью глазного нерва идущего в головной мозг, и в то же время обращаем свой взор в разные стороны при помощи глазных мышц, которые управляются уже спинным мозгом. Да и плачем мы то же по приказу спинного мозга, который «заведует» слезными железами.
Можно сказать, что наши сознательные действия идут от головного мозга, но как только эти действия мы начинаем выполнять уже автоматически и рефлекторно — они передаются в ведение спинного мозга. Так что, когда мы только учимся что-то делать, то, конечно же, сознательно обдумываем и продумываем и осмысливаем каждое движение, а значит, используем головной мозг, но со временем мы уже можем делать это автоматически, и это значит, что головной мозг передает «бразды правления» этим действием спинному, просто ему уже стало скучно и неинтересно….потому как, наш головной мозг очень пытливый, любознательный и любит учиться!
Ну вот, пришло и нам время полюбопытствовать……
Читаем про головной могз! >>
12 * ЭФЕКТОРЫ
4 . Заболевания нервной системы. основные из них:Болезнь Альцгеймера. Это дегенеративное неврологическое заболевание, прогрессирующее и необратимое, который может возникнуть в возрасте 50 лет. Первые симптомы — тяжелая память. потеря и путаница в речи. Это вызывает слабоумие и приводит к неспособность воспринимать происходящее в окружающей среде. Это также может называться « старческое слабоумие », поскольку термин « старческое слабоумие» слабоумие «используется только для слабоумия, которое появляется после возраст 80.
Нервная анорексия. Это постоянное расстройство пищевого поведения, заключающееся в нежелании есть. Обычно это начинается с панического ощущения при мысли о том, что лишний вес. Как ни парадоксально это заболевание сильнее, если у человека сильная воля и требовательность к себе — две характеристики, которые обычно очень положительно для другой человеческой деятельности.
Аутизм. Is склонность уходить во внутренний мир и терять интерес к реальность, сопровождаемая неспособностью к общению.Может быть симптомом шизофрении.
Булимия. Is чрезмерное потребление пищи в ответ на состояние повышенной тревожности. Избыток веса вызывает беспокойство пациента, тем самым усиливая беспокойство и, таким образом, прием, замыкая замкнутый круг.
Зависимость . Это связано с необходимостью регулярного употребления определенного вещества, для того, чтобы восстановить нормальное функционирование организма. Появляется после продолжения использование определенного препарата или лекарства.Это может быть физических (если нет употребляется, может вызвать спазмы, дрожь и потоотделение, называемое отменой синдром ) или психический (есть дискомфорт, но нет реальных симптомов вывода). В зависимости от продукта алкоголизм , курение и наркомании можно выделить . Самые распространенные препараты и их эффекты:
- Кокаин (стимулирующий препарат, вызывающий эйфорию и гиперактивность с последующим период депрессии с раздражительностью и тревогой),
- Героин (обезболивающий препарат, предотвращающий восприятие боли и помогающий забыть проблемы),
- Марихуана, гашиш и ЛСД (галлюциногенные препараты, которые изменяют восприятие реальности, которую мы видим и слышать и вызывать нереалистичные изображения и звуки).
Депрессия. Это постоянное состояние глубокой печали, часто сопровождающееся тревогой, общая незаинтересованность, чувство вины и даже стремление к самоповреждению. Может быть экзогенным , что естественно и вызвано большим несчастьем и ослабнет через несколько дней, или эндогенно , что вызвано изменение производства или реабсорбции определенных нейромедиаторов, и требует нескольких месяцев фармакологического лечения.
Эпилепсия. Это нарушение функции головного мозга, вызывающее кратковременные постоянные приступы мышечных судорог, потери сознания и сенсорных расстройств.
Менингит. Это инфекция мозговых оболочек, вызывающая высокую температуру, скованность шеи и острая боль.
Мигрень. Это болезнь включает повторяющиеся приступы очень сильных головных болей, которые обычно располагается в лобной и височной области.Часто сопровождается светобоязнью, потребностью в тишине и иногда рвотой.
Рассеянный склероз. Заключается в разрушении миелиновой оболочки, покрывающей аксоны. нейронов из-за ошибки собственной иммунной системы пациента (аутоиммунная болезнь). Из-за этого пластины разбросаны по всей ЦНС. Первые симптомы: упадок сил, потеря чувствительности и трудности. в координации движений.
Паранойя. Это постоянное бредовое чувство преследования со стороны окружающих нас. Парадоксально сопровождается большой ясностью и тонкостью восприятия и проанализировать все аспекты жизни, которые влияют на других людей.
Болезнь Паркинсона. Это изменение серых нуклонов, которые существуют в основании мозг, вызванный пока неизвестными причинами. Ранние симптомы включают: дрожание рук, скованность мышц и замедленность движений.Это может произойти от среднего возраста.
Полиомиелит. Это заразная вирусная инфекция, вызывающая паралич и атрофию. конечностей.
Шизофрения. Это тяжелое расстройство личности, мыслей, восприятия эмоции и восприятие окружающего мира. Это может вызвать изолированную агрессивную реакции.
Стресс. Это состояние острого стресса из-за необходимости быстро реагировать на многочисленные раздражители, которые воспринимаются как угрожающие или агрессивные.
Формирование вставочных невромастов. (A) Нормальный эмбрион при 4 dpf ….
Контекст 1
… в конце своего эмбрионального развития PLL рыбок данио обычно состоит из пяти пар боковых и двух-трех пар терминальных невромастов, а также пара дорсальных невромастов близко к первому сомиту (рис. 4А). Этот паттерн, по-видимому, чрезвычайно сохранен среди костистых насекомых (Pichon and Ghysen 2004). Наблюдается не только у относительно базальных остариофизов (напр.g., рыбки данио и слепая тетра, Astyanax fasciatus), но также в группах, которые демонстрируют многие производные признаки, такие как плевронектиды (камбалы; Pichon and Ghysen 2004) и скомбриды (tu- …
Context 2
.. Издательство Spring Harbor Laboratory Press, 24 сентября 2008 г. — Опубликовано genesdev.cshlp.org Загрузка с интеркалярного невромаста? Этот вопрос был решен Грантом и др. (2005) с помощью серии экспериментов, гениально сочетающих хирургию и генетику. показали, что удаление ганглия PLL посредством генетической абляции (рис.4D) или физическая экстирпация (рис. 4E) не влияет на миграцию зачатка или отложение ранних невромастов, но приводит к образованию огромного избытка невромастов на 4 dpf (рис. 4B). Оказывается, что сверхкомплектные невромасты представляют собой сформированные ранее, инкалярные невромасты, происходящие от INC (Grant et al. 2005; …
Context 3
… Press 24 сентября 2008 г. — Опубликовано genesdev. cshlp.org Загружено с интеркалярного невромаста? Этот вопрос был решен Grant et al.(2005) с помощью серии экспериментов, гениально сочетающих хирургию и генетику. Они показали, что удаление ганглия PLL посредством генетической абляции (рис. 4D) или физической экстирпации (рис. 4E) не влияет на миграцию зачатка или отложение ранних невромастов, но приводит к формированию огромный избыток невромастов на 4 dpf (рис. 4B). Сверхкомплектные невромасты оказываются сформированными раньше, происходящими от INC, интеркалярными невромастами (Grant et al. 2005; Lopez-Schier and Hudspeth 2005)….
Context 4
… экспериментов, изобретательно сочетающих хирургию и генетику. Они показали, что удаление ганглия PLL посредством генетической абляции (рис. 4D) или физической экстирпации (рис. 4E) не влияет на миграцию зачатка или отложение ранних невромастов, но приводит к формированию огромный избыток невромастов на 4 dpf (рис. 4B). Оказывается, что сверхкомплектные невромасты представляют собой ранее сформированные интеркалярные невромасты, происходящие из INC (Grant et al.2005; Lopez-Schier and Hudspeth 2005). Очевидной интерпретацией этих данных было бы то, что сенсорные нейроны оказывают тормозящее влияние на формирование новых органов чувств — довольно парадоксальный вывод — …
Контекст 5
… ключ к решению проблемы. этот парадокс исходит из мутационного анализа. Были исследованы три мутанта, представляющие сверхкомплектные невромасты. Один из них имеет типичный дефект нервного гребня и лишен большинства глиальных клеток (рис.4F; Lopez-Schier and Hudspeth 2005), тогда как у двух других оказывается дефицит глиальных клеток вдоль нерва PLL, но не вокруг ганглия PLL (Grant et al. 2005). Таким образом, глиальные клетки, по-видимому, являются основным фактором, контролирующим формирование интеркалярных нейромастов. Этот момент был продемонстрирован трансплантацией дикого типа …
Context 6
… вентральная миграция INC отделяет их от PLL-нерва и его глии и тем самым снимает ингибирование.Однако формирование интеркалярных нейромастов продолжается в течение длительного периода времени, и через 10 недель все еще остаются позиции, в которых локальное утолщение, которое предвещает формирование межкалярного невромаста, только начинается (рис. 4G, звездочки). или все еще отсутствует (рис. 4G, вопросительные знаки). В этом возрасте весь след INC переместился на значительное расстояние от горизонтальной миосепты, подразумевая, что ингибирующий эффект глиальных клеток должен быть опосредован секретируемой молекулой, действующей в течение длительного времени…
Контекст 7
… их от нерва PLL и его глии, и тем самым снимает торможение. Однако формирование интеркалярных нейромастов продолжается в течение длительного периода времени, и через 10 недель все еще остаются позиции, в которых локальное утолщение, которое предвещает формирование межкалярного невромаста, только начинается (рис. 4G, звездочки). или все еще отсутствует (рис. 4G, вопросительные знаки). В этом возрасте весь след INC переместился на значительное расстояние от горизонтальной миосепты, подразумевая, что ингибирующий эффект глиальных клеток должен быть опосредован секретируемой молекулой, действующей в течение длительного времени…
Контекст 8
… окончательно дифференцировать. Как отмечает Whitfield (2005), эта гипотеза д. Предсказывать, что интеркалярные невромасты, продуцируемые с помощью primI, д. Обнаруживать поляризацию D / V, поскольку их вентральная миграция предшествует их дифференцировке. Однако это не так: все первичные невромасты, ранние или поздние (интеркалярные), обнаруживают A / P поляризацию (рис. 4G; A. Ghysen, неопубликовано). Волосковые клетки внутри невромастов также проявляют поляризацию A / P как в эмбриональных, так и в интеркалярных нейромастах, которые зависят от primI (T.Пиотровский, чел. комм.). Реципрокная ситуация обеспечивается спинной линией. Достигнув дорсальной срединной линии, primD мигрирует вперед-назад вдоль …
, определение интеркаляра в The Free Dictionary
in · ter · ca · lar · y
(ĭn-tûr′kə-lĕr′ē, ĭn′tər-kăl′ə-rē) прил. 1.а. Вставляется в календарь, чтобы календарный год соответствовал солнечному году. Используется день или месяц.
б. Указание такого дня или месяца.Б / у год.
2. Вставляется между другими элементами или частями; интерполированный.
[латинское intercalārius, intercalāris, от intercalāre, до интеркалята ; см. , вставка .]
Словарь английского языка American Heritage®, пятое издание. Авторские права © 2016 Издательская компания Houghton Mifflin Harcourt. Опубликовано Houghton Mifflin Harcourt Publishing Company. Все права защищены.
интеркалярный
(ɪnˈtɜːkələrɪ) илиинтеркалярный
adj1. (Часы) (дня, месяца и т. Д.), Вставленные в календарь
2. (Часы) (определенного года) с вставкой одного или нескольких дней
3. вставленные, введенные или интерполированные
4. (Ботаника) ботаника растет между верхними ветвями и нижними ветвями или прицветниками на стебле
[C17: от латинского intercalārius; см. Вставку]
внутренне adv
Словарь английского языка Коллинза — полный и несокращенный, 12-е издание 2014 г. © HarperCollins Publishers 1991, 1994, 1998, 2000, 2003, 2006, 2007, 2009, 2011, 2014
• ter • ca • lar • y
(ɪnˈtɜr kəˌlɛr i, ˌɪn tərˈkæl ə ri)прил.
1. интерполированный; вставил.
2. вставлено или интерполировано в календаре как дополнительный день или месяц.
3. (года) с таким вставленным днем, месяцем и т. Д.
в • терьца • лар • лы, нар.
Random House Словарь колледжа Кернермана Вебстера © 2010 K Dictionaries Ltd. Авторские права 2005, 1997, 1991 принадлежат Random House, Inc. Все права защищены.
вставкавставляется в календарь, как двадцать девятый день февраля в високосном году.- интеркаляция, п. — интеркаляционный, прил.
См. Также: Календарь-Ologies & -Isms. Copyright 2008 The Gale Group, Inc. Все права защищены.
Сегментарно-проводниковый аппарат спинного мозга
Сегментарно-проводниковый аппарат спинного мозга
Сегментарный аппарат спинного мозга — это совокупность функционально связанных между собой структур спинного мозга, обеспечивающих выполнение безусловных рефлексов, морфологической основой которых являются простые рефлекторные дуги.
Сегментарный аппарат спинного мозга включает следующие структуры.
1. Заднекорошковые волокна — центральные отростки сенсорных узлов спинномозговых нервов, расположенные в корешковой зоне и заканчивающиеся синаптическими окончаниями на вставочных нейронах.
2. Вставные нейроны, роль которых играют разрозненные клетки, клетки студенистого вещества, губчатые и концевые зоны. Разбросанные клетки обеспечивают передачу нервного импульса на уровне своего сегмента к нейронам собственных ядер передних рогов спинного мозга своей стороны.Ячейки концевой и губчатой зон передают информацию разрозненным ячейкам одного или двух верхних и нижних сегментов. Клетки студенистого вещества передают информацию разбросанным клеткам трех-четырех высших и низших сегментов. Таким образом, распространение информации с сильными стимулами происходит по шести-семи сегментам (рис. 2.7).
3. Задний, латеральный и передний собственные пучки спинного мозга представляют собой аксоны вставочных нейронов студенистого вещества, губчатую и терминальную зоны, разделяющиеся на восходящие и нисходящие ветви и доходящие до верхнего и нижнего сегментов.Собственные пучки спинного мозга на поперечном срезе имеют вид узкой каймы белого вещества, которая непосредственно прикрепляется к серому веществу. Аксоны интеркалярных нейронов имеют многочисленные коллатерали, которые оканчиваются на нейронах ядер передних рогов спинного мозга верхних и нижних сегментов.
4. Крупные мультиполярные нейроны собственных ядер передних рогов и начальной части их аксонов, которые составляют передние корешковые волокна до того, как они покинут спинной мозг.
Остальные элементы рефлекторных дуг безусловных рефлексов относятся к периферической нервной системе (передние и задние корешки, чувствительные узлы спинномозговых нервов, спинномозговые нервы и их ветви).
Большинство вставочных нейронов сегментарного аппарата направляют свои аксоны к эффекторным нейронам своей стороны, однако есть нейроны, аксоны которых следуют за противоположной половиной спинного мозга и вызывают сокращение мускулатуры противоположной половины тела.
Когда нервный импульс возникает даже в одном рецепторе, в нервный процесс вовлекаются десятки (и сотни) ассоциативных нейронов, каждый из которых вызывает возбуждение сотен или тысяч эффекторных нейронов в ядрах передних рогов. Аксоны каждого из последних вызывают сокращение более 100 мышечных волокон. Таким образом, в результате раздражения одного рецептора сокращаются не только отдельные мышцы, но и несколько групп мышц. Объясняется это лавинообразным нарастанием нервных импульсов в сегментарном аппарате спинного мозга.
Таким образом, сегментированный аппарат спинного мозга включает в себя все структуры серого вещества, за исключением ядер вставочных нейронов (относящихся к проводящему аппарату), анатомически и функционально связанных с ним собственных пучков спинного мозга и соответствующих этим сегментам. задних и передних корешковых волокон.Рек. 2.7. Сегментарный аппарат спинного мозга:
1 — собственно связки; 2 — рецептор; 3 — чувствительный узел спинномозгового нерва; 4 — оси студенистого вещества; 5 — аксоны губчатой зоны; 6 — аксоны пограничной зоны; 7 — третий вышележащий (нижний) сегмент; 8 — второй высший (нижний) сегмент; 9 — первый вышележащий (нижний) сегмент; 10 — вставочные нейроны губчатого, пограничного и студенистого вещества; 11 — разрозненные ячейки
Проводник спинного мозга обеспечивает двустороннюю связь между спинным мозгом и центрами интеграции головного мозга, которые расположены в коре мозжечка, верхних бугорках молочных желез, среднем мозге и коре головного мозга.Интеграционный центр вегетативного отдела нервной системы находится в промежуточном мозге.
Исследовательский аппарат спинного мозга представлен афферентными (восходящими) и эфферентными (нисходящими) путями. Афферентные пути начинаются от нейронов сенсорных узлов спинномозговых нервов и проводят нервные импульсы в центры интеграции головного мозга. По афферентным путям проходят интеркалярные нейроны, скопления которых образуют коммуникационные нервные центры.Эфферентные нервные пути образованы аксонами нейронов ядер мозга. Эфферентный путь заканчивается на нейронах ядер передних рогов спинного мозга или двигательных ядрах черепных нервов.
Таким образом, афферентные и эфферентные нервные пути (тракты) и коммуникационные центры, расположенные вдоль пути афферентных путей (собственное ядро рога, грудное и медиально-медиальное ядро), называются интеграционным аппаратом в спинном мозге.
Роль белков семейства CPEB в функции нервной системы в норме и патологии | Cell & Bioscience
Ким Дж. Х., Рихтер Дж. Д. Противоположные активности полимеразы-деаденилазы регулируют цитоплазматическое полиаденилирование. Mol Cell. 2006; 24: 173–83. https://doi.org/10.1016/j.molcel.2006.08.016.
CAS Статья PubMed Google Scholar
Cao Q, Richter JD. Растворение комплекса маскин-eIF4E за счет цитоплазматического полиаденилирования и поли (A) -связывающего белка контролирует трансляцию мРНК циклина B1 и созревание ооцитов. EMBO J. 2002; 21: 3852–62.https://doi.org/10.1093/emboj/cdf353.
CAS Статья PubMed PubMed Central Google Scholar
Удагава Т., Свангер С.А., Такеучи К., Ким Дж. Х., Налавади В., Шин Дж. И др. Двунаправленный контроль трансляции мРНК и синаптической пластичности цитоплазматическим комплексом полиаденилирования. Mol Cell. 2012; 47: 253–66. https://doi.org/10.1016/j.molcel.2012.05.016.
CAS Статья PubMed PubMed Central Google Scholar
Келеман К., Крюттнер С., Алениус М., Диксон Б.Дж. Функция белка Orb2 CPEB дрозофилы в долговременной памяти ухаживания. Nat Neurosci. 2007; 10: 1587–93. https://doi.org/10.1038/nn1996.
CAS Статья PubMed Google Scholar
Mendez R, Richter JD. Трансляционный контроль со стороны CPEB: средство для достижения цели. Nat Rev Mol Cell Biol. 2001; 2: 521–9. https://doi.org/10.1038/35080081.
CAS Статья PubMed Google Scholar
Lantz V, Ambrosio L, Schedl P. Предполагается, что ген orb дрозофилы кодирует специфичные для пола РНК-связывающие белки зародышевой линии и имеет локализованные транскрипты в яичниках и ранних эмбрионах. Разработка. 1992; 115: 75–88.
CAS PubMed Google Scholar
Пай Т.П., Чен С.К., Линь Х.Х., Чин А.Л., Лай Дж.Си., Ли П.Т. и др. Белок Orb дрозофилы в двух выходных нейронах грибовидного тела необходим для формирования долговременной памяти. Proc Natl Acad Sci U S A.2013; 110: 7898–903. https://doi.org/10.1073/pnas.1216336110.
Артикул PubMed PubMed Central Google Scholar
Хуанг И-С, Кан М.-К, Лин С-Л, Рихтер Дж. Д.. CPEB3 и CPEB4 в нейронах: анализ специфичности связывания РНК и контроль трансляции мРНК GluR2 рецептора AMPA. EMBO J. 2006; 25: 4865–76. https://doi.org/10.1038/sj.emboj.7601322.
CAS Статья PubMed PubMed Central Google Scholar
Piqué M, López JM, Foissac S, Guigó R, Méndez R. Комбинаторный код для CPE-опосредованного контроля трансляции. Клетка. 2008. 132: 434–48. https://doi.org/10.1016/j.cell.2007.12.038.
CAS Статья PubMed Google Scholar
Stepien BK, Oppitz C, Gerlach D, Dag U, Novatchkova M, Krüttner S, et al. Профили связывания РНК белков Orb и Orb2 CPEB дрозофилы. Proc Natl Acad Sci. 2016; 113: E7030–8. https: // doi.org / 10.1073 / pnas.1603715113.
CAS Статья PubMed Google Scholar
Charlesworth A, Meijer HA, De Moor CH. Факторы специфичности цитоплазматического полиаденилирования. Wiley Interdiscip Rev RNA. 2013; 4: 437–61. https://doi.org/10.1002/wrna.1171.
CAS Статья PubMed PubMed Central Google Scholar
Тай Дж., Ходжман Р., Рихтер Дж. Д..Контроль трансляции мРНК циклина B1 во время созревания ооцитов мыши. Dev Biol. 2000; 221: 1–9. https://doi.org/10.1006/dbio.2000.9669.
CAS Статья PubMed Google Scholar
Баркофф А.Ф., Диксон К.С., Грей Н.К., Виккенс М. Трансляционный контроль мРНК циклина B1 во время мейотического созревания: скоординированная репрессия и цитоплазматическое полиаденилирование. Dev Biol. 2000; 220: 97–109. https://doi.org/10.1006/dbio.2000.9613.
CAS Статья PubMed Google Scholar
Афроз Т., Скрисовская Л., Беллок Е., Гильен-Бойше Дж., Мендес Р., Аллен ФХТ. Механизм ловушки для мух обеспечивает специфичное для последовательности распознавание РНК белками CPEB. Genes Dev. 2014; 28: 1498–514. https://doi.org/10.1101/gad.241133.114.
CAS Статья PubMed PubMed Central Google Scholar
Novoa I, Gallego J, Ferreira PG, Mendez R.Прогрессирование митотического клеточного цикла регулируется CPEB1 и CPEB4-зависимым трансляционным контролем. Nat Cell Biol. 2010; 12: 447–56. https://doi.org/10.1038/ncb2046.
CAS Статья PubMed Google Scholar
Proudfoot NJ. Завершение сообщения: poly (A) сигнализирует тогда и сейчас. Genes Dev. 2011; 25: 1770–82. https://doi.org/10.1101/gad.17268411.
CAS Статья PubMed PubMed Central Google Scholar
Mendez R, Murthy KGK, Ryan K, Manley JL, Richter JD. Фосфорилирование CPEB с помощью Eg2 опосредует рекрутирование CPSF в активный цитоплазматический комплекс полиаденилирования. Mol Cell. 2000; 6: 1253–9. https://doi.org/10.1016/S1097-2765(00)00121-0.
CAS Статья PubMed Google Scholar
Mount SM, Salz HK. Факторы процессинга пре-мессенджера РНК в геноме дрозофилы. J Cell Biol. 2000 г. https://doi.org/10.1083/jcb.150.2.f37.
Артикул PubMed PubMed Central Google Scholar
Shorter J, Lindquist S. Прионы как адаптивные каналы памяти и наследования. Nat Rev Genet. 2005 г. https://doi.org/10.1038/nrg1616.
Артикул PubMed Google Scholar
Majumdar A, Cesario WC, White-Grindley E, Jiang H, Ren F, Khan MR, et al. Критическая роль амилоидоподобных олигомеров Drosophila Orb2 в сохранении памяти.Клетка. 2012; 148: 515–29. https://doi.org/10.1016/j.cell.2012.01.004.
CAS Статья PubMed Google Scholar
Крюттнер С., Степьен Б., Нордермер Дж. Н., Моммаас М.А., Мехтлер К., Диксон Б.Дж. и др. Drosophila CPEB Orb2A обеспечивает память независимо от его РНК-связывающего домена. Нейрон. 2012; 76: 383–95. https://doi.org/10.1016/j.neuron.2012.08.028.
CAS Статья PubMed Google Scholar
Hervas R, Rau MJ, Park Y, Zhang W., Murzin AG, Fitzpatrick JAJ и др. Крио-ЭМ структура нейронального функционального амилоида, участвующего в сохранении памяти у Drosophila. Наука. 2020; 367: 1230–4. https://doi.org/10.1126/science.aaw1114.
CAS Статья PubMed PubMed Central Google Scholar
Стефан Дж. С., Фиорити Л., Ламба Н., Колнаги Л., Карл К., Деркач И. Л. и др. Белок CPEB3 — это функциональный прион, который взаимодействует с актиновым цитоскелетом.Cell Rep. 2015; 11: 1772–85. https://doi.org/10.1016/j.celrep.2015.04.060.
CAS Статья PubMed Google Scholar
де Минго Д. Р., Лопес-Гарсия П., Эрвас Р., Лоуренс Д., Каррион-Васкес М. Молекулярные детерминанты расслоения жидкости и амилоидогенеза в CPEB3 человека. bioRxiv. 2020. https://doi.org/10.1101/2020.06.02.129783.
Артикул Google Scholar
Член парламента Джексона, Хьюитт Е.В. Почему функциональные амилоиды нетоксичны для человека? Биомолекулы. 2017; 7: 71. https://doi.org/10.3390/biom7040071.
CAS Статья PubMed Central Google Scholar
Хан М.Р., Ли Л., Перес-Санчес К., Сараф А., Флоренс Л., Слотер Б.Д. и др. Амилоидогенная олигомеризация превращает Drosophila Orb2 из репрессора трансляции в активатор. Клетка. 2015; 163: 1468–83. https://doi.org/10.1016/j.ячейка.2015.11.020.
CAS Статья PubMed Google Scholar
Охаши Р., Шиина Н. Каталогизация и отбор мРНК, локализованных в дендритах в нейронах и регулируемых РНК-связывающими белками в гранулах РНК. Биомолекулы. 2020; 10: 167. https://doi.org/10.3390/biom10020167.
CAS Статья PubMed Central Google Scholar
Kundel M, Jones KJ, Shin CY, Wells DG.Цитоплазматический белок, связывающий элемент полиаденилирования, регулирует нейротрофин-3-зависимую трансляцию мРНК β-катенина в развивающихся нейронах гиппокампа. J Neurosci. 2009. 29: 13630–9. https://doi.org/10.1523/JNEUROSCI.2910-08.2009.
CAS Статья PubMed PubMed Central Google Scholar
Sousa-Nunes R, Somers WG. Механизмы асимметричных делений предков в центральной нервной системе дрозофилы. Adv Exp Med Biol.2013; 786: 79–102. https://doi.org/10.1007/978-94-007-6621-1_6.
CAS Статья PubMed Google Scholar
Hafer N, Xu S, Bhat KM, Schedl P. Белок Orb2 CPEB дрозофилы имеет новый паттерн экспрессии и важен для асимметричного деления клеток и функции нервной системы. Генетика. 2011; 189: 907–21. https://doi.org/10.1534/genetics.110.123646.
CAS Статья PubMed PubMed Central Google Scholar
Сантана Э., Касас-Тинто С. Orb2 как модулятор Brat и их роль в нервно-мышечном соединении. J Neurogenet. 2017; 31: 181–8. https://doi.org/10.1080/01677063.2017.1393539.
CAS Статья PubMed Google Scholar
Oruganty-Das A, Ng T, Udagawa T., Goh ELK, Richter JD. Трансляционный контроль производства митохондриальной энергии опосредует морфогенез нейронов. Cell Metab. 2012; 16: 789–800. https://doi.org/10.1016 / j.cmet.2012.11.002.
CAS Статья PubMed PubMed Central Google Scholar
Alves-Sampaio A, Troca-Marín JA, Montesinos ML. NMDA-опосредованная регуляция дендритной локальной трансляции DSCAM потеряна в модели синдрома Дауна на мышах. J Neurosci. 2010. 30: 13537–48. https://doi.org/10.1523/JNEUROSCI.3457-10.2010.
CAS Статья PubMed PubMed Central Google Scholar
Kandel ER, Dudai Y, Mayford MR. Молекулярная и системная биология памяти. Клетка. 2014; 157: 163–86. https://doi.org/10.1016/j.cell.2014.03.001.
CAS Статья PubMed Google Scholar
Гланцман Д.Л. Общие механизмы синаптической пластичности у позвоночных и беспозвоночных. Curr Biol. 2010; 20: R31–6. https://doi.org/10.1016/j.cub.2009.10.023.
CAS Статья PubMed PubMed Central Google Scholar
Кляйн М, Кандел ER. Механизм модуляции кальциевого тока, лежащий в основе пресинаптического облегчения и поведенческой сенсибилизации при аплизии. Proc Natl Acad Sci USA. 1980; 77: 6912–6. https://doi.org/10.1073/pnas.77.11.6912.
CAS Статья PubMed Google Scholar
Martin KC, Casadio A, Zhu H, Yaping E, Rose JC, Chen M, et al. Специфичное для синапсов, долгосрочное облегчение сенсорной аплизии к моторным синапсам: функция локального синтеза белка в памяти.Клетка. 1997; 91: 927–38. https://doi.org/10.1016/S0092-8674(00)80484-5.
CAS Статья PubMed Google Scholar
Дойл М., Киблер Массачусетс. Механизмы дендритного транспорта мРНК и его роль в синаптической маркировке. EMBO J. 2011; 30: 3540–52. https://doi.org/10.1038/emboj.2011.278.
CAS Статья PubMed PubMed Central Google Scholar
Фрей У., Моррис РГМ. Синаптическая маркировка и долгосрочное потенцирование. Природа. 1997. 385: 533–6. https://doi.org/10.1038/385533a0.
CAS Статья PubMed Google Scholar
Судхакаран И.П., Рамасвами М. Консолидация долговременной памяти: роль РНК-связывающих белков с прионоподобными доменами. RNA Biol. 2017; 14: 568–86. https://doi.org/10.1080/15476286.2016.1244588.
Артикул PubMed Google Scholar
Томпа П., Фридрих П. Прионные белки как молекулы памяти: гипотеза. Неврология. 1998; 86: 1037–43. https://doi.org/10.1016/S0306-4522(98)00148-1.
CAS Статья PubMed Google Scholar
Хуанг Ю.С., Карсон Дж. Х., Барбарезе Е., Рихтер Дж. Д.. Облегчение транспорта дендритной мРНК с помощью CPEB. Genes Dev. 2003. 17: 638–53. https://doi.org/10.1101/gad.1053003.
CAS Статья PubMed PubMed Central Google Scholar
Л Да, Кристиан К, Лу Б. BDNF: ключевой регулятор для синтеза белка-зависимой БПЛ и долговременную память? Neurobiol Learn Mem. 2008; 89: 312–23. https://doi.org/10.1016/j.nlm.2007.08.018.
CAS Статья PubMed Google Scholar
Oe S, Yoneda Y. Цитоплазматические последовательности элементов полиаденилирования участвуют в дендритном нацеливании мРНК BDNF в нейронах гиппокампа. FEBS Lett. 2010; 584: 3424–30. https: // doi.org / 10.1016 / j.febslet.2010.06.040.
CAS Статья PubMed Google Scholar
Ма Б., Калвер Б. П., Бадж Г., Тонгиорги Э., Чао М. В., Танезе Н. Локализация мРНК BDNF с белком болезни Хантингтона в мозге крысы. Mol Neurodegener. 2010; 5: 22. https://doi.org/10.1186/1750-1326-5-22.
CAS Статья PubMed PubMed Central Google Scholar
Krüttner S, Traunmüller L, Dag U, Jandrasits K, Stepien B, Iyer N, et al. Synaptic Orb2A соединяет получение памяти и позднюю консолидацию памяти у Drosophila. Cell Rep. 2015; 11: 1953–65. https://doi.org/10.1016/j.celrep.2015.05.037.
CAS Статья PubMed PubMed Central Google Scholar
Ford L, Asok A, Tripp AD, Parro C, Fitzpatrick M, de Solis CA, et al. Мотив низкой сложности цитоплазматического белка, связывающего элемент полиаденилирования 3 (CPEB3), является критическим для доставки его мишеней в нейроны.bioRxiv. 2020. https://doi.org/10.1101/2020.05.16.100032.
Артикул Google Scholar
Chao HW, Tsai LY, Lu YL, Lin PY, Huang WH, Chou HJ, et al. Делеция CPEB3 усиливает зависимую от гиппокампа память за счет увеличения экспрессии PSD95 и рецепторов NMDA. J Neurosci. 2013; 33: 17008–22. https://doi.org/10.1523/JNEUROSCI.3043-13.2013.
CAS Статья PubMed PubMed Central Google Scholar
Хуанг Ю.С., Юнг М.Ю., Саркисян М., Рихтер Д.Д. Передача сигналов рецептора N-метил-D-аспартата приводит к катализируемому киназой Aurora фосфорилированию CPEB и полиаденилированию мРНК αCaMKII в синапсах. EMBO J. 2002; 21: 2139–48. https://doi.org/10.1093/emboj/21.9.2139.
CAS Статья PubMed PubMed Central Google Scholar
Макэвой М., Цао Г., Ллопис П.М., Кундель М., Джонс К., Хофлер С. и др. Цитоплазматический элемент полиаденилирования, связывающий белок 1, опосредованный трансляцией мРНК в нейронах Пуркинье, необходим для долговременной депрессии мозжечка и координации движений.J Neurosci. 2007. 27: 6400–11. https://doi.org/10.1523/JNEUROSCI.5211-06.2007.
CAS Статья PubMed PubMed Central Google Scholar
Уайт-Гриндли Э., Ли Л., Мохаммад Хан Р., Рен Ф., Сараф А., Флоренс Л. и др. Вклад стабильности Orb2A в регулируемую амилоидоподобную олигомеризацию Orb2 дрозофилы. PLoS Biol. 2014; 12: e1001786. https://doi.org/10.1371/journal.pbio.1001786.
CAS Статья PubMed PubMed Central Google Scholar
Fioriti L, Myers C, Huang YY, Li X, Stephan JS, Trifilieff P и др. Сохранение памяти на основе гиппокампа требует синтеза белка, опосредованного прионоподобным белком CPEB3. Нейрон. 2015; 86: 1433–48. https://doi.org/10.1016/j.neuron.2015.05.021.
CAS Статья PubMed Google Scholar
Дрисальди Б., Колнаги Л., Фиорити Л., Рао Н., Майерс С., Снайдер А.М. и др. SUMOylation — это тормозящее ограничение, которое регулирует прионоподобную агрегацию и активность CPEB3.Cell Rep. 2015; 11: 1694–702. https://doi.org/10.1016/j.celrep.2015.04.061.
CAS Статья PubMed PubMed Central Google Scholar
Павлопулос Э., Трифилиев П., Шевалейре В., Фиорити Л., Заирис С., Пагано А. и др. Neuralized1 активирует CPEB3: функцию непротеолитического убиквитина в синаптической пластичности и хранении памяти. Клетка. 2011; 147: 1369–83. https://doi.org/10.1016/j.cell.2011.09.056.
CAS Статья PubMed PubMed Central Google Scholar
Павлопулос Э., Анезаки М., Скулакис EMC. Neuralized экспрессируется в α / β долях грибовидных тел взрослых дрозофилы и способствует формированию обонятельной долговременной памяти. Proc Natl Acad Sci USA. 2008; 105: 14674–9. https://doi.org/10.1073/pnas.0801605105.
Артикул PubMed Google Scholar
Chater TE, Goda Y. Роль рецепторов AMPA в постсинаптических механизмах синаптической пластичности. Front Cell Neurosci.2014; 8: 401. https://doi.org/10.3389/fncel.2014.00401.
Артикул PubMed PubMed Central Google Scholar
Vogler C, Spalek K, Aerni A, Demougin P, Müller A, Huynh KD, et al. CPEB3 связан с эпизодической памятью человека. Front Behav Neurosci. 2009; 3: 4. https://doi.org/10.3389/neuro.08.004.2009.
CAS Статья PubMed PubMed Central Google Scholar
Li L, Sanchez CP, Slaughter BD, Zhao Y, Khan MR, Unruh JR, et al. Предполагаемая биохимическая инграмма долговременной памяти. Curr Biol. 2016; 26: 3143–56. https://doi.org/10.1016/j.cub.2016.09.054.
CAS Статья PubMed PubMed Central Google Scholar
Лу WH, Yeh NH, Huang YS. CPEB2 активирует трансляцию мРНК GRASP1 и способствует поверхностной экспрессии рецептора AMPA, долговременной потенциации и памяти. Cell Rep.2017; 21: 1783–94. https://doi.org/10.1016/j.celrep.2017.10.073.
CAS Статья PubMed Google Scholar
Savtchouk I, Sun L, Bender CL, Yang Q, Szabó G, Gasparini S, et al. Топологическая регуляция экспрессии синаптического рецептора AMPA с помощью РНК-связывающего белка CPEB3. Cell Rep. 2016; 17: 86–103. https://doi.org/10.1016/j.celrep.2016.08.094.
CAS Статья PubMed PubMed Central Google Scholar
Шин Дж., Саламех Дж. С., Рихтер Дж. Д.. Нарушение нейроразвития из-за домена низкой сложности CPEB4 обнаруживает конвергентный путь с нейродегенерацией. Научный доклад 2016; 6: 29395. https://doi.org/10.1038/srep29395.
CAS Статья PubMed PubMed Central Google Scholar
Кан М.К., Оруганти-Дас А., Купер-Морган А., Джин Дж., Свангер С.А., Басселл Г.Дж. и др. CPEB4 представляет собой белок выживания клетки, который сохраняется в ядре при ишемии или истощении кальция в эндоплазматическом ретикулуме.Mol Cell Biol. 2010; 30: 5658–71. https://doi.org/10.1128/mcb.00716-10.
CAS Статья PubMed PubMed Central Google Scholar
Паррас А., Анта Х, Сантос-Галиндо М., Сваруп В., Элорза А., Нието-Гонсалес Дж. Л. и др. Аутизмоподобный фенотип и деаденилирование мРНК гена риска из-за неправильного сплайсинга CPEB4. Природа. 2018; 560: 441–6. https://doi.org/10.1038/s41586-018-0423-5.
CAS Статья PubMed PubMed Central Google Scholar
Galardi S, Petretich M, Pinna G, D’Amico S, Loreni F, Michienzi A, et al. CPEB1 сдерживает пролиферацию клеток глиобластомы посредством регуляции трансляции мРНК p27Kip1. Научный доклад 2016; 6: 1–9. https://doi.org/10.1038/srep25219.
CAS Статья Google Scholar
Инь Дж., Пак Дж., Ли Дж. Э., Пак Дж. Й., Ким Т. Х., Ким Ю. Дж. И др. CPEB1 модулирует дифференцировку стволовых клеток глиомы путем подавления экспрессии HES1 и SIRT1.Oncotarget. 2014; 5: 6756–69. https://doi.org/https://doi.org/10.18632/oncotarget.2250
Чен У, Ху З, Ли Х, Ли Дж, Сюй XK, Ли Х и др. CPEB4 взаимодействует с виментином и влияет на прогрессирующие функции и плохой прогноз пациентов с астроцитарными опухолями. Tumor Biol. 2016; 37: 5075–87. https://doi.org/10.1007/s13277-015-3975-0.
CAS Статья Google Scholar
Ортис-Сапатер Е., Пинеда Д., Мартинес-Бош Н., Фернандес-Миранда Г., Иглесиас М., Аламеда Ф. и др.Ключевой вклад CPEB4-опосредованного контроля трансляции в прогрессирование рака. Nat Med. 2012; 18: 83–90. https://doi.org/10.1038/nm.2540.
CAS Статья Google Scholar
Zhijun L, Dapeng W., Hong J, Guicong W., Bingjian Y, Honglin L. Сверхэкспрессия CPEB4 в глиоме указывает на плохой прогноз, способствуя миграции и инвазии клеток. Tumor Biol. 2017; 39: 1010428317694538. https://doi.org/10.1177/1010428317694538.
CAS Статья Google Scholar
Вильянуэва Э., Наварро П., Ровира-Ригау М., Сибилио А., Мендес Р., Филлат С. Трансляционное репрограммирование в опухолевых клетках может вызывать онкоселективность при вирусной терапии. Nat Commun. 2017; 8: 1–9. https://doi.org/10.1038/ncomms14833.
CAS Статья Google Scholar
Santoro MR, Bray SM, Warren ST. Молекулярные механизмы синдрома ломкой Х-хромосомы: 20-летняя перспектива. Annu Rev Pathol Mech Dis. 2012; 7: 219–45. https: // doi.org / 10.1146 / annurev-pathol-011811-132457.
CAS Статья Google Scholar
Пиеретти М., Чжан Ф., Фу Й.Х., Уоррен С.Т., Остра Б.А., Каски СТ и др. Отсутствие экспрессии гена FMR-1 при синдроме ломкой Х-хромосомы. Клетка. 1991; 66: 817–22. https://doi.org/10.1016/0092-8674(91)-I.
CAS Статья PubMed Google Scholar
Цинь М., Кан Дж., Бурлин ТВ, Цзян К., Смит С.Б.Постподростковые изменения в региональном синтезе церебрального белка: исследование in vivo на мышах с нулевым Fmr1. J Neurosci. 2005. 25: 5087–95. https://doi.org/10.1523/JNEUROSCI.0093-05.2005.
CAS Статья PubMed PubMed Central Google Scholar
Удагава Т., Фарни Н.Г., Яковцевски М., Кафзан Х., Аларкон Дж. М., Анилкумар С. и др. Генетическое и острое истощение CPEB1 улучшает патофизиологию ломкой Х-хромосомы. Nat Med. 2013; 19: 1473–7.https://doi.org/10.1038/nm.3353.
CAS Статья PubMed PubMed Central Google Scholar
Шу Х, Доннард Э, Лю Б., Юнг С, Ван Р, Рихтер Дж. FMRP связывает оптимальные кодоны со стабильностью мРНК в нейронах. Proc Natl Acad Sci U S. A. 2020; 117: 30400–11. https://doi.org/10.1073/pnas.2009161117.
CAS Статья PubMed Google Scholar
Ханна Дж, Хоссейн Г.С., Кочерха Дж. Потенциал для терапии микроРНК и клинических исследований. Фронт Жене. 2019; 10: 478. https://doi.org/10.3389/fgene.2019.00478.
CAS Статья PubMed PubMed Central Google Scholar
Морган М., Иаконциг А., Муро А.Ф. CPEB2, CPEB3 и CPEB4 координированно регулируются miRNA, распознающими консервативные сайты связывания в положениях паралог своих 3′-UTR. Nucleic Acids Res.2010; 38: 7698–710. https://doi.org/10.1093/nar/gkq635.
CAS Статья PubMed PubMed Central Google Scholar
Fiumara F, Rajasethupathy P, Antonov I, Kosmidis S, Sossin WS, Kandel ER. MicroRNA-22 обеспечивает долгосрочную гетеросинаптическую пластичность при аплизии посредством пресинаптической регуляции CPEB и нижестоящих мишеней. Cell Rep. 2015; 11: 1866–75. https://doi.org/10.1016/j.celrep.2015.05.034.
CAS Статья PubMed Google Scholar
Bates GP, Dorsey R, Gusella JF, Hayden MR, Kay C., Leavitt BR, et al. Болезнь Хантингтона Nat Rev Dis Prim. 2015; 1: 15005. https://doi.org/10.1038/nrdp.2015.5.
Артикул PubMed Google Scholar
Moncke-Buchner E, Reich S, Mücke M, Reuter M, Messer W, Wanker EE, et al. Подсчет повторов CAG в гене болезни Хантингтона путем расщепления эндонуклеазой рестрикции EcoP15I. Nucleic Acids Res. 2002; 30: 83e – 83. https: // doi.org / 10.1093 / nar / gnf082.
Артикул Google Scholar
Joag H, Ghatpande V, Desai M, Sarkar M, Raina A, Shinde M, et al. Роль регуляции клеточной трансляции, связанная с токсичным белком Хантингтина. Cell Mol Life Sci. 2020; 77: 3657–70. https://doi.org/10.1007/s00018-019-03392-y.
CAS Статья PubMed Google Scholar
Барр Дж., Чарания С., Гильмутдинов Р., Яковлев К., Шидловский Ю., Щедл П.Регулятор трансляции CPEB, Orb, функционирует вместе с белками Par для поляризации ооцитов дрозофилы. PLOS Genet. 2019; 15: e1008012. https://doi.org/10.1371/journal.pgen.1008012.
CAS Статья PubMed PubMed Central Google Scholar
Барр Дж., Яковлев К.В., Шидловский Ю., Щедл П. Установление и поддержание клеточной полярности с локализацией мРНК у дрозофилы. BioEssays. 2016; 38: 244–53. https: // doi.org / 10.1002 / bies.201500088.
CAS Статья PubMed PubMed Central Google Scholar
Xu S, Hafer N, Agunwamba B., Schedl P. Белок CPEB Orb2 выполняет несколько функций во время сперматогенеза у Drosophila melanogaster. PLoS Genet. 2012; 8: e1003079. https://doi.org/10.1371/journal.pgen.1003079.
CAS Статья PubMed PubMed Central Google Scholar
Caldeira J, Simões-Correia J, Paredes J, Pinto MT, Sousa S, Corso G и др. CPEB1, новый ген, подавляемый при раке желудка: подход дрозофилы. Кишечник. 2012; 61: 1115–23. https://doi.org/10.1136/gutjnl-2011-300427.
CAS Статья PubMed Google Scholar
Фернандес-Миранда Г., Мендес Р. Семейство белков CPEB, контроль трансляции при старении и раке. Aging Res Rev.2012; 11: 460–72. https: // doi.org / 10.1016 / j.arr.2012.03.004.
CAS Статья PubMed Google Scholar
Giangarrà V, Igea A, Castellazzi CL, Bava FA, Mendez R. Глобальный анализ CPEB показывает последовательные и неизбыточные функции в митотическом клеточном цикле. PLoS ONE. 2015; 10: e0138794. https://doi.org/10.1371/journal.pone.0138794.
CAS Статья PubMed PubMed Central Google Scholar
Майло К., Мартин Дж., Себастьян Д., Эрнандес-Альварес М., Гарсия-Роча М., Рейна О. и др. Циркадианный и UPR-зависимый контроль CPEB4 опосредует трансляционный ответ для противодействия стеатозу печени при стрессе ER. Nat Cell Biol. 2017; 19: 94–105. https://doi.org/10.1038/ncb3461.
CAS Статья PubMed Google Scholar
Интерактивная муха, Drosophila
Андриоли, Л. П., Дигиампьетри, Л.А., де Баррос, Л. П. и Мачадо-Лима, А. (2012). Гекебейн является частью комбинаторного репрессивного кода в передней бластодерме. Dev. Биол. 361 (1): 177-85. PubMed Ссылка: 22027434
Aspöck, G., Ruvkun, G. and Bürglin. Р. (2003). Гомеобоксный ген ceh-2 класса ems Caenorhabditis elegans необходим для функции мотонейрона глотки M3. Разработка 130: 3369-3378. 12810585
Barbieri, A.M., et al. (2002). Инактивация Vax2 у мышей определяет изменение дорсально-вентральной оси глаза, неправильное направление оптических волокон и колобому глаза.Развитие 129: 805-813. 11830579
Bell, E., et al. (2001). Динамические домены экспрессии генов в раннем переднем мозге птиц. Dev. Bio. 236: 76-88. 11456445
Бишоп, К. М., Гудро, Г. и О’Лири, Д. Д. (2000). Регуляция идентичности ареалов в неокортексе млекопитающих с помощью Emx2 и Pax6. Наука 288 (5464): 344-9. Медлайн аннотация: 10764649
Бишоп, К. М., Рубинштейн, Дж. Л. Р. и О’Лири. Д. Д. М.(2002). Определенные действия Emx1, Emx2 и Pax6 в регуляции спецификации областей в развивающемся неокортексе. J. Neurosci. 22 (17): 7627-7638. 12196586
Бишоп, К. М., Гарел, С., Накагава, Ю., Рубенштейн, Дж. Л. и О’Лири, Д. Д. (2003). Emx1 и Emx2 взаимодействуют, чтобы регулировать размер коры, расслоение, дифференцировку нейронов, развитие кортикальных эфферентов и нахождение таламокортикальных путей. J. Comp. Neurol. 457: 345-360. Медлайн аннотация: 12561075
Бончинелли, Э., Гулисано М. и Паннезе М. (1993). Консервированные гены гомеобокса в развивающемся мозге. C. R. Acad. Sci. III 316: 972-984. PubMed Ссылка: 7915635
Bordogna, W., et al. (2005). Гены гомеобокса EMX регулируют микрофтальмию и изменяют биологию меланоцитов. Exp. Cell Res. 311 (1): 27-38. Медлайн аннотация: 16197942
Boyl, P. P., et al. (2001): Развитие переднего и среднего мозга требует ограниченного эпибластами контроля трансляции Otx2, опосредованного его 3 ‘UTR.Развитие 128: 2989-3000. Ссылка PubMed: 11532921
Breiling, A., et al. (2001). Общие факторы транскрипции связывают промоторы, репрессированные белками группы Polycomb. Nature 412: 651-655. 11493924
Capellini, T. D., et al. (2010). Развитие лопатки регулируется генетическими взаимодействиями Pbx1 с членами его семейства и с Emx2 через их совместный контроль над Alx1 . Разработка 137 (15): 2559-69.Ссылка PubMed: 20627960
Кастелли-Гейр, Дж. (1998). линий гена Drosophila необходимы для определенных функций белка HOX Abdominal-B. Development 125, 1269-1274. PubMed Ссылка: 9477325
Chiang, C., et al. (1996). Циклопия и дефект осевого паттерна у мышей, лишенных функции гена Sonic hedgehog . Nature 383: 407-413. PubMed Ссылка: 8837770
Коэн, С. и Юргенс Г. (1990).Опосредование развития головы Drosophila с помощью генов сегментации, подобной разрыву. Nature 346: 482-485. Ссылка PubMed: 1974035
Crozatier, M., et al. (1996). collier , новый регулятор развития головы Drosophila , экспрессируется в единственном митотическом домене. Curr. Биол. 6 (6): 707-718. PubMed Ссылка: 8793297
Crozatier, M., et al. (1999). Формирование паттерна голова по сравнению с туловищем у эмбриона Drosophila ; колье требование для формирования вставочного сегмента.Разработка 126: 4385-4394. PubMed Ссылка: 10477305
Dalton, D., Chadwick, R. и McGinnis, W. (1989). Экспрессия и эмбриональная функция пустых дыхалец : ген гомео-бокса дрозофилы с двумя функциями формирования паттерна на передне-задней оси зародыша. Genes Dev 3: 1940-56. PubMed Ссылка: 2576012
Энрикес, Дж., Венкатасубраманян, Л., Бэк, М., Петерсон, М., Агаева, У. и Манн, Р. С. (2015). Спецификация морфологии отдельных взрослых мотонейронов с помощью кодов факторов комбинаторной транскрипции.Нейрон 86 (4): 955-70. PubMed ID: 25959734
Fernandez, A. S., et al. (1998). Экспрессия генов гомеобокса Emx-1 и Dlx-1 определяет три молекулярно различных домена в конечном мозге эмбрионов мышей, кур, черепах и лягушек: последствия для эволюции конечных подразделений у амниот. Разработка 125: 2099-2111. PubMed Ссылка: 9570774
Gall, A., et al. (2003). Глипикан 4 модулирует передачу сигналов FGF и регулирует формирование паттерна дорсовентрального отдела переднего мозга у эмбрионов Xenopus.Развитие 130: 4919-4929. 12930779
Galli, R., et al. (2002). Emx2 регулирует пролиферацию стволовых клеток центральной нервной системы взрослых млекопитающих. Развитие 129: 1633-1644. 11923200
Galliot, B., et al. (2009). Истоки нейрогенеза, книдарианский взгляд. Dev. Биол. 332: 2-24. PubMed Ссылка: 19465018
Gauchat, D., et al. (2000). Эволюция генов класса Antp и дифференциальная экспрессия генов Hydra Hox / paraHox в формировании переднего паттерна.Proc. Natl. Акад. Sci. 97: 4493-4498. PubMed Ссылка: 10781050
Goldstein, R.E., et al. (2005). Eh2-подобный мотив в Odd-skipped опосредует рекрутирование Groucho и репрессию in vivo. Мол. Клетка. Биол. 25 (24): 10711-20. 16314497
Gont, L. K., et al. (1996). Сверхэкспрессия гена гомеобокса Xnot-2 приводит к образованию хорды у Xenopus . Dev. Биол. 174: 174-178. Ссылка PubMed: 8626017
Grossniklaus, U., Кэдиган, К. М. и Геринг, В. Дж. (1994). Три материнские системы координат взаимодействуют в формировании паттерна головы дрозофилы. Развитие 120: 3155-3171. PubMed Ссылка: 7720559
Groves, J. A., Gillman, C., DeLay, C. N. и Kroll, T. T. (2019). Идентификация новых партнеров связывания для фактора транскрипции Emx2. Протеин J. в PubMed ID: 30628007
Gulisano, M., et al. (1996). Emx1 и Emx2 демонстрируют разные паттерны экспрессии во время пролиферации и дифференцировки развивающейся коры головного мозга у мышей.Евро. J. Neurosci. 8 (5): 1037-1050. PubMed Ссылка: 8743751
Hallonet, M., et al. (1998). Vax1 представляет собой новый ген, содержащий гомеобокс, экспрессирующийся в развивающейся передней брюшной части переднего мозга. Разработка 125 (14): 2599-2610. PubMed Ссылка: 9636075
Halpern, M. E., et al. (1995). Клеточно-автономный сдвиг от аксиального к параксиальному развитию мезодермы у мутантов с плавающей головой у рыбок данио. Разработка 121: 4257-4264. PubMed Ссылка: 8575325
Хамасаки, Т., Лейнгартнер, А., Рингстедт, Т. и О’Лири, Д. Д. (2004). EMX2 регулирует размеры и расположение первичных сенсорных и моторных областей в неокортексе путем прямой спецификации корковых предшественников. Нейрон 43 (3): 359-72. 15294144
Hartmann, B., Reichert, H. and Walldorf, U. (2001). Взаимодействие генов gap в головке дрозофилы: tailless регулирует экспрессию пустых дыхалец на ранних этапах формирования эмбрионального паттерна и развития мозга. Мех. Dev. 109 (2): 161-72.11731230
Хасэгава, Ю., Таката, Н., Окуда, С., Кавада, М., Эйраку, М. и Сасай, Ю. (2016). Возникновение дорсально-вентральной полярности в ткани сетчатки, происходящей из ЭСК. Разработка 143 (21): 3895-3906. Идентификатор PubMed: 27633992
Hassan, B, et al. (1997). Просперо — фактор панневральной транскрипции, который модулирует активность гомеодоменного белка. Proc. Natl. Акад. Sci. 94 (20): 10991-10996. PubMed Ссылка: 9380747
Херт, Ф., (1995). Дефекты развития сегментации мозга, вызванные мутациями генов гомеобокса orthodenticle и пустых дыхалец у Drosophila. Нейрон 15: 769-778. PubMed Ссылка: 7576627
Херт Ф., Хартманн Б. и Райхерт Х. (1998). Действие гомеотического гена в эмбриональном развитии мозга дрозофилы. Развитие 125: 1579-1589. PubMed Ссылка: 9521896
Херт, Ф. (2010). О происхождении и эволюции трехчастного мозга.Brain Behav. Evol. 76 (1): 3-10. PubMed Ссылка: 20926853
Hitoshi, S., et al. (2002). Клоны нервных стволовых клеток регионально специфицированы, но не коммитируются в разных компартментах развивающегося мозга. Развитие 129: 233-244. 11782416
Hong, S. M., et al. (2007). Снижение нейрогенеза в гиппокампе и повышение производительности у взрослых мутантных мышей Emx1. Exp Neurol. [Epub перед печатью]. Медлайн аннотация: 174
Ху, Н.и Кастелли-Гейр, Дж. (1999). Изучение задних дыхалец дрозофилы как модели для понимания генетических и клеточных механизмы, контролирующие морфогенез. Dev. Биол. 214 (1): 197-210. PubMed Ссылка: 10491268
Джонс, Б. и МакГиннис, В. (1993). Регулирование пустых дыхалец с помощью Abdominal-B опосредует функцию идентичности брюшного сегмента. Гены Дев 7: 229-40. PubMed Ссылка: 8094700
Ким, Н., Мин, К. В., Кан, К.Х., Ли, Э. Дж., Ким, Х. Т., Мун, К., Чой, Дж., Ле, Д., Ли, С. Х. и Ким, Дж. У. (2014). Регуляция роста аксонов сетчатки секретируемым гомеодоменным белком Vax1. Элиф 3: e02671. Идентификатор PubMed: 25201875
Ли, Х., Бишоп, К. М. и О’Лири, Д. Д. (2006). Возможные гены-мишени EMX2 включают Odz / Ten-M и другие семейства генов, влияющие на формирование кортикального паттерна. Мол. Cell Neurosci. 33 (2): 136-49. Медлайн аннотация: 16919471
Lichtneckert, R., Белло, Б. и Райхерт, Х. (2007). Специфическая для клеточного происхождения экспрессия и функция гена пустых дыхалец в развитии мозга взрослых особей Drosophila melanogaster. Разработка 134 (7): 1291-300. Медлайн аннотация: 17314131
Lichtneckert, R., Nobs, L. и Reichert, H. (2008). пустых дыхалец необходим для развития контуров нейронов обонятельной проекции у дрозофилы. Развитие 135: 2415-2424. PubMed Ссылка: 18550709
Лигон, К.L., et al. (2003). Утрата функции Emx2 приводит к эктопической экспрессии Wnt1 в развивающемся конечном мозге и корковой дисплазии. Разработка 130: 2275-2287. 12668639
Lord, P. C., et al. (1995). Нормальная экспрессия и эффекты эктопической экспрессии гена гомеобокса (msh) мышечного сегмента Drosophila предполагают роль в дифференциации и формировании паттерна эмбриональные мышцы. Дев Биол 171: 627-640. PubMed Ссылка: 7556942
Лоу, К.J., et al. (2003). Переднезадний паттерн у полухордовых и происхождение хордовой нервной системы. Ячейка 113: 853-865. 12837244
Масиас А. и Мората Г. (1996). Функциональная иерархия и фенотипическое подавление среди гомеотических генов Drosophila : лабиальных и пустых дыхальцев генов. EMBO J. 15: 334-343. PubMed Ссылка: 8617208
Mallamaci, A., et al. (2000). Сдвиг ареала в ранней коре головного мозга мышей с мутантом Emx2 — / — .Nature Neurosci. 3: 679-686. PubMed Ссылка: 10862700
Masai, I., et al. (1997). с плавающей головкой и masterblind регулируют формирование нейронального паттерна в верхней части переднего мозга. Нейрон 18: 43-57
Мелби, А. Э., Варга, Р. М. и Киммел, К. Б. (1996). Спецификация судьбы клеток на дорсальном крае гаструлы рыбок данио. Развитие 122: 2225-2237
Merabet, S., et al. (2002). Генетический скрининг репортера зеленого флуоресцентного белка, который определяет модификаторы функции гена Hox в эмбрионе дрозофилы.Генетика 162: 189-202. 12242233
Miyamoto, N., et al. (1997). Дефекты урогенитального развития у мышей, лишенных Emx2. Развитие 124, 1653-1664
Mohler, J. (1995). Пространственная регуляция экспрессии гена полярности сегментов в передней терминальной области бластодермы дрозофилы эмбрион. Механический Дев 50: 151-161
Morita, T., et al. (1995). Дифференциальная экспрессия мРНК двух гомеопротеинов emx рыбок данио в развивающийся мозг.Neurosci. Lett. 198: 131-134
Морис-Санс, М., Эстасио-Гомес, А., Альварес-Риверо, Дж. И Диас-Бенджумеа, Ф. Дж. (2014). Спецификация подтипов нейронов по разным уровням горбатого. Разработка 141: 4366-4374. Идентификатор PubMed: 25344076
Mui, S.H., et al. (2002). Белок гомеодомена Vax2 формирует дорсовентральную и назотемпоральную оси глаза. Развитие 129: 797-804. 11830578
Муи, С. Х., Ким, Дж. У., Лемке, Г.и Бертуцци, С. (2005). Гены Vax вентрализует глаз эмбриона. Genes Dev. 19 (10): 1249-59. 15
1Муцио, Л. и Малламачи, А. (2003). Emx1, emx2 и pax6 в спецификации, регионализации и ареализации коры головного мозга. Цереб. Кора 13 (6): 641-7. Медлайн аннотация: 12764040
Очоа-Эспиноза, А., Ю., Д., Циригос, А., Струффи, П. и Смолл, С. (2009). Информация о передне-заднем положении при отсутствии сильного бикоидного градиента.Proc. Natl. Акад. Sci. 106 (10): 3823-8. PubMed Ссылка: 19237583
Ода И. и Сайга Х. (2001). Hremx , асцидийный гомолог ems / emx , экспрессируется в переднем и заднебоковом эпидермисе, но не в центральной нервной системе во время эмбриогенеза. Dev. Genes Evol. 211: 291-298. 11466524
Палссон А., Весоловска Н., Рейнисдоттир С., Людвиг М. З. и Крейтман М. (2014). Естественные делеции сайтов связывания Hunchback в энхансере Even-Skipped stripe 3 + 7.PLoS One 9: e91924. PubMed ID: 24786295
Pannese, M., et al. (1998). Гены Xenopus emx идентифицируют предположительный дорсальный конечный мозг и индуцируются организатором головы. сигналы. Мех. Dev. 73 (1): 73-83
Pellegrini, M., et al. (1996). Для образования зубчатой извилины требуется Emx2. Развитие 122: 3893-3898
Pellegrini, M., et al. (2001). Агенез лопатки у гомозиготных мутантов Emx2 . Dev.Биол. 232: 149-156. Идентификатор PubMed: 11254354
Peterson, K.J., et al. (1999), Образец экспрессии Brachyury и Not у морского ежа: сравнительные последствия для происхождения мезодермы в базальных дейтеростомы. Dev. Биол. 207 (2): 419-31
Schock, F., et al. (2000). Фенотипическое подавление пустых дыхалец предотвращается круглой головкой . Nature 405: 351-354.
Шульте, Д., и другие. (1999). Неправильная экспрессия генов гомеобокса, связанных с Emx cVax и mVax2 вентрализует сетчатку и нарушает ее ретинотектальная карта. Нейрон 24: 541-553.
Seibert, J., Volland, D. and Urbach. Р. (2009). Ems и Nkx6 являются центральными регуляторами дорсовентрального паттерна мозга дрозофилы. Разработка 136 (23): 3937-47. Ссылка PubMed: 19
1
Shimogori, T., et al. (2004). Эмбриональные сигнальные центры, экспрессирующие белки BMP, WNT и FGF, взаимодействуют для формирования структуры коры головного мозга, Development 131: 5639-5647.15509764
Shinozaki, K., et al. (2002). Отсутствие клеток Кахаля-Ретциуса и субпластинчатых нейронов, связанное с дефектами тангенциальной миграции клеток из ганглиозных узлов. превосходство в Emx1 / 2 двойной мутант коры головного мозга. Развитие 129: 3479-3492. 12091317
Шинозаки К., Ёсида М., Накамура М., Айзава С., Суда Ю. (2004). Emx1 и Emx2 взаимодействуют на начальной стадии развития архипаллия. Мех.Dev. 121 (5): 475-89. 15147765
Скоттхейм Хонн, Дж., Йоханссон, Л. и Расмусон Лестандер, А. (2016). Регуляция близнеца у безглазого во время развития дрозофилы. Паттерны экспрессии генов. Идентификатор PubMed: 26976323
Spahn, P., Huelsmann, S., Rehorn, K. P., Mischke, S., Mayer, M., Casali, A. and Reuter, R. (2013). Множественные регуляторные меры ограничивают экспрессию змеиного фактора GATA зачатком гемоцитов в мезодерме Drosophila.Дев Биол 386 (1): 272-9. PubMed ID: 24360907
Strutt, H. and Paro, R. (1997). Белковый комплекс поликомб-группы Drosophila melanogaster имеет различный состав в разных генах-мишенях. Мол. Клетка. Биол. 17 (12): 6773-6783
Suda, Y., et al. (2001). Emx2 руководит развитием промежуточного мозга в сотрудничестве с Otx2. Развитие 128: 2433-2450. 11493561
Suda, Y., et al. (2001). Emx2 руководит развитием промежуточного мозга в сотрудничестве с Otx2.Развитие 128: 2433-2450. 11493561
Suda, Y., et al. (2010). Тот же самый энхансер регулирует самую раннюю экспрессию Emx2 в зачатке каудального переднего мозга, последующую экспрессию в дорсальном конечном мозге и более позднюю экспрессию в кортикальной зоне желудочков. Разработка 137 (17): 2939-49. PubMed Ссылка: 20667915
Taglialatela, P., et al. (2004). Скомпрометированная генерация ГАМКергических интернейронов в мозге мышей Vax1 — / — .Развитие 131: 4239-4249. 15280216
Такэ-учи, М., Кларк, Дж. Д. У. и Уилсон, С. В. (2003). Передача сигналов Hedgehog поддерживает интерфейс зрительного стебля и сетчатки посредством регуляции активности гена Vax. Разработка 130: 955-968. 12538521
Talbot, W. S., et al. (1995). Ген гомеобокса, необходимый для развития хорды рыбок данио. Nature 378: 150-157. Ссылка PubMed: 7477317
Тейлор, Х. С. (1998). Регуляторный элемент гена гомеобокса пустых дыхалец состоит из трех отдельных консервативных областей, которые связывают регуляторные белки.Molec. Репрод. Dev. 49: 246-253. 9491376
Theil, T., et al. (1999). Gli3 необходим для экспрессии гена Emx во время развития дорсального конечного мозга. Разработка 126: 3561-3571. Медлайн аннотация: 10409502
Theil, T., et al. (2002). Передача сигналов Wnt и Bmp кооперативно регулирует ступенчатую экспрессию Emx2 в дорсальном конечном мозге. Развитие 129: 3045-3054. 12070081
Урбах, Р. и Технау, Г.М. (2003). Молекулярные маркеры идентифицированных нейробластов в развивающемся мозге дрозофилы. Разработка 130: 3621-3637. 12835380
Вацик Т., Стаббс Дж. Л. и Лемке Г. (2011). Новый механизм транскрипционной регуляции передачи сигналов Wnt в процессе развития. Genes Dev. 25 (17): 1783-95. PubMed Ссылка: 21856776
фон Фровейн, Дж., Визенманн, А. и Гоц, М. (2006). Факторы транскрипции Emx1 и Emx2 подавляют развитие сосудистого сплетения и способствуют судьбе нейроэпителиальных клеток.Dev. Биол. 296 (1): 239-52. Медлайн аннотация: 16793035
Вагнер, Г. П. (2007). Генетика развития гомологии. Nat. Преподобный Жене. 8: 473-479. PubMed Ссылка: 17486120
Уоллдорф У. и Геринг У. Дж. (1992). Пустые дыхальца , ген пробела, содержащий гомеобокс участвует в развитии головы дрозофилы. EMBO J 11: 2247-59. PubMed Ссылка: 1376248
Yoshida, M., et al. (1997). Функции Emx1 и Emx2 при разработке задний конечный мозг.Развитие 124: 101-111. Ссылка PubMed:
71
Younossi-Hartenstein, A., et al. (1997). Контроль раннего нейрогенеза в головном мозге дрозофилы с помощью генов разрыва головы tll, otd, ems, и btd . Dev. Биол 182: 270-283. PubMed Ссылка:
Zembrzycki, A., et al. (2007). Генетическое взаимодействие между факторами транскрипции Sp8 и Emx2 в формировании паттерна переднего мозга. Neural Devel. 2: 8. Аннотация в Medline: 17470284
Интеркалированный диск — обзор
5 Другие внутриклеточные механочувствительные структуры
Чтобы саркомерное укорочение трансформировалось в сокращение клеток, а сократительный аппарат ощущал изменения внешних нагрузок, им требуется механическое соединение с мембраной.Это достигается с помощью сарколеммы, которая состоит из высокоупорядоченных соединений и кортикальной цитоскелетной сети. Кардиомиоциты связаны продольно электромеханически активным соединительным комплексом, известным как интеркалированный диск (ICD), а поперечно Z-диском — специальными фокальными адгезионными бляшками, известными как костамеры. 12 Ощущение и приложение внешних нагрузок саркомерами обеспечивается этими трансмембранными контактами и связанными с ними цитоскелетными сетями, что делает их первичными ответчиками на силы, ведущие к сократительному аппарату и от него.Здесь мы обсудим эти структуры, их влияние на передачу сигналов ниже по течению и их влияние на сократительную функцию.
5.1 Актин-ассоциированный интеркалированный диск и костамерные белки
Недавняя работа определила интеркалированный диск и костамеры, показанные на рис. 9.1 на клеточной мембране, обеспечивающие связи клетка-клетка и клетка-внеклеточный матрикс (ВКМ), как чрезвычайно чувствительные к механочувствительности. т. е. способны реагировать и производить силы и обеспечивать передачу биохимических сигналов через множество сенсорных молекул. 88–93 Большинство этих исследований было сосредоточено на in vitro, системах, 89 или некардиальных клетках, 88,92 , которые имеют те же компоненты, но по-разному организованы. Например, Ле Дюк и его коллеги прикладывали циклическое напряжение к покрытым E-кадгерином магнитным шарикам, связанным с эндотелиальными клетками, и измеряли изменения в отклонении шариков, когда клеточный цитоскелет реагировал. 93 Они обнаружили, что кадгерин-опосредованное связывание является механочувствительным и приводит к локальному усилению жесткости, потенциально управляемому актином усиления кортикального цитоскелета.Спайки в конечном итоге асимптотически приобретают новую жесткость, скорость и величина которой зависят от присутствия актин-связывающей механочувствительной молекулы винкулина. 90,93 Таким образом, актиновый цитоскелет ниже по ходу связей кадгерин-кадгерин реагирует аналогично тем, которые проксимальнее связей интегрин-ЕСМ. 91,94 Разумно предположить, что контакты кадгерина действуют сходным образом в кардиомиоцитах, поскольку существует весь критический механизм и межклеточные контакты в культивируемых миоцитах, как известно, ремоделируются во время циклического и статического растяжения. 95 Однако механочувствительный механизм не всегда присутствует или высокоорганизован. На раннем этапе развития кадгерины экспрессируются повсеместно по всей мембране. 12 Только по мере развития саркомерогенеза и удлинения и гипертрофии клеток кадгерины становятся поляризованными и ограничиваются продольными концами. После того, как он полностью сформирован, вставной диск не остается статичным; пожилой возраст и расширение связаны с сверткой ультраструктуры ICD и обогащением механотрансдуктивными молекулами. 96,97 Предполагается, что ремоделирование актин-связывающих механосенсоров приводит к кортикальному или поперечному усилению жесткости во время старения, 38 , хотя окончательное влияние этого события на сердечную функцию остается неясным.
Помимо нормальных изменений с развитием и возрастом, исследования показывают, что кортикальное ремоделирование, вызванное болезнью или мутацией, может играть функциональную роль в модулировании саркомерной функции. Например, Tangney с соавторами показали, что винкулин-нулевые кардиомиоциты новорожденных мышей испытывают снижение кортикальной жесткости и увеличение расстояния между филаментами, расстояния между тонкими и толстыми филаментами в решетке миофиламентов. 98 Расстояние между нитями влияет на чувствительность к кальцию и выходную мощность саркомеров, 99–101 , возможно, из-за отсутствия кортикальной поперечной компрессии миофиламентов. Кортикальное сжатие посредством осмотической нагрузки in vitro , как было показано, влияет на сократительную функцию в миокарде крысы. 101,102
Наконец, важно учитывать механотрансдукцию при фокальных сращениях, которые играют ведущую роль в сократимости 103,104 и механической индукции миогенной дифференцировки. 88 В частности, в кардиомиоцитах экспрессия интегрина и передача сигналов, а также фокальная адгезия и интегрин-связанные киназы были в центре внимания понимания возрастной гипертрофической передачи сигналов. 105–111 Экспрессия интегрина регулируется как пассивными, так и активными механическими силами, а также лигандами и плотностью лигандов, представленных клетке. 112 Кластеризация интегрина и экспрессия изоформы зависят от белков ЕСМ и их расположения. 113–115 Другим важным компонентом латерального связывания с мембраной является актин-связывающая молекула дистрофина, которая, как полагают, стабилизирует мембрану и обеспечивает дополнительное механическое соединение с кортикальным актиновым цитоскелетом. 116–118 Нокаут дистрофина или экспрессия мутировавшей изоформы приводит к нарушению передачи саркомерной силы на мембрану. Более того, без дистрофина в качестве структурной поддержки, как полагают, происходят конформационные изменения в каналах, активируемых растяжением, что приводит к патологической внутриклеточной утечке кальция и, в конечном итоге, к гибели клеток. 119–121
5.2 Сети промежуточных филаментов и микротрубочек
В то время как актин играет доминирующую роль в регуляции развития и функции кардиомиоцитов, как промежуточные филаменты (IF), так и сети микротрубочек играют важную вспомогательную роль.Например, IFs чрезвычайно деформируемы, их можно растягивать в несколько раз больше своей длины провисания. 122–124 Один из этих IFs, desmin, обеспечивает каркас вокруг Z-диска и связывает их латерально, связывается с десмосомами на интеркалированном диске и связывает ядро с соседними саркомерами. 125 Считается также, что десмин локализуется в костамерах, возможно, обеспечивая дополнительную связь между саркомерами и ЕСМ. 118 Это ранний маркер кардиогенеза. 126,127 Десминопатии возникают в результате мутаций гена десмина и последующей цитоплазматической агрегации десмина и нарушения сборки миофибрилл. 128–130 У пациентов с аритмогенной кардиомиопатией правого желудочка были обнаружены мутации десмина, что позволяет предположить, что это может играть решающую роль в сцеплении ЭК через продольную нагрузку на саркомеры или стабилизацию щелевых контактов на вставочном диске. 131–133
Другой белок ПФ, преобладающий в кардиомиоцитах, — это ламин, который, как и десмин, также чрезвычайно податлив.Например, во время спецификации клонов стволовых клеток, экспрессия ламина в стволовых клетках масштабируется с жесткостью субстрата 134 и является детерминантом деформируемости ядерного цитоскелета. 135,136 Более того, он может действовать как универсальный преобразователь механических сигналов в ядро для регулирования экспрессии генов. 137 Таким образом, дефекты ламината A / C могут отрицательно влиять на передачу сигнала, делая их менее эффективными. Дефекты ламина связаны с поздним старением, 138 предположительно из-за генетической нестабильности и накопленных повреждений ДНК от внешних стрессоров, 139 и дилатационной кардиомиопатии. 15,140,141 Существование механических связей между ядром и сетью цитоскелета подразумевает, что некоторая форма коммуникации необходима во время развития. 142
Обратите внимание, что существуют структурные перекрытия в сетях актина и IF; Также считается, что локализующийся в МКБ винкулин стабилизирует щелевые контакты, 134 и нарушения актин-связывающих неспринов, обнаруженных в ядре, также, как известно, подавляют биомеханическую передачу сигналов и вызывают сердечнососудистую дисфункцию. 143,144 Эти находки предполагают функциональное перекрытие в обеих цитоскелетных сетях.
С другой стороны, микротрубочки имеют решающее значение в управлении цитокинезом 145 и в управлении везикулярным транспортом. Сеть микротрубочек также выполняет несколько функций несения нагрузки и передачи сигналов, влияющих на сердечную функцию. 146–148 Большое внимание было уделено тому, как трансдукция сигнала изменяется после применения колхицина, фармакологического агента, который вызывает разрушение нитей микротрубочек, а также тому, как нарушение регуляции микротрубочек происходит во время гипертрофии сердца. 149–152 Хотя неясно, являются ли механические пертурбации, нарушение везикулярного транспорта или и то, и другое ответственными за изменения функции, известно, что микротрубочки вносят вклад в пассивную жесткость 20 и реконструируются в течение возраста 38 ; оба эти наблюдения указывают на микротрубочки как на важные регуляторы функции кардиомиоцитов.
5.3 Мембрана кардиомиоцитов
Помимо внутриклеточных и трансмембранных механочувствительных компартментов, мембрана сердечных миоцитов также обогащена чувствительными к растяжению структурами. 153–157 Каналы, активируемые растяжением, участвуют в модуляции обращения с кальцием и ритмичности, хотя напряжения, необходимые для наблюдаемой активации растяжения, часто являются супрофизиологическими. 158 Мембрана частично защищена от растяжения за счет кавеолинов, цитоскелетно регулируемых инвагинаций, которые могут добавлять дополнительный материал к мембране за счет быстрой разборки под осмотическими нагрузками 159 или растяжения.