
Биологическая продуктивность это: биологическая продуктивность
9.3. Биологическая продуктивность экосистем. Общая экология
Читайте также
Тема 5. ЕСТЕСТВЕННОЕ РАВНОВЕСИЕ И ЭВОЛЮЦИЯ ЭКОСИСТЕМ
Тема 5. ЕСТЕСТВЕННОЕ РАВНОВЕСИЕ И ЭВОЛЮЦИЯ ЭКОСИСТЕМ Понятие равновесия является одним из основных в науке. Но прежде чем говорить о равновесии в живой природе, выясним, что такое равновесие вообще и равновесие в неживой
Биологическая эволюция и антропогенез
Биологическая эволюция и антропогенез «Как похожа на нас безобразнейшая тварь, обезьяна» — печально констатировал когда-то античный поэт Энний, ещё не подозревавший о том, что говорит о собственных предках. Многие наши современники, видимо, разделяют эстетические вкусы
7. БИОЛОГИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ (БОС)
7. БИОЛОГИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ (БОС) Интерес к исследованиям биологической обратной связи (БОС) в целях произвольного управления ФС на основе объективной информации о динамике психофизиологических показателей возник в связи с данными об изменениях в ФС человека,
Глава 5.
 Биологическая эволюция
Биологическая эволюцияГлава 5. Биологическая эволюция Теория эволюции, объясняющая механизмы исторического развития живой природы, не просто общебиологическая теория. В значительной степени это мировоззренческая система, уровень разработанности которой имеет решающее значение для
8.3. Биологическая структура популяций
8.3. Биологическая структура популяций Основные показатели структуры популяций – численность, распределение организмов в пространстве и соотношение разнокачественных особей.Индивидуальные черты каждого организма зависят от особенностей его наследственной программы
9.4. Динамика экосистем
9.4. Динамика экосистем
Любой биоценоз динамичен, в нем постоянно происходит изменение в состоянии и жизнедеятельности его членов и соотношении популяций. Все многообразные изменения, происходящие в любом сообществе, можно отнести к двум основным типам: циклические и
Все многообразные изменения, происходящие в любом сообществе, можно отнести к двум основным типам: циклические и
Биологическая обусловленность манипуляционных игр
Биологическая обусловленность манипуляционных игр До сих пор мы рассматривали манипуляционные игры только на примере одного вида хищных млекопитающих (лисицы), у которого, однако, эта активность развита слабее, чем у многих других представителей этого отряда, особенно
Глава 6. ПРОДУКТИВНОСТЬ
Глава 6. ПРОДУКТИВНОСТЬ Когда бы знали мы, из какого сора растут стихи, не ведая стыда, то, надо думать, потеряли бы интерес к поэзии. Но в том и заключается работа системы, что продукция на выходе имеет более высокую организацию, чем сырье на входе. Это справедливо как для
7.5. Побочные эффекты и организация экосистем
7.
Продуктивность
Продуктивность Для исследования продуктивности Футс разработал постановку эксперимента, сходного с тем, который проводился при изучении культурной преемственности в условиях частичной изоляции обезьян. Снова в центр внимания шимпанзе ставился некий таинственный, не
24. Структура экосистем
 Сообщество, или биоценоз, – это
Сообщество, или биоценоз, – это26. Причины устойчивости и смены экосистем
26. Причины устойчивости и смены экосистем Вспомните!Какими взаимоотношениями связаны все организмы, входящие в состав одной экосистемы?Какая энергия поддерживает постоянный круговорот веществ в экосистеме?Причины устойчивости экосистем. Каждая экосистема – это
Изучение биологической продуктивности
Биологическая продуктивность представляет собой фундаментальное свойство биосферы и означает способность живого вещества воспроизводить биомассу и образовывать тем самым биотический покров.
Изучение биологической продуктивности — это сравнительно молодое направление биологической и географической науки, которое получило бурное развитие в связи с Международной биологической программой (МБП), начавшейся в конце 60-х годов и сфокусированной в первую очередь именно на понимании продукционных процессов в биотических сообществах и экосистемах.
Результатом работы по МБП послужило большое число специальных исследований по феноменологии продукционного процесса, а также по первичной продуктивности экосистем различных географических типов.
Продукционные процессы в экосистемах традиционно были объектом биогеографических исследований в Институте географии РАН. В отделе, а затем в лаборатории биогеографии в рамках нового направления этой науки, получившего название «География экосистем», разрабатывались методы экспериментального изучения первичной продукции, выполнялись натурные оценки размеров запаса фитомассы и ее годичного воспроизводства в экосистемах разных биомов суши, производилась систематизация опубликованных и оригинальных материалов по первичной продуктивности с целью создания региональных и глобальных баз данных.
Н. И. Базилевич с полным основанием считается организатором этих исследований в нашей стране. Настоящая монография представляет собой обобщение всего фактического материала по первичной продуктивности экосистем Северной Евразии, в пределах территории бывшего СССР. Подобные обзорные работы по этой территории прежде отсутствовали. Автору удалось собрать и унифицировать огромный материал более чем по 2500 пробных площадей, который характеризует с разной степенью детальности обширную территорию.
Подобные обзорные работы по этой территории прежде отсутствовали. Автору удалось собрать и унифицировать огромный материал более чем по 2500 пробных площадей, который характеризует с разной степенью детальности обширную территорию.
Для обеспечения сравнимости многочисленных данных Н. И. Базилевич разработаны специальные методические приемы обработки и унификации материалов. Этот раздел работы представляет особый интерес для всех исследователей, занимающихся составлением глобальных баз данных по продуктивности органического вещества.
В монографии для каждой пробной площади приведены показатели запасов живого растительного вещества (фитомассы) и мертвого растительного вещества (мортмассы) раздельно для надземного и подземного ярусов экосистемы. Кроме этих показателей, для каждого типа экосистем даются характеристики суммарной (надземной и подземной) продукции, создаваемой растительным сообществом за год.
Растительные формации, типология которых разработана в Ботаническом институте им. Комарова РАН, с некоторыми изменениями и дополнениями, сделанными Н. И. Базилевич, описываются в географических ординатах по Н. Н. Розову.
Комарова РАН, с некоторыми изменениями и дополнениями, сделанными Н. И. Базилевич, описываются в географических ординатах по Н. Н. Розову.
Согласно принципам, разработанным Н. Н. Розовым, в пределах Северной Евразии выделяются четыре термических пояса: полярный, бореальный, суббореальный и субтропический. Суббореальный и субтропический пояса в зависимости от степени увлажнения подразделяются на гумидные и семигумидные, семиаридные и аридные биоклиматические области. В пределах термических поясов и биоклиматических областей выделены природные зоны и горные области, которые, в свою очередь, объединяются в пять крупных физико-географических провинций: Европейская территория (ЕТС), Западная, Центральная и Восточная Сибирь, Притикоокеанский регион.
Большой интерес представляют оригинальные карты автора по запасам фито — и мортмассы, годичной продукции, составленные для восстановленного экосистемного покрова.
Таким образом, материалы, представленные в монографии, позволяют по-новому оценить параметры биологической продуктивности экосистем Северной Евразии в соответствии с физико-географическими условиями природной среды и наметить некоторые гидротермические рубежи, обусловливающие географию продукционного потенциала экосистем.
Р. И. Злотый
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
7. Биологическая продуктивность экосистем: первичная и вторичная продукция, правила пирамид, распределение биологической продукции. Биоценоз и экосистема
Похожие главы из других работ:
Влияние мебельного производства ООО «Экос» на окружающую среду г. Дубна
1.3 Мощность предприятия. Сырье и продукция
Основным сырьем на деревообрабатывающем производстве является древесина. Основные породами, используемые в цикле производства ООО «Экос» являются: сосна обыкновенная, береза бородавчатая и дуб черешчатый. Таблица 1…
Воздействие человека на биосферу
2. Продуктивность экосистем
По мере того, как человечество с упрямством, достойным лучшего применения, превращает лицо Земли в сплошной антропогенный ландшафт, все большее практическое значение приобретает оценка продуктивности различных экосистем.
Контроль и улучшение качества услуг, предоставляемых МУП «Водоканал»
2.1 Водозабор и первичная очистка
Для водоснабжения г. Екатеринбурга используются водные ресурсы рек Чусовой, Уфы, Исеть, Ревды с зарегулированными на них водохранилищами: Нязе-Петровским, Верхне-Макаровским, Волчихинским, Верх-Исетским, Ревдинским, Ново-Мариинским…
Надежность биотехнологических систем и охрана окружающей среды
1.4 Правила оптовой торговли GDP и Правила надлежащей аптечной практики GPP
Существуют ещё два стандарта, регулирующие деятельность по оптовой и розничной торговле лекарственными средствами — Правила оптовой торговли GDP и Правила надлежащей аптечной практики GPP. Аналог GDP — ОСТ 915000.05…
Оценка экологичности предприятия ООО «Опытный стекольный завод»
2.2 Продукция ОСЗ
Продукция предприятия сертифицирована, соответствует государственным стандартам и имеет награды многих представительных выставок. Опытные мастера и технологии производства помогут изготовить стеклоизделия по индивидуальному заказу…
Опытные мастера и технологии производства помогут изготовить стеклоизделия по индивидуальному заказу…
Продуктивность природных и антропогенных экосистем
3. Агроэкосистемы и их продуктивность
Агроэкосистема (сельскохозяйственная экосистема, агроценоз, агробиоценоз) — биотическое сообщество, созданное и регулярно поддерживаемое человеком с целью получения сельскохозяйственной продукции. Обычно включает совокупность организмов…
Ранжирование территории по эколого-экономическим критериям на примере Волго-Вятского экономического района
2. Первичная обработка информации
…
Рынок экологически чистой продукции
2. ЭКОЛОГИЧЕСКИ ЧИСТАЯ ПРОДУКЦИЯ В УКРАИНЕ
Значительная часть человечества всегда небезосновательно проявляла заботу об удовлетворении своих потребительских нужд экологически чистыми продуктами питания, выращенными без применения в технологическом цикле компонентов…
Современные проблемы экологии
1.
 Продуктивность биосферы
Продуктивность биосферыВся биосфера с точки зрения глобальных оценок ее биомассы и продуктивности характеризуется следующими цифрами. Биомасса (масса живого вещества) биосферы по сравнению с массой Земли незначительная. Она составляет всего 0…
Сообщество, экосистема, биогеоценоз
Продуктивность сообщества
Продуктивность. Важным функциональным показателем сообществ является их способность к созданию (продуцированию) новой биомассы. Это свойство лежит в основе понятия продуктивность, сходного по смыслу с понятием плодородие…
Строение биосферы. Загрязнение экосистем. Проведение экологической экспертизы
2. Загрязнение экосистем промышленности сточными водами. Преобладающие загрязняющие вещества водных экосистем по отраслям промышленности
Промышленные сточные воды загрязняют экосистемы самыми разнообразными компонентами (табл.1) в зависимости от специфики отраслей промышленности. Следует заметить…
Упаковка и окружающая среда
3.
 2 Вторичная переработка
2 Вторичная переработкаВторичная переработка определяется как некоторая промышленная переработка материала для достижения целей первоначального использования или иных целей. К последним может относиться, например, органическая переработка (получение компоста)…
Утилизация отходов
3. Рециклизация — вторичная переработка отходов
Рециклизация, т.е. вторичная переработка отходов, — очевидный выход из положения. Разумеется, многие предлагали его и раньше. В небольших масштабах стекло, бумага и алюминиевые банки рециклизуются уже десятки лет…
Экологически безопасная продукция
Понятие «экологически безопасная продукция»
Производство экологически безопасной продукции — ключевая задача при экологизации сельскохозяйственной деятельности…
Эстуарии: общая характеристика
1. Биомасса и продуктивность эстуариев
Большинство эстуариев представляют собой заболоченные места. Реки приносят ил, который состоит из смеси очень маленьких кусочков твердой породы с растительным материалом. У моря река течет очень медленно, и ил спокойно оседает на дно…
У моря река течет очень медленно, и ил спокойно оседает на дно…
Биологическая продуктивность экосистемы
Автотрофные экосистемы можно сравнить с промышленным предприятием, ĸᴏᴛᴏᴩᴏᴇ производит различные органические вещества. Используя солнечную энергию, диоксид углерода и элементы минерального питания, экосистемы производят биологическую продукцию – древесину, листовую массу растений, плоды, животную биомассу. Производительность экосистемы, измеряемая количеством органического вещества, ĸᴏᴛᴏᴩᴏᴇ создано за единицу времени на единицу площади, принято называть биологической продуктивностью. Единицы измерения продуктивности: г/м2 в день, кг/м2 в год, т/км2 в год.
На рис. 49 показана структура биологической продукции экосистемы. Различают первичную биологическую продукцию, которую создают растения в процессе фотосинтеза из диоксида углерода, воды и минеральных элементов, и вторичную биологическую продукцию, которую создают гетеротрофы (консументы и редуценты) в результате переработки растительной и животной биомассы. Первичную продукцию подразделяют на валовую – общее количество созданного органического вещества и чистую – то, что осталось после расходов на дыхание и корневые выделения. У большинства растений чистая продукция составляет примерно половину от валовой.
Первичную продукцию подразделяют на валовую – общее количество созданного органического вещества и чистую – то, что осталось после расходов на дыхание и корневые выделения. У большинства растений чистая продукция составляет примерно половину от валовой.
Поскольку консументы лишь используют ранее созданные органические вещества, вторичную продукцию на валовую и чистую не разделяют. Но ее количество также зависит от расходов на дыхание, ĸᴏᴛᴏᴩᴏᴇ тем больше, чем больше энергии затрачивает организм. При интенсивной физической нагрузке (к примеру, у птиц во время миграции) продукция уменьшается. Привесы у сельскохозяйственных животных убывают, в случае если им приходится затрачивать много энергии на переходы с отдаленных пастбищ или согревание тела в холодном помещении.
При переходе энергии с одного трофического уровня на другой (от растений к фитофагам, от фитофагов к хищникам первого порядка, от хищников первого порядка к хищникам второго порядка) с экскрементами и затратами на дыхание теряется примерно 90% энергии.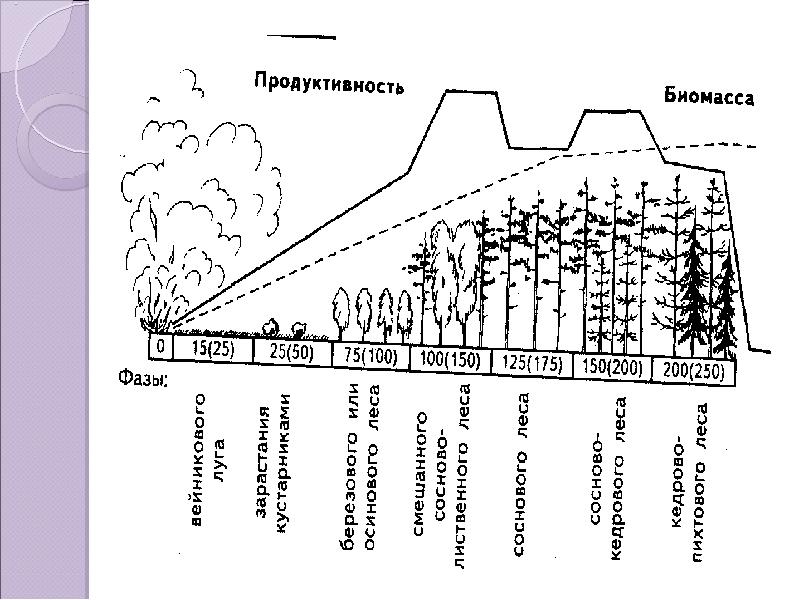 Вместе с тем, фитофаги съедают только часть биомассы растений, а остальная – пополняет запас детрита͵ который затем разрушают редуценты. По этой причине вторичная биологическая продукция в 20–50 раз меньше, чем первичная.
Вместе с тем, фитофаги съедают только часть биомассы растений, а остальная – пополняет запас детрита͵ который затем разрушают редуценты. По этой причине вторичная биологическая продукция в 20–50 раз меньше, чем первичная.
По продуктивности (в сыром весе) экосистемы разделяются на четыре класса.
1. Экосистемы очень высокой биологической продуктивности – свыше 2 кг/м2 в год. К ним относятся заросли тростника в дельтах Волги, Дона и Урала. По продуктивности они близки к экосистемам тропических лесов, коралловых рифов и глубоководных ʼʼоазисовʼʼ рифтовых зон океана.
2. Экосистемы высокой биологической продуктивности – 1–2 кг/м2 в год. Это липово-дубовые леса, прибрежные заросли рогоза или тростника на озере, посевы кукурузы и многолетних трав при орошении и удобрении высокими дозами минеральных удобрений.
3. Экосистемы умеренной биологической продуктивности – 0,25–1 кг/м2 в год. Такую продуктивность имеют многие посевы, сосновые и березовые леса, сенокосные луга и степи, заросшие водными растениями озера, ʼʼморские лугаʼʼ из водорослей в Японском море.
4. Экосистемы низкой биологической продуктивности – менее 0,25 кг/м2 в год. Это арктические пустыни островов Северного Ледовитого океана, тундры, полупустыни Прикаспия, вытоптанные скотом степные пастбища с низким и редким травостоем, горные степи, которые развиваются на почвах толщиной не более 5 см и состоят из растений-камнелюбов, покрывающих поверхность почвы на 20–40%. Такая же низкая продуктивность и у большей части морских экосистем.
Средняя продуктивность экосистем Земли не превышает 0,3 кг/м2 в год, так как на планете преобладают малопродуктивные экосистемы пустынь и океанов.
На рис. 50 и 51 приведены карты первичной продуктивности на суше и в океане (использован показатель – масса сухого вещества, ĸᴏᴛᴏᴩᴏᴇ в 2.5-3 раза ниже сырой биомассы).
Биологическая продуктивность экосистемы отличается от запаса биомассы. Некоторые организмы в экосистеме живут много лет (деревья, крупные животные), и их биомасса переходит из года в год как некоторый капитал. На рис. 52 показано соотношение запаса биомассы и биологической продуктивности в некоторых экосистемах.
На рис. 52 показано соотношение запаса биомассы и биологической продуктивности в некоторых экосистемах.
Биомасса леса велика за счёт многолетних частей деревьев – стволов, веток, корней. По этой причине ежегодный прирост биологической продукции – новые листья, молодые веточки и корни, очередное годичное кольцо древесины и травяной покров – в 30–50 раз меньше, чем запас биомассы. На лугу запас биомассы значительно меньше, и он образован в основном корнями, живущими в почве по несколько лет, и корневищами растений. Он больше биологической продуктивности только в 3–5 раз.
В однолетних агроценозах (посевах культурных растений) биологическая продуктивность и запас биомассы практически равны, так как урожай надземных частей растений (и подземных, в случае если это корнеплоды) убирают, а пожнивные остатки ржи или пшеницы запахивают в почву, где они к весне перегнивают. Как в луговой экосистеме, так и в экосистеме поля длительность жизни многочисленных почвенных беспозвоночных измеряется неделями и месяцами. Их биологическая продуктивность либо равна запасу биомассы, либо больше. Водоросли и мелкие беспозвоночные животные в водоемах живут по несколько дней или недель и потому за лето дают несколько поколений. В каждый конкретный момент биомасса организмов в озере или пруде меньше, чем их биологическая продукция за вегетационный сезон.
Их биологическая продуктивность либо равна запасу биомассы, либо больше. Водоросли и мелкие беспозвоночные животные в водоемах живут по несколько дней или недель и потому за лето дают несколько поколений. В каждый конкретный момент биомасса организмов в озере или пруде меньше, чем их биологическая продукция за вегетационный сезон.
В пресноводных и морских экосистемах за счёт того, что организмы зоопланктона живут дольше, чем водоросли фитопланктона, запас биомассы животных должна быть выше запаса биомассы растений.
Контрольные вопросы
1. Объясните различия первичной, вторичной, чистой и валовой биологической продукций.
2. Какова биологическая продуктивность разных экосистем?
3. Объясните различия понятий ʼʼбиологическая продуктивностьʼʼ и ʼʼзапас биомассыʼʼ.
4. Почему в водных экосистемах запас биомассы больше, чем биологическая продукция?
Справочный материал
На одного жителя России приходится 230 т первичной биологической продукции с экосистем площадью 11,5 га, что намного превышает потребность человека в количестве продукции, которая составляет примерно 40 т. При этом на Дальнем Востоке избыточной продукции много больше, чем в европейской части (соответственно 90% и 60%). При этом для обеспечения человека важно не только количество, но и качество биологической продукции. Таких ее фракций, как зерно, пастбищный корм, ценная древесина хвойных, пригодная для производства пиломатериалов, в большинстве районов России не хватает. Структура производства биологической продукции должна совершенствоваться — ϶ᴛᴏ задачи лесного и сельского хозяйства.
При этом на Дальнем Востоке избыточной продукции много больше, чем в европейской части (соответственно 90% и 60%). При этом для обеспечения человека важно не только количество, но и качество биологической продукции. Таких ее фракций, как зерно, пастбищный корм, ценная древесина хвойных, пригодная для производства пиломатериалов, в большинстве районов России не хватает. Структура производства биологической продукции должна совершенствоваться — ϶ᴛᴏ задачи лесного и сельского хозяйства.
Ответы | Тема 16. Продуктивность экосистем — Биология, 11 класс
1.
а) Количество биомассы, образующейся в экосистеме на единице площади или в единице объема биотопа за единицу времени: продукция, биомасса, чистая продукция, вторичная продукция (нужное подчеркните).
б) Часть продукции экосистемы, которая может быть использована самой экосистемой для ее развития или изъята человеком без ущерба для экосистемы: первичная продукция, вторичная продукция, чистая продукция сообщества (нужное подчеркните).
2.
а) Общее количество органического вещества всех живых организмов, накопившегося в данной экосистеме за предыдущий период ее существования, — биомасса экосистемы.
б) Биомасса органического вещества экосистемы, созданная автотрофными организмами, — первичная продукция.
в) Биомасса органического вещества экосистемы, заключенная в гетеротрофных организмах, — вторичная продукция.
3.
Потому, что некоторые элементы могут накапливаться в сферах Земли и исключаться из круговорота.
4.
Экосистема существует только за счёт направленного потока энергии.
5.
- б) Энергия и элементы питания совершают […]
- в) Биологическая продуктивность тропического леса выше […]
6.
- Прирост мышей составит 8 кг
- 8/0,02% = x/100%;
- x = 8 · 100 : 0,02 = 40 000 кг — урожай пшеницы.


7.
Биологическая продуктивность заливного луга будет выше, это связано с тем, что видовой состав луга представлен в основном однолетними растениями. Скорость их возобновления выше, чем в лиственном лесу.
Биология: Продуктивность экосистем
1. а) Количество биомассы, образующейся в экосистеме на единице площади или в единице объема биотопа за единицу времени: продукция, биомасса, чистая продукция, вторичная продукция (нужное подчеркните).
б) Часть продукции экосистемы, которая может быть использована самой экосистемой для ее развития или изъята человеком без ущерба для экосистемы: первичная продукция, вторичная продукция, чистая продукция сообщества (нужное подчеркните).
2. Закончите предложения, вписав соответствующие термины или понятия.
а) Общее количество органического вещества всех живых организмов, накопившегося в данной экосистеме за предыдущий период ее существования, — биомасса экосистемы.
б) Биомасса органического вещества экосистемы, созданная автотрофными организмами, — первичная продукция.
в) Биомасса органического вещества экосистемы, заключенная в гетеротрофных организмах, — вторичная продукция.
3. Как вы думаете, почему круговорот веществ в экосистеме не бывает полностью замкнутым?
Потому, что некоторые элементы могут накапливаться в сферах Земли и исключаться из круговорота.
4. Почему можно говорить о круговороте веществ, но нельзя говорить о круговороте энергии в экосистеме?
Экосистема существует только за счёт направленного потока энергии.
5. Выберите верные утверждения (буквы с правильными ответами обведите кружком).
б) + Энергия и элементы питания совершают […]
в) + Биологическая продуктивность тропического леса выше […]
6. На одном пшеничном поле площадью 10 га за лето мыши съели 80 кг зерна. Рассчитайте урожай пшеницы, если известно, что прирост биомассы мышей к концу лета составил 0,02% урожая. Переход энергии с одного трофического уровня на другой осуществляется в соответствии с правилом Линдемана.
На одном пшеничном поле площадью 10 га за лето мыши съели 80 кг зерна. Рассчитайте урожай пшеницы, если известно, что прирост биомассы мышей к концу лета составил 0,02% урожая. Переход энергии с одного трофического уровня на другой осуществляется в соответствии с правилом Линдемана.
- прирост мышей составит 8 кг
- 8/0,02% = x/100%; x 8 · 100 : 0,02 = 40 000 кг — урожай пшеницы.
7. Сравните биологическую продуктивность двух экосистем: лиственного леса и заливного луга. Как вы думаете, в какой экосистеме биологическая продуктивность будет выше? Почему? Как это можно проверить?
Биологическая продуктивность заливного луга будет выше, это связано с тем, что видовой состав луга представлен в основном однолетними растениями. Скорость их возобновления выше, чем в лиственном лесу.
Продуктивность сообщества
К основным показателям биогеоценоза относят:
· Видовой состав
· Видовое разнообразие
· Биомассу
·
Продуктивность сообщества.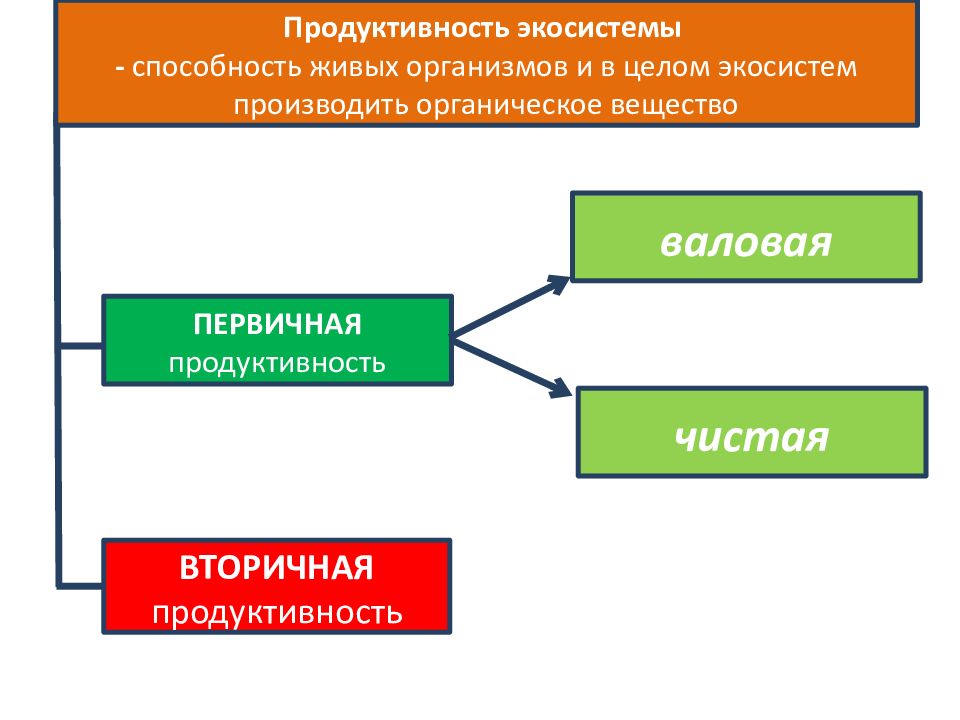
Органическое вещество и энергия переходят от одного уровня к другому. Вещество и энергия необходимы для роста и развития живого организма.
Природное вещество – это любое химическое соединение или элемент, возникающее в ходе спонтанно идущих химических реакций и физических процессов, которое включается в природный круговорот веществ.
Если разобрать все живое вещество планеты, то на долю кислорода приходиться 70%, углерода 17%, водорода 9%, азота 2%, а остальные 2% приходятся на другие химические элементы.
К элементам, которые обязательно входят в состав живого вещества относят: водород, углерод, азот, кислород, фосфор, серу, хлор, йод, натрий, магний, калий, железо, цинк и другие. Однако из всех веществ 98% приходиться на долю кислорода, углерода, водорода и азота.
«Живые организмы… своим
дыханием, своим питанием, своим метаболизмом… непрерывной сменой поколений, фомируют миграцию химических элементов в биосфере».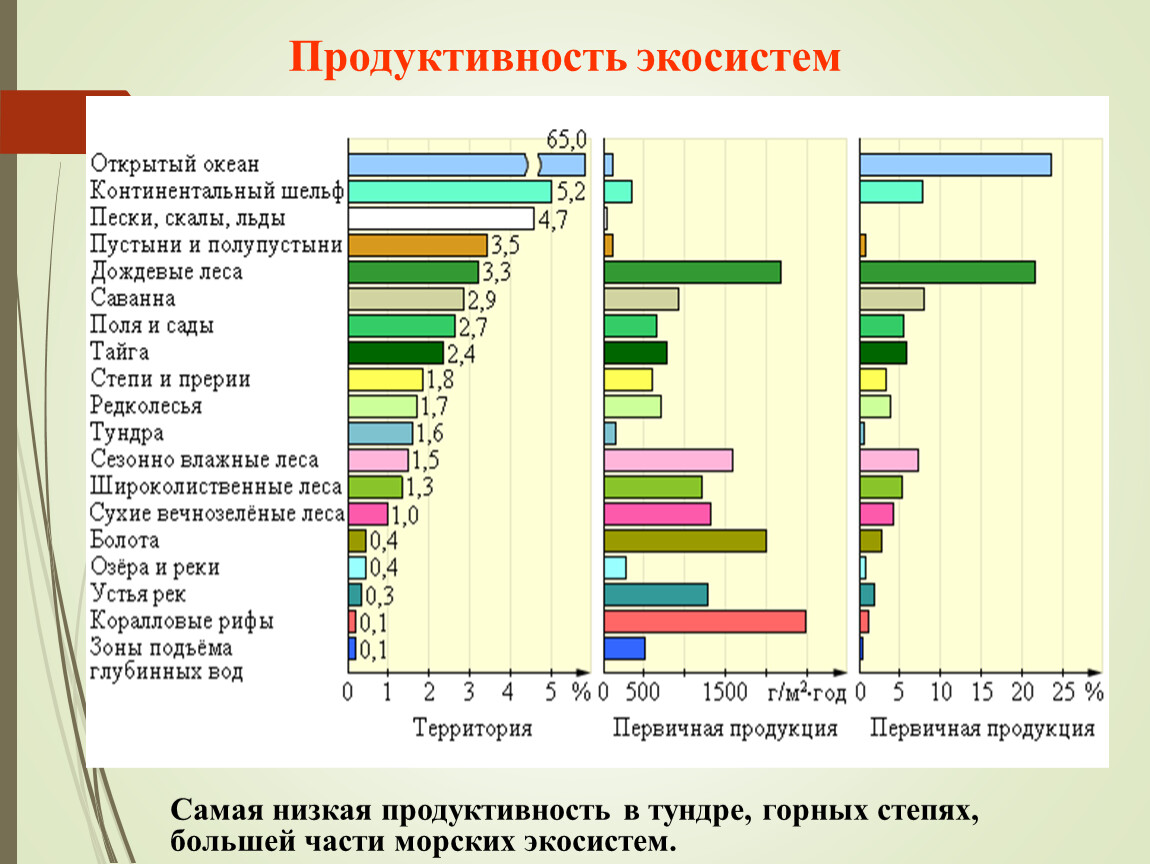
Одним из выдающихся естествоиспытателей, посвятивших себя изучению процессов, протекающих в биосфере, был академик Владимир Иванович Вернадский.
Владимир Иванович Вернадский
Согласно представлениям Вернадского, биосфера состоит из нескольких разнородных компонентов. Главный и основной из них — это живое вещество, совокупность всех живых организмов, населяющих Землю.
Вернадский писал, что «живое вещество в биосфере играет основную роль и по своей мощности, по своей интенсивности и непрерывности во времени, не может быть сравнимо ни с какой геологической силой».
Так совокупность растений образуют (фитобиомассу), а совокупность животных (зообиомассу).
Биомасса — это совокупная масса растительных и животных организмов, присутствующих в биогеоценозе, определённого размера или уровня.
Подсчитано, что биомасса суши составляет примерно 300 миллиарда тонн. При этом организмы континентальной
части образуют фитомассу суши, она
составляет 97% от всей биомассы Земли.
И зоомассу суши (животные и микроорганизмы), которая составляет —3% от всей биомассы.
Большая часть биомассы Земли сосредоточена в лесах. На суше преобладает масса растений.
Общая биомасса населения Мирового океана (без микрофлоры ─ бактерий и простейших) оценивается величиной в 35-38 млрд. т, из них 30-35% составляет фитомасса океана и 65-70% ─ зоомасса океана.
Живые организмы сообщества растут, размножаются, и погибают. Однако за счет роста и размножения биомасса сообщества постоянно возобновляется и пополняется.
Способность организмов создавать (продуцировать) новую биомассу называется продуктивностью.
Биологическая продуктивность – это образование биомассы организмами, выражаемое потоками органического вещества и его химической энергии на единицу площади за единицу времени (например, год).
Для того чтобы узнать
продуктивность, необходимо рассчитать прирост биомассы.
Прирост биомассы — это количество живого вещества, на которое увеличивается особь, популяция, сообщество какой-либо территории за единицу времени.
Например, если взять популяцию зайца-русака и подсчитать их общий вес и количество особей, а затем произвести те же расчёты в конце года, то можно обнаружить что за год их число и общий вес может увеличиться.
Количество, на которое увеличилась популяция зайца за год и есть прирост биомассы популяции.
Биологическая продуктивность существующих экосистем проявляется во многих формах.
Например, продуктивность леса ─ количество различных ресурсов, произведенных лесом за определенный период на единице площади.
Продуктивность леса определяется по отдельным ее составляющим – насаждению и древостою.
Продуктивность леса бывает:
· биологическая ─ количество биомассы, произведенной лесом за определенный период на единице площади;
·
древесная ─ количество древесины, произведенной насаждением за определённый период на единице
площади.
· комплексная ─ совокупная биологическая и экологическая продуктивность участка леса;
Продуктивность водоёмов — это количество органического вещества, производимого в водоёме его обитателями за единицу времени (обычно за год) на единицу площади дна или на единицу объёма воды.
Общая годовая биологическая продуктивность в Мировом океане оценивается более чем 1300 млрд. т, в том числе более 1200 млрд. т дают водоросли и 40-50 млрд. т — животные.
Океан гораздо беднее жизнью, чем суша, его биомасса во много раз меньше. Однако скорость прироста биомассы (оборот) намного больше, в океанах. Отсюда и большая продуктивность.
Чаще всего рассчитывают рыбную продуктивность водоёмов.
Под рыбной продуктивностью надо понимать численность (запасы) общего рыбного стада (промысловых и непромысловых рыб) в нем.
Под промысловой продукцией — количество вылавливаемой рыбы.
Годовая рыбная продукция менее
30 кг с одного гектара считается низкой; 30—60 кг с гектара — средней и более
60 кг с одного гектара — высокой.
Рыбопродуктивность водоёма, как и любая другая продуктивность не является величиной постоянной и изменяется в зависимости от условий.
Изначально, озеро делят на районы (первый район, второй район, третий район, четвёртый район и пятый район, например). В каждом районе забрасывают невод.
Делают несколько закидок в разных районах.
Затем подсчитывают количество выловленных экземпляров рыб. И взвешивают данное количество.
Затем все полученные данные суммируют, сравнивают с прошлым годом, например, и получают необходимые данные о рыбопродуктивности.
Таким образом, для того что бы рассчитать продуктивность сообщества необходимо суммировать прирост биомассы видов (растений и животных), за какой-то промежуток времени.
Так, чтобы узнать продуктивность водоёма, необходимо суммировать прирост зоофауны, прирост фитомассы и прирост других обитателей водоёма.
Заметим, что суммируем не
общее количество организмов, а именно их прирост. В результате чего
получаем общее количество, то есть продуктивность водоёма.
В результате чего
получаем общее количество, то есть продуктивность водоёма.
Продуцирование биомассы – это непрерывный процесс. Так, как и растения и животные непрерывно производят себя в потомстве.
Для устойчивого состояния сообщества, продукция трофического уровня должна покрывать пищевые потребности организмов следующего уровня.
Другими словами… организмам необходимо постоянно воспроизводить себя в потомстве. В противном случае запас биологического вещества снизиться из-за выедания.
Чем выше трофический уровень, тем ниже энергия, общая биомасса и численность составляющих её организмов. Тоже происходит и с продукцией, она убывает при переходе от низших трофических уровней к высшим.
Так на первом трофическом
уровне как вы знаете находятся растения (продуценты), которые в процессе
фотосинтеза образуют органические вещества и кислород. То есть те органические
вещества, из которых и стояться все биологические ткани, составляющие общую
биомассу.
В процессе дыхания растений, часть этих веществ окисляется.
В результате чего образуется углекислый газ, вода и энергия. Таким образом на дыхание растение тратит около 70% образовавшихся в процессе фотосинтеза веществ.
А та часть продукции, которая не израсходована “на дыхание”, представляет собой величину прироста растений и именно эта продукция потребляется консументами и редуцентами.
Таким образом выделяют разные уровни производства органического вещества.
Первичная продукция ─ это скорость образования органического вещества (биомассы) – автотрофами, за единицу времени.
Вторичная продукция ─ это скорость продуцирования биомассы гетеротрофами.
Так гетеротрофы, увеличивают свою массу за счет первичной продукции, т.е. используют ранее созданную продукцию.
Если рассмотреть прирост биомассы к уже имеющей массе, то получаются такие показатели:
За год прирост биомассы
древесной растительности лесов составляет 1,8 %.
Растительности лугов, степей, пашни — 67 %, комплекс растений озёр и рек — 1400 %, морского фитопланктона — 1500 %.
Как вы видите, самый большой прирост биомассы составляет морской фитопланктон.
Быстрый оборот фитомассы океана (1-3 суток) происходит за счет интенсивного деления, быстрого роста и кратковременного существования фитопланктона.
А вот полное обновление фитомассы суши осуществляется за 50 лет и более.
Ежегодно в биосфере в процессе фотосинтеза образуется около 150 млрд тонн сухого органического вещества.
На суше самыми продуктивными являются тропические и субтропические леса.
Низкая продуктивность растений характерна для открытого океана, пустынь и тундры.
Биологическая продуктивность океана
Продуктивность океана в значительной степени относится к производству органического вещества с помощью « фитопланктона », растений, взвешенных в океане, большинство из которых одноклеточные. Фитопланктон — это « фотоавтотрофов », собирающих свет для преобразования неорганического углерода в органический, и они поставляют этот органический углерод различным « гетеротрофам », организмам, которые получают свою энергию исключительно от дыхания органического вещества.Гетеротрофы открытого океана включают бактерии, а также более сложные одноклеточные и многоклеточные « зоопланктон » (плавающие животные), « нектон » (плавучие организмы, включая рыб и морских млекопитающих) и « бентос ». «(сообщество организмов на морском дне).
Фитопланктон — это « фотоавтотрофов », собирающих свет для преобразования неорганического углерода в органический, и они поставляют этот органический углерод различным « гетеротрофам », организмам, которые получают свою энергию исключительно от дыхания органического вещества.Гетеротрофы открытого океана включают бактерии, а также более сложные одноклеточные и многоклеточные « зоопланктон » (плавающие животные), « нектон » (плавучие организмы, включая рыб и морских млекопитающих) и « бентос ». «(сообщество организмов на морском дне).
Множество вложенных циклов углерода, связанных с продуктивностью океана, раскрываются следующими определениями (Bender et al. 1987) (Рисунок 1). «Валовая первичная продукция» ( GPP ) относится к общей скорости производства органического углерода автотрофами , в то время как « дыхание » относится к энергоемкому окислению органического углерода обратно в диоксид углерода. «Чистая первичная продукция» ( НПП ) — ГПЗ минус собственных темпов автотрофов дыхания ; таким образом, это скорость, с которой полный метаболизм фитопланктона производит биомассу. « Вторичная продукция » ( SP ) обычно относится к скорости роста гетеротрофной биомассы. Лишь небольшая часть органического вещества, поглощенного гетеротрофными организмами, используется для роста, большая часть из которых вдыхает обратно до растворенного неорганического углерода и питательных веществ, которые могут повторно использоваться автотрофами .Следовательно, SP в океане мало по сравнению с NPP . Рыболовство полагается на SP ; таким образом, они зависят как от NPP , так и от эффективности, с которой органическое вещество перемещается по пищевой сети (т. е. от соотношения SP / NPP ). «Чистая продукция экосистемы» ( NEP ) составляет GPP минус дыхание всеми организмами в экосистеме.
«Чистая первичная продукция» ( НПП ) — ГПЗ минус собственных темпов автотрофов дыхания ; таким образом, это скорость, с которой полный метаболизм фитопланктона производит биомассу. « Вторичная продукция » ( SP ) обычно относится к скорости роста гетеротрофной биомассы. Лишь небольшая часть органического вещества, поглощенного гетеротрофными организмами, используется для роста, большая часть из которых вдыхает обратно до растворенного неорганического углерода и питательных веществ, которые могут повторно использоваться автотрофами .Следовательно, SP в океане мало по сравнению с NPP . Рыболовство полагается на SP ; таким образом, они зависят как от NPP , так и от эффективности, с которой органическое вещество перемещается по пищевой сети (т. е. от соотношения SP / NPP ). «Чистая продукция экосистемы» ( NEP ) составляет GPP минус дыхание всеми организмами в экосистеме. Значение NEP зависит от границ, определенных для экосистемы.Если рассматривать освещенную солнцем поверхность океана до уровня освещенности 1% (« эвфотическая зона ») в течение всего года, то NEP эквивалентно частиц органического углерода , погружающимся в темные глубины океана. плюс растворенный органический углерод , циркулирующий из эвфотической зоны . В этом случае NEP также часто называют « экспортного производства » (или «нового производства» (Dugdale & Goering 1967), как обсуждается ниже).Напротив, NEP для всего океана, включая его мелкие отложения, примерно эквивалентен медленному захоронению органического вещества в отложениях за вычетом скорости поступления органического вещества с континентов.
Значение NEP зависит от границ, определенных для экосистемы.Если рассматривать освещенную солнцем поверхность океана до уровня освещенности 1% (« эвфотическая зона ») в течение всего года, то NEP эквивалентно частиц органического углерода , погружающимся в темные глубины океана. плюс растворенный органический углерод , циркулирующий из эвфотической зоны . В этом случае NEP также часто называют « экспортного производства » (или «нового производства» (Dugdale & Goering 1967), как обсуждается ниже).Напротив, NEP для всего океана, включая его мелкие отложения, примерно эквивалентен медленному захоронению органического вещества в отложениях за вычетом скорости поступления органического вещества с континентов.
Рис. 1
Производительность в поверхностном океане, определения, используемые для ее описания, и ее связь с круговоротом питательных веществ. Синий цикл для «чистого экосистемного производства» (НЭП) (то есть «нового» или «экспортного» производства) включает в себя «новое» поступление питательных веществ из недр океана, его поглощение путем автотрофного роста фитопланктона, упаковку в крупные частицы гетеротрофными пастбищными организмами. , и опускание органического вещества из поверхности океана.Красный цикл иллюстрирует судьбу большинства органических веществ, образующихся на поверхности океана, которые должны вдыхаться гетеротрофными организмами для удовлетворения своих потребностей в энергии, тем самым высвобождая питательные вещества обратно в поверхностные воды, где они могут быть поглощены фитопланктоном один раз. снова в качестве топлива для «регенерированной продукции». Зеленый цикл представляет собой внутреннее дыхание самого фитопланктона, то есть собственное использование продуктов фотосинтеза не для роста, а для других целей.Эти вложенные циклы объединяются, чтобы дать (1) «валовую первичную продукцию» (GPP), представляющую валовой фотосинтез, и (2) «чистую первичную продукцию» (NPP), которая представляет производство биомассы фитопланктона, которое составляет основу пищевой сети, плюс гораздо меньшую скорость вывоза органического вещества с поверхности. В то время как новые поставки питательных веществ и экспортное производство в конечном итоге связаны балансом массы, возможны дисбалансы в небольших пространственно-временных масштабах, что позволяет кратковременно накапливать биомассу.
, и опускание органического вещества из поверхности океана.Красный цикл иллюстрирует судьбу большинства органических веществ, образующихся на поверхности океана, которые должны вдыхаться гетеротрофными организмами для удовлетворения своих потребностей в энергии, тем самым высвобождая питательные вещества обратно в поверхностные воды, где они могут быть поглощены фитопланктоном один раз. снова в качестве топлива для «регенерированной продукции». Зеленый цикл представляет собой внутреннее дыхание самого фитопланктона, то есть собственное использование продуктов фотосинтеза не для роста, а для других целей.Эти вложенные циклы объединяются, чтобы дать (1) «валовую первичную продукцию» (GPP), представляющую валовой фотосинтез, и (2) «чистую первичную продукцию» (NPP), которая представляет производство биомассы фитопланктона, которое составляет основу пищевой сети, плюс гораздо меньшую скорость вывоза органического вещества с поверхности. В то время как новые поставки питательных веществ и экспортное производство в конечном итоге связаны балансом массы, возможны дисбалансы в небольших пространственно-временных масштабах, что позволяет кратковременно накапливать биомассу.
В морской среде нет скоплений живой биомассы, которые можно сравнить с лесами и лугами на суше (Sarmiento & Bender 1994).Тем не менее, биология океана ответственна за хранение большего количества углерода вдали от атмосферы, чем земная биосфера (Broecker 1982). Это достигается за счет погружения органического вещества с поверхности океана в глубь океана до того, как оно возвращается к растворенному неорганическому углероду и растворенным питательным веществам в результате бактериального разложения. Океанографы часто называют этот процесс «биологическим насосом », поскольку он выкачивает углекислый газ (CO 2 ) из поверхности океана и атмосферы в огромные глубины океана (Volk & Hoffert 1985).
Лишь небольшая часть органического вещества, образующегося на поверхности океана, будет экспортирована в глубины океана. Из органического вещества, производимого фитопланктоном ( NPP ), большая часть вернулась в растворенные неорганические формы на поверхности океана и, таким образом, повторно использовалась для использования фитопланктоном (Eppley & Peterson 1979) (Рисунок 1). Большинство клеток фитопланктона слишком малы, чтобы тонуть по отдельности, поэтому погружение происходит только после того, как они объединяются в более крупные частицы или упаковываются в « фекальных гранул » зоопланктона .Остатки зоопланктона также достаточно велики, чтобы утонуть. Хотя затопление является относительно редкой судьбой для любой конкретной частицы на поверхности океана, биомасса и органическое вещество не накапливаются в поверхностном океане, поэтому экспортирует органического вещества путем погружения — это окончательная судьба всех питательных веществ, которые попадают в океан. поверхностный океан в растворенной форме — за исключением того, что (1) растворенные питательные вещества могут быть возвращены неиспользованными внутрь посредством циркуляции в некоторых полярных регионах (см. ниже), и (2) циркуляция также переносит растворенное органическое вещество из поверхностного океана в интерьер, важный процесс (Hansell et al .
Большинство клеток фитопланктона слишком малы, чтобы тонуть по отдельности, поэтому погружение происходит только после того, как они объединяются в более крупные частицы или упаковываются в « фекальных гранул » зоопланктона .Остатки зоопланктона также достаточно велики, чтобы утонуть. Хотя затопление является относительно редкой судьбой для любой конкретной частицы на поверхности океана, биомасса и органическое вещество не накапливаются в поверхностном океане, поэтому экспортирует органического вещества путем погружения — это окончательная судьба всех питательных веществ, которые попадают в океан. поверхностный океан в растворенной форме — за исключением того, что (1) растворенные питательные вещества могут быть возвращены неиспользованными внутрь посредством циркуляции в некоторых полярных регионах (см. ниже), и (2) циркуляция также переносит растворенное органическое вещество из поверхностного океана в интерьер, важный процесс (Hansell et al . 2009), к которым мы не будем обращаться дальше. По мере того, как органическое вещество оседает через внутреннюю часть океана и на морское дно, оно почти полностью разлагается обратно на растворенные химические вещества (Emerson & Hedges 2003, Martin et al. 1987). Такая высокая эффективность разложения объясняется тем фактом, что организмы, осуществляющие разложение, полагаются на него как на единственный источник химической энергии; в большей части открытого океана гетеротрофов оставляют после себя только органическое вещество, которое является слишком химически стойким, чтобы его разложение стоило вложений.В целом, лишь небольшая часть (обычно намного меньше 1%) органического углерода из АЭС в эвфотической зоне выживает, чтобы быть захороненной в глубоководных отложениях.
2009), к которым мы не будем обращаться дальше. По мере того, как органическое вещество оседает через внутреннюю часть океана и на морское дно, оно почти полностью разлагается обратно на растворенные химические вещества (Emerson & Hedges 2003, Martin et al. 1987). Такая высокая эффективность разложения объясняется тем фактом, что организмы, осуществляющие разложение, полагаются на него как на единственный источник химической энергии; в большей части открытого океана гетеротрофов оставляют после себя только органическое вещество, которое является слишком химически стойким, чтобы его разложение стоило вложений.В целом, лишь небольшая часть (обычно намного меньше 1%) органического углерода из АЭС в эвфотической зоне выживает, чтобы быть захороненной в глубоководных отложениях.
Продуктивность прибрежных экосистем часто отличается от продуктивности открытого океана. Вдоль побережья морское дно неглубокое, и иногда солнечный свет может проникать через толщу воды на дно, что позволяет живущим на дне (« бентосных ») организмам фотосинтезировать. Кроме того, опускающееся органическое вещество задерживается морским дном, где оно поддерживает процветающие сообщества бентосной фауны, в процессе рециркулируя обратно в растворенные питательные вещества, которые затем немедленно становятся доступными для первичной продукции. Близость к суше и ее источникам питательных веществ, перехват опускающегося органического вещества на мелководье морского дна и склонность к прибрежному апвеллингу — все это приводит к высокопродуктивным экосистемам. Здесь мы в основном обращаемся к продуктивности огромного открытого океана; тем не менее, многие из тех же концепций, хотя и в измененной форме, применимы к прибрежным системам.
Кроме того, опускающееся органическое вещество задерживается морским дном, где оно поддерживает процветающие сообщества бентосной фауны, в процессе рециркулируя обратно в растворенные питательные вещества, которые затем немедленно становятся доступными для первичной продукции. Близость к суше и ее источникам питательных веществ, перехват опускающегося органического вещества на мелководье морского дна и склонность к прибрежному апвеллингу — все это приводит к высокопродуктивным экосистемам. Здесь мы в основном обращаемся к продуктивности огромного открытого океана; тем не менее, многие из тех же концепций, хотя и в измененной форме, применимы к прибрежным системам.
Фитопланктон требует набора химикатов, и те, которые потенциально могут быть дефицитными в поверхностных водах, обычно называют «питательными веществами». Кальций является примером элемента, который быстро ассимилируется примерно планктоном (для производства «твердых частей» карбоната кальция), но обычно не считается питательным веществом из-за его равномерно высокой концентрации в морской воде. Растворенный неорганический углерод , который является сырьем для производства органического углерода путем фотосинтеза , также присутствует в большом количестве и поэтому обычно не включается в список питательных веществ.Однако его кислая форма растворенного CO 2 часто имеет достаточно низкие концентрации, чтобы повлиять на рост по крайней мере фитопланктона .
Растворенный неорганический углерод , который является сырьем для производства органического углерода путем фотосинтеза , также присутствует в большом количестве и поэтому обычно не включается в список питательных веществ.Однако его кислая форма растворенного CO 2 часто имеет достаточно низкие концентрации, чтобы повлиять на рост по крайней мере фитопланктона .
Широко важные питательные вещества включают азот (N), фосфор (P), железо (Fe) и кремний (Si). Похоже, что требования к азоту и фосфору среди фитопланктона относительно одинаковы. В начале 1900-х годов океанограф Альфред Редфилд обнаружил, что планктона формируют свою биомассу со стехиометрическими отношениями C: N: P ~ 106: 16: 1, которые мы теперь называем отношениями Редфилда (Redfield 1958).Как заметил Редфилд, растворенный N: P в глубинах океана близко к соотношению 16: 1 биомассы планктона, и ниже мы будем утверждать, что планктон накладывает это соотношение на глубину, а не наоборот. Железо содержится в биомассе только в следовых количествах, но оно используется в организмах для различных основных целей, и за последние 25 лет стало ясно, что дефицит железа часто ограничивает или влияет на продуктивность в открытом океане, особенно в тех регионах, где высокий уровень содержания железа. N и -P глубокие воды быстро выносятся на поверхность (Martin & Fitzwater 1988).Продолжаются исследования, чтобы понять роль других микроэлементов в продуктивности (Morel et al. 2003). Кремний является питательным веществом только для определенных таксонов планктона — диатомовых водорослей ( автотрофных фитопланктона ), силикофлаггеллятов и радиолярий ( гетеротрофных зоопланктона ), которые используют его для образования твердых частей опала. Однако типичное преобладание диатомовых водорослей в содержащих кремний водах и тенденция связанного с диатомовыми водорослей органического вещества опускаться из поверхности океана делают доступность Si основным фактором в более широкой экологии и биогеохимии поверхностных вод.
Железо содержится в биомассе только в следовых количествах, но оно используется в организмах для различных основных целей, и за последние 25 лет стало ясно, что дефицит железа часто ограничивает или влияет на продуктивность в открытом океане, особенно в тех регионах, где высокий уровень содержания железа. N и -P глубокие воды быстро выносятся на поверхность (Martin & Fitzwater 1988).Продолжаются исследования, чтобы понять роль других микроэлементов в продуктивности (Morel et al. 2003). Кремний является питательным веществом только для определенных таксонов планктона — диатомовых водорослей ( автотрофных фитопланктона ), силикофлаггеллятов и радиолярий ( гетеротрофных зоопланктона ), которые используют его для образования твердых частей опала. Однако типичное преобладание диатомовых водорослей в содержащих кремний водах и тенденция связанного с диатомовыми водорослей органического вещества опускаться из поверхности океана делают доступность Si основным фактором в более широкой экологии и биогеохимии поверхностных вод.
Солнечный свет является основным источником энергии — прямо или косвенно — почти для всего живого на Земле, в том числе в глубоких океанских глубинах. Однако свет поглощается и рассеивается так, что очень небольшая его часть проникает ниже глубины ~ 80 м (до 150 м в наименее продуктивных субтропических регионах, и до 10 м в высокопродуктивных и прибрежных регионах) (рис. ). Таким образом, фотосинтез в значительной степени ограничен верхними светопроницаемыми слоями океана. Более того, на большей части площади океана, включая тропики, субтропики и умеренную зону, поглощение солнечного света приводит к тому, что поверхностные воды становятся намного теплее, чем глубинный океан, находящийся под ним, причем последний наполняется водой, которая опускалась с поверхности в глубину. высокие широты.Теплая вода более плавучая, чем холодная, что заставляет верхний залитый солнцем слой плавать в более плотных глубинах океана с переходом между ними, известным как «пикноклин» (для «градиента плотности») или «термоклин» (вертикальный градиент температуры. это приводит к стратификации плотности на большей части океана, рис. 2). Ветер или другой источник энергии необходим для перемешивания через пикноклин, поэтому перенос воды с растворенными в ней химическими веществами между освещенной солнцем поверхностью и темным внутренним пространством является вялым.Этот двойной эффект света на фотосинтез и плавучесть морской воды имеет решающее значение для успеха фитопланктона океана . Если бы в океане не было тонкого плавучего поверхностного слоя, перемешивание уносило бы водоросли за пределы света и, таким образом, от их источника энергии на большую часть времени. Вместо почти нейтрально плавучих одноклеточных водорослей в открытом океане могут доминировать более крупные, положительно плавучие фотосинтезирующие организмы (например, пелагические водоросли). Помимо этого гипотетического случая, хотя жизнеспособные клеток фитопланктона обнаружены (хотя и в низких концентрациях) в более глубоких водах, фотосинтез ограничивает рост активного фитопланктона верхними слоями океана, в то время как стратификация плотности верхнего слоя океана предотвращает их смешивание вниз в темную бездну.
это приводит к стратификации плотности на большей части океана, рис. 2). Ветер или другой источник энергии необходим для перемешивания через пикноклин, поэтому перенос воды с растворенными в ней химическими веществами между освещенной солнцем поверхностью и темным внутренним пространством является вялым.Этот двойной эффект света на фотосинтез и плавучесть морской воды имеет решающее значение для успеха фитопланктона океана . Если бы в океане не было тонкого плавучего поверхностного слоя, перемешивание уносило бы водоросли за пределы света и, таким образом, от их источника энергии на большую часть времени. Вместо почти нейтрально плавучих одноклеточных водорослей в открытом океане могут доминировать более крупные, положительно плавучие фотосинтезирующие организмы (например, пелагические водоросли). Помимо этого гипотетического случая, хотя жизнеспособные клеток фитопланктона обнаружены (хотя и в низких концентрациях) в более глубоких водах, фотосинтез ограничивает рост активного фитопланктона верхними слоями океана, в то время как стратификация плотности верхнего слоя океана предотвращает их смешивание вниз в темную бездну. Таким образом, большая часть биомассы открытого океана, включая фитопланктона , зоопланктона и нектон , находится в пределах ~ 200 м от поверхности океана.
Таким образом, большая часть биомассы открытого океана, включая фитопланктона , зоопланктона и нектон , находится в пределах ~ 200 м от поверхности океана.
Рис. 2
Типичные условия в субтропическом океане, о чем свидетельствуют данные, собранные на станции Бермудских Атлантических временных рядов в июле 2008 года. Термоклин (вертикальный градиент температуры) стратифицирует верхний слой воды. Во время этого конкретного занятия станции поверхностный слой, смешанный с мелким ветром, четко не определен, предположительно из-за сильной инсоляции и отсутствия ветра, что обеспечивало непрерывную стратификацию на всем протяжении до поверхности.Очень мало солнечного света проникает на глубину более ~ 100 м. Новое поступление основных питательных веществ N и P ограничено медленным перемешиванием в верхнем термоклине (здесь показан только нитрат азота, NO 3 — ). В пределах верхней эвфотической зоны медленное поступление питательных веществ полностью потребляется фитопланктоном в процессе своего роста.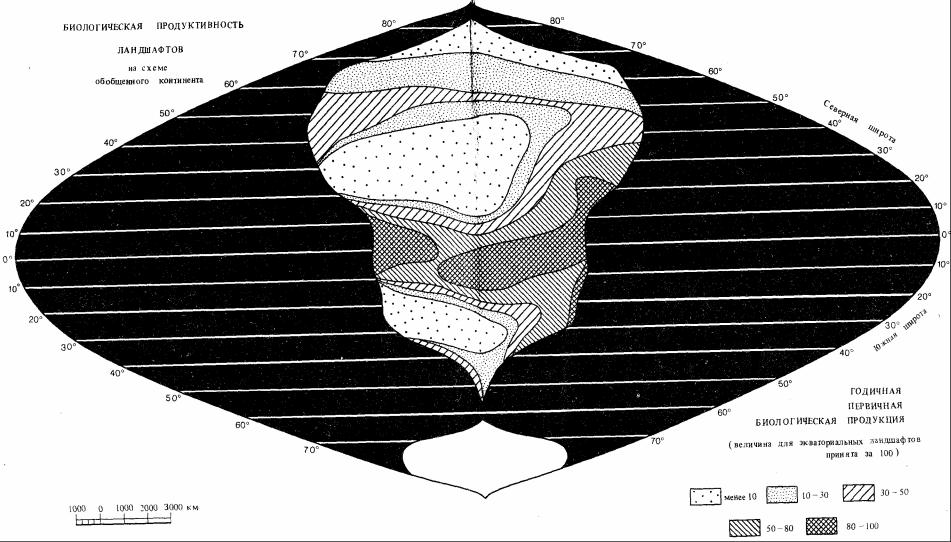 Этот рост приводит к накоплению твердых частиц органического углерода на поверхности океана, часть которых вдыхается бактериями, зоопланктоном и другими гетеротрофами, а часть экспортируется в виде тонущего материала.Глубокий максимум хлорофилла (DCM) происходит в месте контакта, где есть достаточно света для фотосинтеза и при этом значительный приток питательных веществ снизу. DCM не следует строго интерпретировать как максимум глубины биомассы фитопланктона, поскольку фитопланктон в DCM имеет особенно высокую внутреннюю концентрацию хлорофилла. Приведенные здесь данные доступны Бермудскому институту наук об океане (http://bats.bios.edu) и Бермудскому проекту биооптики (http://www.icess.ucsb.edu/bbop/).
Этот рост приводит к накоплению твердых частиц органического углерода на поверхности океана, часть которых вдыхается бактериями, зоопланктоном и другими гетеротрофами, а часть экспортируется в виде тонущего материала.Глубокий максимум хлорофилла (DCM) происходит в месте контакта, где есть достаточно света для фотосинтеза и при этом значительный приток питательных веществ снизу. DCM не следует строго интерпретировать как максимум глубины биомассы фитопланктона, поскольку фитопланктон в DCM имеет особенно высокую внутреннюю концентрацию хлорофилла. Приведенные здесь данные доступны Бермудскому институту наук об океане (http://bats.bios.edu) и Бермудскому проекту биооптики (http://www.icess.ucsb.edu/bbop/).
В то же время наличие тонкого плавучего поверхностного слоя вступает в сговор с другими процессами, накладывая ограничение по питательным веществам на продуктивность океана. Экспорт органического вещества на глубину истощает поверхность океана питательными веществами, заставляя питательные вещества накапливаться в глубоких водах, где нет света для фотосинтеза (рис. 2). Из-за разницы в плотности между поверхностными водами и глубоководным морем на большей части океана циркуляция океана может лишь очень медленно возвращать растворенные питательные вещества в эвфотическую зону .Вытесняя питательные вещества из освещенных солнцем плавучих поверхностных вод, продуктивность океана фактически ограничивает сама себя.
2). Из-за разницы в плотности между поверхностными водами и глубоководным морем на большей части океана циркуляция океана может лишь очень медленно возвращать растворенные питательные вещества в эвфотическую зону .Вытесняя питательные вещества из освещенных солнцем плавучих поверхностных вод, продуктивность океана фактически ограничивает сама себя.
Фитопланктон Ограничение роста традиционно интерпретировалось в контексте закона минимума Либиха, который гласит, что рост растений будет настолько большим, насколько позволяет наименее доступный ресурс, «ограничивающее питательное вещество», которое определяет продуктивность системы (de Баар 1994). Несмотря на то, что эта точка зрения сильна, взаимодействие между питательными веществами и между питательными веществами и светом также может контролировать продуктивность.Простой, но важный пример этого потенциала «совместного ограничения» исходит из полярных регионов, где наклонная солнечная инсоляция сочетается с глубоким перемешиванием поверхностных вод, что приводит к низкой доступности света. В таких средах более высокое содержание железа может повысить эффективность, с которой фитопланктона улавливают световую энергию (Мальдонадо и др. 1999, Sunda & Huntsman 1997). В более широком смысле утверждалось, что фитопланктон , как правило, должен стремиться к совместному ограничению всеми химическими веществами, которые им требуются, включая многие микроэлементы, содержащие металлы (Morel 2008).
В таких средах более высокое содержание железа может повысить эффективность, с которой фитопланктона улавливают световую энергию (Мальдонадо и др. 1999, Sunda & Huntsman 1997). В более широком смысле утверждалось, что фитопланктон , как правило, должен стремиться к совместному ограничению всеми химическими веществами, которые им требуются, включая многие микроэлементы, содержащие металлы (Morel 2008).
Биологическая продуктивность — обзор
VII.D Океаны: Цвет океана
В дополнение к измерению температуры поверхности океанов с помощью ведра ранние океанографы и моряки также использовали другой метод для характеристики океана: его внешний вид. Эта оценка часто была качественной, поскольку видимый цвет океана может варьироваться от темно-синего до различных оттенков зеленого, даже до коричневого и серого в прибрежных районах, нагруженных отложениями. Обнаружение Гольфстрима, которое было важно либо для использования его сильного течения для увеличения скорости плавания на пути из Америки в Европу, либо для того, чтобы избежать его на обратном пути, можно было выполнить, просто отметив изменение цвета воды. Одна из первых качественных оценок оптических свойств воды была сделана с использованием диска Секки . Диск Секки (изобретен отцом Пьетро Анджело Секки, 1818–1878 гг., Научным советником Папы Пинса IX) представляет собой диск шириной 20 см, полностью белый или цветной с чередующимися черными и белыми квадрантами. Диск (с грузом под ним для устойчивости) опускается в толщу воды по линии с отмеченными интервалами глубины. Глубина Секки , мера прозрачности воды, это глубина, на которой наблюдатель с поверхности больше не может различить диск.Самые высокие из когда-либо определенных глубин Секки наблюдались в полыньях в Антарктиде.
Одна из первых качественных оценок оптических свойств воды была сделана с использованием диска Секки . Диск Секки (изобретен отцом Пьетро Анджело Секки, 1818–1878 гг., Научным советником Папы Пинса IX) представляет собой диск шириной 20 см, полностью белый или цветной с чередующимися черными и белыми квадрантами. Диск (с грузом под ним для устойчивости) опускается в толщу воды по линии с отмеченными интервалами глубины. Глубина Секки , мера прозрачности воды, это глубина, на которой наблюдатель с поверхности больше не может различить диск.Самые высокие из когда-либо определенных глубин Секки наблюдались в полыньях в Антарктиде.
Измерение Секки показывает, что фундаментальным фактором, определяющим оптические характеристики водной толщи океана, является присутствие органических или неорганических частиц. (Второстепенным фактором является присутствие некоторых типов растворенных веществ.) Эти частицы рассеивают падающий солнечный свет и выборочно поглощают волны определенных длин. Затем рассеянный свет отражается вверх от поверхности воды.Этот конкретный рассеянный свет упоминается как уходящее воду сияние . Кажущийся цвет океанской воды на самом деле является уходящим от нее сиянием, воспринимаемым в диапазоне видимого света. Поглощение поступающего света фотосинтетическими пигментами (в первую очередь хлорофиллом) в фитопланктоне связано с концентрацией фитопланктона в эвфотической зоне , глубине, которая освещается солнечным светом. Сияние выходящей воды может быть количественно связано с концентрацией фотосинтетических пигментов и, таким образом, указывает на уровень биологической активности в поверхностных водах.
Затем рассеянный свет отражается вверх от поверхности воды.Этот конкретный рассеянный свет упоминается как уходящее воду сияние . Кажущийся цвет океанской воды на самом деле является уходящим от нее сиянием, воспринимаемым в диапазоне видимого света. Поглощение поступающего света фотосинтетическими пигментами (в первую очередь хлорофиллом) в фитопланктоне связано с концентрацией фитопланктона в эвфотической зоне , глубине, которая освещается солнечным светом. Сияние выходящей воды может быть количественно связано с концентрацией фотосинтетических пигментов и, таким образом, указывает на уровень биологической активности в поверхностных водах.
Измерение сияния выходящей воды из-за ее связи с фитопланктоном и, следовательно, основы океанического углеродного цикла и пищевой сети, поэтому представляло соблазнительную проблему для ученых, занимающихся дистанционным зондированием. Задача оказалась более сложной, чем дистанционное зондирование суши или облаков, по двум основным причинам: во-первых, поверхность океана намного темнее, т. Е. Гораздо менее ярка, чем поверхность суши или облака; и во-вторых, потому что поверхность океана настолько темнее, что соответственно больше влияние атмосферной поправки.Хотя небольшой просчет атмосферной коррекции будет иметь минимальное влияние на дистанционное зондирование земли или облаков, он существенно повлияет на расчет яркости выходящей воды, и это повлияет на любую оценку концентрации пигмента или фитопланктона.
Е. Гораздо менее ярка, чем поверхность суши или облака; и во-вторых, потому что поверхность океана настолько темнее, что соответственно больше влияние атмосферной поправки.Хотя небольшой просчет атмосферной коррекции будет иметь минимальное влияние на дистанционное зондирование земли или облаков, он существенно повлияет на расчет яркости выходящей воды, и это повлияет на любую оценку концентрации пигмента или фитопланктона.
VII.D.1 CZCS
Из-за этих факторов миссия CZCS на борту Nimbus 7 была названа миссией «доказательство концепции», призванной продемонстрировать возможность надежных измерений яркости уходящей воды и концентрации фотосинтетических пигментов. из космоса.Первоначальная миссия CZCS должна была продлиться только первый год миссии Nimbus 7. Однако CZCS проработала 8 лет, вернув почти 60 000 изображений поверхности океана и позволив создать первое полное изображение океанской биосферы в глобальном масштабе.
CZCS имел шесть спектральных полос (каналов), четыре из которых были посвящены цвету океана, каждая из которых имела ширину полосы 20 нм и центрировалась на 443, 520, 550 и 670 нм. Канал 5 измерял отраженное солнечное излучение и имел полосу пропускания 100 нм с центром на 750 нм и динамический диапазон, который больше подходил для наземных применений.Канал 6 работал в диапазоне от 10,5 до 12,5 мкм и считывал излучаемое тепловое излучение для определения эквивалентной температуры черного тела. (Канал 6 не предоставил достоверных данных во время миссии.) Разрешение CZCS составляло примерно 0,8 км в надире, что немного выше, чем у AVHRR. CZCS использовала конструкцию сканирующего зеркала, которая обеспечивала ширину полосы обзора 1566 км от Nimbus 7 на высоте 955 км.
Канал 5 измерял отраженное солнечное излучение и имел полосу пропускания 100 нм с центром на 750 нм и динамический диапазон, который больше подходил для наземных применений.Канал 6 работал в диапазоне от 10,5 до 12,5 мкм и считывал излучаемое тепловое излучение для определения эквивалентной температуры черного тела. (Канал 6 не предоставил достоверных данных во время миссии.) Разрешение CZCS составляло примерно 0,8 км в надире, что немного выше, чем у AVHRR. CZCS использовала конструкцию сканирующего зеркала, которая обеспечивала ширину полосы обзора 1566 км от Nimbus 7 на высоте 955 км.
Оптическая технология CZCS не позволяла напрямую определять концентрацию хлорофилла; вместо этого рассчитывалась общая концентрация фотосинтетических пигментов («CZCS-пигмент»).Замечательный успех CZCS и очевидная полезность данных о цвете океана для биологических океанографов привели к осознанию того, что специальный датчик цвета океана является критически важным условием для точной характеристики биологической продуктивности океанов.
VII.D.2 SeaWiFS
Хотя потребность в специализированном датчике цвета океана была признана, НАСА заняло несколько лет, прежде чем новая миссия была запланирована. Эта миссия, датчик с широким полем обзора для обзора моря (SeaWiFS), была создана в рамках государственно-частного партнерства с целью снижения затрат на миссию (частному партнеру будет разрешено продавать данные для коммерческих приложений).НАСА достигло соглашения с Orbital Sciences Corporation (OSC) о создании прибора в соответствии со спецификациями НАСА и о запуске его с помощью своей ракеты-носителя с воздушным сбросом Pegasus. Компания OSC впоследствии заключила контракт с Исследовательским центром Санта-Барбары (SBRC) на создание прибора.
Первоначально запланированный к запуску в 1993 году проблемы с ракетой-носителем Pegasus XL следующего поколения удерживали SeaWiFS на земле (но также позволили усовершенствовать датчики). Тем временем японская космическая программа NASDA смогла завершить создание датчика цвета и температуры океана (OCTS) и запустить его на платформе ADEOS (переименованной в Midori) в 1996 году.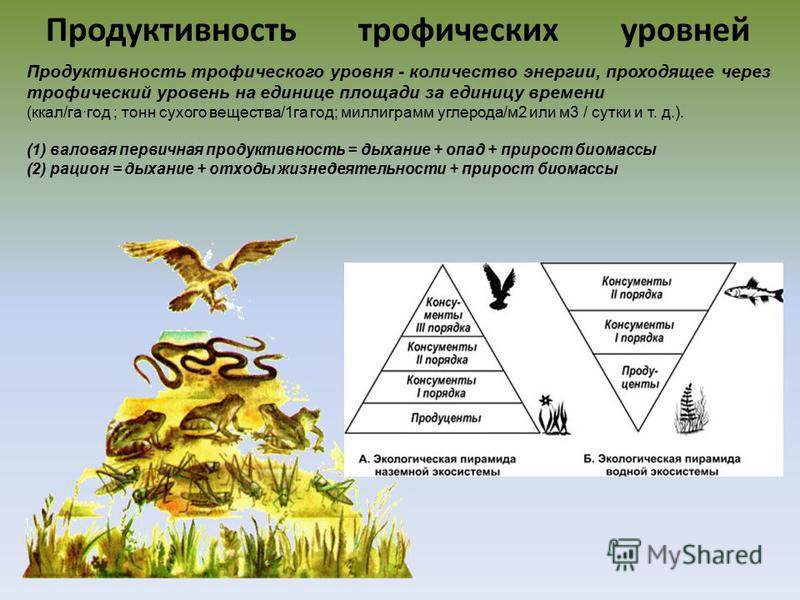 OCTS предоставила первые спутниковые данные о цвете океана после CZCS с дополнительным дополнением параллельных данных SST за 10 месяцев. За неудачной кончиной Midori и OCTS через 2 месяца последовал запуск SeaWiFS на спутнике Orbview-2 1 августа 1997 года. Данные SeaWiFS впервые были распространены среди общественности 23 сентября 1997 года.
OCTS предоставила первые спутниковые данные о цвете океана после CZCS с дополнительным дополнением параллельных данных SST за 10 месяцев. За неудачной кончиной Midori и OCTS через 2 месяца последовал запуск SeaWiFS на спутнике Orbview-2 1 августа 1997 года. Данные SeaWiFS впервые были распространены среди общественности 23 сентября 1997 года.
SeaWiFS использует восемь диапазонов, сосредоточенных в при 412, 443, 490, 510, 555, 670, 735 и 835 нм. Полосы 735 и 835 нм обеспечивают значительно улучшенную атмосферную коррекцию.Оптическая технология обеспечивает гораздо лучшее соотношение сигнал / шум, чем CZCS. Эти улучшения позволяют напрямую рассчитывать концентрацию хлорофилла. Конструкция вращающегося телескопа SeaWiFS обеспечивает ширину сканирования 2800 км с орбитальной высоты 705 км. Эти рабочие параметры позволяют SeaWiFS наблюдать 90% поверхности мирового океана каждые 2 дня (теоретически, потому что облака мешают). SeaWiFS, как и CZCS, также имеет возможность наклоняться вперед или назад, что уменьшает количество морской поверхности, которая не может быть отображена из-за солнечного блика , прямого отражения солнечного света обратно на датчик от поверхности.
SeaWiFS передает данные с разрешением 1 км на любую авторизованную станцию HRPT по всему миру. Субдискретизированные данные с разрешением 4 км записываются на спутник и передаются на объект SeaWiFS Project в Центре космических полетов Годдарда и в Orbimage Inc., дочернюю компанию OSC, которая занимается коммерческими приложениями. Данные SeaWiFS закодированы, так что только коммерческие пользователи и избранные исследовательские пользователи могут получать данные в реальном времени. Все остальные данные обрабатываются проектом SeaWiFS в ежедневные, 8-дневные, ежемесячные и годовые глобальные информационные продукты.
SeaWiFS был запущен в уникальное время, что привело к удивительному совпадению, связанному с CZCS. Одним из первых явлений, которые наблюдал SeaWiFS, было неожиданное цветение кокколитофорид в Беринговом море. Кокколитофориды представляют собой проблему для восприятия цвета океана, потому что они создают микроскопические сферы из карбоната кальция, которые действуют как превосходные светоотражатели, окрашивая воду в блестящий сине-зеленый цвет. Такого масштаба цветения кокколитофорид в Беринговом море ранее не наблюдалось.В то же время Тихий океан демонстрировал признаки (обнаруженные как по данным ТПО, так и по данным альтиметрии) надвигающегося чрезвычайно большого явления Эль-Ниньо. Предыдущий рекордсмен Эль-Ниньо был мощным событием 1982–1983 гг., Которое наблюдало CZCS. Условия Эль-Ниньо имеют отчетливый биологический сигнал в Тихом океане, поскольку продуктивность подавляется вдоль зоны экваториального апвеллинга, что имеет значительные локальные последствия на Галапагосских островах и у побережья Перу и Эквадора. CZCS наблюдала снижение продуктивности в 1983 году, а осенью 1997 года после запуска SeaWiFS наблюдала снижение продуктивности, которое сохранялось до лета 1998 года.Когда условия Эль-Ниньо наконец улеглись, возможности непрерывных наблюдений SeaWiFS показали океанографам удивительную скорость, с которой нормальные, высокопродуктивные условия были восстановлены в Тихом океане во время последующих условий Ла-Нинья.
Такого масштаба цветения кокколитофорид в Беринговом море ранее не наблюдалось.В то же время Тихий океан демонстрировал признаки (обнаруженные как по данным ТПО, так и по данным альтиметрии) надвигающегося чрезвычайно большого явления Эль-Ниньо. Предыдущий рекордсмен Эль-Ниньо был мощным событием 1982–1983 гг., Которое наблюдало CZCS. Условия Эль-Ниньо имеют отчетливый биологический сигнал в Тихом океане, поскольку продуктивность подавляется вдоль зоны экваториального апвеллинга, что имеет значительные локальные последствия на Галапагосских островах и у побережья Перу и Эквадора. CZCS наблюдала снижение продуктивности в 1983 году, а осенью 1997 года после запуска SeaWiFS наблюдала снижение продуктивности, которое сохранялось до лета 1998 года.Когда условия Эль-Ниньо наконец улеглись, возможности непрерывных наблюдений SeaWiFS показали океанографам удивительную скорость, с которой нормальные, высокопродуктивные условия были восстановлены в Тихом океане во время последующих условий Ла-Нинья.
За первые три года работы SeaWiFS предоставила беспрецедентное количество и качество данных о цвете океана с их приложениями в динамике промысла, оценке продуктивности, обнаружении ядовитого и токсичного цветения фитопланктона, а также в многочисленных прибрежных и региональных исследованиях.Хотя разрешение намного ниже, чем у Landsat и коммерческих спутников дистанционного зондирования, таких как Spot или IKONOS, SeaWiFS также отлично зарекомендовал себя в наблюдении земных и атмосферных явлений. В частности, SeaWiFS зафиксировала огромные клубы дыма от пожаров в тропических лесах в Индонезии и на полуострове Юкатан в Мексике во время Эль-Ниньо 1997–1998 годов. Еще одно событие, которое SeaWiFS регулярно наблюдает, — это облака пыли, которые переносятся в верхние слои атмосферы пыльными бурями. В апреле 1998 года SeaWiFS поймал сильный шторм в пустыне Гоби, который возник в Китае и пересек Тихий океан примерно за одну неделю.В феврале 2000 года было замечено облако пыли размером с Испанию, поднимающееся из пустыни Сахара над восточной частью Атлантического океана.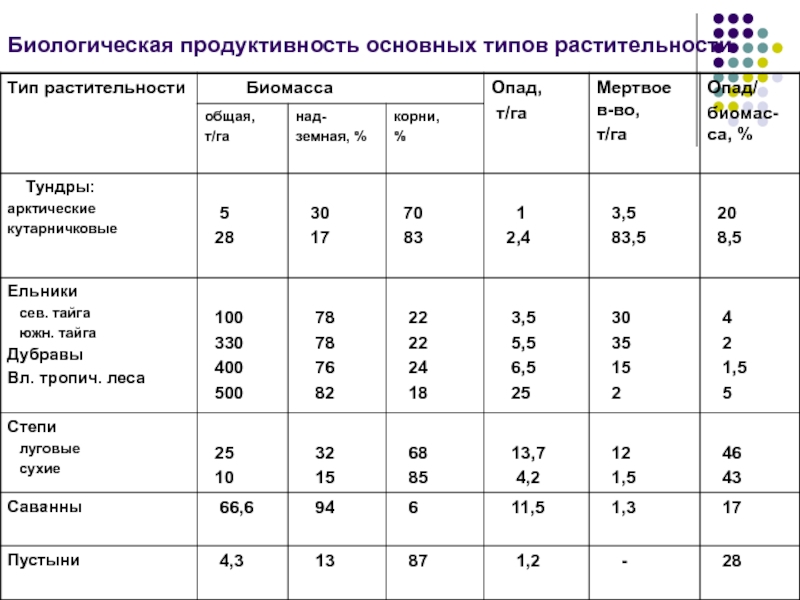 SeaWiFS может отслеживать как ежедневные местоположения ураганов, так и последующую нагрузку наносов, которые выносятся в океан речными наводнениями, вызванными дождями, связанными с ураганами.
SeaWiFS может отслеживать как ежедневные местоположения ураганов, так и последующую нагрузку наносов, которые выносятся в океан речными наводнениями, вызванными дождями, связанными с ураганами.
VII.D.3 После SeaWiFS…
SeaWiFS также знаменует начало эры, когда резко возросло количество миссий по определению цвета океана. Немцы разместили свой модульный оптико-электронный сканер (МОП) на модуле дистанционного зондирования «Природа» космической станции МИР в 1996 году, а также на индийском спутнике дистанционного зондирования IRS-P3.По сравнению с SeaWiFS, MOS имеет 17 диапазонов и разрешение 0,65 км, но ширину полосы обзора всего 85 км. Данные МОП оказались ценными в нескольких экспериментах по взаимному сравнению данных с SeaWiFS. Мидори также носил прибор POLDER (поляризация и направленность отражения Земли), который также обладал способностью окрашивать океан. Корея запустила KOMPSAT-1, а Китайская Республика запустила ROCSAT-1, оба с возможностями получения изображений, аналогичными SeaWiFS, в 1999 году. Орбиты этих спутников расположены таким образом, чтобы обеспечить оптимальные изображения региональных вод вокруг Кореи и Тайваня.
Орбиты этих спутников расположены таким образом, чтобы обеспечить оптимальные изображения региональных вод вокруг Кореи и Тайваня.
Следующей вехой в развитии дистанционного зондирования цвета океана стал запуск первого MODIS на спутнике Terra в декабре 1999 года. MODIS — сканер с вращающимся зеркалом, поддерживающий 36 диапазонов в УФ, видимом и ИК диапазонах. для дистанционного зондирования атмосферы, суши и океана. (Другие приложения MODIS описаны в других разделах.) Для океанов MODIS по существу дублирует набор диапазонов SeaWiFS с небольшими различиями в расположении диапазонов и полосе пропускания 10 нм.Он добавляет новую важную полосу: 683 нм, позволяющую обнаруживать флуоресценцию хлорофилла, которая предоставляет информацию о физиологическом здоровье фитопланктона в дополнение к их концентрации. Данные флуоресценции позволяют гораздо лучше оценить первичную продуктивность (производство нового углерода), относящуюся к фитопланктону. Кроме того, как и OCTS, MODIS получает данные SST в той же области просмотра, что и данные о цвете океана, что позволяет напрямую коррелировать текущую динамику с концентрациями фитопланктона для связанных физико-биологических моделей.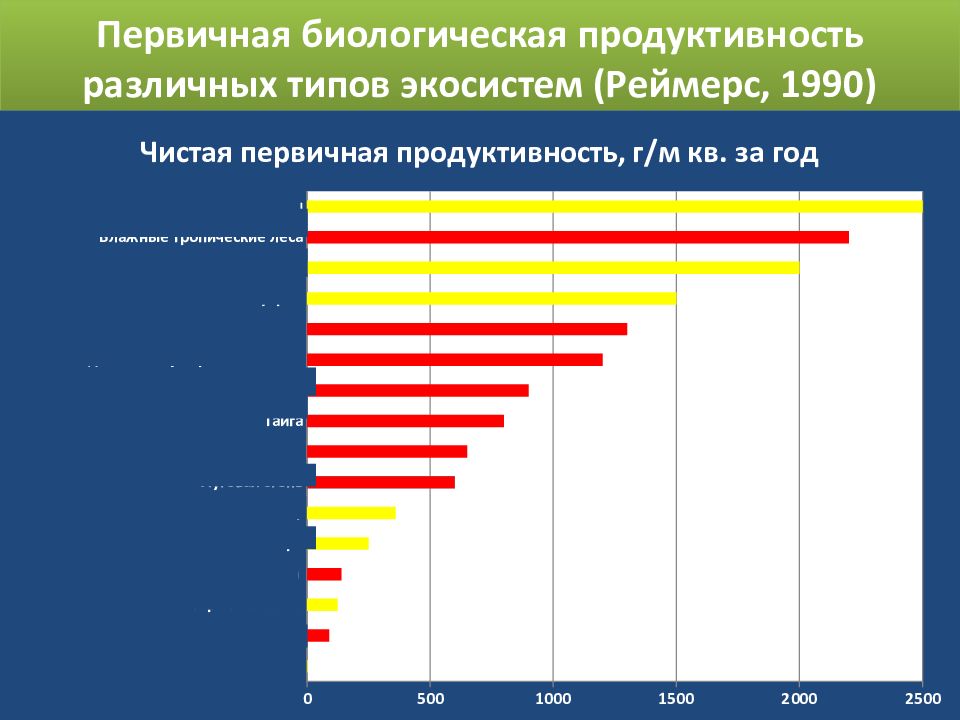 У MODIS есть одно ограничение по сравнению с SeaWiFS: ему не хватает возможности наклона, что означает, что больше данных MODIS будут подвержены влиянию солнечного света (фактически, охват двух инструментов MODIS примерно равен охвату одного SeaWiFS). Второй инструмент MODIS находится на платформе Aqua, запуск которого запланирован на начало 2002 года. Кроме того, Япония и ЕКА имеют инструменты цвета океана на ADEOS-II и ENVISAT; см. Раздел IX.C.
У MODIS есть одно ограничение по сравнению с SeaWiFS: ему не хватает возможности наклона, что означает, что больше данных MODIS будут подвержены влиянию солнечного света (фактически, охват двух инструментов MODIS примерно равен охвату одного SeaWiFS). Второй инструмент MODIS находится на платформе Aqua, запуск которого запланирован на начало 2002 года. Кроме того, Япония и ЕКА имеют инструменты цвета океана на ADEOS-II и ENVISAT; см. Раздел IX.C.
Биологическая продуктивность водных объектов
Выбор площадки для аквакультуры: биологическая продуктивность водных объектов Биологическая продуктивность водных систем, как и суши, определена.
на нескольких уровнях, основная или первичная производительность, которая снова делится
на валовую и чистую первичную продуктивность и вторичную продуктивность на различных трофических уровнях (обсуждение выше).Основная или первичная производительность ,
определяется как скорость, с которой энергия сохраняется в фотосинтетических
деятельность организмов-продуцентов (организмов, несущих хлорофилл, в основном растений
и фитопланктон) в виде органических веществ, которые можно использовать в качестве
пищевые вещества.
Валовая первичная продуктивность — общая скорость фотосинтеза, включая органическое вещество, использованное при дыхании в течение периода измерения (также известный как общий фотосинтез или полная ассимиляция ).Чистая первичная продуктивность скорость хранения энергии в виде пищи, т.е. исключая энергия, рассеиваемая растениями при дыхании (также называемая «кажущейся») фотосинтез или чистая ассимиляция). Обычно значение, полученное для ставки дыхания растений добавляется к очевидному фотосинтезу для получения оценок валовой первичной продуктивности.
Нормы накопления энергии на трофических уровнях потребителей и деструкторов
называются «вторичной продуктивностью»; эти ставки, как уже
указывается все меньше и меньше на каждом последующем трофическом уровне.Чтобы быть правильным,
на вторичном уровне продуктивности происходит только усвоение пищи.
продуцируемые автотрофами (растениями) на каждом последующем трофическом уровне (гетеротрофы)
и как таковой термин «производительность» не должен ассоциироваться с
эти трофические уровни.
Термин «производительность» всегда используется в этом контексте для обозначения «ставка производства »или« скорость потока энергии ». Валовая продукция (P G ) и нетто производство (P N ) вторичные производства (P 2 — P 5 ) указаны в фигура (поток энергии) (рис.10.3).
Следует подчеркнуть, что «производительность» относится к скорости производства. или скорость потока энергии и, следовательно, «богатство» водоема или экосистема; об этом нельзя судить только по «биомассе» урожая. (стоячий урожай) организмов на различных трофических уровнях.
Крупная биомасса или стоячий урожай не обязательно должны быть связаны с высокая урожайность. В этом контексте важна концепция текучести кадров.
Очевидно, что валовая и чистая производительность будут меняться в зависимости от
богатство водоема.Производительность может варьироваться от менее 1 г углерода
(C) / M 2 / день -1 в открытом море (глубокое озеро) до более 60 г C / M 2 / день -1 в естественном
и искусственные системы.
Валовая продуктивность некоторых водоемов, адаптированная из Odum (1959), указаны:
| Открытый океан (долгосрочный) | : | 0,5 г / м 2 / день | |||
| Мелководье у берега | : | 3.2 | |||
| Приливные эстуарии | : | 4,4 | |||
| Коралловые рифы | : | 18,2 « | |||
| Пруд — с неочищенными сточными водами | 9033 | 9033 9033 903 | : | 43,0 « | |
| Загрязненный поток | : | 57,0″ |
Видно, что очень большая часть поверхности Земли (океан и
луга) малопродуктивны.Определенные области, такие как зона шельфа,
arella умеренно или высокопродуктивно, как и коралловые рифы. в
в последнем случае симбиоз растений и животных приводит к очень быстрому обмену питательных веществ
вызывая очень высокую производительность.
Рис. 10.2. Диаграмма, показывающая поток энергии в экосистеме с указанием различные уровни производительности валовой первичной продукции (P G ), чистое первичное производство (P N ) и вторичное производство, P 2 , P 3 , P 4 и P 5 — декомпозер не указан (Изменено из Odum, 1959) (размерные соотношения в эскизе не в масштабе)
(PDF) Биологическая продуктивность океана
природа Образование
15
© 2012 Macmillan Publishers Limited.Все права защищены
Том 3 | Выпуск 6 | 2012
Ссылки и рекомендуемая литература
Altabet, M. A. et al. Связанные с климатом изменения в денитрификации
в Аравийском море по осадкам, соотношение 15N / 14N. Nature 373,
506-509 (1995). doi: 10.1038 / 373506a0
Анбар, А. Д. и Кнолл, А. Х. Протерозойская химия океана и эволюция —
люция: био-неорганический мост? Science 297, 1137-1142 (2002).
DOI: 10.1126 / science.1069651
Арчер Д. «Биологические потоки в океане и атмосферный pCO2»,
в «Трактате по геохимии», том. 6, ред. Х. Д. Холланд и К. К.
Turekian (Elsevier, 2003) 275-291.
Arrigo, K. R. et al. Структура сообщества фитопланктона и уменьшение количества питательных веществ и CO2 в Южном океане на уровне
. Science
283, 365-367 (1999). DOI: 10.1126 / science.283.5400.365
Barbeau, K. et al. Фотохимический цикл железа на поверхности
океана, опосредованный микробными лигандами, связывающими железо (III).Nature
413, 409-413 (2001). DOI: 10.1038 / 35096545
Behrenfeld, M. J. et al. Климатические тенденции в современной продуктивности океана
. Nature 444, 752-755 (2006). DOI: 10.1038 /
nature05317
Bender, M. et al. Сравнение 4 методов определения продукции планктонных сообществ
. Лимнология и океанография —
phy 32, 1085-1098 (1987).
Бойд, П. В. и Эллвуд, М. Дж. Биогеохимический цикл железа в
В. и Эллвуд, М. Дж. Биогеохимический цикл железа в
океане.Nature Geoscience 3, 675-682 (2010). DOI: 10.1038 /
ngeo964
Boyd, P. W. et al. Эксперименты по мезомасштабному обогащению железа 1993-
2005: Синтез и будущие направления. Science 315, 612-617
(2007). DOI: 10.1126 / science.1131669
Брандес, Дж. А. и Девол, А. Х. Глобальный морской изотопный баланс азота
: последствия для круговорота азота в голоцене
. Глобальные биогеохимические циклы 16, 1120 (2002).
doi: 10.1029 / 2001gb001856
Broecker, W. S. Химия океана в ледниковое время. Geochimica et al.,
,, Cosmochimica Acta, 46, 1689–1705 (1982). DOI: 10.1016 / 0016-
7037 (82) -7
Бюсселер, К. О. Разделение производства и экспорта твердых частиц
на поверхность океана. Global Biogeochemical Cycles 12,
297-310 (1998). DOI: 10.1029 / 97gb03366
Chisholm, S. W. et al. Новый свободноживущий прохлорофит, изобилующий
, обитает в океанической эвфотической зоне. Nature 334, 340-343
Nature 334, 340-343
(1988). doi: 10.1038 / 334340a0
Кристенсен, Дж. П. Экспорт углерода с континентальных шельфов, de-
нитрификация и атмосферный углекислый газ. Continen-
tal Shelf Research 14, 547-576 (1994). DOI: 10.1016 / 0278-
4343 (94) -1
Морель, Ф. М. М. и др. Подкисление океана: национальная стратегия
для решения проблем изменяющегося океана. Национальная газета Aca-
demic Press, 2010 г.
Каллен, Дж. Дж. Глубокий максимум хлорофилла: сравнение вертикальных
каловых профилей хлорофилла a. Канадский журнал рыболовства и
Aquatic Sciences 39, 791-803 (1982). DOI: 10.1139 / f82-108
Dalsgaard, T., Thamdrup, B. & Caneld, D. E. Анаэробные аммо-
окисление ниия (анаммокс) в морской среде. Re-
поиск по микробиологии 156, 457-464 (2005). DOI: 10.1016 / j.
resmic.2005.01.011
де Баар, Х.Закон минимума Дж. В. фон Либиха и экология планктона
(1899–1991). Progress in Oceanography 33, 347-386
Progress in Oceanography 33, 347-386
(1994). DOI: 10.1016 / 0079-6611 (94)
-1Deutsch, C. et al. Изотопные ограничения на ледниковые / межледниковые
изменения баланса азота в океане. Global Biogeochemi-
циклов 18, GB4012 (2004). DOI: 10.1029 / 2003gb002189
Deutsch, C. et al. Пространственная взаимосвязь поступления и потерь азота
в океане.Nature 445, 163-167 (2007). DOI: 10.1038 / na-
ture05392
Duce, R.A. et al. Воздействие атмосферного антропогенного азота
на открытый океан. Science 320, 893-897 (2008). doi: 10.1126 /
science.1150369
Dugdale, R.C. & Goering, J. J. Поглощение новых и регенерированных
форм азота в первичной продуктивности. Лимнология и
Океанография 12, 196-206 (1967).
Emerson, S. & Hedges, J.«Диагенез отложений и бентосный поток», №
в «Трактате по геохимии», том. 6, ред. Х. Д. Холланд и К. К.
Turekian (Elsevier, 2003) 293-319.
Эппли, Р. У. и Петерсон, Б. Дж. Твердые органические вещества и новые планктонные образования
в глубоких океанских глубинах. Nature 282,
677-680 (1979). doi: 10.1038 / 282677a0
Фальковски П.Г. Эволюция азотного цикла и ее влияние
на биологическое связывание СО2 в океане.Nature
387, 272-275 (1997). DOI: 10.1038 / 387272a0
Francois, R. et al. Вклад стратификации поверхностных вод Южного океана
в низкие концентрации СО2 в атмосфере в течение
последнего ледникового периода. Nature 389, 929-935 (1997).
Froelich, P. N. et al. Морской цикл фосфора. Американский
Научный журнал 282, 474-511 (1982).
Ganeshram, R. S. et al. Значительные изменения в океанических биогенных веществах —
торией от ледникового до межледникового периода.Nature 376, 755-
758 (1995). doi: 10.1038 / 376755a0
Гейдер, Р.Дж., Макинтайр, Х.Л. и Кана, TM Динамическая модель роста и акклиматизации фитопланктона
: реакция на ускоренную скорость роста балла-
и отношение хлорофилла а: углерода к свету,
ограничение питательных веществ и температуры. Морская экология-Прогресс
Морская экология-Прогресс
Серия 148, 187-200 (1997). DOI: 10.3354 / meps148187
Hansell, D.A. et al. Растворенное органическое вещество в океане: трофей кон-
стимулирует новые идеи.Океанография 22, 202-211
(2009).
Jaccard, S. L. et al. Ледниковые / межледниковые изменения в субарктике
Стратификация Северного Тихого океана. Science 308, 1003-1006 (2005).
doi: 10.1126 / science.1108696
Джин, X. и Грубер, Н. Определение радиационной выгоды от удобрения океаническим железом
за счет увеличения выбросов N2O. Geophysical Re-
search Letters 30, 2249 (2003). DOI: 10.1029 / 2003gl018458
Джонсон, К.С., Гордон, Р. М. и Коул, К. Х. Что контролирует концентрацию растворенного железа
в мировом океане? Морская химия 57,
137-161 (1997). DOI: 10.1016 / s0304-4203 (97) 00043-1
Кеннет, Дж. П. Кайнозойская эволюция антарктического оледенения, круговая полоса
вместе с Антарктическим океаном и их влияние на глобальный палеоокеан. Журнал геофизических исследований океанов и атмосферы —
Журнал геофизических исследований океанов и атмосферы —
сферы 82, 3843-3860 (1977). DOI: 10.1029 / JC082i027p03843
Килхэм, П.И Хекки Р. Э. Сравнительная экология морского и пресноводного фитопланктона. Лимнология и океанография 33,
776-795 (1988).
Kohfeld, K. E. et al. Роль морской биологии в ледниково-интерглинистых циклах СО2. Science 308, 74-78 (2005). doi: 10.1126 / sci-
ence.1105375
Ландри, М. Р. и Хассетт, Р. П. Оценка воздействия пастбищ
морского микрозоопланктона. Морская биология 67, 283-288
(1982).DOI: 10.1007 / bf00397668
Maldonado, M. T. et al. Совместное ограничение роста фитопланктона
светом и Fe зимой в северо-восточной субарктической части Тихого океана.
Deep-Sea Research Part II — Актуальные исследования в океанографии
46, 2475-2485 (1999). doi: 10.1016 / s0967-0645 (99) 00072-7
Мартин, Дж. Х. и Фитцуотер, С. Э. Дефицит железа ограничивает рост фитопланков —
тонн в Северо-восточной части Тихого океана в Субарктике. Nature 331, 341-
Nature 331, 341-
343 (1988).DOI: 10.1038 / 331341a0
Martin, J. H. et al. VERTEX: исследования фитопланктона / железа в заливе
Аляски. Deep-Sea Research Part A: Oceanographic
Research Papers 36, 649-680 (1989). DOI: 10.1016 / 0198-
0149 (89)
-1
Мартин, Дж. Х. и др. ВЕРТЕКС: Круговорот углерода на северо-востоке Тихого океана.
Deep-Sea Research Part A: Oceanographic Research Papers 34,
267-285 (1987). DOI: 10.1016 / 0198-0149 (87) -0
Мартин, Дж.H. Ледниково-межледниковое изменение CO2: биологическая продуктивность железа hy-
в предложении
Эти примеры взяты из корпусов и из источников в Интернете. Любые мнения в примерах не отражают мнение редакторов Cambridge Dictionary, Cambridge University Press или его лицензиаров.
Эти условия существуют в областях высокой биологической продуктивности , связанных с вулканическими островами и зонами апвеллинга, богатыми питательными веществами.
Википедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Концентрация углерода указывает на повышенный уровень биологической продуктивности , возможно, из-за высокого уровня питательных веществ, доступных из переслаивающегося вулканического материала.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA. Доказательства более высокой биологической продуктивности представляют собой биоконцентрированный барий.
Википедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Он предполагает, что система имеет возможности способствовать способности коэволюции увеличивать биологическую продуктивность и стабильность экосистемы.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA. Эти месторождения долгое время считались основным источником железа на поверхности океана и, следовательно, основным источником железа для биологической продуктивности .
Википедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Коэффициенты пересчета могут оказаться большим подспорьем при оценке воздействия поправок на почву на биологическую продуктивность на основе данных по урожайности.
Эти урожаи биологические измерения продуктивности в агроэкосистеме тропических засушливых земель имеют агрономические и экологические последствия.
Практическое беспокойство, инициировавшее это исследование, заключается в том, подвержены ли такие системы значительному повышению их биологической продуктивности .
Настоящее исследование фокусируется на влиянии биологической продуктивности и урожайности сельскохозяйственных культур на параметры плодородия почвы.
Скорость биологическая продуктивность ограничена в полярных условиях, влияя на процессы ослабления, такие как микробная деградация.
Загрязнение атмосферы, океанов, поверхностных и подземных вод уже достигло такого уровня, что биологическая продуктивность начинает заметно снижаться.
Мелководные моря и заросли водорослей западного залива особенно важны для биологической продуктивности как места разведения рыбы и креветок.
Луга водорослей обладают высокой биологической продуктивностью и представляют собой богатые биоразнообразные среды обитания.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Его распространение также ограничено районами с высокой биологической продуктивностью , такими как полярные океаны и зоны апвеллинга вблизи экватора.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.
Большие колебания уровня воды из-за выработки электроэнергии влияют на рекреационные возможности и биологическую продуктивность озера.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Наиболее вероятный метод восстановления предполагает увеличение биологической продуктивности , транспортируя углерод в глубины океана.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA. На протяжении многих лет здесь проводились глубокие исследования процессов функционирования и биологической продуктивности водных объектов.
Википедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Морская вода характеризуется повышенной концентрацией растворенного кислорода, что приводит к высокой биологической продуктивности .
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Он сказал, что изменения в биологической продуктивности были связаны с изменениями доступных питательных веществ.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.
Экономическая рентабельность может быть максимизирована без значительного ущерба биологическая продуктивность за счет сочетания стратегии кормления на основе рыбы и ракообразных.
ИзВикипедия
Этот пример взят из Википедии и может быть повторно использован по лицензии CC BY-SA.Эти примеры взяты из корпусов и из источников в Интернете. Любые мнения в примерах не отражают мнение редакторов Cambridge Dictionary, Cambridge University Press или его лицензиаров.
| Этот проект планируется завершить в сентябре 2007 года. | Благодарности: это исследование является совместным усилием.Финансирование по различным аспектам был предоставлен Геологической службой США, Инженерный корпус армии США и Служба рыболовства и дикой природы США. Часть этой работы спонсируется Long Term Resource Monitoring. Программа системы реки Верхняя Миссисипи, которая является совместной усилия государственных и федеральных агентств, включая рыбную ловлю и дикую природу США Служба, Инженерный корпус армии США, Геологическая служба США, Служба охраны природных ресурсов, U.S. Охрана окружающей среды Агентства и штатов Иллинойс, Айова, Миннесота, Миссури и Висконсин. Университет Висконсин-Ла-Кросс является партнером в этом исследовательская работа. Научный персонал, участвовавший в этом исследовании Рэнди Хайнс, Джон Валлацза, Джим Рогала, Джим Лайон, Боб Кеннеди, Джон Калас и Бэррон Муди.  Для получения дополнительной информации см. Эти публикации: 1 Джонсон, Б.Л., У. Б. Ричардсон и T. J. Naimo. 1995. Прошлое, настоящее и будущее концепции большой реки. экология. Биология 43: 134-141. Деттмерс, Дж. М., С. Гутрейтер, Д. Х. Валь и Д. А. Солук. 2001 г. Закономерности обилия рыбы в основных протоках Верхнего Миссисипи Речная система. Канадский журнал рыболовства и водных наук 58: 933-942. |
 Предоставлено Службой охраны рыболовства и дикой природы США.
Предоставлено Службой охраны рыболовства и дикой природы США. 
Биологическая продуктивность вод
Цитируется по
1. Анализ данных на основе модели влияния зимнего перемешивания на первичную продукцию в озере при реолиготрофикации
2. Влияние субмезомасштабной вертикальной адвекции на первичную продуктивность в южно-восточно-китайском море
3. Энергетический поток при росте и производство
4. Характеристика индивидуальной изменчивости роста мидий (Mytilus galloprovincialis) и тестирование ее физиологических факторов с использованием анализа функциональных данных
5. Биомасса и продуктивность рыб в эстуариях: пример из Южной Африки
Биомасса и продуктивность рыб в эстуариях: пример из Южной Африки
6. Гибкость физиологических признаков, лежащих в основе межиндивидуальных различий в росте литоральных и подливных мидий Mytilusgalloprovincialis
7. Продуктивная способность местообитаний рыб: обзор исследований и перспективных направлений
8. Воздействие частичного демонтажа выведенных из эксплуатации нефтегазовых платформ на рыбную биомассу и продукцию на оставшуюся конструкцию платформы и окружающие насыпи ракушек
9. Наличие и использование мидий Mytilus edulis кормовых отходов рыбных отходов в коммерческой интегрированной многотрофической системе аквакультуры (IMTA): подход к оценке по нескольким показателям
10. Влияние видов и функционального разнообразия: понимание роли взаимодополняемости и конкуренция за функцию экосистемы в скоплении рыб тропических водотоков
11. Нефтяные платформы у побережья Калифорнии являются одними из самых продуктивных местообитаний морских рыб в мире
12. Влияние сезонных колебаний фитопланктона на биоэнергетические реакции мидий (Mytilus galloprovincialis), удерживаемых на плоту вблизи загонов для ловли красного морского леща (Pagellus bogaraveo)
Влияние сезонных колебаний фитопланктона на биоэнергетические реакции мидий (Mytilus galloprovincialis), удерживаемых на плоту вблизи загонов для ловли красного морского леща (Pagellus bogaraveo)
13. Численное моделирование перехода доминирующих водорослей в озере Китаура, Япония
14. Устойчивость молоди Mytilus galloprovincialis к экспериментальному подкислению морской воды
15. Оценки параметров популяции и соотношения потребления / биомассы рыб в водоемах, штат Парана, Бразилия
16. Факторы, влияющие на пространственные и временные вариации в продуктивности и соотношении продуктивности / биомассы кумжи (Salmo trutta), обитающей в ручье, в кантабрийских водотоках
17. Набор как фактор динамики продуктивности обитающей в ручье кумжи ( Сальмо Трутта )
18. Рост нильской тилапии Oreochromis niloticus, скармливаемый рационами с разным уровнем дрожжевых белков
19. Влияние возрастного выбора среды обитания сообществом речных раков (Orconectes spp.) на вторичном производстве
Влияние возрастного выбора среды обитания сообществом речных раков (Orconectes spp.) на вторичном производстве
20. Зависимый от плотности рост речной форели Salmo trutta L.
21. Produtividade de peixes em um riacho costeiro da bacia do Leste, Рио-де-Жанейро, Бразилия
22. Двухфазное самоореживание молоди озерно-перелетной кумжи, живущей в ручье. Сальмо Трутта L. Совместимость линейных и нелинейных моделей в популяциях?
23. Важность пополнения для динамики производства водной форели (Salmo trutta)
24. Пространственно-временная динамика производства кумжи в кантабрийском ручье: влияние плотности и качества среды обитания
25. Атлантический лосось Salmo salar Л., Кумжа Сальмо Трутта L. и арктический голец Salvelinus alpinus (L.): обзор аспектов их жизненных историй
26. Влияние речных процессов и неоднородности местообитаний на распространение, рост и плотность молоди атлантического лосося (
Salmo salar
L.), с последствиями для численности взрослой рыбы
Влияние речных процессов и неоднородности местообитаний на распространение, рост и плотность молоди атлантического лосося (
Salmo salar
L.), с последствиями для численности взрослой рыбы
27. Использование аллометрии с размером рыбы для оценки продукции по биомассе ( п / B ) соотношения популяций лососевых
28. Влияние концентрации пищи в слоях угря на энергетический баланс инвазивных мидий Musculista Senhousia
29. Оптимальное потребление энергии и общая эффективность преобразования энергии для кумжи, Salmo trutta, кормления беспозвоночными или рыбой
30. Рост нильской тилапии Oreochromis niloticus, полученной с использованием различных уровней спиртовых дрожжей
31. Рост, потребление и продуктивность калифорнийского киллифа в лагуне Охо-де-Либре, Мексика
32. Оценка экологических последствий изменения среды обитания: выход за пределы производственной мощности
33. Влияние концентрации пищи на возможности роста и характеристики роста семян Ruditapes decussatus (L.), выращенных в системе открытого потока
Влияние концентрации пищи на возможности роста и характеристики роста семян Ruditapes decussatus (L.), выращенных в системе открытого потока
34. Последовательность катаболического окисления субстрата и энтальпийный баланс развивающихся эмбрионов и личинок желточного сака камбалы (Scophthalmus maximus L.)
35. Рост плавательной мускулатуры и его метаболическая стоимость у личинок сига при разных температурах
36. Физиологическая энергетика развивающихся эмбрионов и личинок желточного мешка атлантической трески (Gadus морхуа).II. Липидный обмен и баланс энтальпии
37. Количественный анализ опубликованных данных о росте, метаболизме, потреблении пищи и связанных с ними характеристиках краснобрюхой пираньи, Serrasalmus nattereri (Characidae)
38. Производство молоди Атлантики лосось, Salmo salar, в природных водах
39. СТОИМОСТЬ РАЗВИТИЯ КЛЕТК И ОРГАНИЗМОВ: ОБЩИЕ ПРАВИЛА И СРАВНИТЕЛЬНЫЕ АСПЕКТЫ
40. Количественный анализ опубликованных данных о росте, метаболизме, потреблении пищи и связанных с ними характеристиках Краснобрюхой пираньи, Serrasalmus nattereri (Characidae)
Количественный анализ опубликованных данных о росте, метаболизме, потреблении пищи и связанных с ними характеристиках Краснобрюхой пираньи, Serrasalmus nattereri (Characidae)
41. Влияние концентрации пищи на энергетический баланс и показатели роста семян Venerupis pullastra, выращенных в системе открытого потока
42. Скорость метаболизма и стоимость роста молоди щуки (Esox lucius L.) и окуня (Perca fluviatilis L. ): использование энергетических бюджетов в качестве индикаторов экологических изменений
43. Рост и производство
44. Структура сообщества рыб, привлеченных мелководными устройствами для скопления рыбы у побережья Южной Каролины, США.SA
45. Биология популяции щуки Esox lucius L. в двух озерах с гравийными карьерами, с особым упором на раннюю историю жизни
46. О взаимодействии пелагической пищевой сети в крупных водоемах
47 Экология кумжи Salmo trutta в английских меловых ручьях
48. Кумжа Salmo trutta L., добыча в прибрежном потоке Ирландии
Кумжа Salmo trutta L., добыча в прибрежном потоке Ирландии
49. Подробнее о преобразовании энергии в эктотермах и эндотермах: биохимические и социальные расходы
50. Рост молоди камбал (Platichthys flesus L.) при солености 0, 5, 15 и 35 %%
51. Темпы использования кормов прибрежными рыбами в мезотрофном озере северного умеренного пояса
52. Отказ от производства: альтернативный метод оценки последствий уноса рыбы и потерь от столкновения на электростанциях и других водозаборах
53. Низкая производительность? Эффективность? гомеотермных популяций: недоразумение
54. Обзор вторичной продукции в пелагических экосистемах
55. Производство молоди лососевых в ручье питомника озера Эри
56. Распределение, продукция и возрастная структура слизистых бычков в арктическом озере
57. Вылов рыбы для пропитания и последствия для управления в системе реки Барам, Саравак, Малайзия
58. Расчеты структуры животноводческого сообщества и энергетического бюджета популяции Daphnia magna (straus) по отношению к окружающей среде горного бассейна
Расчеты структуры животноводческого сообщества и энергетического бюджета популяции Daphnia magna (straus) по отношению к окружающей среде горного бассейна
59. Мезомасштабное пространственное распределение планктона: совместная эволюция концепций и приборов
60. Физиологическая энергия
61. Темпы роста и модели
63. Производство молоди атлантического лосося, Salmo salar in верхний Уай, Уэльс
64. Некоторые факторы, влияющие на потребление водорослей субарктическими эфемероптерами, Plecoptera и simuliidae
65. ЭФФЕКТИВНОСТЬ КОНВЕРСИИ В ГЕТЕРОТРОФНЫХ ОРГАНИЗМАХ
66. Некоторые аспекты биологии питания донных травоядных
67. Вторичная продукция во внутренних водах
68. Экология водорослей, Zostera marina (L.), сообществ рыб. II. Функциональный анализ
69. Полный баланс углерода и азота для плотоядных амфипод Calliopius laeviusculus (KRØYER)
70.