
Физиология мышечной ткани человека: Лекция № 5 физиология мышечной ткани План лекции
Лекция № 5 физиология мышечной ткани План лекции
14
Виды мышечной ткани в организме человека, ее физические и физиологические свойства. Сравнительная характеристика поперечно-полосатой и гладкой мышечной ткани.
Двигательная единица. Классификация.
Современные представления о механизме мышечного сокращения.
Виды и режимы мышечного сокращения. Теория тетанического сокращения.
Утомление в мышечной ткани.
Виды мышечной ткани в организме человека, ее физические и физиологические свойства. Сравнительная характеристика поперечно-полосатой и гладкой мышечной ткани.
Одним из наиболее
распространенных проявлений возбуждения
в возбудимых тканях является изменение
длины – сокращение.
Различают поперечно-полосатую и гладкую мышечную ткань. Гладкая мышечная ткань представлена во внутренних полых органах: мышечном слое стенки сосудов, желудочно-кишечного тракта, мочевого пузыря, матки и некоторых других. Поперечно-полосатая мышечная ткань составляет основу скелетных мышц. Особой разновидностью поперечно-полосатой мускулатуры является сердечная мышца. Физиологические особенности сердечной мышцы мы будем рассмат-ривать в четвертом семестре при изучении системы кровообращения.
Поперечно-полосатая мышечная ткань опорно-двигательного аппарата
Любая скелетная мышца состоит из 3-х типов мышечных волокон:
быстрых, фазических мышечных волокон. В связи с относительно низким содержанием в указанных волокнах пигмента миоглобина данные волокна называют белыми;
медленных, тонических мышечных волокон.
 В связи
с относительно высоким содержанием в
данных волокнах миоглобина, и в связи
с особенностью окраски, данные мышечные
волокна называют красными;
В связи
с относительно высоким содержанием в
данных волокнах миоглобина, и в связи
с особенностью окраски, данные мышечные
волокна называют красными;волокон-рецепторов.
 В связи
с относительно высоким содержанием в
данных волокнах миоглобина, и в связи
с особенностью окраски, данные мышечные
волокна называют красными;
В связи
с относительно высоким содержанием в
данных волокнах миоглобина, и в связи
с особенностью окраски, данные мышечные
волокна называют красными;Основная функция первых двух групп волокон состоит в изменении длины мышц: формирование мышечного тонуса и различных вариантов быстрых (фазических) сокращений. Первые две группы мышечных волокон объединяют в группу экстрафузальных мышечных волокон. У волокон-рецепторов есть название-синоним – интрафузальные мышечные волокна.
На сегодняшней лекции мы рассмотрим особенности строения и физиологических свойств экстрафузальных волокон
Выделяют физические и физиологические свойства мышечной ткани. К физическим свойствам мышечной ткани относят вязкость, эластичность, пластичность. К физиологическим – раздражимость, возбудимость, лабильность, проводимость.
Физические свойства мышц
Растяжимость – способность менять длину под влиянием приложенной силы;
Эластичность – способность мышцы восстанавливать первоначальную форму после прекращения действия сил, вызывающих ее деформацию;
Сила мышц – максимальный груз, который мышца способна еще приподнять;
Работа мышц – произведение поднимаемого груза на высоту подъема.
Измеряется в кгм.
 Измеряется в кгм.
Измеряется в кгм.А = Р х Δh, где А – работа выполненная мышцой, Р – вес груза, Δh – высота, на которую поднят груз.
Максимальный объем выполненной мышцей работы отмечаеся при средних величинах нагрузки (см. рис. 1).
Работа (А)
0 Вес груза (Р)
Рис.1 Объем выполненной работы мышцей в зависимости от величины
груза
Скелетная
поперечно-полосатая мышечная ткань
состоит из мышеч-ных волокон диаметром
от 10 до 100 микрон и длиной от 5 до 400 мм.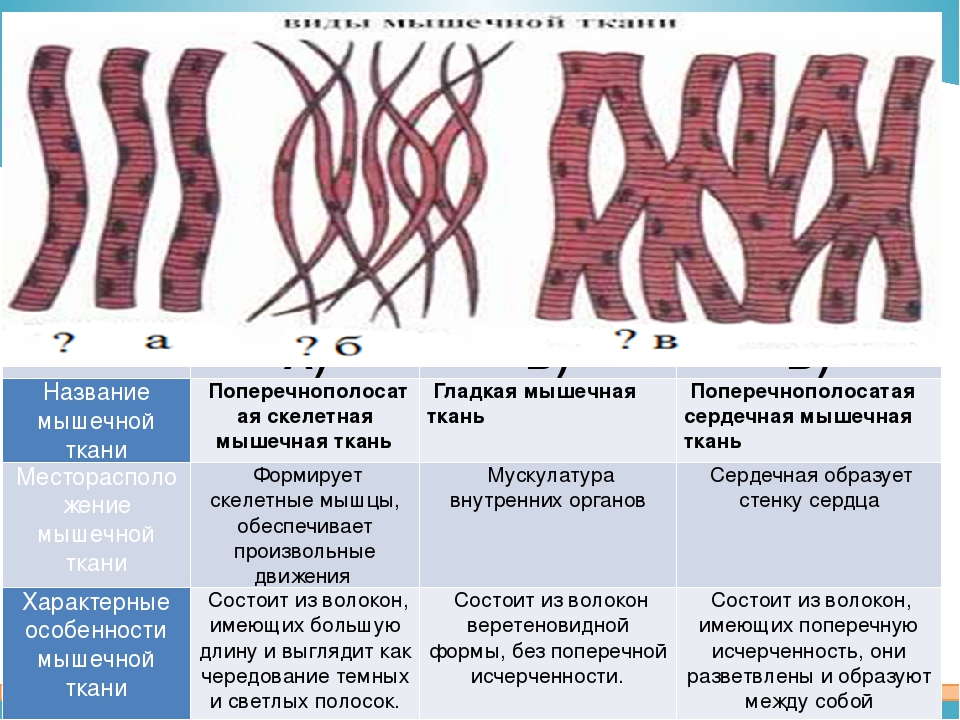 Каждое мышечное волокно содержит до
1000, а в ряде случаев и более сократительных
волокон – миофибрил, толщиной от 1 до 3
микрон. Каж-дая миофибрила состоит из
множества толстых и тонких нитей –
миофи-ламентов (соотношение тонких и
толстых нитей 2 : 1). Каждое мышеч-ное
волокно поперечно-полосатой мускулатуры
содержит большое количество ядер и
представляет из себя симпласт. Толстые
нити состоят из белка миозина, а тонкие
– актина. Кроме того в состав тонких
нитей входят еще белки тропонин и
тропомиозин, формирующими с актомиозином
единый регуляторный комплекс. Миозиновые
нити имеют поперечно-ориентированные
мостики, которые участвуют в механизмах
взаимодействия актиновых и миозиновых
нитей (см. рис. 2).
Каждое мышечное волокно содержит до
1000, а в ряде случаев и более сократительных
волокон – миофибрил, толщиной от 1 до 3
микрон. Каж-дая миофибрила состоит из
множества толстых и тонких нитей –
миофи-ламентов (соотношение тонких и
толстых нитей 2 : 1). Каждое мышеч-ное
волокно поперечно-полосатой мускулатуры
содержит большое количество ядер и
представляет из себя симпласт. Толстые
нити состоят из белка миозина, а тонкие
– актина. Кроме того в состав тонких
нитей входят еще белки тропонин и
тропомиозин, формирующими с актомиозином
единый регуляторный комплекс. Миозиновые
нити имеют поперечно-ориентированные
мостики, которые участвуют в механизмах
взаимодействия актиновых и миозиновых
нитей (см. рис. 2).
Наряду
с миофибрилами в саркоплазме мышечных
волокон имеется система канальцев –
саркоплазматический ретикулум, имеющих
расширения в области биологической
мембраны мышечного волокна (цистерны
саркоплазматического ретикулума). В
цистернах саркоплаз-матического
ретикулума депонируются ионы кальция
– Са++.
Презентация по анатомии и физиологии человека на тему «Мышечная ткань» | Презентация к уроку на тему:
Слайд 1
Теоретическое занятие Тема: «Мышечная ткань» Составила преподаватель анатомии и физиологии человека Губанова А.С.Слайд 2
Ткань — это совокупность клеток и межклеточного вещества, имеющие общее происхождение, строение и выполняющие общие функции. Мышечные ткани- ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Состоят из вытянутых клеток, которые принимают раздражение от нервной системы и отвечают на него сокращением. Они обеспечивают перемещения в пространстве организма в целом, его движение органов внутри организма (сердце, язык, кишечник и др. ) и состоят из мышечных волокон.
) и состоят из мышечных волокон.
Слайд 4
Гладкая мышечная ткань Гладкая мышечная ткань мезенхимного происхождени , располагается в стенке внутренних органов и сосудов. Структурная единица- гладкий миоцит. Это клетка веретеновидной, иногда отростчатой формы (матка, эндокард, аорта). Цитолемма гладкого миоцита образует многочисленные впячивания – кавеолы (мелкие пузырьки). Снаружи цитолемму покрывает тонкая базальная мембрана. В базальной мембране каждого миоцита есть отверстия, где клетки контактируют друг с другом при помощи нексусов , осуществляющих метаболические связи. Органеллы общего значения – комплекс Гольджи, митохондрии, свободные рибосомы, саркоплазматическая сеть – локализуются в основном около полюсов ядра. Наиболее развитыми и многочисленными из них являются митохондрии.
Слайд 5
Иннервация гладкой мышечной ткани осуществляется симпатическими и парасимпатическими нервными волокнами.
Слайд 6
Поперечнополосатая мышечная ткань Скелетная мышечная ткань образует скелетные мышцы двигательного аппарата. Структурным компонентом этой ткани являются миосимпласты и миосателлитоциты , вместе образующие мышечные волокна. Они развиваются из клеток миотомов дорзальной мезодермы. Оболочку волокна называют сарколеммой (от греч. « саркос » – мясо). Она двуслойна: внешний слой образован базальной мембраной, связанной с ретикулярными и тонкими коллагеновыми волокнами, и входит в состав опорного аппарата, а внутренний слой является плазмолеммой симпласта и участвует в проведении нервных импульсов. Между базальной мембраной и плазмолеммой располагаются миосателлитоциты – небольшие клетки с одним ядром, слабо развитыми органеллами и собственной оболочкой. Иннервируются мышцы эфферентными и афферентными волокнами соматической нервной системы.
Структурным компонентом этой ткани являются миосимпласты и миосателлитоциты , вместе образующие мышечные волокна. Они развиваются из клеток миотомов дорзальной мезодермы. Оболочку волокна называют сарколеммой (от греч. « саркос » – мясо). Она двуслойна: внешний слой образован базальной мембраной, связанной с ретикулярными и тонкими коллагеновыми волокнами, и входит в состав опорного аппарата, а внутренний слой является плазмолеммой симпласта и участвует в проведении нервных импульсов. Между базальной мембраной и плазмолеммой располагаются миосателлитоциты – небольшие клетки с одним ядром, слабо развитыми органеллами и собственной оболочкой. Иннервируются мышцы эфферентными и афферентными волокнами соматической нервной системы.
Слайд 7
Сердечная мышечная ткань Напоминает одновременно и на поперечно-полосатую, и на гладкую. Как и гладкая, она регулируется вегетативной нервной системой. Однако сокращается она так же активно, как и поперечно-полосатая. Состоит она из клеток, называющихся кардиомиоцитами . Функции мышечной ткани данного вида: Она всего одна: обеспечение передвижения крови по организму
Функции мышечной ткани данного вида: Она всего одна: обеспечение передвижения крови по организму
Развитие мышечной ткани в онтогенезе Текст научной статьи по специальности «Биологические науки»
ОБЗОРЫ
РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИ В ОНТОГЕНЕЗЕ
Р.В.Тамбовцева1
Институт возрастной физиологии РАО, г.Москва

Ключевые слова: мышечное волокно, постнатальный онтогенез, рост и развитие, дифференцировка, индивидуальное развитие, нейробласты, мотонейроны, анаэробно-гликолитическое энергообеспечение, лабильность, митохондриальное окисление.
The development of muscle tissue in ontogeny. Periodic growth and development of skeletal muscles is determined by the sequence of differentiations during which the structure, contractile and power muscle characteristics change. The sequence of ontogenetic development of skeletal muscles, common for all mammals, is closely connected with their biological nature. Muscular fibers of all types manifest high lability of power metabolism at different stages of postnatal development.

Индивидуальное развитие организма как в фазе становления, так и в старости сопряжено с возрастными изменениями скелетных мышц [11,29,31,37,40,48].
Рост и развитие мышечной ткани в постнатальном онтогенезе идут в форме последовательного ряда дифференцировок [32], в ходе которых происходят изменения морфофункциональной и метаболической организации мышечных волокон. Как известно [6,16,24,46,50], основой процессов дифференцировок является активация нового синтетического аппарата, связанного с вовлечением и усиленным размножением ядер клеток-сателлитов, появлением новой РНК, в то время как рост мышечных клеток опирается на имеющиеся возможности клеточного синтеза в зонах эргастоплазмы на уже подготовленных матрицах. Сигнал о необходимости очередной передифференцировки может поступить как нейрогенно, с синаптических окончаний, так и гуморально [3,4,16,18,46,50]. Нейрогенные влияния проходят сверху вниз, и их последовательность устанавливается по мере созревания вышера-сположенных структур головного мозга в процессе онтогенетического развития.
Контакты: 1 Тамбовцева Р.В.-E-mail:[email protected]
— 81 —
у одного из исследованных нами объектов процесс дифференциации мышечных волокон в период первичного становления локомоций в возрасте 1-2 лет у человека или в возрасте 20-22 дней у крысят, не является окончательным, определяющим дефинитивное развитие периферического двигательного аппарата у взрослых [12,13,14,25,26,27,28,29]. Дифференцировка скелетных мышц — это сложный многоэтапный процесс, в котором уровень дефинитивной организации мышечных структур достигается только к завершению полового созревания [25,29].
Организация управления движениями очень сложна, и ее становление занимает у млекопитающих длительный отрезок постнатального онтогенеза. В это же время происходит созревание мышечной ткани. Эти два процесса (развитие мышечной и нервной ткани) в значительной степени сопряжены. При этом следует обратить внимание на то, что организация управления движениями разделена на уровни, определенным образом взаимодействующими между собой, но четко проявляющими свою самостоятельность как в филогенезе системы построения движения, так и в процессе индивидуального развития. В онтогенезе происходит «….анатомическое дозревание центрально-нервных субстратов двигательных функций, опоздавших к моменту рождения и заканчивающих (по крайней мере в отношении миелинизации) к 2 — 2 1/2 — летнему возрасту. Это дозревание сопровождается функциональным столь же поочередным вступлением в строй мозговых эффекторных систем» [2,3,16,32].
При этом следует обратить внимание на то, что организация управления движениями разделена на уровни, определенным образом взаимодействующими между собой, но четко проявляющими свою самостоятельность как в филогенезе системы построения движения, так и в процессе индивидуального развития. В онтогенезе происходит «….анатомическое дозревание центрально-нервных субстратов двигательных функций, опоздавших к моменту рождения и заканчивающих (по крайней мере в отношении миелинизации) к 2 — 2 1/2 — летнему возрасту. Это дозревание сопровождается функциональным столь же поочередным вступлением в строй мозговых эффекторных систем» [2,3,16,32].
Сопоставляя возрастную динамику композиции мышечных волокон разного типа, можно выделить общие закономерности, свойственные всем исследованным объектам. Несомненная общность характеризует очередность развития волокон разного типа, изменение темпов роста, взаимосвязь с процессами полового созревания, половой диморфизм.
Исследуя возрастную динамику развития мышечных волокон, мы прежде всего имеем дело с закономерностями развития всего периферического звена двигательной системы, включающего в себя определенные мотонейроны и связанные с ними группы мышечных волокон. Таким образом, в процессе онтогенеза развиваются не отдельные мышечные волокна, а суперструктуры, обычно обозначаемые как двигательные единицы (ДЕ), в которых изменения состояния мышечных волокон связано, в первую очередь, с развитием соответствующих мотонейронов. К такому выводу можно прийти на основании многочисленных данных, накопленных в литературе после классических опытов с перекрестной иннервацией Дж.Экклса [39]. Большинство таких работ напрямую не связано с возрастными исследованиями, но все они однозначно доказывают зависимость состояния мышечных волокон от трофических и импульсных влияний со стороны мотонейронов, контролирующих их двигательную активность [16,18].
Перенос акцента при анализе механизмов развития скелетно-мышечной ткани с мышечных волокон на соответствующие мотонейроны, заставляет изменить сложившиеся представления. Так, при рассмотрении возрастных преобразований мышечной ткани в период полового созревания исследователи исходят только из прямого действия гормонов на мышечную ткань [38,43]. В то же время очевидно, что необходимо учитывать и изменения двигательных центров спинного мозга.
Так, при рассмотрении возрастных преобразований мышечной ткани в период полового созревания исследователи исходят только из прямого действия гормонов на мышечную ткань [38,43]. В то же время очевидно, что необходимо учитывать и изменения двигательных центров спинного мозга.
— 82 —
Хорошо известно, что в период полового созревания мотонейроны спинного мозга, особенно в пояснично-крестцовой области, претерпевают существенные изменения [30]. В цитированной выше работе показано, что наиболее значимые биохимические изменения в мышце тазовой области (ш.1еуа1ог аш) наблюдаются в период полового созревания.
На основании полученных нами данных складываются новые представления о развитии скелетных мышц в постнатальном онтогенезе, несколько отличающиеся от сложившихся представлений.
В наших исследованиях постнатального развития мышечных волокон различных мышц мелких лабораторных животных доказано наличие перестроек молекулярной организации АТФазы миозина, наступающих у крыс с 7-8 дня постнатального развития [26,27,28]. Причем сроки наступления этого процесса в различных мышцах имеют отчетливый кранио-каудальный градиент и, по-видимому, связаны с созреванием мотонейронов спинного мозга (4). Процесс этот завершается, как уже указывалось выше, к 20 дню жизни, то есть ко времени выхода крысят из гнезда и началу самостоятельного передвижения. Однако в наших исследованиях было показано, что процесс постнатальной дифференциации мышечных волокон у крыс проходит в два этапа. Первый — в возрасте от 7 до 16 дней определяется выделением структур с «медленным» миозином в мышечных волокнах I типа, а второй период с 16 по 20 день с дифференциацией волокон II типа с «быстрым» и «промежуточным» миозином, что связано с разделением фазных волокон II типа на подтипы МВ11А и МВПВ.
Причем сроки наступления этого процесса в различных мышцах имеют отчетливый кранио-каудальный градиент и, по-видимому, связаны с созреванием мотонейронов спинного мозга (4). Процесс этот завершается, как уже указывалось выше, к 20 дню жизни, то есть ко времени выхода крысят из гнезда и началу самостоятельного передвижения. Однако в наших исследованиях было показано, что процесс постнатальной дифференциации мышечных волокон у крыс проходит в два этапа. Первый — в возрасте от 7 до 16 дней определяется выделением структур с «медленным» миозином в мышечных волокнах I типа, а второй период с 16 по 20 день с дифференциацией волокон II типа с «быстрым» и «промежуточным» миозином, что связано с разделением фазных волокон II типа на подтипы МВ11А и МВПВ.
Первые два этапа молекулярной дифференциации сократительного аппарата сопряжены с изменением тканевой энергетики. Специализация волокон по энергообеспечению начинает отчетливо проявляться у крыс после выхода из гнезда (20-22 день жизни). Еще одна волна передифференцировок связана с периодом полового созревания и, по-видимому, касается только волокон II типа. У крысят в 45-дневном возрасте резко увеличивается относительное число волокон типа НА и лишь затем, к 60-дневному возрасту, происходит перераспределение волокон и увеличивается относительное число МВПВ. На пубертатный период приходится и интенсивный рост поперечника волокон II типа. Как уже говорилось выше, интенсивный рост волокон II типа и развитие механизмов анаэробно-гли-колитического энергообеспечения связаны с влиянием стероидных гормонов — в первую очередь тестостерона [17,20,38,43]. В наших экпериментах показано, что при дефиците мужских половых гормонов, достигаемом кастрацией, у крысят сохраняется большое количество промежуточных волокон типа ПА, в то время как дополнительное введение тестостерона активирует переход промежуточных структур в волокна типа ПВ и значительно увеличивает активность ферментов анаэробного гликолиза, а также интенсивность ростовых процессов [17].
Еще одна волна передифференцировок связана с периодом полового созревания и, по-видимому, касается только волокон II типа. У крысят в 45-дневном возрасте резко увеличивается относительное число волокон типа НА и лишь затем, к 60-дневному возрасту, происходит перераспределение волокон и увеличивается относительное число МВПВ. На пубертатный период приходится и интенсивный рост поперечника волокон II типа. Как уже говорилось выше, интенсивный рост волокон II типа и развитие механизмов анаэробно-гли-колитического энергообеспечения связаны с влиянием стероидных гормонов — в первую очередь тестостерона [17,20,38,43]. В наших экпериментах показано, что при дефиците мужских половых гормонов, достигаемом кастрацией, у крысят сохраняется большое количество промежуточных волокон типа ПА, в то время как дополнительное введение тестостерона активирует переход промежуточных структур в волокна типа ПВ и значительно увеличивает активность ферментов анаэробного гликолиза, а также интенсивность ростовых процессов [17].
Полученные нами данные согласуются с исследованиями A.C.Platzer [51]. Электронно-микроскопическое изучение мышц мыши и крысы позволило автору придти к выводу о стадийности в развитии скелетных мышц. Анализ спектра изоферментов ЛДГ на разных сроках развития свидетельствует о том, что в ходе онтогенеза происходит изменение активности отдельных изоферментов, что
— 83 —
позволяет судить об изменении метаболизма мышц. На ранних стадиях постна-тального онтогенеза в мышцах отмечается преобладание активности ЛДГ1 и ЛДГ2, что говорит о функционировании органа в условиях окислительного метаболизма. На более поздних этапах постнатального онтогенеза в мышцах усиливается активность изоферментов ЛДГ4 и ЛДГ5, характеризующих преобладание в мышцах анаэробного метаболизма.
Волокна II типа достаточно изменчивы, и в ходе индивидуального развития мы не раз видим во время дифференцировок довольно значительные изменения их числа. Эти изменения связаны либо с усилением, либо с ослаблением окислительного потенциала клетки. При этом очень часто можно заметить, что число исчезнувших волокон типа ПА соответствует количеству появившихся волокон типа ПВ, что может быть истолковано как результат прямого перехода структур друг в друга. Видимо, поэтому многие исследователи придерживаются точки зрения, что не существует разнообразия мышечных волокон, как отдельных структур, а имеется большой волоконный симпласт, в котором происходит постоянное изменение энергетического профиля в зависимости от эндо-и экзогенных факторов [21,22,23]. Согласно нашим исследованиям, существует несколько мало меняющихся волокон, например: тип I , тип I — «гигантские» (до сих пор исследователи не могут определить: какова же функция данных структур [20], и небольшой процент волокон типа ПВ, сохраняющихся до старости как у млекопитающих, так и у человека. Весь остальной объем занимает большое количество волокон типа ПА, постоянно меняющих свой энергетический профиль.
Эти изменения связаны либо с усилением, либо с ослаблением окислительного потенциала клетки. При этом очень часто можно заметить, что число исчезнувших волокон типа ПА соответствует количеству появившихся волокон типа ПВ, что может быть истолковано как результат прямого перехода структур друг в друга. Видимо, поэтому многие исследователи придерживаются точки зрения, что не существует разнообразия мышечных волокон, как отдельных структур, а имеется большой волоконный симпласт, в котором происходит постоянное изменение энергетического профиля в зависимости от эндо-и экзогенных факторов [21,22,23]. Согласно нашим исследованиям, существует несколько мало меняющихся волокон, например: тип I , тип I — «гигантские» (до сих пор исследователи не могут определить: какова же функция данных структур [20], и небольшой процент волокон типа ПВ, сохраняющихся до старости как у млекопитающих, так и у человека. Весь остальной объем занимает большое количество волокон типа ПА, постоянно меняющих свой энергетический профиль.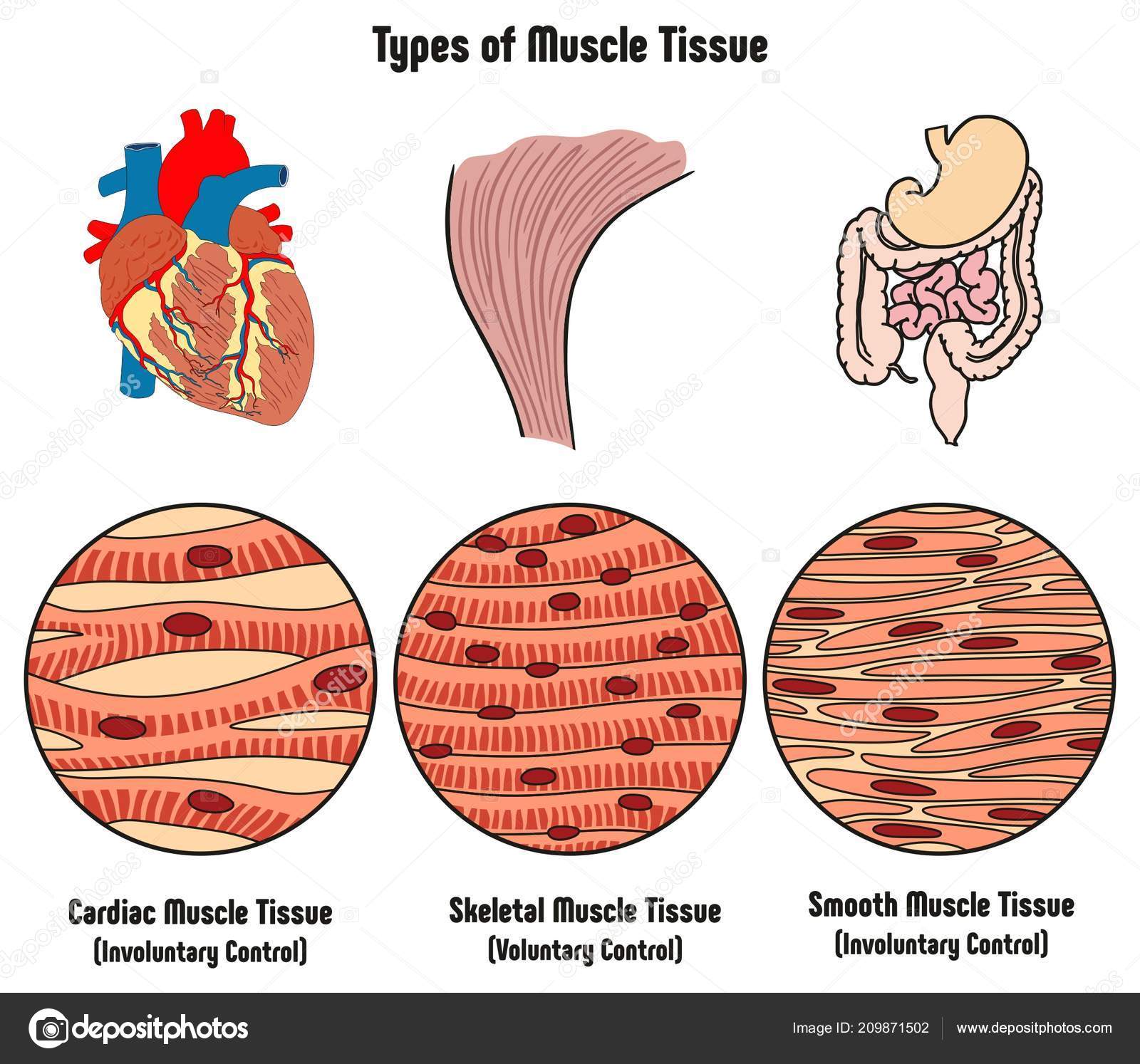 Сюда же мы отнесли и волокна типа I — вторично окислительные (камбаловидная мышца).
Сюда же мы отнесли и волокна типа I — вторично окислительные (камбаловидная мышца).
Сравнение с данными, полученными на зрелорождающихся морских свинках, показывает, что основные фазы развития мышечных волокон полностью сохраняются, однако начало дифференциации тонических волокон I типа смещено к более раннему возрасту и происходит еще пренатально, а начало дифференциации волокон II типа наступает уже в возрасте 6 дней.
Гистохимические исследования проведенные на крысах-самках показали, что динамика развития мышечных волокон у самок и самцов совпадает, но основные различия определяются сроками созревания и относительным содержанием различных мышечных структур. У самок крыс и морских свинок дифференцировки фазных волокон начинаются на 5 дней раньше, чем у самцов. К концу пубертатного периода устанавливается окончательное количество различных типов мышечных волокон, при этом выделяется полный набор МВ! МВПА, МВПВ. Однако количество МВI и МВПА у самок на 20-30% больше, чем у самцов.
Нестандартной мышцей в плане развития является камбаловидная мышца. Эта мышца у взрослых состоит, преимущественно, из волокон одного типа. Обычно считается, что это медленные окислительные волокна I типа. В то же время, знакомство с особенностями возрастного развития волокон этой мышцы заставляет усомниться в правильности этих утверждений. Действительно, в отличие от смешанных мышц, здесь отсутствует первая волна перестроек, в результате которой образуются волокна I типа. На основании наших данных можно предположить, что в камбаловидной мышце дифференциация соответствующих структур начинается с формирования
— 84 —
волокон II типа. При этом образуются структуры с «промежуточным», а затем с «быстрым» миозином. В дальнейшем происходит постепенное увеличение популяции окислительных волокон с «медленным» миозином. Такие структуры, в отличие от медленных волокон I типа смешанной мышцы, сохраняют высокие темпы роста. Развитие этих структур также находится под гормональным контролем, но это гормоны щитовидной железы. В работе Н.П.Резвякова [20] показано, что после тирео-идэктомии у крыс в камбаловидной мышце уменьшается содержание медленных волокон. К тому же известно, что содержание в крови растущих крыс тироксина и трийодтиронина с 6 по 9 неделю жизни значительно увеличивется [20]. Волокна такого же типа имеются и в смешанных мышцах крыс, а также в глубоких слоях круглых мышц человека. Во всех случаях это крупные окислительные волокна с «медленным» миозином, они отчетливо выявляются уже после полового созревания и были обозначены нами как вторичные окислительные волокна (к). Считаем, что появление всех этих структур связано с адаптацией к определенному роду нагрузок. У взрослых крыс изменение уровня того или иного типа нагрузок приводит к нарушению в этой мышце организации волокон типа к и переходу их в один из подтипов промежуточных волокон ПА и далее в ПВ, с появлением молекулярных фрагментов «быстрого» миозина. Такие изменения происходят также после длительного устранения гравитации при полете на биоспутниках [19], при гиподинамии и при адаптации к холоду [20]. Имеющиеся данные позволяют полагать, что и в ходе индивидуального развития появление структур типа к связано с ростом организма и увеличением нагрузок на соответствующие мышцы. Вполне возможно, что подобные превращения происходят под влиянием гормонов щитовидной железы, контролирующих развитие механизмов окислительной энергетики.
Сопоставление с данными, полученными на человеке, показывает, что основные принципы развития скелетной мускулатуры в процессе индивидуального развития человека полностью соответствуют таковым, изученным на животных. У человека первый этап дифференциации тонических волокон с «медленным» миозином, сохраняющим высокую АТФазную активность после преинкубации в кислой среде, обнаруживается у плода на 5-6 месяце внутриутробной жизни [29]. Сопоставление этих результатов с данными исследований, проведенных на экспериментальных животных, позволяет признать, что у человека развитие скелетной мускулатуры идет по типу зрелорождающихся. Очень ограниченный набор двигательных возможностей новорожденного ребенка, казалось бы, противоречит этому выводу. Можно полагать, что у человека сохраняется общий тип развития, свойственный приматам, характеризующийся высокими возможностями тонической мускулатуры конечностей у новорожденных. Особенность человека связана с более длительным развитием высших уровней регуляции движений [1,3]. В первые месяцы постнатальной жизни во многих областях мозга еще преобладают незрелые нервные элементы и даже нейробласты [5]. В то же время, к моменту рождения оказывается довольно хорошо сформированным спинальный уровень регуляции. Функциональные возможности этого уровня не идут дальше реализации простых форм движения типа сгибательного рефлекса и его вариантов. Однако уровень развития мотонейронов, по-видимому, достаточен для инициации
— 85 —
первой волны мышечных дифференцировок. При этом необходимо отметить большое число волокон, начинающих дифференциацию по I типу — в разных смешанных мышцах их число составляет не менее 50-60%. В то время, как относительное количество волокон I типа в смешанных мышцах конечностей мелких лабораторных животных не превышает 10-15%. Можно полагать, что число волокон I типа определяется появлением гравитационных нагрузок. На мускулатуру человека они действуют в значительно большей степени, что связано с большей массой тела по сравнению с мелкими лабораторными животными [12,13,14,29].
Общая возрастная тенденция проявляется в снижении относительного количества волокон I типа. Наиболее выражены эти изменения в четырехглавой мышце бедра.
Другой четко проявляющейся закономерностью является неуклонное возрастание количества волокон с гликолитическим типом энергообеспечения. Наиболее выражены эти изменения в трехглавой мышце плеча и в четырехглавой мышце бедра. При этом, если иметь в виду, что волокна I типа обладают преимущественно окислительными механизмами энергообеспечения, а волокна типа ПВ — анаэробно-гликолитическими, то можно представить, что описанная в ряде работ [15,26,27,28] возрастная тенденция, связанная с увеличением роли анаэробно-гликолитического энергообеспечения, находит свое подтверждение и в особенностях перераспределения спектра мышечных волокон.
Таким образом, выявляются две волны перестройки энергетики скелетных мышц в период полового созревания — на первой стадии некоторое увеличение аэробной энергетики, а затем, к последней стадии пубертатного периода, резкое увеличение активности анаэробно-гликолитических энергетических источников. Это во многом определяется перестройкой спектра волокон в скелетных мышцах человека. Общая тенденция (за исключением 14-летнего возраста) постепенного увеличения с возрастом доли быстрых волокон с анаэробно-гликолитическим энергообеспечением в еще большей степени выявляется при анализе размеров мышечных волокон. Полученные нами результаты показывают еще более выраженную динамику снижения общей площади, занимаемой волокнами I типа от 7 к 17 годам и значительное возрастание представительства волокон типа ПВ. Это в значительной степени связано с различиями в темпах роста поперечника мышечных волокон I и II типов. Если в 7-8-летнем возрасте МВI характеризуются большей величиной суммарной площади поперечного сечения, то в пубертатный период отмечается резкое увеличение темпов роста поперечника волокон II типа, особенно МВПВ, что является одной из причин значительной перестройки энергетического профиля исследованных мышц. В четырехглавой мышце бедра наряду с увеличением площади мы видим и одновременное увеличение числа МВПВ. Можно полагать, что изменение соотношения количества МВ идет только за счет интенсивной передифференцировки волокон в возрасте 14-16 лет. Как было сказано выше, все эти процессы в значительной степени контролируются мужскими половыми гормонами [13,17,25,40].
Роль пубертатного периода в перестройке энергетики мышечных волокон в еще большей степени проявилась при исследовании возрастных изменений
— 86 —
активности СДГ. Как указывалось выше, активность этого фермента, как маркера, характеризующего митохондриальное окисление, широко используется в практике гистохимических исследований мышечной ткани. Проведенные нами исследования показали, что СДГ-активность в различных мышечных волокнах исследованных мышц претерпевает в возрастном диапазоне от 7 до 17 лет неоднозначные изменения.о11шк [41], и с тех пор считается прочно устоявшимся в физиологии спорта. В то же время, различия волоконного состава мышц, установленные с использованием биопсии у спортсменов разной специализации (особенно при сопоставлениях бегунов на короткие и длинные дистанции), еще не дают возможности утверждать, что эти различия были исходными -не исключена возможность, что они возникли в ходе адаптации к соответствующим нагрузкам. Об этом свидетельствует огромный экспериментальный материал, показывающий большую лабильность различных типов мышечных волокон в результате воздействия как эндогенных, так и экзогенных факторов [7,17,24,23,36,40,42,44,48]. Такие эксперименты, как перекрестная иннервация «быстрой» и «медленной» мышц реципрокными нервами, электрическая стимуляция нерва, идущего в быстрой мышце с частотой импульсов, характерных для нерва к медленной мышце (10Гц) и наоборот, приводят к перепрограммированию фенотипов скелетных мышц по признаку скорости сокращения [36,38,39,47], что связано со сменой синтезируемых миозинов. Согласно некоторым литературным данным, имеет место и обратное влияние — от мышечного волокна на дифференциацию мотонейрона [20,38]. Смена фенотипических признаков скелетной мышцы наблюдается и при действии других факторов, введение различных гормонов, изменение характера питания, воздействие различных физических нагрузок [36,37,40,42,43,47,48,49].
Различный уровень физической нагрузки на мышцу вызывает изменение гистохимических характеристик и структуры скелетной мышцы [33,34,43,45] и приводит к развитию неоднозначных изменений в скелетных мышцах. При этом разные типы мышечных волокон неодинаково реагируют на физическую нагрузку, что особенно показательно для белых гликолитических мышечных волокон [33]. О широких возможностях взаимного перехода между различными типами мышечных волокон свидетельствуют эксперименты с бета-гуанидинпропионовой кислотой. Систематическое введение этого вещества блокирует синтез креатина и способствует вымыванию его из организма. Известно, что креатин играет очень большую роль в энергетике мышечных волокон. Однако этим не исчерпывается его значение для мышечной ткани. На определенной стадии эмбрионального развития креатин прямым воздействием на геном мышечных ядер активирует
— 87 —
синтез молекул миозина, креатинкиназы и ферментов анаэробного гликолиза [18,20]. Оказалось, что влияние креатина на синтетические процессы в мышечном волокне поддерживается и во взрослом состоянии. Вызванный действием бета-гуанидинпропионовой кислоты дефицит креатина в мышечной ткани приводит у взрослых животных к снижению относительного количества волокон ПВ и к увеличению числа волокон I типа [20]. Это свидетельствует о способности креатина индуцировать переход волокон ПВ в тип I.
Размах фенотипического выражения отдельных признаков, характеризующих различные типы мышечных волокон, при воздействии различных экзогенных и эндогенных факторов, хорошо объясняется в терминах адаптивных вариантов признака и связано с рабочими (эргонтическими) корреляциями по И.И.Шмаль-гаузену [32], при наличии которых происходит согласованная перестройка функционально связанных частей. Рабочие корреляции касаются тонких структур, обеспечивающих функционирование органов и систем, «значительно повышают пластичность организма и позволяют ему с гораздо большей скоростью перестраиваться в целом соответственно требованиям изменяющейся среды. Изменение относительного содержания различных типов мышечных волокон разных типов подтверждается данными об отсутствии отличий в транскриптатах белков мышечных волокон. Допускается, что реализация изменений в содержании мышечных волокон различных типов происходит через посредство нервных элементов нервно-мышечной системы, и, таким образом, эти изменения следует рассматривать в рамках вопроса о нейротрофическом контроле скелетной мышцы [7,18,38,42,47].
Как показано в экспериментах Н.П.Резвякова [20] с использованием различных физических нагрузок, с дефицитом белка в поедаемом корме, с аппликацией колхицина на двигательный нерв, дефинитивное мышечное волокно не окончательно детерминировано быть быстрым или медленным, и эта серия факторов может приводить к серьезным изменениям характеристик фазных волокон. Это свойство и выражает пластичность мышечных волокон. По мнению многих авторов [20,24,], полиморфизм мышечных волокон и их способность к адаптивным пластическим перестройкам объясняется не только полиморфизмом миозинов и других контрак-тильных белков, синтезируемых в одном мышечном волокне [20], но и лабильностью энергетического метаболизма. Признание факта сосуществования в одном мышечном волокне различных миозинов и разрешенность их транскрипции позволяет объяснить трансформации мышечных волокон, и это свидетельствует о динамичности фенотипа мышечных волокон, его постоянном подстраивании к меняющимся условиям функционирования скелетной мышцы [43,48].
Наш экспериментальный материал еще раз доказывает возможности перехода между волокнами различных по функциональной специализации типов. Это свидетельствует, что дифференцировки в скелетных мышцах отличаются от сходных процессов в других тканях достаточно легкой обратимостью, и соответственно лучшей способностью приспособления к меняющимся условиям функционирования. Основные процессы перестройки мышечных волокон проходят в период полового созревания. При этом появление дефинитивных форм организации
— 88 —
энергетики мышечных волокон связано лишь с последней стадией пубертатного процесса и, по-видимому, эти изменения определяют окончательное становление периферического эффекторного аппарата двигательной системы.
Ряд последовательных передифференцировок, очень схожих по своим проявлениям у всех исследованных нами объектов, свидетельствует о наличии общих закономерностей развития скелетных мышц в постнатальном онтогенезе. Внутренний механизм, обуславливающий их периодические проявления, остается пока неясным. Очевидно, что немалую роль в этом играют возрастные изменения мотонейронов спинного мозга, осуществляющих трофические влияния на соответствующие ДЕ. Можно представить, что по ходу онтогенеза центральные и периферические нейро-гуморальные влияния перестраивают мотонейроны, которые в свою очередь вызывают своими трофическими воздействиями изменения в мышечных волокнах.
Значительно более важным представляется анализ факторов, определяющих возможность осуществления всех этих изменений периферического двигательного аппарата, включающего в себя ДЕ, формирующие ту или иную мышцу, и соответствующие двигательные ядра спинного мозга. Происходящие при этом возрастные изменения мышечной ткани можно подразделить на постепенные (градуальные), которые можно видеть на примере роста мышечных волокон, и быстрые (дискретные), в виде волн последовательных передифференцировок, резко меняющих организацию мышечных волокон. Первые связаны с функционированием синтетического аппарата (эргастоплазмы) мышечных волокон на уже подготовленных матрицах, вторые — определяются активацией ядерного аппарата, появлением новой РНК и изменением вида синтезируемых молекул. Сигнал о необходимости очередной перестройки двигательных единиц можно получить нейрогенно, как при трофических влияниях с синаптических окончаний нервного волокна, так и гуморально, с более широким гормональным воздействиями. Нейрогенные влияния приходят, очевидно, сверху и их очередность связана с созреванием нервных центров, определяющих возрастные особенности и координацию двигательной деятельности. Известно, что организация управления движениями очень сложна и занимает у млекопитающих длительный отрезок постнатального онтогенеза, включающий и половое созревание (и даже некоторое время и после него). Примерно такое же время приходится и на созревание мышечной ткани. Можно представить, что эти два процесса в какой-то степени сопряжены. При этом следует обратить внимание на то, что система управления движениями разделена на дискретные уровни, определенным образом взаимодействующие между собой, но четко проявляющиеся в филогенезе двигательной активности у позвоночных, при нервной патологии и в процессах онтогенеза. В своей работе Н.А.Бернштейн [3] выделяет несколько уровней управления двигательной деятельностью.
Самый нижний уровень «А», рубро-спинальный, у высших позвоночных активность этого уровня проявляется только при распределении мышечного тонуса, с ним связано появление тремора и других типов мышечной дрожи. Интересно, что у незрелорождающихся крысят способность к проявлению терморегу-
— 89 —
ляционного мышечного тонуса и дрожи наблюдается только с 11-12-дневного возраста [8,9,10], что дает возможность полагать, что только к этому сроку у крысят складывается функционирование уровня «А». Необходимо напомнить, что именно в эти сроки начинается формирование в смешанных мышцах (пока на стадии промежуточного волокна) тонических волокон I типа. Можно полагать, что это начало первой волны мышечных дифференцировок, связанной с прорастанием (миэлинизацией) рубро-спинального пути и появлением нижнего уровня регуляции.
Следующий уровень «В» — таламо-паллидарный. Уровень синергий, ритмических движений и штампов. В чистом виде функционирует у рыб, определяя плавательные, синхронные движения мускулатуры всего тела. У незрелорождаю-щихся крысят этот уровень начинает функционировать почти одновременно с уровнем «А». Зрелорождающаяся морская свинка уже с момента рождения проявляет все признаки функционирования уровня «В». У ребенка первые признаки активности этого уровня также проявляются с первого дня после рождения и полностью формируются к 5-6-месячному возрасту. Эфферентные влияния с globus pallidus так же, как и при функционировании рубро-спинального уровня, осуществляются через красное ядро, поэтому активация этого уровня не ведет к существенным переменам характера нисходящих влияний на мотонейроны спинного мозга. По всей вероятности, поэтому функционирование уровня «В» не связано со значительными перестройками периферических ДЕ.
Уровень «С», пирамидно-стриальный, или уровень пространственного поля, включает полосатое тело — striatum и двигательную кору, эффекторные клетки которой образуют кортико-спинальный (пирамидный) путь, заканчивающийся на мотонейронах спинного мозга. С появлением новых нисходящих влияний связана вторая волна мышечных дифференцировок и образование на втором году жизни волокон II типа, необходимых для осуществления сложных локомоций, определяющих возможность удержания позы стояния и элементарной ходьбы. У крысят эти изменения наступают к 16 дню жизни.
Дальнейшее развитие кортикальных уровней регуляции связано с привлечением других зон коры — премоторной, ассоциативно-теменной, а в лобных долях полей 10, 11 и чисто человеческих лобных полей 44 и 47. Все они являются морфологическим субстратом для новых, все более совершенных уровней регуляции движений и осуществления психических функций. Н.А.Бернштейн [3] отмечает, что в пубертатном периоде происходит даже некоторое «выпячивание» пирамидно-коркового аппарата в ущерб экстрапирамидным фоновым уровням, отсюда наблюдаемая угловатость, неловкость глобальных движений у подростков, неустойчивость тонуса. Отсюда деавтоматизация движений и большая утомляемость. В то же время, именно на этот возрастной период приходится коренная перестройка периферического двигательного аппарата, формирование дефинитивной организации периферических двигательных единиц и морфофункционального состояния мышечных волокон. С течением времени эти диспропорции выравниваются и устанавливается индивидуальный психомоторный профиль взрослого человека.
— 90 —
Таким образом, поуровневое дискретное созревание центральных нервных регуляционных механизмов приводит к последовательному (поэтапному) развитию моторной функции, проявляющейся и в дискретных этапах развития мышечной ткани.
Наличие закономерно сменяющих друг друга этапов возрастного развития должно определить наличие определенных возрастных закономерностей развития энергетики скелетных мышц.
СПИСОК ЛИТЕРАТУРЫ
1. Аршавский И.А. Роль энергетических факторов в качестве ведущих закономерностей онтогенеза. // Киев: Наукова Думка, 1972. — С.43-72.
2. Безобразова В.Н., Догадкина С.Б. Особенности центрального и периферического кровообращения юношей 16-17 лет с разными типами мануальной асимметрии. // Физиология школьника юношеского возраста. — 1987. -С. 145-153.
3. Бернштейн Н.А. Очерки по физиологии движений и физиологии активности. // М.: Медицина, 1965. 349 с.
4. Буйкис И.М. Гистохимия дегидрогеназ развивающегося спинного мозга. // Рига, 1975. 230 с.
5. Глезер И.И. Количественный анализ роста и развития пирамидных клеток коры лобной доли в постнатальном онтогенезе. // Автореф.дис…канд.биол.наук. -М. — 1959. — 22 с.
6. Данилов Р.К. Дивергентная дифференцировка в эмбриональном гистогенезе скелетной мышечной ткани. // Архив анатомии, гистологии и эмбриологии. -1985. — №10. — С.77-82.
7. Итина Н.А. Влияние мотонейронов на функциональные характеристики скелетных мышц позвоночных. Эффекты денервации и перекрестной реинерва-ции. // Успехи современной биологии. — 1973. — Т.75. — №3. — С.419-440.
8. Корниенко И.А., Маслова Г.М., Гохблит И.И. О роли бурого жира в химической терморегуляции на различных стадиях онтогенеза крыс. // Эволюционная биохимия и физиология. — 1971. — Т.7. — №4. — С.285-391.
9. Корниенко И.А. Возрастные изменения энергетического обмена и терморегуляции. // М.: 1979. 157 с.
10. Корниенко И.А. Возрастные изменения энергетического обмена. // Авто-реф.дис….д-ра биол.наук. — М. — 1980. — 54 с.
11. Корниенко И.А., Сонькин В.Д. «Биологическая надежность», онтогенез и возрастная динамика мышечной работоспособности // Физиология человека. -1999. — Т.25. — №1. — С.98.
12. Корниенко И.А., Сонькин В.Д., Тамбовцева Р.В. Возрастное развитие скелетных мышц и физической работоспособности. // Физиология развития ребенка: теоретические и прикладные аспекты. М., 2000. С.209.
13. Корниенко И.А., Сонькин В.Д., Тамбовцева Р.В. Возрастная периодизация развития скелетных мышц в онтогенезе человека. // Новые исследования. Альманах. — 2001. — №1. — С.44.
— 91 —
14. Корниенко И.А., Сонькин В.Д., Тамбовцева Р.В. Возрастное развитие энергетики мышечной деятельности: Итоги 30-летнего исследования. Сообщение 1. Структурно-функциональные перестройки. // Физиология человека. — 2005. -Т.31. — №4. — С.402-406.
15. Корниенко И.А., Сонькин В.Д., Тамбовцева Р.В. Возрастное развитие энергетики мышечной деятельности: Итоги 30-летнего исследования. Сообщение 3. Эндогенные и экзогенные факторы, влияющие на развитие энергетики скелетных мышц. // Физиология человека. — 2007. — Т.33. — №6. — С.94-99.
16. Механизмы контроля мышечной деятельности. // Под ред. Ушакова В.Б., Л.: 1985, 255с.
17. Мусаева З.Т., Тамбовцева Р.В. Влияние половых гормонов на развитие биоэнергетики скелетных мышц. // V Всес.конф.по биохимии мышц. Телави. -1985. — С.156.
18. Наследов Г.А. Некоторые вопросы нейротрофической регуляции свойств скелетной мышцы // В кн.: Механизмы контроля мышечной деятельности. Л.: Наука, 1985. — С.209.
19. Онтогенез млекопитающих в невесомости. // Под ред. О.Г.Газенко. М.: Наука, 1988. 178 с.
20. Резвяков Н.П. Общие закономерности дифференцировки и пластичности скелетных мышц. // Автореф.дис….д-ра мед.наук. — Казань. — 1982. — 33с.
21. Рехачева И.П. Возрастные особенности активности некоторых ферментов в развивающихся мышечных волокнах. // Архив, анатомии, гистологии, эмбриологии. — 1981. — Т.81. — №10. — С.77- 88.
22. Рехачева И.П., Сапроненкова И.Н., Суфарина С.В. и др. Возрастные изменения активности сукцинатдегидрогеназы в функционально различных мышцах у молодых крыс // Архив анатомии, гистохимии, эмбриологии. — 1984. — Т.87. — №2. — С.79-82.
23. Рехачева И.П., Катинас Г.С., Сапроненкова И.П. Изменение активности сукцинатдегидрогеназы в мышечных волокнах функционально различных мышц у крыс после снижения силовой нагрузки — гиподинамии. // Архив анатомии, гистологии и эмбриологии. — 1985. — Т.59. — №9. — С.49-55.
24. Румянцев П.П., Ерохина И.Л. Морфологические аспекты дифференциров-ки и пролиферации в гистогенезе скелетных, сердечной и гладких мышц позвоночных. // В кн.: Проблемы миогенеза. — Л.: Наука, 1981, С.22-50.
25. Сонькин В.Д. Энергетическое обеспечение мышечной деятельности школьников. // Автореф.дис….д-ра биол.наук. — М. — 1990. — 50с.
26. Тамбовцева Р.В., Корниенко И.А. Развитие различных мышечных волокон четырехглавой мышцы бедра и камбаловидной мышцы в онтогенезе человека. // Архив анатомии, гистологии и эмбриологии. — 1986. — Т.91. — №9. — С.96.
27. Тамбовцева Р.В., Корниенко И.А. Развитие различных типов мышечных волокон камбаловидной мышцы в постнатальном онтогенезе крысы. // Архив анатомии, гистологии и эмбриологии. — 1986. — Т.93. — №7. — С.77.
28. Тамбовцева Р.В., Корниенко И.А. Развитие мышечных волокон разного типа в постнатальном онтогенезе морской свинки. // Архив анатомии, гистологии и эмбриологии. — 1987. — Т.93. — №7. — С.55.
— 92 —
29. Тамбовцева Р.В. Возрастные и типологические особенности энергетики мышечной деятельности. // Автореф.дис….д-ра биол.наук. — М. — 2002. — 48с.
30. Фарбер Д.А., Корниенко И.А., Сонькин В.Д. Физиология школьника. // М.: Педагогика, 1990, 64с.
31. Харрисон Дж., Уайнер Дж, Таннер Дж. Биология человека. // М.: Мир, 1968, 422с.
32. Шмальгаузен И.И. Рост и дифференцировка. // Избранные труды в 2х томах. Киев, 1984, С.176-186.
33. Язвиков В.В. Состав скелетно-мышечных волокон мышц конечностей человека и способность к выполнению различных видов физической работы. // Автореф.дис….д-ра мед.наук. — М. — 1991. — 48с.
34. Яковлев Н.Н., Макарова Т.Н. Функциональная и метаболическая дифференциация волокон скелетной мышцы. // Физиологический журнал СССР. -1980. — Т.66. — №8. — С.1129-1144.
35. Bell R.D., MacDougall J.D., Billeter R.et al. Muscle fiber types and morphomet-ric analysis of skeletal muscle in six-year-old children.// Med. Sci. Sports Exerc. -1980. — V.12. — P.28.
36. Cullen M.J., Harris J.B., Marshall W.M. An electrophysiological and morphological study of normal and denervated chicken latissimus dorsi muscles. // J.Physiol. -1975. — P.371-385.
37. Colling-Saltin A-S. Skeletal muscle development in the human fetus and during childhood. / Children and exercise / Eds. Berg K., Eriksson B.O. Baltimore (MD). // University Park Press, 1980. — P. 193.
38. Dux L., Dux E. A non-neurol regulatory effect of the metabolic differentiation of the skeletal muscles effect of castration and testosterone administration the skeletal muscle of the rat. // Comp. Biochem. And Physiol. — 1970. — V.64. — N1. -P.177-183.
39. Tccles J.C., Eccles R.M., Lundbog A. The action potentialis of the alpha moton-eurones Supplying fast and slow muscle. // J.Physiol. — 1958. — V.142. — P.275-291.
40. Elder G.C.B., Kakulas B.A. Histochemical and contractile property changes during human development. // Muscle Nerv. — 1993. — V.16. — P1246.
41. Gollnick P.D., Hodgson D.R. The identification of fiber types in skeletal muscle: acontinual dilemma. // Med.Sci in Sport. — 1986. — V.14. — P.81-108.
42. Guth L., Samaha F.J., Albers R.W. The neurol regulation of some phenotypic differences between the fiber types of mammalian skeletal muscle. // Exptl.neurol. -1970. — V.26. — N1. — H.126-135.
43. Gutman E., Hanzlikova-La Dar V. Effect of androgens on histochemical fibre type. // Histochemie. — 1970. — V.24. — P.287-291.
44. Hinriksson L.K. Distribution, number and size of different types of fibres in uhole eross Sections of femore m.tibiales anterior. An enzyme histochemical stady. // Acta physiol Scand. — 1985. — V.123. — N3. — P.229-235.
45. Hinriksson L.K., Friden J. Distribution of fibre sizes in human skeletal muscle. An enzyme Histochemical study in m.tibial anterior. // Acta Physiol Scand. — 1985. -V.123. — N2. — P.171-177.
-93-
46. Jeffrey P.L., Austin L. Axoplasmie transport. // Progr.Neurobiol. — 1973. -N2. — P.107-255.
47. Landon D.N. Skeletal muscle-normal morphology: development and innervation. // In: Skeletal muscle pathology. London. — 1982. — P.8-121.
48. Larsson L., Sjodin B., Karlsson J. Histochemical and biochemical changes in human skeletal muscle with age in sedentary males age 22-65 yars. // Acta Physiol. Scand. Scand. — 1978. — V.103. — N1. — P.31-39/
49. Lexell J., Sjostrom M., Nordlund A.-S. Growth and development of human muscle: a quantitative morphological study of whole vastus lateralis from childhood to adult age. // Muscle Nerve. — 1922. — V. 15. — P. 404.
50. Paulson J.C., McClure W.O. Microtubules and axoplasmic transport. // Rrain Rec. — 1974. — V. 73. — N2. — P.333-337.
51. Platzer A.C. The ultrastructure of normal myogenesis in the limb of the mouse. // Anat.Rec. — 1978. — V.190. — N3. — P.639-657.
-Q-
Опровергнуты представления о механизме утраты тонуса мышц
Долгое время физиологи были уверены, что длительное снижение физической активности или утрата мышечного тонуса в силу травм, болезней или иных естественных причин приводит к необратимой атрофии мышц. Профессор Лоуренс М. Шварц (Lawrence M. Schwartz) из Университета Массачусетса опроверг эту аксиому, сообщается в журнале Frontiers in Physiology.
Скелетные мышцы – уникальное формирование, их волокна состоят из наиболее крупных клеток в организме – синцитиев. Это гигантские многоядерные клетки, образующиеся в результате многочисленных слияний одноядерных клеток. Некоторые волокна могут достигать 600 мм в длину и в 100 000 раз превосходить по объему одноядерные клетки. Долгое время бытовала так называемая «гипотеза ядер мышечных клеток», согласно которой каждое ядро в синцитии поддерживало определенный максимальный объем цитоплазмы, а в процессе роста или атрофии волокон количество ядер изменялось соответственно. Но если рост волокна сопровождался включением дополнительных ядер из стволовых клеток, то однозначного ответа о том, что происходит с ядрами клеток мышц при атрофии, не было. Главенствовало представление об их утрате мышцами в результате апоптоза.
Несколько групп исследователей недавно продемонстрировали, что уменьшение объема мышечной ткани характерное для атрофии не сопровождается уменьшением числа ядер мышечных клеток. Для этого Дадди и коллеги в экспериментах in vitro изолировали мышечные волокна мышей и содержали их в состоянии покоя в условиях приближенных к физиологическим в течение 14 дней. Из изолированных волокон выделяли белковую фракцию и оценивали изменение содержания белка с помощью флуоресцентной микроскопии по количеству белка f-актина. Ученые заметили, что его содержание уменьшается на 35 процентов в неделю. Количество ядер в делящихся и покоящихся клетках проследили с помощью иммуногистохимического окрашивания мышечных волокон. Оказалось, что при уменьшении количества белка и, соответственно, объема мышечных волокон утраты ядер мышечных клеток не происходит.
В более ранних исследованиях автора статьи, профессора Лоуренса Шварца, показаны аналогичные результаты. В качестве модели в этих экспериментах использовались межсегментные мышцы табачного бражника Manduca sexta. Эти гигантские мышцы обеспечивают способность личинок бабочки ползать, но подвергаются программированной гормональной атрофии с 15 по 18 день превращения личинки во взрослое насекомое. Проведя измерения поперечных размеров волокон межсегментных мышц бражника, ученые обнаружили, что потеря мышечной массы в ходе программируемой атрофии составила 49 процентов, но количество ядер мышечных клеток осталось неизменным.
Противоположный процесс – появление дополнительных ядер в процессе роста мышц — был ранее неоднократно доказан. Чаще всего источником дополнительных ядер становится определенный тип стволовых клеток мышц – клетки-спутницы. Их слияние с волокнами скелетных мышц приводит к увеличению количества ядер, необходимому для поддержания метаболических нужд растущей мышцы и ее способности к сокращению.
Недавние наблюдения, свидетельствующие о том, что даже при уменьшении объема скелетных мышц и их атрофии, способность мышц к сохранению и восстановлению физической активности «депонируется» представляют большое значение для здравоохранения. Фактически это обозначает необходимость формирования новой культуры восприятия физической активности, прививаемой с раннего детства, когда возможности для создания долговременного мышечного ресурса оптимальны. Ресурс, приобретенный в детстве, способен продлить физическую активность человека в зрелом возрасте и даже предотвратить инвалидизацию и тяжелый ущерб здоровью, вызванный травмами, падениями и переломами.
Екатерина КорзеневаМышечные ткани — Ченцов Ю.С.
Год выпуска: 2001Автор: Ю.С. Ченцов
Жанр: Гистология
Формат: DjVu
Качество: Отсканированные страницы
Описание: В современной медицине все больший интерес вызывают вопросы лечения и профилактики сердечно-сосудистых заболеваний, возникновение которых в значительной мере связано с нарушением структуры и функций мышечных тканей (атеросклероз, инфаркт миокарда, гипертензия, астма и др.). Патология беременности и родов также связана с нарушениями деятельности и строения мускулатуры миометрия. Повышенная мышечная нагрузка и гипокинезия, а также ряд заболеваний (миастения, дистрофия Дюшенна и др.) обусловлены в значительной степени изменениями в соматической мускулатуре. В связи с необходимостью более глубокого изучения этиологии и патогенеза заболеваний мышечной системы, познания механизмов, лежащих в основе этих состояний, возрос интерес к фундаментальным исследованиям структурно-функциональных особенностей и биохимии мышечных тканей. Это и побудило авторов к написанию данной книги. Ее содержание легло в основу расширенного курса лекций, читаемых в университетах на биологических и медицинских факультетах для студентов старших курсов в течение многих лет.
В книге «Мышечные ткани» излагаются современные сведения о микроскопическом и ультрамикроскопическом строении, развитии и регенерации мышечных тканей, их физиологических и биохимических особенностях и специфических изменениях этих тканей при некоторых патологических состояниях. Приведены также сведения по сравнительной гистологии и рассмотрены вопросы эволюции мышечных тканей.
Книга «Мышечные ткани» написана опытными преподавателями, содержит как данные отечественной и зарубежной литературы, так и оригинальные материалы; снабжена многочисленными микрофотографиями, схемами и графиками.
Часть первая, посвященная микроскопической и электронно-микроскопической организации мышечных тканей, их гистогенезу, регенерации, а также вопросам сравнительной гистологии и эволюции, написана доктором биологических наук, профессором кафедры цитологии и гистологии биологического факультета МГУ Е.А. Шубниковой. Глава по гладкой мускулатуре написана в соавторстве с доктором медицинских наук, членом-корреспондентом РАЕН, заведующим кафедрой гистологии и эмбриологии медицинского факультета Российского университета дружбы народов профессором Н.А. Юриной. Ими же в соавторстве с кандидатом биологических наук, заведующей лабораторией морфологического сканирования и пластинации Института морфологии человека РАМН Г.Б. Большаковой изложены вопросы патологической гистологии поперечнополосатой мускулатуры. Вторая часть, в которой рассматриваются физиология и патофизиология мышечных тканей, написана доктором биологических наук, ведущим научным сотрудником кафедры физиологии человека и животных биологического факультета МГУ О.П. Балезиной. В третьей части пособия, написанной доктором биологических наук, профессором кафедры биохимии биологического факультета МГУ Н.Б. Гусевым, рассматриваются основы биохимии мышечных тканей. Цветные иллюстрации микропрепаратов выполнены Г.Б. Большаковой.
Содержание учебного пособия
«Мышечные ткани»
Строение, развитие, сравнительная и патологическая гистология мышечных тканей
Общая характеристика и классификация мышечных тканей
Соматические (скелетные) мышечные ткани
- Общая характеристика соматической поперечнополосатой (исчерченная, локомоторная, скелетная) мышечной ткани позвоночных животных
- Ультраструктура поперечнополосатого мышечного волокна и основные белки миофибрилл
- Саркоплазматическая сеть и Т-система
- Неспецифические органеллы мышечного волокна
- Тоническая и фазная мышечные ткани позвоночных животных
- Иннервация мышечных волокон
- Исчерченные мышечные ткани нелокомоторного аппарата
- Интрафузальные мышечные волокна, или мышечные веретена
- Дифференцировка мезодермы и ее производных
- Гистогенез поперечнополосатой соматической мышечной ткани (стриомиогистогенез)
- Регенерация и патологическая гистология поперечнополосатой мышечной ткани
- Элементы сравнительной гистологии скелетной поперечнополосатой мышечной ткани
- Скелетная поперечнополосатая мышечная ткань позвоночных животных
- Круглоротые и рыбы
- Земноводные
- Пресмыкающиеся
- Птицы
- Млекопитающие
- Локомоторные мышцы беспозвоночных животных и низших хордовых
- Низшие беспозвоночные
- Черви
- Моллюски
- Членистоногие
- Вторичноротые (иглокожие и низшие хордовые)
- Скелетная поперечнополосатая мышечная ткань позвоночных животных
- Рабочие кардиомиоциты
- Развитие сердца и кардиомиогенез
- Регенерация и патологическая гистология сердечной мышечной ткани
- Элементы сравнительной гистологии сердечной мышечной ткани
- Кардиомиоциты птиц, рептилий и низших позвоночных животных
- Кардиомиоциты низших хордовых и беспозвоночных животных
- Гладкая мышечная ткань позвоночных животных
- Строение гладкой мышечной ткани и ее классификация
- Иннервация гладкой мышечной ткани и передача возбуждения
- Развитие гладкой мышечной ткани (лейомиогистогенез)
- Регенерация и патологическая гистология гладкой мышечной ткани
- Элементы сравнительной гистологии гладкой мышечной ткани
- Висцеральная мышечная ткань позвоночных животных
- Висцеральная мышечная ткань беспозвоночных животных
Физиология мышечных тканей
Клеточные механизмы запуска мышечного сокращения
- Внутриклеточный метаболизм ионов Са2+ и методы его анализа
- Уровень ионизированного кальция в покое
- Механизмы, поддерживающие низкий уровень [Са2+]вн в покое
- Повышение уровня [Са2+]вн при сокращении
- Источники повышения содержания внутриклеточного кальция в мышечных клетках и волокнах при сокращении
- Участие электрической активности мембраны в сокращении мышечных клеток и волокон
- Особенности потенциала покоя мышечных клеток и волокон
- Особенности генерации потенциала действия в разных типах мышечных клеток и волокон
- Натриевые и кальциевые каналы мышечных клеток и волокон
- Механизмы электромеханического сопряжения в разных типа мышечных клеток и волокон
- Первично-мышечные миопатии
- Миодистрофии
- Вторично-мышечные (нейрогенные) миопатии
Сочетательный аппарат мышц
- Структурная организация сократительного аппарата поперечнополосатых мышц
- Структура и свойства миозина
- Структура и свойства актина
- Актинсвязывающие белки
Регуляция взаимодействия ангина и миозина на уровне сократительного аппарата
Рекомендуемая литература
Физиология, мышцы — StatPearls — Книжная полка NCBI
Введение
В человеческом теле встречаются три основных типа мышц: скелетные, сердечные и гладкие. Каждый тип мышц имеет уникальные клеточные компоненты, физиологию, определенные функции и патологию. Скелетная мышца — это орган, который в первую очередь контролирует движение и осанку. Сердечная мышца охватывает сердце, которое поддерживает человеческое тело. Гладкая мускулатура присутствует во всей желудочно-кишечной, репродуктивной, мочевой, сосудистой и дыхательной системах.
Клеточная
Скелетные мышцы составляют примерно 40% от общей массы тела человека. В его состав входит множество отдельных волокон, которые объединены в мышечное веретено; это то, что придает скелетным мышцам полосатый вид. Одно мышечное волокно состоит в основном из актиновых и миозиновых волокон, которые покрыты клеточной мембраной (сарколемма). Эти волокна являются функциональной единицей органа, ведущей к сокращению и расслаблению. Есть две основные классификации скелетных мышц; к ним относятся Тип I (медленное окисление) и Тип II (быстро сокращающийся).Огромное разнообразие скелетных мышц приводит к различиям в скорости и продолжительности сокращений в разных группах мышц в зависимости от их конкретной функции [1].
Сердечная мышца или миокард — это непроизвольно поперечно-полосатая мышца, охватывающая камеры сердца. Он состоит из отдельных кардиомиоцитов, которые по структуре похожи на скелетные мышцы. Каждый кардиомиоцит содержит цитоскелетные и сократительные элементы, все из которых связаны между собой вставными дисками.Это хорошо прилипающие комплексы, которые позволяют клеткам сердечной мышцы получать быструю электрическую передачу и сокращаться как единое целое. [2] Сердечная мышца также содержит специализированные клетки кардиостимулятора, которые находятся в миокарде. Эти клетки позволяют сердечной ткани внутренне деполяризоваться без внешних стимулов. [3]
Клетки гладкой мускулатуры также состоят из актиновых и миозиновых волокон; однако они расположены в виде листов, а не веретен, что придает этому типу мышц гладкий вид.Эти клетки присутствуют в стенках многих органов, таких как легкие, желудочно-кишечный тракт, репродуктивные органы, кровеносные сосуды и даже кожа. [4]
Функция
Мышцы человеческого тела, будь то скелетные, сердечные или гладкие, создают силу и движение. Мышцы скелета поддерживают кости, чтобы поддерживать осанку, а также контролировать произвольные движения. Скелетные мышцы также способствуют метаболизму и хранению энергии. Сердечная мышца продвигает кровь и приводит к надлежащему насыщению кислородом и поддержанию каждой клетки, составляющей человеческое тело.Гладкая мышца расположена по всему телу и использует сократительную силу для укорачивания и продвижения различного содержимого через просвет множества систем органов, в которых она задействована.
Механизм
Потенциалы действия нервных волокон центральной нервной системы деполяризуют мышцу по длине сарколеммы к самым внутренним волокнам через систему поперечных канальцев (Т-канальцев). Потенциал действия отвечает дигидропиридиновым рецептором на Т-канальце; это действует как датчик напряжения, позволяющий высвобождать кальций.Впоследствии кальций активирует рианодиновые рецепторы в саркоплазматическом ретикулуме, чтобы высвободить еще больше кальция. Затем более высокие количества кальция могут связываться с протеином тропонином, который расположен на актиновых филаментах. Кальций-тропониновый комплекс вытесняет белок тропомиозин из активного центра актиновой нити и обеспечивает связывание миозина и сокращение мышц. Аденозинтрифосфат (АТФ) необходим для отделения миозина от актиновых волокон и обеспечения расслабления мышц. [1]
Подобно скелетным мышцам, сердечная мышца запускается связыванием кальция с тропонином в актиновых филаментах кардиомиоцитов.Это связывание затем удаляет тропомиозин и делает возможным связывание миозина с актиновыми филаментами и возможное сокращение. Существенная разница между сердечной и скелетной мышцами заключается в автоматизме кардиомиоцитов. Специализированные кардиостимуляторы, расположенные в синоатриальном (SA) узле, отвечают за сокращение сердечной мышцы. Они запускают потенциалы действия, которые обеспечивают приток натрия и калия, а также высвобождение кальция из саркоплазматической сети. Тогда сердечная мышца может сокращаться как единое скоординированное целое.[5]
Сокращение гладких мышц не находится под произвольным контролем и осуществляется за счет вегетативной регуляции взаимодействия кальция и кальмодулина. Сокращение начинается за счет изменения потенциала действия или активации механических рецепторов растяжения в плазматической мембране. Внутриклеточный кальций увеличивается и соединяется с белком кальмодулином. Именно этот комплекс активирует киназу легкой цепи миозина (MLC) для фосфорилирования и образования перекрестных мостиков между миозином и актином, что приводит к сокращению мышц.Некоторые гладкие мышцы поддерживают тонус, что обусловлено постоянным уровнем фосфорилирования при отсутствии внешних потенциалов. Снижение уровня внутриклеточного кальция вызывает расслабление. [4]
Клиническая значимость
Мышечная дистрофия — это прогрессирующая генетическая миопатия, которая приводит к нарушению нормальной анатомии и физиологии клеток скелетных мышц. Полное или частичное отсутствие белка дистрофина является патологическим механизмом мышечной дистрофии Беккера и Дюшенна.Дистрофин — это белок, связанный с филаментами скелетных мышц. Дистрофин обеспечивает структуру и поддержку сарколеммы моноволокна. Недостаток белка дистрофина приводит к повреждению поддерживающей сарколеммы, слабости и, в конечном итоге, атрофии здоровых мышечных волокон. Мышечная дистрофия Дюшенна поражает до 1 из 3600 мальчиков, что делает ее самой распространенной среди всех типов мышечных дистрофий. Многие с Дюшенном имеют низкую продолжительность жизни, потому что в настоящее время лечение недоступно.Лечение этих расстройств носит исключительно поддерживающий характер. Наиболее частой причиной смерти этих людей является кардиореспираторная недостаточность. [6]
Саркопения — это потеря мышечной массы и атрофия, связанная со старением. Это результат уменьшения размера мышц, а также уменьшения количества клеток-сателлитов, количества митохондрий и эластичности. Саркопения растет с возрастом, но не универсальна. Саркопения различается по степени физической активности, полу и расе.Это может быть связано с потерей мышечной силы и проблемами с неподвижностью, такими как падения, которые часто наблюдаются у стареющих людей. [1]
Клетки гладкой мускулатуры выстилают всю сосудистую систему человека. Они проявляют пластичность при повреждении сосудов. Именно эта пластичность влияет на патологический процесс атеросклероза. Зрелые гладкомышечные клетки участвуют в сокращении и тонусе сосудистой системы. Нагрузка холестерина явно увеличивает нагрузку на эндотелиальные клетки, что приводит к повреждению сосудов.Это повреждение переводит гладкую мускулатуру сосудов из неактивного сократительного состояния в состояние провоспалительного ответа. Результатом является пролиферация и ремоделирование гладкомышечных клеток; это приводит к образованию фиброзной капсулы, наблюдаемой при атеросклерозе. [7]
Гипертрофическая обструктивная кардиомиопатия (HOCM) — аутосомно-доминантное заболевание, вызываемое генетическими вариантами, кодирующими часть сократительного элемента кардиомиоцита. Эти мутации приводят к повышенной чувствительности миофиламентов к кальцию, утолщению межжелудочковой перегородки и, в конечном итоге, к затруднению кровотока.Хотя обычно симптомы обструкции протекают бессимптомно, они могут проявляться болью в груди при физической нагрузке, тахикардией с одышкой, обмороками и внезапной сердечной смертью. HOCM — это наиболее часто наследуемое сердечное заболевание с распространенностью 1 из 500. Это основная причина внезапной смерти у молодых людей, и в настоящее время от нее нет лечения. [8]
Непрерывное обучение / обзорные вопросы
Ссылки
- 1.
- Фронтера В.Р., Очала Дж. Скелетные мышцы: краткий обзор структуры и функций.Calcif Tissue Int. 2015 Март; 96 (3): 183-95. [PubMed: 25294644]
- 2.
- Roth GM, Bader DM, Pfaltzgraff ER. Выделение и физиологический анализ кардиомиоцитов мыши. J Vis Exp. 2014 7 сентября; (91): e51109. [Бесплатная статья PMC: PMC4828048] [PubMed: 25225886]
- 3.
- Буркхард С., ван Эйф В., Гаррик Л., Кристоффельс В. М., Баккерс Дж. Об эволюции кардиостимулятора. J Cardiovasc Dev Dis. 27 апреля 2017 г .; 4 (2) [Бесплатная статья PMC: PMC5715705] [PubMed: 29367536]
- 4.
- Уэбб RC. Плавное сокращение и расслабление мышц. Adv Physiol Educ. 2003 декабрь; 27 (1-4): 201-6. [PubMed: 14627618]
- 5.
- Севриева И., Ноулз А.С., Кампуракис Т., Сан Ю.Б. Регуляторный домен тропонина динамически перемещается во время активации сердечной мышцы. J Mol Cell Cardiol. 2014 Октябрь; 75: 181-7. [Бесплатная статья PMC: PMC4169182] [PubMed: 25101951]
- 6.
- Shieh PB. Мышечные дистрофии и другие генетические миопатии. Neurol Clin. 2013 ноя; 31 (4): 1009-29.[PubMed: 24176421]
- 7.
- Чистяков Д.А., Орехов А.Н., Бобрышев Ю.В. Гладкомышечные клетки сосудов при атеросклерозе. Acta Physiol (Oxf). 2015 Май; 214 (1): 33-50. [PubMed: 25677529]
- 8.
- Робинсон П., Лю X, Воробей А., Патель С., Чжан Ю. Х., Касадей Б., Уоткинс Н., Редвуд С. Мутации при гипертрофической кардиомиопатии увеличивают миофиламент Са 2+ буферизация, изменяют внутриклеточный Са 2+ и стимулируют Ca 2+ -зависимую передачу сигналов.J Biol Chem. 6 июля 2018 г .; 293 (27): 10487-10499. [Бесплатная статья PMC: PMC6036197] [PubMed: 29760186]
Физиология, мышцы — StatPearls — Книжная полка NCBI
Введение
В человеческом теле встречаются три основных типа мышц: скелетные, сердечные и гладкие. Каждый тип мышц имеет уникальные клеточные компоненты, физиологию, определенные функции и патологию. Скелетная мышца — это орган, который в первую очередь контролирует движение и осанку. Сердечная мышца охватывает сердце, которое поддерживает человеческое тело.Гладкая мускулатура присутствует во всей желудочно-кишечной, репродуктивной, мочевой, сосудистой и дыхательной системах.
Клеточная
Скелетные мышцы составляют примерно 40% от общей массы тела человека. В его состав входит множество отдельных волокон, которые объединены в мышечное веретено; это то, что придает скелетным мышцам полосатый вид. Одно мышечное волокно состоит в основном из актиновых и миозиновых волокон, которые покрыты клеточной мембраной (сарколемма). Эти волокна являются функциональной единицей органа, ведущей к сокращению и расслаблению.Есть две основные классификации скелетных мышц; к ним относятся Тип I (медленное окисление) и Тип II (быстро сокращающийся). Огромное разнообразие скелетных мышц приводит к различиям в скорости и продолжительности сокращений в разных группах мышц в зависимости от их конкретной функции [1].
Сердечная мышца или миокард — это непроизвольно поперечно-полосатая мышца, охватывающая камеры сердца. Он состоит из отдельных кардиомиоцитов, которые по структуре похожи на скелетные мышцы.Каждый кардиомиоцит содержит цитоскелетные и сократительные элементы, все из которых связаны между собой вставными дисками. Это хорошо прилипающие комплексы, которые позволяют клеткам сердечной мышцы получать быструю электрическую передачу и сокращаться как единое целое. [2] Сердечная мышца также содержит специализированные клетки кардиостимулятора, которые находятся в миокарде. Эти клетки позволяют сердечной ткани внутренне деполяризоваться без внешних стимулов. [3]
Клетки гладкой мускулатуры также состоят из актиновых и миозиновых волокон; однако они расположены в виде листов, а не веретен, что придает этому типу мышц гладкий вид.Эти клетки присутствуют в стенках многих органов, таких как легкие, желудочно-кишечный тракт, репродуктивные органы, кровеносные сосуды и даже кожа. [4]
Функция
Мышцы человеческого тела, будь то скелетные, сердечные или гладкие, создают силу и движение. Мышцы скелета поддерживают кости, чтобы поддерживать осанку, а также контролировать произвольные движения. Скелетные мышцы также способствуют метаболизму и хранению энергии. Сердечная мышца продвигает кровь и приводит к надлежащему насыщению кислородом и поддержанию каждой клетки, составляющей человеческое тело.Гладкая мышца расположена по всему телу и использует сократительную силу для укорачивания и продвижения различного содержимого через просвет множества систем органов, в которых она задействована.
Механизм
Потенциалы действия нервных волокон центральной нервной системы деполяризуют мышцу по длине сарколеммы к самым внутренним волокнам через систему поперечных канальцев (Т-канальцев). Потенциал действия отвечает дигидропиридиновым рецептором на Т-канальце; это действует как датчик напряжения, позволяющий высвобождать кальций.Впоследствии кальций активирует рианодиновые рецепторы в саркоплазматическом ретикулуме, чтобы высвободить еще больше кальция. Затем более высокие количества кальция могут связываться с протеином тропонином, который расположен на актиновых филаментах. Кальций-тропониновый комплекс вытесняет белок тропомиозин из активного центра актиновой нити и обеспечивает связывание миозина и сокращение мышц. Аденозинтрифосфат (АТФ) необходим для отделения миозина от актиновых волокон и обеспечения расслабления мышц. [1]
Подобно скелетным мышцам, сердечная мышца запускается связыванием кальция с тропонином в актиновых филаментах кардиомиоцитов.Это связывание затем удаляет тропомиозин и делает возможным связывание миозина с актиновыми филаментами и возможное сокращение. Существенная разница между сердечной и скелетной мышцами заключается в автоматизме кардиомиоцитов. Специализированные кардиостимуляторы, расположенные в синоатриальном (SA) узле, отвечают за сокращение сердечной мышцы. Они запускают потенциалы действия, которые обеспечивают приток натрия и калия, а также высвобождение кальция из саркоплазматической сети. Тогда сердечная мышца может сокращаться как единое скоординированное целое.[5]
Сокращение гладких мышц не находится под произвольным контролем и осуществляется за счет вегетативной регуляции взаимодействия кальция и кальмодулина. Сокращение начинается за счет изменения потенциала действия или активации механических рецепторов растяжения в плазматической мембране. Внутриклеточный кальций увеличивается и соединяется с белком кальмодулином. Именно этот комплекс активирует киназу легкой цепи миозина (MLC) для фосфорилирования и образования перекрестных мостиков между миозином и актином, что приводит к сокращению мышц.Некоторые гладкие мышцы поддерживают тонус, что обусловлено постоянным уровнем фосфорилирования при отсутствии внешних потенциалов. Снижение уровня внутриклеточного кальция вызывает расслабление. [4]
Клиническая значимость
Мышечная дистрофия — это прогрессирующая генетическая миопатия, которая приводит к нарушению нормальной анатомии и физиологии клеток скелетных мышц. Полное или частичное отсутствие белка дистрофина является патологическим механизмом мышечной дистрофии Беккера и Дюшенна.Дистрофин — это белок, связанный с филаментами скелетных мышц. Дистрофин обеспечивает структуру и поддержку сарколеммы моноволокна. Недостаток белка дистрофина приводит к повреждению поддерживающей сарколеммы, слабости и, в конечном итоге, атрофии здоровых мышечных волокон. Мышечная дистрофия Дюшенна поражает до 1 из 3600 мальчиков, что делает ее самой распространенной среди всех типов мышечных дистрофий. Многие с Дюшенном имеют низкую продолжительность жизни, потому что в настоящее время лечение недоступно.Лечение этих расстройств носит исключительно поддерживающий характер. Наиболее частой причиной смерти этих людей является кардиореспираторная недостаточность. [6]
Саркопения — это потеря мышечной массы и атрофия, связанная со старением. Это результат уменьшения размера мышц, а также уменьшения количества клеток-сателлитов, количества митохондрий и эластичности. Саркопения растет с возрастом, но не универсальна. Саркопения различается по степени физической активности, полу и расе.Это может быть связано с потерей мышечной силы и проблемами с неподвижностью, такими как падения, которые часто наблюдаются у стареющих людей. [1]
Клетки гладкой мускулатуры выстилают всю сосудистую систему человека. Они проявляют пластичность при повреждении сосудов. Именно эта пластичность влияет на патологический процесс атеросклероза. Зрелые гладкомышечные клетки участвуют в сокращении и тонусе сосудистой системы. Нагрузка холестерина явно увеличивает нагрузку на эндотелиальные клетки, что приводит к повреждению сосудов.Это повреждение переводит гладкую мускулатуру сосудов из неактивного сократительного состояния в состояние провоспалительного ответа. Результатом является пролиферация и ремоделирование гладкомышечных клеток; это приводит к образованию фиброзной капсулы, наблюдаемой при атеросклерозе. [7]
Гипертрофическая обструктивная кардиомиопатия (HOCM) — аутосомно-доминантное заболевание, вызываемое генетическими вариантами, кодирующими часть сократительного элемента кардиомиоцита. Эти мутации приводят к повышенной чувствительности миофиламентов к кальцию, утолщению межжелудочковой перегородки и, в конечном итоге, к затруднению кровотока.Хотя обычно симптомы обструкции протекают бессимптомно, они могут проявляться болью в груди при физической нагрузке, тахикардией с одышкой, обмороками и внезапной сердечной смертью. HOCM — это наиболее часто наследуемое сердечное заболевание с распространенностью 1 из 500. Это основная причина внезапной смерти у молодых людей, и в настоящее время от нее нет лечения. [8]
Непрерывное обучение / обзорные вопросы
Ссылки
- 1.
- Фронтера В.Р., Очала Дж. Скелетные мышцы: краткий обзор структуры и функций.Calcif Tissue Int. 2015 Март; 96 (3): 183-95. [PubMed: 25294644]
- 2.
- Roth GM, Bader DM, Pfaltzgraff ER. Выделение и физиологический анализ кардиомиоцитов мыши. J Vis Exp. 2014 7 сентября; (91): e51109. [Бесплатная статья PMC: PMC4828048] [PubMed: 25225886]
- 3.
- Буркхард С., ван Эйф В., Гаррик Л., Кристоффельс В. М., Баккерс Дж. Об эволюции кардиостимулятора. J Cardiovasc Dev Dis. 27 апреля 2017 г .; 4 (2) [Бесплатная статья PMC: PMC5715705] [PubMed: 29367536]
- 4.
- Уэбб RC. Плавное сокращение и расслабление мышц. Adv Physiol Educ. 2003 декабрь; 27 (1-4): 201-6. [PubMed: 14627618]
- 5.
- Севриева И., Ноулз А.С., Кампуракис Т., Сан Ю.Б. Регуляторный домен тропонина динамически перемещается во время активации сердечной мышцы. J Mol Cell Cardiol. 2014 Октябрь; 75: 181-7. [Бесплатная статья PMC: PMC4169182] [PubMed: 25101951]
- 6.
- Shieh PB. Мышечные дистрофии и другие генетические миопатии. Neurol Clin. 2013 ноя; 31 (4): 1009-29.[PubMed: 24176421]
- 7.
- Чистяков Д.А., Орехов А.Н., Бобрышев Ю.В. Гладкомышечные клетки сосудов при атеросклерозе. Acta Physiol (Oxf). 2015 Май; 214 (1): 33-50. [PubMed: 25677529]
- 8.
- Робинсон П., Лю X, Воробей А., Патель С., Чжан Ю. Х., Касадей Б., Уоткинс Н., Редвуд С. Мутации при гипертрофической кардиомиопатии увеличивают миофиламент Са 2+ буферизация, изменяют внутриклеточный Са 2+ и стимулируют Ca 2+ -зависимую передачу сигналов.J Biol Chem. 6 июля 2018 г .; 293 (27): 10487-10499. [Бесплатная статья PMC: PMC6036197] [PubMed: 29760186]
Физиология, мышцы — StatPearls — Книжная полка NCBI
Введение
В человеческом теле встречаются три основных типа мышц: скелетные, сердечные и гладкие. Каждый тип мышц имеет уникальные клеточные компоненты, физиологию, определенные функции и патологию. Скелетная мышца — это орган, который в первую очередь контролирует движение и осанку. Сердечная мышца охватывает сердце, которое поддерживает человеческое тело.Гладкая мускулатура присутствует во всей желудочно-кишечной, репродуктивной, мочевой, сосудистой и дыхательной системах.
Клеточная
Скелетные мышцы составляют примерно 40% от общей массы тела человека. В его состав входит множество отдельных волокон, которые объединены в мышечное веретено; это то, что придает скелетным мышцам полосатый вид. Одно мышечное волокно состоит в основном из актиновых и миозиновых волокон, которые покрыты клеточной мембраной (сарколемма). Эти волокна являются функциональной единицей органа, ведущей к сокращению и расслаблению.Есть две основные классификации скелетных мышц; к ним относятся Тип I (медленное окисление) и Тип II (быстро сокращающийся). Огромное разнообразие скелетных мышц приводит к различиям в скорости и продолжительности сокращений в разных группах мышц в зависимости от их конкретной функции [1].
Сердечная мышца или миокард — это непроизвольно поперечно-полосатая мышца, охватывающая камеры сердца. Он состоит из отдельных кардиомиоцитов, которые по структуре похожи на скелетные мышцы.Каждый кардиомиоцит содержит цитоскелетные и сократительные элементы, все из которых связаны между собой вставными дисками. Это хорошо прилипающие комплексы, которые позволяют клеткам сердечной мышцы получать быструю электрическую передачу и сокращаться как единое целое. [2] Сердечная мышца также содержит специализированные клетки кардиостимулятора, которые находятся в миокарде. Эти клетки позволяют сердечной ткани внутренне деполяризоваться без внешних стимулов. [3]
Клетки гладкой мускулатуры также состоят из актиновых и миозиновых волокон; однако они расположены в виде листов, а не веретен, что придает этому типу мышц гладкий вид.Эти клетки присутствуют в стенках многих органов, таких как легкие, желудочно-кишечный тракт, репродуктивные органы, кровеносные сосуды и даже кожа. [4]
Функция
Мышцы человеческого тела, будь то скелетные, сердечные или гладкие, создают силу и движение. Мышцы скелета поддерживают кости, чтобы поддерживать осанку, а также контролировать произвольные движения. Скелетные мышцы также способствуют метаболизму и хранению энергии. Сердечная мышца продвигает кровь и приводит к надлежащему насыщению кислородом и поддержанию каждой клетки, составляющей человеческое тело.Гладкая мышца расположена по всему телу и использует сократительную силу для укорачивания и продвижения различного содержимого через просвет множества систем органов, в которых она задействована.
Механизм
Потенциалы действия нервных волокон центральной нервной системы деполяризуют мышцу по длине сарколеммы к самым внутренним волокнам через систему поперечных канальцев (Т-канальцев). Потенциал действия отвечает дигидропиридиновым рецептором на Т-канальце; это действует как датчик напряжения, позволяющий высвобождать кальций.Впоследствии кальций активирует рианодиновые рецепторы в саркоплазматическом ретикулуме, чтобы высвободить еще больше кальция. Затем более высокие количества кальция могут связываться с протеином тропонином, который расположен на актиновых филаментах. Кальций-тропониновый комплекс вытесняет белок тропомиозин из активного центра актиновой нити и обеспечивает связывание миозина и сокращение мышц. Аденозинтрифосфат (АТФ) необходим для отделения миозина от актиновых волокон и обеспечения расслабления мышц. [1]
Подобно скелетным мышцам, сердечная мышца запускается связыванием кальция с тропонином в актиновых филаментах кардиомиоцитов.Это связывание затем удаляет тропомиозин и делает возможным связывание миозина с актиновыми филаментами и возможное сокращение. Существенная разница между сердечной и скелетной мышцами заключается в автоматизме кардиомиоцитов. Специализированные кардиостимуляторы, расположенные в синоатриальном (SA) узле, отвечают за сокращение сердечной мышцы. Они запускают потенциалы действия, которые обеспечивают приток натрия и калия, а также высвобождение кальция из саркоплазматической сети. Тогда сердечная мышца может сокращаться как единое скоординированное целое.[5]
Сокращение гладких мышц не находится под произвольным контролем и осуществляется за счет вегетативной регуляции взаимодействия кальция и кальмодулина. Сокращение начинается за счет изменения потенциала действия или активации механических рецепторов растяжения в плазматической мембране. Внутриклеточный кальций увеличивается и соединяется с белком кальмодулином. Именно этот комплекс активирует киназу легкой цепи миозина (MLC) для фосфорилирования и образования перекрестных мостиков между миозином и актином, что приводит к сокращению мышц.Некоторые гладкие мышцы поддерживают тонус, что обусловлено постоянным уровнем фосфорилирования при отсутствии внешних потенциалов. Снижение уровня внутриклеточного кальция вызывает расслабление. [4]
Клиническая значимость
Мышечная дистрофия — это прогрессирующая генетическая миопатия, которая приводит к нарушению нормальной анатомии и физиологии клеток скелетных мышц. Полное или частичное отсутствие белка дистрофина является патологическим механизмом мышечной дистрофии Беккера и Дюшенна.Дистрофин — это белок, связанный с филаментами скелетных мышц. Дистрофин обеспечивает структуру и поддержку сарколеммы моноволокна. Недостаток белка дистрофина приводит к повреждению поддерживающей сарколеммы, слабости и, в конечном итоге, атрофии здоровых мышечных волокон. Мышечная дистрофия Дюшенна поражает до 1 из 3600 мальчиков, что делает ее самой распространенной среди всех типов мышечных дистрофий. Многие с Дюшенном имеют низкую продолжительность жизни, потому что в настоящее время лечение недоступно.Лечение этих расстройств носит исключительно поддерживающий характер. Наиболее частой причиной смерти этих людей является кардиореспираторная недостаточность. [6]
Саркопения — это потеря мышечной массы и атрофия, связанная со старением. Это результат уменьшения размера мышц, а также уменьшения количества клеток-сателлитов, количества митохондрий и эластичности. Саркопения растет с возрастом, но не универсальна. Саркопения различается по степени физической активности, полу и расе.Это может быть связано с потерей мышечной силы и проблемами с неподвижностью, такими как падения, которые часто наблюдаются у стареющих людей. [1]
Клетки гладкой мускулатуры выстилают всю сосудистую систему человека. Они проявляют пластичность при повреждении сосудов. Именно эта пластичность влияет на патологический процесс атеросклероза. Зрелые гладкомышечные клетки участвуют в сокращении и тонусе сосудистой системы. Нагрузка холестерина явно увеличивает нагрузку на эндотелиальные клетки, что приводит к повреждению сосудов.Это повреждение переводит гладкую мускулатуру сосудов из неактивного сократительного состояния в состояние провоспалительного ответа. Результатом является пролиферация и ремоделирование гладкомышечных клеток; это приводит к образованию фиброзной капсулы, наблюдаемой при атеросклерозе. [7]
Гипертрофическая обструктивная кардиомиопатия (HOCM) — аутосомно-доминантное заболевание, вызываемое генетическими вариантами, кодирующими часть сократительного элемента кардиомиоцита. Эти мутации приводят к повышенной чувствительности миофиламентов к кальцию, утолщению межжелудочковой перегородки и, в конечном итоге, к затруднению кровотока.Хотя обычно симптомы обструкции протекают бессимптомно, они могут проявляться болью в груди при физической нагрузке, тахикардией с одышкой, обмороками и внезапной сердечной смертью. HOCM — это наиболее часто наследуемое сердечное заболевание с распространенностью 1 из 500. Это основная причина внезапной смерти у молодых людей, и в настоящее время от нее нет лечения. [8]
Непрерывное обучение / обзорные вопросы
Ссылки
- 1.
- Фронтера В.Р., Очала Дж. Скелетные мышцы: краткий обзор структуры и функций.Calcif Tissue Int. 2015 Март; 96 (3): 183-95. [PubMed: 25294644]
- 2.
- Roth GM, Bader DM, Pfaltzgraff ER. Выделение и физиологический анализ кардиомиоцитов мыши. J Vis Exp. 2014 7 сентября; (91): e51109. [Бесплатная статья PMC: PMC4828048] [PubMed: 25225886]
- 3.
- Буркхард С., ван Эйф В., Гаррик Л., Кристоффельс В. М., Баккерс Дж. Об эволюции кардиостимулятора. J Cardiovasc Dev Dis. 27 апреля 2017 г .; 4 (2) [Бесплатная статья PMC: PMC5715705] [PubMed: 29367536]
- 4.
- Уэбб RC. Плавное сокращение и расслабление мышц. Adv Physiol Educ. 2003 декабрь; 27 (1-4): 201-6. [PubMed: 14627618]
- 5.
- Севриева И., Ноулз А.С., Кампуракис Т., Сан Ю.Б. Регуляторный домен тропонина динамически перемещается во время активации сердечной мышцы. J Mol Cell Cardiol. 2014 Октябрь; 75: 181-7. [Бесплатная статья PMC: PMC4169182] [PubMed: 25101951]
- 6.
- Shieh PB. Мышечные дистрофии и другие генетические миопатии. Neurol Clin. 2013 ноя; 31 (4): 1009-29.[PubMed: 24176421]
- 7.
- Чистяков Д.А., Орехов А.Н., Бобрышев Ю.В. Гладкомышечные клетки сосудов при атеросклерозе. Acta Physiol (Oxf). 2015 Май; 214 (1): 33-50. [PubMed: 25677529]
- 8.
- Робинсон П., Лю X, Воробей А., Патель С., Чжан Ю. Х., Касадей Б., Уоткинс Н., Редвуд С. Мутации при гипертрофической кардиомиопатии увеличивают миофиламент Са 2+ буферизация, изменяют внутриклеточный Са 2+ и стимулируют Ca 2+ -зависимую передачу сигналов.J Biol Chem. 6 июля 2018 г .; 293 (27): 10487-10499. [Бесплатная статья PMC: PMC6036197] [PubMed: 29760186]
Типы мышечной ткани | Изучите мышечную анатомию
Примерно половину веса вашего тела составляют мышцы. В мышечной системе мышечная ткань подразделяется на три различных типа: скелетную, сердечную и гладкую. Каждый тип мышечной ткани в организме человека имеет уникальную структуру и определенную роль. Скелетная мышца перемещает кости и другие структуры. Сердечная мышца сокращает сердце, чтобы перекачивать кровь.Гладкая мышечная ткань, образующая такие органы, как желудок и мочевой пузырь, меняет форму, чтобы облегчить функции организма. Вот более подробная информация о структуре и функциях каждого типа мышечной ткани в мышечной системе человека.
1. Человеческое тело имеет более 600 скелетных мышц, которые перемещают кости и другие структуры
Скелетные мышцы прикрепляются к костям и перемещают их, сокращаясь и расслабляясь в ответ на произвольные сообщения нервной системы. Ткань скелетных мышц состоит из длинных клеток, называемых мышечными волокнами, которые имеют поперечно-полосатый вид.Мышечные волокна организованы в пучки, снабжаемые кровеносными сосудами и иннервируемые мотонейронами.
2. Стены многих человеческих органов сжимаются и автоматически расслабляются
Гладкая мускулатура находится в стенках полых органов по всему телу. Сокращения гладких мышц — это непроизвольные движения, вызванные импульсами, которые проходят через вегетативную нервную систему к гладкой мышечной ткани. Расположение клеток в гладкой мышечной ткани позволяет сокращаться и расслабляться с большой эластичностью.Гладкие мышцы стенок таких органов, как мочевой пузырь и матка, позволяют этим органам расширяться и расслабляться по мере необходимости. Гладкая мышца пищеварительного тракта (пищеварительного тракта) способствует перистальтическим волнам, которые перемещают проглоченную пищу и питательные вещества. В глазу гладкие мышцы изменяют форму линзы, чтобы сфокусировать объекты. Стенки артерий включают гладкие мышцы, которые расслабляются и сокращаются для перемещения крови по телу
3. Сокращения сердечной мышцы в ответ на сигналы от системы сердечной проводимости
Стенка сердца состоит из трех слоев.Средний слой, миокард, отвечает за работу сердца. Сердечная мышца, находящаяся только в миокарде, сокращается в ответ на сигналы сердечной проводящей системы, заставляющие сердце биться. Сердечная мышца состоит из клеток, называемых кардиоцитами. Кардиоциты, как и клетки скелетных мышц, имеют полосатый вид, но их общая структура короче и толще. Кардиоциты разветвлены, что позволяет им соединяться с несколькими другими кардиоцитами, образуя сеть, которая способствует скоординированному сокращению.
мышечной ткани | Безграничная анатомия и физиология
Характеристики мышечной ткани
Три типа мышечной ткани: скелетная, гладкая и сердечная.
Цели обучения
Опишите виды мышечной ткани
Основные выводы
Ключевые моменты
- Мышечная ткань может быть разделена функционально, находится ли она под произвольным или непроизвольным контролем; и морфологически, с бороздками или без бороздок.
- Применяя эти классификации, можно описать три типа мышц; скелетные, сердечные и гладкие.
- Скелетная мышца является произвольной и поперечно-полосатой, сердечная мышца — непроизвольной и растянутой, а гладкая мышца — непроизвольной и не поперечнополосатой.
Ключевые термины
- скелетная мышца : произвольная мышца позвоночных, поперечно-полосатая и прикрепленная сухожилиями к кости, используется для осуществления скелетных движений, таких как локомоция.
- непроизвольное : движение мышцы, не контролируемое сознанием, например биение сердца.
- полосатый : полосатый вид определенных типов мышц, в которых миофибриллы выровнены для создания постоянного направленного напряжения.
- произвольное : движение мышцы под сознательным контролем, например решает переместить предплечье.
- гладкая мышца : Непроизвольная мышца, которая находится в кишечнике, горле, матке и стенках кровеносных сосудов.
- сердечная мышца : поперечнополосатая мышца сердца позвоночного животного.
Мышечная ткань
Мышца — это мягкая ткань, которая в высокой степени специализируется на создании напряжения, которое приводит к созданию силы. Мышечные клетки или миоциты содержат миофибриллы, состоящие из миофиламентов актина и миозина, которые скользят друг мимо друга, создавая напряжение, изменяющее форму миоцита. Многочисленные миоциты составляют мышечную ткань, и контролируемое производство напряжения в этих клетках может генерировать значительную силу.
Типы мышечной ткани
Мышечную ткань можно разделить на функциональную, произвольную или непроизвольную, а также морфологически поперечно-полосатую или не полосатую. Произвольный относится к тому, находится ли мышца под сознательным контролем, полосатость относится к присутствию видимых полос внутри миоцитов, которые возникают из-за организации миофибрилл, обеспечивающих постоянное направление напряжения.
Применяя вышеупомянутые классификации, можно описать три формы мышечной ткани, которые выполняют широкий спектр описанных функций.
Скелетные мышцы
Скелетная мышца в основном прикрепляется к скелетной системе через сухожилия, чтобы поддерживать осанку и контролировать движение, например, сокращение двуглавой мышцы, прикрепленной к лопатке и лучевой кости, поднимет предплечье. Некоторые скелетные мышцы могут прикрепляться непосредственно к другим мышцам или коже, как это видно на лице, где многочисленные мышцы контролируют выражение лица.
Скелетные мышцы находятся под произвольным контролем, хотя это может быть подсознательным, например, при поддержании позы или равновесия.Морфологически скелетные миоциты имеют удлиненную, трубчатую форму и имеют поперечно-полосатую форму с множеством периферических ядер.
Ткань сердечной мышцы
Ткань сердечной мышцы находится только в сердце, где сердечные сокращения перекачивают кровь по всему телу и поддерживают кровяное давление.
Как и в случае со скелетными мышцами, сердечная мышца имеет поперечно-полосатую форму, но не контролируется сознательно и поэтому является непроизвольной. Сердечная мышца может быть дополнительно дифференцирована от скелетной мышцы по наличию вставных дисков, которые контролируют синхронизированное сокращение сердечных тканей.Сердечные миоциты короче скелетных эквивалентов и содержат только одно или два ядра, расположенных в центре.
Гладкая мышечная ткань
Гладкая мышечная ткань связана с множеством других органов и тканевых систем, таких как пищеварительная система или дыхательная система. Он играет важную роль в регуляции потока в таких тканях, например, помогая перемещению пищи через пищеварительную систему через перистальтику.
Гладкая мышца не имеет поперечно-полосатой части, хотя содержит те же миофиламенты, но они устроены по-разному и непроизвольно.Гладкомышечные миоциты имеют веретенообразную форму с одним центрально расположенным ядром.
Типы мышц : Сердечные и скелетные мышцы имеют поперечнополосатый вид, а гладкие — нет. И сердечные, и гладкие мышцы являются непроизвольными, а скелетные — произвольными.
мышечной системы человека | Функции, схемы и факты
В следующих разделах представлена основная структура для понимания общей анатомии мышц человека с описанием крупных групп мышц и их действий.Различные группы мышц работают согласованно, чтобы контролировать движения человеческого тела.
Шея
Движение шеи описывается в терминах вращения, сгибания, разгибания и бокового сгибания (т. Е. Движения, используемого для прикосновения уха к плечу). Направление действия может быть ипсилатеральным, что относится к движению в направлении сокращающейся мышцы, или контралатеральным, что относится к движению от стороны сокращающейся мышцы.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту.Подпишитесь сейчасВращение — одно из важнейших действий шейного отдела позвоночника. Вращение в основном осуществляется грудинно-ключично-сосцевидной мышцей, которая сгибает шею в ипсилатеральную сторону и вращает шею в противоположную сторону. Вместе грудинно-ключично-сосцевидные мышцы по обеим сторонам шеи сгибают шею и поднимают грудину, чтобы способствовать принудительному вдоху. Передняя и средняя лестничные мышцы, которые также расположены по бокам шеи, действуют ипсилатерально, поворачивая шею, а также поднимая первое ребро.Сплениус головы и сплениус шеи, расположенные в задней части шеи, вращают голову.
Боковое сгибание также является важным действием шейного отдела позвоночника. В сгибание шейной стороны вовлекаются грудинно-ключично-сосцевидные мышцы. Задние лестничные мышцы, расположенные на нижних сторонах шеи, ипсилатерально сгибают шею в сторону и приподнимают второе ребро. Сплениус головы и шейный позвонок также помогают при сгибании шеи в стороны. Мышцы, выпрямляющие позвоночник (подвздошно-костная, длинная и спинальная) — это большие глубокие мышцы, которые увеличивают длину спины.Все три действуют для ипсилатерального изгиба шеи.
Сгибание шеи относится к движению, используемому для прикосновения подбородка к груди. Это достигается в первую очередь грудинно-ключично-сосцевидными мышцами с помощью длинных коленных и длинных мышц головы, которые находятся в передней части шеи. Разгибание шеи противоположно сгибанию и выполняется многими из тех же мышц, которые используются для других движений шеи, включая шейную шейку шеи, звездочную мышцу головы, подвздошно-костную, длинную и спинную мышцы.
Спина
Послушайте, как врач объяснит причины и методы лечения боли в спине, называемой лордозом.
Узнайте о причинах и способах лечения боли в спине.
Contunico © ZDF Enterprises GmbH, Майнц Посмотреть все видео по этой статьеСпина является источником многих мышц, которые участвуют в движении шеи и плеч. Кроме того, осевой скелет, который проходит через спину вертикально, защищает спинной мозг, который иннервирует почти все мышцы тела.
Множественные мышцы спины функционируют именно при движениях спины. Например, мышцы, выпрямляющие позвоночник, разгибают спину (сгибают ее назад) и сгибают спину в стороны. Мышцы semispinalis dorsi и semispinalis capitis также расширяют спину. Маленькие мышцы позвонков (мультифиди и вращатели) помогают вращать, разгибать и сгибать спину в стороны. Квадратная мышца поясницы в нижней части спины сгибает поясничный отдел позвоночника и помогает вдыхать воздух благодаря своим стабилизирующим воздействиям в месте прикрепления к 12-му ребру (последнему из плавающих ребер).Лопатка (лопатка) поднимается трапециевидной мышцей, которая проходит от задней части шеи до середины спины, большими и малыми ромбовидными мышцами в верхней части спины и мышцей, поднимающей лопатку, которая проходит по бокам и сзади на шее.
Обзор мышечных тканей — анатомия и физиология
OpenStaxCollege
Цели обучения
К концу этого раздела вы сможете:
- Опишите различные типы мышц
- Объясните сжимаемость и расширяемость
Мышцы — это один из четырех основных типов тканей тела, и тело содержит три типа мышечной ткани: скелетные мышцы, сердечные мышцы и гладкие мышцы ([ссылка]).Все три мышечные ткани обладают некоторыми общими свойствами; все они проявляют свойство, называемое возбудимостью, поскольку их плазматические мембраны могут изменять свое электрическое состояние (с поляризованного на деполяризованное) и посылать электрическую волну, называемую потенциалом действия, по всей длине мембраны. В то время как нервная система может в некоторой степени влиять на возбудимость сердечной и гладкой мускулатуры, скелетные мышцы полностью зависят от сигналов нервной системы для правильной работы. С другой стороны, и сердечная мышца, и гладкие мышцы могут реагировать на другие раздражители, такие как гормоны и местные раздражители.
Три типа мышечной ткани
Тело состоит из трех типов мышечной ткани: (а) скелетная мышца, (б) гладкая мышца и (в) сердечная мышца. Сверху, LM × 1600, LM × 1600, LM × 1600. (Микрофотографии предоставлены Медицинской школой Риджентс Мичиганского университета © 2012)
Все мышцы начинают фактический процесс сокращения (укорачивания), когда белок актин притягивается белком миозином. Это происходит в поперечно-полосатых мышцах (скелетных и сердечных) после того, как специфические сайты связывания на актине подвергаются воздействию в ответ на взаимодействие между ионами кальция (Ca ++ ) и белками (тропонин и тропомиозин), которые «защищают» связывающий актин места.Ca ++ также необходим для сокращения гладкой мускулатуры, хотя его роль иная: здесь Ca ++ активирует ферменты, которые, в свою очередь, активируют миозиновые головки. Все мышцы нуждаются в аденозинтрифосфате (АТФ) для продолжения процесса сокращения, и все они расслабляются, когда Ca ++ удаляется и сайты связывания актина повторно экранируются.
Мышца может вернуться к своей исходной длине при расслаблении благодаря качеству мышечной ткани, называемому эластичностью.Он может вернуться к исходной длине за счет эластичных волокон. Мышечная ткань также обладает свойством растяжимости; он может растягиваться или расширяться. Сократимость позволяет мышечной ткани натягивать точки прикрепления и с силой укорачиваться.
Различия между тремя типами мышц включают микроскопическую организацию их сократительных белков — актина и миозина. Белки актина и миозина очень регулярно располагаются в цитоплазме отдельных мышечных клеток (называемых волокнами) как в скелетных, так и в сердечных мышцах, что создает узор или полосы, называемые полосами.Полосы видны в световой микроскоп при большом увеличении (см. [Ссылка]). Волокна скелетных мышц — это многоядерные структуры, из которых состоят скелетные мышцы. Каждое волокно сердечной мышцы имеет от одного до двух ядер и физически и электрически связаны друг с другом, так что все сердце сокращается как единое целое (так называемое синцитий).
Поскольку актин и миозин не расположены в гладких мышцах таким регулярным образом, цитоплазма гладкомышечного волокна (которое имеет только одно ядро) имеет однородный, гладкий вид (отсюда и название «гладкая мышца»).Однако менее организованный вид гладкой мускулатуры не следует рассматривать как менее эффективный. Гладкая мускулатура стенок артерий является важным компонентом, регулирующим кровяное давление, необходимое для проталкивания крови по кровеносной системе; гладкие мышцы кожи, внутренних органов и внутренних проходов необходимы для перемещения всех материалов по телу.
Мышца — это ткань у животных, которая обеспечивает активное движение тела или материалов внутри тела.Есть три типа мышечной ткани: скелетная мышца, сердечная мышца и гладкая мышца. Большая часть скелетных мышц тела движется, воздействуя на скелет. Сердечная мышца находится в стенке сердца и перекачивает кровь по кровеносной системе.
Гладкая мышца находится в коже, где она связана с волосяными фолликулами; он также находится в стенках внутренних органов, кровеносных сосудах и внутренних проходах, где помогает перемещать материалы.
Мышца, имеющая полосатый вид, описывается как ________.
- резинка
- гладкий
- возбудимый
- полосатая
Какой элемент важен для непосредственного запуска сокращения?
- натрий (Na + )
- кальций (Ca ++ )
- калий (K + )
- хлорид (класс — )
Какое из следующих свойств не является общим для всех трех мышечных тканей?
- возбудимость
- потребность в АТФ
- в состоянии покоя, использует защитные белки для покрытия актин-связывающих сайтов
- эластичность
Почему эластичность — важное качество мышечной ткани?
Позволяет мышцам вернуться к исходной длине во время расслабления после сокращения.
Глоссарий
- сердечная мышца
- поперечно-полосатых мышц в сердце; соединены друг с другом в вставных дисках и под контролем пейсмекерных клеток, которые сокращаются как одна единица, перекачивая кровь через систему кровообращения. Сердечная мышца находится под непроизвольным контролем.
- сократимость
- Возможность принудительного сокращения (сокращения)
- эластичность
- способность растягиваться и отскакивать
- возбудимость
- Способность к нервной стимуляции
- расширяемость
- возможность удлинения (удлинения)
- скелетная мышца
- поперечно-полосатая, многоядерная мышца, требующая передачи сигналов от нервной системы для запуска сокращения; большинство скелетных мышц относятся к произвольным мышцам, которые перемещают кости и производят движение
- гладкая мускулатура
- гладкая одноядерная мышца кожи, связанная с волосяными фолликулами; помогает перемещать материалы в стенках внутренних органов, кровеносных сосудах и внутренних проходах