
Для какого типа ткани характерны свойства возбудимости и проводимости: Для какого типа ткани характерны свойства возбудимости и проводимости?
Страница не найдена |
Страница не найдена |404. Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
78910111213
14151617181920
21222324252627
28
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Мар
Апр
Май
Июн
Июл
Авг
Сен
Окт
Ноя
Дек
Метки
Настройки
для слабовидящих
Амиодарон инструкция по применению: показания, противопоказания, побочное действие – описание Amiodarone таб.
 200 мг: 30 шт. (8932)
200 мг: 30 шт. (8932)Лекарственное взаимодействие амиодарона с другими препаратами возможно даже через несколько месяцев после окончания его применения за счет длительного T1/2.
При одновременном применении амиодарона и антиаритмических средств класса I А (в т.ч. дизопирамида) увеличивается интервал QT вследствие аддитивного действия на его величину и увеличивается риск развития желудочковой тахикардии типа «пируэт».
При одновременном применении амиодарона со слабительными средствами, которые могут вызывать гипокалиемию, повышается риск развития желудочковой аритмии.
Средства, вызывающие гипокалиемию, включая диуретики, кортикостероиды, амфотерицин В (в/в), тетракозактид при одновременном применении с амиодароном вызывают увеличение интервала QT и повышение риска развития желудочковой аритмии (в т.ч. типа «пируэт»).
При одновременном применении средств для общей анестезии, оксигенотерапии возникает риск развития брадикардии, артериальной гипотензии, нарушений проводимости, уменьшения ударного объема сердца, что, по-видимому, обусловлено аддитивным кардиодепрессивным и вазодилатирующим эффектами.
При одновременном применении трициклические антидепрессанты, фенотиазины, астемизол, терфенадин вызывают увеличение интервала QT и повышение риска развития желудочковой аритмии, особенно типа «пируэт».
При одновременном применении варфарина, фенпрокумона, аценокумарола усиливается антикоагулянтное действие и повышается риск развития кровотечений.
При одновременном применении винкамина, сультоприда, эритромицина (в/в), пентамидина (в/в, в/м) повышается риск развития желудочковой аритмии типа «пируэт».
При одновременном применении возможно повышение концентрации декстрометорфана в плазме крови вследствие уменьшения скорости его метаболизма в печени, что обусловлено ингибированием активности изофермента CYP2D6 системы цитохрома P450 под влиянием амиодарона и замедлением выведения декстрометорфана из организма.
При одновременном применении дигоксина значительно повышается концентрация дигоксина в плазме крови за счет снижения его клиренса и, вследствие этого, увеличивается риск развития дигиталисной интоксикации.
При одновременном применении дилтиазема, верапамила усиливается отрицательное инотропное действие, брадикардия, нарушение проводимости, AV-блокада.
Описан случай повышения концентрации амиодарона в плазме крови при его одновременном применении с индинавиром. Полагают, что ритонавир, нелфинавир, саквинавир будут оказывать подобное действие.
При одновременном применении колестирамина уменьшается концентрация амиодарона в плазме крови вследствие его связывания с колестирамином и уменьшения абсорбции из ЖКТ.
Имеются сообщения о повышении концентрации лидокаина в плазме крови при одновременном применении с амиодароном и развитии судорог, по-видимому, вследствие ингибирования метаболизма лидокаина под влиянием амиодарона.
Полагают, что возможен синергизм в отношении угнетающего действия на синусовый узел.
При одновременном применении лития карбоната возможно развитие гипотиреоидизма.
При одновременном применении прокаинамида увеличивается интервал QT вследствие аддитивного действия на его величину и риск развития желудочковой тахикардии типа «пируэт». Повышение концентрации в плазме крови прокаинамида и его метаболита N-ацетилпрокаинамида и усиление побочных эффектов.
Повышение концентрации в плазме крови прокаинамида и его метаболита N-ацетилпрокаинамида и усиление побочных эффектов.
При одновременном применении пропранолола, метопролола, соталола возможны артериальная гипотензия, брадикардия, фибрилляция желудочков, асистолия.
При одновременном применении тразодона описан случай развития аритмии типа «пируэт».
При одновременном применении хинидина увеличивается интервал QT вследствие аддитивного действия на его величину и риск развития желудочковой тахикардии типа «пируэт». Повышение концентрации хинидина в плазме крови и усиление его побочных эффектов.
При одновременном применении описан случай усиления побочных эффектов клоназепама, что, по-видимому, обусловлено его кумуляцией вследствие ингибирования окислительного метаболизма в печени под влиянием амиодарона.
При одновременном применении цизаприда значительно увеличивается интервал QT вследствие аддитивного действия, риск развития желудочковой аритмии (в т.ч. типа «пируэт»).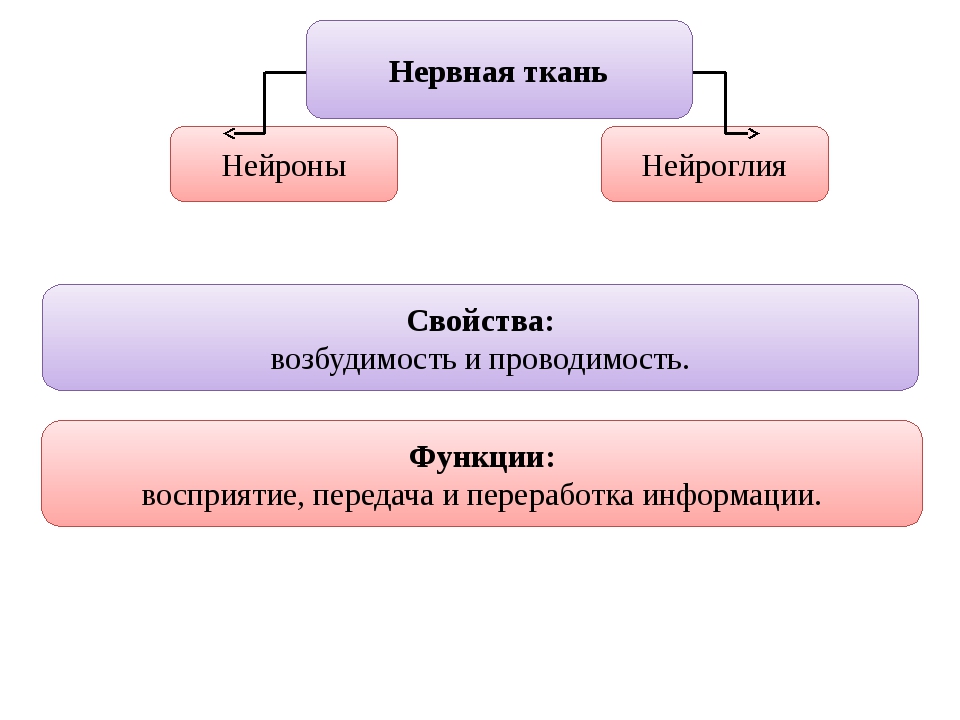
При одновременном применении повышается концентрация циклоспорина в плазме крови, риск развития нефротоксичности.
Описан случай легочной токсичности при одновременном применении циклофосфамида в высоких дозах и амиодарона.
Повышается концентрация амиодарона в плазме крови вследствие замедления его метаболизма под влиянием циметидина и других ингибиторов микросомальных ферментов печени.
Полагают, что вследствие ингибирования под влиянием амиодарона ферментов печени, при участии которых происходит метаболизм фенитоина, возможно повышение концентрации последнего в плазме крови и усиление его побочных эффектов.
Вследствие индукции микросомальных ферментов печени под влиянием фенитоина повышается скорость метаболизма амиодарона в печени и происходит уменьшение его концентрации в плазме крови.
Типы тканей и их свойства
Тема: Ткани. Типы тканей и их свойства
Цель: познакомить учащихся со строением и функциями тканей организма человека, развивать навыки самостоятельной работы с учебником, составления таблиц, распознавания микроструктур по описанию.
Ход урока
Организационный момент
Проверка домашнего задания.
Отчеты учащихся о заполнении таблицы «Функции органоидов и частей клетки».
Изучение нового материала
Познавательные вопросы по теме урока:
Что вам известно о тканях?
Какие ткани составляют организм человека?
Каковы их особенности строения, свойства и функции?
/Ответы обучающихся
Слайд 1 — Во многом вы правы. Давайте подробно остановимся на особенностях тканей человека. Запишите тему нашего урока.
Слайд 2 — В процессе изучения материала вам необходимо заполнить следующую таблицу:
Вспомним основные понятия:Ткань – группа клеток и межклеточного вещества, объединенные общим строением, происхождением и функцией.
Тканевая жидкость пополняется из вышедшей из кровеносного сосуда жидкой части крови, состав которой при этом изменяется, межклеточное вещество выделяется клетками
Слайд 3 – В организме человека выделяются четыре группы тканей.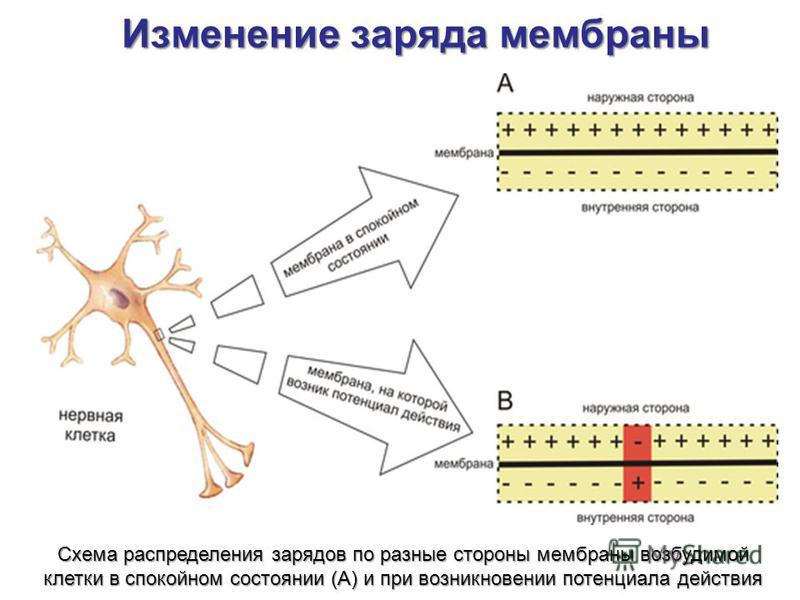
Эпителиальная. Клетки плотно прилегают друг к другу, межклеточного вещества мало
Соединительная. Клетки расположены рыхло, сильно развито межклеточное вещество
Мышечная. Образована мышечными волокнами, способна возбуждаться и сокращаться
Нервная. Состоит из клеток с отростками. Способна возбуждаться и передавать возбуждение
Слайды 4-9 – Эпителиальная (покровная) ткань.
Однослойная. Расположение: смежные оболочки внутренних органов. Функции: защитная, всасывающая
Многослойная. Расположение: покровы тела. Функции: защитная.
Железистая. Расположение: железы внешней и внутренней секреции. Функции: секреторная
Эпителиальная ткань – ткань, покрывающая тело и выстилающая его полости в виде пласта.
1) Образуется в онтогенезе раньше всех других тканей из зародышевых листков.
2) Способна к регенерации.
3) Лишена кровеносных сосудов.
4) Клетки плотно прилегают друг к другу.
5) Имеет мало межклеточного вещества.
6) Может состоять из нескольких слоев клеток.
7) Основные функции — защита (кожа), всасывание (кишечник), избирательный транспорт (почки, сосуды).
Слайды 10-16 – Соединительная ткань.
Костная. Расположение: скелет. Функции: Опорная, защитная, кроветворная
Хрящевая. Расположение: скелет, органы дыхания, ушная раковина. Функции: опорная, защитная
Волокнистая. Расположение: связки, сухожилия, дерма, прослойки между органами. Функции: опорно-защитная
Жировая. Расположение: подкожная клетчатка, между внутренними органами.
Функции: запасающая, защитная
К соединительной ткани относится и жидкая ткань – кровь.
Соединительная ткань — ткань, развивающаяся из мезодермы и выполняющая следующие функции: опорную (костная и хрящевая), трофическую (жировая и лимфа), защитную (лимфоидная и кровь).
1) Клетки не прилегают друг к другу.
2) Много межклеточного вещества.
3) Отличается большим разнообразием клеток.
Слайды 17-22 – Мышечная ткань.
Поперечно-полосатая скелетная . Расположение: опорно-двигательный аппарат тела и некоторых внутренних органов (язык, глотка, начальная часть пищевода). Функции: сократительная
Состоят из многоядерных мышечных волокон, покрытых возбудимой мембраной.
а) Волокна объединяются в мышечные пучки, из которых состоит мышца.
б) Основа скелетной мускулатуры.
в) Белые поперечнополосатые мышцы содержат много миофибрилл, сильно сокращаются, но быстро утомляются.
г) Красные — мало миофибрилл, имеют меньшую силу, но могут долго работать.
д) Клетки имеют поперечную исчерченность за счет миофибрилл.
МИОФИБРИЛЛЫ — мышечные нити,состоящие из саркомеров, способные к сокращению за счет актино-миозинового взаимодействия.
Сердечная мышца. Состоит из прямоугольных сократительных поперечнополосатых клеток.
а) Сокращения более медленные, чем у скелетных мышц.
б) Волокна переплетены в пучки.
в) Клетки не сливаются как в поперечнополосатой мышце.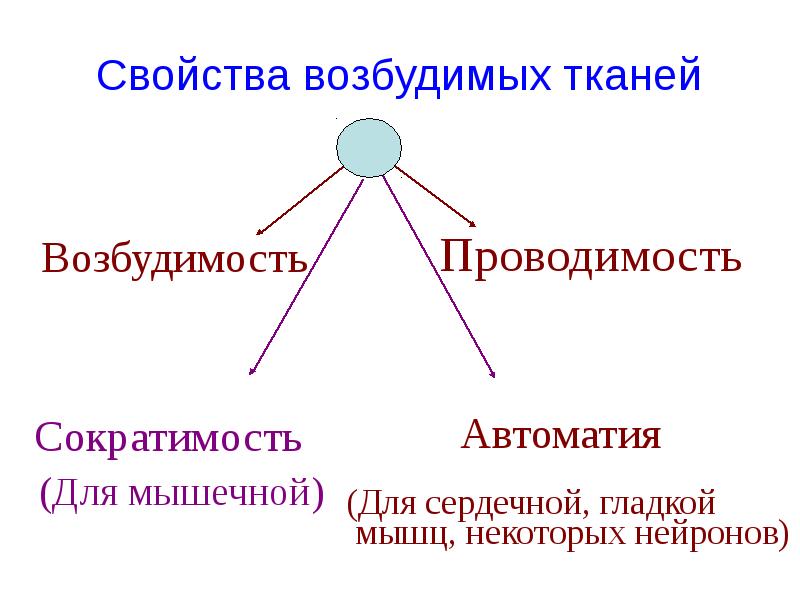
г) Способны к автоматическим сокращениям.
д) Имеет большой рефрактерный период (не может сокращаться).
е) Может управляться вегетативной нервной системой.
ж) Сокращается в объеме, уменьшая просвет полостей сердца.
Мышечная ткань — ткань, состоящая из клеток мезодермального происхождения, способных к возбуждению и сокращению.
Гладкая мышечная ткань — сократимая ткань, состоящая из отдельных клеток и не имеющая поперечной исчерченности.
Клетки сильно вытянуты. Способны к медленным длительным сокращениям.
Управляется вегетативной нервной системой. Входят в состав внутренних органов и сосудов.
Слайды 23-27 – Нервная ткань. Расположение: Головной и спинной мозг, нервные узлы и волокна Функции: Обеспечение согласованной деятельности разных систем органов, обеспечение связи организма с внешней средой, приспособление обмена веществ к изменяющимся условиям
Строение нервной ткани:
Нейроглия – вспомогательная роль (опора, питание)
Нейрон = тело + отростки (дендриты + аксон)
Дендрит – отросток, передающий возбуждение к телу нейрона.
Аксон – длинный единственный отросток, передающий информацию от тела нейрона к другому нейрону или рабочему органу.
Нервная ткань образована клетками (нейронами) эктодермального происхождения.
Нейроны не делятся.
Они способны к возбуждению и проведению нервного импульса.
Образуют стабильные контакты с другими клетками.
Образуют группы — ганглии, серое и белое вещество, нервные волокна.
Рассмотреть строение нейрона по схеме.
Закрепление изученного материала.
Провести фронтальное уточнение по заполнению таблицы
Покровы тела
Защита
Железистая
Железы внешней и внутренней секреции
Секреторная
Соединительная – клетки расположены рыхло, сильно развито межклеточное вещество.
Костная
Скелет
Опорная, защитная, кроветворная
Хрящевая
Скелет, органы дыхания, ушная раковина
Опорная, защитная
Волокнистая
Связки, сухожилия, дерма, прослойки между органами
Опорно-защитная
Жировая
Подкожная клетчатка, между внутренними органами
Запасающая, защитная
Кровь
Полости сердца и кровеносные сосуды
Дыхательная, транспортная, защитная
Мышечная – образована мышечными волокнами, способна возбуждаться и сокращаться.
Поперечно-полосатая скелетная
Опорно-двигательный аппарат тела и некоторых внутренних органов (язык, глотка, пищевод)
Сократительная
Поперечно-полосатая сердечная
Сердце
Сократительная
Гладкая
Мускулатура пищеварительного тракта, мочевого пузыря, кровеносных и лимфатических сосудов и др. внутренних органов
Сократительная
Нервная – состоит из клеток с отростками. Способна возбуждаться и передавать возбуждение.
Нейроны + нейроглия
Головной и спинной мозг, нервные узлы, волокна
Согласованная работа всех систем органов, связь с окружающей средой
Слайды 28-30 — Проверочная работа:
Запишите номера суждений, напротив верных поставьте «+», напротив ошибочных — «—»:
1. Эпителий желудка и кишечника относится к эпителиальным тканям.
2. для эпителиальной ткани характерно слабое развитие межклеточного вещества.
3. для эпителиальной ткани характерны свойства возбудимости и проводимости. 4. В эпителии отсутствуют кровеносные сосуды.
4. В эпителии отсутствуют кровеносные сосуды.
5. Эндотелий кровеносных сосудов относится к эпителиальной ткани.
6. Подкожная жировая клетчатка относится к эпителиальной ткани.
7. Для соединительных тканей характерно наличие хорошо развитого межклеточного вещества.
8. У соединительных тканей межклеточное вещество может быть твердым, жидким, эластичным.
9. К клеткам соединительной ткани относятся клетки крови, жировые клетки, клетки хряща.
10. Для мышечной ткани характерны свойства: возбудимость и сократимость.
11. Гладкая мышечная ткань входит в состав внутренних органов.
12. Поперечно-полосатая мышечная ткань образована мышечными клетками.
13. Сердечная мышца образована гладкой мышечной тканью.
14. Скелетные мышцы образованы мышечными волокнами, имеющими длину около 4 сантиметров, в каждом волокне сотни ядер находятся на периферии.
15. Возбуждение по аксону может идти только от тела нейрона.
16. По двигательному нейрону возбуждение от тела нейрона передается по аксону.
17. Нейрон всегда имеет только один аксон.
18. С одним нейроном могут контактировать более тысячи нервных клеток.
/По окончании работы проводится взаимопроверка/
Слайд 31 – Ответы:
Критерии оценивания:16-18 правильных ответов – «5»,
12-15 правильных ответов – «4»,
8-11 правильных ответов – «3»,
До 7 правильных ответов – «2».
Подведение итога урока. Выставление оценок
Слайд 32 — Домашнее задание: § 8 прочитать. Ответить устно на вопросы к параграфу. Закончить оформление таблицы схемами.
ФУНКЦИИ И СВОЙСТВА МЫШЕЧНОЙ ТКАНИ
ФУНКЦИИ И СВОЙСТВА МЫШЕЧНОЙ ТКАНИ
Функции мышечной ткани
- Движение: Скелет нашего тела придает телу достаточную жесткость, чтобы скелетные мышцы могли дергать и тянуть его, что приводит к движениям тела, таким как ходьба, жевание, бег, подъем, манипулирование предметами руками , и ковыряемся в носу.

- Поддержание осанки: Без особого сознательного контроля наши мышцы генерируют постоянную силу сокращения, которая позволяет нам сохранять вертикальное или сидячее положение, или позу.
- Дыхание: Наша мышечная система автоматически управляет движением воздуха в наше тело и из него.
- Выработка тепла: Сокращение мышечной ткани вырабатывает тепло, необходимое для поддержания температурного гомеостаза.Например, если температура нашего тела падает, мы дрожим, чтобы вырабатывать больше тепла.
- Связь: Мышечная ткань позволяет нам говорить, жестикулировать, писать и передавать свое эмоциональное состояние, например, улыбаясь или хмурясь.
- Сужение органов и кровеносных сосудов: Питательные вещества проходят через наш пищеварительный тракт, моча выводится из организма, а выделения выводятся из желез за счет сокращения гладкой мускулатуры. Сужение или расслабление кровеносных сосудов регулирует артериальное давление и распределение крови по всему телу.
- Перекачка крови: Кровь движется по кровеносным сосудам, потому что наше сердце неустанно принимает кровь и доставляет ее ко всем тканям и органам тела .
- Это не полный список. Среди множества возможных примеров можно привести тот факт, что мышцы помогают защитить хрупкие внутренние органы, окружая их, а также имеют решающее значение для поддержания целостности полостей тела.Например, у плодов с неполностью сформированной диафрагмой содержимое брюшной полости выпячивается (выпячивается) вверх в грудную полость, что препятствует нормальному росту и развитию легких. Несмотря на то, что это неполный список, понимание некоторых из этих основных мышечных функций поможет вам в дальнейшем.

 Сужение или расслабление кровеносных сосудов регулирует артериальное давление и распределение крови по всему телу.
Сужение или расслабление кровеносных сосудов регулирует артериальное давление и распределение крови по всему телу.Свойства мышечной ткани
Все мышечные клетки имеют несколько общих свойств: сократимость, возбудимость, растяжимость и эластичность:
- Сократимость — это способность мышечных клеток принудительно сокращаться. Например, чтобы согнуть (уменьшить угол сустава) локоть, вам нужно сократить (укоротить) двуглавую мышцу плеча и другие мышцы-сгибатели локтя в передней части руки. Обратите внимание, что для того, чтобы разогнуть локоть, задние мышцы-разгибатели руки должны сокращаться. Таким образом, мышц могут только тянуть, но не толкать.
- Возбудимость — это способность реагировать на раздражитель, который может исходить от двигательного нейрона или гормона.
- Растяжимость — способность мышцы растягиваться. Например, давайте пересмотрим наше сгибание локтей, которое мы обсуждали ранее. Чтобы иметь возможность согнуть локоть, мышцы-разгибатели локтя должны разгибаться, чтобы обеспечить сгибание. Отсутствие растяжимости известно как спастичность.
- Эластичность – это способность сокращаться или возвращаться к первоначальной длине мышцы после растяжения.
 Например, чтобы согнуть (уменьшить угол сустава) локоть, вам нужно сократить (укоротить) двуглавую мышцу плеча и другие мышцы-сгибатели локтя в передней части руки. Обратите внимание, что для того, чтобы разогнуть локоть, задние мышцы-разгибатели руки должны сокращаться. Таким образом, мышц могут только тянуть, но не толкать.
Например, чтобы согнуть (уменьшить угол сустава) локоть, вам нужно сократить (укоротить) двуглавую мышцу плеча и другие мышцы-сгибатели локтя в передней части руки. Обратите внимание, что для того, чтобы разогнуть локоть, задние мышцы-разгибатели руки должны сокращаться. Таким образом, мышц могут только тянуть, но не толкать. 
**Вы можете использовать кнопки ниже, чтобы перейти к следующему или предыдущему чтению в этом модуле**
Распечатать эту страницуГлава IНЕЙРОФИЗИОЛОГИЯ, ОБЗОР Нейрофизиология изучает нейронов , нервов и нервных систем , их функции и
как они это делают.Нейрон — это клетка, которая специализируется на двух фундаментальных свойствах.
живого вещества, а именно возбудимость и проводимость . Возбудимость – это способность
реагировать на изменения в окружающей среде; проводимость – это способность передавать импульс или действие
потенциала от одной части клетки к другой. В большинстве нейронов часть клетки,
несет потенциалы действия от тела клетки, называется аксоном или нервным волокном . При обсуждении любого столь сложного явления, как нервная система, почти
невозможно представить каждую тему в соответствующей последовательности, не упомянув некоторые идеи, события,
или конструкции вышли из строя. Краткий обзор тем, которые будут обсуждаться, может помочь читателю
поддерживать чувство порядка, даже когда темы упоминаются до того, как они подробно обсуждаются. Если расстояние для связи небольшое, локальные изменения напряжения могут быть
сообщается напрямую; однако, поскольку нервная оболочка является плохим проводником электричества,
сигналы местного напряжения не будут охватывать расстояния, превышающие один или два миллиметра.Таким образом,
специальное устройство связи, потенциал действия, используется для связи на большие расстояния,
у крупных животных на много метров. Клетки общаются друг с другом посредством специальных соединений, называемых синапсами .Синапсы
бывают двух видов: химические и электрические. В химических синапсах напряжение сигнала в одной клетке
запускает высвобождение химического вещества-трансмиттера в другую клетку. Этот передатчик
вещество может либо повышать активность принимающей клетки, т. е. повышать вероятность того, что оно
разрядит потенциалы действия или снизит активность принимающей клетки, т. е. уменьшит
Вероятность того, что он разрядит потенциалы действия. Мы называем первый эффект возбуждением ,
последний эффект как ингибирование .В дополнение к этим очевидным и прямым эффектам вещества-трансмиттеры
также может воздействовать на нейроны косвенно, воздействуя на нейроны на некотором расстоянии от точки выброса
вещества или изменения эффективности других веществ-передатчиков. Электрические синапсы не используют передающие вещества, но сообщают об изменениях в мембранное напряжение напрямую, и они обычно связаны только с возбуждением. У млекопитающих хим. синапсы, по-видимому, образуют большинство синаптических соединений, но это может быть просто потому, что они получили больше изучения. На принимающую клетку обычно оказывает синаптическое влияние не одна клетка, а многие. Это интегрирует возбуждающие и тормозные влияния ряда других клеток и на основании этого
интеграции, генерирует свои собственные сигналы, которые, в свою очередь, влияют на другие клетки. Понимание этого
интегративное свойство нервных клеток чрезвычайно важно для понимания того, как нервная
система ведет себя нормально и как она работает со сбоями. Некоторым клеткам требуется много синаптических влияний.
генерировать вывод; другие нет.Большинству нервных клеток требуется большое количество синаптических сигналов для генерации
выход; мышечных клеток нет. Некоторые нервные структуры, рецепторы , специализируются на приеме сигналов от внешних
окружающей среды или из внутренней среды, т. е. сигнализирует о состоянии самого организма.Рецепторы используют потенциальную энергию своих мембранных потенциалов, чтобы реагировать на эти воздействия окружающей среды.
сигналы и генерировать собственные закодированные сигналы, которые сообщают остальной нервной системе, что произошло.
ощущались в окружающей среде. Сигналы обычно представляют собой последовательности потенциалов действия, которые различаются по
шаблон или частота в зависимости от качества сигналов окружающей среды. Эти действия
потенциальные сигналы передаются по аксонам в центральную нервную систему (ЦНС). Часть кода
сигнал определяется тем, какой именно аксон или какой конкретный путь в центральном
нервную систему он пересекает. Чувствительность рецепторов различна. Это варьируется между людьми и между полами. Он меняется в зависимости от времени суток или времени месяца. Это зависит от сознательного состояние человека и то, что он делает. Следовательно, то, что мы чувствуем, есть не константа, а мы, тем не менее, поймите, что мы и наш мир относительно постоянны.Визуальный мир создается несколько миллионов крошечных точечных приемников света (палочек и колбочек), но мы воспринимаем зрительный мир как если бы оно было непрерывным. За эту трансформацию отвечает мозг. Движения генерируются в центральной нервной системе, как прямые, так и автоматические
реакции на сенсорные сигналы (рефлексы) и как реакции на окружающую среду, более независимые
отдельных сенсорных сигналов. Спинной мозг играет большую роль в обоих видах движения, как в
потому что он обеспечивает единственную эффективную связь между нервной системой и скелетными мышцами
и потому что он содержит нервный аппарат, который генерирует большую часть рефлекторной активности и других
более сложная деятельность, такая как ходьба. «Высшие функции» — обучение, эмоции, речь, мышление — предположительно производятся используя те же клеточные процессы, которые участвуют в простом рефлекторном поведении, но с участием этих процессы в различных структурах. Имея дело с нервной системой, естественно задаться вопросом, почему у животных, включая человека,
прибегли к использованию нервов вместо какого-либо другого механизма для направления своей деятельности. Давайте кратко сравним нервную и гормональную системы в отношении некоторых
существенные признаки нервной системы. Основная функция как нервной системы, так и
Гормональная система предназначена для передачи информации от одной части тела к другой. нерв
Клетки способны передавать сигнал от пальца ноги шестифутового человека к коре головного мозга примерно за
25 мс, то есть примерно за 25/1000 секунды. Если гормон выделяется в кровь в
пальцев ног, он не смог бы достичь коры головного мозга менее чем за 17 секунд. Скорость находится на
премия в поведении большинства животных, и очевидно, что преимущество в скорости принадлежит нервной
система. Те же функции, которые выполняет гормональная система в медленном организме или в организме, не
нуждаются в быстрых ответах, могут обслуживаться нервной системой в организме, которому действительно нужно реагировать
быстро. Например, чувствительность глаза у большинства организмов изменчива таким образом, что в
при ярком свете он менее чувствителен, а при тусклом свете более чувствителен. Другая важная функция нервной системы заключается в том, чтобы точно координировать сокращение
различные мышцы тела. Гормональные системы также играют роль в координации, но только
для действий, которые не требуют очень близкого времени или резкого начала и завершения.Нервная система
использует короткие потенциалы действия для передачи сигналов. Поскольку эти сигналы длятся лишь долю
секунды и двигаться на высоких скоростях: движения можно начинать и останавливать за долю секунды.
второй. Напротив, гормоны имеют период полураспада в секундах или минутах и путешествуют около тысячи лет.
раз медленнее, так как они должны доставляться к тканям по сосудистой системе. Еще одно различие между нервной и гормональной регуляцией заключается в количестве мишеней.
органы активируются одновременно. Если данное поведение требует действия нескольких органов для обеспечения
генерализованный ответ, гормональная система может хорошо подходить, но если только один или несколько
органы должны реагировать определенным образом, тогда нервный контроль может быть более подходящим. Еще одной важной функцией нервной системы является сбор и интеграция сенсорных сигналов.
информации, то есть объединять, анализировать и обрабатывать информацию об окружающей среде как
внутри и снаружи тела, чтобы выполнить соответствующий ответ.
Нервный контроль поведения наименее эффективен при инициировании и организации генерализованных или затянутые ответы. До этого момента в нашем обсуждении мы имели дело с нервной системой и нейронами, как если бы
их единственная функция состояла в том, чтобы производить потенциалы действия.Многие учебники, на самом деле многие
нейрофизиологи рассматривают нервную систему так, как если бы это было правдой. Сейчас есть много
свидетельство других функций. Например, удаление иннервации поперечно-полосатой мышцы приводит к
в конечном итоге в истощении или атрофии мышц. Другой функцией нейронов является секреция . Мы узнаем, что большинство нейронов общаются
друг с другом посредством химических веществ-переносчиков. Эти вещества синтезируются в
тело клетки нейрона, вероятно, сохраняется в аппарате Гольджи, а затем транспортируется ( аксоплазматический транспорт ) вниз
аксон.В этом прямоугольном направлении есть две скорости транспорта; один очень быстрый, из
порядка 410 мм/сутки (это одна из самых высоких скоростей клеточного транспорта для всех клеток в
тело; быстрый аксоплазматический транспорт ), а другой относительно медленный, порядка 1-12 мм/сутки ( медленный аксоплазматический транспорт ). Существует также вероятность того, что нейроны могут играть роль в процессах контроля заболеваний . За
например, пересечение тройничного нерва или его части для лечения хронической боли или для любого другого заболевания. Насколько важна эта неспайковая активность нейронов для функционирования нервная система неизвестна. Возможно, они являются основным путем межнейронных коммуникация.Возможно также, что они являются лишь проявлениями основного клеточного метаболизма. процессы. Ближайшее будущее может содержать ответ. Как возникла нервная система человека? Нервная система подвержена
эволюционное давление так же, как и любая другая часть организма. Филогенетическое развитие нервной системы можно охарактеризовать по аналогии с
деревянные блоки, с которыми вы играли в детстве. Сначала на дороге было разбросано всего несколько блоков.
пол, но вы быстро положите их, чтобы сформировать каркас. Затем вы полезли в коробку и
выбрал еще несколько блоков и добавил их в свою конструкцию по мере необходимости.То
основной фундамент остался, вы просто добавили больше блоков сверху. Таким же образом каркас
спинного мозга развились его основные нервные связи, обеспечивающие рефлексов . Подводя итог, функции нервной системы следующие:
В нашем исследовании нервной системы мы рассматриваем эти функции примерно в таком порядке. |
 Некоторые аксоны
очень короткие, несколько микрометров в длину; некоторые аксоны длинные, у человека до шести футов в
длина.Нерв представляет собой пучок этих аксонов, происходящих из различных клеток. Наконец, нервная система представляет собой совокупность всех нервных клеток в пределах одного организма. Случилось так, что в ходе
эволюции все животные, кроме простейших, прибегали к использованию какой-либо нервной системы.
организовывать и осуществлять свое поведение. Таким образом, изучение нервной системы, другими словами,
слов нейрофизиология , имеет основополагающее значение для изучения поведения.
Некоторые аксоны
очень короткие, несколько микрометров в длину; некоторые аксоны длинные, у человека до шести футов в
длина.Нерв представляет собой пучок этих аксонов, происходящих из различных клеток. Наконец, нервная система представляет собой совокупность всех нервных клеток в пределах одного организма. Случилось так, что в ходе
эволюции все животные, кроме простейших, прибегали к использованию какой-либо нервной системы.
организовывать и осуществлять свое поведение. Таким образом, изучение нервной системы, другими словами,
слов нейрофизиология , имеет основополагающее значение для изучения поведения.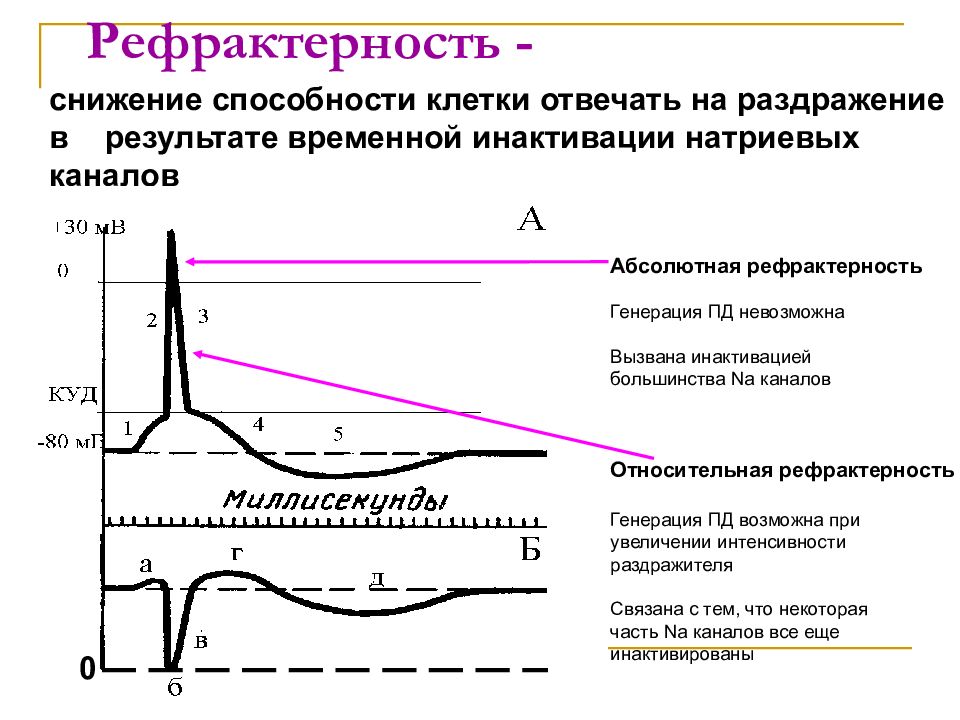 То
Нервная система состоит из миллиардов отдельных нервных клеток, каждая из которых имеет свою мембрану.
который разделяет электрические заряды; в биологических тканях эти заряды имеют форму ионов.То
Результатом является то, что каждая клетка имеет напряжение или потенциал покоящейся мембраны, который действует как источник
потенциальная энергия для работы клетки. Мембранные напряжения нервных клеток могут изменяться при
время, и эти изменения могут передаваться в виде сигналов другим нервным клеткам или исполнительным органам.
таких как железы и мышцы. На самом деле, это одно из основных направлений деятельности нервной системы.
То
Нервная система состоит из миллиардов отдельных нервных клеток, каждая из которых имеет свою мембрану.
который разделяет электрические заряды; в биологических тканях эти заряды имеют форму ионов.То
Результатом является то, что каждая клетка имеет напряжение или потенциал покоящейся мембраны, который действует как источник
потенциальная энергия для работы клетки. Мембранные напряжения нервных клеток могут изменяться при
время, и эти изменения могут передаваться в виде сигналов другим нервным клеткам или исполнительным органам.
таких как железы и мышцы. На самом деле, это одно из основных направлений деятельности нервной системы. Потенциал действия хорошо подходит для этой цели, поскольку
его короткая продолжительность (0,5 мс или около того) и быстрая проводимость (1-120 м/с) без декремента. То
короткая продолжительность потенциала действия позволяет генерировать его на высоких частотах, 500/сек или
более.
Потенциал действия хорошо подходит для этой цели, поскольку
его короткая продолжительность (0,5 мс или около того) и быстрая проводимость (1-120 м/с) без декремента. То
короткая продолжительность потенциала действия позволяет генерировать его на высоких частотах, 500/сек или
более.
 Одиночный потенциал действия, поступающий на синаптическое соединение
Аксон на мышце, нервно-мышечное соединение, всегда заставляет здоровую мышцу генерировать одиночный импульс.
собственный потенциал действия и сокращаться.
Одиночный потенциал действия, поступающий на синаптическое соединение
Аксон на мышце, нервно-мышечное соединение, всегда заставляет здоровую мышцу генерировать одиночный импульс.
собственный потенциал действия и сокращаться. Сигналы в определенных путях часто связаны с определенными видами
сенсорные события. Большая часть изучения нейрофизиологии посвящена рассмотрению того, как сенсорные системы
кодировать сенсорную информацию.
Сигналы в определенных путях часто связаны с определенными видами
сенсорные события. Большая часть изучения нейрофизиологии посвящена рассмотрению того, как сенсорные системы
кодировать сенсорную информацию.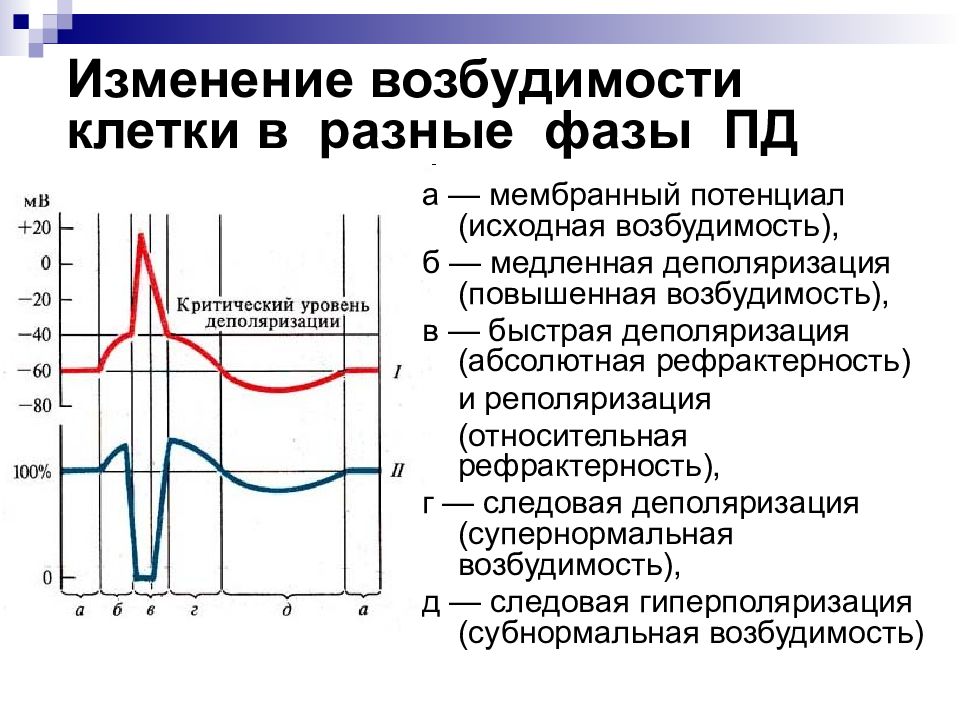 Однако спинной мозг обычно не работает.
независимо от остальной нервной системы. Значительные нарушения движения и поведения
протекают с нарушением деятельности очень многих участков нервной системы. Эти дефициты могут
варьироваться от небольших изменений в точной настройке движений, которые происходят после некоторого повреждения
пирамидного тракта, к отсутствию координации и синергии мышечных сокращений, как это бывает при
поражений мозжечка, вплоть до полного отсутствия какой-либо нормальной активности, как это бывает при мелких сосудистых
поражения ствола головного мозга.
Однако спинной мозг обычно не работает.
независимо от остальной нервной системы. Значительные нарушения движения и поведения
протекают с нарушением деятельности очень многих участков нервной системы. Эти дефициты могут
варьироваться от небольших изменений в точной настройке движений, которые происходят после некоторого повреждения
пирамидного тракта, к отсутствию координации и синергии мышечных сокращений, как это бывает при
поражений мозжечка, вплоть до полного отсутствия какой-либо нормальной активности, как это бывает при мелких сосудистых
поражения ствола головного мозга. это
нетрудно придумать множество альтернативных схем, некоторые из которых действительно встречаются в
природы, но вскоре становится ясно, что для крупных организмов единственный практический путь, кроме
нервной системы, чтобы общаться или активировать удаленную часть тела с помощью
химические вещества, называемые гормонами, переносимые кровью.
это
нетрудно придумать множество альтернативных схем, некоторые из которых действительно встречаются в
природы, но вскоре становится ясно, что для крупных организмов единственный практический путь, кроме
нервной системы, чтобы общаться или активировать удаленную часть тела с помощью
химические вещества, называемые гормонами, переносимые кровью. Вы можете себе представить, что случилось бы с кошкой, чей восстановительный рефлекс опосредован
гормоны? Гормон, выделяемый вестибулярным аппаратом, который определяет ориентацию,
дойти до мышц шеи, где начинается выпрямление, примерно за две секунды. в земле
сила тяжести, объект падает на 64 фута за две секунды. Это означает, что кошка не сможет правильно
себя при падении с высоты менее 64 футов над землей.Понятно, производительность нормальных кошек
превышает этот уровень. Конечно, там, где скорость не важна, например, при регулировании роста,
гормональная система работает отлично и, действительно, используется для этой цели.
Вы можете себе представить, что случилось бы с кошкой, чей восстановительный рефлекс опосредован
гормоны? Гормон, выделяемый вестибулярным аппаратом, который определяет ориентацию,
дойти до мышц шеи, где начинается выпрямление, примерно за две секунды. в земле
сила тяжести, объект падает на 64 фута за две секунды. Это означает, что кошка не сможет правильно
себя при падении с высоты менее 64 футов над землей.Понятно, производительность нормальных кошек
превышает этот уровень. Конечно, там, где скорость не важна, например, при регулировании роста,
гормональная система работает отлично и, действительно, используется для этой цели. В относительно медленном
ракообразных, которые не реагируют быстро на изменение интенсивности света, эта чувствительность находится под
гормональный контроль, в то время как у человека и относительно быстро движущихся насекомых необходимо реагировать
быстро меняется интенсивность света, тот же механизм контролируется нервной системой.
В относительно медленном
ракообразных, которые не реагируют быстро на изменение интенсивности света, эта чувствительность находится под
гормональный контроль, в то время как у человека и относительно быстро движущихся насекомых необходимо реагировать
быстро меняется интенсивность света, тот же механизм контролируется нервной системой. Поэтому они не могут быстро начинать действия или быстро их заканчивать. С другой стороны, где скорость
ответ не является существенным, гормоны могут адекватно координировать активность во многих тканях в
в разных местах, как, например, в случае событий, связанных с овуляцией.Где
требуется длительная активность, гормоны снова имеют преимущество. Они могут действовать долго,
потому что гормоны имеют более длительный период полураспада по сравнению с потенциалами действия. Чтобы сделать ту же работу,
потребуется длительная нервная деятельность, что означает непрерывную генерацию действия
потенциалы при, возможно, более высоких метаболических затратах.
Поэтому они не могут быстро начинать действия или быстро их заканчивать. С другой стороны, где скорость
ответ не является существенным, гормоны могут адекватно координировать активность во многих тканях в
в разных местах, как, например, в случае событий, связанных с овуляцией.Где
требуется длительная активность, гормоны снова имеют преимущество. Они могут действовать долго,
потому что гормоны имеют более длительный период полураспада по сравнению с потенциалами действия. Чтобы сделать ту же работу,
потребуется длительная нервная деятельность, что означает непрерывную генерацию действия
потенциалы при, возможно, более высоких метаболических затратах. Нужно ли было контролировать каждое движение нормального человека с помощью гормонов, гормона или
комбинация гормонов потребуется для каждого движения, а рецепторы будут
требуется для нескольких гормонов на один и тот же эффекторный орган. Иначе орган не знал бы, что
вид ответа на дай-маленький или большой, быстрый или медленный. Ясно, что количество гормонов будет
огромна, а система непрактична. С другой стороны. если бы было необходимо регулировать рост
с нервной системой потребовалась бы нейронная связь с каждой клеткой тела, а также
непрактичное состояние.Нервная система участвует в некоторых генерализованных реакциях, но в
вообще нервная система обеспечивает высокоспецифическое управление только одним или несколькими органами.
Нужно ли было контролировать каждое движение нормального человека с помощью гормонов, гормона или
комбинация гормонов потребуется для каждого движения, а рецепторы будут
требуется для нескольких гормонов на один и тот же эффекторный орган. Иначе орган не знал бы, что
вид ответа на дай-маленький или большой, быстрый или медленный. Ясно, что количество гормонов будет
огромна, а система непрактична. С другой стороны. если бы было необходимо регулировать рост
с нервной системой потребовалась бы нейронная связь с каждой клеткой тела, а также
непрактичное состояние.Нервная система участвует в некоторых генерализованных реакциях, но в
вообще нервная система обеспечивает высокоспецифическое управление только одним или несколькими органами. Потому что общее
знаменателем активности всех сенсорных систем являются изменения электрического потенциала,
добавить их вместе.Легко суммировать электрические потенциалы. Если бы интеграция была выполнена
с химической системой дело было бы намного сложнее. Можно было бы, например,
хотят отличить визуальные события от слуховых, поэтому каждое чувство должно использовать отдельный
гормон. Затем возникает проблема, как сложить их вместе, что-то вроде сложения яблок.
и апельсины; молекулы должны были бы соединиться химически, и тогда разные клетки были бы
необходимо реагировать на все возможные способы сочетания гормонов.Этот вид химического
интеграция происходит в гормональных системах, например, в контроле активности жировых клеток, но
количество вовлеченных гормонов и общий репертуар реакций органа ограничен
по сравнению с количеством различных способов, которыми нейрон может реагировать, и общим поведением
организм. На уровне целого организма трудная проблема химической интеграции заключается в том, как
для обработки чего-то вроде математических вычислений.
Потому что общее
знаменателем активности всех сенсорных систем являются изменения электрического потенциала,
добавить их вместе.Легко суммировать электрические потенциалы. Если бы интеграция была выполнена
с химической системой дело было бы намного сложнее. Можно было бы, например,
хотят отличить визуальные события от слуховых, поэтому каждое чувство должно использовать отдельный
гормон. Затем возникает проблема, как сложить их вместе, что-то вроде сложения яблок.
и апельсины; молекулы должны были бы соединиться химически, и тогда разные клетки были бы
необходимо реагировать на все возможные способы сочетания гормонов.Этот вид химического
интеграция происходит в гормональных системах, например, в контроле активности жировых клеток, но
количество вовлеченных гормонов и общий репертуар реакций органа ограничен
по сравнению с количеством различных способов, которыми нейрон может реагировать, и общим поведением
организм. На уровне целого организма трудная проблема химической интеграции заключается в том, как
для обработки чего-то вроде математических вычислений. Сейчас, без сомнения, очевидно, что нервный
контроль над поведением дает определенные преимущества перед всеми альтернативами.Эти:
Сейчас, без сомнения, очевидно, что нервный
контроль над поведением дает определенные преимущества перед всеми альтернативами.Эти: Это означает, что контакт нерва
требуется мышце для сохранения целостности. Не только это, но нерв определяет, по крайней мере
в некоторой степени сократительные свойства мышцы. Характерная скорость сокращения
мышца может быть систематически изменена путем ее денервации и реиннервации вместе с нервом.
из другой мышцы.Также оказывается, что именно нейрон индуцирует образование миелина.
вокруг аксона шванновскими клетками. Кроме того, аксоны, вероятно, снабжают питательными веществами или некоторыми
другие факторы, жизненно важные для поддержания миелина. Нервная система также тесно взаимодействует с иммунной системой.
Это означает, что контакт нерва
требуется мышце для сохранения целостности. Не только это, но нерв определяет, по крайней мере
в некоторой степени сократительные свойства мышцы. Характерная скорость сокращения
мышца может быть систематически изменена путем ее денервации и реиннервации вместе с нервом.
из другой мышцы.Также оказывается, что именно нейрон индуцирует образование миелина.
вокруг аксона шванновскими клетками. Кроме того, аксоны, вероятно, снабжают питательными веществами или некоторыми
другие факторы, жизненно важные для поддержания миелина. Нервная система также тесно взаимодействует с иммунной системой. В терминальной части
аксон, вещество-передатчик сохраняется для последующего высвобождения потенциалами действия на близлежащие
нейроны, на активность которых он влияет характерным образом.Кроме того, теперь известно, что
аксоны транспортируют вещества в ретроградном направлении, т. е. от терминалей обратно к соме ,
со скоростью 40-100 мм/сут ( ретроградный аксоплазматический транспорт ). Концевые части аксона собирают вещества из своих
окружающая обстановка; белки поглощаются нервными окончаниями и транспортируются обратно в сому, где
они могут быть использованы для получения других соединений или расщеплены и использованы в дальнейшем синтезе или
устраняются как отходы.Удивительно, но аксон может транспортировать два разных вещества.
в противоположных направлениях вдоль своего аксона одновременно.
В терминальной части
аксон, вещество-передатчик сохраняется для последующего высвобождения потенциалами действия на близлежащие
нейроны, на активность которых он влияет характерным образом.Кроме того, теперь известно, что
аксоны транспортируют вещества в ретроградном направлении, т. е. от терминалей обратно к соме ,
со скоростью 40-100 мм/сут ( ретроградный аксоплазматический транспорт ). Концевые части аксона собирают вещества из своих
окружающая обстановка; белки поглощаются нервными окончаниями и транспортируются обратно в сому, где
они могут быть использованы для получения других соединений или расщеплены и использованы в дальнейшем синтезе или
устраняются как отходы.Удивительно, но аксон может транспортировать два разных вещества.
в противоположных направлениях вдоль своего аксона одновременно. по другой причине часто следует вспышка вирусной инфекции опоясывающего герпеса в зоне
лицо, обслуживаемое этим нервом. Некоторые исследователи считают, что вирус можно контролировать путем поглощения
в нейроны, и когда нейроны повреждаются, они высвобождают захваченные вирусы в свои
терминалы в коже.(Мой друг, доктор Роберт Гримм, называет это теорией мусорной свалки).
(герпетические вирусные инфекции) в одном и том же месте или в соседних местах в течение многих лет. Там, из
Конечно, могут быть и другие объяснения, но существует возможность, что нервы играют некоторую роль в
контроль заболеваний.
по другой причине часто следует вспышка вирусной инфекции опоясывающего герпеса в зоне
лицо, обслуживаемое этим нервом. Некоторые исследователи считают, что вирус можно контролировать путем поглощения
в нейроны, и когда нейроны повреждаются, они высвобождают захваченные вирусы в свои
терминалы в коже.(Мой друг, доктор Роберт Гримм, называет это теорией мусорной свалки).
(герпетические вирусные инфекции) в одном и том же месте или в соседних местах в течение многих лет. Там, из
Конечно, могут быть и другие объяснения, но существует возможность, что нервы играют некоторую роль в
контроль заболеваний. Его структура может быть
изменены или дополнены, чтобы позволить видам выжить в измененной или новой среде, которая
требует нового образа жизни. Таким образом, можно ожидать, что мы сможем проследить эволюцию нервных
системы в летописи окаменелостей.Это оказалось трудно сделать, потому что нервная ткань мягкая.
не сохранились так, как кости. Однако мы имеем некоторое представление о развитии нервной
системы. Первые организмы, скорее всего, были одиночными клетками, такими как амебы. Потому что одноклеточный
животные обладают всеми характеристиками материи, они должны обладать проводимостью и возбудимостью. В
таким образом, эти первые одноклеточные организмы были также и первой нервной системой, выполнявшей многие из
вещи, которые делают более сложные системы.По мере того, как животные становились более сложными, стало более эффективно
дифференцировать клетки на функциональные типы. Появились разные ткани, а вместе с ними и нервные клетки. В
во-первых, нервные клетки могли быть плохо организованы, возможно, в виде рыхлых пучков без четкого порядка;
позже они более тесно объединились в организованные группы, называемые ганглиями.
Его структура может быть
изменены или дополнены, чтобы позволить видам выжить в измененной или новой среде, которая
требует нового образа жизни. Таким образом, можно ожидать, что мы сможем проследить эволюцию нервных
системы в летописи окаменелостей.Это оказалось трудно сделать, потому что нервная ткань мягкая.
не сохранились так, как кости. Однако мы имеем некоторое представление о развитии нервной
системы. Первые организмы, скорее всего, были одиночными клетками, такими как амебы. Потому что одноклеточный
животные обладают всеми характеристиками материи, они должны обладать проводимостью и возбудимостью. В
таким образом, эти первые одноклеточные организмы были также и первой нервной системой, выполнявшей многие из
вещи, которые делают более сложные системы.По мере того, как животные становились более сложными, стало более эффективно
дифференцировать клетки на функциональные типы. Появились разные ткани, а вместе с ними и нервные клетки. В
во-первых, нервные клетки могли быть плохо организованы, возможно, в виде рыхлых пучков без четкого порядка;
позже они более тесно объединились в организованные группы, называемые ганглиями. Еще позже ганглии
стали взаимосвязаны и слились, чтобы сформировать то, что мы знаем как спинной мозг и ствол головного мозга. В
в ходе филогенеза добавлялись какие-то новые группы клеток или ядер; позже в разных
животных эти ядра увеличились в размере и сложности, в то время как другие уменьшились или остались прежними.Кора головного мозга впервые появилась в эволюции у рептилий, а затем постепенно
изменился в размере и сложности, чтобы достичь своей нынешней конфигурации в человеческом мозгу.
Еще позже ганглии
стали взаимосвязаны и слились, чтобы сформировать то, что мы знаем как спинной мозг и ствол головного мозга. В
в ходе филогенеза добавлялись какие-то новые группы клеток или ядер; позже в разных
животных эти ядра увеличились в размере и сложности, в то время как другие уменьшились или остались прежними.Кора головного мозга впервые появилась в эволюции у рептилий, а затем постепенно
изменился в размере и сложности, чтобы достичь своей нынешней конфигурации в человеческом мозгу. В виде
окружающая среда изменилась, например, с влажной на сухую, когда ранее водные виды появились на
на суше на виды оказывалось давление, заставляя их менять свое поведение или вымирать.
У многих успешных видов это означало модификацию основного каркаса спинного мозга и
ствол мозга и, в некоторых случаях, добавление новых структур.Каждая новая структура была способна
изменение активности основного каркаса. Даже в человеческом мозгу так называемые высшие центры,
такие как кора головного мозга, работают, изменяя активность низших центров, но эти высшие
центры не способны контролировать поведение сами по себе. Именно эта модификация основного
рефлекторная деятельность, а не непосредственная регуляция мышечного сокращения, что приводит к плавному, непрерывному
активность свойственна высшим млекопитающим, в том числе и человеку.
В виде
окружающая среда изменилась, например, с влажной на сухую, когда ранее водные виды появились на
на суше на виды оказывалось давление, заставляя их менять свое поведение или вымирать.
У многих успешных видов это означало модификацию основного каркаса спинного мозга и
ствол мозга и, в некоторых случаях, добавление новых структур.Каждая новая структура была способна
изменение активности основного каркаса. Даже в человеческом мозгу так называемые высшие центры,
такие как кора головного мозга, работают, изменяя активность низших центров, но эти высшие
центры не способны контролировать поведение сами по себе. Именно эта модификация основного
рефлекторная деятельность, а не непосредственная регуляция мышечного сокращения, что приводит к плавному, непрерывному
активность свойственна высшим млекопитающим, в том числе и человеку. Под термином «коммуникация» мы подразумеваем просто передачу информации из одного места тела в другое.
другой, как правило, путем передачи нервных импульсов. Чтобы понять этот процесс, мы должны
знать, как генерируются и распространяются импульсы, т. е. как нервные клетки несут сигналы и передают
информацию другим нервным клеткам, мышцам и железам. Изучение ощущений включает
рассмотрение того, какие особенности окружающего мира можно ощущать, какие органы используются для
смысл; процесса трансдукции, т.е., преобразование энергии (будь то механическая, тепловая или световая)
в форму, которую может использовать нервная система, а именно в потенциал действия; и как эти
процессы в конце концов приводят к «ощущению». Эта последняя тема включает некоторое рассмотрение того, как
деятельность нервной системы связана с тем, что мы чувствуем. Интеграция – это сочетание и
сравнение информации от различных органов чувств и из памяти для использования в создании
решения о поведении. Движение и секреция, конечно, являются конечными продуктами всех
махинации нервной системы, и когда многочисленные движения или секреции собраны
вместе в сложные паттерны, которые можно модифицировать с течением времени, результат называется поведение .
Под термином «коммуникация» мы подразумеваем просто передачу информации из одного места тела в другое.
другой, как правило, путем передачи нервных импульсов. Чтобы понять этот процесс, мы должны
знать, как генерируются и распространяются импульсы, т. е. как нервные клетки несут сигналы и передают
информацию другим нервным клеткам, мышцам и железам. Изучение ощущений включает
рассмотрение того, какие особенности окружающего мира можно ощущать, какие органы используются для
смысл; процесса трансдукции, т.е., преобразование энергии (будь то механическая, тепловая или световая)
в форму, которую может использовать нервная система, а именно в потенциал действия; и как эти
процессы в конце концов приводят к «ощущению». Эта последняя тема включает некоторое рассмотрение того, как
деятельность нервной системы связана с тем, что мы чувствуем. Интеграция – это сочетание и
сравнение информации от различных органов чувств и из памяти для использования в создании
решения о поведении. Движение и секреция, конечно, являются конечными продуктами всех
махинации нервной системы, и когда многочисленные движения или секреции собраны
вместе в сложные паттерны, которые можно модифицировать с течением времени, результат называется поведение . Именно понимание поведения является конечной целью нейрофизиологии.
таким образом, это тема, о которой меньше всего можно сказать в всеобъемлющем смысле. Мы обсудим
стереотипное поведение, такое как рефлексы и некоторые другие
сложное поведение, такое как сон, речь и обучение, но то, что мы можем сказать о них, будет
поверхностны по сравнению со сложностью самого поведения. Мы относительно мало знаем о процессе обучения или координации, которая позволяет стрижу летать на расстоянии 140 миль.
в час через щель в стене, лишь немного превышающую его тело.Мы также не можем много сказать
о механизмах личности или сознания. Хотя наше понимание этих
явления находятся в зачаточном состоянии, все время обнаруживаются новые «факты»; и существующие знания
одновременно завораживает и завораживает.
Именно понимание поведения является конечной целью нейрофизиологии.
таким образом, это тема, о которой меньше всего можно сказать в всеобъемлющем смысле. Мы обсудим
стереотипное поведение, такое как рефлексы и некоторые другие
сложное поведение, такое как сон, речь и обучение, но то, что мы можем сказать о них, будет
поверхностны по сравнению со сложностью самого поведения. Мы относительно мало знаем о процессе обучения или координации, которая позволяет стрижу летать на расстоянии 140 миль.
в час через щель в стене, лишь немного превышающую его тело.Мы также не можем много сказать
о механизмах личности или сознания. Хотя наше понимание этих
явления находятся в зачаточном состоянии, все время обнаруживаются новые «факты»; и существующие знания
одновременно завораживает и завораживает.10.1: Обзор мышечных тканей
Цели обучения
- Описать различные типы мышц
- Объяснить сократимость и растяжимость
Мышцы — это один из четырех основных типов тканей тела, а тело содержит три типа мышечных тканей: скелетные мышцы, сердечные мышцы и гладкие мышцы (Рисунок \(\PageIndex{1}\)). Все три мышечные ткани имеют некоторые общие свойства; все они обладают качеством, называемым возбудимостью , поскольку их плазматические мембраны могут изменять свое электрическое состояние (с поляризованного на деполяризованное) и посылать электрическую волну, называемую потенциалом действия, по всей длине мембраны. В то время как нервная система может в некоторой степени влиять на возбудимость сердечной и гладкой мускулатуры, скелетные мышцы полностью зависят от сигналов нервной системы для правильной работы. С другой стороны, как сердечная мышца, так и гладкая мышца могут реагировать на другие раздражители, такие как гормоны и локальные раздражители.
Все три мышечные ткани имеют некоторые общие свойства; все они обладают качеством, называемым возбудимостью , поскольку их плазматические мембраны могут изменять свое электрическое состояние (с поляризованного на деполяризованное) и посылать электрическую волну, называемую потенциалом действия, по всей длине мембраны. В то время как нервная система может в некоторой степени влиять на возбудимость сердечной и гладкой мускулатуры, скелетные мышцы полностью зависят от сигналов нервной системы для правильной работы. С другой стороны, как сердечная мышца, так и гладкая мышца могут реагировать на другие раздражители, такие как гормоны и локальные раздражители.
Рисунок \(\PageIndex{1}\): Три типа мышечной ткани. Тело содержит три типа мышечной ткани: (а) скелетные мышцы, (б) гладкие мышцы и (в) сердечную мышцу. Сверху, LM × 1600, LM × 1600, LM × 1600. (Микрофотографии предоставлены Regents of Michigan Medical School © 2012) притягивается белком, называемым миозином.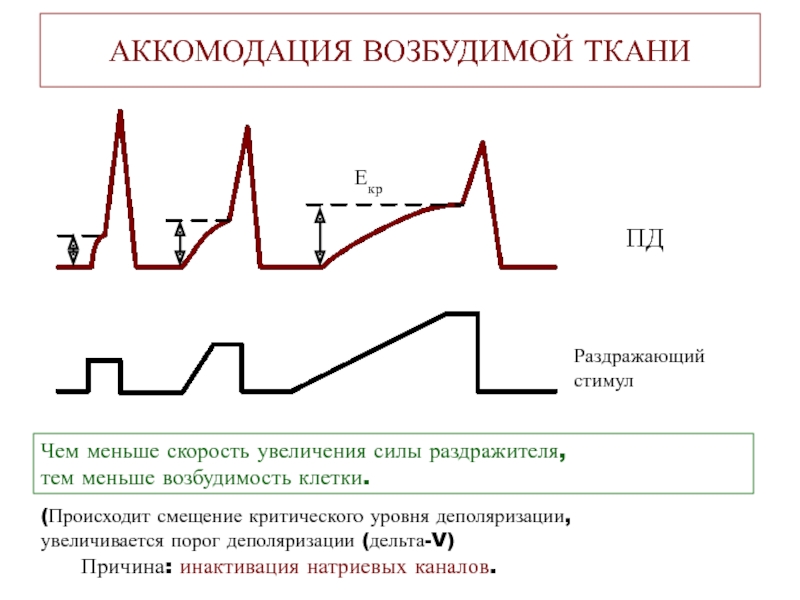 Это происходит в поперечнополосатых мышцах (скелетных и сердечных) после того, как специфические участки связывания на актине обнажаются в ответ на взаимодействие между ионами кальция (Ca ++ ) и белками (тропонином и тропомиозином), которые «защищают» актин-связывающие места. Ca ++ также необходим для сокращения гладких мышц, хотя его роль иная: здесь Ca ++ активирует ферменты, которые, в свою очередь, активируют головки миозина. Всем мышцам требуется аденозинтрифосфат (АТФ) для продолжения процесса сокращения, и все они расслабляются, когда Са ++ удаляется, а участки связывания актина повторно экранируются.
Это происходит в поперечнополосатых мышцах (скелетных и сердечных) после того, как специфические участки связывания на актине обнажаются в ответ на взаимодействие между ионами кальция (Ca ++ ) и белками (тропонином и тропомиозином), которые «защищают» актин-связывающие места. Ca ++ также необходим для сокращения гладких мышц, хотя его роль иная: здесь Ca ++ активирует ферменты, которые, в свою очередь, активируют головки миозина. Всем мышцам требуется аденозинтрифосфат (АТФ) для продолжения процесса сокращения, и все они расслабляются, когда Са ++ удаляется, а участки связывания актина повторно экранируются.
Мышца может вернуться к своей первоначальной длине при расслаблении благодаря свойству мышечной ткани, называемому эластичностью . Он может вернуться к своей первоначальной длине благодаря эластичным волокнам. Мышечная ткань также имеет качество растяжимость ; он может растягиваться или растягиваться. Сократимость позволяет мышечной ткани натягиваться в точках прикрепления и укорачиваться с силой.
Сократимость позволяет мышечной ткани натягиваться в точках прикрепления и укорачиваться с силой.
Различия между тремя типами мышц включают микроскопическую организацию их сократительных белков — актина и миозина.Белки актина и миозина расположены очень регулярно в цитоплазме отдельных мышечных клеток (называемых волокнами) как в скелетных мышцах, так и в сердечной мышце, что создает узор или полосы, называемые исчерченностью. Исчерченность видна в световой микроскоп при большом увеличении (см. рисунок \(\PageIndex{1}\).c). Скелетная мышца Волокна представляют собой многоядерные структуры, составляющие скелетную мышцу. Сердечная мышца волокон, каждое из которых имеет от одного до двух ядер и физически и электрически связаны друг с другом, так что все сердце сокращается как единое целое (называемое синцитием).
Поскольку актин и миозин не располагаются в гладкой мышце таким регулярным образом, цитоплазма волокна гладкой мышцы (которое имеет только одно ядро) имеет однородный, неисчерченный вид (что приводит к названию гладкой мышцы). Однако менее организованный внешний вид гладких мышц не следует интерпретировать как менее эффективный. Гладкие мышцы в стенках артерий являются важным компонентом, который регулирует кровяное давление, необходимое для проталкивания крови по системе кровообращения; а гладкие мышцы кожи, внутренних органов и внутренних путей необходимы для перемещения всех материалов по телу.
Однако менее организованный внешний вид гладких мышц не следует интерпретировать как менее эффективный. Гладкие мышцы в стенках артерий являются важным компонентом, который регулирует кровяное давление, необходимое для проталкивания крови по системе кровообращения; а гладкие мышцы кожи, внутренних органов и внутренних путей необходимы для перемещения всех материалов по телу.
Обзор главы
Мышцы — это ткани животных, которые обеспечивают активное движение тела или материалов внутри тела. Существует три типа мышечной ткани: скелетные мышцы, сердечная мышца и гладкие мышцы. Большая часть скелетных мышц тела производит движение, воздействуя на скелет. Сердечная мышца находится в стенке сердца и перекачивает кровь по кровеносной системе.
Гладкие мышцы находятся в коже, где они связаны с волосяными фолликулами; он также находится в стенках внутренних органов, кровеносных сосудов и внутренних проходов, где он помогает перемещать материалы.
Контрольные вопросы
Q. Мышца, имеющая полосатый вид, описывается как ________.
Мышца, имеющая полосатый вид, описывается как ________.
А. эластичный
Б. неполосатый
С. возбудимый
Г. поперечнополосатый
В. Какой элемент важен для непосредственного запуска сокращения?
A. натрий (Na + )
B. кальций (Ca ++ )
C. калий (K + )
D. хлорид (Cl — )
В.Какое из следующих свойств не является общим для всех трех мышечных тканей?
A. возбудимость
B. потребность в АТФ
C. в состоянии покоя использует экранирующие белки для прикрытия мест связывания актина
D. эластичность
Вопросы критического мышления
В. Почему эластичность является важным качеством мышечной ткани?
A. Позволяет мышце вернуться к своей первоначальной длине во время расслабления после сокращения.
Глоссарий
- сердечная мышца
- поперечнополосатая мышца сердца; соединены друг с другом во вставочных дисках и регулируются пейсмекерными клетками, которые сокращаются как единое целое, чтобы перекачивать кровь по системе кровообращения. Сердечная мышца находится под непроизвольным контролем.
 Сердечная мышца находится под непроизвольным контролем.
Сердечная мышца находится под непроизвольным контролем.- сократимость
- способность насильственно укорачиваться (сокращаться)
- эластичность
- способность растягиваться и восстанавливаться
- возбудимость
- способность подвергаться нервной стимуляции
- растяжимость
- способность удлиняться (удлиняться)
- скелетная мышца
- поперечно-полосатая многоядерная мышца, для запуска сокращения которой требуется сигнал от нервной системы; большинство скелетных мышц называют произвольными мышцами, которые двигают кости и производят движение
- гладкая мышца
- одноядерная гладкая мышца в коже, связанная с волосяными фолликулами; способствует перемещению материалов в стенках внутренних органов, кровеносных сосудов и внутренних проходов
Авторы и авторство
Идентификация системы эндогенных глутаматергических передатчиков, контролирующих возбудимость и проводимость предсердных кардиомиоцитов
Животные
С. Национальные институты здоровья. Все эксперименты на животных были одобрены Комитетом по уходу и использованию животных Медицинской школы Университета Тунцзи.
Национальные институты здоровья. Все эксперименты на животных были одобрены Комитетом по уходу и использованию животных Медицинской школы Университета Тунцзи.
Этика и получение образцов человеческих предсердий
Получение образцов человеческих предсердий было одобрено комитетом по этике больницы Чжуншань Университета Фудань и соответствовало Хельсинкской декларации. Образцы человеческих предсердий были получены во время операции в больнице Чжуншань Университета Фудань. Образцы предсердного миокарда были взяты у взрослых пациентов с нормальным синусовым ритмом, перенесших операцию на клапанах сердца.Все участники дали письменное информированное согласие.
Электронная микроскопия
Свежий миокард предсердий разрезали на блоки ткани толщиной 1 мм и немедленно погружали в охлажденный льдом 2,5% глутаровый альдегид (забуференный 0,1 М фосфатным буфером, рН 7,4). Образцы фиксировали не менее 1 ч при 4 °С. Фиксированные ткани предсердий неоднократно промывали фосфатным буфером и постфиксировали в 1% четырехокиси осмия в течение 1 часа. Зафиксированные образцы обезвоживали в этаноле возрастающей концентрации (10%, 50%, 70% и 90%), затем заливали 100% эпоксидной смолой и оставляли полимеризоваться при 55°С в 5% СО 2 на 36 ч. .Затем полимерные блоки разрезали на ультрамикротоме. Ультратонкие срезы помещали на сетки, окрашивали уранилацетатом и раствором цитрата свинца для наблюдения с помощью ПЭМ (JOEL TEM1230, Япония).
Зафиксированные образцы обезвоживали в этаноле возрастающей концентрации (10%, 50%, 70% и 90%), затем заливали 100% эпоксидной смолой и оставляли полимеризоваться при 55°С в 5% СО 2 на 36 ч. .Затем полимерные блоки разрезали на ультрамикротоме. Ультратонкие срезы помещали на сетки, окрашивали уранилацетатом и раствором цитрата свинца для наблюдения с помощью ПЭМ (JOEL TEM1230, Япония).
Выделение предсердных кардиомиоцитов крысы и человека
Отдельные предсердные кардиомиоциты получали из правого предсердия взрослых самцов крыс Sprague-Dawley (6–8 недель) путем ферментативного расщепления, как описано ранее, с небольшими изменениями. 31 Вкратце, крысу анестезировали пентобарбиталом (25 мг/кг) внутрибрюшинно, грудную клетку вскрыли для доступа к сердцу.Нисходящую аорту перерезали, и сердце немедленно промывали введением 15 мл холодного раствора ЭДТА-Тироде: 140 мМ NaCl, 5,4 мМ KCl, 1,2 мМ KH 2 PO 4 , 1,8 мМ CaCl 2 , 1,0 мМ MgCl 2 , 5,5 мМ глюкозы, 5 мМ HEPES и 5 мМ ЭДТА (pH доводили до 7,4 с помощью NaOH) в правый желудочек.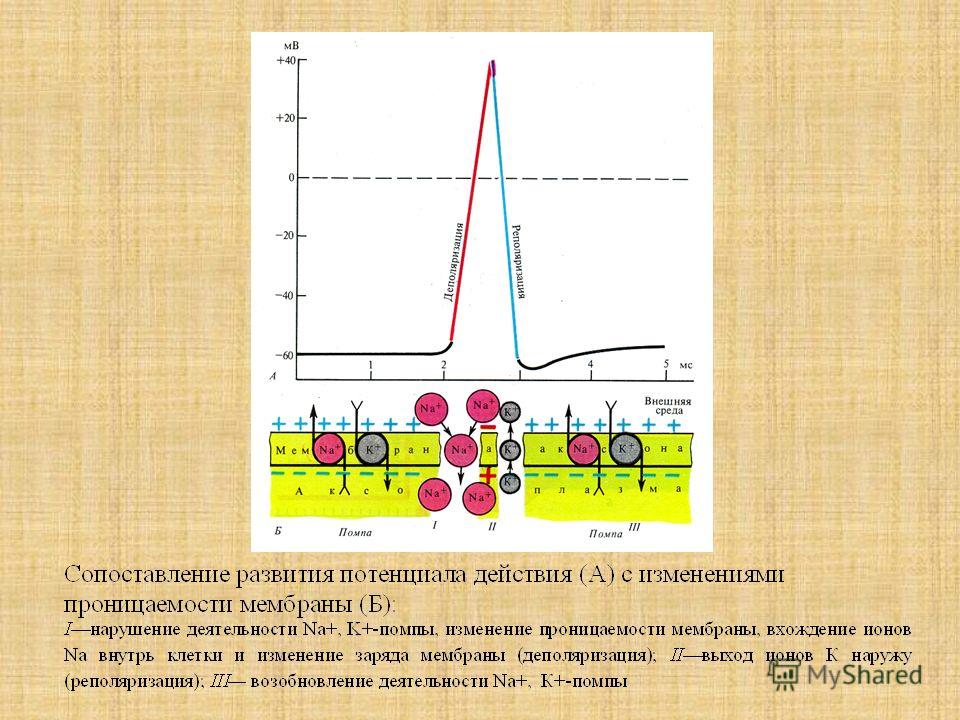 Сердце вырезали и переносили в чашку диаметром 60 мм, содержащую свежий буфер ЭДТА, для удаления жира и соединительной ткани. Ткани предсердий разрезали на мелкие кусочки и дважды промывали раствором 2+ с низким содержанием Са, содержащим: 140 мМ NaCl, 5.4 мМ KCl, 1,2 мМ KH 2 PO 4 , 0,2 мМ CaCl 2 , 18,5 мМ глюкозы, 50 мМ таурина и 1,0 % BSA (pH доводили до 6,9 с помощью NaOH) и затем расщепляли в течение 10–15 мин в растворе low-Ca 2+ , содержащем коллагеназу типа II (0,8 мг/мл; Worthington, США) при осторожном перемешивании. Супернатант переносили в модифицированный раствор Kraftbruhe (KB), содержащий: 100 мМ глутамата калия, 10 мМ аспартата калия, 25 мМ KCl, 10 мМ KH 2 PO 4 , 2 мМ MgSO 4 , 20 мМ таурина, 5 мМ креатин, 0.5 мМ EGTA, 20 мМ глюкозы, 5 мМ HEPES и 1,0% BSA (pH доводили до 7,2 с помощью KOH) для прекращения переваривания. Суспензии клеток фильтровали через 100-мкм фильтр и центрифугировали при 100× g в течение 1 мин для осаждения предсердных кардиомиоцитов.
Сердце вырезали и переносили в чашку диаметром 60 мм, содержащую свежий буфер ЭДТА, для удаления жира и соединительной ткани. Ткани предсердий разрезали на мелкие кусочки и дважды промывали раствором 2+ с низким содержанием Са, содержащим: 140 мМ NaCl, 5.4 мМ KCl, 1,2 мМ KH 2 PO 4 , 0,2 мМ CaCl 2 , 18,5 мМ глюкозы, 50 мМ таурина и 1,0 % BSA (pH доводили до 6,9 с помощью NaOH) и затем расщепляли в течение 10–15 мин в растворе low-Ca 2+ , содержащем коллагеназу типа II (0,8 мг/мл; Worthington, США) при осторожном перемешивании. Супернатант переносили в модифицированный раствор Kraftbruhe (KB), содержащий: 100 мМ глутамата калия, 10 мМ аспартата калия, 25 мМ KCl, 10 мМ KH 2 PO 4 , 2 мМ MgSO 4 , 20 мМ таурина, 5 мМ креатин, 0.5 мМ EGTA, 20 мМ глюкозы, 5 мМ HEPES и 1,0% BSA (pH доводили до 7,2 с помощью KOH) для прекращения переваривания. Суспензии клеток фильтровали через 100-мкм фильтр и центрифугировали при 100× g в течение 1 мин для осаждения предсердных кардиомиоцитов.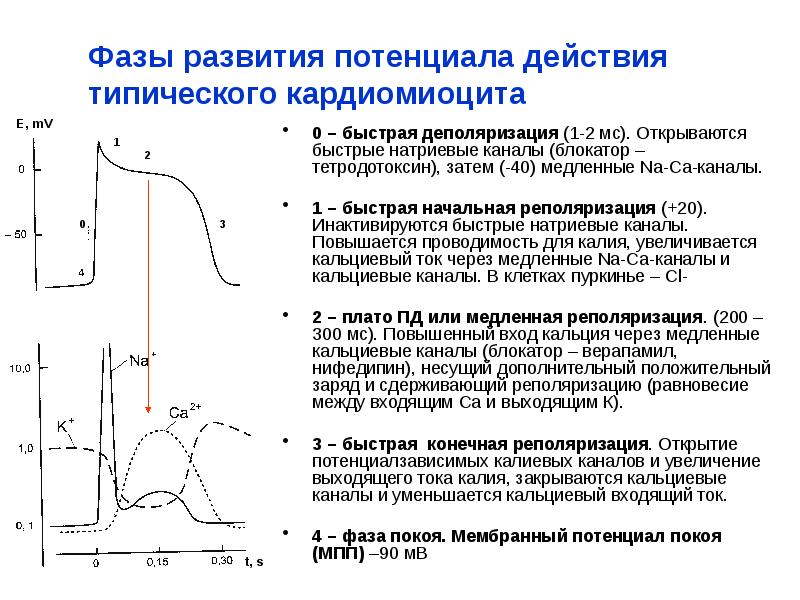 Супернатант удаляли, а осажденные предсердные кардиомиоциты ресуспендировали в 4 мл (в зависимости от выхода) раствора КБ. Для электрофизиологических экспериментов градиентная рекальцинация остро изолированных клеток была выполнена с использованием 1 мМ раствора Ca 2+ в течение 30 минут до конечной концентрации 1.8 мМ.
Супернатант удаляли, а осажденные предсердные кардиомиоциты ресуспендировали в 4 мл (в зависимости от выхода) раствора КБ. Для электрофизиологических экспериментов градиентная рекальцинация остро изолированных клеток была выполнена с использованием 1 мМ раствора Ca 2+ в течение 30 минут до конечной концентрации 1.8 мМ.
Выделение предсердных кардиомиоцитов человека проводили, как описано ранее. 32 Образцы человеческих предсердий сначала помещали в ледяной раствор Тирода, не содержащий Ca 2+ (100 мМ NaCl, 10 мМ KCl, 1,2 мМ KH 50 мМ таурина, 5 мМ MOPS и 20 мМ глюкозы, рН доводили до 7,0 с помощью NaOH) с добавлением моноксима 2,3-бутандиона (BDM, 30 мМ, Sigma, США), нарезанного на мелкие кусочки и промытого 3 раза в течение 5 мин с раствором Тирода, не содержащим Ca 2+ .На всех стадиях растворы оксигенировали 100% O 2 при 35 °C. Затем небольшие кусочки переносили в раствор Tyrode, не содержащий Ca 2+ , содержащий 286 ед/мл коллагеназы типа I (Worthington, США) и 3,5–5 ед/мл протеазы типа XXIV (Sigma, США), и осторожно расщепляли в течение 15 мин. , затем супернатант отбрасывали. Последующие расщепления включали только коллагеназу типа I, а концентрация Ca 2+ повышалась до 0,2 мМ. Перемешивание продолжали до тех пор, пока не были видны палочковидные поперечно-полосатые кардиомиоциты.Суспензию осторожно центрифугировали (100× г ), кардиомиоциты ресуспендировали и хранили до использования в растворе Тирода, содержащем Ca 2+ (0,5 мМ) (без BDM) при комнатной температуре.
, затем супернатант отбрасывали. Последующие расщепления включали только коллагеназу типа I, а концентрация Ca 2+ повышалась до 0,2 мМ. Перемешивание продолжали до тех пор, пока не были видны палочковидные поперечно-полосатые кардиомиоциты.Суспензию осторожно центрифугировали (100× г ), кардиомиоциты ресуспендировали и хранили до использования в растворе Тирода, содержащем Ca 2+ (0,5 мМ) (без BDM) при комнатной температуре.
Иммунофлуоресцентный анализ
Для иммунофлуоресцентного анализа предсердных кардиомиоцитов крысы клетки фиксировали свежим 4% параформальдегидом в течение 15 минут и дважды промывали PBS в течение 5 минут. Затем фиксированные клетки пермеабилизировали в 0,1% тритоне Х-100 в PBS (PBST) в течение 10 мин, блокировали 4% нормальной козьей сывороткой в течение 1 ч при комнатной температуре и инкубировали с первичными антителами при 4 °C в течение ночи.Используемые первичные антитела перечислены ниже: анти-ANP (Санта-Круз, США), анти-GLS (Abcam, США), анти-GRIA3 (Abcam, США), анти-GRIN1 (Novus, США), анти-EAAT1 (Abcam , США), анти-CAST (Invitrogen, США) и анти-глутамат (Sigma, США). Затем клетки промывали PBST и инкубировали со вторичными антителами, конъюгированными с Alexa Fluor (Abcam, США), в течение 1 ч при комнатной температуре. После трех промывок PBST ядра окрашивали DAPI. Конфокальные изображения одного среза или стопки z были получены на конфокальном лазерном сканирующем микроскопе Leica SP8 (Leica, США) с использованием 63-кратного иммерсионного объектива.Трехмерную реконструкцию и флуоресцентный анализ изображений проводили с помощью программ LAS X (Leica, США) и ImageJ 2.0 (NIH, США).
Затем клетки промывали PBST и инкубировали со вторичными антителами, конъюгированными с Alexa Fluor (Abcam, США), в течение 1 ч при комнатной температуре. После трех промывок PBST ядра окрашивали DAPI. Конфокальные изображения одного среза или стопки z были получены на конфокальном лазерном сканирующем микроскопе Leica SP8 (Leica, США) с использованием 63-кратного иммерсионного объектива.Трехмерную реконструкцию и флуоресцентный анализ изображений проводили с помощью программ LAS X (Leica, США) и ImageJ 2.0 (NIH, США).
Для иммунофлуоресцентного окрашивания срезов ткани предсердий образцы фиксировали 4% параформальдегидом, заливали парафином и делали срезы, срезы ткани депарафинизировали ксилолом и регидратировали с помощью ряда уменьшающихся концентраций этанола. Извлечение антигена осуществляли путем нагревания в цитратном растворе в течение 10 мин в микроволновой печи; затем предметные стекла блокировали 10% козьей сывороткой и инкубировали с первичными антителами при 4 °C в течение ночи.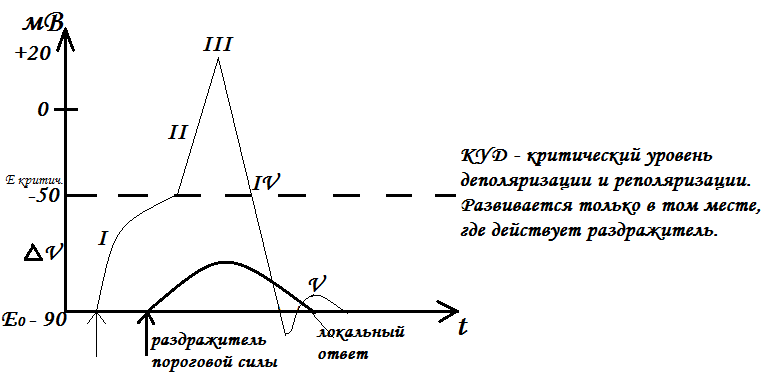 На следующий день предметные стекла трижды промывали в PBST для удаления несвязавшихся антител, а затем инкубировали с соответствующими флуоресцентно-конъюгированными вторичными антителами в PBST с 10% козьей сывороткой в течение 1 ч при комнатной температуре. Предметные стекла трижды промывали в PBST, а затем проводили окрашивание DAPI (Sigma, США) для визуализации ядер.
На следующий день предметные стекла трижды промывали в PBST для удаления несвязавшихся антител, а затем инкубировали с соответствующими флуоресцентно-конъюгированными вторичными антителами в PBST с 10% козьей сывороткой в течение 1 ч при комнатной температуре. Предметные стекла трижды промывали в PBST, а затем проводили окрашивание DAPI (Sigma, США) для визуализации ядер.
Экстракция РНК и количественная ПЦР
Для выделения РНК из одной клетки отдельные клетки собирали вручную под микроскопом и переносили в 1.5 мкл буфера для лизиса клеток для извлечения общей РНК. РНК превращали в первую цепь кДНК в течение 9 циклов с использованием Superscript III (Invitrogen, США), а затем амплифицировали в течение 18 циклов ПЦР с помощью КАРА-полимеразы (Kapa Biosystems, США). Количественную оценку кДНК проводили с помощью программы Qubit 3.0 (Thermo Fisher, США). кДНК правильно разбавляли и использовали в качестве матрицы для количественной ПЦР.
Все реакции количественной ПЦР проводили с использованием набора SYBR Premix Ex Taq Kit (TAKARA, Япония) на установке QuantStudio 6 Flex (Applied Biosystems, США). Данные количественной ПЦР были сгенерированы с помощью системного программного обеспечения QuantStudio Real-Time PCR. Кроме того, продукты полу-КПЦР анализировали с помощью электрофореза в 2% агарозном геле. Используемые праймеры перечислены в дополнительной информации, Таблица S1.
Данные количественной ПЦР были сгенерированы с помощью системного программного обеспечения QuantStudio Real-Time PCR. Кроме того, продукты полу-КПЦР анализировали с помощью электрофореза в 2% агарозном геле. Используемые праймеры перечислены в дополнительной информации, Таблица S1.
Вестерн-блот-анализ
Суммарные белки экстрагировали традиционным методом с использованием буфера для лизиса RIPA (Beyotime Biotechnology, Китай) с коктейлем ингибиторов протеаз (Beyotime Biotechnology, Китай) при 4 °C. Концентрацию белков определяли с помощью анализа белков BCA (Beyotime Biotechnology, Китай), и равные количества общих белков разделяли с помощью 10% SDS-PAGE (Thermo Fisher Scientific, США), а затем переносили на мембраны PVDF (Millipore, США).Далее мембраны блокировали 5 % обезжиренным молоком в 0,1 % промывочном буфере Tween в течение 1 ч при комнатной температуре, а затем инкубировали с первичными антителами, включая анти-GLS (Abcam, США), анти-GRIA3 (Abcam, США), анти -GRIN1 (Novus, США), анти-EAAT1 (Abcam, США) и анти-GAPDH (Abcam, США) в течение ночи при 4 °C. На следующий день мембраны инкубировали с конъюгированным вторичным антителом (Invitrogen, США) в течение 1 ч при комнатной температуре и визуализировали полосы с помощью ChemiDoc Touch Gel Imaging System (Bio-Rad, США).Блоты количественно определяли денситометрией с использованием программного обеспечения ImageJ (NIH).
На следующий день мембраны инкубировали с конъюгированным вторичным антителом (Invitrogen, США) в течение 1 ч при комнатной температуре и визуализировали полосы с помощью ChemiDoc Touch Gel Imaging System (Bio-Rad, США).Блоты количественно определяли денситометрией с использованием программного обеспечения ImageJ (NIH).
Пэтч-кламп
Пэтч-клампы для целых клеток применяли для регистрации мембранных потенциалов и токов. Использовали два микроэлектрода из боросиликатного стекла: один для электрической стимуляции и регистрации с сопротивлением 2–5 МОм, заполненный пипеточным раствором, а другой для пульсирующего введения с наконечником с внутренним диаметром 10 мкМ, заполненным раствором для ванн или раствором для ванн, содержащим различные фармакологические агенты.Давление в каналах аппликационных пипеток регулировали с помощью микроинъектора пикоинжектора PL1-100 (Harvard Apparatus, США). Импульсы стимуляции подавались через усилитель EPC-10 (HEKA, Германия), синхронизированный с микроинъектором для синхронного высвобождения лекарственного средства. Потенциал действия или ток регистрировали в режиме фиксации тока или фиксации напряжения, все данные собирали с помощью усилителя EPC-10 с помощью стандартных методов пэтч-кламп при комнатной температуре. Раствор для ванны содержал следующие ингредиенты: 135 мМ NaCl, 10 мМ глюкозы, 5.4 мМ KCl, 1,8 мМ CaCl 2 , 0,3 мМ Na 2 HPO 4 , 0,3 мМ KH 2 PO 4 , 10 мМ HEPES (рН 7,35, отрегулировано с помощью NaOH 7,35). Стандартный раствор пипетки содержал следующие ингредиенты: 140 мМ KCl, 10 мМ EGTA, 10 мМ HEPES, 5 мМ глюкозы, 3 мМ Na 2 АТФ, рН доводили до 7,2 с помощью КОН. Данные анализировали с помощью программного обеспечения Patchmaster 2.42 (HEKA, Германия) и Clampfit 10.7 (Molecular Devices, CA).
Потенциал действия или ток регистрировали в режиме фиксации тока или фиксации напряжения, все данные собирали с помощью усилителя EPC-10 с помощью стандартных методов пэтч-кламп при комнатной температуре. Раствор для ванны содержал следующие ингредиенты: 135 мМ NaCl, 10 мМ глюкозы, 5.4 мМ KCl, 1,8 мМ CaCl 2 , 0,3 мМ Na 2 HPO 4 , 0,3 мМ KH 2 PO 4 , 10 мМ HEPES (рН 7,35, отрегулировано с помощью NaOH 7,35). Стандартный раствор пипетки содержал следующие ингредиенты: 140 мМ KCl, 10 мМ EGTA, 10 мМ HEPES, 5 мМ глюкозы, 3 мМ Na 2 АТФ, рН доводили до 7,2 с помощью КОН. Данные анализировали с помощью программного обеспечения Patchmaster 2.42 (HEKA, Германия) и Clampfit 10.7 (Molecular Devices, CA).
Измерение скорости проводимости предсердий in vivo
Для имплантации сердечного катетера в предсердия через правую яремную вену был сделан 5-сантиметровый разрез на правой стороне шеи, и наружная яремная вена была открыта путем тупой диссекции. Затем в правое предсердие вводили электрофизиологический катетер 1.1F (Millar Instruments, США) и регистрировали электрические сигналы с помощью 16-канальной PowerLab (ADInstruments, США). Электрокардиограммы собирали и анализировали с помощью LabChart 7 (ADInstruments, США), а скорость проведения рассчитывали по уравнению: CV = Δs/Δt, в котором Δs — расстояние между двумя электродами, а Δt — время между точками с максимальным наклоном деполяризации на электрокардиограммах.
Затем в правое предсердие вводили электрофизиологический катетер 1.1F (Millar Instruments, США) и регистрировали электрические сигналы с помощью 16-канальной PowerLab (ADInstruments, США). Электрокардиограммы собирали и анализировали с помощью LabChart 7 (ADInstruments, США), а скорость проведения рассчитывали по уравнению: CV = Δs/Δt, в котором Δs — расстояние между двумя электродами, а Δt — время между точками с максимальным наклоном деполяризации на электрокардиограммах.
Оптическое картирование
Оптическое картирование на основе красителя, чувствительного к напряжению, выполняли, как описано ранее. 33 Вкратце, взрослых самцов крыс Sprague-Dawley (возраст 6–8 недель) анестезировали пентобарбиталом (25 мг/кг), внутрибрюшинно содержащим 120 МЕ гепарина. После срединного разреза сердце быстро вырезали и подвергали перфузии по Лангендорфу при 37°C оксигенированным (95% O 2 и 5% CO 2 ) стандартным раствором Тирода. Сердце окрашивали потенциалчувствительным красителем Rh337 (30 мкМ, ААТ, США) в течение 5 мин. В перфузат добавляли блеббистатин (10 мкМ, MCE, США) для устранения артефактов движения при оптических записях. Краситель возбуждали с помощью светодиодных источников света с центром на 550 нм. Изображения были получены с помощью высокоскоростной камеры с разрешением 10000 пикселей (SciMedia, MiCAM ULTIMA, США) со скоростью 1000 кадров/с. Анализ данных проводили с помощью программ Brainvision 16.04 (SciMedia, США), предназначенных для анализа оптически регистрируемых потенциалов действия. Локальную активацию определяли как момент времени максимального изменения флуоресценции во времени для каждого флуоресцентного сигнала на изображении.Скорость проводимости рассчитывали по градиенту карт активации.
В перфузат добавляли блеббистатин (10 мкМ, MCE, США) для устранения артефактов движения при оптических записях. Краситель возбуждали с помощью светодиодных источников света с центром на 550 нм. Изображения были получены с помощью высокоскоростной камеры с разрешением 10000 пикселей (SciMedia, MiCAM ULTIMA, США) со скоростью 1000 кадров/с. Анализ данных проводили с помощью программ Brainvision 16.04 (SciMedia, США), предназначенных для анализа оптически регистрируемых потенциалов действия. Локальную активацию определяли как момент времени максимального изменения флуоресценции во времени для каждого флуоресцентного сигнала на изображении.Скорость проводимости рассчитывали по градиенту карт активации.
Запись МЭА в предсердных кардиомиоцитах, полученных из иПСК человека
Предсердные кардиомиоциты, полученные из иПСК человека, были получены из Института биофизики Китайской академии наук (Пекин, Китай). После восстановления клетки культивировали в чашке, предварительно покрытой витронектином (0,01 мкг/мкл, Cauliscell, Китай) с поддерживающей средой для кардиомиоцитов (Cauliscell, Китай) при 37 °C и 5% CO 2 . Среду меняли каждые 2 дня. На 5-й день клетки диссоциировали и ресуспендировали в поддерживающей сердечной среде с плотностью 1 × 10 7 клеток/мл. Аликвоты по 10 мкл клеточной суспензии высевали на планшеты CytoView MEA 24 (Axion BioSystems, США) для возможной регистрации. Устройство MEA автоматически регулировало и контролировало окружающую среду (37 °C и 5% CO 2 ) для поддержания температуры и pH среды. Данные были получены с использованием многолуночной платформы МЭА Maestro Pro (Axion BioSystems, США).
Среду меняли каждые 2 дня. На 5-й день клетки диссоциировали и ресуспендировали в поддерживающей сердечной среде с плотностью 1 × 10 7 клеток/мл. Аликвоты по 10 мкл клеточной суспензии высевали на планшеты CytoView MEA 24 (Axion BioSystems, США) для возможной регистрации. Устройство MEA автоматически регулировало и контролировало окружающую среду (37 °C и 5% CO 2 ) для поддержания температуры и pH среды. Данные были получены с использованием многолуночной платформы МЭА Maestro Pro (Axion BioSystems, США).
Трансфекция siRNA
Опосредованный siRNA нокдаун GRIA3 и GRIN1 проводили с помощью липофектамина RNAiMAX (Invitrogen, США) в соответствии с рекомендациями производителя. Вкратце, культивированные предсердные кардиомиоциты, полученные из иПСК человека, трансфицировали конечной концентрацией миРНК ~20 пмоль вместе с 3 мкл липофектамина RNAiMAX на 500 мкл. Последующие эксперименты проводили через 72 ч после трансфекции. Все последовательности миРНК перечислены в дополнительной информации, таблица S2.
Перфузия сердца по Лангендорфу и индукция ФП
Модель индукции ФП у крыс была основана на ранее описанном исследовании. 34 Вкратце, крысу анестезировали пентобарбиталом (25 мг/кг) внутрибрюшинно, грудную клетку вскрыли для доступа к сердцу. Затем сердце быстро удаляли и помещали в перфузионную систему Лангендорфа, перфузируемую оксигенированным перфузионным раствором, содержащим 135 мМ NaCl, 10 мМ глюкозы, 5,4 мМ KCl, 1,8 мМ CaCl 2 , 0,3 мМ Na 2 HPO 4 , 0.3 мМ KH 2 PO 4 , 10 мМ HEPES (pH 7,35, отрегулировано NaOH) при 37 °C. Электрокардиограмму (ЭКГ) и интракардиальные записи правого предсердия (РАЭ) непрерывно записывали из сердца с перфузией по Лангендорфу с помощью усилителя Powerlab (Powerlab, AD Instruments, США). Электрофизиологический катетер 1,1F (Millar Instruments, США) вводили через яремную вену для стимуляции внутренней стенки правого предсердия. Пакетную стимуляцию генерировали внешним стимулятором (STG-3008-FA, Multi-Channel Systems, Германия). Порог стимуляции определяли путем постепенного увеличения напряжения до тех пор, пока не возникал предсердный захват; затем стимуляция выполнялась при 2-кратном пороге стимуляции. Для индуцирования ФП использовали снижение перфузии и импульсную стимуляцию. После баланса перфузии в течение 30 минут мы уменьшили коронарный поток с 6 мл/мин до 3 мл/мин и стимулировали предсердия импульсной стимуляцией с частотой 40 Гц в течение 1 минуты (повторяли до десяти раз с интервалом в 1 минуту). вызвать ФП. Каждая крыса подвергалась идентичному протоколу кардиостимуляции и запрограммированной стимуляции.Успешная индукция ФП определялась как нерегулярная предсердная частота при РАЭ продолжительностью более 2 минут, а частота индукции определялась как число успешно индуцированных сердечных сокращений по отношению к общему числу стимулированных сердец.
Порог стимуляции определяли путем постепенного увеличения напряжения до тех пор, пока не возникал предсердный захват; затем стимуляция выполнялась при 2-кратном пороге стимуляции. Для индуцирования ФП использовали снижение перфузии и импульсную стимуляцию. После баланса перфузии в течение 30 минут мы уменьшили коронарный поток с 6 мл/мин до 3 мл/мин и стимулировали предсердия импульсной стимуляцией с частотой 40 Гц в течение 1 минуты (повторяли до десяти раз с интервалом в 1 минуту). вызвать ФП. Каждая крыса подвергалась идентичному протоколу кардиостимуляции и запрограммированной стимуляции.Успешная индукция ФП определялась как нерегулярная предсердная частота при РАЭ продолжительностью более 2 минут, а частота индукции определялась как число успешно индуцированных сердечных сокращений по отношению к общему числу стимулированных сердец.
Статистический анализ
Нормальность проверялась с помощью критерия Колмогорова–Смирнова. Двусторонний критерий Стьюдента t или непараметрический критерий использовали для сравнения между двумя группами. Скорость индукции ФП анализировали с помощью теста х 2 .Статистическая значимость определялась как значение P <0,05. Нормально распределенные данные показаны как среднее ± стандартная ошибка среднего (SEM), а ненормально распределенные данные показаны как медианы ± межквартильный размах (IQR). Все статистические анализы проводились с помощью GraphPad Prism 8.
Скорость индукции ФП анализировали с помощью теста х 2 .Статистическая значимость определялась как значение P <0,05. Нормально распределенные данные показаны как среднее ± стандартная ошибка среднего (SEM), а ненормально распределенные данные показаны как медианы ± межквартильный размах (IQR). Все статистические анализы проводились с помощью GraphPad Prism 8.
Типы мышечных клеток человека | Интерактивное руководство по анатомии
Мышечная ткань, один из четырех основных типов тканей, играет жизненно важную роль в обеспечении движения и выработки тепла органами тела.Внутри мышечной ткани выделяют три отдельные группы тканей: скелетные мышцы , сердечные мышцы и гладкие мышцы . Каждая из этих групп тканей состоит из специализированных клеток, которые придают ткани ее уникальные свойства. Продолжите прокрутку, чтобы прочитать больше ниже…
Нажмите, чтобы просмотреть большое изображение
Продолжение сверху. ..
..Скелетная мышца
Скелетные мышцы — это наиболее распространенная и широко распространенная мышечная ткань в организме, составляющая около 40% общей массы тела. Он образует все скелетные мышцы, такие как двуглавая мышца плеча и большая ягодичная мышца, и находится в глазах, горле, диафрагме и заднем проходе. Четыре характеристики определяют клетки ткани скелетных мышц: они произвольные, поперечно-полосатые, неразветвленные и многоядерные.
Скелетная мышечная ткань является единственной мышечной тканью, находящейся под непосредственным сознательным контролем коры головного мозга, что дает ей название произвольной мышцы.Все осознанные движения тела, включая движения конечностей, мимику, движения глаз и глотание, являются продуктами скелетной мышечной ткани. Сокращение скелетных мышц также производит большую часть тепла тела как побочный продукт клеточного метаболизма.
Структура клеток скелетных мышц также делает их уникальными среди мышечных тканей. Скелетные мышечные клетки развиваются в результате слияния множества более мелких клеток во время внутриутробного развития, в результате чего образуются длинные прямые мышечные волокна, содержащие много ядер.При рассмотрении под микроскопом клетки скелетных мышц выглядят полосатыми или исчерченными узорами из светлых и темных областей. Эти полосы вызваны правильным расположением белков актина и миозина внутри клеток в структуры, известные как миофибриллы. Миофибриллы отвечают за огромную силу скелетных мышц и способность тянуть с невероятной силой и толкать тело.
Скелетные мышечные клетки развиваются в результате слияния множества более мелких клеток во время внутриутробного развития, в результате чего образуются длинные прямые мышечные волокна, содержащие много ядер.При рассмотрении под микроскопом клетки скелетных мышц выглядят полосатыми или исчерченными узорами из светлых и темных областей. Эти полосы вызваны правильным расположением белков актина и миозина внутри клеток в структуры, известные как миофибриллы. Миофибриллы отвечают за огромную силу скелетных мышц и способность тянуть с невероятной силой и толкать тело.
Сердечная мышца
Клетки сердечной мышцы находятся только в сердце и специализируются на мощном и эффективном перекачивании крови на протяжении всей нашей жизни.Четыре характеристики определяют клетки сердечной мышечной ткани: они непроизвольны и внутренне контролируются, поперечно-полосатые, разветвленные и одноядерные.
Сердечная мышца считается непроизвольной тканью, поскольку она бессознательно контролируется областями ствола головного мозга и гипоталамуса. Он также считается внутренней или самоконтролируемой тканью, поскольку нормальный сердечный ритм задается специализированными кардиостимуляторными клетками сердечной мышцы в самом сердце. Клетки сердечной мышечной ткани короче скелетной мышечной ткани и образуют между собой сеть из множества разветвлений.Между клетками сердечной мышцы образуются вставочные диски из перекрывающихся клеточных мембран, которые плотно скрепляют их и обеспечивают быстрое прохождение электрохимических сигналов между клетками. Клетки не сливаются во время развития, оставляя каждую клетку с одним ядром. Одной из общих черт скелетной и сердечной мышц является наличие исчерченности из-за расположения актина и миозина в регулярные миофибриллы. Наличие миофибрилл и множества митохондрий в клетках сердечной мышцы обеспечивает им большую силу и выносливость, чтобы перекачивать кровь на протяжении всей жизни.
Он также считается внутренней или самоконтролируемой тканью, поскольку нормальный сердечный ритм задается специализированными кардиостимуляторными клетками сердечной мышцы в самом сердце. Клетки сердечной мышечной ткани короче скелетной мышечной ткани и образуют между собой сеть из множества разветвлений.Между клетками сердечной мышцы образуются вставочные диски из перекрывающихся клеточных мембран, которые плотно скрепляют их и обеспечивают быстрое прохождение электрохимических сигналов между клетками. Клетки не сливаются во время развития, оставляя каждую клетку с одним ядром. Одной из общих черт скелетной и сердечной мышц является наличие исчерченности из-за расположения актина и миозина в регулярные миофибриллы. Наличие миофибрилл и множества митохондрий в клетках сердечной мышцы обеспечивает им большую силу и выносливость, чтобы перекачивать кровь на протяжении всей жизни.
Висцеральная мышца
Висцеральные мышечные клетки находятся в органах, кровеносных сосудах и бронхиолах тела для перемещения веществ по всему телу. Висцеральные мышцы также широко известны как гладкие мышцы из-за отсутствия исчерченности. Четыре характеристики определяют клетки гладкой мышечной ткани: они непроизвольно контролируются, не исчерчены, не разветвлены и имеют одно ядро.
Висцеральные мышцы также широко известны как гладкие мышцы из-за отсутствия исчерченности. Четыре характеристики определяют клетки гладкой мышечной ткани: они непроизвольно контролируются, не исчерчены, не разветвлены и имеют одно ядро.
Бессознательные области мозга контролируют висцеральные мышцы через вегетативную и кишечную нервные системы.Таким образом, висцеральные мышцы непроизвольно контролируются. Об этом свидетельствует наша неспособность сознательно контролировать многие физиологические процессы, такие как кровяное давление или пищеварение. Каждая висцеральная мышечная клетка длинная и тонкая, с одним центральным ядром и множеством белковых волокон. Белковые волокна организованы в нити, называемые промежуточными нитями, и массы, известные как плотные тела. Промежуточные филаменты сокращаются, сближая плотные тела и сокращая висцеральные мышечные клетки. Каждая висцеральная мышечная клетка очень слаба, но работая вместе, эти клетки могут производить мощные и продолжительные сокращения. Например, многие висцеральные мышечные клетки в матке способны сокращаться вместе, чтобы вытолкнуть плод из матки во время родов.
Например, многие висцеральные мышечные клетки в матке способны сокращаться вместе, чтобы вытолкнуть плод из матки во время родов.
2. Нервные и мышечные клетки
Berne RM, Levy MN (1993): Physiology , 3-е изд., 1091 стр. CV Mosby, Сент-Луис.
Буллок Т.Х. (1959): Доктрина нейронов и электрофизиология. Наука 129:(3355) 997-1002.
Дэвис Л.Дж., Лоренте де Н.Р. (1947): Вклад в математическую теорию электротонуса. Шпилька. Институт Рокфеллера. Мед. Рез. 131: 442-96.
Эльсберг, Калифорния (1931): Хирургический папирус Эдвина Смита. Энн. Мед. История 3: 271-9.
Ganong WF (1991): Review of Medical Physiology , 15-е изд., Appleton & Lange, Norwalk, Conn.
Guyton AC (1992): Human Physiology and Mechanisms of Disease , 5-е изд., 690 стр. Saunders, Philadelphia.
Герман Л. (1872 г.): Grundriss der Physiologie , 4-е изд., (Цитируется по L Hermann (1899): Zur Theorie der Erregungsleitung und der elektrischen Erregung. Pflger Arch. ges. Physiol. 75: 574-90.)
Pflger Arch. ges. Physiol. 75: 574-90.)
Hermann L (1905): Lehrbuch der Physiologie , 13-е изд., 762 стр. Август Хиршвальд, Берлин.
Kandel ER, Schwartz JH (1985): Принципы нейронауки , Elsevier Publishing, Нью-Йорк.
Lorente de NR (1947): A Study of Nerve Physiology , 293 стр. Институт медицинских исследований Рокфеллера, Нью-Йорк.
Мулер А.Л., Маркин В.С. (1978): Электрические свойства анизотропных нервно-мышечных синцитиев — II. Распространение плоского фронта возбуждения. Биофиз. 22: 536-41.
Нуньес П.Л. (1981): Электрические поля мозга: нейрофизика ЭЭГ , 484 стр. Издательство Оксфордского университета, Нью-Йорк.
Patton HD, Fuchs AF, Hille B, Scher AM, Steiner R (eds.) (1989): Учебник физиологии , 21-е изд., 1596 стр. WB Saunders, Philadelphia.
Руч Т.К., Паттон HD (ред.) (1982): Physiology and Biophysics , 20-е изд., 1242 стр. WB Saunders, Philadelphia.
Schad JP, Ford DH (1973): Basic Neurology , 2-е изд. , 269 стр. Elsevier Scientific Publishing, Амстердам.
, 269 стр. Elsevier Scientific Publishing, Амстердам.
Thompson CF (1985): Мозг — Введение в неврологию , 363 стр. WH Freeman, Нью-Йорк.
%PDF-1.5
%
7 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
16 0 объект
>>>/BBox[0 0 595.44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
13 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
14 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
1 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
6 0 объект
>>>/BBox[0 0 595.44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
12 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
17 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
3 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
5 0 объект
>>>/BBox[0 0 595. 44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
8 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
15 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
4 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
9 0 объект
>>>/BBox[0 0 595.44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
10 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
11 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
18 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
20 0 объект
>поток
; изменено с помощью iText 4.
44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
8 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
15 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
4 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
9 0 объект
>>>/BBox[0 0 595.44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
10 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
11 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
18 0 объект
>>>/BBox[0 0 595,44 841,68]/длина 114>>поток
х%А
P_CBZFyI o[~7P3*HOg#»E|)2n:hgspoken˨W
| ڛ2ү/bj
конечный поток
эндообъект
20 0 объект
>поток
; изменено с помощью iText 4. 2.0 от 1T3XT2022-02-01T18:56:07-08:00
2.0 от 1T3XT2022-02-01T18:56:07-08:00
