
Ч 2 ст 243 тк рф: ТК РФ Статья 243. Случаи полной материальной ответственности / КонсультантПлюс
Статья 243 ТК РФ 2016-2019. Случаи полной материальной ответственности. ЮрИнспекция
Порядок выдачи спецодежды работникам строительных организаций регулируется Приказом Минздравсоцразвития России от 16.07.2007 N 477 . Поскольку работодатель обязан сообщить работникам о полагающихся им средствах индивидуальной защиты (ч. 2 ст. 212 ТК РФ) , каждого работника необходимо ознакомить с правилами и нормами выдачи средств индивидуальной защиты при заключении с ним трудового договора (п. 9 Межотраслевых правил, утвержденных Приказом Минздравсоцразвития России от 01.06.2009 N 290н) .После того как работодатель определил, по какой профессии, каким работникам, в каком количестве и на какой срок выдают СИЗ, он должен зафиксировать это во внутренних документах. Правила и нормы выдачи СИЗ могут быть установлены приложением к коллективному договору или самостоятельным локальным актом.Выдача и возврат специальной одежды должны отражаться в личных карточках работников (п. 61 Методических указаний, утвержденных Приказом Минфина России от 26. 12.2002 N 135н) .При выдаче СИЗ с работником необходимо заключить договор о передаче ему СИЗ в пользование или оформить разовый документ приема-передачи, поскольку в соответствии с п. 2 ч. 1 ст. 243 ТК РФ материальная ответственность в полном объеме причиненного ущерба возлагается на работника в случае недостачи ценностей, вверенных ему на основании специального письменного договора или полученных им по разовому документу. Согласно п. 64 Методических указаний специальная одежда, выданная работникам, является собственностью организации и подлежит возврату при увольнении. Обязанность работника возмещать причиненный работодателю прямой действительный ущерб предусмотрена ст. 238 ТК РФ, поэтому расторжение трудового договора не влечет освобождения работника от материальной ответственности за невозврат спецодежды. Таким образом, в случае правильного оформления выдачи спецодежды при невозвращении ее работником, увольняющимся из организации, работодатель вправе требовать возмещения стоимости спецодежды (срок использования которой не истек) с учетом степени ее износа (ст.
12.2002 N 135н) .При выдаче СИЗ с работником необходимо заключить договор о передаче ему СИЗ в пользование или оформить разовый документ приема-передачи, поскольку в соответствии с п. 2 ч. 1 ст. 243 ТК РФ материальная ответственность в полном объеме причиненного ущерба возлагается на работника в случае недостачи ценностей, вверенных ему на основании специального письменного договора или полученных им по разовому документу. Согласно п. 64 Методических указаний специальная одежда, выданная работникам, является собственностью организации и подлежит возврату при увольнении. Обязанность работника возмещать причиненный работодателю прямой действительный ущерб предусмотрена ст. 238 ТК РФ, поэтому расторжение трудового договора не влечет освобождения работника от материальной ответственности за невозврат спецодежды. Таким образом, в случае правильного оформления выдачи спецодежды при невозвращении ее работником, увольняющимся из организации, работодатель вправе требовать возмещения стоимости спецодежды (срок использования которой не истек) с учетом степени ее износа (ст. 246 ТК РФ) . Согласно ст. 241 ТК РФ за причиненный организации ущерб работник несет материальную ответственность в пределах среднего месячного заработка, если иное не установлено федеральным законом. В случае умышленного причинения ущерба устанавливается полная материальная ответственность работника, предполагающая возмещение в полном размере (ст. ст. 242 и 243 ТК РФ) . Обязанность по установлению размера и причин возникновения ущерба возложена на работодателя ст. 247 ТК РФ.Работодатель, согласно ст. 248 ТК РФ, вправе удержать из заработной платы работника сумму, не превышающую размера среднего месячного заработка работника. Ст. 137 ТК РФ введены ограничения оснований удержаний. Удержания за причиненный ущерб в ней не поименованы.В связи с этим необходимо заручиться письменным согласием работника на удержание из его заработной платы суммы, не превышающей размера его среднего месячного заработка.Если работодатель и работник не пришли к согласию о возмещении ущерба или о его размере, то они вправе обратиться в суд.
246 ТК РФ) . Согласно ст. 241 ТК РФ за причиненный организации ущерб работник несет материальную ответственность в пределах среднего месячного заработка, если иное не установлено федеральным законом. В случае умышленного причинения ущерба устанавливается полная материальная ответственность работника, предполагающая возмещение в полном размере (ст. ст. 242 и 243 ТК РФ) . Обязанность по установлению размера и причин возникновения ущерба возложена на работодателя ст. 247 ТК РФ.Работодатель, согласно ст. 248 ТК РФ, вправе удержать из заработной платы работника сумму, не превышающую размера среднего месячного заработка работника. Ст. 137 ТК РФ введены ограничения оснований удержаний. Удержания за причиненный ущерб в ней не поименованы.В связи с этим необходимо заручиться письменным согласием работника на удержание из его заработной платы суммы, не превышающей размера его среднего месячного заработка.Если работодатель и работник не пришли к согласию о возмещении ущерба или о его размере, то они вправе обратиться в суд. При увольнении работника, получившего спецодежду по правильно оформленным первичным документам, работодатель вправе требовать ее возврата или оплаты ущерба. В случае если работник не согласен возместить причиненный ущерб в добровольном порядке, работодатель может обратиться в суд с иском о возмещении ущерба.Срок исковой давности для обращения работодателя в суд равен одному году (ч. 2 ст. 392 ТК РФ).
При увольнении работника, получившего спецодежду по правильно оформленным первичным документам, работодатель вправе требовать ее возврата или оплаты ущерба. В случае если работник не согласен возместить причиненный ущерб в добровольном порядке, работодатель может обратиться в суд с иском о возмещении ущерба.Срок исковой давности для обращения работодателя в суд равен одному году (ч. 2 ст. 392 ТК РФ).Материальная ответственность работников
Сотрудник отвечает за деньги и товары, с которыми работает. Чтобы закрепить обязанность на бумаге, заключают договоры о полной материальной ответственности.
Они пригодятся, если будет недоимка, а сотрудник откажется платить.
Если предпочитаете юридический язык, перейдите по ссылкам ниже, а мы постараемся объяснить всё простыми словами.
Глава 39 Трудового кодекса РФ
Постановление Минтруда РФ от 31.12.2002 г. № 85
Постановление Пленума ВС РФ от 17.03.2004 г. № 2
Обзор практики судов о материальной ответственности работников от 05. 12.2018 г.
12.2018 г.
Зачем нужен договор о полной материальной ответственности работника
Сотрудники отвечают за ущерб даже без договора. Устроили человека на работу — он обязан не вредить и быть внимательным по ст. 232 ТК РФ.
Это ограниченная материальная ответственность. Ограниченная — потому что сотрудник отвечает в пределах среднего заработка по ст. 241 ТК РФ. Если он угробит автомобиль компании ценой 1 млн, спросить с него можно только небольшую часть суммы.
Мастер пролил клей на десять рулонов дизайнерских обоев. На обои ушло 30 000 ₽, а зарплата мастера 20 000 ₽. Значит, работодатель выложит 10 000 ₽ из своего кармана.
В ст. 243 ТК РФ есть исключения. Если сотрудник причинил ущерб в пьяном виде, патихардил или воровал, он всё равно платит полную сумму. Но возникает другая проблема: доказать, что он действительно был пьян или что своровал именно он.
По договору работник платит полностьюДоговор о материальной ответственности снимает ограничения Трудового кодекса. Сотрудник компенсирует столько ущерба, сколько нанёс, без уменьшения суммы до размера одной зарплаты.
Сотрудник компенсирует столько ущерба, сколько нанёс, без уменьшения суммы до размера одной зарплаты.
Общие ограничения полной материальной ответственности
Конечно, работодатель не взыскивает ущерб абсолютно свободно. Ограничения всё равно есть, об этом сказано в Обзоре практики судов от 05.12.2018 г.
Реальный ущербРаботник оплачивает только реальный ущерб работодателя — ст. 238 ТК РФ. Это недостача денег, порча продуктов, кражи, выплаты по претензиям клиентов или штрафы от госорганов. Упущенную прибыль с сотрудника взыскать нельзя.
Курьер потерял планшеты на 100 000 ₽, которые вёз клиенту. Работодателю пришлось заказать новые и отменить запланированную закупку на ещё 500 000 ₽. Исчезновение планшетов — реальный ущерб, за него сотрудник отвечает. Сорвавшийся заказ и последующие продажи — упущенная выгода, которую работник компенсировать не должен.
Суд имеет право уменьшить долг работника. Он принимает во внимание, какая зарплата у сотрудника и был ли кто-то ещё виноват в ущербе. Исключение — если человек совершил преступление: украл деньги или организовал сбыт товаров на сторону. Тогда уменьшить сумму нельзя по 250 ТК РФ.
Он принимает во внимание, какая зарплата у сотрудника и был ли кто-то ещё виноват в ущербе. Исключение — если человек совершил преступление: украл деньги или организовал сбыт товаров на сторону. Тогда уменьшить сумму нельзя по 250 ТК РФ.
Суд не даёт скидку всем подряд. Будет ли она — зависит от искусности работника во время процесса.
Вина работникаНельзя брать деньги у сотрудника, если он не виноват — ст. 233 ТК РФ. Сотрудник не отвечает за форс-мажоры вроде потопа, пожара и ограбления. Он не следит за товарами во время чужой смены или ночью. Все похожие обстоятельства перечислены в ст. 239 ТК РФ.
🎁
Новым ИП — год Эльбы в подарок
Год онлайн-бухгалтерии на тарифе Премиум для ИП младше 3 месяцев
Попробовать бесплатноС кем заключают договоры о полной материальной ответственности
Должны выполняться три условия:
— Человеку есть 18 лет;
— Он непосредственно работает с деньгами и товарами. Непосредственно — значит, берёт в руки деньги, стоит за прилавком с товарами или развозит заказы.
Непосредственно — значит, берёт в руки деньги, стоит за прилавком с товарами или развозит заказы.
— Должность или выполняемая работа есть в перечне в Постановлении Минтруда РФ от 31.12.2002 № 85. Поэтому важно правильно называть должность в штатном расписании и трудовом договоре.
Популярные должности из перечня:
- кассир;
- администратор в торговле, общепите, гостиницах;
- продавец;
- товаровед;
- мастер строительных и монтажных работ;
- заведующий складом;
- кладовщик;
- экспедитор.
Популярные работы:
- приём платежей;
- отпуск товаров и услуг;
- отпуск материальных ценностей со склада;
- доставка грузов.
Если в перечне нет подходящей должности или работы, оформлять полную ответственность нельзя. Например, охранников Минтруд не упоминает. Договор с ними не имеет юридической силы.
Директора и главного бухгалтера нет в перечне, но они тоже несут полную ответственность. Нужно только включить пункт в трудовой договор — ст. 243 ТК РФ.
Нужно только включить пункт в трудовой договор — ст. 243 ТК РФ.
Люди не любят брать на себя лишние риски. Но сотрудники обязаны подписать договор о полной материальной ответственности, если в первую очередь работают с деньгами или товарами, а должность входит в перечень Минтруда. За бунт можно сделать выговор и даже уволить, что разрешил Пленум ВС РФ в п. 36 Постановления от 17.03.2004 г. № 2.
Статья про ответственность работников
Как оформить договор о полной материальной ответственности
Договоры о материальной ответственности бывают индивидуальные и коллективные (бригадные).
Коллективные — когда люди работают вместе и невозможно понять, с кого спрашивать. Например, на складе работает три человека. Предприниматель проводил ежемесячную инвентаризацию и обнаружил, что 5 коробок с туфлями пропали. Каждый сотрудник заплатит треть.
По коллективному договору действует презумпция виновности. По умолчанию работник виноват, но он может доказать обратное — ст. 245 ТК РФ. Например, он не заходил на склад целый месяц, а при прошлой инвентаризации туфли были на месте. Тогда взыскивать деньги не за что.
По умолчанию работник виноват, но он может доказать обратное — ст. 245 ТК РФ. Например, он не заходил на склад целый месяц, а при прошлой инвентаризации туфли были на месте. Тогда взыскивать деньги не за что.
Как ввести коллективную ответственность:
— Издать приказ.
— Попросить сотрудников подписать договор (типовая форма).
— Назначить руководителя «бригады».
Пример: как договоры о материальной ответственности помогли в суде
Предприниматель нанял двух продавцов в магазин одежды. Он оформил трудовые договоры и договор о коллективной материальной ответственности.
Однажды предприниматель заметил, что денег в онлайн-кассе слишком мало. Инвентаризация показала, что не хватает 62 880 ₽. Продавцы должны были выплатить недостачу, но они отказались.
Дальше был долгий спор с заявлением в милицию, судом и обвинением комиссии по инвентаризации в необъективности. Но в итоге суд сказал, что продавцы одежды отвечают за кассу полностью, а с инвентаризацией порядок. Придётся заплатить по 32 483 ₽.
Придётся заплатить по 32 483 ₽.
Дело № 33-4711/2019
Типовая форма договора о полной индивидуальной материальной ответственности
Типовая форма договора о полной коллективной материальной ответственности
Товар и деньги работнику передают по акту или до начала работы проводят инвентаризацию.
Как взыскать ущерб с сотрудника
Главное правило взыскания ущерба с сотрудников — следовать Трудовому кодексу. Правила прописаны в ст. 246, 247 и 248. Если не соблюдать их или вообще не оформлять документы, суд отменит взыскание, а Трудовая инспекция оштрафует по 5.27 КоАП.
Здесь расскажем общий порядок действий, а подробности читайте в статье,
— Соберите независимую комиссию.
— Проведите с ней инвентаризацию и посчитайте убыток.
— Возьмите с сотрудника объяснения.
— Издайте приказ об удержании из зарплаты.
— Если сотрудник согласен платить сразу или больше 20 % от зарплаты, сделайте соглашение о компенсации.
— Если сотрудник отказывается платить, идите в суд.
Увольнение не снимает с сотрудника ответственность. Взыскать ущерб можно, даже если человек уже не работает на вас — ст. 232 ГК РФ. Проблема в том, что работник может скрыться, а потом оспорить сумму в суде. Подстраховаться просто: проводите инвентаризацию перед каждым увольнением.
Ошибки договоров о полной материальной ответственности
По этим договорам уже сложилась большая судебная практика. Мы проанализировали её и расскажем о популярных ошибках работодателей.
1. Работник не участвовал в инвентаризацииНельзя признать сотрудника виновным без объяснения причин. Работодатель должен ознакомить его с заключением комиссии и дать возможность высказаться. Иначе получится необъективно. Суд отменит взыскание, следуя Обзору практики судов от 05.12.2018.
Полагаться на интуицию не стоит. Даже если уверены, что сотрудник должен нести полную материальную ответственность, проверьте перечень Минтруда. Нет подходящего пункта — нет договора.
Даже если уверены, что сотрудник должен нести полную материальную ответственность, проверьте перечень Минтруда. Нет подходящего пункта — нет договора.
Автослесарь поменял масло на автомобиле клиента. Он плохо закрутил сливную пробку и со временем масло вылилось. Автомобиль встал посреди дороги, потому что двигатель заглох. Автосервис признал свою вину и компенсировал расходы водителя на вызов эвакуатора и ремонт. А потом стал разбираться с сотрудником.
Автослесарь подписал договор о полной материальной ответственности. Предприниматель оформил документы и пошёл в суд. Суд сказал: ни должности слесаря, ни работ по ремонту автомобилей в перечне Минтруда нет. Договор незаконный, поэтому сотрудник отвечает в пределах месячного заработка.
Дело № 33-4390/2015.
Если ваш сотрудник работает с дорогими товарами, оформляйте получение разовыми документами — актами и доверенностями. Тогда ответственность тоже будет полной по ст. 243 ТК РФ.
Без договора полная ответственность не работает.
Женщина устроилась работать барменом, а через месяц уволилась. Работодатель заметил недостачу в кассе и обратился в суд. Суд сказал: отпуск товаров и услуг есть в перечне, но договора нет. Инвентаризацию тоже провели неправильно, поэтому даже часть суммы женщина платить не обязана.
Дело № 33-144.
Статья актуальна на
Образец приказа о назначении материально ответственного лица
С точки зрения трудового права материальную ответственность можно представить как обязанность стороны трудового договора возместить причиненный по ее вине ущерб (ст. 232 ТК РФ). Ответственность работника наступает в результате его противоправного поведения. При этом размер компенсации подлежит доказыванию (ст.
 233 ТК РФ).
233 ТК РФ).Правовой основой матответственности работника является ст. 238 ТК РФ, устанавливающая обязанность работника по возмещению реального уменьшения имущества работодателя. При этом в силу прямого указания закона упущенную выгоду с работника взыскать нельзя.
Закон обязывает работодателя участвовать в определении сумм убытков и установлении причин, по которым они возникли. Для этого работодателю законом предоставлено право на проведение проверки и создание специальной комиссии с привлечением экспертов.
Сумма ущерба рассчитывается по фактическим потерям, исчисляемым исходя из рыночных цен (ст. 246 ТК РФ). Материальный ущерб подлежит возмещению вне зависимости от того, был ли специалист привлечен к ответственности иного вида (дисциплинарные взыскания, административные штрафы или уголовные санкции).
В зависимости от размера возмещаемого ущерба матответственность сотрудника бывает полной (ст. 243 ТК РФ) и ограниченной.
Ограничение матответственности подразумевает, что работник возместит ущерб в сумме не более среднего заработка в месяц (ст. 241 ТК РФ). Взыскание работодатель может назначить в течение месяца со дня установления суммы убытков, издав приказ – ст. 248 ТК РФ. Если сотрудник не возместил ущерб в месячный срок или вовсе отказывается от обязательств, взыскать средства можно только через суд – ч. 2 ст. 248 ТК РФ.
241 ТК РФ). Взыскание работодатель может назначить в течение месяца со дня установления суммы убытков, издав приказ – ст. 248 ТК РФ. Если сотрудник не возместил ущерб в месячный срок или вовсе отказывается от обязательств, взыскать средства можно только через суд – ч. 2 ст. 248 ТК РФ.
Полная матответственность работника (ст. 243 ТК РФ) состоит в установленной законом обязанности вернуть работодателю всю сумму убытков в следующих случаях:
— при прямом указании на обязанность такого возмещения в законе;
— выявлена недостача вверенных сотруднику ценностей;
— умышленное причинение убытков, в том числе в состоянии опьянения;
— по приговору суда;
— по вердикту о привлечении к административной ответственности;
— при разглашении тайны.
Возмещение ущерба в размере причиненных убытков может быть предусмотрено трудовым соглашением с заместителем руководителя организации, главным бухгалтером.
При этом работодателю следует помнить, что работники в возрасте до восемнадцати лет могут быть подвергнуты полной материальной ответственности только в случаях, прямо указанных в законе, а именно за ущерб причиненный:
— умышленно,
— в состоянии опьянения (алкогольного или токсического),
— в результате участия в преступлении или правонарушении по КоАП (ст. 242 ТК РФ).
242 ТК РФ).
Перечень должностей работников, с которыми может быть заключен договор о полной материальной ответственности, приведен в Постановлении Минтруда РФ от 31.12.2002 № 85. Например, такой договор может быть заключен:
— с кассиром;
— руководителями и замами организаций, ведущих деятельность в области экспертизы, оборота денежных знаков и ценных бумаг, кассового обслуживания, инкассации и некоторых иных;
— директорами и администраторами торговых предприятий, общепита, гостиниц;
— начальниками и производителями работ на строительных площадках;
— заведующими складов и другими работниками, обслуживающими материальные ценности, в том числе c кладовщиком, продавцом, товароведом и c курьером.
Кроме разделения по объему возмещения разделяют также коллективную (бригадную) материальная ответственность (ст. 245 ТК РФ) и возмещение затрат на обучение специалиста (ст. 249 ТК РФ).
В случае совместного выполнения сотрудниками работ, связанных с оборотом переданных им ценностей, когда невозможно определить меру ответственности каждого специалиста, может устанавливаться бригадная ответственность за убыток, причиненный всем коллективом.
Договор о порядке и содержании бригадной ответственности должен заключаться с каждым членом бригады. Чтобы освободиться от обязанности по возмещению убытка, член бригады должен доказать работодателю, что его вины в возникновении ущерба нет. В случае добровольного возмещения ущерба степень вины члена бригады определяется по соглашению между бригадой и работодателем. Если дело доходит до суда, вину каждого специалиста определяют арбитры. (ст. 245 ТК РФ).
В случае если работник уволится без уважительных причин до истечения срока, обусловленного соглашением об обучении за счет средств работодателя, для сотрудника также могут наступить последствия в виде обязанности возместить затраты работодателю, понесенные на его обучение и исчисленные пропорционально фактически не отработанному после окончания обучения времени (ст. 249 ТК РФ).
Следует указать, что матответственность сотрудника должна исключаться в случаях возникновения форс-мажорных обстоятельств либо, если компания не позаботилась о создании безопасных условий хранения активов, в которых обнаружена недостача (ст. 239 ТК РФ).
239 ТК РФ).
В заключение необходимо сказать, что привлечение сотрудника к ответственности является правом, а не обязанностью работодателя, который вправе с учетом конкретных обстоятельств отказаться от взыскания. Однако стоит учесть, что такое право ограничено правом собственника имущества организации на определение его объема (ст. 240 ТК РФ).
Нужно ли с работником, который заменяет на время отпуска материально ответственное лицо, заключать договор о полной материальной ответственности?
Согласно ч. 1 ст. 244 ТК РФ с работником может быть заключен договор о полной индивидуальной материальной ответственности за недостачу имущества, вверенного работникам, непосредственно обслуживающим или использующим денежные, товарные ценности или иное имущество. Целью заключения такого договора является возмещение работодателю вероятного ущерба в полном размере. Отсутствие данного договора не позволяет работодателю привлечь работника к полной материальной ответственности.
Заключение договора о полной индивидуальной материальной ответственности является правом работодателя, а не обязанностью. Незаключение указанного договора не является нарушением. Однако незаключение таких договоров с работниками означает освобождение последних от обязанности нести полную материальную ответственность.
Незаключение указанного договора не является нарушением. Однако незаключение таких договоров с работниками означает освобождение последних от обязанности нести полную материальную ответственность.
Обратите внимание: работодатель не вправе заключать письменные договоры о материальной ответственности, если должность работника или конкретная поручаемая ему работа не предусмотрена Перечнем, утв. Постановлением Минтруда России от 31.12.2002 N 85 (ст. 244 ТК РФ, Письмо Роструда от 19.10.2006 N 1746-6-1).
Если в отпуск уходит работник на должности (работе), включенной в Перечень, рекомендуем заключить с замещающим его сотрудником договор о полной материальной ответственности, используя типовую форму такого договора. Тогда при необходимости будет возможно привлечь этого работника к полной материальной ответственности (п. 2 ч. 1 ст. 243, ст. 244 ТК РФ).
Одновременно с заключением с замещающим работником договора о материальной ответственности необходимо провести инвентаризацию, так как надо убедиться в сохранности имущества, за которое отвечал отпускник (п. 27 Положения по бухучету N 34н).
27 Положения по бухучету N 34н).
Инвентаризации подлежат только те активы, которые вверены материально ответственному лицу.
Провести инвентаризацию нужно на день приемки-передачи дел между материально ответственными лицами. Например, при уходе материально ответственного лица в отпуск с понедельника – в его последний рабочий день перед отпуском, то есть в пятницу (п. 1.5 Методических указаний по инвентаризации).
На заметку: как провести инвентаризацию при смене материально ответственного лица, читайте в Типовой ситуации в СПС КосультантПлюс.
Полный текст документа смотрите в СПС КонсультантПлюс Ссылки на документы доступны только пользователям КонсультантПлюс — клиентам компании «ЭЛКОД». Дополнительную информацию по приобретению СПС КонсультантПлюс Вы можете получить
ЗДЕСЬ.
Дополнительную информацию по приобретению СПС КонсультантПлюс Вы можете получить
ЗДЕСЬ.
Возможна ли полная материальная ответственность главбуха
Материальная ответственность стороны трудового договора наступает за ущерб, причиненный ею другой стороне этого договора в результате ее виновного противоправного поведения (действий или бездействия), если иное не предусмотрено Трудовым кодексом РФ или иными федеральными законами (ст. 233 ТК РФ).
По общему правилу за причиненный ущерб работник несет материальную ответственность в пределах своего среднего месячного заработка, если иное не предусмотрено Трудовым кодексом РФ или иными федеральными законами (ст. 241 ТК РФ).
Полная материальная ответственность работника состоит в его обязанности возмещать причиненный работодателю прямой действительный ущерб в полном размере. При этом материальная ответственность в полном размере причиненного ущерба может возлагаться на работника лишь в случаях, предусмотренных Трудовым кодексом РФ или иными федеральными законами (ч. 1, 2 ст. 242 ТК РФ).
При этом материальная ответственность в полном размере причиненного ущерба может возлагаться на работника лишь в случаях, предусмотренных Трудовым кодексом РФ или иными федеральными законами (ч. 1, 2 ст. 242 ТК РФ).
Письменные договоры о полной материальной ответственности, то есть о возмещении работодателю причиненного ущерба в полном размере за недостачу вверенного работникам имущества, могут заключаться с работниками, достигшими возраста восемнадцати лет и непосредственно обслуживающими или использующими денежные, товарные ценности или иное имущество (ст. 244 ТК РФ).
Перечни работ и категорий работников, с которыми могут заключаться указанные договоры, а также типовые формы этих договоров утверждаются в порядке, устанавливаемом Правительством РФ.
В соответствии с постановлением Правительства РФ от 14.11.2002 № 823 «О порядке утверждения перечней должностей и работ, замещаемых или выполняемых работниками, с которыми работодатель может заключать письменные договоры о полной индивидуальной или коллективной бригадной) материальной ответственности, а также типовых форм договоров о полной материальной ответственности» Перечень должностей и работ, замещаемых или выполняемых работниками, с которыми работодатель может заключать письменные договоры о полной индивидуальной материальной ответственности за недостачу вверенного имущества, утвержден Постановлением Минтруда России от 31. 12.2002 № 85 (далее — Перечень).
12.2002 № 85 (далее — Перечень).
Должность главного бухгалтера в названном Перечне не указана.
Письменные договоры о полной материальной ответственности могут заключаться только с теми работниками и на выполнение тех видов работ, которые предусмотрены Перечнем.
Названный Перечень является исчерпывающим и расширительному толкованию не подлежит (письмо Роструда от 19.10.2006 № 1746-6-1).
Учитывая изложенное, заключить отдельный договор о полной материальной ответственности с главным бухгалтером нельзя.
Материальная ответственность в полном размере причиненного ущерба возлагается на работника в случаях, когда в соответствии с Трудовым кодексом РФ или иными федеральными законами на работника возложена материальная ответственность в полном размере за ущерб, причиненный работодателю при исполнении работником трудовых обязанностей (п. 1 ч. 1 ст. 243 ТК РФ).
1 ч. 1 ст. 243 ТК РФ).
Трудовым договором, заключаемым с заместителями руководителя организации, главным бухгалтером, может быть установлена материальная ответственность в полном размере причиненного работодателю ущерба (ч. 2 ст. 243 ТК РФ, п. 10 постановления Пленума Верховного Суда РФ от 16.11.2006 № 52).
Таким образом, условие о полной материальной ответственности главного бухгалтера может быть предусмотрено только в трудовом договоре с ним или в дополнительном соглашении к трудовому договору.
В этом случае главный бухгалтер несет полную материальную ответственность за причиненный ущерб имуществу организации с учетом положений статей 233, 239, 243 ТК РФ. При отсутствии в трудовом договоре с главным бухгалтером такого условия (и при отсутствии иных оснований, дающих право на привлечение к такой ответственности) главный бухгалтер несет ответственность за причиненный ущерб в пределах своего среднего месячного заработка (ст. 241 ТК РФ, п. 10 постановления Пленума Верховного Суда РФ от 16.11.2006 № 52).
241 ТК РФ, п. 10 постановления Пленума Верховного Суда РФ от 16.11.2006 № 52).
Заключение договора о полной материальной ответственности
По общему правилу работник несет материальную ответственность перед работодателем в размере, не превышающем его среднемесячный заработок. В случаях, прямо предусмотренных Трудовым кодексом РФ (далее — ТК РФ), на работника может быть возложена полная материальная ответственность. Подробности – в статье юриста «Пепеляев Групп» Екатерины Мартемьяновой.
Если работы, связанные с хранением, обработкой, продажей, перевозкой или иным использованием товарно-материальных ценностей, выполняются совместно несколькими работниками и невозможно разграничить ответственность каждого работника за причинение ущерба, работодатель может вводить коллективную материальную ответственность. В этом случае ценности вверяются заранее установленной группе лиц, на которую возлагается полная материальная ответственность за их недостачу.
В этом случае ценности вверяются заранее установленной группе лиц, на которую возлагается полная материальная ответственность за их недостачу.
Правомерность заключения с работником (коллективом работников) договора о полной материальной ответственности должен доказать работодатель.
Законодательные требований к заключению договора о полной материальной ответственности можно разделить на три группы:
- требования к субъектному составу;
- требования к форме и порядку заключения договора;
- требование к предмету договора.
Субъектный состав
Договоры о полной материальной ответственности могут заключаться с работниками, достигшими возраста восемнадцати лет и непосредственно обслуживающими или использующими денежные, товарные ценности или иное имущество (ст.ст. 243, 244 ТК РФ). Перечни работ и категорий работников, с которыми могут заключаться указанные договоры, а также типовые формы этих договоров утверждаются в порядке, устанавливаемом Правительством РФ (далее – Перечень работ и категорий работников) . Договор о полной материальной ответственности, заключенный с лицом, чья должность и/или выполняемая работа не поименованы в указанном перечне, не имеют юридической силы.
Договор о полной материальной ответственности, заключенный с лицом, чья должность и/или выполняемая работа не поименованы в указанном перечне, не имеют юридической силы.
Так, работодателем был заключен договор о полной материальной ответственности с водителем автомобиля, согласно которому работник принял на себя ответственность «за сохранность автомобилей, обязался обеспечить корректное плавное профессиональное вождение автомобиля, максимально обеспечивающее технически исправленное состояние самого автомобиля, следить за его техническим состоянием, выполнять самостоятельно необходимые работы по обеспечению его безопасной эксплуатации».
Занимаемая работником должность, равно как и выполняемая им работа, в Перечень работ и категорий работников не включены, в связи с чем отсутствуют законные основания для возложения на ответчика полной материальной ответственности. Суд апелляционной инстанции поддержал позицию суда первой инстанции «о взыскании с работника суммы причиненного ущерба в соответствии со ст.
 241 ТК РФ в пределах его среднемесячного заработка».
241 ТК РФ в пределах его среднемесячного заработка».При заключении с работником договора о полной материальной ответственности на него возлагается как обязанность по обеспечению сохранности вверенного имущества, так и обязанность осуществлять конкретные необходимые для него действия (или наоборот воздерживаться от определенного поведения), что должно быть предусмотрено его должностной инструкцией. Работник может быть привлечен к материальной ответственности только в том случае, если будет установлено нарушение возложенных на него должностных обязанностей.
В связи с этим, при переводе работника на новую должность и, соответственно, изменении его должностных обязанностей с ним должен быть заключен новый договор о полной материальной ответственности.
Так, судом было установлено, что договор о полной материальной ответственности был заключен, когда «ответчик занимал должность ведущего инженера по ремонту в бюро вакуумного оборудования [общества].
 Истец же вменяет [работнику] в вину ненадлежащее исполнение должностных обязанностей начальника бюро подготовки производства в отделе по ремонту и наладке оборудования [общества], связанных с сохранностью вверенного ему имущества. Однако в указанной должности договор о полной материальной ответственности с ответчиком не заключался, обязанности по возмещении ущерба [работник] на себя не принимал».
Истец же вменяет [работнику] в вину ненадлежащее исполнение должностных обязанностей начальника бюро подготовки производства в отделе по ремонту и наладке оборудования [общества], связанных с сохранностью вверенного ему имущества. Однако в указанной должности договор о полной материальной ответственности с ответчиком не заключался, обязанности по возмещении ущерба [работник] на себя не принимал».Как следует из приведенных выше положений ТК РФ, договор о полной материальной ответственности, заключенный с лицом, не состоящим с работодателем в трудовых отношениях, является недействительным. Включение такого лица в члены коллектива также приведет к недействительности договора и невозможности привлечения к материальной ответственности всех членов коллектива.
Так, суд апелляционной инстанции признал ошибочным вывод суда первой инстанции о том, что если один из членов коллектива на момент подписания договора не состоял в трудовых отношениях с работодателем, на него не может быть возложена материальная ответственность, но другие работники должны нести ответственность в полном объеме. «Коллективная материальная ответственность наступает в случае, когда все члены бригады принимают на себя такую ответственность, что подтверждается подписанием договора. В данном случае все члены бригады такую ответственность на себя не приняли, соответствующий договор не подписали. Вывод суда о возложении ответственности на некоторых из членов бригады противоречит положениям ст. 245 ТК РФ».
«Коллективная материальная ответственность наступает в случае, когда все члены бригады принимают на себя такую ответственность, что подтверждается подписанием договора. В данном случае все члены бригады такую ответственность на себя не приняли, соответствующий договор не подписали. Вывод суда о возложении ответственности на некоторых из членов бригады противоречит положениям ст. 245 ТК РФ».
Форма и порядок заключения договора
До подписания договора о коллективной ответственности работодатель должен издать приказ о формировании коллектива работников, назначении руководителя коллектива и возложения на них материальной ответственности за сохранность определенного имущества, все члены коллектива должны быть ознакомлены с указанным приказом. Перед заключением договора и передачей имущества коллективу работников работодателем в установленном законом порядке должна быть проведена инвентаризация.
Договор о коллективной материальной ответственности за причинение ущерба заключается в письменной форме. Непосредственно при заключении договора он должен быть подписан всеми членами коллектива, которые принимают на себя ответственность за сохранность вверенного имущества. Подписание договора членами коллектива в разные даты рассматривается судами как несоблюдение порядка заключения договора.
Непосредственно при заключении договора он должен быть подписан всеми членами коллектива, которые принимают на себя ответственность за сохранность вверенного имущества. Подписание договора членами коллектива в разные даты рассматривается судами как несоблюдение порядка заключения договора.
В дальнейшем, договор перезаключается при смене руководителя бригады или при выбытии из коллектива более 50% от его первоначального состава. При выбытии отдельных членов коллектива или приеме в коллектив новых работников договор не перезаключается. В этих случаях против подписи выбывшего члена коллектива указывается дата его выбытия, а вновь принятый работник подписывает договор и указывает дату вступления в коллектив.
Отсутствие единого договора с членами коллектива является обстоятельством, исключающим материальную ответственность всех входивших в состав коллектива работников, поскольку соблюдение правил заключения договора о коллективной материальной ответственности в силу ст. 245 ТК РФ является обязательным. Включение в текст трудового договора условия о возложении на работника полной материальной ответственности является недостаточным.
245 ТК РФ является обязательным. Включение в текст трудового договора условия о возложении на работника полной материальной ответственности является недостаточным.
Ненадлежащее оформление договора является безусловным основанием для отказа в удовлетворении исковых требований о взыскании причиненного ущерба.
Так, апелляционная инстанция поддержала позицию суда первой инстанции о признании договора недопустимым доказательством, указав следующее. «Договоры не прошиты и не скреплены печатью работодателя, а в судебном заседании ответчицы … отрицали факт заключения с ними указанного договора, пояснив, что с текстом договора они ознакомлены не были, подписывали только одну страницу, на которой указаны члены коллектива и их подписи, полагая, что заполняют данные для бухгалтерии для предоставления отчетов в налоговый орган. В ходе судебного разбирательства стороной истца пояснения ответчиц, отрицавших заключение с ними указанного договора, опровергнуты не были».
Если работники отрицают факт ознакомления с текстом договора, оспаривают принадлежность им подписей в договоре или приложениях к нему, обязанность доказать обратное лежит на работодателе.
Предмет договора
Предметом договора о полной коллективной материальной ответственности должно быть именно обеспечение сохранности вверенного имущества.
Между банком и сотрудниками операционного подразделения был заключен договор, в соответствии с которым коллектив принял на себя «материальную ответственность за … неправомерные и мошеннические действия, совершенные сотрудниками подразделения, повлекшие причинение материального ущерба банку, клиентам банка, а также наносящих урон репутации банка при осуществлении операций по открытию/закрытию и ведению банковских счетов, счетов по вкладам, депозитам, по работе с банковскими картами, по выдаче потребительских, ипотечных кредитов, кредитов в рамках программы малого и среднего бизнеса, по осуществлению банковских и системных переводов, по выполнению иных операций, а также за ущерб, возникший у работодателя в результате возмещения им ущерба иным лицам».
Суд пришел к выводу о том, что «указанное условие договора о материальной ответственности, по существу возлагающее на работника (члена коллектива) полную материальную ответственность за совершение противоправных и мошеннических действий другим работником, как противоречащее требованиям трудового законодательства о материальной ответственности и ухудшающее положение работника по сравнению с установленным трудовым законодательством, не подлежит применению (ч. 2 ст. 9 ТК РФ)».
Возложение на работников обязанности по обеспечению сохранности инструментов, оборудования и других средств труда, предоставленных работодателем для надлежащего выполнения работников своих должностных обязанностей, противоречит действующему законодательству.
Суд апелляционный инстанции отменил решение суда первой инстанции, указав следующее. «Договор о материальной ответственности с водителем-экспедитором не может заключаться в отношении транспортного средства, на котором осуществляется перевозка груза, поскольку транспортное средство не является вверенным ему для транспортировки или доставки имуществом предприятия, а представляет собой материально-техническое средство, используемое и необходимое для исполнения трудовой функции водителя.
Таким образом, решение о взыскании … полной стоимости восстановительного ремонта автомобиля принято без учета незаконности заключения договора о полной материальной ответственности водителя за эксплуатируемое транспортное средство. Материальная ответственность ответчика за поврежденный автомобиль могла наступить лишь в ограниченном размере, установленном ст. 241 ТК РФ».
Аналогичным образом, будут нести ограниченную материальную ответственность: кассир – в случае причинения ущерба кассовому аппарату, продавец — в случае причинения ущерба торговому оборудованию.
Действующим трудовым законодательством строго определены категории работниках, на которых договором может быть возложена полная материальная ответственность, форма и порядок заключения договора, а также его предмет. Регулирование носит императивный, обязательный характер и не допускает произвольного толкования. Нарушение работодателем установленных требований приводит к признанию договора полной материальной ответственности недействительным.
Статья 243 ТК РФ. Случаи полной материальной ответственности. Комментарии
Статья 243 ТК описывает ситуации, когда для сотрудника наступает полная материальная ответственность. Этот перечень носит закрытый характер.
Трудовой кодекс Российской Федерации от 30.12.2001 N 197-ФЗ
Характеристика ст. 243 ТК
В ней указано, что полная ответственность за причинённый сотрудником ущерб возлагается на него, если:
- в соответствии с положениями ТК или федеральных законов он полностью отвечает за вред, причинённый им нанимателю при выполнении должностных функций;
- обнаруживается недостача ценностей, доверенных ему в соответствии со специальным договором или одноразовым документом;
- обнаруживается умышленный характер нанесённого им вреда;
- ущерб образуется по причине алкогольного или другого токсического опьянения;
- вред возникает из-за его неправомерных действий, удостоверенных в ходе судебных разбирательств;
- ущерб образуется из-за административного правонарушения, обнаруженного профильной госструктурой;
- им разглашаются данные, составляющие тайну, в ситуациях, предусмотренных ТК или федеральными законами;
- ущерб образуется не из-за исполнения им трудовых функций.


Отдельно указано, что полная ответственность может быть прописана в трудовом договоре, оформленном с заместителями главы организации и главбухом.
Разобранная статья детально освещает параметры полной материальной компенсации, взыскиваемой с сотрудника за причинённый им вред. Стоит отметить, что ТК предусмотрены и иные ситуации, предполагающие подобную компенсацию (статьи 244, 245, 277, 346). Они затрагивают особые обстоятельства полной компенсации материального ущерба.
Основные вопросы по положениям ст. 243 ТК
О каких конфиденциальных данных упоминает комментируемая статья?
Актуальное законодательство предусматривает довольно много разновидностей конфиденциальной информации. И это относится не только к общеизвестным тайным сведениям: военным, коммерческим, врачебным. Есть и более специализированная информация:
- о сиротах и связанных с ними людях (лица, принимающие их в семьи, лишённые родительских прав и пр.) – ФЗ № 44 от 16. 04.01;
- связанная с проведением следственных мероприятий – ст. 161 УПК;
- связанная с мероприятиями по закупкам товаров для государственных и местных органов – ФЗ № 44 05.04.13.
 04.01;
04.01;Всего законодательством предусмотрено около 120 разновидностей конфиденциальных сведений как общего, так и специализированного характера. Разглашение этих данных, даже не связанных непосредственно с экономическими взаимоотношениями, может привести к негативным последствиям в плане нанесения ущерба, предусмотренного гл. 39 ТК.
О каком специальном договоре, предусматривающем полную компенсацию, упоминает данная статья?
Имеются в виду договоры, нюансы которых описаны в последующих статьях ТК: 244 и 245. Подобные договоры могут заключаться как в коллективном, так и индивидуальном формате.
Следует отметить, что указанные договорённости могут заключаться не со всеми сотрудниками. Список тех, для кого она разрешены, утверждается на уровне Правительства. Оно же утверждает и стандартные формы этих договоров.
Важное отличие этих договоров – они предусматривают полную компенсацию лишь в случае утраты ценностей, доверенных сотруднику. При их повреждении ответственность носит частичный характер, описанный в ст. 241.
Каковы нюансы получения компенсации от сотрудника, чьи неправомерные действия привели к ущербу?
Для получения полной компенсации в этой ситуации, как подчёркнуто в статье, необходимо соответствующее судебное решение. Т.е. возбуждение дела против сотрудника, проведение следственных мероприятий и иные подобные действия не подпадают под п. 5 ст. 243 ТК. Для её применения нужно обвинительное решение суда. Естественно, если сотрудник оправдан судом или его дело прекращено на предварительном этапе, это освобождает его от обязанности оплачивать полный ущерб. Однако для нанимателя остаётся открытой возможность добиваться полной компенсации по иным основаниям, предусмотренным ст. 243.
Перечисленные нюансы разъяснены в п. 11 Постановления Пленума ВС № 52 от 16. 11.06.
11.06.
Проблемы и новые рубежи в аналитической характеристике конъюгатов антитело-лекарство
Определение отношения лекарственное средство-антитело
DAR, который представляет собой среднее количество лекарств, конъюгированных с антителом, является важным атрибутом качества, который напрямую связан с эффективностью и безопасность ADC, поскольку низкая нагрузка лекарством (низкий DAR) может снизить эффективность ADC, в то время как высокая нагрузка лекарством (высокая DAR) может изменить фармакокинетику и токсичность молекулы ADC. 34 Следовательно, контроль реакции конъюгации, при которой изменения концентраций реагентов могут приводить к изменению DAR (особенно в случае случайной конъюгации), является наиболее важным шагом в разработке.Для предотвращения появления неконъюгированных антител, а также видов с низким / высоким содержанием DAR, для постоянного достижения желаемой цели DAR должны быть предусмотрены производственные и постпроизводственные средства обработки / обработки. 35 С точки зрения стабильности продукта, степень загрузки лекарственного средства может изменить стабильность и склонность ADC к агрегации. 36 Следовательно, загрузка и распределение лекарств — это CQA, которые следует контролировать при производстве ADC. Для определения DAR использовались различные аналитические методы с использованием спектроскопических, 37 радиологических, 38 хроматографических, 39 , 40 и масс-спектрометрических методов, 41 , 42 .Пригодность этих методов основана на химии конъюгации и физико-химических характеристиках линкера или молекул лекарственного средства. Методы, используемые для определения DAR, включая их сильные стороны и ограничения, обсуждаются в следующих разделах.
35 С точки зрения стабильности продукта, степень загрузки лекарственного средства может изменить стабильность и склонность ADC к агрегации. 36 Следовательно, загрузка и распределение лекарств — это CQA, которые следует контролировать при производстве ADC. Для определения DAR использовались различные аналитические методы с использованием спектроскопических, 37 радиологических, 38 хроматографических, 39 , 40 и масс-спектрометрических методов, 41 , 42 .Пригодность этих методов основана на химии конъюгации и физико-химических характеристиках линкера или молекул лекарственного средства. Методы, используемые для определения DAR, включая их сильные стороны и ограничения, обсуждаются в следующих разделах.
Среди всех этих методов УФ-видимая спектроскопия является относительно простым и часто используемым методом определения DAR. Предпосылками для применения метода являются: 1) препарат должен содержать хромофор УФ-видимого диапазона; 2) лекарственное средство и антитело должны демонстрировать различные и отдельные максимумы поглощения в своих УФ-видимых спектрах; и 3) присутствие лекарственного средства не должно влиять на светопоглощающие свойства фрагмента антитела в образце ADC и наоборот. На основе измеренных коэффициентов поглощения и экстинкции можно рассчитать индивидуальные концентрации белка и лекарственного средства в соответствии с принципом Бера-Ламберта. Среднее значение DAR (количество молей лекарства на моль антитела) можно определить по уравнению 1:
На основе измеренных коэффициентов поглощения и экстинкции можно рассчитать индивидуальные концентрации белка и лекарственного средства в соответствии с принципом Бера-Ламберта. Среднее значение DAR (количество молей лекарства на моль антитела) можно определить по уравнению 1:
Avg DAR = Cdrug CA Antibody
(1)
, где C лекарство и C антитело относится к молярному концентрация лекарства и антитела соответственно. Поскольку для любой данной длины волны полное поглощение является суммой индивидуальных значений поглощения для каждого компонента, значение поглощения антитела и лекарственного средства на их соответствующих длинах волн (λmax) необходимо скорректировать, чтобы ограничить завышение полезной нагрузки.Благодаря своей простоте спектроскопический метод в УФ-видимом диапазоне обычно используется для определения DAR для различных АЦП. Например, трастузумаб эмтанзин (Кадсила®) представляет собой ADC, в котором гуманизированное антитело против HER2 IgG1 трастузумаб (Герцептин®) ковалентно конъюгировано с цитотоксическим агентом DM-1 с использованием 4- (N-малеимидометил) циклогексан-1-карбоксилата ( MCC) в качестве линкерной молекулы. 37 Свободный DM-1 имеет максимум поглощения при 252 нм, что значительно отличается от максимумов поглощения (при 280 нм) основных ароматических хромофоров (остатки триптофана и тирозина) трастузумаба ().Об успешной конъюгации DM-1 с трастузумабом свидетельствует наличие дополнительного пика при 252 нм в конъюгате и дифференциально расположенные полосы поглощения (при 252 нм и 280 нм), которые используются для расчета DAR.
37 Свободный DM-1 имеет максимум поглощения при 252 нм, что значительно отличается от максимумов поглощения (при 280 нм) основных ароматических хромофоров (остатки триптофана и тирозина) трастузумаба ().Об успешной конъюгации DM-1 с трастузумабом свидетельствует наличие дополнительного пика при 252 нм в конъюгате и дифференциально расположенные полосы поглощения (при 252 нм и 280 нм), которые используются для расчета DAR.
УФ-видимые спектры поглощения (A) свободного эмтанзина (DM-1), (B) трастузумаба и (C) конъюгата трастузумаба эмтанзина. Молярные концентрации антитела и лекарственного средства определяются на основе измеренных коэффициентов поглощения и экстинкции (перепечатано с разрешения из ссылки 37).
Другие конъюгированные молекулы лекарственного средства, которые были проанализированы с помощью спектроскопии УФ-видимого диапазона для определения DAR, приведены в. Несмотря на простоту и легкость использования, применение спектроскопического метода в УФ-видимом диапазоне ограничивается активными лекарствами в УФ-видимом диапазоне, которые обладают полосами поглощения, отличными от полос поглощения ароматических остатков (триптофана или тирозина) антитела. Кроме того, присутствие свободного лекарственного средства в образце ADC может потенциально помешать измерению и, следовательно, привести к завышению значения DAR.Кроме того, спектроскопический метод в УФ-видимом диапазоне не дает никакой информации о распределении нагрузки лекарственного средства.
Кроме того, присутствие свободного лекарственного средства в образце ADC может потенциально помешать измерению и, следовательно, привести к завышению значения DAR.Кроме того, спектроскопический метод в УФ-видимом диапазоне не дает никакой информации о распределении нагрузки лекарственного средства.
Таблица 3.
Список ADC с их значением λmax для молекулы антитела и лекарства, используемых для определения DAR.
| Нацеливающий компонент | Терапевтический компонент | ||||
|---|---|---|---|---|---|
| Конъюгат антитело-лекарство | Молекула антитела | λmax (нм) | Молекула лекарственного средства | λmax (нм) | |
| Брентуксимаб ведотин | Anti-CD30 | 280 | Монометил ауристатин E | 248 | 45 |
| Трастузумаб эмтанзин | Трастузумаб | 280 | DM10 | 280 | DM10 |
| MTX-791T / 36 | 791T / 36 | 280 | Метотрексат | 307 | 47 |
| Моноклональные антитела-винка | IgG | 280 | Барвинок Алкалоид | 900 48||
| Анти-B4-DC1 | 900 85 Anti-B4280 | DC1 * | 314 | 49 | |
| Anti-N901-DC1 | Anti-N901 | 280 | DC1 * | 314 | 49 |
| 5E9-адриамицин | 5E9 | 280 | Адриамицин | 495 | 50 |
| BT96- доксорубицин | BT96 | 86 0505 | |||
| CT-M-01-калихеамицин | CT-M-01 | 280 | Аналоги калихеамицина | 333 | 38 |
| MOPC-21-calicheamicin | MOPC-21 | 280 | Аналоги калихеамицина | 333 | 38 |
| Анти-CD79b-MCC-DM1 | Анти-CD79b | 280 | DM1 | 252 | 52 |
| Анти-CD79b-MC-MMAF | Анти-CD79b | 280 | Монометил ауристатин F | 248 | 53 |
Из-за этих ограничений, связанных с методом спектроскопии в УФ-видимом диапазоне, были исследованы другие подходы, такие как мечение молекул лекарства или антител.![]() Например, Cohen et al. использовал подход двойного мечения, в котором и лекарственный фрагмент (варианты тубулизина), и антитело (трастузумаб) были четко конъюгированы с двумя разными радиоактивными метками перед реакцией конъюгации. Просто измеряя радиоактивность, этот подход облегчает мониторинг ADC во время реакции конъюгации и процессов очистки. Кроме того, радиоактивная метка может помочь в изучении in vivo поведения конъюгата ( 131 I-TUB-OMOM- 89 Zr-трастузумаб конъюгат 43 ) для прогнозирования фармакокинетики и таргетного потенциала ADC.
Например, Cohen et al. использовал подход двойного мечения, в котором и лекарственный фрагмент (варианты тубулизина), и антитело (трастузумаб) были четко конъюгированы с двумя разными радиоактивными метками перед реакцией конъюгации. Просто измеряя радиоактивность, этот подход облегчает мониторинг ADC во время реакции конъюгации и процессов очистки. Кроме того, радиоактивная метка может помочь в изучении in vivo поведения конъюгата ( 131 I-TUB-OMOM- 89 Zr-трастузумаб конъюгат 43 ) для прогнозирования фармакокинетики и таргетного потенциала ADC.
В другом исследовании сообщается о новом подходе, основанном на флуоресценции, включающем дериватизацию двух майтанзиноидов, DM1 и DM4, с использованием флуорофора BODIPY FL. 44 Дериватизация включает конъюгацию BODIPY с тиолсодержащим лекарством в стехиометрии 1: 1. Преимущества этого подхода заключаются в том, что его можно использовать для количественного определения аналитов лекарственных средств в микромасштабе (шкала 50 мкг), и, следовательно, его можно использовать на ранней стадии исследования, когда точное взвешивание часто затруднено из-за нехватки материалов. Кроме того, теоретически простое измерение флуоресценции может облегчить его потенциальное использование для количественной оценки DAR для ADC, особенно для соединений, в которых поглощение лекарственного средства и белка перекрывается. Однако применение может быть ограничено из-за проблем, связанных с фотообесцвечиванием и тушением флуоресценции.
Кроме того, теоретически простое измерение флуоресценции может облегчить его потенциальное использование для количественной оценки DAR для ADC, особенно для соединений, в которых поглощение лекарственного средства и белка перекрывается. Однако применение может быть ограничено из-за проблем, связанных с фотообесцвечиванием и тушением флуоресценции.
Некоторые из новых подходов к измерению DAR включают ферментативное отщепление лекарства от антитела и последующую квалификацию с использованием хроматографических методов.Было показано, что скорость и степень расщепления лекарственного средства ферментом катепсина B из ADC на основе vcMMAE, конъюгированных по восстановленным дисульфидным связям, не зависят от местоположения лекарственного средства и носителя моноклонального антитела. 54 На основе аналогичной концепции был описан новый подход с использованием фермента катепсина B для количественной оценки DAR для случайных конъюгированных с лизином ADC. 55 , 56 Этот метод применим как для случайных, так и для сайт-специфичных ADC, конъюгированных с использованием гидролитически стабильного расщепляемого линкера катепсина-B. 55 Он включает серию обработок ферментами и восстановителем для полного высвобождения конъюгированных лекарственных средств с последующим хроматографическим разделением. Во-первых, ADC расщепляются протеазой IdeS в сайте ниже шарнирной области с образованием фрагментов F (ab ‘) 2 и Fc с последующим восстановлением 2-меркаптоэтиламином (2-MEA), чтобы позволить дальнейшую фрагментацию с образованием Fdʹ, Fc и Lc. фрагменты (). Эта предварительная обработка IdeS и 2-MEA обеспечивает доступ к сайтам расщепления и, таким образом, способствует полному высвобождению конъюгированного лекарственного средства.После предварительной обработки образцы обрабатывают ферментом катепсином B для облегчения высвобождения лекарства из фрагментов антител. Высвободившееся лекарство хроматографически отделяют от белковых компонентов с помощью RP-HPLC с количественным определением по УФ-поглощению. Количественная оценка расщепленного лекарственного средства позволяет точно и точно определять общий DAR для ADC на основе расщепляемого линкера катепсина B независимо от химии конъюгации.
55 Он включает серию обработок ферментами и восстановителем для полного высвобождения конъюгированных лекарственных средств с последующим хроматографическим разделением. Во-первых, ADC расщепляются протеазой IdeS в сайте ниже шарнирной области с образованием фрагментов F (ab ‘) 2 и Fc с последующим восстановлением 2-меркаптоэтиламином (2-MEA), чтобы позволить дальнейшую фрагментацию с образованием Fdʹ, Fc и Lc. фрагменты (). Эта предварительная обработка IdeS и 2-MEA обеспечивает доступ к сайтам расщепления и, таким образом, способствует полному высвобождению конъюгированного лекарственного средства.После предварительной обработки образцы обрабатывают ферментом катепсином B для облегчения высвобождения лекарства из фрагментов антител. Высвободившееся лекарство хроматографически отделяют от белковых компонентов с помощью RP-HPLC с количественным определением по УФ-поглощению. Количественная оценка расщепленного лекарственного средства позволяет точно и точно определять общий DAR для ADC на основе расщепляемого линкера катепсина B независимо от химии конъюгации. Этот метод позволяет обойти наиболее распространенные ограничения, связанные с другими методами определения DAR, такими как УФ-метод, требующий различных длин волн поглощения для лекарства и антитела, масс-спектрометрические методы, в которых на массовый сигнал влияет гидрофобная природа и чистый заряд белка, а также гидрофобное взаимодействие. Метод хроматографии (HIC), который часто приводит к низкой разрешающей способности для произвольно сопряженных АЦП.Однако, как и метод УФ-видимого излучения, метод катепсина B не обеспечивает профиля распределения нагрузки лекарственного средства, поскольку метод основан на сумме общего поглощения всех расщепленных молекул лекарственного средства. Кроме того, расщепление белка на более мелкие фрагменты с использованием IdeS и 2-MEA запрещает прямое количественное определение концентрации белка. Метод катепсина B включает в себя множество этапов предварительной обработки перед количественной оценкой лекарственного средства, поэтому перенос метода и внедрение в среду контроля качества может быть сложной задачей.
Этот метод позволяет обойти наиболее распространенные ограничения, связанные с другими методами определения DAR, такими как УФ-метод, требующий различных длин волн поглощения для лекарства и антитела, масс-спектрометрические методы, в которых на массовый сигнал влияет гидрофобная природа и чистый заряд белка, а также гидрофобное взаимодействие. Метод хроматографии (HIC), который часто приводит к низкой разрешающей способности для произвольно сопряженных АЦП.Однако, как и метод УФ-видимого излучения, метод катепсина B не обеспечивает профиля распределения нагрузки лекарственного средства, поскольку метод основан на сумме общего поглощения всех расщепленных молекул лекарственного средства. Кроме того, расщепление белка на более мелкие фрагменты с использованием IdeS и 2-MEA запрещает прямое количественное определение концентрации белка. Метод катепсина B включает в себя множество этапов предварительной обработки перед количественной оценкой лекарственного средства, поэтому перенос метода и внедрение в среду контроля качества может быть сложной задачей.
Графическое представление расщепления IdeS, восстановления 2-MEA и расщепления катепсином B ADC. Перепечатано с разрешения, из ссылки 55.
Масс-спектрометрия (МС) — еще один часто используемый метод для определения характеристик АЦП. 57 Доступные в настоящее время MS-спектрометры, использующие ионизацию электрораспылением (ESI) в сочетании с времяпролетным (TOF) или Orbitrap с расширенным диапазоном масс, могут стандартно дифференцировать гетерогенные молекулярные разновидности ADC с достаточным разрешением.Например, полный профиль массы молекул ADC может быть получен с помощью измерения неповрежденной массы в восстанавливающих и невосстанавливающих условиях. Молекулярные виды, такие как молекулы ADC с различным количеством полезной нагрузки и нулевого линкера (линкер без полезной нагрузки), а также неконъюгированные молекулы антител, могут быть разделены, идентифицированы и количественно определены. В масс-спектре площадь пика или высота пика представляют относительное содержание каждого вида DAR. Следовательно, среднее значение DAR может быть получено из средневзвешенного значения обнаруженных видов с разными номерами DAR.В качестве примеров показан развернутый масс-спектр конъюгированного с цистеином антитела и показан развернутый масс-спектр конъюгата лизина из ЖХ-МС. Масс-спектр молекулы ADC намного сложнее, чем у антитела, из-за дополнительных слоев неоднородности, вносимых конъюгацией. Чтобы получить больше специфической для ADC структурной информации с меньшим вмешательством со стороны свойств антител, полезно обрабатывать молекулы ADC гликозидазой, такой как PNGase F, для удаления гетерогенности, вызванной N-связанными гликанами, чтобы выявить подробные структурные особенности ADC, включая конъюгированные виды и связанные с линкером неоднородности.
Следовательно, среднее значение DAR может быть получено из средневзвешенного значения обнаруженных видов с разными номерами DAR.В качестве примеров показан развернутый масс-спектр конъюгированного с цистеином антитела и показан развернутый масс-спектр конъюгата лизина из ЖХ-МС. Масс-спектр молекулы ADC намного сложнее, чем у антитела, из-за дополнительных слоев неоднородности, вносимых конъюгацией. Чтобы получить больше специфической для ADC структурной информации с меньшим вмешательством со стороны свойств антител, полезно обрабатывать молекулы ADC гликозидазой, такой как PNGase F, для удаления гетерогенности, вызванной N-связанными гликанами, чтобы выявить подробные структурные особенности ADC, включая конъюгированные виды и связанные с линкером неоднородности.
(A) Деконволютированный масс-спектр дегликозилированного конъюгата mcMMAF, ADC-A, и соответствующего исходного материала, Mab-A (ссылка 67) (B) Деконволютированный масс-спектр дегликозилированного конъюгата mc-vc-тубулизин. Методы, основанные на
Методы, основанные на
MS, особенно полезны для характеристики неспецифической конъюгации через Lys, поскольку из примерно 80 остатков Lys на типичном антителе более 20 остатков Lys находятся на поверхности и, таким образом, имеют большую склонность к конъюгации. 58 Методы, основанные на хроматографии, не очень эффективны для разделения случайных конъюгированных ADC с разной нагрузкой лекарственного средства и точного количественного определения DAR, поскольку высокая степень гетерогенности конъюгирования подавляет хроматографическое разделение.На измерение неповрежденной массы, напротив, гораздо меньше влияет неоднородность различных сайтов конъюгации для одних и тех же видов DAR, и он может напрямую разделять и количественно определять молекулы ADC с различными нагрузками лекарственного средства. На практике дегликозилирование обработкой PNGase-F перед анализом неповрежденной массы может значительно упростить масс-спектры. 59 Другим способом упрощения масс-спектров является восстановление молекул ADC до видов тяжелой и легкой цепей или расщепление молекул ADC на его компоненты Fc и Fd путем сочетания ферментативной обработки и восстановления. 60 Этот подход позволяет получать более подробную структурную информацию и обнаруживать низкие уровни вариантов, так что DAR может быть определен с высокой точностью и чувствительностью. Стратегия анализа молекул ADC на уровне фрагментов и последующего объединения информации значительно улучшает массовое разрешение и чувствительность. Еще одно преимущество фрагментного анализа состоит в том, что данные, полученные по фрагментам, могут быть использованы для перекрестного исследования полученных на молекулах в целом для измерения DAR.
60 Этот подход позволяет получать более подробную структурную информацию и обнаруживать низкие уровни вариантов, так что DAR может быть определен с высокой точностью и чувствительностью. Стратегия анализа молекул ADC на уровне фрагментов и последующего объединения информации значительно улучшает массовое разрешение и чувствительность. Еще одно преимущество фрагментного анализа состоит в том, что данные, полученные по фрагментам, могут быть использованы для перекрестного исследования полученных на молекулах в целом для измерения DAR.
Обычно образцы АЦП представлены в виде матриц, несовместимых с масс-спектрометрами. Следовательно, перед анализом МС необходима обработка образца. Если ADC необходимо обогатить из биологических образцов, таких как плазма или сыворотка, иммунозахват является мощным методом. 61 , 62 Образцы ADC могут быть обессолены и разделены с помощью обращенно-фазовой хроматографии или эксклюзионной хроматографии (SEC), 41 или непосредственно введены наноспреем. 63 , 64 В качестве альтернативы, капиллярный электрофорез (КЭ), сопряженный с МС, обеспечивает ортогональный вариант разделения. 65 , 66
63 , 64 В качестве альтернативы, капиллярный электрофорез (КЭ), сопряженный с МС, обеспечивает ортогональный вариант разделения. 65 , 66
Для конъюгированных с цистеином ADC, где межцепочечные дисульфидные связи восстанавливаются и используются в качестве сайтов конъюгации, молекулы антител диссоциируют на легкие и тяжелые цепи в условиях обращенно-фазовой хроматографии, используемых в ЖХ- Анализ МС. Следовательно, для получения полного профиля неповрежденной массы для молекул ADC, конъюгированных с цистеином, следует использовать нативный аэрозольный МС без высоких уровней органического растворителя.В МС с естественным распылением образцы могут быть непосредственно распылены с помощью источника наноразмеров или обессолены и разделены с помощью SEC с помощью MS-совместимых подвижных фаз, таких как ацетат аммония (~ 200 мМ) при нейтральном pH. 67 Пример неповрежденного масс-спектра ADC с межцепочечными дисульфидными цистеинами в качестве сайтов конъюгации показан на. Интересно отметить, что для этого класса ADC преобладающими видами являются молекулы с четным числом конъюгированных лекарств, что указывает на то, что почти все свободные тиолы на остатках цистеина конъюгированы.
Интересно отметить, что для этого класса ADC преобладающими видами являются молекулы с четным числом конъюгированных лекарств, что указывает на то, что почти все свободные тиолы на остатках цистеина конъюгированы.
Интенсивность сигнала MS зависит от эффективности ионизации аналита. В режиме положительных ионов МС-анализа белки обычно ионизируются протонами, предпочтительно прикрепленными к боковым цепям основных аминокислотных остатков. Для конъюгированных с лизином ADC эффективность ионизации снижается из-за присоединенного гидрофобного лекарственного средства, особенно с большим числом загрузки, вероятно, потому, что конъюгация снижает общее количество заряженных остатков лизина, доступных для протонирования. Такая корреляция согласуется с наблюдением, что DAR, измеренный с помощью MS, обычно ниже, чем DAR с помощью УФ-метода.На эффективность ионизации, естественно, влияет степень гидрофобности. Чен и соавторы сообщили, что после ферментативного удаления гидрофобной части лекарственной нагрузки (PABC-MMAE) DAR, измеренный с помощью MS (на основе оставшегося линкера MC-VC), хорошо соответствовал DAR, измеренному с помощью HIC. 64
64
Большинство цитотоксических лекарств, используемых при создании ADC, являются гидрофобными по природе, и поэтому конъюгация этих лекарств с антителом изменяет общие физико-химические свойства, такие как баланс гидрофобности и гидрофильности на поверхности.Изменение гидрофобности молекул можно использовать для определения DAR и распределения нагрузки лекарственного средства. 40 , 68 Одним из таких методов является HIC, который основан на разделении конъюгатов на основе количества прикрепленных к антителу гидрофобных лекарственных средств. Следовательно, виды, нагруженные лекарственным средством, разделяются на основе увеличения гидрофобности, причем наименее гидрофобное неконъюгированное антитело элюируется первым из колонки, а конъюгат с наибольшим DAR элюируется последним.Важно отметить, что разрешение видов, обеспечиваемое столбцом HIC, может быть достаточным для определения различий в полезной нагрузке, и, следовательно, может быть проанализирован уровень конъюгации в ADC. Процент площади индивидуальных пиков представляет собой относительное распределение конъюгатов, нагруженных лекарственным средством. Среднее значение DAR можно рассчитать с помощью следующего уравнения 2:
Процент площади индивидуальных пиков представляет собой относительное распределение конъюгатов, нагруженных лекарственным средством. Среднее значение DAR можно рассчитать с помощью следующего уравнения 2:
A v e r a g e D A R = ∑ ( W e i g h t e d p e a k a r e a ) / 100
(2)
Где взвешенная площадь пика — это индивидуальная процентная площадь пика, умноженная на соответствующую нагрузку лекарственного средства.
HIC — это метод выбора для определения DAR и распределения нагрузки лекарственного средства для ADC, полученных с помощью химии сайт-специфической конъюгации, включая сконструированные остатки цистеина 68 и вставленную неестественную аминокислоту с помощью трансглутаминазы. 25 ADC, полученные с помощью сайт-специфической конъюгации, в которой лекарство прикрепляется к определенным участкам на антителе, демонстрируют относительно низкий уровень гетерогенности по сравнению с ADC, полученными с помощью химии случайной конъюгации, 69 и, следовательно, более поддаются анализу с использованием ИК.Одним из таких примеров является THIOMAB, в котором сайт-специфическая конъюгация монометилауристатина E с сконструированными заменами цистеина в положениях на легкой и тяжелой цепях дает композицию с высокой степенью гомогенности, которую можно легко разделить с помощью HIC-UV. 68 Кроме того, HIC можно также использовать для ADC, синтезированных путем частичного восстановления межцепочечных дисульфидных связей, которые менее гетерогены по сравнению с конъюгацией лизина. Конъюгаты, полученные путем частичного восстановления четырех межцепочечных дисульфидных связей антитела (такие как Mab-vc-MMAE 40 ), имеют переменную стехиометрию, варьирующуюся от нуля до восьми молекул лекарственного средства на одно антитело.
25 ADC, полученные с помощью сайт-специфической конъюгации, в которой лекарство прикрепляется к определенным участкам на антителе, демонстрируют относительно низкий уровень гетерогенности по сравнению с ADC, полученными с помощью химии случайной конъюгации, 69 и, следовательно, более поддаются анализу с использованием ИК.Одним из таких примеров является THIOMAB, в котором сайт-специфическая конъюгация монометилауристатина E с сконструированными заменами цистеина в положениях на легкой и тяжелой цепях дает композицию с высокой степенью гомогенности, которую можно легко разделить с помощью HIC-UV. 68 Кроме того, HIC можно также использовать для ADC, синтезированных путем частичного восстановления межцепочечных дисульфидных связей, которые менее гетерогены по сравнению с конъюгацией лизина. Конъюгаты, полученные путем частичного восстановления четырех межцепочечных дисульфидных связей антитела (такие как Mab-vc-MMAE 40 ), имеют переменную стехиометрию, варьирующуюся от нуля до восьми молекул лекарственного средства на одно антитело. HIC позволяет хроматографически разделить эти нагруженные лекарственным средством конъюгаты, чтобы получить пять преобладающих пиков, которые соответствуют антителам, содержащим ноль, две, четыре, шесть и восемь молекул лекарства. Недавно Zhou et al. сообщил об определении DAR и распределения нагрузки лекарственного средства с помощью HIC-UV для новых сайт-специфичных ADC, созданных с помощью гликоинженерии, то есть путем нацеливания на нативные гликаны на Asn-297 антитела (глико ADC). 70 Сначала две единицы сиаловой кислоты вводятся в нативные гликаны антитела посредством модификации in vitro с последующим мягким окислением новых сиаловых кислот периодатом натрия.Наконец, желаемый ADC получают путем конъюгирования антител, функционализированных альдегидом, с линкером аминоокси-лекарственного средства через стабильную оксимную связь. Глико-ADC, синтезированные из трех разных антител и двух разных типов линкеров, показали хорошее разделение с помощью HIC (
С по ), и полученные значения DAR хорошо коррелировали с результатами ЖХ-МС.
HIC позволяет хроматографически разделить эти нагруженные лекарственным средством конъюгаты, чтобы получить пять преобладающих пиков, которые соответствуют антителам, содержащим ноль, две, четыре, шесть и восемь молекул лекарства. Недавно Zhou et al. сообщил об определении DAR и распределения нагрузки лекарственного средства с помощью HIC-UV для новых сайт-специфичных ADC, созданных с помощью гликоинженерии, то есть путем нацеливания на нативные гликаны на Asn-297 антитела (глико ADC). 70 Сначала две единицы сиаловой кислоты вводятся в нативные гликаны антитела посредством модификации in vitro с последующим мягким окислением новых сиаловых кислот периодатом натрия.Наконец, желаемый ADC получают путем конъюгирования антител, функционализированных альдегидом, с линкером аминоокси-лекарственного средства через стабильную оксимную связь. Глико-ADC, синтезированные из трех разных антител и двух разных типов линкеров, показали хорошее разделение с помощью HIC (
С по ), и полученные значения DAR хорошо коррелировали с результатами ЖХ-МС. Более того, авторы дополнительно демонстрируют, что эти глико-конъюгированные ADC могут генерировать более гомогенный и четко определенный профиль продукта по сравнению с конъюгатами на основе тиол-малеимида ().
Более того, авторы дополнительно демонстрируют, что эти глико-конъюгированные ADC могут генерировать более гомогенный и четко определенный профиль продукта по сравнению с конъюгатами на основе тиол-малеимида ().
Хроматография гидрофобного взаимодействия (HIC) Glyco ADC, полученных с 3 различными сиалированными антителами и 2 различными линкерами аминоокси-лекарственного средства (AO-MMAE и AO-PEG8 Dol10). (A) Трастузумаб, конъюгированный с АО-MMAE. (B) Трастузумаб, конъюгированный с AO-PEG8-Dol10. (C) Антитело B11 против белка активации фибробластов (FAP), конъюгированное с AO-MMAE. (D) Анти-FAP-антитело G11, конъюгированное с AO-MMAE. (E) Трастузумаб, конъюгированный с MC-VC-PABC-MMAE (тиоловый конъюгат). Цифры под каждым пиком указывают количество конъюгированных препаратов.АО-MMAE: аминоокси-Cys-MC-VC-PABC-MMAE; AO-PEG8-Dol10: аминоокси-Cys-MC-VCPABC-PEG8-Dol10. (Печатается с разрешения автора из ссылки 70).
Одной из проблем с методами на основе HIC является неспособность разделить позиционные изомеры видов ADC, содержащих одинаковое количество конъюгированных лекарств, включая лекарства, конъюгированные с тяжелыми и легкими цепями. Разработка метода HIC, способного разделять различные виды DAR для конъюгированных с лизином ADC, является серьезной проблемой. Использование высокой концентрации солей в методе HIC может ограничить его прямую совместимость с анализом МС.
Разработка метода HIC, способного разделять различные виды DAR для конъюгированных с лизином ADC, является серьезной проблемой. Использование высокой концентрации солей в методе HIC может ограничить его прямую совместимость с анализом МС.
Определение распределения лекарственной нагрузки
Как описано в предыдущих разделах, общее распределение лекарственной нагрузки может быть охарактеризовано анализом интактной массы или HIC для менее гетерогенных ADC, генерируемых с помощью цистеина или других сайт-специфических конъюгаций. Чтобы определить распределение лекарственной нагрузки на каждом сайте конъюгации, необходим более полный анализ, такой как картирование пептидов. В зависимости от сложности каждого сценария могут применяться несколько стратегий.Для ADC с сайт-специфической конъюгацией относительно просто определить или подтвердить сайты конъюгации пептидным картированием с обнаружением MS.
Если достигается хорошее хроматографическое разделение, точную информацию о занятости сайта конъюгации можно получить методом ЖХ.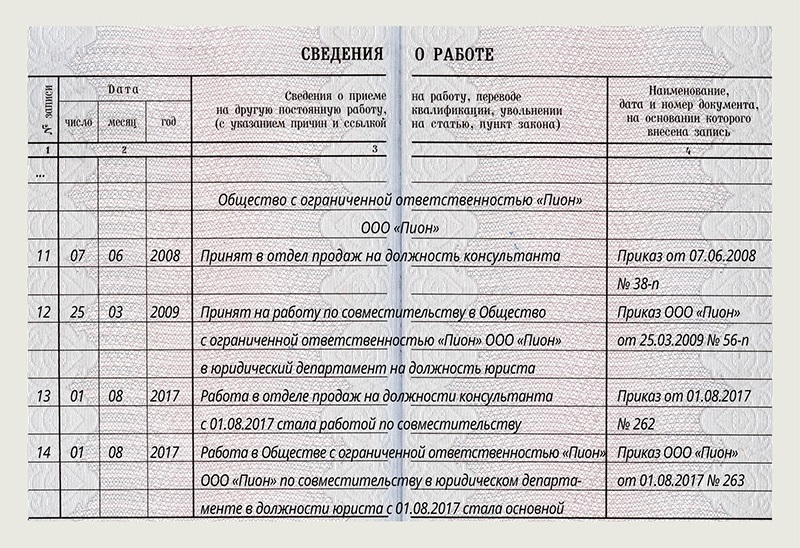 Следует отметить, что конъюгированные соединения могут влиять на эффективность протеолиза из-за стерических затруднений и конформационных изменений. Следовательно, для точного определения важно достичь полного охвата всех родственных видов.Часто необходимо использовать второй фермент в последовательной или параллельной обработке, чтобы захватить все вовлеченные виды. Если удовлетворительное хроматографическое разделение невозможно, можно использовать сигнал MS для достижения относительной количественной оценки конъюгированных и неконъюгированных пептидов с точностью, сильно зависящей от каждого конкретного случая.
Следует отметить, что конъюгированные соединения могут влиять на эффективность протеолиза из-за стерических затруднений и конформационных изменений. Следовательно, для точного определения важно достичь полного охвата всех родственных видов.Часто необходимо использовать второй фермент в последовательной или параллельной обработке, чтобы захватить все вовлеченные виды. Если удовлетворительное хроматографическое разделение невозможно, можно использовать сигнал MS для достижения относительной количественной оценки конъюгированных и неконъюгированных пептидов с точностью, сильно зависящей от каждого конкретного случая.
Для ADC со случайной конъюгацией, если конъюгированное лекарство демонстрирует пик поглощения, не показанный неконъюгированным антителом, карты пептидов ADC и неконъюгированного антитела на этой уникальной длине волны могут использоваться для сравнения и определения местоположения пептидов, конъюгированных с лекарственным средством, как продемонстрировано Ким и соавторами (). 46 Пептиды, конъюгированные с молекулами лекарственного средства, и местоположения сайтов конъюгации затем могут быть определены с помощью анализа ЖХ-МС. Аналогичный подход был использован для характеристики конъюгата лизина (). Если для обнаружения пептидного картирования используется типичная длина волны 214 нм (длина волны поглощения общих пептидных связей), идентификация отдельных сайтов конъюгации и их уровней занятости может оказаться более сложной задачей. Например, как сообщается Wang et al, 71 , параллельное сравнение конъюгированного и неконъюгированного антитела обычно показывает небольшую разницу между двумя картами из-за низкого содержания каждого модифицированного пептида по сравнению с его нативной неконъюгированной формой.Кроме того, многие из незначительных различий, наблюдаемых на карте, являются потенциальными артефактами, возникающими из-за изменений, вызванных методами. В этом случае MS-анализ посредством поиска ожидаемого увеличения массы на конъюгированной карте в корреляции с отсутствием этого увеличения массы на неконъюгированной карте является наиболее эффективным способом идентификации конъюгированных пептидов, который может привести к определению сайтов конъюгации.
46 Пептиды, конъюгированные с молекулами лекарственного средства, и местоположения сайтов конъюгации затем могут быть определены с помощью анализа ЖХ-МС. Аналогичный подход был использован для характеристики конъюгата лизина (). Если для обнаружения пептидного картирования используется типичная длина волны 214 нм (длина волны поглощения общих пептидных связей), идентификация отдельных сайтов конъюгации и их уровней занятости может оказаться более сложной задачей. Например, как сообщается Wang et al, 71 , параллельное сравнение конъюгированного и неконъюгированного антитела обычно показывает небольшую разницу между двумя картами из-за низкого содержания каждого модифицированного пептида по сравнению с его нативной неконъюгированной формой.Кроме того, многие из незначительных различий, наблюдаемых на карте, являются потенциальными артефактами, возникающими из-за изменений, вызванных методами. В этом случае MS-анализ посредством поиска ожидаемого увеличения массы на конъюгированной карте в корреляции с отсутствием этого увеличения массы на неконъюгированной карте является наиболее эффективным способом идентификации конъюгированных пептидов, который может привести к определению сайтов конъюгации. .
.
(A) Наложенные УФ-следы при 252 нм триптических гидролизатов трастузумаба эмтанзина и герцептина, разделенных обращенно-фазовой ВЭЖХ.Аннотирована примерная пара пиков диастериомеров, соответствующих одному и тому же сайту конъюгации. На рисунке представлены только части каждой хроматограммы, показывающие область элюирования пептидов, содержащих лекарственное средство. (ссылка 46). (B) Наложенные УФ-следы при 252 нм конъюгата лизин mc-vc-тубулизин и соответствующие триптические гидролизаты антител, разделенные с помощью обращенно-фазовой UPLC.
Предел обнаружения при таком подходе может регулярно снижаться до менее 1%. Когда для количественной оценки конъюгированных пептидов используется сигнал MS, необходимо учитывать влияние присоединенных соединений на эффективность ионизации из-за потери первичной аминогруппы и введения гидрофобной части для достижения точной количественной оценки.Однако влияние конъюгации на эффективность ионизации, вероятно, будет менее заметным для интактного белка. Кроме того, эндопротеиназы трипсин / Lys-C, которые обычно выбирают для картирования пептидов, не расщепляют модифицированные лизины, что затрудняет измерение занятости сайта.
Кроме того, эндопротеиназы трипсин / Lys-C, которые обычно выбирают для картирования пептидов, не расщепляют модифицированные лизины, что затрудняет измерение занятости сайта.
Еще одна трудность при анализе пептидного картирования ADC — это большая склонность конъюгированных пептидов к осаждению и адсорбции на поверхности флакона, хотя иногда это можно уменьшить, добавив ацетонитрил или изопропаноловый спирт в раствор образца. 72
Отображение пептидов можно использовать для оценки согласованности процесса конъюгации между партиями, поэтому это очень полезный метод для поддержки разработки процесса. демонстрирует занятость сайта случайным лизин-конъюгированным ADC. Интенсивности MS-сигналов конъюгированных пептидов демонстрируют согласованность между четырьмя партиями ADC, произведенными одним и тем же способом, для каждого конъюгированного остатка лизина.
Площади пиков на экстрагированных ионных хроматограммах всех обнаруженных конъюгированных пептидов в триптическом гидролизате случайного конъюгата лизин mc-vc-тубулизин. Четыре партии, произведенные с помощью одного и того же процесса, были проанализированы бок о бок, чтобы продемонстрировать согласованность процесса.
Четыре партии, произведенные с помощью одного и того же процесса, были проанализированы бок о бок, чтобы продемонстрировать согласованность процесса.
ВЭЖХ с обращенной фазой (RPC) — это ортогональный метод, который предоставляет информацию о распределении лекарств по легкой и тяжелой цепям. Если включение линкера или лекарственного средства вызывает заметное увеличение гидрофобности антитела, конгенеры лекарственного средства (различные виды DAR) могут быть разделены градиентом RPC. В зависимости от химического состава и характера конъюгации необработанные образцы можно вводить непосредственно в колонку или образцы можно предварительно обработать восстанавливающим агентом для получения информации о неповрежденном и восстановленном DAR.Например, в случае Cys-связанных ADC, где остатки цистеина из межцепочечных дисульфидных связей конъюгированы с линкером и лекарством, даже небольшое количество органического растворителя оказалось слишком разрушительным и может привести к диссоциации антител. 40 Поэтому образцы обычно обрабатывают восстановителями, такими как дитиотреитол (DTT), перед анализом на RPC. Wiggins и др. . сообщили о характеристике связанных с цистеином конъюгатов IgG1 и IgG2 с лекарственными средствами с использованием RPC-UV, где каждый из конъюгатов восстанавливали с помощью DTT с последующим разделением на обращенную фазу. 73 В другом исследовании Xu et al . разработал метод RPC-UV для DAR-характеристики сайт-специфичных ADC на основе неприродных аминокислот, который был применим как для восстановленных, так и для интактных конъюгатов (). 74
40 Поэтому образцы обычно обрабатывают восстановителями, такими как дитиотреитол (DTT), перед анализом на RPC. Wiggins и др. . сообщили о характеристике связанных с цистеином конъюгатов IgG1 и IgG2 с лекарственными средствами с использованием RPC-UV, где каждый из конъюгатов восстанавливали с помощью DTT с последующим разделением на обращенную фазу. 73 В другом исследовании Xu et al . разработал метод RPC-UV для DAR-характеристики сайт-специфичных ADC на основе неприродных аминокислот, который был применим как для восстановленных, так и для интактных конъюгатов (). 74
(A) DAR для полного конъюгации. Используя метод восстановленной RP-HPLC в качестве инструмента мониторинга, для конъюгации MMAF на HC-F404 использовали диапазон молярных соотношений лекарственное средство-антитело от 1: 4 до 10: 1. Конъюгации проводили при комнатной температуре в течение ночи.(B) Характеристика интактного DAR с помощью RP-HPLC. Хроматограммы ОФ-ВЭЖХ выявили исходное разрешение видов DAR0, DAR1 и DAR2 для неконъюгированных (черный), частично конъюгированных (красный) и полностью конъюгированных (синий) HC-F404-MMAF. (Печатается с разрешения автора из ссылки 74).
(Печатается с разрешения автора из ссылки 74).
Одним из ограничений метода на основе RPC является то, что он основан на различиях гидрофобности между видами с различными уровнями конъюгации; следовательно, он может не подходить для мониторинга определенных конъюгатов лекарственных средств с низкой гидрофобностью.Этот метод применим к конъюгированным с цистеином ADC, но может не подходить для сильно гетерогенных ADC, таких как случайные конъюгаты лизина. Кроме того, использование денатурирующих условий в этом методе может вызвать диссоциацию молекул антител, лишенных межцепочечных дисульфидных связей.
Определение неконъюгированного антитела
В общем, типичный профиль распределения ADC содержит смесь неконъюгированных антител и конгенеров лекарственное средство-антитело. 75 «Голое» антитело конкурирует с ADC за антиген-мишень и в конечном итоге снижает количество лекарства, доставляемого к клетке-мишени.Уровень неконъюгированного антитела в составе ADC является критическим параметром в управлении процессом, поскольку он может напрямую влиять на эффективность ADC. Следовательно, уровень неконъюгированных антител является важным показателем качества, который необходимо контролировать на протяжении всего срока годности продукта. Масс-спектрометрические методы продолжают оставаться универсальными инструментами, которые успешно применяются для количественного определения неконъюгированных антител. MS-анализ антитела huc242, конъюгированного с молекулами майтанзиноидов (DM4), показал типичное распределение, при котором ADC содержит смесь неконъюгированных антител huc242 (D0) и конгенеров антитело-лекарство (от D1 до D7) со средним значением DAR 3.6 (). 41 Процент неконъюгированного антитела рассчитывается на основе площади под кривой для каждого пика и общей площади. В целом, ЖХ-МС — лучший метод для полной характеристики АЦП, хотя основанные на МС методы для АЦП включают в себя дорогостоящее оборудование и очень сложный анализ данных.
Следовательно, уровень неконъюгированных антител является важным показателем качества, который необходимо контролировать на протяжении всего срока годности продукта. Масс-спектрометрические методы продолжают оставаться универсальными инструментами, которые успешно применяются для количественного определения неконъюгированных антител. MS-анализ антитела huc242, конъюгированного с молекулами майтанзиноидов (DM4), показал типичное распределение, при котором ADC содержит смесь неконъюгированных антител huc242 (D0) и конгенеров антитело-лекарство (от D1 до D7) со средним значением DAR 3.6 (). 41 Процент неконъюгированного антитела рассчитывается на основе площади под кривой для каждого пика и общей площади. В целом, ЖХ-МС — лучший метод для полной характеристики АЦП, хотя основанные на МС методы для АЦП включают в себя дорогостоящее оборудование и очень сложный анализ данных.
Масс-спектр дегликозилированного конъюгата huc242-DM4. Метки пиков указывают количество молекул DM4, прикрепленных к антителу: D0 — неконъюгированное антитело, а Dn соответствует молекулам антитела, несущим n молекул DM4 (для n = 1, 2,…, 8) (перепечатано с разрешения, из ссылки 41 ).
Поскольку конъюгация молекул лекарства с антителом может значительно изменить поверхность и общий электростатический профиль, методы разделения на основе заряда могут быть полезны для улучшенной характеристики, включая обнаружение неконъюгированного антитела. Изображение капиллярной изоэлектрической фокусировки (iCIEF) привлекло значительное внимание в последние годы как аналитический метод для анализа вариантов заряда АЦП. В отличие от традиционных методов разделения, таких как ионообменная хроматография (IEX) и изоэлектрическое фокусирование на основе пластин-гелей (IEF), iCIEF более точно разделяет варианты заряда на основе их изоэлектрической точки (pI).Если антитело и ADC имеют разные значения pI, антитело можно отделить от различных видов ADC и количественно оценить с помощью iCIEF. Например, Lin et al. успешно продемонстрировал разделение неконъюгированного антитела и конъюгированного с лекарственным средством антитела на основе их различных значений pI с использованием iCIEF. 75 Виды, нагруженные лекарственным средством (конъюгированные антитела), дополнительно разделяются на основании снижения поверхностного заряда из-за конъюгации с Lys. С увеличением количества лекарств, конъюгированных с антителом, pI видов, нагруженных лекарством, смещается в кислую сторону по сравнению с неконъюгированным антителом (как показано на рисунке).Каждый отдельный пик представляет собой конъюгат с определенной полезной нагрузкой. Концентрация неконъюгированного антитела определяется по калибровочной кривой. Кроме того, iCIEF может также служить в качестве быстрого и удобного метода снятия отпечатков пальцев для АЦП для отслеживания любых изменений, связанных с конъюгацией или процессом. показывает согласованность 4 различных партий ADC, как определено iCIEF. Однако характеристика распределения нагрузки лекарственного средства с помощью iCIEF может быть очень сложной задачей из-за того, что конъюгация линкера без лекарственного средства также может сдвигать профиль заряда в сторону кислой стороны и, таким образом, переоценивать распределение.
75 Виды, нагруженные лекарственным средством (конъюгированные антитела), дополнительно разделяются на основании снижения поверхностного заряда из-за конъюгации с Lys. С увеличением количества лекарств, конъюгированных с антителом, pI видов, нагруженных лекарством, смещается в кислую сторону по сравнению с неконъюгированным антителом (как показано на рисунке).Каждый отдельный пик представляет собой конъюгат с определенной полезной нагрузкой. Концентрация неконъюгированного антитела определяется по калибровочной кривой. Кроме того, iCIEF может также служить в качестве быстрого и удобного метода снятия отпечатков пальцев для АЦП для отслеживания любых изменений, связанных с конъюгацией или процессом. показывает согласованность 4 различных партий ADC, как определено iCIEF. Однако характеристика распределения нагрузки лекарственного средства с помощью iCIEF может быть очень сложной задачей из-за того, что конъюгация линкера без лекарственного средства также может сдвигать профиль заряда в сторону кислой стороны и, таким образом, переоценивать распределение. Таким образом, iCIEF можно применять для измерения уровня неконъюгированного антитела и распределения нагрузки лекарственного средства, но он не может различать конъюгаты, промежуточные продукты процесса и примеси, такие как антитело только с линкером и антитело с конъюгатом линкер / лекарственное средство. Кроме того, анализ iCIEF может быть неприменим к другим химическим соединениям конъюгации, таким как цистеин, гликаны или сайт-специфичные химические соединения, которые не приводят к значительным изменениям заряда и чистого pI.
Таким образом, iCIEF можно применять для измерения уровня неконъюгированного антитела и распределения нагрузки лекарственного средства, но он не может различать конъюгаты, промежуточные продукты процесса и примеси, такие как антитело только с линкером и антитело с конъюгатом линкер / лекарственное средство. Кроме того, анализ iCIEF может быть неприменим к другим химическим соединениям конъюгации, таким как цистеин, гликаны или сайт-специфичные химические соединения, которые не приводят к значительным изменениям заряда и чистого pI.
(A) Электрофореграмма модельного АЦП на основе лизина.Количество неконъюгированного антитела определяли с помощью калибровочной кривой. (B) ICIEF как метод отпечатка пальца для контроля согласованности процесса от партии к партии.
Подобно изменениям поверхностного заряда, конъюгация (гидрофобного) лекарственного средства также изменяет гидрофобность белковых молекул из-за присущей ему липофильной природы лекарственного средства или линкера. Такие изменения поверхностной гидрофобности можно использовать для определения уровня неконъюгированного антитела в образцах ADC с помощью методов хроматографии на основе гидрофобности, таких как HIC, когда сначала элюируется наименее гидрофобное неконъюгированное антитело, а затем молекулы ADC с увеличивающейся поверхностной гидрофобностью.Взаимодействия между гидрофобными пятнами на поверхности молекулы ADC и неподвижной фазой HIC, как правило, обратимы по природе и в значительной степени зависят от присутствия соли в подвижной фазе. Высокая концентрация соли усиливает гидрофобные взаимодействия, тогда как низкая концентрация соли их ослабляет. Следовательно, удерживание и элюирование молекул ADC из колонки HIC можно регулировать содержанием соли.
Такие изменения поверхностной гидрофобности можно использовать для определения уровня неконъюгированного антитела в образцах ADC с помощью методов хроматографии на основе гидрофобности, таких как HIC, когда сначала элюируется наименее гидрофобное неконъюгированное антитело, а затем молекулы ADC с увеличивающейся поверхностной гидрофобностью.Взаимодействия между гидрофобными пятнами на поверхности молекулы ADC и неподвижной фазой HIC, как правило, обратимы по природе и в значительной степени зависят от присутствия соли в подвижной фазе. Высокая концентрация соли усиливает гидрофобные взаимодействия, тогда как низкая концентрация соли их ослабляет. Следовательно, удерживание и элюирование молекул ADC из колонки HIC можно регулировать содержанием соли.
Гидрофобность лекарственного средства или линкера и гетерогенность ADC оказывают значительное влияние на отделение неконъюгированных антител от видов, конъюгированных с лекарственным средством.Например, виды с DAR, отличными от сайт-специфичных или основанных на цистеине ADC, обычно хорошо разделяются в HIC. Напротив, виды ADC с разными DAR от случайной конъюгации по остаткам лизина редко хорошо разделяются в HIC из-за высокой степени молекулярной гетерогенности. Однако по-прежнему возможно хотя бы частично отделить неконъюгированные антитела от конъюгированных. показывает профили HIC-UV двух партий ADC на основе лизина на колонке MAbPac HIC-20 с использованием подвижной фазы фосфат натрия / сульфат аммония.Несмотря на сложный профиль HIC-UV из-за случайной конъюгации, неконъюгированное антитело разделяется достаточно хорошо, чтобы отличить его от видов, конъюгированных с лекарственным средством. Уровень неконъюгированного антитела можно рассчитать с помощью уравнения 3:
Напротив, виды ADC с разными DAR от случайной конъюгации по остаткам лизина редко хорошо разделяются в HIC из-за высокой степени молекулярной гетерогенности. Однако по-прежнему возможно хотя бы частично отделить неконъюгированные антитела от конъюгированных. показывает профили HIC-UV двух партий ADC на основе лизина на колонке MAbPac HIC-20 с использованием подвижной фазы фосфат натрия / сульфат аммония.Несмотря на сложный профиль HIC-UV из-за случайной конъюгации, неконъюгированное антитело разделяется достаточно хорошо, чтобы отличить его от видов, конъюгированных с лекарственным средством. Уровень неконъюгированного антитела можно рассчитать с помощью уравнения 3:
% неконъюгированного Mab в ADC = неконъюгированное Mab (мг / мл) Общая концентрация белка (мг / мл) × 100
(3)
Где определяется количество неконъюгированного антитела. на основе стандартной кривой для свободных антител при различных концентрациях.
HIC-профиль лизин-конъюгированных ADC, партия 1 (черный) и партия 2 (синий), дающая один заметный пик, соответствующий неконъюгированному антителу (красный), и несколько неразрешенных пиков для видов антител, конъюгированных с лекарственным средством. Расчетный уровень неконъюгированных антител составлял около 4%.
Расчетный уровень неконъюгированных антител составлял около 4%.
Антиоксиданты | Бесплатный полнотекстовый | Функции АФК в макрофагах и противомикробный иммунитет
4.1.1. АФК против бактерий
Распознавание бактерий макрофагами приводит к продукции АФК в разных клеточных компартментах, где они выполняют разные антибактериальные функции. Одной из первых функциональных ролей, описанных для АФК, продуцируемых макрофагами, была инактивация фагоцитированных бактерий за счет окислительного взрыва, создаваемого Nox2.Распознавание вторгшихся бактерий вызывает быстрое и устойчивое производство АФК во внеклеточное пространство и просвет фагосомы (внеклеточные АФК) (см. Раздел 5.2) [30,58,233,234,235]. Это полностью устранено в перитонеальных макрофагах (PM) с дефицитом Nox2 и макрофагах, происходящих из костного мозга (BMDM) [30,58,232], благодаря чему Nox2 является исключительным источником внеклеточных ROS, продуцируемых макрофагами в ответ на бактериальную инфекцию. Лица с хронической гранулематозной болезнью (ХГБ), генетическим заболеванием, вызванным гипо- или аморфными мутациями в генах, кодирующих Nox2 или его субъединицы, не могут продуцировать достаточное количество АФК с помощью Nox2 и, как следствие, страдают от повышенной восприимчивости к инфекциям [236]. важность АФК, производных от Nox2, для противомикробного иммунитета.В зависимости от инвазивного патогена также могут быть активированы другие изоформы Nox или митохондрии для производства ROS либо непосредственно в фагосому для инактивации фагоцитированных бактерий, либо ROS продуцируются в цитозоле для борьбы с бактериями, которые уже покинули фагосому. Listeria monocytogenes (Lm ), патоген пищевого происхождения, специализируется на побеге из фагосомы через порообразующий токсин листериолизин O и две фосфолипазы PlcA и PlcB [237 238 239]. При заражении L.m.тканевые макрофаги активируют высоко антимикробный фагоцитарный путь, связанный с LC3 фагоцитоз (LAP), во время которого фагосомы украшаются LC3 [240, 241, 242].
Лица с хронической гранулематозной болезнью (ХГБ), генетическим заболеванием, вызванным гипо- или аморфными мутациями в генах, кодирующих Nox2 или его субъединицы, не могут продуцировать достаточное количество АФК с помощью Nox2 и, как следствие, страдают от повышенной восприимчивости к инфекциям [236]. важность АФК, производных от Nox2, для противомикробного иммунитета.В зависимости от инвазивного патогена также могут быть активированы другие изоформы Nox или митохондрии для производства ROS либо непосредственно в фагосому для инактивации фагоцитированных бактерий, либо ROS продуцируются в цитозоле для борьбы с бактериями, которые уже покинули фагосому. Listeria monocytogenes (Lm ), патоген пищевого происхождения, специализируется на побеге из фагосомы через порообразующий токсин листериолизин O и две фосфолипазы PlcA и PlcB [237 238 239]. При заражении L.m.тканевые макрофаги активируют высоко антимикробный фагоцитарный путь, связанный с LC3 фагоцитоз (LAP), во время которого фагосомы украшаются LC3 [240, 241, 242]. Эти так называемые LAPosomes демонстрируют улучшенное слияние с лизосомами, что приводит к улучшенному уничтожению L.m. и, как следствие, существенно улучшают иммунитет к L.m. in vitro и in vivo (см. рисунок 5) [58 243]. Активация LAP строго требует продукции внеклеточных ROS, происходящих из Nox2, но, в частности, производные от Nox2 ROS, по-видимому, сами по себе не обладают листерицидным действием [58].Продукция АФК Nox2 при листериальной инфекции регулируется на разных уровнях. Опосредованное рецептором распознавание вторгшейся бактерии часто является первым шагом в каскаде, который запускает антибактериальную способность макрофагов. Для L.m. интегрин Mac-1 является критическим для инициации продукции ROS с помощью Nox2, тогда как передача сигналов, опосредованная Toll-подобным рецептором (TLR), совершенно не обязательна [58]. Было показано, что на цитозольном уровне деубиквитиназы (DUB) отрицательно влияют на продукцию АФК, опосредованную Nox2, во время L.м. инфекция в иммортализованной линии макрофагоподобных клеток RAW246.
Эти так называемые LAPosomes демонстрируют улучшенное слияние с лизосомами, что приводит к улучшенному уничтожению L.m. и, как следствие, существенно улучшают иммунитет к L.m. in vitro и in vivo (см. рисунок 5) [58 243]. Активация LAP строго требует продукции внеклеточных ROS, происходящих из Nox2, но, в частности, производные от Nox2 ROS, по-видимому, сами по себе не обладают листерицидным действием [58].Продукция АФК Nox2 при листериальной инфекции регулируется на разных уровнях. Опосредованное рецептором распознавание вторгшейся бактерии часто является первым шагом в каскаде, который запускает антибактериальную способность макрофагов. Для L.m. интегрин Mac-1 является критическим для инициации продукции ROS с помощью Nox2, тогда как передача сигналов, опосредованная Toll-подобным рецептором (TLR), совершенно не обязательна [58]. Было показано, что на цитозольном уровне деубиквитиназы (DUB) отрицательно влияют на продукцию АФК, опосредованную Nox2, во время L.м. инфекция в иммортализованной линии макрофагоподобных клеток RAW246. 7 (RAW) [244]. Химическое ингибирование DUB в клетках RAW, инфицированных L.m. привело к увеличению общих клеточных уровней АФК (см. раздел 5.1) и снижению бактериальной нагрузки. Клетки RAW с дефицитом Nox2 не показали индукции общей клеточной продукции ROS после ингибирования DUB и отсутствия снижения бактериальной нагрузки, что указывает на то, что DUB ограничивают зависимую от Nox2 продукцию ROS и бактериальный клиренс. Негативный регулятор ROS (NRROS), регуляторный фактор, непосредственно взаимодействующий с Nox2, отрицательно влияет на продукцию ROS через Nox2, конкурируя с p22 phox в связывании gp91 phox [245].Праймирование с помощью LPS, IFNγ или фактора некроза опухоли (TNF) приводило к подавлению NRROS и, следовательно, к увеличению продукции общих клеточных ROS в BMDM, совпадающих с убитыми нагреванием L.m. Соответственно, BMDM с дефицитом NRROS показал повышенные уровни общих клеточных ROS после совпадения с убитыми нагреванием L.m. Не только Nox2, но и митохондрии могут быть задействованы в качестве источника фагосомных АФК во время бактериальной инфекции [61].
7 (RAW) [244]. Химическое ингибирование DUB в клетках RAW, инфицированных L.m. привело к увеличению общих клеточных уровней АФК (см. раздел 5.1) и снижению бактериальной нагрузки. Клетки RAW с дефицитом Nox2 не показали индукции общей клеточной продукции ROS после ингибирования DUB и отсутствия снижения бактериальной нагрузки, что указывает на то, что DUB ограничивают зависимую от Nox2 продукцию ROS и бактериальный клиренс. Негативный регулятор ROS (NRROS), регуляторный фактор, непосредственно взаимодействующий с Nox2, отрицательно влияет на продукцию ROS через Nox2, конкурируя с p22 phox в связывании gp91 phox [245].Праймирование с помощью LPS, IFNγ или фактора некроза опухоли (TNF) приводило к подавлению NRROS и, следовательно, к увеличению продукции общих клеточных ROS в BMDM, совпадающих с убитыми нагреванием L.m. Соответственно, BMDM с дефицитом NRROS показал повышенные уровни общих клеточных ROS после совпадения с убитыми нагреванием L.m. Не только Nox2, но и митохондрии могут быть задействованы в качестве источника фагосомных АФК во время бактериальной инфекции [61]. Гэн и его коллеги показали, что это также относится к L.m. инфекция [246].Исследование выявило две киназы, расположенные в фагосомах, стерильные 20-подобные киназы 1/2 млекопитающих (MST1 / MST2), которые регулируют рекрутирование митохондрий в фагосомы, содержащие L.m. BMDM с дефицитом MST1 / 2 показал снижение продукции ROS в фагосомы и увеличение бактериальной нагрузки. Escherichia coli (E.c.), связанный с зондом для АФК CellROX (см. Раздел 5.1), использовали для измерения вклада митохондрий в уровни фагосомных АФК. Примечательно, что MST1 / 2 напрямую не регулирует продукцию mtROS, поскольку уровни mtROS в матрице остаются неизменными, как было определено с помощью MitoSOX (см. Раздел 5.2). Эти исследования показывают, что продукция ROS либо из Nox2, либо из митохондрий играет решающую роль в антистериальной защите (Рисунок 5). В отличие от очевидной роли NOx2-dervied ROS в антистериальной активности макрофагов, роль Nox2 во время инфицирования другим грампластом. положительный возбудитель, золотистый стафилококк (Sa), является спорным.
Гэн и его коллеги показали, что это также относится к L.m. инфекция [246].Исследование выявило две киназы, расположенные в фагосомах, стерильные 20-подобные киназы 1/2 млекопитающих (MST1 / MST2), которые регулируют рекрутирование митохондрий в фагосомы, содержащие L.m. BMDM с дефицитом MST1 / 2 показал снижение продукции ROS в фагосомы и увеличение бактериальной нагрузки. Escherichia coli (E.c.), связанный с зондом для АФК CellROX (см. Раздел 5.1), использовали для измерения вклада митохондрий в уровни фагосомных АФК. Примечательно, что MST1 / 2 напрямую не регулирует продукцию mtROS, поскольку уровни mtROS в матрице остаются неизменными, как было определено с помощью MitoSOX (см. Раздел 5.2). Эти исследования показывают, что продукция ROS либо из Nox2, либо из митохондрий играет решающую роль в антистериальной защите (Рисунок 5). В отличие от очевидной роли NOx2-dervied ROS в антистериальной активности макрофагов, роль Nox2 во время инфицирования другим грампластом. положительный возбудитель, золотистый стафилококк (Sa), является спорным. Этот патоген устойчив к антибактериальным механизмам фагоцитов и может использовать их в качестве репликативной ниши [247]. Роль Nox2 in vivo при заражении S.а. был предложен в модели легочной инфекции [248]. В то время как мыши с дефицитом Nox2 показали повышенную бактериальную нагрузку, BMDM с дефицитом Nox2 in vitro не различалась по их бактериальной нагрузке. Это неудивительно, потому что BMDM экспрессирует намного меньше Nox2 и демонстрирует сильно сниженную внеклеточную продукцию ROS по сравнению с тканевыми макрофагами, такими как PM [58]. Кроме того, измерения АФК не проводились, поэтому остается выяснить, является ли антибактериальный эффект производных от NOx2 АФК, наблюдаемый in vivo, и как он связан с продуцированием АФК макрофагами.В соответствии с этим, другое исследование также показало, что BMDM с дефицитом Nox2 не отличался по бактериальной нагрузке от BMDM дикого типа (WT) после S.a. инфекция [249]. Однако лечение BMDM с дефицитом WT или Nox2 глобальным поглотителем ROS N-ацетил-цистеином (NAC) или ингибитором флавопротеинов DPI (см.
Этот патоген устойчив к антибактериальным механизмам фагоцитов и может использовать их в качестве репликативной ниши [247]. Роль Nox2 in vivo при заражении S.а. был предложен в модели легочной инфекции [248]. В то время как мыши с дефицитом Nox2 показали повышенную бактериальную нагрузку, BMDM с дефицитом Nox2 in vitro не различалась по их бактериальной нагрузке. Это неудивительно, потому что BMDM экспрессирует намного меньше Nox2 и демонстрирует сильно сниженную внеклеточную продукцию ROS по сравнению с тканевыми макрофагами, такими как PM [58]. Кроме того, измерения АФК не проводились, поэтому остается выяснить, является ли антибактериальный эффект производных от NOx2 АФК, наблюдаемый in vivo, и как он связан с продуцированием АФК макрофагами.В соответствии с этим, другое исследование также показало, что BMDM с дефицитом Nox2 не отличался по бактериальной нагрузке от BMDM дикого типа (WT) после S.a. инфекция [249]. Однако лечение BMDM с дефицитом WT или Nox2 глобальным поглотителем ROS N-ацетил-цистеином (NAC) или ингибитором флавопротеинов DPI (см. Раздел 6 и Раздел 7) привело к снижению общих клеточных уровней ROS, снижению бактериальной нагрузки и снижению индукции. инозитол-требующего фермента 1α (IRE1), который является датчиком стресса эндоплазматического ретикулума.Поскольку IRE1-дефицитный BMDM показал снижение бактериальной нагрузки и общих клеточных уровней ROS, была предложена антибактериальная роль IRE1, опосредованная ROS. Источники этого всплеска ROS не анализировались, но поскольку DPI ингибирует не только ферменты Nox, но также влияет на ETC [63,250,251] (см. Раздел 7) и, следовательно, на продукцию mtROS, могут быть задействованы как другие изоформы Nox, так и митохондрии. В продолжающемся исследовании той же группы была выявлена комплексная антибактериальная роль митохондрий во время инфицирования S.а. [252]. В то время как другие исследования [61,246,253] продемонстрировали, что митохондрии могут быть задействованы в бактериальных фагосомах и увеличивают продукцию mtROS в антибактериальном матриксе, исследование Abuaita et al.
Раздел 6 и Раздел 7) привело к снижению общих клеточных уровней ROS, снижению бактериальной нагрузки и снижению индукции. инозитол-требующего фермента 1α (IRE1), который является датчиком стресса эндоплазматического ретикулума.Поскольку IRE1-дефицитный BMDM показал снижение бактериальной нагрузки и общих клеточных уровней ROS, была предложена антибактериальная роль IRE1, опосредованная ROS. Источники этого всплеска ROS не анализировались, но поскольку DPI ингибирует не только ферменты Nox, но также влияет на ETC [63,250,251] (см. Раздел 7) и, следовательно, на продукцию mtROS, могут быть задействованы как другие изоформы Nox, так и митохондрии. В продолжающемся исследовании той же группы была выявлена комплексная антибактериальная роль митохондрий во время инфицирования S.а. [252]. В то время как другие исследования [61,246,253] продемонстрировали, что митохондрии могут быть задействованы в бактериальных фагосомах и увеличивают продукцию mtROS в антибактериальном матриксе, исследование Abuaita et al. предположили гораздо более сложное вовлечение митохондрий во время S.a. инфекционное заболевание. Зараженные S.a. клетки RAW показали повышенную продукцию mtROS в матриксе, и обработка Necrox-5, редко используемым поглотителем ROS митохондриального матрикса, не только снижала уровни mtROS в матриксе, но также увеличивала бактериальную нагрузку, что указывает на роль матричных mtROS в защите от S.а. TLR-опосредованная передача сигналов может индуцировать продукцию mtROS [30,61], но в отличие от TLR2 / 4/9-дефицитных RAW-клеток, инфицированных S.a. показали неизменную продукцию mtROS в матриксе, в то время как бактериальная нагрузка была снижена. Abuaita et al. предположили, что передача сигналов TLR, вместо запуска продукции mtROS, индуцирует доставку происходящих из митохондрий везикул, содержащих белок митохондриального матрикса SOD2, к S.a.-содержащим фагосомам. Затем в просвете фагосомы доставленный SOD2 усиливает преобразование O 2 ● — в H 2 O 2 .
предположили гораздо более сложное вовлечение митохондрий во время S.a. инфекционное заболевание. Зараженные S.a. клетки RAW показали повышенную продукцию mtROS в матриксе, и обработка Necrox-5, редко используемым поглотителем ROS митохондриального матрикса, не только снижала уровни mtROS в матриксе, но также увеличивала бактериальную нагрузку, что указывает на роль матричных mtROS в защите от S.а. TLR-опосредованная передача сигналов может индуцировать продукцию mtROS [30,61], но в отличие от TLR2 / 4/9-дефицитных RAW-клеток, инфицированных S.a. показали неизменную продукцию mtROS в матриксе, в то время как бактериальная нагрузка была снижена. Abuaita et al. предположили, что передача сигналов TLR, вместо запуска продукции mtROS, индуцирует доставку происходящих из митохондрий везикул, содержащих белок митохондриального матрикса SOD2, к S.a.-содержащим фагосомам. Затем в просвете фагосомы доставленный SOD2 усиливает преобразование O 2 ● — в H 2 O 2 . К сожалению, уровни АФК в Sa-содержащих фагосомах не были определены, например, с бактериями, связанными с зондом АФК [246] или с непроницаемыми для клеток внеклеточными зондами АФК (см. Раздел 5.2) [30,58,99,254], и поэтому явные доказательства отсутствует тот факт, что продукция фагосомных АФК действительно была увеличена за счет SOD2, доставленного в фагосомы через везикулы, происходящие из митохондрий. Эксперименты с SOD2-дефицитными макрофагами также могли усилить это предположение. Остается исследовать, индуцируется ли этот очень сложный путь усиления продукции антибактериальных АФК только S.а. или также другими патогенами, и влияет ли это на антибактериальный иммунитет in vivo. По крайней мере, для L.m., внеклеточная продукция ROS полностью независима от передачи сигналов TLR [58], исключая роль TLR-опосредованной доставки SOD2 в L.m.-содержащие фагосомы. Явная роль либо производных Nox2 ROS, либо mtROS в прямой антибактериальной защите макрофагов против S.a. поэтому остается не полностью охарактеризованным. Для прояснения этой темы потребуются будущие исследования с хорошо зарекомендовавшими себя измерениями ROS и твердыми генетическими данными.Как и S.a., Mycobacterium tuberculosis (Mtb) использует фагосомы макрофагов в качестве пролиферативной ниши. Механизмы выживания в фагосомах — это ингибирование слияния с лизосомами и избегание ксенофагии [255]. Благодаря своему фактору вирулентности CpsA, Mtb также активно ингибирует рекрутирование Nox2 в фагосомы. Предотвращая продукцию ROS, производных от Nox2, Mtb эффективно избегает нацеливания и уничтожения LAP [241,256]. Поскольку продукция АФК, происходящая из Nox2, эффективно подавляется и Mtb ускользает в цитозоль, макрофаги должны активировать другие цитозольные источники АФК, чтобы инактивировать этот патоген.Антибактериальная роль цитозольного mtROS против Mtb была исследована в двух исследованиях in vivo на рыбках данио [129, 257]. Одной инфекции Mtb было недостаточно для увеличения общих клеточных уровней ROS или для индукции матричной продукции mtROS в макрофагах рыбок данио. Однако прайминг TNF индуцировал продукцию антибактериального матрикса mtROS двумя сходящимися сигнальными путями. Во-первых, TNF-опосредованная индукция продукции mtROS в матриксе с помощью рианоид-рецептор-взаимодействующей с рецептором серин / треонин-протеинкиназы 1 (RIPK1) -RIPK3-опосредованной перегрузки митохондрий кальцием [257].Во-вторых, поскольку матрикс mtROS не может выйти из матрикса интактных митохондрий [127,128,129], TNF открыл mPTP через сигнальный путь, включающий RIPK1, RIPK3 и домен киназы смешанного происхождения, такой как псевдокиназа (MLKL). Ранний эффект цитозольного mtROS, высвобождаемого открытым mPTP, приводил к гибели Mtb, но позже он приводил к некроптозу макрофагов [129]. Авторы неоднозначно предположили продукцию mtROS в фагосомах, однако в двух исследованиях это не было показано. Как упоминалось выше, Mtb уклоняется от деградации в фагосоме и быстро ускользает в цитозоль, где, как хорошо показано в двух статьях, цитозольные mtROS способствовали их уничтожению.В отличие от исследований на рыбках данио, Ким и его коллеги не обнаружили какой-либо антибактериальной роли матриксного mtROS во время инфекции Mtb при BMDM [258]. Они также наблюдали лишь незначительное увеличение продукции mtROS в матриксе только за счет инфекции. Однако это было сильно увеличено при BMDM, дефицитном по митохондриальной деацетилазе сиртуина 3 (SIRT3). Дефицит SIRT3 приводит к митохондриям с набухшими и поврежденными кристами, а поврежденные митохондрии, как известно, продуцируют большое количество матрикса mtROS [127,129,130,131].Однако эти индуцированные повреждением матричные mtROS в SIRT3-дефицитном BMDM были только побочным эффектом и не способствовали прямой антибактериальной защите. Вместо этого SIRT3 индуцировал антибактериальную ксенофагию Mtb, ускользнувшего из фагосомы. Таким образом, эти исследования показывают, что, поскольку Mtb эффективно избегает фагосомной деградации и инактивации за счет продукции ROS, происходящих из Nox2, макрофаги активируют чрезмерную продукцию mtROS, переходя к гибели клеток, чтобы минимизировать распространение инфекции Mtb (см. Рисунок 5).Salmonella typhimurium (S.t.), патоген, ответственный за тяжелые пищевые заболевания и основная причина диарейных заболеваний [259], также может уклоняться от деградации в фагосомах и размножаться в макрофагах [260]. Важность АФК при заражении S.t. подчеркивается сильно повышенной восприимчивостью мышей с дефицитом Nox2 к S.t. инфекция [261]. Антибактериальная роль производных Nox2 ROS, продуцируемых макрофагами во время S.t. инфекция была исследована во многих исследованиях [262, 263, 264], однако S.т. имеет большой набор средств для противодействия окислительному стрессу и даже его использования в своих целях [265]. В то время как деструктивный потенциал нейтрофилов за счет генерации OCl — с помощью миелопероксидазы (МПО) очевиден [266], макрофаги, похоже, борются в сбалансированной борьбе с S.t., в результате чего некоторые S.t. убиваются, в то время как другим успешно удается ремоделировать фаголизосомы в репликативную нишу [266, 267]. Помимо Nox2, Уэст и его коллеги представили митохондрии в качестве новых игроков в производстве антибактериальных АФК [61].Они впервые продемонстрировали, что С. Инфекция BMDM индуцировала рекрутирование митохондрий и продукцию mtROS в матриксе рядом с фагосомами в зависимости от фактора 6 (TRAF6), связанного с первичным ответом 88 (MyD88) -TNF рецептора дифференцировки TLR-миелоида. Производство матричных АФК затем способствовало инактивации S.t. Другое исследование также наблюдало кластеризацию митохондрий вокруг фагосом и индукцию матричной продукции mtROS в макрофагах рыбок данио, инфицированных S.t. [253]. Кластеризация митохондрий вокруг фагосом и продукция mtROS зависели от иммунно-чувствительного белка гена 1 (IRG1), 2-метилцитратдегидратазы.Было высказано предположение, что IRG1 питает цитратный цикл и тем самым усиливает поток электронов через ETC и, как следствие, также увеличивает производство mtROS в матрице. Соответственно, в макрофагах с дефицитом IRG1 кластеризация митохондрий и продукция mtROS в матриксе не индуцировались, а бактериальная нагрузка увеличивалась. Как и Mtb, S.t. предлагает множество методов, позволяющих выжить в фагосоме, зависимой от Nox2-зависимых АФК. Однако, в то время как Mtb ускользает в цитозоль, S.t. остается в фагосоме. В ответ макрофаги привлекают митохондрии к фагосоме, чтобы увеличить продукцию АФК в фагосоме, наконец, преодолевая антиоксидантные способности S.т. (см. рисунок 5) .Garaude et al. также исследовали перестройку комплексов ETC при инфицировании грамотрицательными бактериями [268]. В BMDM, инфицированном E.c., митохондрии проявляли пониженную активность комплекса I, но повышенную активность комплекса II. Хотя индукция матричного mtROS с ротеноном [269], обычно используемый положительный контроль для измерения матричного mtROS (см. Раздел 5.2 и Раздел 7) [30,270,271], все еще была возможна, E.c. инфекция не индуцировала продукцию mtROS в матриксе, что позволяет предположить, что сдвиг активности на комплекс II не приводит к усилению продукции mtROS в матрице.Напротив, внеклеточная продукция ROS, индуцированная инфекцией E.c, была вызвана Nox2, поскольку BMDM с дефицитом Nox2 полностью неспособен продуцировать внеклеточные ROS после заражения. Примечательно, что BMDM с дефицитом Nox2 также демонстрирует сниженную активность комплекса II, что указывает на сигнальную функцию производных от Nox2 внеклеточных ROS в перестройке ETC. Расположенная в митохондриях тирозинкиназа кошачьих гарднер-рашид (FGR) имела решающее значение для увеличения активности комплекса II, поскольку FGR-дефицитный BMDM показал сниженную активность комплекса II.К сожалению, механистические представления о том, как и как происходящие из Nox2 внеклеточные ROS достигают и регулируют FGR внутри митохондрий, отсутствуют, оставляя эти два основных открытия в корреляционной связи. Кроме того, не рассматривался вопрос о том, имеет ли комплекс II прямые антибактериальные функции. Недавнее исследование также изучило роль митохондриальных FGR в макрофагах в контексте ожирения [272]. В отличие от исследования Garaude et al., Матриксные mtROS, а не производные от Nox2 ROS имели решающее значение для активации FGR, подтверждая наблюдение других исследований, согласно которым ROS должны продуцироваться в непосредственной близости от регулируемой мишени и не насыщают чрезмерно ячейку, пока они не поразят свою цель случайным образом [29,30,37,61,125,273].
К сожалению, уровни АФК в Sa-содержащих фагосомах не были определены, например, с бактериями, связанными с зондом АФК [246] или с непроницаемыми для клеток внеклеточными зондами АФК (см. Раздел 5.2) [30,58,99,254], и поэтому явные доказательства отсутствует тот факт, что продукция фагосомных АФК действительно была увеличена за счет SOD2, доставленного в фагосомы через везикулы, происходящие из митохондрий. Эксперименты с SOD2-дефицитными макрофагами также могли усилить это предположение. Остается исследовать, индуцируется ли этот очень сложный путь усиления продукции антибактериальных АФК только S.а. или также другими патогенами, и влияет ли это на антибактериальный иммунитет in vivo. По крайней мере, для L.m., внеклеточная продукция ROS полностью независима от передачи сигналов TLR [58], исключая роль TLR-опосредованной доставки SOD2 в L.m.-содержащие фагосомы. Явная роль либо производных Nox2 ROS, либо mtROS в прямой антибактериальной защите макрофагов против S.a. поэтому остается не полностью охарактеризованным. Для прояснения этой темы потребуются будущие исследования с хорошо зарекомендовавшими себя измерениями ROS и твердыми генетическими данными.Как и S.a., Mycobacterium tuberculosis (Mtb) использует фагосомы макрофагов в качестве пролиферативной ниши. Механизмы выживания в фагосомах — это ингибирование слияния с лизосомами и избегание ксенофагии [255]. Благодаря своему фактору вирулентности CpsA, Mtb также активно ингибирует рекрутирование Nox2 в фагосомы. Предотвращая продукцию ROS, производных от Nox2, Mtb эффективно избегает нацеливания и уничтожения LAP [241,256]. Поскольку продукция АФК, происходящая из Nox2, эффективно подавляется и Mtb ускользает в цитозоль, макрофаги должны активировать другие цитозольные источники АФК, чтобы инактивировать этот патоген.Антибактериальная роль цитозольного mtROS против Mtb была исследована в двух исследованиях in vivo на рыбках данио [129, 257]. Одной инфекции Mtb было недостаточно для увеличения общих клеточных уровней ROS или для индукции матричной продукции mtROS в макрофагах рыбок данио. Однако прайминг TNF индуцировал продукцию антибактериального матрикса mtROS двумя сходящимися сигнальными путями. Во-первых, TNF-опосредованная индукция продукции mtROS в матриксе с помощью рианоид-рецептор-взаимодействующей с рецептором серин / треонин-протеинкиназы 1 (RIPK1) -RIPK3-опосредованной перегрузки митохондрий кальцием [257].Во-вторых, поскольку матрикс mtROS не может выйти из матрикса интактных митохондрий [127,128,129], TNF открыл mPTP через сигнальный путь, включающий RIPK1, RIPK3 и домен киназы смешанного происхождения, такой как псевдокиназа (MLKL). Ранний эффект цитозольного mtROS, высвобождаемого открытым mPTP, приводил к гибели Mtb, но позже он приводил к некроптозу макрофагов [129]. Авторы неоднозначно предположили продукцию mtROS в фагосомах, однако в двух исследованиях это не было показано. Как упоминалось выше, Mtb уклоняется от деградации в фагосоме и быстро ускользает в цитозоль, где, как хорошо показано в двух статьях, цитозольные mtROS способствовали их уничтожению.В отличие от исследований на рыбках данио, Ким и его коллеги не обнаружили какой-либо антибактериальной роли матриксного mtROS во время инфекции Mtb при BMDM [258]. Они также наблюдали лишь незначительное увеличение продукции mtROS в матриксе только за счет инфекции. Однако это было сильно увеличено при BMDM, дефицитном по митохондриальной деацетилазе сиртуина 3 (SIRT3). Дефицит SIRT3 приводит к митохондриям с набухшими и поврежденными кристами, а поврежденные митохондрии, как известно, продуцируют большое количество матрикса mtROS [127,129,130,131].Однако эти индуцированные повреждением матричные mtROS в SIRT3-дефицитном BMDM были только побочным эффектом и не способствовали прямой антибактериальной защите. Вместо этого SIRT3 индуцировал антибактериальную ксенофагию Mtb, ускользнувшего из фагосомы. Таким образом, эти исследования показывают, что, поскольку Mtb эффективно избегает фагосомной деградации и инактивации за счет продукции ROS, происходящих из Nox2, макрофаги активируют чрезмерную продукцию mtROS, переходя к гибели клеток, чтобы минимизировать распространение инфекции Mtb (см. Рисунок 5).Salmonella typhimurium (S.t.), патоген, ответственный за тяжелые пищевые заболевания и основная причина диарейных заболеваний [259], также может уклоняться от деградации в фагосомах и размножаться в макрофагах [260]. Важность АФК при заражении S.t. подчеркивается сильно повышенной восприимчивостью мышей с дефицитом Nox2 к S.t. инфекция [261]. Антибактериальная роль производных Nox2 ROS, продуцируемых макрофагами во время S.t. инфекция была исследована во многих исследованиях [262, 263, 264], однако S.т. имеет большой набор средств для противодействия окислительному стрессу и даже его использования в своих целях [265]. В то время как деструктивный потенциал нейтрофилов за счет генерации OCl — с помощью миелопероксидазы (МПО) очевиден [266], макрофаги, похоже, борются в сбалансированной борьбе с S.t., в результате чего некоторые S.t. убиваются, в то время как другим успешно удается ремоделировать фаголизосомы в репликативную нишу [266, 267]. Помимо Nox2, Уэст и его коллеги представили митохондрии в качестве новых игроков в производстве антибактериальных АФК [61].Они впервые продемонстрировали, что С. Инфекция BMDM индуцировала рекрутирование митохондрий и продукцию mtROS в матриксе рядом с фагосомами в зависимости от фактора 6 (TRAF6), связанного с первичным ответом 88 (MyD88) -TNF рецептора дифференцировки TLR-миелоида. Производство матричных АФК затем способствовало инактивации S.t. Другое исследование также наблюдало кластеризацию митохондрий вокруг фагосом и индукцию матричной продукции mtROS в макрофагах рыбок данио, инфицированных S.t. [253]. Кластеризация митохондрий вокруг фагосом и продукция mtROS зависели от иммунно-чувствительного белка гена 1 (IRG1), 2-метилцитратдегидратазы.Было высказано предположение, что IRG1 питает цитратный цикл и тем самым усиливает поток электронов через ETC и, как следствие, также увеличивает производство mtROS в матрице. Соответственно, в макрофагах с дефицитом IRG1 кластеризация митохондрий и продукция mtROS в матриксе не индуцировались, а бактериальная нагрузка увеличивалась. Как и Mtb, S.t. предлагает множество методов, позволяющих выжить в фагосоме, зависимой от Nox2-зависимых АФК. Однако, в то время как Mtb ускользает в цитозоль, S.t. остается в фагосоме. В ответ макрофаги привлекают митохондрии к фагосоме, чтобы увеличить продукцию АФК в фагосоме, наконец, преодолевая антиоксидантные способности S.т. (см. рисунок 5) .Garaude et al. также исследовали перестройку комплексов ETC при инфицировании грамотрицательными бактериями [268]. В BMDM, инфицированном E.c., митохондрии проявляли пониженную активность комплекса I, но повышенную активность комплекса II. Хотя индукция матричного mtROS с ротеноном [269], обычно используемый положительный контроль для измерения матричного mtROS (см. Раздел 5.2 и Раздел 7) [30,270,271], все еще была возможна, E.c. инфекция не индуцировала продукцию mtROS в матриксе, что позволяет предположить, что сдвиг активности на комплекс II не приводит к усилению продукции mtROS в матрице.Напротив, внеклеточная продукция ROS, индуцированная инфекцией E.c, была вызвана Nox2, поскольку BMDM с дефицитом Nox2 полностью неспособен продуцировать внеклеточные ROS после заражения. Примечательно, что BMDM с дефицитом Nox2 также демонстрирует сниженную активность комплекса II, что указывает на сигнальную функцию производных от Nox2 внеклеточных ROS в перестройке ETC. Расположенная в митохондриях тирозинкиназа кошачьих гарднер-рашид (FGR) имела решающее значение для увеличения активности комплекса II, поскольку FGR-дефицитный BMDM показал сниженную активность комплекса II.К сожалению, механистические представления о том, как и как происходящие из Nox2 внеклеточные ROS достигают и регулируют FGR внутри митохондрий, отсутствуют, оставляя эти два основных открытия в корреляционной связи. Кроме того, не рассматривался вопрос о том, имеет ли комплекс II прямые антибактериальные функции. Недавнее исследование также изучило роль митохондриальных FGR в макрофагах в контексте ожирения [272]. В отличие от исследования Garaude et al., Матриксные mtROS, а не производные от Nox2 ROS имели решающее значение для активации FGR, подтверждая наблюдение других исследований, согласно которым ROS должны продуцироваться в непосредственной близости от регулируемой мишени и не насыщают чрезмерно ячейку, пока они не поразят свою цель случайным образом [29,30,37,61,125,273].4.1.2. АФК против паразитов
В то время как Nox2 и митохондрии до сих пор являются единственными источниками АФК, которые, как было установлено, активируются макрофагами при бактериальном заражении, было высказано предположение, что несколько других ферментов Nox участвуют в защите от инфекции простейшими паразитами.
Toxoplasma gondii (T.g.) — такой простейший паразит, ответственный за токсоплазмоз, поражающий одну треть населения [274, 275]. Макрофаги играют центральную роль в иммунной защите от этого патогена [276].Хотя производство антимикробных АФК в целом было одним из первых ответов, выявленных в макрофагах, инфицированных T.g. [277, 278], относительный вклад различных источников АФК все еще неясен. Роль in vivo в продукции АФК с помощью Nox1 и Nox2 была предложена, поскольку мыши с дефицитом Nox1 или Nox2, инфицированные T.g. показали повышенную паразитарную нагрузку [279]. Т. Г. Инфекция также увеличивала общие клеточные уровни ROS при BMDM дикого типа in vitro, что отменялось при BMDM с дефицитом Nox1 или Nox2, в то время как паразитарное бремя увеличивалось, что предполагает роль Nox1 или Nox2 как важных источников ROS в противопаразитарной защите.Однако уровни ROS были только минимально снижены при BMDM с дефицитом Nox1 или Nox2 по сравнению с BMDM дикого типа, что вряд ли объясняет сильно сниженное паразитарное бремя, наблюдаемое in vitro и in vivo. Поскольку mtROS, которые могут вносить вклад во внеклеточную и цитозольную продукцию ROS, не исследовались, нельзя исключать роль их противопаразитарной защиты. Следует отметить, что другое исследование показало, что BMDM с дефицитом Nox2, инфицированный T.g. не показали изменений в паразитарной нагрузке [280]. Nox4 был предложен в качестве еще одного источника противопаразитарных АФК, поскольку у мышей с дефицитом Nox4 было меньше паразитарных кист, а паразитарная нагрузка увеличивалась при BMDM с дефицитом Nox4.Nox4 в основном обнаруживается на внутриклеточных органеллах, таких как ER и митохондрии [37,83,281,282], в то время как доказательства локализации Nox4 на фагосомных мембранах пока отсутствуют. Поскольку измерения ROS в клетках с дефицитом Nox2 или Nox4 не проводились, местоположение (фагосома, содержащая T.g. или другие клеточные органеллы) и функция (прямо или косвенно противопаразитарная) производных от Nox4 ROS оставались неуловимыми. Способность паразитов Toxoplasma детоксифицировать фагосомные АФК была продемонстрирована с помощью эволюционно родственной Toxoplasma cruzii в фагосомах J774A.1 макрофагоподобные клетки (J774) и BMDM [283]. Никаких измерений ROS с помощью классических зондов не проводилось, но изящные и технически сложные прямые измерения O 2 ● — в паразитарном цитозоле и фагосомах были выполнены в WT и Nox2-дефицитных клетках J774. Супероксиддисмутаза Fe-SODB в цитозоле паразитов была идентифицирована как основной механизм защиты от противопаразитарных АФК. Fe-SODB является консервативным среди паразитов Toxoplasma, что позволяет предположить, что этой способностью могут обладать и другие штаммы Toxoplasma, что ставит под сомнение до сих пор спорно обсуждаемую прямую противопаразитарную роль ROS у T.g.-инфицированные макрофаги. Паразиты простейших из рода Leishmania вызывают тропическое заболевание Leishamaniasis [284]. Нейтрофилы фагоцитируют и убивают 80–90% вторгшихся паразитов Leishmania [285, 286]. Макрофаги также участвуют в удалении паразитов, однако их роль имеет двойственную природу, как и у S.t. инфекционное заболевание. Макрофагам удается убить некоторых фагоцитированных паразитов, но они также составляют репликативную нишу [287, 288]. По крайней мере, один подвид Leishmania, Leishmania major, активно нарушает рекрутирование Nox2 в фагосомы, что приводит к снижению внеклеточной продукции ROS и снижению активации LAP [241,289].Возникает соблазн предположить, что все подвиды Leishmania способны снижать прямую продукцию антипаразитарных АФК за счет ингибирования рекрутирования Nox2. Инфекция Leishmania amazonensis в клетках J774 вызвала повышение уровней белка Nox2 и общих клеточных уровней АФК [290]. Однако для подтверждения этого предположения не использовались макрофаги с дефицитом Nox2. Производство ROS было подавлено с помощью DPI, который не является специфическим для Nox2 (см. Раздел 7), оставляя вопрос о точном источнике ROS без ответа.Хотя антимикробная роль АФК против паразитов в целом не подвергается сомнению, источники АФК остаются спорными и нуждаются в дальнейших исследованиях.SEC.gov | Превышен порог скорости запросов
Чтобы обеспечить равный доступ для всех пользователей, SEC оставляет за собой право ограничивать запросы, исходящие от необъявленных автоматизированных инструментов. Ваш запрос был идентифицирован как часть сети автоматизированных инструментов за пределами допустимой политики и будет обрабатываться до тех пор, пока не будут приняты меры по объявлению вашего трафика.
Укажите свой трафик, обновив свой пользовательский агент, включив в него информацию о компании.
Чтобы узнать о передовых методах эффективной загрузки информации с SEC.gov, в том числе о последних документах EDGAR, посетите sec.gov/developer. Вы также можете подписаться на рассылку обновлений по электронной почте о программе открытых данных SEC, включая передовые методы, которые делают загрузку данных более эффективной, и улучшения SEC.gov, которые могут повлиять на процессы загрузки по сценарию. Для получения дополнительной информации свяжитесь с opendata @ sec.губ.
Для получения дополнительной информации см. Политику конфиденциальности и безопасности веб-сайта SEC. Благодарим вас за интерес к Комиссии по ценным бумагам и биржам США.
Код ссылки: 0.5dfd733e.1640217005.695a8e21
Дополнительная информация
Политика интернет-безопасности
Используя этот сайт, вы соглашаетесь на мониторинг и аудит безопасности. В целях безопасности и для обеспечения того, чтобы общедоступная услуга оставалась доступной для пользователей, эта правительственная компьютерная система использует программы для мониторинга сетевого трафика для выявления несанкционированных попыток загрузки или изменения информации или иного причинения ущерба, включая попытки отказать пользователям в обслуживании.
Несанкционированные попытки загрузить информацию и / или изменить информацию в любой части этого сайта строго запрещены и подлежат судебному преследованию в соответствии с Законом о компьютерном мошенничестве и злоупотреблениях 1986 года и Законом о защите национальной информационной инфраструктуры 1996 года (см. Раздел 18 USC §§ 1001 и 1030).
Чтобы обеспечить хорошую работу нашего веб-сайта для всех пользователей, SEC отслеживает частоту запросов на контент SEC.gov, чтобы гарантировать, что автоматический поиск не влияет на возможность доступа других пользователей к SEC.содержание правительства. Мы оставляем за собой право блокировать IP-адреса, которые отправляют чрезмерное количество запросов. Текущие правила ограничивают пользователей до 10 запросов в секунду, независимо от количества машин, используемых для отправки запросов.
Если пользователь или приложение отправляет более 10 запросов в секунду, дальнейшие запросы с IP-адреса (-ов) могут быть ограничены на короткий период. Как только количество запросов упадет ниже порогового значения на 10 минут, пользователь может возобновить доступ к контенту на SEC.губ. Эта практика SEC предназначена для ограничения чрезмерного автоматического поиска на SEC.gov и не предназначена и не ожидается, чтобы повлиять на людей, просматривающих веб-сайт SEC.gov.
Обратите внимание, что эта политика может измениться, поскольку SEC управляет SEC.gov, чтобы гарантировать, что веб-сайт работает эффективно и остается доступным для всех пользователей.
Примечание: Мы не предлагаем техническую поддержку для разработки или отладки процессов загрузки по сценарию.
Споруляция в сольвентогенных и ацетогенных клостридиях
Абд-Алла М.Х., Зохри АНА, Эль-Энани AWE, Али С.М. (2017) Преобразование отходов пищевой промышленности в биотопливо с использованием клостридий.Анаэроб 48: 135–143. https://doi.org/10.1016/j.anaerobe.2017.08.011
CAS Статья PubMed Google Scholar
Abrini J, Naveau H, Nyns E-J (1994) Clostridium autoethanogenum , sp. nov., анаэробная бактерия, производящая этанол из окиси углерода. Arch Microbiol 161 (4): 345–351. https://doi.org/10.1007/BF00303591
CAS Статья Google Scholar
Афрамиан Н., Эльдар А. (2020) Бактериальная вавилонская башня: разнообразие сигналов, воспринимающее кворум, и его эволюция.Анну Рев Микробиол 74 (1): annurev-micro-012220-063740. https://doi.org/10.1146/annurev-micro-012220-063740
CAS Статья Google Scholar
Aklujkar M, Leang C, Shrestha PM, Shrestha M, Lovley DR (2017) Транскриптомные профили Clostridium ljungdahlii во время литотрофного роста с сингазом или H 2 и CO 2 по сравнению с органотрофным ростом с фруктами. Научный журнал 7 (1): 13135. https: // doi.org / 10.1038 / s41598-017-12712-w
CAS Статья PubMed PubMed Central Google Scholar
Al-Hinai MA, Jones SW, Papoutsakis ET (2014) σ K из Clostridium acetobutylicum — первый известный сигма-фактор, специфичный для споруляции, с двумя разделенными в процессе развития ролями: одной ранней и одной поздней в споруляции. J Bacteriol 196 (2): 287–299. https://doi.org/10.1128/JB.01103-13
CAS Статья PubMed PubMed Central Google Scholar
Аль-Хинаи М.А., Джонс С.В., Папуцакис Е.Т. (2015) Программы споруляции Clostridium : разнообразие и сохранение дифференцировки эндоспор.Microbiol Mol Biol Rev 79 (1): 19–37. https://doi.org/10.1128/MMBR.00025-14
Статья PubMed PubMed Central Google Scholar
Alsaker KV, Spitzer TR, Papoutsakis ET (2004) Транскрипционный анализ сверхэкспрессии spo0A в Clostridium acetobutylicum и его влияние на реакцию клетки на бутаноловый стресс. J Bacteriol 186 (7): 1959–1971. https://doi.org/10.1128/JB.186.7.1959-1971.2004
CAS Статья PubMed PubMed Central Google Scholar
Annous BA, Blaschek HP (1991) Выделение и характеристика мутантов Clostridium acetobutylicum с повышенной амилолитической активностью.Appl Environ Microbiol 57 (9)
Antunes A, Martin-Verstraete I, Dupuy B (2011) CcpA-опосредованная репрессия экспрессии гена токсина Clostridium difficile . Мол микробиол 79 (4): 882–899. https://doi.org/10.1111/j.1365-2958.2010.07495.x
CAS Статья PubMed Google Scholar
Атмаджая А.Н., Холби В., Хардинг А.Дж., Краббен П., Смит Х.К., Дженкинсон Э.Р. (2019) CRISPR-Cas, высокоэффективный инструмент для редактирования генома в Clostridium saccharoperbutylacetonicum N1-4 (HMT).FEMS Microbiol Lett 366. https://doi.org/10.1093/femsle/fnz059
Аврова Н.П., Зубко И.К., Алексеева Е.Г. (1981) Продукты брожения и активность пектолитических ферментов у штаммов Clostridium felsineum , различающихся в скорости образования спор. Микробиология 50 (3): 458–466 https://europepmc.org/article/med/7278720
CAS Google Scholar
Awang GM, Ingledew WM, Jones GA (1992) Влияние сбраживаемых углеводов на споруляцию и производство бутанола Clostridium acetobutylicum P262.Appl Microbiol Biotechnol 38 (1): 12–16. https://doi.org/10.1007/BF00169411
CAS Статья Google Scholar
Barák I, Muchová K, Labajová N (2019) Асимметричное деление клеток во время споруляции Bacillus subtilis . Future Microbiol 14 (4): 353–363. https://doi.org/10.2217/fmb-2018-0338
CAS Статья PubMed Google Scholar
Basu A, Xin F, Lim TK, Lin Q, Yang KL, He J (2017) Количественные профили протеома помогают выявить эффективные механизмы использования ксилозы в сольвентогенных Clostridium sp.штамм BOh4. Biotechnol Bioeng 114 (9): 1959–1969. https://doi.org/10.1002/bit.26332
CAS Статья PubMed Google Scholar
Bayat H, Modarressi MH, Rahimpour A (2018) Заметность системы CRISPR-Cpf1 как значительного прорыва в редактировании генома. Curr Microbiol 75 (1): 107–115. https://doi.org/10.1007/s00284-017-1406-8
CAS Статья PubMed Google Scholar
Behrens S, Meyer U, Schankin H, Lonetto MA, Fischer RJ, Bahl H (2000) Идентификация двух генов, кодирующих предполагаемых новых членов подсемейства ECF сигма-факторов эубактериальной РНК-полимеразы в Clostridium acetobutylicum .J Mol Microbiol Biotechnol 2 (3): 265–269
CAS PubMed Google Scholar
Березина О., Захарова Н., Яроцкий С., Зверлов В. (2012) Микробные продуценты бутанола. Appl Biochem Microbiol 48 (7): 625–638. https://doi.org/10.1134/S0003683812070022
CAS Статья Google Scholar
Bi C, Jones SW, Hess DR, Tracy MBP, Papoutsakis ET (2011) SpoIIe необходим для асимметричного деления, споруляции и экспрессии σ F , σ E и σ G , но не контролирует производство растворителей в Clostridium acetobutylicum ATCC 824.J Bacteriol 193 (19): 5130–5137. https://doi.org/10.1128/JB.05474-11
CAS Статья PubMed PubMed Central Google Scholar
Bischofs IB, Hug JA, Liu AW, Wolf DM, Arkin AP (2009) Сложность в коммуникации бактериальная клетка-клетка: интеграция сигнала кворума и передача сигналов субпопуляции в фосфореле Bacillus subtilis . Proc Natl Acad Sci 106 (16): 6459–6464. https://doi.org/10.1073/pnas.0810878106
Статья PubMed Google Scholar
Cañadas IC, Groothuis D, Zygouropoulou M, Rodrigues R, Minton NP (2019) RiboCas: универсальный инструмент редактирования на основе CRISPR для Clostridium .ACS Synth Biol 8 (6): 1379–1390. https://doi.org/10.1021/acssynbio.9b00075
CAS Статья PubMed Google Scholar
Cartman ST (2011) Пора подумать о пробиотиках Clostridium ? Future Microbiol 6 (9): 969–971. https://doi.org/10.2217/fmb.11.86
Статья PubMed Google Scholar
Chen Y, Zhou T, Liu D, Li A, Xu S, Liu Q, Li B, Ying H (2013) Производство бутанола из глюкозы и ксилозы с иммобилизованными клетками Clostridium acetobutylicum .Biotechnol Bioprocess Eng 18 (2): 234–241. https://doi.org/10.1007/s12257-012-0573-5
CAS Статья Google Scholar
Chen Y, Yang Y, Ji X, Zhao R, Li G, Gu Y, Shi A, Jiang W, Zhang Q (2020) Рантипептиды, производные SCIFF, участвуют в распознавании кворума в сольвентогенных клостридиях. Biotechnol J 15 (10): 1–8. https://doi.org/10.1002/biot.202000136
CAS Статья Google Scholar
Cheng C, Bao T, Yang ST (2019) Разработка Clostridium для улучшенного производства растворителей: недавний прогресс и перспективы.Appl Microbiol Biotechnol 103 (14): 5549–5566. https://doi.org/10.1007/s00253-019-09916-7
CAS Статья PubMed Google Scholar
Church BD (1959) Зависимость термостойкости бактериальных эндоспор от содержания в них дипиколиновой кислоты. Nature 183: 124–125
CAS Статья Google Scholar
Clark SW, Bennett GN, Rudolph FB (1989) Выделение и характеристика мутантов Clostridium acetobutylicum ATCC 824, дефицитных по ацетоацетил-коферменту a: ацетат / бутират: кофермент a-трансфераза (EC 2.8.3.9) и в других ферментах пути растворителя. Appl Environ Microbiol 55 (4): 970–976. https://doi.org/10.1128/AEM.55.4.970-976.1989
CAS Статья PubMed PubMed Central Google Scholar
Collins M, Lawson P, Willems A, Cordoba JJ, Fernandez-Garayzabal J, Garcia P, Cai J, Hippe H, Farrow J (1994) Филогения рода Clostridium : предложение пяти новых родов и одиннадцать новых комбинаций видов.Int J Syst Bacteriol 44 (4): 812–826. https://doi.org/10.1099/00207713-44-4-812
CAS Статья PubMed Google Scholar
Cornillot E, Nair RV, Papoutsakis ET, Soucaille P (1997) Гены образования бутанола и ацетона в Clostridium acetobutylicum ATCC 824 расположены на большой плазмиде, потеря которой приводит к дегенерации штамма. J Bacteriol 179 (17): 5442–5447 http://jb.asm.org/content/179/17/5442.аннотация
CAS Статья Google Scholar
Cruz-Morales P, Orellana CA, Moutafis G, Moonen G, Rincon G, Nielsen LK, Marcellin E, Bapteste E (2019) Пересмотр эволюции и таксономии клостридий, обновленное филогеномное сообщение. Genome Biol Evol 11 (7): 2035–2044. https://doi.org/10.1093/gbe/evz096
CAS Статья PubMed PubMed Central Google Scholar
Davidson P, Eutsey R, Redler B, Hiller NL, Laub MT, Durand D (2018) Гибкость и ограничение: эволюционное ремоделирование пути инициации споруляции в Firmicutes .PLoS Genet 14 (9): e1007470. https://doi.org/10.1371/journal.pgen.1007470
CAS Статья PubMed PubMed Central Google Scholar
Dawes IW, Mandelstam J (1970) Споруляция Bacillus subtilis в непрерывной культуре. J Bacteriol 103 (3): 529–535. https://doi.org/10.1128/jb.103.3.529-535.1970
CAS Статья PubMed PubMed Central Google Scholar
de Vrije T, Budde M, van der Wal H, Claassen PAM, López-Contreras AM (2013) Удаление изопропанола, бутанола и этанола из ферментационного бульона путем газовой отгонки «на месте».Биоресурсы Technol 137: 153–159. https://doi.org/10.1016/j.biortech.2013.03.098
CAS Статья PubMed Google Scholar
Decker AR, Ramamurthi KS (2017) Путь гибели клеток, который контролирует морфогенез спор. Trends Microbiol 25 (8): 637–647. https://doi.org/10.1016/j.tim.2017.03.005
CAS Статья PubMed PubMed Central Google Scholar
Dembek M, Willing SE, Hong HA, Hosseini S, Salgado PS, Cutting SM (2017) Индуцируемая экспрессия spo0A как универсального инструмента для изучения споруляции у Clostridium difficile .Front Microbiol 8: 1793. https://doi.org/10.3389/fmicb.2017.01793
Статья PubMed PubMed Central Google Scholar
Desai RP, Papoutsakis ET (1999) Стратегии антисмысловой РНК для метаболической инженерии Clostridium acetobutylicum . Appl Environ Microbiol 65 (3): 936–945. https://doi.org/10.1128/aem.65.3.936-945.1999
CAS Статья PubMed PubMed Central Google Scholar
Diallo M, Simons AD, van der Wal H, Collas F, Houweling-Tan B, Kengen SWM, López-Contreras AM (2018) метаболизм L-рамнозы в штамме Clostridium beijerinckii DSM 6423.Appl Environ Microbiol 85 (5). https://doi.org/10.1128/AEM.02656-18
Diallo M, Hocq R, Collas F, Chartier G, Wasels F, Wijaya HS, Werten MWT, Wolbert EJH, Kengen SWM, van der Oost J , Феррейра Н.Л., Лопес-Контрерас А.М. (2020a) Адаптация и применение системы CRISPR-Cas9, индуцируемой двумя плазмидами, в Clostridium beijerinckii . Методы 172: 51–60. https://doi.org/10.1016/j.ymeth.2019.07.022
CAS Статья PubMed Google Scholar
Diallo M, Kint N, Monot M, Collas F, Martin-Verstraete I, van der Oost J, Kengen SWM, López-Contreras AM (2020b) Транскриптомный и фенотипический анализ мутанта spoIIE в Clostridium beijerinckii .Front Microbiol 11 (сентябрь): 1–19. https://doi.org/10.3389/fmicb.2020.556064
Статья Google Scholar
Diender M, Parera Olm I, Gelderloos M, Koehorst JJ, Schaap PJ, Stams AJM, Sousa DZ (2019) Метаболический сдвиг, вызванный совместным синтетическим культивированием, способствует высокому выходу кислот с удлиненной цепью из синтез-газа. Научный журнал 9 (1): 18081. https://doi.org/10.1038/s41598-019-54445-y
CAS Статья PubMed PubMed Central Google Scholar
Долейш И., Краснян В., Стлоукал Р., Розенберг М., Реброш М. (2014) Производство бутанола с помощью иммобилизованного Clostridium acetobutylicum в режимах повторной периодической, периодической с подпиткой и непрерывной ферментации.Bioresour Technol 169: 723–730. https://doi.org/10.1016/j.biortech.2014.07.039
CAS Статья PubMed Google Scholar
Dürre P (2005) Справочник по клостридиям. В: Duerre P (ed) Handbook on Clostridia. CRC Press. https://doi.org/10.1201/9780203489819
Dürre P (2014) Физиология и споруляция Clostridium . В книге «Спора бактерий: от молекул к системам» (том 2, выпуск 4, стр.315–329). Американское общество микробиологов. https://doi.org/10.1128/microbiolspec.TBS-0010-2012
Эдвардс А.Н., Кролл Э.Г., МакБрайд С.М. (2020) Штамм-зависимая регуляция RstA выработки и споруляции токсина Clostridioides difficile . Дж. Бактериол 202 (2). https://doi.org/10.1128/JB.00586-19
Ehsaan M, Kuit W, Zhang Y, Cartman ST, Heap JT, Winzer K, Minton NP (2016) Генерация мутантов путем обмена аллелей и повторного секвенирования генома биобутанольного организма Clostridium acetobutylicum ATCC 824.Биотехнология Биотопливо 9 (1): 4. https://doi.org/10.1186/s13068-015-0410-0
CAS Статья PubMed PubMed Central Google Scholar
Ezeji T., Qureshi N, Blaschek HP (2007) Производство бутанола из сельскохозяйственных остатков: влияние продуктов разложения на рост Clostridium beijerinckii и ферментацию бутанола. Biotechnol Bioeng 97 (6): 1460–1469. https://doi.org/10.1002/bit.21373
CAS Статья PubMed Google Scholar
Falahati-Pour SK, Lotfi AS, Ahmadian G, Baghizadeh A (2015) Ковалентная иммобилизация рекомбинантной фосфорорганической гидролазы на спорах Bacillus subtilis .J Appl Microbiol 118 (4): 976–988. https://doi.org/10.1111/jam.12744
CAS Статья PubMed Google Scholar
Feng J, Zong W, Wang P, Zhang ZT, Gu Y, Dougherty M, Borovok I, Wang Y (2020) Системы контроля кворума RRNPP-типа регулируют образование растворителя, споруляцию и подвижность клеток в Clostridium saccharoperbutylacetonicum . Биотехнология Биотопливо 13 (1): 84. https://doi.org/10.1186/s13068-020-01723-x
Fimlaid KA, Shen A (2015) Различные механизмы регулируют активность сигма-фактора споруляции в Firmicutes .Curr Opin Microbiol 24: 88–95. https://doi.org/10.1016/j.mib.2015.01.006
CAS Статья PubMed PubMed Central Google Scholar
Forsberg CW (1987) Производство 1,3-пропандиола из глицерина с помощью Clostridium acetobutylicum и других видов Clostridium . Appl Environ Microbiol 53 (4): 639–643
CAS Статья Google Scholar
Foulquier C, Huang CN, Nguyen NPT, Thiel A, Wilding-Steel T, Soula J, Yoo M, Ehrenreich A, Meynial-Salles I, Liebl W, Soucaille P (2019) Эффективный метод безмаркерного создания мутантов путем аллельного обмена в Clostridium acetobutylicum и Clostridium saccharobutylicum с использованием суицидных векторов.Биотехнология Биотопливо 12 (1): 31. https://doi.org/10.1186/s13068-019-1364-4
CAS Статья PubMed PubMed Central Google Scholar
Freedman JC, Li J, Mi E, McClane BA (2019) Идентификация важной орфанской гистидинкиназы для инициации споруляции и продукции энтеротоксина Clostridium perfringens Тип F штамм SM101. MBio 10 (1): e02674 – e02618. https://doi.org/10.1128/mbio.02674-18
CAS Статья PubMed PubMed Central Google Scholar
Fujita M, González-Pastor JE, Losick R (2005) Гены высокого и низкого порога в регулоне Spo0A Bacillus subtilis .J Bacteriol 187 (4): 1357–1368. https://doi.org/10.1128/JB.187.4.1357-1368.2005
CAS Статья PubMed PubMed Central Google Scholar
Gallazzi A, Branska B, Marinelli F, Patakova P (2015) Непрерывное производство н-бутанола Clostridium pasteurianum DSM 525 с использованием суспендированных и иммобилизованных на поверхности клеток. J Biotechnol 216: 29–35. https://doi.org/10.1016/j.jbiotec.2015.10.008
CAS Статья PubMed Google Scholar
Galperin MY (2013) Разнообразие генома спорообразующих Firmicutes .Microbiol Spectr 1 (2). https://doi.org/10.1128/microbiolspectrum.TBS-0015-2012
Гальперин М.Ю., Мехедов С.Л., Пуигбо П., Смирнов С., Вольф Ю.И., Ригден Д.И. (2012) Геномные детерминанты споруляции у бацилл и клостридий: к минимальному набору генов, специфичных для споруляции. Environ Microbiol 14 (11): 2870–2890. https://doi.org/10.1111/j.1462-2920.2012.02841.x
CAS Статья PubMed PubMed Central Google Scholar
Джордж HA, Johnson JL, Moore WE, Holdeman LV, Chen JS (1983) Производство ацетона, изопропанола и бутанола с помощью Clostridium beijerinckii (син. Clostridium butylicum ) и Clostridium aurantibutyricum . Appl Environ Microbiol 45 (3): 1160–1163 http://aem.asm.org/content/45/3/1160.abstract
CAS Статья Google Scholar
Gong F, Bao G, Zhao C, Zhang Y, Li Y, Dong H (2016) Ферментация и геномный анализ производства не связанного с ацетоном бутанола с помощью Clostridium tetanomorphum . Appl Microbiol Biotechnol 100 (3): 1523–1529. https: // doi.org / 10.1007 / s00253-015-7121-0
CAS Статья PubMed Google Scholar
Green EM (2011) Ферментативное производство бутанола — промышленная перспектива. Curr Opin Biotechnol 22 (3): 337–343. https://doi.org/10.1016/j.copbio.2011.02.004
CAS Статья PubMed Google Scholar
Green EM, Boynton ZL, Harris LM, Rudolph FB, Papoutsakis ET, Bennett GN (1996) Генетическая манипуляция путей образования кислоты путем инактивации гена в Clostridium acetobutylicum ATCC 824.Микробиология 142 (8): 2079–2086. https://doi.org/10.1099/13500872-142-8-2079
CAS Статья PubMed Google Scholar Clostridium acetobutylicum . J Mol Microbiol Biotechnol 20 (1): 1–15. https: // doi.org / 10.1159 / 000320973
CAS Статья PubMed Google Scholar
Grossman AD, Losick R (1988) Внеклеточный контроль образования спор у Bacillus subtilis . Proc Natl Acad Sci U S A 85 (12): 4369–4373. https://doi.org/10.1073/pnas.85.12.4369
CAS Статья PubMed PubMed Central Google Scholar
Guo P, Zhang K, Ma X, He P (2020) Clostridium видов в качестве пробиотиков: возможности и проблемы.J Anim Sci Biotechnol 11 (1): 24. https://doi.org/10.1186/s40104-019-0402-1
Статья PubMed PubMed Central Google Scholar
Gupta RS, Gao B (2009) Филогеномный анализ клостридий и идентификация новых белковых сигнатур, специфичных для рода Clostridium sensu stricto (кластер I). Int J Syst Evol Microbiol 59 (2): 285–294. https://doi.org/10.1099/ijs.0.001792-0
CAS Статья PubMed Google Scholar
Harris LM, Desai RP, Welker NE, Papoutsakis ET (2000) Характеристика рекомбинантных штаммов мутанта по инактивации бутираткиназы Clostridium acetobutylicum : потребность в новых феноменологических моделях для сольвентогенеза и ингибирования бутанола? Biotechnol Bioeng 67 (1): 1–11.https://doi.org/10.1002/(SICI)1097-0290(20000105)
CAS Статья PubMed Google Scholar
Harris LM, Welker NE, Papoutsakis ET (2002) Северный, морфологический и ферментационный анализ инактивации и сверхэкспрессии spo0A в Clostridium acetobutylicum ATCC 824. J Bacteriol 184 (13): 3586–3597. https://doi.org/10.1128/JB.184.13.3586-3597.2002
CAS Статья PubMed PubMed Central Google Scholar
Hecker M, Völker U (2001) Общая стрессовая реакция Bacillus subtilis и других бактерий.В достижениях микробной физиологии (том 44, стр. 35–91). Академическая пресса. https://doi.org/10.1016/S0065-2911(01)44011-2
Heluane H, Dagher MRE, Bruno-Bárcena JM, Evans MR, Dagher SF, Bruno-Bárcena JM (2011) Мета-анализ и функциональная проверка потребностей в питании сольвентогенных клостридий, растущих в условиях бутанольного стресса и котилизации D-глюкозы и D-ксилозы. Appl Environ Microbiol 77 (13): 4473–4485. https://doi.org/10.1128/AEM.00116-11
CAS Статья PubMed PubMed Central Google Scholar
Herman NA, Kim SJ, Li JS, Cai W., Koshino H, Zhang W. (2017) Промышленный анаэроб Clostridium acetobutylicum использует поликетиды для регулирования клеточной дифференциации.Нац Коммуна 8 (1): 1514. https://doi.org/10.1038/s41467-017-01809-5
CAS Статья PubMed PubMed Central Google Scholar
Hocq R, Bouilloux-Lafont M, Lopes Ferreira N, Wasels F (2019) σ 54 (σ L ) играет центральную роль в метаболизме углерода в промышленно значимой Clostridium beijerinckii . Научный журнал 9 (1): 7228. https://doi.org/10.1038/s41598-019-43822-2
CAS Статья PubMed PubMed Central Google Scholar
Hosseini-Abari A, Kim B-G, Lee S-H, Emtiazi G, Kim W, Kim J-H (2016) Поверхностное отображение бактериальной тирозиназы на спорах Bacillus subtilis с использованием CotE в качестве якорного белка.J Basic Microbiol 56 (12): 1331–1337. https://doi.org/10.1002/jobm.201600203
CAS Статья PubMed Google Scholar
Hu S, Zheng H, Gu Y, Zhao J, Zhang W, Yang Y, Wang S, Zhao G, Yang S, Jiang W. (2011) Сравнительный геномный и транскриптомный анализ выявил генетические характеристики, связанные с образованием растворителя и ксилозы использование в Clostridium acetobutylicum EA 2018. BMC Genomics 12 (1): 93. https: // doi.org / 10.1186 / 1471-2164-12-93
CAS Статья PubMed PubMed Central Google Scholar
Huang SS, Chen D, Pelczar PL, Vepachedu VR, Setlow P, Li YQ (2007) Уровни Ca 2+ -дипиколиновой кислоты в отдельных спорах Bacillus , определенные с помощью микрожидкостного пинцета комбинационного рассеяния. J Bacteriol 189 (13): 4681–4687. https://doi.org/10.1128/JB.00282-07
CAS Статья PubMed PubMed Central Google Scholar
Huang H, Chai C, Yang S, Jiang W, Gu Y (2019) Геномная инженерия, опосредованная фаговой сериновой интегразой, для эффективной экспрессии пути химического биосинтеза при газовой ферментации Clostridium ljungdahlii .Metab Eng 52 (сентябрь 2018 г.): 293–302. https://doi.org/10.1016/j.ymben.2019.01.005
CAS Статья PubMed Google Scholar
Huhnke RL, Lewis RS, Tanner RS (2008) Выделение и характеристика новых видов клостридий (патент № US7704723 B2). В патенте (US7704723 B2). Бюро патентов и товарных знаков США. https://shareok.org/handle/11244/15409
Jabbari S, Steiner E, Heap JT, Winzer K, Minton NP, King JR (2013) Предполагаемое влияние оперона agr на используемые механизмы выживания по Clostridium acetobutylicum .Math Biosci 243 (2): 223–239. https://doi.org/10.1016/j.mbs.2013.03.005
CAS Статья PubMed Google Scholar
Jain MK, Beacom D, Datta R, Lansing E (1994) Мутантный штамм C. acetobutylicum и способ получения бутанола (Патент № US5192673A). В издании Biotechnology Advances (№ US5192673A). Бюро патентов и товарных знаков США. https://doi.org/10.1016/0734-9750(94)
Jamroskovic J, Chromikova Z, List C, Bartova B, Barák I, Bernier-Latmani R (2016) Изменчивость DPA и кальция содержание в спорах вида Clostridium .Front Microbiol 7 (NOV): 1791. https://doi.org/10.3389/fmicb.2016.01791
Статья PubMed PubMed Central Google Scholar
Jiao S, Zhang Y, Wan C, Lv J, Du R, Zhang R, Han B (2016) Транскрипционный анализ вырожденного штамма Clostridium beijerinckii DG-8052 показывает плейотропный ответ на CaCO 3-, связанный с восстановление сольвентного производства. Sci Rep 6: 38818. https://doi.org/10.1038/srep38818
CAS Статья PubMed PubMed Central Google Scholar
Джонсон EA (2019) Clostridia.Энцикл Microbiol 690–695. https://doi.org/10.1016/B978-0-12-801238-3.98821-4
Jones DT (2001) Применяемая ацетон-бутаноловая ферментация. В H Bahl & P Dürre (ред.) Clostridia (том 2, выпуск 1, стр. 125–168). Wiley-VCH Verlag GmbH. https://doi.org/10.1002/3527600108.ch5
Jones DT, Keis S (1995) Происхождение и взаимосвязь промышленных штаммов клостридиалов, продуцирующих растворитель. FEMS Microbiol. Ред. 17 (3): 223–232. https://doi.org/10.1111/j.1574-6976.1995.tb00206.x
CAS Статья Google Scholar
Jones DT, Woods DR (1986) Ацетон-бутаноловая ферментация еще раз. Microbiol Rev 50 (4): 484–524 3540574
CAS Статья Google Scholar
Jones SW, Paredes CJ, Tracy B, Cheng N, Sillers R, Senger RS, Papoutsakis ET (2008) Программа транскрипции, лежащая в основе физиологии споруляции клостридиумов.Геном Биол 9 (7): R114. https://doi.org/10.1186/gb-2008-9-7-r114
CAS Статья PubMed PubMed Central Google Scholar
Jones SW, Tracy BP, Gaida SM, Papoutsakis ET (2011) Инактивация σ F в Clostridium acetobutylicum ATCC 824 блокирует споруляцию до асимметричного деления и отменяет экспрессию σ E и σ G G но не блокирует образование растворителя.J Bacteriol 193 (10): 2429–2440. https://doi.org/10.1128/JB.00088-11
Джозеф Р.К., Ким Н.М., Сандовал Н.Р. (2018) Последние разработки инструментария синтетической биологии для Clostridium . Front Microbiol 9: 154. https://doi.org/10.3389/fmicb.2018.00154
CAS Статья PubMed PubMed Central Google Scholar
Кашкет Э. Р., Цао Z-Y (1995) Дегенерация клостридиального штамма. FEMS Microbiol.Ред. 17 (3): 307–315. https://doi.org/10.1016/0168-6445(95)00012-2
CAS Статья Google Scholar
Kay D, Warren SC (1968) Споруляция у Bacillus subtilis . Морфологические изменения. Biochem J 109 (5): 819–824. https://doi.org/10.1042/bj10
CAS Статья PubMed PubMed Central Google Scholar
Кейс С., Беннетт К.Ф., Уорд В.К., Джонс Д.Т. (1995) Таксономия и филогения промышленных клостридий, продуцирующих растворитель.Int J Syst Bacteriol 45 (4): 693–705. https://doi.org/10.1099/00207713-45-4-693
CAS Статья PubMed Google Scholar
Кейс С., Шахин Р., Джонс Д.Т. (2001) Измененные описания Clostridium acetobutylicum и Clostridium beijerinckii и описания Clostridium saccharoperbutylacetonicum 905charoperbutylacetonicum 905charoperbutylacetonicum. Int J Syst Evol Microbiol 51 (2001): 2095–2103.https://doi.org/10.1099/00207713-51-6-2095
CAS Статья PubMed Google Scholar
Kihm DJ, Hutton MT, Hanlin JH, Johnson EA (1988) Цинк стимулирует споруляцию в Clostridium botulinum 113B. Curr Microbiol 17 (4): 193–198. https://doi.org/10.1007/BF01589451
CAS Статья Google Scholar
Kiyoshi K, Kawashima S, Nobuki K, Kadokura T., Nakazato A, Suzuki KI, Nakayama S (2017) Добавление аденина восстанавливает жизнеспособность клеток и производство бутанола у Clostridium saccharoperbutylacetonicum , культивируемых при культивировании 13564 N1-4 (ATCC 37583). ° C.Appl Environ Microbiol 83 (7): e02960 – e02916. https://doi.org/10.1128/AEM.02960-16
CAS Статья PubMed PubMed Central Google Scholar
Knaysi G (1948) Эндоспора бактерий. Bacteriol Rev 12 (1): 19–77. https://doi.org/10.1128/mmbr.12.1.19-77.1948
CAS Статья PubMed PubMed Central Google Scholar C.beijerinckii NRRL B-598 и его мутантный штамм, сверхэкспрессирующий spo0A . Appl Microbiol Biotechnol 101 (22): 1–13. https://doi.org/10.1007/s00253-017-8555-3
CAS Статья Google Scholar
Косака Т., Накаяма С., Накая К., Йошино С., Фурукава К. (2007) Характеристика зольного оперона в гиперпродуцирующем бутанол штамме N1-4 Clostridium saccharoperbutylacetonicum и механизм его дегенерации.Biosci Biotechnol Biochem 71 (1): 58–68. https://doi.org/10.1271/bbb.60370
CAS Статья PubMed Google Scholar
Kosono S, Tamura M, Suzuki S, Kawamura Y, Yoshida A, Nishiyama M, Yoshida M (2015) Изменения ацетилома и сукцинилома Bacillus subtilis в ответ на источник углерода. PLoS One 10 (6): e0131169. https://doi.org/10.1371/journal.pone.0131169
CAS Статья PubMed PubMed Central Google Scholar
Kotte A-K, Severn O, Bean Z, Schwarz K, Minton NP, Winzer K (2020) Определение кворума RRNPP-типа влияет на образование растворителя и споруляцию в Clostridium acetobutylicum .Микробиология 166 (6): 579–592. https://doi.org/10.1099/mic.0.000916
CAS Статья PubMed PubMed Central Google Scholar
Król E, De Sousa BA, Kopacz M, Scheffers DJ (2017) Металл-зависимая олигомеризация SpoIIE стабилизирует FtsZ во время асимметричного деления в Bacillus subtilis . PLoS One 12 (3): e0174713. https://doi.org/10.1371/journal.pone.0174713
CAS Статья PubMed PubMed Central Google Scholar
Krouwel PG, Groot WJ, Kossen NWF, van der Laan WFM (1983) Непрерывная ферментация изопропанол-бутанол-этанол с иммобилизованными клетками Clostridium beijerinckii в ферментере с уплотненным слоем.Enzym Microb Technol 5: 46–54. https://doi.org/10.1016/0141-0229(83)-9
CAS Статья Google Scholar
Кубяк А.М., Минтон Н.П. (2015) Потенциал клостридиальных спор в качестве терапевтических средств доставки в терапии опухолей. Res Microbiol 166 (4): 244–254. https://doi.org/10.1016/j.resmic.2014.12.006
CAS Статья PubMed Google Scholar
Куит В. (2013) Метаболическая инженерия образования кислоты в Clostridium acetobutylicum Воутер Куит [Университет Вагенингена].http://library.wur.nl/WebQuery/wurpubs/fulltext/241021
Kuit W, Minton NP, López-Contreras AM, Eggink G (2012) Нарушение гена ацетаткиназы ( ack ) из Clostridium acetobutylicum приводит к задержке выработки ацетата. Appl Microbiol Biotechnol 94 (3): 729–741. https://doi.org/10.1007/s00253-011-3848-4
LanzaTech (2019) LanzaTech | Улавливание углерода. Подпитывая рост. LanzaTech. https://www.lanzatech.com/
Lawson PA, Rainey FA (2016) Предложение ограничить род Clostridium Prazmowski Clostridium butyricum и родственными видами.Int J Syst Evol Microbiol 66 (2): 1009–1016. https://doi.org/10.1099/ijsem.0.000824
Lee KY, Juang TC, Lee KC (1978) Влияние ионов металлов на рост и споруляцию Clostridium perfringens в синтетической среде. Chin J Microbiol Immunol 11 (2): 50–61
CAS Google Scholar
Lee AJ, Byun BY, Kang D-H, Tang J, Kim Y-W, Hwang H-J, Mah J-H (2011) Способность цинка ингибировать споруляцию и жизнеспособность Clostridium sporogenes и рост других бактерий.Int J Food Sci Technol 46 (7): 1494–1501. https://doi.org/10.1111/j.1365-2621.2011.02644.x
Lee S, Lee JH, Mitchell RJ (2015) Анализ транскрипционного ответа Clostridium beijerinckii NCIMB 8052 на феруловую кислоту и ее применение для повышения устойчивости к деформации. Биотехнология Биотопливо 8 (1): 68. https://doi.org/10.1186/s13068-015-0252-9
CAS Статья PubMed PubMed Central Google Scholar
Леггетт MJ, Mcdonnell G, Denyer SP, Setlow P, Maillard JY (2012) Бактериальные споровые структуры и их защитная роль в устойчивости к биоцидам.J Appl Microbiol 113 (3): 485–498. https://doi.org/10.1111/j.1365-2672.2012.05336.x
CAS Статья PubMed Google Scholar
Ленц С.А., Фогель РФ (2014) Влияние споруляционной среды и содержания в ней двухвалентных катионов на нагрев и устойчивость к высокому давлению спор Clostridium botulinum типа E. Пищевой микробиол 44: 156–167. https://doi.org/10.1016/j.fm.2014.05.010
CAS Статья PubMed Google Scholar
Letzel A-C, Pidot SJ, Hertweck C (2013) Геномный подход к загадочному вторичному метаболому анаэробного мира.Nat Prod Rep 30 (3): 392–428. https://doi.org/10.1039/C2NP20103H
CAS Статья PubMed Google Scholar
Li T, He J (2016) Одновременное осахаривание и ферментация гемицеллюлозы до бутанола с помощью неспорулирующих видов Clostridium . Bioresour Technol 219: 430–438. https://doi.org/10.1016/j.biortech.2016.07.138
CAS Статья PubMed Google Scholar
Li J, Chen J, Vidal JE, McClane BA (2011) Agr-подобная система определения кворума регулирует споруляцию и производство энтеротоксина и бета2-токсина Clostridium perfringens типа a непищевое желудочно-кишечное заболевание человека штамм F5603.Заражение иммунной 79 (6): 2451–2459. https://doi.org/10.1128/IAI.00169-11
CAS Статья PubMed PubMed Central Google Scholar
Li Q, Chen J, Minton NP, Zhang Y, Wen Z, Liu J, Yang H, Zeng Z, Ren X, Yang J, Gu Y, Jiang W, Jiang Y, Yang S (2016) CRISPR- основанные на системах редактирования генома и контроля экспрессии в Clostridium acetobutylicum и Clostridium beijerinckii . Biotechnol J 11 (7): 961–972.https://doi.org/10.1002/biot.201600053
CAS Статья PubMed Google Scholar
Li Q, Seys FM, Minton NP, Yang J, Jiang Y, Jiang W., Yang S (2019) Редактирование оснований CRISPR-Cas9 D10A с помощью никазы в производителе растворителя Clostridium beijerinckii . Biotechnol Bioeng 116: 1475–1483. https://doi.org/10.1002/bit.26949
CAS Статья PubMed Google Scholar
Li JS, Barber CC, Herman NA, Cai W., Zafrir E, Du Y, Zhu X, Skyrud W., Zhang W. (2020a) Исследование вторичного метаболизма промышленного гипер-продуцента бутанола Clostridium saccharoperbutylacetonicum N1- 4.J Ind Microbiol Biotechnol 47 (3): 319–328. https://doi.org/10.1007/s10295-020-02266-8
CAS Статья PubMed PubMed Central Google Scholar
Li S, Huang L, Ke C, Pang Z, Liu L (2020b) Рассечение пути, регулирование, разработка и применение: уроки, извлеченные из производства биобутанола с помощью сольвентогенных клостридий. Биотехнология Биотопливо 13 (1): 39. https://doi.org/10.1186/s13068-020-01674-3
CAS Статья PubMed PubMed Central Google Scholar
Лев Ф., Мартин М.Э., Таппель Р.С., Хейстра Б.Д., Михалча С., Кёпке М. (2016) Газовая ферментация — гибкая платформа для промышленного производства низкоуглеродного топлива и химикатов из отходов и возобновляемого сырья.Front Microbiol 7 (694). https://doi.org/10.3389/fmicb.2016.00694
Liou JS-C, Balkwill DL, Drake GR, Tanner RS (2005) Clostridium carboxidivorans sp. nov., продуцирующий растворитель Clostridium , выделенный из сельскохозяйственных отстойников, и реклассификация штамма acetogen Clostridium scatologenes SL1 как Clostridium drakei sp. ноя Int J Syst Evol Microbiol 55 (5): 2085–2091. https://doi.org/10.1099/ijs.0.63482-0
CAS Статья PubMed Google Scholar
List C, Hosseini Z, Lederballe Meibom K, Hatzimanikatis V, Bernier-Latmani R (2019) Влияние восстановления железа на метаболизм Clostridium acetobutylicum .Environ Microbiol 21 (10): 3548–3563. https://doi.org/10.1111/1462-2920.14640
CAS Статья PubMed PubMed Central Google Scholar
Liu Z, Qiao K, Tian L, Zhang Q, Liu Z-Y, Li F-L (2015) Спонтанный крупномасштабный автолиз у Clostridium acetobutylicum способствует образованию большего количества спор. Фронтальный микробиол 6: 950. https://doi.org/10.3389/fmicb.2015.00950
Статья PubMed PubMed Central Google Scholar
Liu H, Zhang J, Yuan J, Jiang X, Jiang L, Li Z, Yin Z, Du Y, Zhao G, Liu B, Huang D (2020) Сетевой анализ коэкспрессии генов показывает новый метаболический механизм Clostridium acetobutylicum реагирует на фенольные ингибиторы лигноцеллюлозных гидролизатов.Биотехнология Биотопливо 13 (1): 163. https://doi.org/10.1186/s13068-020-01802-z
CAS Статья PubMed PubMed Central Google Scholar
Long S, Jones DT, Woods DR (1983) Споруляция Clostridium acetobutylicum P262 в определенной среде. Appl Environ Microbiol 45 (4): 1389–1393. https://doi.org/10.1128/AEM.45.4.1389-1393.1983
CAS Статья PubMed PubMed Central Google Scholar
Long S, Jones D, Woods D (1984a) Инициирование образования растворителя, клостридиальной стадии и образования эндоспор в Clostridium acetobutylicum P262.Appl Microbiol Biotechnol 20 (4). https://doi.org/10.1007/BF00250635
Long S, Jones DT, Woods DR (1984b) Взаимосвязь между споруляцией и производством растворителя в Clostridium acetobutylicum P262. Biotechnol Lett 6 (8): 529–534. https://doi.org/10.1007/BF00139997
CAS Статья Google Scholar
Lund BM, Brocklehurst TF, Wyatt GM (1981) Характеристика штаммов Clostridium puniceum sp.nov., пектолитическая бактерия с розовым пигментом. J Gen Microbiol 122 (1): 17–26. https://doi.org/10.1099/00221287-122-1-17
Статья Google Scholar
Lv J, Jiao S, Du R, Zhang R, Zhang Y, Han B (2016) Протеомный анализ для выяснения дегенерации Clostridium beijerinckii NCIMB 8052 и роли Ca 2+ в восстановлении штамма после дегенерации. J Ind Microbiol Biotechnol 43 (6): 741-750. https://doi.org/10.1007 / с10295-016-1754-6
CAS Статья PubMed Google Scholar
Macek B, Forchhammer K, Hardouin J, Weber-Ban E, Grangeasse C, Mijakovic I (2019) Посттрансляционные модификации белков у бактерий. Nat Rev Microbiol 17 (11): 651–664. https://doi.org/10.1038/s41579-019-0243-0
CAS Статья PubMed Google Scholar
Mah JH, Kang DH, Tang J (2008) Влияние минералов на споруляцию и термостойкость Clostridium sporogenes .Int J Food Microbiol 128 (2): 385–389. https://doi.org/10.1016/j.ijfoodmicro.2008.10.002
CAS Статья PubMed Google Scholar
Malaviya A, Jang YS, Lee SY (2012) Непрерывное производство бутанола с уменьшенным образованием побочных продуктов из глицерина с помощью мутанта-гиперпродуцента Clostridium pasteurianum . Appl Microbiol Biotechnol 93 (4): 1485–1494. https://doi.org/10.1007/s00253-011-3629-0
CAS Статья PubMed Google Scholar
Мамане-Гравец Х., Линден К.Г. (2004) УФ-дезинфекция местных аэробных спор: значение для проверки УФ-реактора в нефильтрованной воде.Water Res 38 (12): 2898–2906. https://doi.org/10.1016/j.watres.2004.03.035
CAS Статья PubMed Google Scholar
Martínez-Lumbreras S, Alfano C, Evans NJ, Collins KM, Flanagan KA, Atkinson RA, Krysztofinska EM, Vydyanath A, Jackter J, Fixon-Owoo S, Camp AH, Isaacson RL (2018) Взгляды на структурные и функциональные аспекты в Bacillus subtilis ингибитор сигма-фактора, CsfB. Структура 26 (4): 640–648.e5. https: // doi.org / 10.1016 / j.str.2018.02.007
CAS Статья PubMed PubMed Central Google Scholar
Mate de Gerando H, Wasels F, Bisson A, Clement B, Bidard F, & Jourdier E. (2018). Последовательность генома природного продуцента изопропанола Clostridium beijerinckii DSM 6423. https://www.ebi.ac.uk/ena/browser/view/PRJEB11626
Máté de Gérando H, Wasels F, Bisson A, Clement , Bidard F, Jourdier E, López-Contreras AM, Lopes Ferreira N (2018) Геном и транскриптом природного продуцента изопропанола Clostridium beijerinckii DSM 6423.BMC Genomics 19 (1): 242. https://doi.org/10.1186/s12864-018-4636-7
CAS Статья PubMed PubMed Central Google Scholar
Meinecke B, Bahl H, Gottschalk G (1984) Выбор аспорогенного штамма Clostridium acetobutylicum в непрерывной культуре при ограничении фосфата. Appl Environ Microbiol 48 (5): 1064–1065. https://doi.org/10.1128/aem.48.5.1064-1065.1984
CAS Статья PubMed PubMed Central Google Scholar
Mukherjee M, Sarkar P, Goswami G, Das D (2019) Регулирование биосинтеза бутанола в Clostridium acetobutylicum ATCC 824 под влиянием цинковых добавок и магниевого голодания.Enzym Microb Technol 129: 109352. https://doi.org/10.1016/j.enzmictec.2019.05.009
CAS Статья Google Scholar
Mutschlechner O, Swoboda H, Gapes JR (2000) Непрерывная двухступенчатая ферментация ABE с использованием Clostridium beijerinckii NRRL B592, работающая со скоростью роста в емкости первой стадии, близкой к максимальной. J Mol Microbiol Biotechnol 2 (1): 101–105
CAS PubMed Google Scholar
Нарула Дж., Деви С.Н., Фуджита М., Игошин О.А. (2012) Сверхчувствительность решения споруляции Bacillus subtilis .Proc Natl Acad Sci 109 (50): E3513 – E3522. https://doi.org/10.1073/pnas.1213974109
Статья PubMed Google Scholar
Nimbalkar PR, Khedkar MA, Parulekar RS, Chandgude VK, Sonawane KD, Chavan PV, Bankar SB (2018) Роль микроэлементов как кофактора: эффективная стратегия увеличения производства биобутанола. ACS Sustain Chem Eng 6 (7): 9304–9313. https://doi.org/10.1021/acssuschemeng.8b01611
CAS Статья PubMed PubMed Central Google Scholar
Nimbalkar PR, Khedkar MA, Chavan PV, Bankar SB (2019) Повышенное производство биобутанола в среде, индуцированной фолиевой кислотой, с использованием Clostridium acetobutylicum NRRL B-527.ACS Omega 4 (7): 12978–12982. https://doi.org/10.1021/acsomega.9b00583
CAS Статья PubMed PubMed Central Google Scholar
Paidhungat M, Setlow B, Driks A, Setlow P (2000) Характеристика спор Bacillus subtilis , в которых отсутствует дипиколиновая кислота. J Bacteriol 182 (19): 5505–5512. https://doi.org/10.1128/JB.182.19.5505-5512.2000
CAS Статья PubMed PubMed Central Google Scholar
Papoutsakis ET (2008) Разработка сольвентогенных клостридий.Curr Opin Biotechnol 19 (5): 420–429. https://doi.org/10.1016/j.copbio.2008.08.003
CAS Статья PubMed Google Scholar
Paredes CJ, Alsaker KV, Papoutsakis ET (2005) Сравнительный геномный взгляд на споруляцию и физиологию клостридий. Nat Rev Microbiol 3 (12): 969–978. https://doi.org/10.1038/nrmicro1288
CAS Статья PubMed Google Scholar
Paredes-Sabja D, Setlow B, Setlow P, Sarker MR (2008) Характеристика спор Clostridium perfringens , в которых отсутствуют белки SpoVA и дипиколиновая кислота.J Bacteriol 190 (13): 4648–4659. https://doi.org/10.1128/JB.00325-08
CAS Статья PubMed PubMed Central Google Scholar
Paredes-Sabja D, Shen A, Sorg JA (2014) Clostridium difficile биология спор: споруляция, прорастание и структурные белки спор. Тенденции Microbiol 22 (7): 406–416. https://doi.org/10.1016/j.tim.2014.04.003
CAS Статья PubMed PubMed Central Google Scholar
Патакова П., Линхова М., Рихтера М., Паулова Л., Мелзох К. (2013) Новые и забытые проблемы ацетон-бутанол-этанольной ферментации клостридиями: Метаболическое разнообразие Clostridium , инструменты для картирования процессов и непрерывного системы ферментации.Biotechnol Adv 31 (1): 58–67. https://doi.org/10.1016/j.biotechadv.2012.01.010
CAS Статья PubMed Google Scholar
Патакова П., Линхова М., Выкидалова П., Бранска Б., Рихтера М., Мелзоч К. (2014) Использование флуоресцентного окрашивания и проточной цитометрии для мониторинга физиологических изменений в сольвентогенных клостридиях. Анаэроб 29: 113–117. https://doi.org/10.1016/j.anaerobe.2013.10.006
CAS Статья PubMed Google Scholar
Патакова П., Бранска Б., Седлар К., Васильковская М., Юрецкова К., Колек Дж., Косцова П., Провазник И. (2019) Ацидогенез, сольвентогенез, метаболическая реакция на стресс и изменения жизненного цикла у Clostridium beijerinckii NRRL B-59583 на транскриптомном уровне.Научный журнал 9 (1): 1371. https://doi.org/10.1038/s41598-018-37679-0
CAS Статья PubMed PubMed Central Google Scholar
Paul C, Filippidou S, Jamil I, Kooli W, House GL, Estoppey A, Hayoz M, Junier T, Palmieri F, Wunderlin T, Lehmann A, Bindschedler S, Vennemann T, Chain PSG, Junier P (2019 ) Споры бактерий, от экологии до биотехнологии. Adv Appl Microbiol 106: 79–111. https://doi.org/10.1016/bs.aambs.2018.10.002
CAS Статья PubMed Google Scholar
Peng F, Zheng B, Zhang Y, Faheem A, Chai Y, Jiang T, Chen X, Hu Y (2020) Биокаталитическое окисление ароматических соединений системой на основе спор. ACS Sustain Chem Eng 8 (37): 14159–14165. https://doi.org/10.1021/acssuschemeng.0c04884
CAS Статья Google Scholar
Philips J, Rabaey K, Lovley DR, Vargas M (2017) Формирование биопленки с помощью Clostridium ljungdahlii индуцируется стрессом хлорида натрия: экспериментальная оценка и анализ транскриптома.PLoS One 12 (1): e0170406. https://doi.org/10.1371/journal.pone.0170406
CAS Статья PubMed PubMed Central Google Scholar
Piggot PJ, Hilbert DW (2004) Споруляция Bacillus subtilis . Curr Opin Microbiol 7 (6): 579–586. https://doi.org/10.1016/j.mib.2004.10.001
CAS Статья PubMed Google Scholar
Poehlein A, Solano JDM, Flitsch SK, Krabben P, Winzer K, Reid SJ, Jones DT, Green E, Minton NP, Daniel R, Dürre P (2017) Повторное рассмотрение образования микробного растворителя с помощью сравнительного анализа генома.Биотехнология Биотопливо 10 (1): 58. https://doi.org/10.1186/s13068-017-0742-z
CAS Статья PubMed PubMed Central Google Scholar
Prazmowski A (1880) Untersuchungen über die Entwickelungsgeschichte und Fermentwirkung einiger Bacterien-Arten. Voigt.
Qureshi N, Blaschek HP (2001) Последние достижения в ферментации ABE: получение гипербутанола Clostridium beijerinckii BA101.J Ind Microbiol Biotechnol 27 (5): 287–291. https://doi.org/10.1038/sj.jim.7000114
CAS Статья PubMed Google Scholar
Raedts J, Siemerink MAJ, Levisson M, van der Oost J, Kengen SWM (2014) Молекулярная характеристика НАДФН-зависимой ацетоинредуктазы / 2,3-бутандиолдегидрогеназы из Clostridium beijerinckii NCIMB 8052. 80 (6): 2011–2020. https://doi.org/10.1128/AEM.04007-13
CAS Статья PubMed PubMed Central Google Scholar
Rathore S, Wan Sia Heng P, Chan LW (2015) Микрокапсулирование спор Clostridium acetobutylicum ATCC 824 в микросферах геллановой камеди для производства биобутанола. J Microencapsul 32 (3): 290–299. https://doi.org/10.3109/02652048.2015.1017617
CAS Статья PubMed Google Scholar
Ravagnani A, Jennert KCB, Steiner E, Grünberg R, Jefferies JR, Wilkinson SR, Young DI, Tidswell EC, Brown DP, Youngman P, Gareth Morris J, Young M (2000) Spo0A напрямую управляет переключением с кислоты к получению растворителя в клостридиях, образующих растворитель.Mol Microbiol 37 (5): 1172–1185. https://doi.org/10.1046/j.1365-2958.2000.02071.x
CAS Статья PubMed Google Scholar
Reeve BWP, Reid SJ (2016) Глутамат и гистидин улучшают как выход растворителя, так и реакцию устойчивости к кислоте Clostridium beijerinckii NCP 260. J Appl Microbiol 120 (5): 1271–1281. https://doi.org/10.1111/jam.13067
CAS Статья PubMed Google Scholar
Ren C, Gu Y, Wu Y, Zhang W, Yang C, Yang S, Jiang W (2012) Плейотропные функции белка контроля катаболита CcpA в продуцирующем бутанол Clostridium acetobutylicum .BMC Genomics 13 (1): 349. https://doi.org/10.1186/1471-2164-13-349
CAS Статья PubMed PubMed Central Google Scholar
Ricca E, Cutting SM (2003) Новые применения бактериальных спор в нанобиотехнологии. Дж. Нанобиотехнология 1 (1): 6. https://doi.org/10.1186/1477-3155-1-6
Статья Google Scholar
Rogers P, Palosaari N (1987) Мутанты Clostridium acetobutylicum , которые продуцируют масляный альдегид и измененное количество растворителей.Appl Environ Microbiol 53 (12): 2761–2766 http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=204194&tool=pmcentrez&rendertype=abstract
CAS Статья Google Scholar
Сандовал Н.Р., Венкатараманан К.П., Грот Т.С., Папуцакис Е.Т. (2015) Полногеномная последовательность эволюционировавшего штамма Clostridium pasteurianum выявляет дефицит Spo0A, ответственный за повышенное производство бутанола и превосходный рост.Биотехнология Биотопливо 8 (1): 227. https://doi.org/10.1186/s13068-015-0408-7
CAS Статья PubMed PubMed Central Google Scholar
Sandoval-Espinola WJ, Makwana ST, Chinn MS, Thon MR, Azcárate-Peril MA, Bruno-Bárcena JM (2013) Сравнительный фенотипический анализ и последовательность генома Clostridium beijerinckii SA-1, потомство NCIMB 8052 NCIMB 8052 Microbiology 159 (Pt_12): 2558–2570. https://doi.org/10.1099/mic.0,069534-0
CAS Статья PubMed PubMed Central Google Scholar
Sauer M (2016) Промышленное производство ацетона и бутанола путем ферментации — 100 лет спустя. FEMS Microbiol Lett 363 (13). https://doi.org/10.1093/femsle/fnw134
Sauer U, Santangelo JD, Treuner A, Buchholz M, Dürre P (1995) Сигма-фактор и гены споруляции в Clostridium . FEMS Microbiol Rev 17 (3): 331–340.https://doi.org/10.1111/j.1574-6976.1995.tb00216.x
CAS Статья PubMed Google Scholar
Schmeisser F, Brannigan JA, Lewis RJ, Wilkinson AJ, Youngman P, Barák I (2000) Новая мутация в spo0A с внутригенными супрессорами в эффекторном домене. FEMS Microbiol Lett 185 (2): 123–128. https://doi.org/10.1111/j.1574-6968.2000.tb09049.x
CAS Статья PubMed Google Scholar
Schuster KC, Van Den Heuvel R, Gutierrez NA, Maddox IS (1998) Разработка маркеров образования продукта и клеточного цикла при периодическом культивировании Clostridium acetobutylicum ATCC 824.Appl Microbiol Biotechnol 49 (6): 669–676. https://doi.org/10.1007/s002530051230
CAS Статья Google Scholar
Schuster KC, Goodacre R, Gapes J, Young M (2001) Дегенерация сольвентогенных штаммов Clostridium , отслеживаемая с помощью инфракрасной спектроскопии с преобразованием Фурье бактериальных клеток. J Ind Microbiol Biotechnol 27: 314–321. https://link.springer.com/content/pdf/10.1038/sj.jim.7000146.pdf
CAS Статья Google Scholar
Schwarz KM, Kuit W, Grimmler C, Ehrenreich A, Kengen SWM (2012) Исследование транскрипции ацидогенных клеток хемостата Clostridium acetobutylicum — клеточное поведение при адаптации к н-бутанолу.J. Biotechnol. 161 (3): 366–377. https://doi.org/10.1016/j.jbiotec.2012.03.018
CAS Статья PubMed Google Scholar
Schwarz KM, Grosse-Honebrink A, Derecka K, Rotta C, Zhang Y, Minton NP (2017) На пути к увеличению производства бутанола за счет целевой генетической модификации Clostridium pasteurianum . Metab Eng 40: 124–137. https://doi.org/10.1016/j.ymben.2017.01.009
CAS Статья PubMed PubMed Central Google Scholar
Scotcher MC, Bennett GN (2005) SpoIIE регулирует споруляцию, но не влияет напрямую на сольвентогенез в Clostridium acetobutylicum ATCC 824.J Bacteriol 187 (6): 1930–1936. https://doi.org/10.1128/JB.187.6.1930-1936.2005
CAS Статья PubMed PubMed Central Google Scholar
Scotcher MC, Rudolph FB, Bennett GN (2005) Экспрессия abrB310, и sinR , а также эффекты снижения экспрессии abrB310 на переходе от ацидогенеза к сольвентогенезу в Clostridium acetobutylicum Clostridium acetobutylicum. Environ Microbiol 71 (4): 1987–1995.https://doi.org/10.1128/AEM.71.4.1987-1995.2005
CAS Статья PubMed PubMed Central Google Scholar
Sedlar K, Koscova P, Vasylkivska M, Branska B, Kolek J, Kupkova K, Patakova P, Provaznik I (2018) Профилирование транскрипции продуцента бутанола Clostridium beijerinckii NRRL B-598 с использованием RNA-Seq. BMC Genomics 19 (1): 415. https://doi.org/10.1186/s12864-018-4805-8
CAS Статья PubMed PubMed Central Google Scholar
Sedlar K, Kolek J, Gruber M, Jureckova K, Branska B, Csaba G, Vasylkivska M, Zimmer R, Patakova P, Provaznik I (2019) Транскрипционный ответ Clostridium beijerinckii NRRL B-598 бутаноловый шок.Биотехнология Биотопливо 12 (1): 243. https://doi.org/10.1186/s13068-019-1584-7
CAS Статья PubMed PubMed Central Google Scholar
Седлар К., Васильковская М., Мусилова Ю., Бранска Б., Провазник И., Патакова П. (2021) Фенотипический и геномный анализ продуцента изопропанола и 1,3-пропандиола Clostridium diolis DSM 15410. Геномика 113 (1): 1109–1119. https://doi.org/10.1016/j.ygeno.2020.11.007
Seo SO, Janssen H, Magis A, Wang Y, Lu T, Price ND, Jin YS, Blaschek HP (2017a) Геномный, транскрипционный и фенотипический анализ глюкозо-дерепрессированного мутанта Clostridium beijerinckii , проявляющего фенотип кислотного краша.Biotechnol J 12 (11): 1700182. https://doi.org/10.1002/biot.201700182
CAS Статья Google Scholar
Seo SO, Wang Y, Lu T, Jin YS, Blaschek HP (2017b) Характеристика мутанта Clostridium beijerinckii spo0A и его применение для производства бутилбутирата. Biotechnol Bioeng 114 (1): 106–112. https://doi.org/10.1002/bit.26057
CAS Статья PubMed Google Scholar
Seo SO, Lu T, Jin Y-S, Blaschek HP (2021) Сравнительный фенотипический и геномный анализ мутанта Clostridium beijerinckii с повышенным продуцированием растворителя.J Biotechnol 329 (октябрь 2020 г.): 49–55. https://doi.org/10.1016/j.jbiotec.2021.02.002
CAS Статья PubMed Google Scholar
Serrano M, Crawshaw AD, Dembek M, Monteiro JM, Pereira FC, Pinho MG, Fairweather NF, Salgado PS, Henriques AO (2016) Комплекс SpoIIQ-SpoIIIAH Clostridium difficile контролирует поглощение предспор и поздние стадии экспрессия генов и морфогенез спор. Mol Microbiol 100 (1): 204–228.https://doi.org/10.1111/mmi.13311
CAS Статья PubMed PubMed Central Google Scholar
Seys FM, Rowe P, Bolt EL, Humphreys CM, Minton NP (2020) Золотой стандарт, стратегия комплементации на основе CRISPR / Cas9, основанная на 24 нуклеотидных последовательностях закладок. Гены (Базель) 11 (4): 458. https://doi.org/10.3390/genes11040458
CAS Статья Google Scholar
Шахин Р., Ширли М., Джонс Д. Т. (2000) Сравнительные исследования ферментации промышленных штаммов, принадлежащих к четырем видам клостридий, продуцирующих растворитель.J Mol Microbiol Biotechnol 2 (1): 115–124 10937496
CAS PubMed Google Scholar
Шен А., Эдвардс А.Н., Саркер М.Р., Паредес-Сабжа Д. (2019) Споруляция и прорастание клостридиальных патогенов. Microbiol Spectr 7 (6). https://doi.org/10.1128/microbiolspec.gpp3-0017-2018
Shi Z, Blaschek HP (2008) Транскрипционный анализ Clostridium beijerinckii NCIMB 8052 и мутанта BA101, продуцирующего гипербутанол, во время смены от ацидогенеза к сольвентогенезу.Appl Environ Microbiol 74 (24): 7709–7714. https://doi.org/10.1128/AEM.01948-08
CAS Статья PubMed PubMed Central Google Scholar
Sjolander NO, Langlykke AF, Peterson WH (1938) Брожение древесного сахара бутиловым спиртом. Ind Eng Chem 30 (11): 1251–1255. https://doi.org/10.1021/ie50347a011
CAS Статья Google Scholar
Song T, Wang F, Xiong S, Jiang H (2019) Поверхностное отображение фосфорорганических ферментов на рекомбинантной споре Bacillus subtilis .Biochem Biophys Res Commun 510 (1): 13–19. https://doi.org/10.1016/j.bbrc.2018.12.077
CAS Статья PubMed Google Scholar
Steiner E, Dago AE, Young DI, Heap JT, Minton NP, Hoch JA, Young M (2011) Множественные орфанные гистидинкиназы напрямую взаимодействуют с Spo0A, чтобы контролировать начало образования эндоспор в Clostridium acetobutylicum . Мол микробиол 80 (3): 641–654. https://doi.org/10.1111/j.1365-2958.2011.07608.x
CAS Статья PubMed PubMed Central Google Scholar
Steiner E, Scott J, Minton NP, Winzer K (2012) Система контроля кворума agr, которая регулирует образование гранул и споруляцию в Clostridium acetobutylicum . Appl Environ Microbiol 78 (4): 1113–1122. https://doi.org/10.1128/AEM.06376-11
CAS Статья PubMed PubMed Central Google Scholar
Sullivan L, Bennett GN (2006) Протеомный анализ и сравнение вариантов штаммов Clostridium acetobutylicum ATCC 824 и Spo0A.J Ind Microbiol Biotechnol 33 (4): 298–308. https://doi.org/10.1007/s10295-005-0050-7
CAS Статья PubMed Google Scholar
Таширо Ю., Йошида Т., Ногучи Т., Сономото К. (2013) Последние достижения и перспективы увеличения производства бутанола путем ацетон-бутанол-этанольной ферментации. Eng Life Sci 13 (5): 432–445. https://doi.org/10.1002/elsc.201200128
CAS Статья Google Scholar
Terracciano JS, Kashket ER (1986) Внутриклеточные условия, необходимые для инициирования продукции растворителя Clostridium acetobutylicum .Appl Environ Microbiol 52 (1): 86–91. https://doi.org/10.1128/aem.52.1.86-91.1986
CAS Статья PubMed PubMed Central Google Scholar
Theys J, Lambin P (2015) Clostridium для лечения рака: мечта или реальность? Ann Transl Med 3 (Приложение 1): S21. https://doi.org/10.3978/j.issn.2305-5839.2015.03.39
CAS Статья PubMed PubMed Central Google Scholar
Thorn GJ, King JR, Jabbari S (2013) pH-индуцированная генная регуляция продукции растворителя с помощью Clostridium acetobutylicum в непрерывной культуре: оценка параметров и моделирование споруляции.Math Biosci 241 (2): 149–166. https://doi.org/10.1016/J.MBS.2012.11.004
CAS Статья PubMed PubMed Central Google Scholar
Tokuyasu K, Yamada E (1959) Тонкая структура Bacillus subtilis . J Biophys Biochem Cytol 5 (1): 129–133. https://doi.org/10.1083/jcb.5.1.129
CAS Статья PubMed PubMed Central Google Scholar
Tomas CA, Beamish J, Papoutsakis ET (2004) Транскрипционный анализ бутанольного стресса и толерантности у Clostridium acetobutylicum .Журнал Бактериол 186 (7): 2006–2018. https://doi.org/10.1128/JB.186.7.2006-2018.2004
CAS Статья PubMed PubMed Central Google Scholar
Traag BA, Pugliese A, Eisen JA, Losick R (2013) Консервация генов среди эндоспорообразующих бактерий выявляет дополнительные гены споруляции в Bacillus subtilis . J Bacteriol 195 (2): 253–260. https://doi.org/10.1128/JB.01778-12
CAS Статья PubMed PubMed Central Google Scholar
Tracy BP, Gaida SM, Papoutsakis ET (2008) Разработка и применение методов проточной цитометрии для анализа и сортировки эндоспорообразующих клостридий.Appl Environ Microbiol 74 (24): 7497–7506. https://doi.org/10.1128/AEM.01626-08
CAS Статья PubMed PubMed Central Google Scholar
Tracy BP, Jones SW, Papoutsakis ET (2011) Инактивация σ E и σ G в Clostridium acetobutylicum проливает свет на их роль в биогенезе клостридиальных клеточных форм, синтезе гранулез, сольвентогенезе и морфогенезе спор . J Bacteriol 193 (6): 1414–1426.https://doi.org/10.1128/JB.01380-10
CAS Статья PubMed PubMed Central Google Scholar
Tracy BP, Jones SW, Fast AG, Indurthi DC, Papoutsakis ET (2012) Clostridia: важность их исключительного разнообразия субстратов и метаболитов для биотоплива и биопереработки. Curr Opin Biotechnol 23 (3): 364–381. https://doi.org/10.1016/j.copbio.2011.10.008
CAS Статья PubMed Google Scholar
Ugwuodo CJ, Nwagu TN (2020) Стабилизация ферментов путем иммобилизации на бактериальных спорах: обзор литературы.Int J Biol Macromol 166: 238–250. https://doi.org/10.1016/j.ijbiomac.2020.10.171
CAS Статья PubMed Google Scholar
Варга Дж., Стируолт В.Л., Мелвилл С.Б. (2004) Белок CcpA необходим для эффективной регуляции споруляции и регуляции гена энтеротоксина ( cpe ) в Clostridium perfringens . J Bacteriol 186 (16): 5221–5229. https://doi.org/10.1128/JB.186.16.5221-5229.2004
CAS Статья PubMed PubMed Central Google Scholar
Vasylkivska M, Jureckova K, Branska B, Sedlar K, Kolek J, Provaznik I, Patakova P (2019) Транскрипционный анализ поглощения аминокислот, ионов металлов, витаминов и углеводов в бутанол-продуцирующих продуктах Clostridium beijerinckii NRRL -598.PLoS One 14 (11): e0224560. https://doi.org/10.1371/journal.pone.0224560
CAS Статья PubMed PubMed Central Google Scholar
Vees CA, Neuendorf CS, Pflügl S (2020) На пути к непрерывной промышленной биопереработке с сольвентогенными и ацетогенными клостридиями: проблемы, прогресс и перспективы. J Ind Microbiol Biotechnol 1–35: 753–787. https://doi.org/10.1007/s10295-020-02296-2
CAS Статья Google Scholar
Wang P, Feng J, Guo L, Fasina O, Wang Y (2019) Engineering Clostridium saccharoperbutylacetonicum для получения высокого уровня изопропанол-бутанол-этанола (IBE) из просо проса, предварительно обработанного уксусной кислотой, с использованием системы CRISPR-Cas9.ACS Sustain Chem Eng 7 (21): 18153–18164. https://doi.org/10.1021/acssuschemeng.9b05336
CAS Статья Google Scholar
Wasels F, Chartier G, Hocq R, Lopes Ferreira N (2020) Подход к редактированию генома CRISPR / Anti-CRISPR подчеркивает синергию бутанолдегидрогеназ в Clostridium acetobutylicum DSM 792. Appl Environ Microbiol 86. https: / /doi.org/10.1128/aem.00408-20
Whitham JM, Tirado-Acevedo O, Chinn MS, Pawlak JJ, Grunden AM (2015) Метаболический ответ Clostridium ljungdahlii на воздействие кислорода.Appl Environ Microbiol 81 (24): 8379–8391. https://doi.org/10.1128/AEM.02491-15
CAS Статья PubMed PubMed Central Google Scholar
Wilkinson SR, Young M (1994) Нацеленная интеграция генов в хромосому Clostridium acetobutylicum . Микробиология 140 (1): 89–95. https://doi.org/10.1099/13500872-140-1-89
CAS Статья Google Scholar
Wilkinson SR, Young M, Goodacre R, Morris JG, Farrow JAE, Collins MD (1995) Фенотипические и генотипические различия между некоторыми штаммами Clostridium acetobutylicum .FEMS Microbiol Lett 125 (2–3): 199–204. https://doi.org/10.1111/j.1574-6968.1995.tb07358.x
Williams RHN, Whitworth DE (2010) Генетическая организация сигнальных путей двухкомпонентной системы прокариот. BMC Genomics 11 (1): 720. https://doi.org/10.1186/1471-2164-11-720
CAS Статья PubMed PubMed Central Google Scholar
Wolken WAM, Tramper J, van der Werf MJ (2003) Что споры могут сделать для нас? Trends Biotechnol 21 (8): 338–345.https://doi.org/10.1016/S0167-7799(03)00170-7
CAS Статья PubMed Google Scholar
Woo JE, Lee SY, Jang YS (2018) Влияние обогащения питательными веществами на производство кислоты из дегенерированного (не содержащего растворителей) штамма M5. Clostridium acetobutylicum . Appl Biol Chem 61 (4): 469–472. https://doi.org/10.1007/s13765-018-0372-6
CAS Статья Google Scholar
Woods DR, Jones DT (1987) Физиологические реакции штаммов Bacteroides и Clostridium на факторы стресса окружающей среды.Adv Microb Physiol 28 (C): 1–64. https://doi.org/10.1016/S0065-2911(08)60236-2
Статья Google Scholar
Xin F, Wang C, Dong W, Zhang W, Wu H, Ma J, Jiang M (2016) Комплексные исследования производства биобутанола неацетоном и 1,3-пропандиолом, производящим штамм Clostridium из глицерина и полисахариды. Биотехнология Биотопливо 9 (1): 220. https://doi.org/10.1186/s13068-016-0641-8
CAS Статья PubMed PubMed Central Google Scholar
Xin F, Yan W, Zhou J, Wu H, Dong W, Ma J, Zhang W, Jiang M (2018) Использование новых сольвентогенных штаммов дикого типа для производства бутанола.Биотехнология Биотопливо 11 (1): 252. https://doi.org/10.1186/s13068-018-1252-3
CAS Статья PubMed PubMed Central Google Scholar
Xin X, Cheng C, Du G, Chen L, Xue C (2020) Метаболическая инженерия гистидинкиназ в Clostridium beijerinckii для увеличения производства бутанола. Фронт Bioeng Biotechnol 8: 214. https://doi.org/10.3389/fbioe.2020.00214
Статья PubMed PubMed Central Google Scholar
Xu M, Zhao J, Yu L, Tang I-C, Xue C, Yang S-T (2015) Engineering Clostridium acetobutylicum с нокаутом гистидинкиназы для повышения толерантности к н-бутанолу и его продукции.Appl Microbiol Biotechnol 99 (2): 1011–1022. https://doi.org/10.1007/s00253-014-6249-7
CAS Статья PubMed Google Scholar
Xu M, Zhao J, Yu L, Yang S-T (2017) Сравнительный геномный анализ Clostridium acetobutylicum для понимания мутаций, способствующих повышенной толерантности и выработке бутанола. J Biotechnol 263: 36–44. https://doi.org/10.1016/J.JBIOTEC.2017.10.010
CAS Статья PubMed Google Scholar
Xu J-Y, Xu Z, Liu X, Tan M, Ye B-C (2018) Ацетилирование и бутирилирование белков регулируют фенотип и метаболические сдвиги образующих эндоспоры Clostridium acetobutylicum .Протеомика клеток Mol 17 (6): 1156–1169. https://doi.org/10.1074/mcp.RA117.000372
CAS Статья PubMed PubMed Central Google Scholar
Xue Q, Yang Y, Chen J, Chen L, Yang S, Jiang W, Gu Y (2016) Роли трех AbrB в регулировании двухфазной ферментации Clostridium acetobutylicum . Appl Microbiol Biotechnol 100 (21): 9081–9089. https://doi.org/10.1007/s00253-016-7638-x
CAS Статья PubMed Google Scholar
Yang X, Tu M, Xie R, Adhikari S, Tong Z (2013) Сравнение трех методов контроля pH для выявления эффектов недиссоциированной масляной кислоты на удельную скорость производства бутанола при периодической ферментации Clostridium acetobutylicum .АМБ Экспресс 3 (1): 3. https://doi.org/10.1186/2191-0855-3-3
CAS Статья PubMed PubMed Central Google Scholar
Yang Y, Lang N, Zhang L, Wu H, Jiang W, Gu Y (2020) Новый регуляторный путь, состоящий из двухкомпонентной системы и транспортера типа ABC, способствует толерантности к бутанолу у Clostridium acetobutylicum . Appl Microbiol Biotechnol 1–13: 5011–5023. https://doi.org/10.1007/s00253-020-10555-6
CAS Статья Google Scholar
Yu HY, Meade A, Liu SJ (2019) Филогения Clostridium spp.На основе консервативных генов и сравнений с другими деревьями. Микробиология 88 (4): 469–478. https://doi.org/10.1134/s0026261712x
CAS Статья Google Scholar
Ютин Н., Гальперин М.Ю. (2013) Обновленная информация о геноме клостридиальной филогении: грамотрицательные спорообразователи и другие неуместные клостридии. Environ Microbiol 15 (10): 2631–2641. https://doi.org/10.1111/1462-2920.12173
CAS Статья PubMed PubMed Central Google Scholar
Зан Дж. А., Саксена Дж. (2011) Новый этанологенный Clostridium видов, Clostridium coskatii. в патенте. http://www.google.com/patents/US20110229947
Zhang Y, Jiao S, Lv J, Du R, Yan X, Wan C, Zhang R, Han B (2017) Сигма-фактор регулирует клеточный ответ в нерастворитель, продуцирующий вырожденный штамм Clostridium beijerinckii : сравнительный анализ транскриптома. Фронтальный микробиол 8 (ЯНВАРЬ): 23. https://doi.org/10.3389/fmicb.2017.00023
Статья PubMed PubMed Central Google Scholar
Zhang X, Al-Dossary A, Hussain M, Setlow P, Li J (2020) Применение спор Bacillus subtilis в биотехнологии и передовых материалах.Appl Environ Microbiol 86 (17): 1–22. https://doi.org/10.1128/AEM.01096-20
Статья Google Scholar
Zhao H, Msadek T, Zapf J, Madhusudan HJA, Varughese KI (2002) ДНК-комплексная структура ключевого фактора транскрипции, инициирующего развитие спорообразующих бактерий. Структура 10 (8): 1041–1050. https://doi.org/10.1016/S0969-2126(02)00803-1
CAS Статья PubMed Google Scholar
Zhao Y, Tomas CA, Rudolph FB, Papoutsakis ET, Bennett GN (2005) Внутриклеточные концентрации бутирилфосфата и ацетилфосфата в Clostridium acetobutylicum и их значение для образования растворителя.Appl Environ Microbiol 71 (1): 530–537. https://doi.org/10.1128/AEM.71.1.530-537.2005
CAS Статья PubMed PubMed Central Google Scholar
Zhao R, Liu Y, Zhang H, Chai C, Wang J, Jiang W, Gu Y (2019) CRISPR-Cas12a-опосредованная делеция и регуляция гена в Clostridium ljungdahlii и его применение в перенаправлении потока углерода в синтезе газовое брожение. ACS Synth Biol 8 (10): 2270–2279. https: // doi.org / 10.1021 / acssynbio.9b00033
CAS Статья PubMed Google Scholar
Zheng Y-N, Li L-Z, Xian M, Ma Y-J, Yang J-M, Xu X, He D-Z (2009) Проблемы с микробным производством бутанола. J Ind Microbiol Biotechnol 36 (9): 1127–1138. https://doi.org/10.1007/s10295-009-0609-9
CAS Статья PubMed Google Scholar
Gale Apps — Технические трудности
Технические трудности
Приложение, к которому вы пытаетесь получить доступ, в настоящее время недоступно.Приносим свои извинения за доставленные неудобства. Пожалуйста, попробуйте еще раз через несколько секунд.
Если проблемы с доступом не исчезнут, обратитесь за помощью в наш отдел технического обслуживания по телефону 1-800-877-4253. Еще раз спасибо за выбор Gale, обучающей компании Cengage.
org.springframework.remoting.RemoteAccessException: невозможно получить доступ к удаленной службе [authorizationService @ theBLISAuthorizationService]; вложенное исключение — Ice.UnknownException unknown = «java.lang.IndexOutOfBoundsException: Индекс 0 выходит за границы для длины 0 в java.base / jdk.internal.util.Preconditions.outOfBounds (Preconditions.java:64) в java.base / jdk.internal.util.Preconditions.outOfBoundsCheckIndex (Preconditions.java:70) в java.base / jdk.internal.util.Preconditions.checkIndex (Preconditions.java:248) в java.base / java.util.Objects.checkIndex (Objects.java:372) в java.база / java.util.ArrayList.get (ArrayList.java:458) в com.gale.blis.data.subscription.dao.LazyUserSessionDataLoaderStoredProcedure.populateSessionProperties (LazyUserSessionDataLoaderStoredProcedure.java:60) в com.gale.blis.data.subscription.dao.LazyUserSessionDataLoaderStoredProcedure.reQuery (LazyUserSessionDataLoaderStoredProcedure.java:53) в com.gale.blis.data.model.session.UserGroupEntitlementsManager.reinitializeUserGroupEntitlements (UserGroupEntitlementsManager.java: 30) в com.gale.blis.data.model.session.UserGroupSessionManager.getUserGroupEntitlements (UserGroupSessionManager.java:17) в com.gale.blis.api.authorize.contentmodulefetchers.CrossSearchProductContentModuleFetcher.getProductSubscriptionCriteria (CrossSearchProductContentModuleFetcher.java:244) в com.gale.blis.api.authorize.contentmodulefetchers.CrossSearchProductContentModuleFetcher.getSubscribedCrossSearchProductsForUser (CrossSearchProductContentModuleFetcher.java: 71) в com.gale.blis.api.authorize.contentmodulefetchers.CrossSearchProductContentModuleFetcher.getAvailableContentModulesForProduct (CrossSearchProductContentModuleFetcher.java:52) в com.gale.blis.api.authorize.strategy.productentry.strategy.AbstractProductEntryAuthorizer.getContentModules (AbstractProductEntryAuthorizer.java:130) на com.gale.blis.api.authorize.strategy.productentry.strategy.CrossSearchProductEntryAuthorizer.isAuthorized (CrossSearchProductEntryAuthorizer.java: 82) в com.gale.blis.api.authorize.strategy.productentry.strategy.CrossSearchProductEntryAuthorizer.authorizeProductEntry (CrossSearchProductEntryAuthorizer.java:44) в com.gale.blis.api.authorize.strategy.ProductEntryAuthorizer.authorize (ProductEntryAuthorizer.java:31) в com.gale.blis.api.BLISAuthorizationServiceImpl.authorize_aroundBody0 (BLISAuthorizationServiceImpl.java:57) на com.gale.blis.api.BLISAuthorizationServiceImpl.authorize_aroundBody1 $ advice (BLISAuthorizationServiceImpl.java: 61) в com.gale.blis.api.BLISAuthorizationServiceImpl.authorize (BLISAuthorizationServiceImpl.java:1) в com.gale.blis.auth._AuthorizationServiceDisp._iceD_authorize (_AuthorizationServiceDisp.java:141) в com.gale.blis.auth._AuthorizationServiceDisp._iceDispatch (_AuthorizationServiceDisp.java:359) в IceInternal.Incoming.invoke (Incoming.java:209) в Ice.ConnectionI.invokeAll (ConnectionI.java:2800) на льду.ConnectionI.dispatch (ConnectionI.java:1385) в Ice.ConnectionI.message (ConnectionI.java:1296) в IceInternal.ThreadPool.run (ThreadPool.java:396) в IceInternal.ThreadPool.access 500 долларов (ThreadPool.java:7) в IceInternal.ThreadPool $ EventHandlerThread.run (ThreadPool.java:765) в java.base / java.lang.Thread.run (Thread.java:834) » org.springframework.remoting.ice.IceClientInterceptor.convertIceAccessException (IceClientInterceptor.java:365) org.springframework.remoting.ice.IceClientInterceptor.invoke (IceClientInterceptor.java:327) org.springframework.remoting.ice.MonitoringIceProxyFactoryBean.invoke (MonitoringIceProxyFactoryBean.java:71) org.springframework.aop.framework.ReflectiveMethodInvocation.proceed (ReflectiveMethodInvocation.java:186) org.springframework.aop.framework.JdkDynamicAopProxy.invoke (JdkDynamicAopProxy.java:212) com.sun.proxy. $ Proxy130.authorize (Неизвестный источник) com.gale.auth.service.BlisService.getAuthorizationResponse (BlisService.java:61) com.gale.apps.service.impl.MetadataResolverService.resolveMetadata (MetadataResolverService.java:65) com.gale.apps.controllers.DiscoveryController.resolveDocument (DiscoveryController.java:57) com.gale.apps.controllers.DocumentController.redirectToDocument (DocumentController.java:22) jdk.internal.reflect.GeneratedMethodAccessor315.invoke (Неизвестный источник) java.base / jdk.internal.reflect.DelegatingMethodAccessorImpl.invoke (DelegatingMethodAccessorImpl.java:43) java.base / java.lang.reflect.Method.invoke (Method.java:566) org.springframework.web.method.support.InvocableHandlerMethod.doInvoke (InvocableHandlerMethod.java: 215) org.springframework.web.method.support.InvocableHandlerMethod.invokeForRequest (InvocableHandlerMethod.java:142) org.springframework.web.servlet.mvc.method.annotation.ServletInvocableHandlerMethod.invokeAndHandle (ServletInvocableHandlerMethod.java:102) org.springframework.web.servlet.mvc.method.annotation.RequestMappingHandlerAdapter.invokeHandlerMethod (RequestMappingHandlerAdapter.java:895) org.springframework.web.servlet.mvc.method.annotation.RequestMappingHandlerAdapter.handleInternal (RequestMappingHandlerAdapter.java:800) org.springframework.web.servlet.mvc.method.AbstractHandlerMethodAdapter.handle (AbstractHandlerMethodAdapter.java:87) org.springframework.web.servlet.DispatcherServlet.doDispatch (DispatcherServlet.java:1038) org.springframework.web.servlet.DispatcherServlet.doService (DispatcherServlet.java:942) орг.springframework.web.servlet.FrameworkServlet.processRequest (FrameworkServlet.java:998) org.springframework.web.servlet.FrameworkServlet.doGet (FrameworkServlet.java:890) javax.servlet.http.HttpServlet.service (HttpServlet.java:626) org.springframework.web.servlet.FrameworkServlet.service (FrameworkServlet.java:875) javax.servlet.http.HttpServlet.service (HttpServlet.java:733) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:227) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.apache.tomcat.websocket.server.WsFilter.doFilter (WsFilter.java:53) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java: 162) org.apache.catalina.filters.HttpHeaderSecurityFilter.doFilter (HttpHeaderSecurityFilter.java:126) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.servlet.resource.ResourceUrlEncodingFilter.doFilter (ResourceUrlEncodingFilter.java:63) орг.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:101) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:101) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:101) орг.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.boot.web.servlet.support.ErrorPageFilter.doFilter (ErrorPageFilter.java:130) org.springframework.boot.web.servlet.support.ErrorPageFilter.access $ 000 (ErrorPageFilter.java:66) org.springframework.boot.web.servlet.support.ErrorPageFilter $ 1.doFilterInternal (ErrorPageFilter.java:105) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.springframework.boot.web.servlet.support.ErrorPageFilter.doFilter (ErrorPageFilter.java:123) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java: 162) org.springframework.boot.actuate.web.trace.servlet.HttpTraceFilter.doFilterInternal (HttpTraceFilter.java:90) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) орг.springframework.web.filter.RequestContextFilter.doFilterInternal (RequestContextFilter.java:99) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.FormContentFilter.doFilterInternal (FormContentFilter.java:92) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.HiddenHttpMethodFilter.doFilterInternal (HiddenHttpMethodFilter.java: 93) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.boot.actuate.metrics.web.servlet.WebMvcMetricsFilter.filterAndRecordMetrics (WebMvcMetricsFilter.java:154) орг.springframework.boot.actuate.metrics.web.servlet.WebMvcMetricsFilter.filterAndRecordMetrics (WebMvcMetricsFilter.java:122) org.springframework.boot.actuate.metrics.web.servlet.WebMvcMetricsFilter.doFilterInternal (WebMvcMetricsFilter.java:107) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) орг.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.springframework.web.filter.CharacterEncodingFilter.doFilterInternal (CharacterEncodingFilter.java:200) org.springframework.web.filter.OncePerRequestFilter.doFilter (OncePerRequestFilter.java:107) org.apache.catalina.core.ApplicationFilterChain.internalDoFilter (ApplicationFilterChain.java:189) org.apache.catalina.core.ApplicationFilterChain.doFilter (ApplicationFilterChain.java:162) org.apache.catalina.core.StandardWrapperValve.invoke (StandardWrapperValve.java:202) org.apache.catalina.core.StandardContextValve.invoke (StandardContextValve.java:97) org.apache.catalina.authenticator.AuthenticatorBase.invoke (AuthenticatorBase.java:542) org.apache.catalina.core.StandardHostValve.invoke (StandardHostValve.java:143) org.apache.catalina.вентили.ErrorReportValve.invoke (ErrorReportValve.java:92) org.apache.catalina.valves.AbstractAccessLogValve.invoke (AbstractAccessLogValve.java:687) org.apache.catalina.core.StandardEngineValve.invoke (StandardEngineValve.java:78) org.apache.catalina.connector.CoyoteAdapter.service (CoyoteAdapter.java:357) org.apache.coyote.http11.Http11Processor.service (Http11Processor.java:374) org.apache.coyote.AbstractProcessorLight.process (AbstractProcessorLight.java:65) org.apache.coyote.AbstractProtocol $ ConnectionHandler.process (AbstractProtocol.java:893) org.apache.tomcat.util.net.NioEndpoint $ SocketProcessor.doRun (NioEndpoint.java:1707) org.apache.tomcat.util.net.SocketProcessorBase.run (SocketProcessorBase.java:49) java.base / java.util.concurrent.ThreadPoolExecutor.runWorker (ThreadPoolExecutor.java:1128) Ява.base / java.util.concurrent.ThreadPoolExecutor $ Worker.run (ThreadPoolExecutor.java:628) org.apache.tomcat.util.threads.TaskThread $ WrappingRunnable.run (TaskThread.java:61) java.base / java.lang.Thread.run (Thread.java:834)
| Острые последствия физических упражнений | • PA вызывает повышенное поглощение глюкозы активными мышцами, уравновешенное производством глюкозы в печени, с большей зависимостью от углеводов для поддержания мышечной активности по мере увеличения интенсивности. | A / * |
| • Инсулино-стимулированное поглощение глюкозы в скелетных мышцах преобладает в состоянии покоя и нарушается при диабете 2 типа, в то время как мышечные сокращения стимулируют транспорт глюкозы с помощью отдельного, аддитивного механизма, не нарушенного инсулинорезистентностью или типом 2 сахарный диабет. | A / * | |
| • Хотя умеренные аэробные упражнения резко улучшают уровень глюкозы в крови и действие инсулина, риск гипогликемии, вызванной физической нагрузкой, минимален без использования экзогенного инсулина или стимуляторов секреции инсулина.Преходящая гипергликемия может следовать за интенсивной ПА. | C / * | |
| • Об острых эффектах упражнений с отягощениями при диабете 2 типа не сообщалось, но они приводят к снижению уровня глюкозы натощак в течение по крайней мере 24 часов после тренировки у лиц с НГГ. | C / * | |
| • Комбинация аэробных тренировок и упражнений с отягощениями может быть более эффективной для улучшения контроля уровня глюкозы в крови, чем любая из них по отдельности; тем не менее, необходимы дополнительные исследования, чтобы определить, является ли причиной этого общий расход калорий, продолжительность упражнений или режим упражнений. | B / * | |
| • Более мягкие формы упражнений (например, тай-чи, йога) дали смешанные результаты. | C / * | |
| • PA может привести к резкому улучшению системного действия инсулина, продолжающемуся от 2 до 72 часов. | A / * | |
| Хронические эффекты физических упражнений | • Как аэробные тренировки, так и тренировки с отягощениями улучшают действие инсулина, контроль уровня глюкозы в крови, а также окисление и накопление жира в мышцах. | B / * |
| • Упражнения с отягощениями увеличивают массу скелетных мышц. | A / * | |
| • Липидные реакции крови на тренировку неоднозначны, но могут привести к небольшому снижению холестерина ЛПНП без изменения холестерина ЛПВП или триглицеридов. Комбинированная потеря веса и ПА могут быть более эффективными, чем только аэробные упражнения на липидах. | C / * | |
| • Аэробная тренировка может немного снизить систолическое АД, но снижение диастолического АД встречается реже у людей с диабетом 2 типа. | C / * | |
| • Наблюдательные исследования показывают, что более высокая PA и физическая подготовка связаны с более низким риском смертности от всех причин и сердечно-сосудистых заболеваний. | C / * | |
| • Рекомендуемые уровни PA могут способствовать снижению веса. Однако может потребоваться до 60 минут в день, если для похудания полагаются только на упражнения. | C / * | |
| • Лица с сахарным диабетом 2 типа, участвующие в контролируемых тренировках, демонстрируют более высокую комплаентность и контроль уровня глюкозы в крови, чем те, кто занимается физическими упражнениями без присмотра. | B / * | |
| • Повышенная физическая нагрузка и физическая подготовка могут уменьшить симптомы депрессии и улучшить качество жизни, связанное со здоровьем, у людей с диабетом 2 типа. | B / * | |
| ПА и профилактика сахарного диабета 2 типа | • Не менее 2,5 часов в неделю на умеренную или интенсивную ПА следует проводить как часть изменения образа жизни для предотвращения развития диабета 2 типа у взрослых из группы высокого риска. | A / A |
| PA в профилактике и контроле GDM | • Эпидемиологические исследования показывают, что более высокие уровни PA могут снизить риск развития GDM во время беременности. | C / * |
| • РКИ показывают, что умеренная физическая нагрузка может снизить уровень глюкозы крови у матери при ГСД. | B / * | |
| Оценка перед тренировкой | • Перед тем, как приступить к упражнениям, более интенсивным, чем быстрая ходьба, лицам, ведущим сидячий образ жизни, с диабетом 2 типа, вероятно, будет полезно обследование у врача. ЭКГ-тестирование с физической нагрузкой для бессимптомных лиц с низким риском ИБС не рекомендуется, но может быть показано при более высоком риске. | C / C |
| Рекомендуемое участие в ПА для лиц с диабетом 2 типа | • Лица с диабетом 2 типа должны выполнять не менее 150 минут в неделю умеренных и интенсивных аэробных упражнений, распределенных по крайней мере 3 дня в течение недели. , с не более чем 2 днями подряд между приступами аэробной активности. | B / B |
| • Помимо аэробных тренировок, люди с диабетом 2 типа должны проводить тренировки с отягощениями от умеренного до интенсивного не менее 2–3 дней в неделю. | B / B | |
| • Контролируемые и комбинированные аэробные тренировки и тренировки с отягощениями могут принести дополнительную пользу для здоровья, хотя более легкие формы ПА (например, йога) показали смешанные результаты. Людям с диабетом 2 типа рекомендуется увеличивать общую суточную неструктурированную ФА. Тренинг по гибкости может быть включен, но его не следует проводить вместо других рекомендуемых типов PA. | B / C | |
| Упражнения с неоптимальным контролем ГК | • Лица с диабетом 2 типа могут участвовать в ПА, соблюдая осторожность при выполнении упражнений с уровнем ГК, превышающим 300 мг / дл (16.7 ммоль / л) без кетоза при условии хорошего самочувствия и достаточной гидратации. | C / E |
| • Люди с диабетом 2 типа, не использующие инсулин или средства, стимулирующие секрецию инсулина, вряд ли будут испытывать гипогликемию, связанную с ПА. Пользователям инсулина и средств, усиливающих секрецию инсулина, рекомендуется принимать добавки с углеводами по мере необходимости для предотвращения гипогликемии во время и после упражнений. | C / C | |
| Влияние лекарств на реакцию на упражнения | • Людям, принимающим инсулин или некоторые средства, стимулирующие секрецию инсулина, может потребоваться корректировка дозировки для предотвращения гипогликемии, связанной с физической нагрузкой.Большинство других лекарств, назначаемых при сопутствующих проблемах со здоровьем, не влияют на упражнения, за исключением β-адреноблокаторов, некоторых диуретиков и статинов. | C / C |
| Физические упражнения при отдаленных осложнениях диабета | • Известные ССЗ не являются абсолютным противопоказанием к упражнениям. Людям со стенокардией, отнесенным к категории умеренного или высокого риска, вероятно, следует начинать упражнения в рамках контролируемой программы кардиологической реабилитации. PA рекомендуется для всех с PAD. | C / C |
| • Лица с периферической невропатией и без острой язвы могут выполнять упражнения с умеренной нагрузкой.Для предотвращения и раннего выявления ран и язв рекомендуется комплексный уход за ногами, включая ежедневный осмотр ступней и использование соответствующей обуви. Умеренная ходьба, скорее всего, не увеличивает риск возникновения язв стопы или рецидива периферической невропатии. | B / B | |
| • Лица с CAN должны пройти обследование и получить одобрение врача и, возможно, тест с физической нагрузкой перед началом физических упражнений. Интенсивность упражнений лучше всего назначать методом резерва ЧСС с прямым измерением максимальной ЧСС. | C / C | |
| • Людям с неконтролируемой пролиферативной ретинопатией следует избегать действий, которые значительно повышают внутриглазное давление и риск кровотечения. | D / E | |
| • Физические упражнения улучшают физическую функцию и качество жизни у людей с заболеванием почек, и их можно даже проводить во время сеансов диализа. Наличие микроальбуминурии само по себе не требует ограничения физических упражнений. | C / C | |
| Принятие и поддержание физических упражнений людьми с диабетом | • Усилия по продвижению ФА должны быть сосредоточены на развитии самоэффективности и усилении социальной поддержки со стороны семьи, друзей и поставщиков медицинских услуг.Поощрение легкой или умеренной ПА может быть наиболее полезным для принятия и поддержания регулярного участия в ПА. Вмешательства в образ жизни могут иметь некоторую эффективность в продвижении поведения ПА. | B / B |
Мэри Хоун, Мэриленд, FACS | Стэнфордское здравоохранение
- Резиденция: Мичиганский университет (1998 г.) MI
- Стажировка: Мичиганский университет (1992) MI
- Сертификация совета: Американский совет хирургии, общая хирургия (1999)
- Стипендия: Орегонский университет здоровья и науки (2001) ИЛИ Соединенные Штаты Америки
- Медицинское образование: Мичиганский университет (1991), MI
- М.P.H., Школа общественного здравоохранения Мичиганского университета, MPH Epidemiology (1996)
- Доктор медицины, Медицинская школа Мичиганского университета (1991)
- B.S., Мичиганский университет, биомедицинские науки (1987)