
Государство в широком и узком смысле: Лекция 7. Государство в политической системе
Лекция 7. Государство в политической системе
Лекция 7. ГОСУДАРСТВО
1. Сущность государства
1.1 Широкое и узкое значение термина «государство».
Центральным институтом политической системы является государство. В его деятельности концентрируется основное содержание политики. Сам термин «государство» обычно употребляется в двух значениях. В широком смысле государство понимается как общность людей, представляемая и организуемая органом высшей власти и проживающая на определенной территории. Оно тождественно стране и политически организованному народу. В этом значении говорят, например, о российском, американском, немецком государстве, имея в виду все представляемое им общество.
Примерно до XVII в. государство обычно трактовалось широко и не отделялось от общества. Для обозначения государства использовались многие конкретные термины: «полития», «княжество», «королевство», «империя», «республика», «деспотия», «правление» и др.
Четкое разграничение государства и общества было обосновано в контрактных (договорных) теориях государства Гоббсом, Локком, Руссо и другими представителями либерализма. В них эти понятия разделяются не только содержательно, но и историйки, поскольку утверждается, что существовавшие первоначально в свободном и неорганизованном состоянии индивиды в результате хозяйственного и иного взаимодействия вначале образовали общество, а затем для защиты своей безопасности и естественных прав договорным путем создали специальный орган государство. В современной науке  п.
п.
1.2. Общие признаки государства.
Государства разных исторических эпох и народов мало схожи между собой. И все же они имеют некоторые черты, которые в большей или меньшей степени присущи каждому из них, хотя у современных государств, подверженных интеграционным процессам, они порою достаточно размыты. Общими для государства являются следующие признаки:
1. Отделение публичной власти от общества, ее несовпадение с организацией всего населения, появление слоя профессионалов-управленцев. Этот признак отличает государство от родоплеменной организации, основанной на принципах самоуправления
2. Территория, очерчивающая границы государства. Законы и полномочия государства распространяются на людей, проживающих на определенной территории. Само оно строится не по кровнородственному или религиозному признаку, а на основе территориальной и, обычно, этнической общности людей.
3. Суверенитет, т.е. верховная власть на определенной территории. В любом современном обществе имеется множество властей: семейная, производственная, партийная и т.д. Но высшей властью, решения которой обязательны для всех граждан, организаций и учреждений, обладает государство. Лишь ему принадлежит право на издание законов и норм, обязательных для всего населения.
В любом современном обществе имеется множество властей: семейная, производственная, партийная и т.д. Но высшей властью, решения которой обязательны для всех граждан, организаций и учреждений, обладает государство. Лишь ему принадлежит право на издание законов и норм, обязательных для всего населения.
4. Монополия на легальное применение силы, физического принуждения. Диапазон государственного принуждения простирается от ограничения свободы до физического уничтожения человека. Возможность лишить граждан высших ценностей, каковыми являются жизнь и свобода, определяет особую действенность государственной власти. Для выполнения функций принуждения у государства имеются специальные средства (оружие, тюрьмы и т.д.), а также органы — армия, полиция, службы 6езопасности, суд, прокуратура.
5. Право на взимание налогов и сборов с населения. Налоги необходимы для содержания многочисленных служащих и для материального обеспечения государственной политики: оборонной, экономической, социальной и т. д.
д.
6. Обязательность членства в государстве. В отличие, например от такой политической организации, как партия, пребывание в которой добровольно и не обязательно для населения, государственное гражданство человек получает с момента рождения.
7. Претензия на представительство общества как целого и защиту общих интересов и общего блага. Ни одна другая организация кроме разве что тоталитарных партий-государств, не претендует на представительство и защиту всех граждан и не обладает для этого необходимыми средствами.
Определение общих признаков государства имеет не только научное, но и практическое политическое значение, особенно для международного права. Государство — субъект международных отношений. Лишь на основе обладания качествами государства те или иные организации признаются субъектами международного права и наделяются соответствующими правами и обязанностями. В современном международном праве выделяются три минимальных признака государства: территория, народ, объединенный правовым союзом граждан (гражданством), и суверенная власть, осуществляющая эффективный контроль хотя бы над большинством территории и населения.
Отмеченные выше признаки отличают государство от других организаций и объединений, однако еще не раскрывают его связь с обществом, факторы, лежащие в основе его возникновения и эволюции.
1.3. Причины возникновения государства.Государство появляется в результате разложения родоплеменного строя, постепенного обособления от общества вождей и их приближенных и сосредоточения у них управленческих функций, ресурсов власти и социальных привилегий под действием ряда факторов, важнейшие из которых:
1) развитие общественного разделения труда, выделение управленческого труда в целях повышения его эффективности в специальную отрасль и образование для этого специального орана — государства;
2) возникновение в ходе развития производства частной собственности, классов и эксплуатации (марксизм). Не отрицая влияние этих факторов, большинство современных ученых все же не связывают существование государства непосредственно с возникновением частной собственности и классов.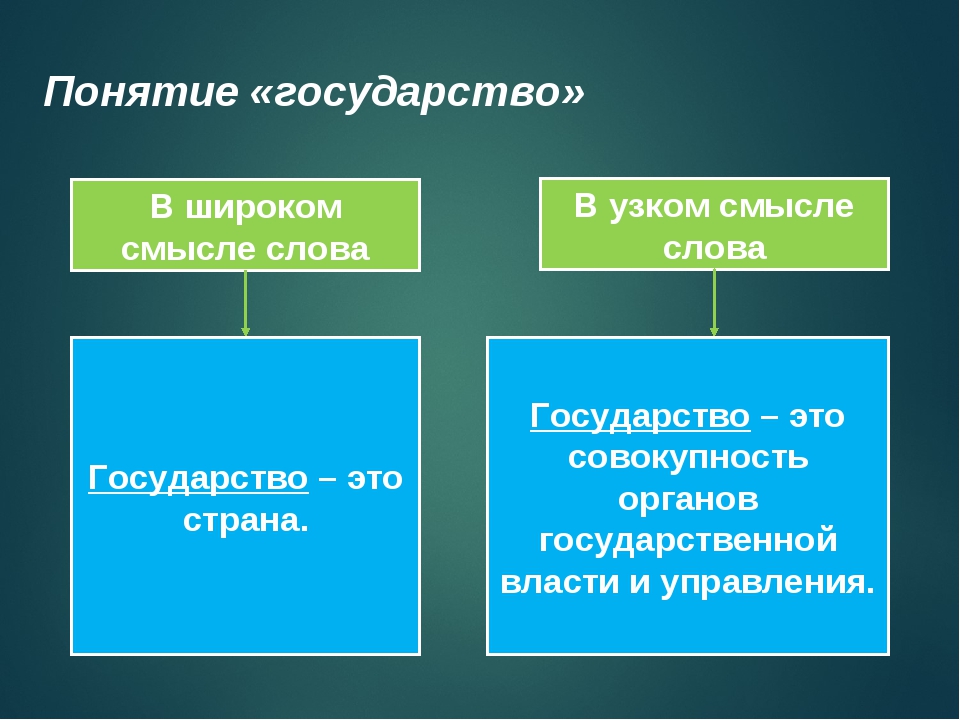 В некоторых странах его образование исторически предшествовало и способствовало классовому расслоению общества. В ходе исторического развития по мере стирания классовых противоположностей и демократизации общества государство все более становится надклассовой, общенациональной организацией;
В некоторых странах его образование исторически предшествовало и способствовало классовому расслоению общества. В ходе исторического развития по мере стирания классовых противоположностей и демократизации общества государство все более становится надклассовой, общенациональной организацией;
3) завоевание одних народов другими (Ф. Оппенгеймер, Л. Гумплович и др.). Влияние завоеваний на образование и развитие государства несомненно. Однако его не следует абсолютизировать, упуская из виду другие, часто более важные факторы;
4) демографические факторы, изменения в воспроизводстве самого человеческого рода. Имеется в виду прежде всего рост численности и плотности населения, переход народов от кочевого к оседлому образу жизни, а также запрет кровосмешения и упорядочение брачных отношений между родами. Все это повышало потребность общностей в регулировании взаимосвязей этнически близких людей;
5) психологические (рациональные и эмоциональные) факторы. Одни авторы (Гоббс) сильнейшим мотивом, побуждающим человека к созданию государства, считают страх перед агрессией со стороны других людей, опасение за жизнь и имущество. Другие же (Локк) ставят на первый план разум людей, приведший их к соглашению о создании специального органа государства, способного лучше обеспечить права людей, чем традиционные формы общежития.
Другие же (Локк) ставят на первый план разум людей, приведший их к соглашению о создании специального органа государства, способного лучше обеспечить права людей, чем традиционные формы общежития.
Контректные теории государства подтверждаются некоторыми реальными фактами. Так, например, договорная система княжения существовала в Древнем Новгороде, где с приглашаемым на определенный срок князем заключался договор, невыполнение которого могло повлечь за собой его изгнание. Под прямым влиянием теории «общественного договора» создавалось американское государство — США. И все же, несмотря на эти и некоторые другие исторические факты, реальное государство возникло не в результате добровольной передачи индивидам части своих прав специально созданному для защиты граждан и общества органу, а в ходе длительного естественноисторического развития общества;
6) антропологические факторы. Они означают, что государственная форма организации коренится в самой общественной природе человека, ее развитии.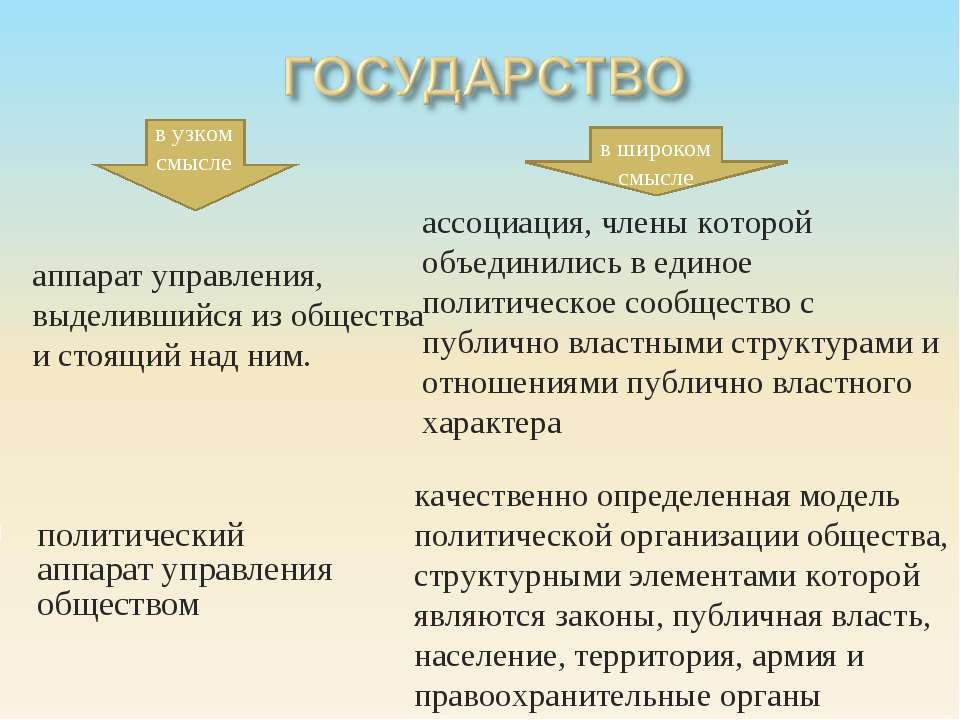
1.4. Исторические рубежи в развитии государства.
Существуя на протяжении многих тысячелетий, государство изменяется вместе с развитием всего общества, частью которого оно является. С точки зрения особенностей взаимоотношений государства и личности, воплощения в государственном устройстве рациональности, принципов свободы и прав человека, в развитии государства можно выделить два глобальных этапа: традиционный и конституционный, а также промежуточные стадии, причудливо сочетающие черты традиционных и конституционных государств, например тоталитарная государственность.
С точки зрения особенностей взаимоотношений государства и личности, воплощения в государственном устройстве рациональности, принципов свободы и прав человека, в развитии государства можно выделить два глобальных этапа: традиционный и конституционный, а также промежуточные стадии, причудливо сочетающие черты традиционных и конституционных государств, например тоталитарная государственность.
Традиционные государства возникли и существовали преимущественно стихийно, на основе обычаев и норм, уходящих корнями в глубокую древность. Они имели институционально не ограниченную власть над подданными, отрицали равноправие всех людей, не признавали личность как источник государственной власти. Типичным воплощением такого государства являлись Анархии.
Некоторые авторы, принимая во внимание глубокие отличия конституционных и неконституционных государств, предлагаю в соответствии с античной традицией, различающей государство и деспотию, называть государством лишь «организацию публичной власти, производную от гражданского общества и так или иначе ему подконтрольную».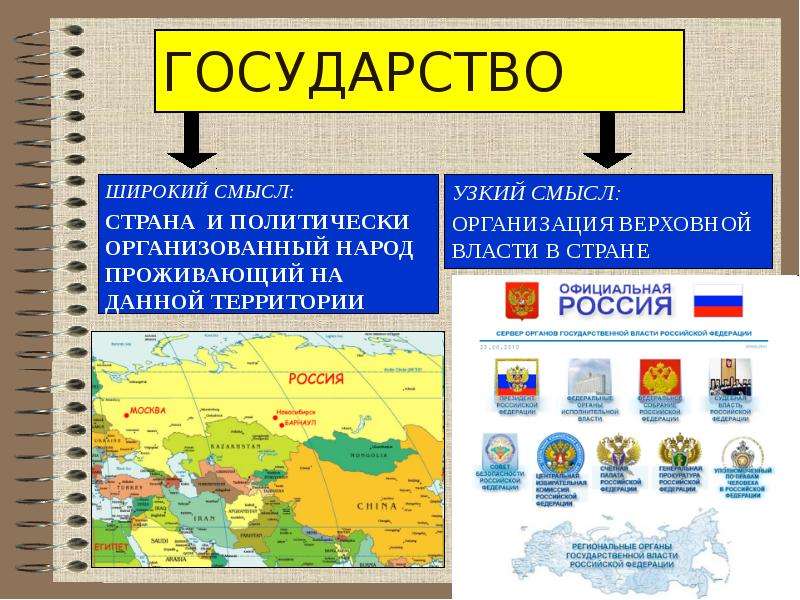 Хотя такая трактовка не учитывает разные типы государств и не разделяется большинством ученых она обоснованно указывает на принципиальную границу, отделяющую современные конституционные государства от государств по типу своего устройства и функционирования связанных с прошлым.
Хотя такая трактовка не учитывает разные типы государств и не разделяется большинством ученых она обоснованно указывает на принципиальную границу, отделяющую современные конституционные государства от государств по типу своего устройства и функционирования связанных с прошлым.
Конституционное государство является объектом сознательного человеческого формирования, управления и регулирования. Оно не стремится охватить своим регулятивным воздействием все проявления жизнедеятельности человека — его экономическую, культурную, религиозную и политическую активность и ограничивается лишь выполнением функций, делегированных гражданами и не нарушающих свободу личности.
В целом конституционный этап в развитии государства связан с его подчиненностью обществу и гражданам, с юридической очерченностью полномочий и сферы государственного вмешательства, с правовой регламентацией деятельности государства и созданием институциональных и иных гарантий прав человека. Одним словом, он связан с появлением конституции.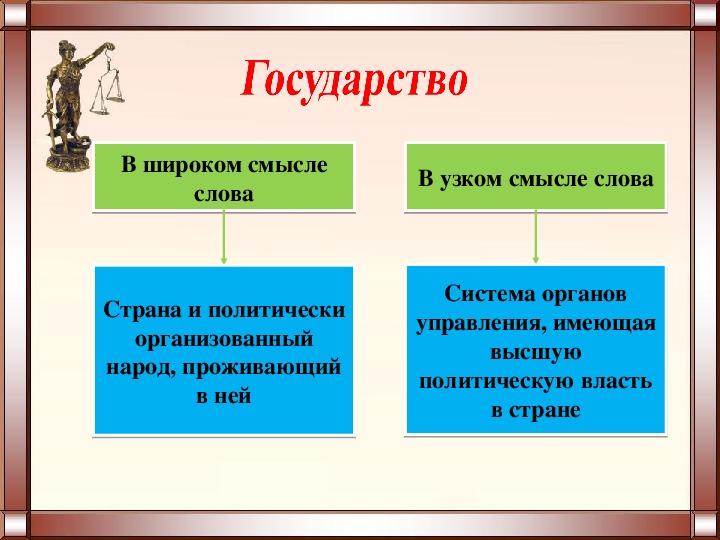
1.5. Конституция.
Сам термин «конституция» в науке употребляется в двух значениях. Первое из них, часто обозначаемое термином «реальная конституция», восходит к Аристотелю, который в своей знаменитой «Политике» трактовал конституцию как «определенный порядок для жителей одного государства». Иными словами, реальная конституция — это государственный строй, устойчивая модель государственной деятельности, определяемая тем или иным ценностно-нормативным кодексом. Этот кодекс не обязательно носит форму свода законов, присущего современным государствам. Он может иметь характер религиозно-политических заповедей или неписанных вековых традиций, которым подчиняются текущие законы государства. Понимаемый в этом значении конституционализм уходит корнями в далекое прошлое и не связывается лишь с появлением либеральных конституций в эпоху Просвещения.
Во втором, наиболее распространенном значении термин конституция» — это свод законов, юридический или нормативный акт. Она представляет собой систему зафиксированных специальном документе (или нескольких документах) относительно стабильных правил (законов), которые определяют основания, цели и устройство государства, принципы его организации и функционирования, способы политического волеобразования и принятия решений, а также положение личности в государстве.
Она представляет собой систему зафиксированных специальном документе (или нескольких документах) относительно стабильных правил (законов), которые определяют основания, цели и устройство государства, принципы его организации и функционирования, способы политического волеобразования и принятия решений, а также положение личности в государстве.
Конституция выступает как бы текстом «общественного договора», заключаемого между гражданами и государством и регламентирующего его деятельность. Она придает государству современного, конституционного типа необходимую легитимность. Обычно принимаемая при согласии подавляющего (квалифицированного) большинства населения, она фиксирует тот минимум общественного согласия, без которого невозможна свободная совместная жизнь людей в едином государстве и который обязуются уважать все граждане.
Конституции, как правило, состоят из двух важнейших частей, В первой определяются нормы взаимоотношений граждан и государства, права личности, утверждается правовое равенство всех граждан; во второй части описываются характер государства (республика, монархия, федерация и т. п.), статус различных властей, правила взаимоотношения парламента, президента, правительства и суда, а также структура и порядок функционирования органов управления.
п.), статус различных властей, правила взаимоотношения парламента, президента, правительства и суда, а также структура и порядок функционирования органов управления.
Первые конституции были приняты в 1789 г. в США (в 1791 г. Билль о правах) и во Франции (в 1789 г. «Декларация прав человека и гражданина» и в 1791 г. конституция), хотя ряд правовых документов, фактически носящих характер конституционных актов, появился еще раньше — в 1215, 1628, 1679, 1689 гг. в Англии. В современном мире лишь несколько государств (Великобритания, Израиль, Саудовская Аравия, Бутан и Оман) не имеют конституционных сводов законов.
Наличие демократической конституции — показатель подлинной конституционности государства лишь в том случае, если она реально воплощена в государственной организации и неукоснительно исполняется органами власти, учреждениями и гражданами. Завершенность процесса формирования конституционного государства, закрепление принципа ограничения его компетенций с помощью специальных институтов и законов, исходящих от народа, характеризует понятие «правовое государство».
2. Правовое социальное государство
2.1. Понятие правового государства.
Правовое государство — реальное воплощение идей и принципов конституционализма. В его основе лежи стремление оградить человека от государственного террора, насилия над совестью, мелочной опеки со стороны органов власти гарантировать индивидуальную свободу и основополагающие права личности. Это государство, ограниченное в своих действиях правом, защищающим свободу, безопасность и достоинство личности и подчиняющим власть воле суверенного народа. Взаимоотношения между личностью и властью определяются в нем конституцией, утверждающей приоритет прав человека, которые не могут быть нарушены законами государства и его действиями. Для того чтобы народ мог контролировать государство и оно не превратилось в этакого Левиафана, чудовище, господствующее над обществом, существует разделение властей: законодательной, исполнительной и судебной. Независимый суд призван защищать примат права, которое обладает всеобщностью, распространяется в равной мере на всех граждан, государственные и общественные институты.
Правовое государство формировалось постепенно на базе соответствующих идей и элементов государственности, некоторые из которых появились еще в глубокой древности. Так, о власти закона, одинакового для всех граждан, говорил в VI в. до н.э. древнегреческий архонт Солон. О соотношении естественных прав человека и законов государства писали Аристотель и Цицерон. В Древнем Риме возникли, например, такие зачатки правовой государственности, как разделение властей. Концепция же правового государства в основополагающих чертах сложилась в XVII—XIX вв. в работах Локка, Монтескье, Канта, Джефферсона и других теоретиков либерализма. Сам термин «правовое государство» окончательно утвердился в трудах немецких юристов — Т. К. Велькера, Р. фон Моля и других. Различные теории правового государства базируются на концепции гражданского общества.
2.2. Гражданское общество.
Появление правовой государственности было обусловлено развитием самого общества и стало возможным в результате вызревания гражданского общества. Сам термин «гражданское общество» употребляется как в широком, так и в узком значениях. В широком смысле гражданское общество включает всю непосредственно не охватываемую государством, его структурами часть общества, т.е. до чего «не доходят руки» государства. Оно возникает и изменяется в ходе естественно-исторического развития как автономная непосредственно не зависимая от государства сфера. Гражданское общество в широком значении совместимо не только с демократией, но и с авторитаризмом, и лишь тоталитаризм означает его полное, а чаще частичное поглощение политической властью.
Сам термин «гражданское общество» употребляется как в широком, так и в узком значениях. В широком смысле гражданское общество включает всю непосредственно не охватываемую государством, его структурами часть общества, т.е. до чего «не доходят руки» государства. Оно возникает и изменяется в ходе естественно-исторического развития как автономная непосредственно не зависимая от государства сфера. Гражданское общество в широком значении совместимо не только с демократией, но и с авторитаризмом, и лишь тоталитаризм означает его полное, а чаще частичное поглощение политической властью.
Гражданское общество в узком, собственном значении неразрывно связано с правовым государством, они не существуют друг без друга. Гражданское общество представляет собой многообразие не опосредованных государством взаимоотношений свободных и равноправных индивидов в условиях рынка и демократической правовой государственности. Это сфера свободной игры частных интересов и индивидуализма. Гражданское общество — продукт буржуазной эпохи и формируется преимущественно снизу, спонтанно, как результат раскрепощения индивидов, их превращения из подданных государства в свободных граждан-собственников, обладающих чувством личного достоинства и готовых взять на себя хозяйственную и политическую ответственность.
Гражданское общество — продукт буржуазной эпохи и формируется преимущественно снизу, спонтанно, как результат раскрепощения индивидов, их превращения из подданных государства в свободных граждан-собственников, обладающих чувством личного достоинства и готовых взять на себя хозяйственную и политическую ответственность.
Гражданское общество имеет сложную структуру, включает хозяйственные, экономические, семейно-родственные, этнические, религиозные и правовые отношения, мораль, а также не опосредованные государством политические отношения между индивидами как первичными субъектами власти, партиями, группами интересов и т.д. В гражданском обществе в отличие от государственных структур преобладают не вертикальные (подчиненности), а горизонтальные связи — отношения конкуренции и солидарности между юридически свободными и равноправными партнерами.
2.3. Отличительные черты правового государства.
Обобщая опыт возникновения и развития различных правовых государств можно выделить их следующие общие признаки: 1) наличие развитого гражданского общества; 2) ограничение сферы деятельности правового государства охраной прав и свобод личности, общественного порядка, созданием благоприятных правовых условий для хозяйственной деятельности; 3) мировоззренческий индивидуализм, ответственность каждого за собственное благополучие; 4) правовое равенство всех граждан приоритет прав человека над законами государства; 5) всеобщность права, его распространение на всех граждан, все организации и учреждения, в том числе органы государственной власти; 6) суверенитет народа, конституционно-правовая регламентация государственного суверенитета. Это означает, что именно народ является конечным источником власти, государственный же суверенитет носит представительный характер; 7) разделение законодательной, исполнительной и судебной властей государства, что не исключает единства их действий на основе процедур, предусмотренных конституцией, а также определенного верховенства законодательной власти, не нарушающие конституцию решения которой обязательны для всех; 8) приоритет в государственном регулировании гражданских отношений метода запрета над методом дозволения. Это означает, что в правовом государстве по отношению к гражданам действует принцип: «Разрешено все то, что не запрещено законом». Метод же дозволения применяется здесь лишь по отношению к самому государству, которое обязано действовать в пределах дозволенного — формально зафиксированных полномочий; 9) свобода и права других людей как единственный ограничитель свободы индивида. Правовое государство не создает абсолютной свободы личности. Свобода каждого кончается там, где нарушается свобода других.
Это означает, что именно народ является конечным источником власти, государственный же суверенитет носит представительный характер; 7) разделение законодательной, исполнительной и судебной властей государства, что не исключает единства их действий на основе процедур, предусмотренных конституцией, а также определенного верховенства законодательной власти, не нарушающие конституцию решения которой обязательны для всех; 8) приоритет в государственном регулировании гражданских отношений метода запрета над методом дозволения. Это означает, что в правовом государстве по отношению к гражданам действует принцип: «Разрешено все то, что не запрещено законом». Метод же дозволения применяется здесь лишь по отношению к самому государству, которое обязано действовать в пределах дозволенного — формально зафиксированных полномочий; 9) свобода и права других людей как единственный ограничитель свободы индивида. Правовое государство не создает абсолютной свободы личности. Свобода каждого кончается там, где нарушается свобода других.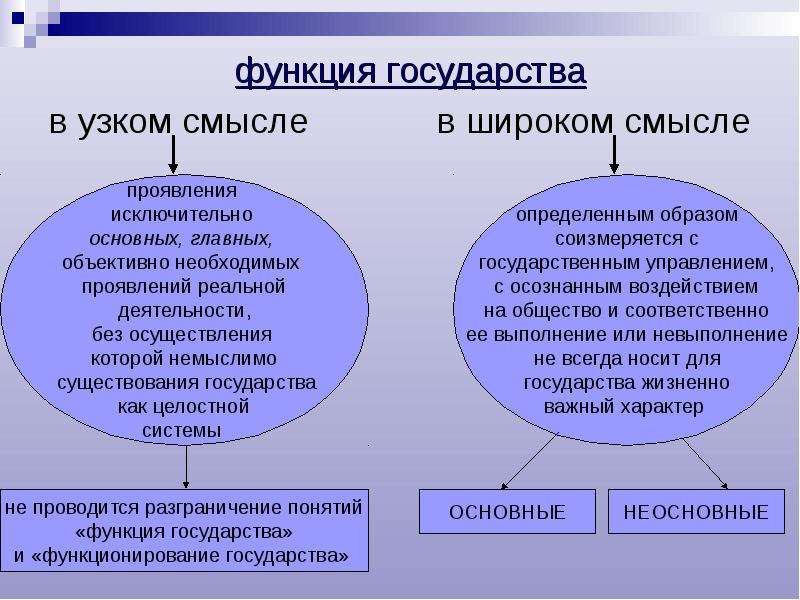
2.4. Социальное государство.
Утверждение правового государства явилось важным этапом в расширении свободы индивида и общества. Его создатели полагали, что обеспечение каждому негативной свободы (свободы от ограничений) и поощрение конкуренции пойдут на пользу всем, сделай частную собственность доступной для каждого, максимизируй индивидуальную ответственность и инициативу и приведут в конечном счете к всеобщему благополучию. Однако этого не произошло. Провозглашенные в правовых государствах индивидуальная свобода, равноправие и невмешательство государства дела гражданского общества не препятствовали монополизации экономики и ее периодическим кризисам, жестокой эксплуатации, обострению социального неравенства и классовой борьбы. глубокое фактическое неравенство обесценивало равноправие граждан, превращало использование конституционных прав в привилегию имущих классов.
Конструктивным ответом на несовершенство правового государства в его классическом либеральном варианте, а также на неудавшуюся попытку административного социализма обеспечить каждому материальную свободу и установить в обществе социальную справедливость и равенство явились теория и практика социального государства или государства всеобщего благоденствия.
Социальное государство —- это государство, стремящееся к обеспечению каждому гражданину достойных условий существования, социальной защищенности, соучастия в управлении производством, а в идеале примерно одинаковых жизненных шансов, возможностей для самореализации личности в обществе. Деятельность такого государства направлена на всеобщее благо, утверждение в обществе социальной справедливости. Оно сглаживает имущественное и иное социальное неравенство, помогает слабым и обездоленным, заботится о предоставлении каждому работы или иного источника существования, о сохранении мира в обществе, формировании благоприятной для человека жизненной среды.
Истоки социального государства восходят к социальной политике, зародившейся в далеком прошлом. Еще в древности многие правители, например римские цезари, заботились о наиболее бедных гражданах, о предоставлении плебсу «хлеба и зрелищ». Однако основное бремя обеспечения слабых и обездоленных лежало в то время на семьях и общинах. Индустриализация, урбанизация и индивидуализация общества, бесконтрольное развитие капитализма разрушили традиционные формы социального обеспечения, обострили социальные противоречия и классовую борьбу. Решение этого вопроса потребовало резкого расширения объектов социальной политики и превращения ее в одно из ведущих направлений государства. В результате этого примерно в 60-х гг. XX в. и возникли социальные государства. Их необходимой материальной предпосылкой явился высокий уровень экономического развития ведущих стран Запада, позволяющий обеспечивать прожиточный минимум каждому нуждающемуся.
Индустриализация, урбанизация и индивидуализация общества, бесконтрольное развитие капитализма разрушили традиционные формы социального обеспечения, обострили социальные противоречия и классовую борьбу. Решение этого вопроса потребовало резкого расширения объектов социальной политики и превращения ее в одно из ведущих направлений государства. В результате этого примерно в 60-х гг. XX в. и возникли социальные государства. Их необходимой материальной предпосылкой явился высокий уровень экономического развития ведущих стран Запада, позволяющий обеспечивать прожиточный минимум каждому нуждающемуся.
Деятельность современного социального государства многогранна. Это перераспределение национального дохода в пользу менее обеспеченных слоев населения, политика занятости и охрана прав работника на предприятии, социальное страхование, поддержка семьи и материнства, забота о безработных, престарелых, молодежи, развитие доступного для всех образования, здравоохранения, культуры и т.д.
Социальное государство осуществляет свои цели и принципы в форме правовой государственности, однако идет значительно дальше по пути гуманизации общества — стремится расширить права личности и наполнить правовые нормы более справедливым содержанием.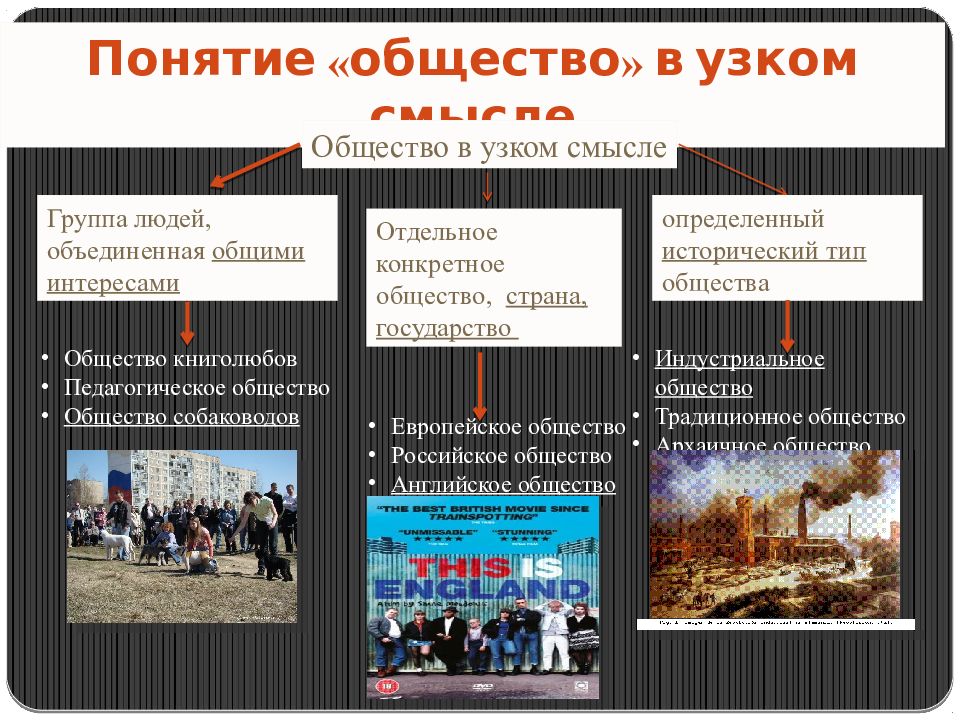 Между правовым и социальным принципами государственного устройства есть как единство, так и противоречия. Их единство состоит в том, что оба они призваны обеспечивать благо индивида: первый — физическую безопасность граждан по отношению к власти и друг к другу индивидуальную свободу и основополагающие, главным образом гражданские и политические права личности с помощью установления четких границ государственного вмешательства и гарантий против деспотии, второй — социальную безопасность материальные условия свободы и достойного существования каждого человека. Противоречия же между ними проявляются в том, что правовое государство по своему замыслу не должно вмешиваться в вопросы распределения общественного богатства, обеспечения материального и культурного благосостояния граждан, социальное же государство непосредственно занимается этим, хотя и стремится не подрывать такие основы рыночного хозяйства, как частная собственность, конкуренция, предприимчивость, индивидуальная ответственность и т.
Между правовым и социальным принципами государственного устройства есть как единство, так и противоречия. Их единство состоит в том, что оба они призваны обеспечивать благо индивида: первый — физическую безопасность граждан по отношению к власти и друг к другу индивидуальную свободу и основополагающие, главным образом гражданские и политические права личности с помощью установления четких границ государственного вмешательства и гарантий против деспотии, второй — социальную безопасность материальные условия свободы и достойного существования каждого человека. Противоречия же между ними проявляются в том, что правовое государство по своему замыслу не должно вмешиваться в вопросы распределения общественного богатства, обеспечения материального и культурного благосостояния граждан, социальное же государство непосредственно занимается этим, хотя и стремится не подрывать такие основы рыночного хозяйства, как частная собственность, конкуренция, предприимчивость, индивидуальная ответственность и т. п., не порождать массовое социальное иждивенчество. В отличие от социализма советского типа, который пытался установить благополучие всех с помощью уравнительного распределения благ, социальное государство ориентируется на обеспечение каждому достойных условий жизни в первую очередь в результате повышения эффективности производства, индивидуальной ответственности и активности.
п., не порождать массовое социальное иждивенчество. В отличие от социализма советского типа, который пытался установить благополучие всех с помощью уравнительного распределения благ, социальное государство ориентируется на обеспечение каждому достойных условий жизни в первую очередь в результате повышения эффективности производства, индивидуальной ответственности и активности.
В наши дни демократические государства стремятся найти меру оптимального сочетания правового и социального принципов. При этом консерваторы обычно делают больший акцент на правовом, а социал-демократы и близкие к ним либералы — на социальном принципе.
2.6. Тенденции развития государства.
Правовой и социальный этапы не завершают развитие конституционного государства. Некоторые политологи считают, что современные демократические государства вступают в новую, экологическую стадию. Для нее характерно выдвижение на первый план проблемы обеспечения экологической безопасности и экологических (экзистенциальных) прав личности, выживания всего человечества. В новых условиях государство вместе с общественностью призвано предотвратить ядерную и экологическую катастрофы, наладить адаптивный, поддерживающий экологическое равновесие образ жизни.
В новых условиях государство вместе с общественностью призвано предотвратить ядерную и экологическую катастрофы, наладить адаптивный, поддерживающий экологическое равновесие образ жизни.
В развитии современных государств наблюдаются две тенденции. Первая из них — деэтатистская — состоит в активизации гражданского общества, его контроля над государством, расширении влияния на него политических партий и групп интересов, ‘централизации ряда функций государства, усилении в деятельности некоторых его органов самоуправленческих начал. Вторая тенденция — этатистская — проявляется в повышении роли государства как регулятивного и интеграционного инструмента общества. Современное государство активно вмешивается в экономические, социальные и информационные процессы, с помощью налогов, инвестиционной, кредитной и иной политики стимулирует развитие производства, устраняет диспропорции в народном хозяйстве. Все более важное место в его деятельности занимает разработка стратегии и планирование общественного развития.
В государственной деятельности заметно сокращается применение принуждения. Оно все реже используется для решения крупных общественных проблем, уступая место кооперации различных социальных сил, хотя и остается важным средством в борьбе с нарушителями закона, криминальными и экстремистскими элементами. В целом же изменения, происходящие в государстве и обществе, не дают достаточных оснований говорить об отмирании государства в обозримом будущем, как это утверждают анархизм и марксизм.
Еще более велика роль государства в переходные этапы общественного развития, как это имеет место в России и других постсоциалистических странах. Здесь государство выступает главным орудием реформирования общества, поддержания стабильности и порядка. При этом оно само подвергается глубоким изменениям, приобретает новые формы организации.
3. Устройство современного государства.
3.1. Формы правления.
Строение государства традиционно характеризуется через формы правления и формы территориального (государственного) устройства. В них воплощается организация верховной власти, структура и порядок взаимоотношений высших государственных органов, должностных лиц и граждан. Формы правления делятся по способу организации власти, ее формальному источнику на монархии и республики. В монархии формальным источником власти является одно лицо. Глава государства получает свой пост по наследству независимо от избирателей или представительных органов власти
В них воплощается организация верховной власти, структура и порядок взаимоотношений высших государственных органов, должностных лиц и граждан. Формы правления делятся по способу организации власти, ее формальному источнику на монархии и республики. В монархии формальным источником власти является одно лицо. Глава государства получает свой пост по наследству независимо от избирателей или представительных органов власти
Существует несколько разновидностей монархической формы правления: абсолютная монархия (Саудовская Аравия, Катар, Оман) — всевластие главы государства; конституционная монархия — государство, в котором полномочия монарха ограничены конституцией. Конституционная монархия делится на дуалистическую (Иордания, Кувейт, Марокко), в которой монарх наделен преимущественно исполнительной властью и лишь частично -законодательной, и парламентскую, здесь монарх, хотя и считается главой государства, но фактически обладает представительскими функциями и лишь частично исполнительскими, а иногда имеет также право вето на решения парламента, которым практически не пользуется. Подавляющее большинство современных демократических монархий — парламентские монархии. Правительство формируется в них парламентским большинством и подотчетно не монарху, а парламенту.
Подавляющее большинство современных демократических монархий — парламентские монархии. Правительство формируется в них парламентским большинством и подотчетно не монарху, а парламенту.
Монархия была господствующей формой правления на протяжении тысячелетий. В специфической форме она сохраняется и сегодня почти в трети стран мира (например в Великобритании, Швеции, Дании, Испании).
3.2. Парламентская республика.
Современные парламентские монархии, отдавая дань политической традиции и поддерживая тем самым уважение граждан к государству, фактически мало отличаются от республик — второй основной формы правления. В республиках источником власти является народное большинство, высшие органы государства избираются гражданами. В современном мире существуют три основные разновидности республики: парламентская, президентская и смешанная, или полупрезидентская, хотя история знает и многие другие разновидности республик: рабовладельческую, аристократическую, советскую, теократическую и др.
Главной отличительной чертой парламентской республики является образование правительства на парламентской основе (обычно парламентским большинством) и его формальная ответственность перед парламентом. Он осуществляет по отношению к правительству ряд функций: формирует и поддерживает его; издает законы, принимаемые правительством к исполнению; вотирует (утверждает) государственный бюджет и тем самым устанавливает финансовые рамки деятельности правительства; осуществляет контроль над правительством и в случае необходимости может выразить ему вотум недоверия, что влечет за собой либо отставку правительства, либо роспуск парламента и проведение урочных выборов; критикует правительственную политику, представляет альтернативные варианты правительственных решений и всего политического курса.
Правительство обладает исполнительной властью, а нередко и законодательной инициативой, а также правом ходатайства перед президентом о роспуске парламента. (Такие ходатайства президент обычно удовлетворяет.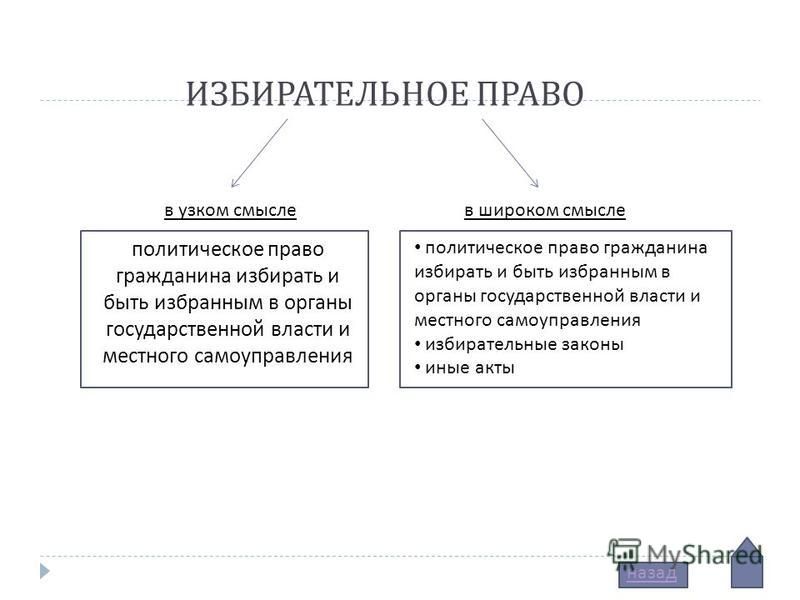 ) В большинстве стран членство в правительстве совместимо с сохранением депутатского мандата. Это позволяет привлекать в правительство не только лидеров правящих партий, но и других наиболее влиятельных депутатов парламентского большинства и тем самым контролировать парламент, получая одновременно массовую партийную поддержку. Хотя руководитель правительства (премьер-министр, канцлер) официально не является главой государства, реально он — первое лицо в политической иерархии. Президент же фактически занимает в ней более скромное место. Он может избираться либо парламентом, либо собранием выборщиков, либо непосредственно народом. Его политический вес почти не зависит от характера выборов и обычно ограничивается представительскими функциями, мало чем, отличаясь от функций главы государства в парламентских монархиях.
) В большинстве стран членство в правительстве совместимо с сохранением депутатского мандата. Это позволяет привлекать в правительство не только лидеров правящих партий, но и других наиболее влиятельных депутатов парламентского большинства и тем самым контролировать парламент, получая одновременно массовую партийную поддержку. Хотя руководитель правительства (премьер-министр, канцлер) официально не является главой государства, реально он — первое лицо в политической иерархии. Президент же фактически занимает в ней более скромное место. Он может избираться либо парламентом, либо собранием выборщиков, либо непосредственно народом. Его политический вес почти не зависит от характера выборов и обычно ограничивается представительскими функциями, мало чем, отличаясь от функций главы государства в парламентских монархиях.
3.3. Президентская республика.
Второй достаточно распространенной формой республиканского правления является президентская республика. Ее отличительный признак состоит в том, что в ней президент одновременно выступает и главой государства, и главой правительства. Он руководит внутренней и внешней политикой и является верховным главнокомандующим вооруженных сил. Президент чаще всего избирается прямо народом. Он сам (в США с одобрения сената) назначает членов кабинета министров, которые ответственны перед ним, а не перед парламентом.
Он руководит внутренней и внешней политикой и является верховным главнокомандующим вооруженных сил. Президент чаще всего избирается прямо народом. Он сам (в США с одобрения сената) назначает членов кабинета министров, которые ответственны перед ним, а не перед парламентом.
В президентской республике правительство отличается стабильностью. В ней существует жесткое разделение законодательной и исполнительной ветвей власти, их значительная самостоятельность. Парламент не может вынести правительству вотум недоверия, президент же не вправе распустить парламент. Лишь в случае серьезных антиконституционных действий или преступления со стороны президента ему может быть выражен импичмент — он досрочно отстраняется от власти. Однако процедура импичмента очень затруднена.
Отношения между парламентом и президентом основываются на системе сдержек, противовесов и взаимозависимости. Парламент может ограничивать действия президента с помощью законов и через утверждение бюджета. Президент же обычно обладает правом отлагательного вето на решения парламента.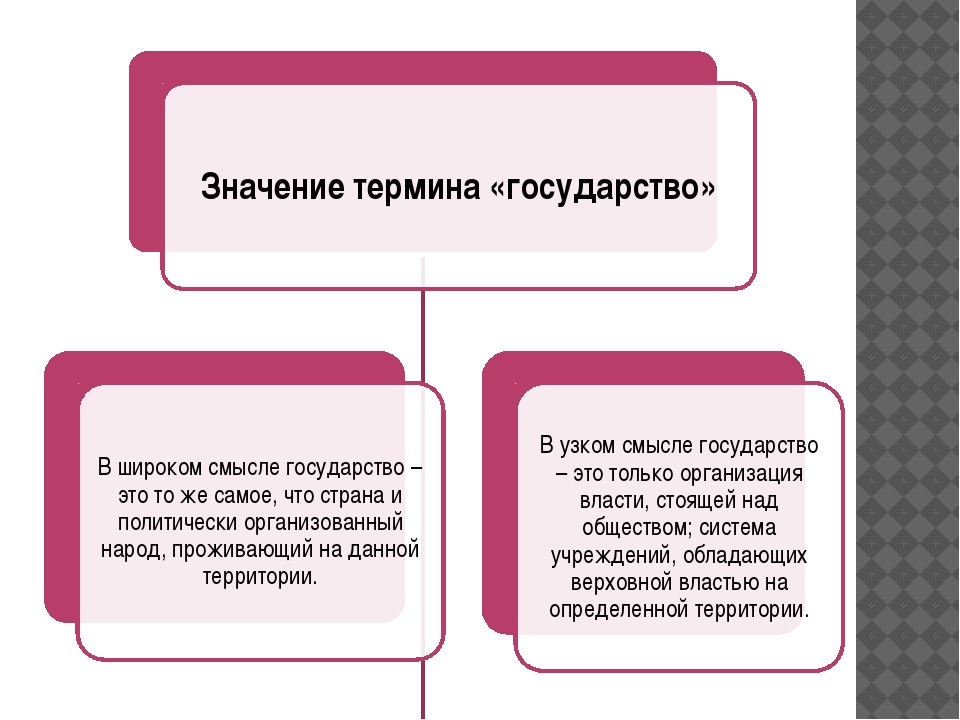 Чтобы нормально выполнять свои обязанности, и парламент, и президент вынуждены сотрудничать, находить общий язык, даже если оба этих института контролируются различными партиями.
Чтобы нормально выполнять свои обязанности, и парламент, и президент вынуждены сотрудничать, находить общий язык, даже если оба этих института контролируются различными партиями.
Президентская республика не получила распространения в Западной Европе. В странах же с длительными авторитарными традициями, прежде всего в Латинской Америке, Азии и Африке, а также на территории бывшего СССР, эта форма правления нередко вырождается в «суперпрезидентскую республику». В ней почти вся реальная власть сосредоточивается у президента, который выходит из-под контроля парламента и судебных органов и фактически обладает полудиктаторскими полномочиями, а кое-где (Заир, Малави и др.) даже объявляется пожизненным главой государства.
3.4. Полупрезидентская республика.Третьей основной разновидностью республики является полупрезидентская, или смешанная республика. Она существует в Австрии, Ирландии, Португалии, Польше, Финляндии, Франции, Болгарии и некоторых других странах. При этой форме правления сильная президентская власть сочетается с эффективным контролем парламента за деятельностью правительства. Полупрезидентская республика не имеет таких устойчивых типичных черт, как парламентская и президентская, и в различных странах тяготеет к одной из этих форм. Ее главная характерная черта — двойная ответственность правительства: перед президентом и перед парламентом.
При этой форме правления сильная президентская власть сочетается с эффективным контролем парламента за деятельностью правительства. Полупрезидентская республика не имеет таких устойчивых типичных черт, как парламентская и президентская, и в различных странах тяготеет к одной из этих форм. Ее главная характерная черта — двойная ответственность правительства: перед президентом и перед парламентом.
Классическим образцом полупрезидентской республики является Франция. В ней президент и парламент избираются независимо друг от друга. Парламент не может сместить президента, который, в свою очередь вправе распустить парламент с обязательным условием объявления даты внеочередных парламентских выборов. Президент является главой государства и верховным главнокомандующим, представляет страну на международной арене, обладает правом отлагательного вето на решения парламента, а также правом единоличного введения чрезвычайного положения, но в период действия такового он утрачивает право распустить парламент.
Президент, без согласования с парламентом, но учитывая расклад в нем политических сил, назначает главу правительства, вместе с которым они формируют кабинет министров. Глава государства председательствует на заседаниях правительства, утверждает его решения и тем самым контролирует его деятельность. Сам президент не обладает правом законодательной инициативу но таким правом пользуется премьер-министр, несущий всю ответственность за деятельность правительства. Парламент имеет возможность контролировать правительство через утверждение ежегодного бюджета, а также с помощью вынесения ему вотума недоверия.
3.5. Референдум.
Разнообразные республиканские и монархические формы государства не исчерпывают всех механизмов правления. Одним из них является институт референдума. Он предусматривает решение наиболее важных для общества вопросов посредством всенародного голосования, результаты которого имеют высший правовой статус и обязательны для исполнения всеми государственными органами. Референдум используется в качестве законодательного механизма большинством демократических государств мира, особенно на местном уровне, хотя в целом он имеет подчиненное значение по отношению к законотворческой деятельности парламента.
Референдум используется в качестве законодательного механизма большинством демократических государств мира, особенно на местном уровне, хотя в целом он имеет подчиненное значение по отношению к законотворческой деятельности парламента.
В разных странах имеются существенные расхождения в области права инициирования референдума. В одних государствах (Великобритания, Швеция, Норвегия и др.) инициаторами его являются лишь парламент и правительство, в других (например — Франция) — и президент, в третьих (Швейцария, Австрия, Италия) — непосредственно народ. В Швейцарии граждане могут добиться проведения референдума по тому или иному закону, собрав 50 тысяч подписей. Для того чтобы референдум состоялся, вовсе не обязательно участие в нем большинства населения. Всенародные голосования используются не только для принятия законов, но и для их отмены. В ряде стран (Франция, Австралия и др.) они обязательны для принятия конституционных поправок.
Хотя проведение референдумов, особенно в масштабах всей страны, — дело достаточно сложное и дорогостоящее, с их помощью народ способен непосредственно выразить свою волю, стать борцом законов, проявить инициативу. Кроме того, возможность доведения всенародного голосования заставляет государственные органы и правительство больше ориентироваться на мнение народа. Опыт многих стран показывает, что наиболее эффективно использование референдумов на региональном уровне, где агитационные и мобилизационные кампании не требуют больших финансовых затрат и поддержки со стороны крупных организаций и где люди лучше разбираются в сути решаемых вопросов. В некоторых странах (США, ФРГ и др.) этот демократический институт используется лишь субъектами федерации и более мелкими административно-территориальными единицами.
Кроме того, возможность доведения всенародного голосования заставляет государственные органы и правительство больше ориентироваться на мнение народа. Опыт многих стран показывает, что наиболее эффективно использование референдумов на региональном уровне, где агитационные и мобилизационные кампании не требуют больших финансовых затрат и поддержки со стороны крупных организаций и где люди лучше разбираются в сути решаемых вопросов. В некоторых странах (США, ФРГ и др.) этот демократический институт используется лишь субъектами федерации и более мелкими административно-территориальными единицами.
3.6. Территориальное устройство. Унитарное государство.
Территориальная организация государства характеризует соотношение целого и частей, центральных и региональных органов власти. Различают две основные формы территориального устройства государства: унитарную и федеративную. Унитарное государство представляет собой единую, политически однородную организацию, состоящую из административно-территориальных единиц, не обладающих собственной государственностью. Оно имеет единую конституцию и гражданство. Все государственные, в том числе судебные, органы составляют единую систему, действуют на основе единых правовых норм. Унитарные государства сформировались преимущественно в странах с мононациональным населением, хотя некоторые из них, например Испания, имеют в своем составе инонациональные образования, пользующиеся автономией, компетенции которой определяются центральной властью.
Оно имеет единую конституцию и гражданство. Все государственные, в том числе судебные, органы составляют единую систему, действуют на основе единых правовых норм. Унитарные государства сформировались преимущественно в странах с мононациональным населением, хотя некоторые из них, например Испания, имеют в своем составе инонациональные образования, пользующиеся автономией, компетенции которой определяются центральной властью.
Унитарные государства бывают централизованными (Великобритания, Швеция, Дания и др.) и децентрализованными (Франция, Италия, Испания). Централизованные государства могут предоставлять достаточно широкую самостоятельность (самоуправление) местным, низовым органам управления. Однако в них средние уровни управления не обладают значительной автономией и непосредственно ориентированы на выполнение решений центра. В децентрализованных же унитарных государствах крупные регионы пользуются широкой автономией и даже располагают собственными парламентами, правительствами, административно-управленческими структурами и самостоятельно решают переданные им в ведение центральными органами вопросы, как правило, в области образования, коммунального хозяйства, охраны общественного порядка и т. п. Однако в отличие от субъектов федерации в области налогооблажения их компетенции сильно ограничены, что ставит их в сильную финансовую зависимость от центра.
п. Однако в отличие от субъектов федерации в области налогооблажения их компетенции сильно ограничены, что ставит их в сильную финансовую зависимость от центра.
3.7. Федерация и конфедерация..
Главное отличие федерации от унитарного государства состоит в том, что источником власти, субъектами государственного суверенитета выступают в ней как крупные территориальные образования (штаты земли, кантоны), так и весь народ, состоящий из равноправных граждан. (В унитарном же государстве существует лишь один субъект суверенитета — народ.) Федерация — это устойчивый союз государств, самостоятельных в пределах распределенных между ними и центром компетенции, имеющих собственные законодательные, исполнительные и судебные органы и, как правило, конституцию, а часто и двойное гражданство.
По своему замыслу федеральный принцип государственного устройства призван обеспечить свободное объединение и равноправное взаимодействие общностей, обладающих значительными этническими, историко-культурными, религиозными, лингвистическими и другими особенностями; создать оптимальные возможности для выражения региональных и других интересов меньшинств, для постепенной подготовки оппозиции к выполнению общесоюзных правительственных функций; приблизить власть и управление к гражданам.
Федерация строится на основе распределения функций между ее субъектами и центром, зафиксированного в союзной конституции, которая может быть изменена только с согласия субъектов федерации. При этом одна часть вопросов является исключительной компетенцией союзных органов, другая — субъектов федерации, третья — совместной компетенцией союза и его членов. Уважение союзным руководством и всеми субъектами федерации прав и полномочий друг друга контролирует независимый суд, а также обычно двухпалатный парламент, верхняя палата которого формируется из представителей штатов (земель).
Члены федерации — соучастники общегосударственного суверенитета — фактически не обладают индивидуальным суверенитетом и правом одностороннего выхода из союзного государства. В большинстве союзных конституций содержится право федеральных органов вмешиваться во внутренние дела членов федерации в случае возникновения там чрезвычайных ситуаций: стихийных бедствий или массовых беспорядков.
Федерация как форма территориального устройства государства показала свою жизнеспособность. Этого нельзя сказать о конфедерации — постоянном союзе самостоятельных государств для осуществления конкретных совместных целей. Ее члены полностью сохраняют государственный суверенитет и передают в компетенцию союза решение лишь ограниченного числа вопросов, чаще всего в области обороны, внешней политики, транспорта и связи, денежной системы. Конфедерации существовали в США (1776—1787), Швейцарии ( до 1848 г.), Германии (1815-1867) и некоторых других странах. Эта форма государственного объединения непрочна и обычно либо эволюционирует в федерацию, либо распадается.
Этого нельзя сказать о конфедерации — постоянном союзе самостоятельных государств для осуществления конкретных совместных целей. Ее члены полностью сохраняют государственный суверенитет и передают в компетенцию союза решение лишь ограниченного числа вопросов, чаще всего в области обороны, внешней политики, транспорта и связи, денежной системы. Конфедерации существовали в США (1776—1787), Швейцарии ( до 1848 г.), Германии (1815-1867) и некоторых других странах. Эта форма государственного объединения непрочна и обычно либо эволюционирует в федерацию, либо распадается.
В последние годы на территории бывшего СССР сделана по пытка создать Содружество Независимых Государств (СНГ) — союз суверенных государств, координирующих свою деятельность в различных сферах. Такая форма государственных образований не может быть устойчивой и эффективной, поскольку не обеспечивает единство деятельности содружества, не создает властных гарантий выполнения ими своих обязательств. Опыт развития Европейского Союза свидетельствует, что долговременное объединение и сближение государств возможно на пути экономической интеграции и постепенной реализации федеративных принципов.
Формы территориального устройства и формы правления влияют на строение законодательной и исполнительной властей государства. Не менее важную значимость для нормального функционирования всего государственного механизма имеет судебная власть. Независимое правосудие призвано контролировать соблюдение конституции и законов всеми государственными и общественными учреждениями и гражданами, разрешать возникающие между ними споры, обеспечивать стабильность государственного и общественного строя.
Вся деятельность современного демократического государства строится на тесном взаимодействии с такими важнейшими институтами политической системы, как партии и группы интересов.
Литература
Геллнэр Э. Условия свободы: гражданское общество и его исторические соперники. – М., 1995.
Ильин В.В., Ахиезер А.С. Российская государственность: истоки, традиции, перспективы. – М., 1995.
Купряшин Г.Л., Соловьев А.И. Государственное управление. – М., 1996.
Макеев А. В. Политология. – М., 2000.
В. Политология. – М., 2000.
Пугачев В.П., Соловьев А.И. Введение в политологию. – М., 2000.
Соловьев А.И. Политология: Политическая теория, политические технологии. – М., 2000.
Глоссарий договорной терминологии
Акт официального подтверждения
«Акт официального подтверждения» используется в качестве эквивалента термину «ратификация», когда международная организация выражает свое согласие на обязательность для нее договора.
[Статьи 2 (1) (b-бис) и 14 Венской конвенции о праве договоров между государствами и международными организациями или между международными организациями 1986 г.]
Возражение
Любое подписавшее договор государство или договаривающееся государство может возразить против оговорки, помимо прочего, если, по его мнению, такая оговорка несовместима с объектом и целями договора. Возражающее против оговорки государство может далее заявить, что его возражение призвано препятствовать вступлению договора в силу между государством, возражающим против оговорки, и государством, сформулировавшим оговорку.
[Статьи 20-23 Венской конвенции о праве международных договоров 1969 г.]
Временное применение
Все более широкое использование положений о временном применении договоров обусловлено тем, что государства стремятся придать юридическую силу обязательствам по договору до его официальной ратификации /присоединения к нему государства. Государство обязуется соблюдать эти обязательства, относящиеся к временному применению, на сознательной и добровольной основе в соответствии с его внутренней правовой структурой.
Временное применение договора, который уже вступил в силу:
Временное применение договора, который уже вступил в силу, может произойти, когда государство обязуется придать юридическую силу обязательствам по договору на временной основе, несмотря на то, что его внутренние процедуры ратификации/присоединения еще не завершены. Как правило, государство намерено ратифицировать договор/присоединиться к нему после выполнения своих внутренних процедурных требований.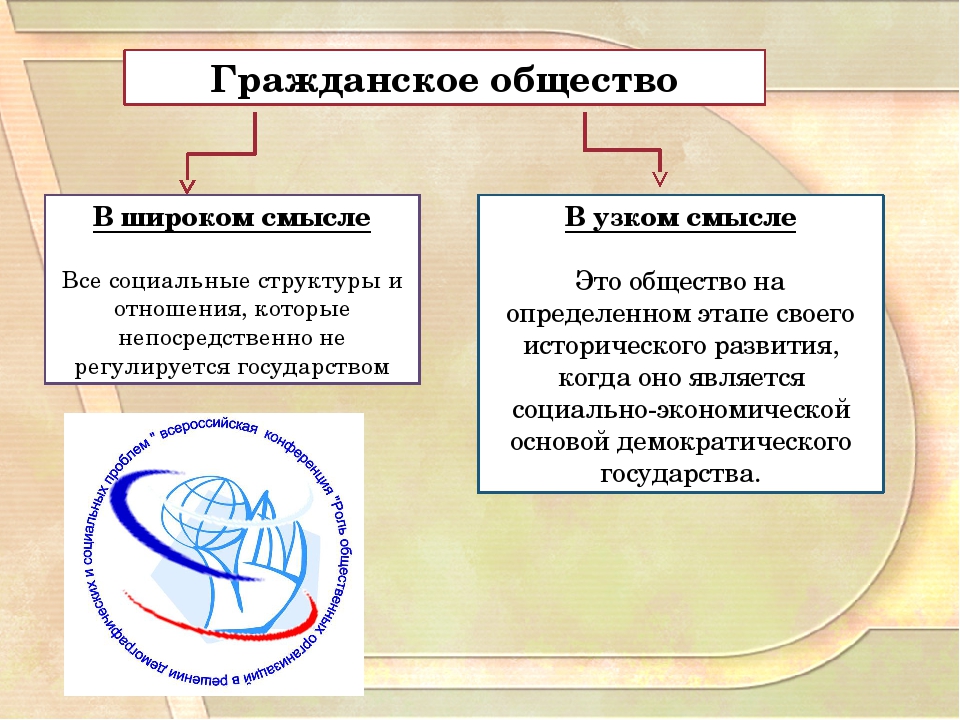 Временное применение может быть прекращено в любой момент. Напротив, государство, которое согласилось на обязательность для него договора путем ратификации/ присоединения или окончательного подписания, может отозвать свое согласие только в соответствии с правилами, изложенными в этом договоре (статьи 54, 56 Венской конвенции о праве международных договоров 1969 г.).
Временное применение может быть прекращено в любой момент. Напротив, государство, которое согласилось на обязательность для него договора путем ратификации/ присоединения или окончательного подписания, может отозвать свое согласие только в соответствии с правилами, изложенными в этом договоре (статьи 54, 56 Венской конвенции о праве международных договоров 1969 г.).
[Статья 25 Венской конвенции о праве международных договоров 1969 г.]
Временное применение договора, который еще не вступил в силу:
Временное применение договора, который еще не вступил в силу, может произойти, когда государство уведомляет государства, подписавшие договор, который еще не вступил в силу, что оно будет соблюдать юридические обязательства, предусмотренные в таком договоре, на временной основе. Государство обязуется соблюдать эти юридические обязательства на сознательной и добровольной основе в соответствии с его внутренней правовой структурой. Временное применение может быть прекращено в любой момент. Напротив, государство, которое согласилось на обязательность для него договора путем ратификации/ присоединения или окончательного подписания, может отозвать свое согласие только в соответствии с правилами, изложенными в этом договоре (статьи 54, 56 Венской конвенции о праве международных договоров 1969 г.). Государство может продолжить временное применение договора даже после его вступления в силу в отношении этого государства, до тех пор пока это государство не ратифицирует договор. Временное применение договора государством прекращается, если это государство уведомит другие государства, в отношениях между которыми временно применяется договор, о своем намерении не становиться участником договора.
Напротив, государство, которое согласилось на обязательность для него договора путем ратификации/ присоединения или окончательного подписания, может отозвать свое согласие только в соответствии с правилами, изложенными в этом договоре (статьи 54, 56 Венской конвенции о праве международных договоров 1969 г.). Государство может продолжить временное применение договора даже после его вступления в силу в отношении этого государства, до тех пор пока это государство не ратифицирует договор. Временное применение договора государством прекращается, если это государство уведомит другие государства, в отношениях между которыми временно применяется договор, о своем намерении не становиться участником договора.
[Статья 25 (2) Венской конвенции о праве международных договоров 1969 г.]
Вступление в силу
Обычно дата вступления договора в силу определяется положениями данного договора. Если в договоре не указано такой даты, предполагается, что договор должен вступить в силу, как только будет выражено согласие всех участвовавших в переговорах государств на обязательность для них договора. В двусторонних договорах может предусматриваться их вступление в силу на определенную дату, на дату подписания последней стороной, после обмена ратификационными грамотами или после обмена уведомлениями. Когда речь идет о многосторонних договорах, обычно предусматривается, что определенное число государств выражают свое согласие на вступление договора в силу. В некоторых договорах предусмотрены дополнительные условия, которые должны быть выполнены, например, в число государств, выразивших свое согласие, должна входить определенная категория государств. Кроме того, в договоре может предусматриваться, что после того, как требуемое число стран выразят свое согласие либо будут выполнены необходимые условия, должен пройти дополнительный период времени. В договоре может также предусматриваться, что после выполнения определенных условий договор вступает в силу временно.
В двусторонних договорах может предусматриваться их вступление в силу на определенную дату, на дату подписания последней стороной, после обмена ратификационными грамотами или после обмена уведомлениями. Когда речь идет о многосторонних договорах, обычно предусматривается, что определенное число государств выражают свое согласие на вступление договора в силу. В некоторых договорах предусмотрены дополнительные условия, которые должны быть выполнены, например, в число государств, выразивших свое согласие, должна входить определенная категория государств. Кроме того, в договоре может предусматриваться, что после того, как требуемое число стран выразят свое согласие либо будут выполнены необходимые условия, должен пройти дополнительный период времени. В договоре может также предусматриваться, что после выполнения определенных условий договор вступает в силу временно.
[Статья 24 Венской конвенции о праве международных договоров 1969 г.]
Двусторонние договоры
Двусторонние договоры заключаются между двумя государствами или сторонами. Тем не менее, возможно участие в двусторонних договорах более двух сторон, как, например, в двусторонних договорах между Швейцарией и Европейским союзом (ЕС), которые были заключены после отказа Швейцарии от участия в Соглашении о Европейском экономическом пространстве. Каждый из этих договоров насчитывает семнадцать сторон, однако они по-прежнему являются двусторонними, а не многосторонними, договорами. Стороны разделены на две группы: Швейцария («с одной стороны») и ЕС и его государства-члены («с другой стороны»). В договоре определены права и обязательства в отношениях между Швейцарией и ЕС и его государствами-членами по отдельности, в нем не определены никакие права и обязательства в отношениях между ЕС и его государствами-членами.
Тем не менее, возможно участие в двусторонних договорах более двух сторон, как, например, в двусторонних договорах между Швейцарией и Европейским союзом (ЕС), которые были заключены после отказа Швейцарии от участия в Соглашении о Европейском экономическом пространстве. Каждый из этих договоров насчитывает семнадцать сторон, однако они по-прежнему являются двусторонними, а не многосторонними, договорами. Стороны разделены на две группы: Швейцария («с одной стороны») и ЕС и его государства-члены («с другой стороны»). В договоре определены права и обязательства в отношениях между Швейцарией и ЕС и его государствами-членами по отдельности, в нем не определены никакие права и обязательства в отношениях между ЕС и его государствами-членами.
Декларации
Термин «декларация» употребляется для обозначения различных международных документов. Однако декларации не во всех случаях являются юридически обязательными. Этот термин часто намеренно выбирается для демонстрации того, что стороны не собираются устанавливать юридически обязательных обязательств, а лишь хотят декларировать определенные чаяния. Примером является Декларация Рио 1992 года. В то же время декларации также могут представлять собой договоры в общем смысле слова, которые должны иметь обязательную силу по международному праву. В этой связи в каждом отдельном случае необходимо определять, намеревались ли стороны устанавливать юридически обязательные обязательства. Выяснение намерений сторон зачастую может быть сложной задачей. Некоторые документы, озаглавленные «декларации», как изначально предполагалось, не должны были иметь обязательной силы, однако позднее их положения могли стать отражением международного обычного права или приобрести обязательный характер в качестве нормы обычного права. Это произошло со Всеобщей декларацией прав человека 1948 года. Декларации, которые должны иметь обязательную силу, можно классифицировать следующим образом:
Примером является Декларация Рио 1992 года. В то же время декларации также могут представлять собой договоры в общем смысле слова, которые должны иметь обязательную силу по международному праву. В этой связи в каждом отдельном случае необходимо определять, намеревались ли стороны устанавливать юридически обязательные обязательства. Выяснение намерений сторон зачастую может быть сложной задачей. Некоторые документы, озаглавленные «декларации», как изначально предполагалось, не должны были иметь обязательной силы, однако позднее их положения могли стать отражением международного обычного права или приобрести обязательный характер в качестве нормы обычного права. Это произошло со Всеобщей декларацией прав человека 1948 года. Декларации, которые должны иметь обязательную силу, можно классифицировать следующим образом:
- декларация может быть договором в собственном смысле слова. Показательным примером является Совместная декларация Соединенного Королевства и Китая по вопросу о Сянгане 1984 года;
- декларация о толковании представляет собой документ, который прилагается к договору с целью дать толкование или разъяснение его положений;
- декларацией может также называться неофициальное соглашение по какому-либо маловажному вопросу;
- ряд односторонних деклараций (заявлений) могут составлять соглашения, имеющие обязательную силу.
 Типичным примером являются декларации (заявления) в соответствии с факультативным положением Статута Международного Суда, которые создают правовую связь между делающими их сторонами, хотя они прямо и не обращены друг к другу. Еще одним примером является сделанная Египтом в 1957 году односторонняя Декларация о Суэцком канале и порядок ее действия, которая рассматривалась как обязательство международного характера.
Типичным примером являются декларации (заявления) в соответствии с факультативным положением Статута Международного Суда, которые создают правовую связь между делающими их сторонами, хотя они прямо и не обращены друг к другу. Еще одним примером является сделанная Египтом в 1957 году односторонняя Декларация о Суэцком канале и порядок ее действия, которая рассматривалась как обязательство международного характера.
 Типичным примером являются декларации (заявления) в соответствии с факультативным положением Статута Международного Суда, которые создают правовую связь между делающими их сторонами, хотя они прямо и не обращены друг к другу. Еще одним примером является сделанная Египтом в 1957 году односторонняя Декларация о Суэцком канале и порядок ее действия, которая рассматривалась как обязательство международного характера.
Типичным примером являются декларации (заявления) в соответствии с факультативным положением Статута Международного Суда, которые создают правовую связь между делающими их сторонами, хотя они прямо и не обращены друг к другу. Еще одним примером является сделанная Египтом в 1957 году односторонняя Декларация о Суэцком канале и порядок ее действия, которая рассматривалась как обязательство международного характера.Депонирование
После заключения договора письменные документы, предоставляющие официальное доказательство согласия на обязательность договора, а также оговорки и заявления, передаются на хранение депозитарию. Если договором не предусмотрено иное, то депонирование ратификационных грамот, документов о принятии, утверждении или присоединении означает согласие государства на обязательность для него договора. Для договоров с небольшим числом сторон депозитарием обычно является правительство государства, на чьей территории был подписан этот договор. Иногда в качестве депозитария выбираются различные государства. Обычно для многосторонних договоров в качестве депозитария назначается международная организация или Генеральный секретарь Организации Объединенных Наций. В обязанности депозитария входит получение всех уведомлений и документов, относящихся к договору, изучение вопроса о том, выполнены ли все официальные требования, их депонирование, регистрация договора и уведомление соответствующих сторон обо всех документах, имеющих отношение к договору.
Обычно для многосторонних договоров в качестве депозитария назначается международная организация или Генеральный секретарь Организации Объединенных Наций. В обязанности депозитария входит получение всех уведомлений и документов, относящихся к договору, изучение вопроса о том, выполнены ли все официальные требования, их депонирование, регистрация договора и уведомление соответствующих сторон обо всех документах, имеющих отношение к договору.
[Статьи 16, 76 и 77 Венской конвенции о праве международных договоров 1969 г.]
Договоры
Термин «договор» может использоваться и в общем смысле, и как специальный термин для обозначения документа с определенными характеристиками.
- Термин «договор» в общем смысле: обычно термин «договор» используется как общий термин, который охватывает все документы, связывающие участников обязательствами согласно международному праву, независимо от их официального наименования, и заключенные между субъектами международного права. Использование термина «договор» в таком широком смысле подтверждается Венской конвенцией 1969 года и Венской конвенцией 1986 года. Венская конвенция 1969 года определяет договор как «международное соглашение, заключенное между государствами в письменной форме и регулируемое международным правом, независимо от того, содержится ли такое соглашение в одном документе, в двух или нескольких связанных между собой документах, а также независимо от его конкретного наименования». Венская конвенция 1986 года включает в определение договора также международные соглашения с участием международных организаций. Для использования термина «договор» в общем смысле необходимо, чтобы документ отвечал нескольким критериям. Во-первых, он должен связывать участников обязательствами, что означает, что договаривающиеся стороны намерены создать юридические права и обязанности. Во-вторых, этот документ должен быть заключен между государствами или международными организациями, обладающими правоспособностью заключать договоры. В-третьих, он должен регулироваться международным правом. Наконец, это соглашение должно быть составлено в письменной форме. Даже до принятия в 1969 году Венской конвенции о праве международных договоров слово «договор» в общем смысле обычно применялось к соглашениям, заключенным в письменной форме.
- Договор как специальный термин: нет никаких правил, которые устанавливали бы, в каком случае международный документ должен называться договором. Обычно термин «договор» используется в отношении документов, обладающих определенным весом и требующих более официального исполнения. Типичными примерами международных документов, называемых «договорами», служат мирные договоры, договоры о границе, договоры о делимитации, договоры об экстрадиции, а также договоры о дружбе, торговле и сотрудничестве. В последние несколько десятилетий термин «договор» для международных документов используется значительно реже, вместо него используются другие термины.
 Венская конвенция 1969 года определяет договор как «международное соглашение, заключенное между государствами в письменной форме и регулируемое международным правом, независимо от того, содержится ли такое соглашение в одном документе, в двух или нескольких связанных между собой документах, а также независимо от его конкретного наименования». Венская конвенция 1986 года включает в определение договора также международные соглашения с участием международных организаций. Для использования термина «договор» в общем смысле необходимо, чтобы документ отвечал нескольким критериям. Во-первых, он должен связывать участников обязательствами, что означает, что договаривающиеся стороны намерены создать юридические права и обязанности. Во-вторых, этот документ должен быть заключен между государствами или международными организациями, обладающими правоспособностью заключать договоры. В-третьих, он должен регулироваться международным правом. Наконец, это соглашение должно быть составлено в письменной форме.
Венская конвенция 1969 года определяет договор как «международное соглашение, заключенное между государствами в письменной форме и регулируемое международным правом, независимо от того, содержится ли такое соглашение в одном документе, в двух или нескольких связанных между собой документах, а также независимо от его конкретного наименования». Венская конвенция 1986 года включает в определение договора также международные соглашения с участием международных организаций. Для использования термина «договор» в общем смысле необходимо, чтобы документ отвечал нескольким критериям. Во-первых, он должен связывать участников обязательствами, что означает, что договаривающиеся стороны намерены создать юридические права и обязанности. Во-вторых, этот документ должен быть заключен между государствами или международными организациями, обладающими правоспособностью заключать договоры. В-третьих, он должен регулироваться международным правом. Наконец, это соглашение должно быть составлено в письменной форме. Даже до принятия в 1969 году Венской конвенции о праве международных договоров слово «договор» в общем смысле обычно применялось к соглашениям, заключенным в письменной форме.
Даже до принятия в 1969 году Венской конвенции о праве международных договоров слово «договор» в общем смысле обычно применялось к соглашениям, заключенным в письменной форме.Договоры, административное управление которыми осуществляет ВОИС
Договоры, заключенные под эгидой Всемирной организации интеллектуальной собственности (ВОИС) по итогам международной конференции.
Договоры по вопросам региональной экономической интеграции
Соглашение об экономической интеграции между суверенными государствами одного и того же географического региона, например, соглашения о свободной торговле.
Изменение
Термин «изменение» подразумевает изменение определенных положений договора только во взаимоотношениях между конкретными сторонами этого договора. В отношениях между другими сторонами применяются первоначальные положения. Если в договоре ничего не говорится об изменении, они допустимы только в той мере, в какой они не затрагивают прав и обязательств других сторон договора и не противоречат объекту и целям этого договора.
[Статья 41 Венской конвенции о праве международных договоров 1969 г.]
Исправление ошибок
Если после установления аутентичности текста договора подписавшие его государства и договаривающиеся государства констатируют с общего согласия, что в нем содержится ошибка, то эта ошибка может быть исправлена путем парафирования исправленного текста договора, составления документа с изложением исправления или обмена такими документами, или исполнения исправленного текста всего договора в том же порядке, как и при исполнении первоначального текста. Если существует депозитарий, он должен уведомить все подписавшие договор государства и договаривающиеся государства о предлагаемых исправлениях. В практике Организации Объединенных Наций Генеральный секретарь как депозитарий информирует все стороны договора об ошибке и предложении исправит ее. Если по истечении соответствующего периода времени от подписавших договор государств и договаривающихся государств не последует возражений, депозитарий рассылает протокол об исправлении и обеспечивает внесение исправлений в аутентичный текст (аутентичные тексты).
Если существует депозитарий, он должен уведомить все подписавшие договор государства и договаривающиеся государства о предлагаемых исправлениях. В практике Организации Объединенных Наций Генеральный секретарь как депозитарий информирует все стороны договора об ошибке и предложении исправит ее. Если по истечении соответствующего периода времени от подписавших договор государств и договаривающихся государств не последует возражений, депозитарий рассылает протокол об исправлении и обеспечивает внесение исправлений в аутентичный текст (аутентичные тексты).
[Статья 79 Венской конвенции о праве международных договоров 1969 г.]
Конвенции
Термин «конвенция» также может иметь как общее, так и специальное значение.
- Термин «конвенция» в общем смысле: в статье 38(1)(a) Статута Международного Суда говорится о «международных конвенциях, как общих, так и специальных» как источниках права, помимо международных обычных норм и общих принципов международного права, а в качестве вторичного источника названы судебные решения и доктрины наиболее квалифицированных специалистов по публичному праву. В этом общем употреблении термин «конвенция» включает все международные соглашения так же, как и общий термин «договор». Общепризнанные принципы и нормы права также традиционно называют «конвенционным правом», с тем чтобы отличать их от других источников международного права, таких, как обычное право и общие принципы международного права. Таким образом, общий термин «конвенция» является синонимом общего термина «договор».
- Конвенция как специальный термин: если в прошлом веке термин «конвенция» традиционно употреблялся для обозначения двусторонних соглашений, то в настоящее время он, как правило, используется для обозначения официальных многосторонних договоров с участием большого числа сторон. Конвенции, как правило, открыты для участия международного сообщества в целом или большого числа государств. Обычно документы, заключаемые под эгидой международных организаций, называются «конвенции» (например, Конвенция о биологическом разнообразии 1992 года, Конвенция Организации Объединенных Наций по морскому праву 1982 года, Венская конвенция о праве международных договоров1969 года). То же самое относится к документам, принимаемым органом международной организации (например, Конвенция МОТ относительно равного вознаграждения мужчин и женщин за труд равной ценности 1951 года, принятая Международной конференцией труда, или Конвенция о правах ребенка 1989 года, принятая Генеральной Ассамблеей Организации Объединенных Наций).
 В этом общем употреблении термин «конвенция» включает все международные соглашения так же, как и общий термин «договор». Общепризнанные принципы и нормы права также традиционно называют «конвенционным правом», с тем чтобы отличать их от других источников международного права, таких, как обычное право и общие принципы международного права. Таким образом, общий термин «конвенция» является синонимом общего термина «договор».
В этом общем употреблении термин «конвенция» включает все международные соглашения так же, как и общий термин «договор». Общепризнанные принципы и нормы права также традиционно называют «конвенционным правом», с тем чтобы отличать их от других источников международного права, таких, как обычное право и общие принципы международного права. Таким образом, общий термин «конвенция» является синонимом общего термина «договор». То же самое относится к документам, принимаемым органом международной организации (например, Конвенция МОТ относительно равного вознаграждения мужчин и женщин за труд равной ценности 1951 года, принятая Международной конференцией труда, или Конвенция о правах ребенка 1989 года, принятая Генеральной Ассамблеей Организации Объединенных Наций).
То же самое относится к документам, принимаемым органом международной организации (например, Конвенция МОТ относительно равного вознаграждения мужчин и женщин за труд равной ценности 1951 года, принятая Международной конференцией труда, или Конвенция о правах ребенка 1989 года, принятая Генеральной Ассамблеей Организации Объединенных Наций).Меморандумы о взаимопонимании
Меморандум о взаимопонимании представляет собой международный документ менее официального характера. В нем часто излагаются практические мероприятия, которые осуществляются в рамках международного соглашения. Он также используется для регулирования технических вопросов или конкретных деталей. Меморандум о взаимопонимании обычно состоит из одного документа и не требует ратификации. Он заключается между государствами и/или международными организациями. Как правило, Организация Объединенных Наций заключает меморандумы о взаимопонимании с государствами-членами, чтобы организовывать свои операции по поддержанию мира или проводить конференции Организации Объединенных Наций. Кроме того, Организация Объединенных Наций заключает меморандумы о взаимопонимании в отношении сотрудничества с другими международными организациями.
Кроме того, Организация Объединенных Наций заключает меморандумы о взаимопонимании в отношении сотрудничества с другими международными организациями.
Многосторонние договоры
Многосторонние договоры представляют собой соглашения между двумя и более сторонами. Часто они заключаются в результате международной конференции или встречи государств, которая проводится под эгидой международной организации.
Модус вивенди
Модус вивенди представляет собой документ, содержащий временное или предварительное международное соглашение, которое предполагается заменить другим, более постоянного характера и более подробным. Обычно составляется неофициально и никогда не требует ратификации.
Обмен письмами или нотами
Государства могут выразить свое согласие на обязательность для них договора путем обмена письмами или нотами. Основная особенность этой процедуры состоит в том, что подписи обеих сторон проставляются не на одном письме или ноте, а на двух отдельных письмах или нотах. Поэтому соглашение заключается в обмене этими письмами или нотами, при этом каждая из сторон оставляет у себя одно письмо или ноту, подписанное(ую) представителем другой стороны. На практике второе письмо или нота (обычно ответное письмо или нота) воспроизводит текст первого(ой). В двустороннем договоре стороны могут также обменяться письмами или нотами, указывающими на то, что они завершили все необходимые внутригосударственные процедуры.
Поэтому соглашение заключается в обмене этими письмами или нотами, при этом каждая из сторон оставляет у себя одно письмо или ноту, подписанное(ую) представителем другой стороны. На практике второе письмо или нота (обычно ответное письмо или нота) воспроизводит текст первого(ой). В двустороннем договоре стороны могут также обменяться письмами или нотами, указывающими на то, что они завершили все необходимые внутригосударственные процедуры.
[Статья 13 Венской конвенции о праве международных договоров 1969 г.]
Оговорка
Оговорка — это заявление, сделанное государством, посредством которого оно намерено исключить или изменить юридические последствия определенных положений договора в процессе их применения к этому государству. Оговорка может позволить государству участвовать в многостороннем договоре, в котором в ином случае оно не могло бы или не желало бы участвовать. Государства могут делать оговорки к договору в момент его подписания, ратификации, принятия, утверждения или присоединения к нему. Оговорки не должны быть несовместимыми с объектом и целями договора. Кроме того, договор может запрещать делать оговорки или разрешать лишь некоторые оговорки.
Оговорки не должны быть несовместимыми с объектом и целями договора. Кроме того, договор может запрещать делать оговорки или разрешать лишь некоторые оговорки.
[Статьи 2 (1) (d) и 19-23 Венской конвенции о праве международных договоров 1969 г.]
Окончательное подписание
Если договор не предполагает ратификации, принятия или утверждения, окончательное подписание выражает согласие государства на обязательность для него договора. Большинство двусторонних договоров, которые касаются вопросов скорее повседневного, чем политического характера, вступают в силу путем окончательного подписания, без процедуры ратификации.
[Статья 12 Венской конвенции о праве международных договоров 1969 г.]
Организация-инициатор
Организация, ставшая инициатором создания договора.
Пересмотр
Пересмотр по сути означает поправку. Однако в некоторых договорах положения о пересмотре предусматриваются отдельно от положений, касающихся поправок (см., в частности, статью 109 Устава Организации Объединенных Наций). В таком случае пересмотр обычно означает общую корректировку договора в соответствии с меняющимися обстоятельствами, в то время как термин «поправка» подразумевает изменение конкретных положений.
В таком случае пересмотр обычно означает общую корректировку договора в соответствии с меняющимися обстоятельствами, в то время как термин «поправка» подразумевает изменение конкретных положений.
Подписание ad referendum
Представитель может подписать договор «ad referendum», т.е. при условии подтверждения подписания его государством. В таком случае подписание становится полным после подтверждения ответственным органом.
[Статья 12 (2) (b) Венской конвенции о праве международных договоров 1969 г.]
Подписание, подлежащее ратификации, принятию или утверждению
Если подписание подлежит ратификации, принятию или утверждению, такое подписание не служит выражением согласия на обязательность договора. Тем не менее, оно является способом установления аутентичности текста и выражения готовности подписавшего договор государства к продолжению процесса заключения договора. Подписание договора дает подписавшему его государству право на дальнейшую ратификацию, принятие или утверждение. Оно такое создает обязательство добросовестно воздерживаться от действий, которые лишили бы договор его объекта и цели.
Оно такое создает обязательство добросовестно воздерживаться от действий, которые лишили бы договор его объекта и цели.
[Статьи 10 и 18 Венской конвенции о праве международных договоров 1969 г.]
Полномочия
«Полномочия» означают документ, который исходит от компетентного органа государства и посредством которого одно или несколько лиц назначаются представлять это государство в целях ведения переговоров, принятия текста договора или установления его аутентичности, выражения согласия этого государства на обязательность для него договора или в целях совершения любого другого акта, относящегося к договору. Главы государств, главы правительств и министры иностранных дел считаются представляющими свое государство в целях всех актов, относящихся к договору, без необходимости предъявлять полномочия. От глав дипломатических представительств не требуется предъявлять полномочия в целях принятия текста договора между аккредитующим государством и государством, при котором они аккредитованы. Аналогичным образом не требуется предъявлять полномочия представителям, уполномоченным государствами представлять их на международной конференции или в международной организации, или в одном из ее органов, в целях принятия текста договора на такой конференции, в такой организации или в таком органе.
Аналогичным образом не требуется предъявлять полномочия представителям, уполномоченным государствами представлять их на международной конференции или в международной организации, или в одном из ее органов, в целях принятия текста договора на такой конференции, в такой организации или в таком органе.
[Статьи 2 (1) (c) и 7 Венской конвенции о праве международных договоров 1969 г.]
Поправка
Термин «поправка» означает официальное изменение положений договора, которое затрагивает все стороны данного соглашения. Такие изменения должны быть произведены с соблюдением тех же формальностей, что и при составлении первоначального текста договора. Во многих многосторонних договорах предусмотрены конкретные требования, которые необходимо выполнить для того, чтобы внесенные поправки были приняты. В отсутствие таких положений для внесения поправок необходимо согласие всех сторон.
[Статья 40 Венской конвенции о праве международных договоров 1969 г.]
Правопреемство
Правопреемство возникает в случаях, когда одно государство прекращает существование или утрачивает контроль над частью своей территории, а другое государство возникает или приобретает контроль над территорией, утраченной первым государством. В таком случае главный вопрос состоит в том, примет ли государство-правопреемник международные обязательства первого государства. Изменения в форме государственного управления одного государства, например, демократическая форма правления вместо монархии, не меняют и не прекращают действия обязательств, принятых на себя прежним правительством.
В таком случае главный вопрос состоит в том, примет ли государство-правопреемник международные обязательства первого государства. Изменения в форме государственного управления одного государства, например, демократическая форма правления вместо монархии, не меняют и не прекращают действия обязательств, принятых на себя прежним правительством.
Однако в случае прекращения существования государства заключенные им договора, как правило, прекращают действие, а договора, заключенные государством-правопреемником, применяются к этой территории. Это включает в себя такие политические договоры, как альянсы, для которых требуется существование заключившего их государства. Однако некоторые обязательства, такие как соглашения, касающиеся границ или других вопросов местного значения, переходят к государству-правопреемнику. Сложнее обстоит вопрос с сохранением юридической силы договоров, предоставляющих концессию или права по контракту. Мнения исследователей по этому аспекту правопреемства расходятся, и практика государств также различается. Таким образом, в каждом конкретном случае необходимо рассматривать вопрос по существу, чтобы определить, являются ли права и обязанности по контракту или концессии такими, что государство-правопреемник является связанным обязательствами государства-предшественника.
Таким образом, в каждом конкретном случае необходимо рассматривать вопрос по существу, чтобы определить, являются ли права и обязанности по контракту или концессии такими, что государство-правопреемник является связанным обязательствами государства-предшественника.
Принятие
«Принятие» представляет собой официальный акт, посредством которого устанавливаются форма и содержание предложенного текста договора. Как правило, принятие текста договора происходит путем выражения согласия государств, участвующих в процессе заключения этого договора. Договоры, переговоры по которым ведутся в рамках международной организации, обычно принимаются посредством резолюции представительного органа организации, членский состав которой в той или иной степени соответствует потенциальному составу участников указанного договора. Договор может быть также принят международной конференцией, специально созданной для утверждения договора, путем голосования за него двух третей государств, присутствующих и участвующих в голосовании, если тем же большинством голосов они не решили применить иное правило.
[Статья 9 Венской конвенции о праве международных договоров 1969 г.]
Принятие или утверждение
Документы о «принятии» или «утверждении» договора имеют такую же юридическую силу, как и ратификация, и, следовательно, выражают согласие государства на обязательность для него договора. В практике некоторых государств принятие и утверждение используются вместо ратификации в случаях, когда конституционное право на национальном уровне не содержит требования о ратификации договора главой государства.
[Статьи 2 (1) (b) и 14 (2) Венской конвенции о праве международных договоров 1969 г.]
Присоединение
«Присоединение» означает акт, посредством которого государство принимает предложение или возможность стать участником договора, по которому другие государства уже провели переговоры и которое подписали другие государства. Оно имеет такую же юридическую силу, как и ратификация. Присоединение к договору обычно происходит после его вступления в силу. Кроме того, Генеральный секретарь Организации Объединенных Наций как депозитарий принимал присоединение к некоторым конвенциям до их вступления в силу. Условия, по которым присоединение может иметь место, и соответствующая процедура зависят от положений договора. Договор может предусматривать присоединение всех других государств либо ограниченного или определенного числа государств. В отсутствие такого положения присоединение может иметь место только в случае, если участвующие в переговорах государства договорились или впоследствии договорятся по этому вопросу в отношении указанного государства.
Условия, по которым присоединение может иметь место, и соответствующая процедура зависят от положений договора. Договор может предусматривать присоединение всех других государств либо ограниченного или определенного числа государств. В отсутствие такого положения присоединение может иметь место только в случае, если участвующие в переговорах государства договорились или впоследствии договорятся по этому вопросу в отношении указанного государства.
[Статьи 2 (1) (b) и 15 Венской конвенции о праве международных договоров 1969 г.]
Протоколы
Термин «протокол» употребляется для обозначения соглашений, имеющих менее официальный характер, чем соглашения, называемые «договорами» или «конвенциями». Этот термин мог бы использоваться для обозначения следующих видов документов:
- протокол о подписании представляет собой субсидиарный по отношению к договору документ, составленный теми же сторонами. Такой протокол касается вспомогательных вопросов, таких, как толкование отдельных положений договора, официальных положений, которые не были включены в договор, или решения технических вопросов. Ратификация договора, как правило, будет ipso facto включать ратификацию такого протокола;
- факультативный протокол к договору представляет собой документ, который предусматривает дополнительные по отношению к договору права и обязательства. Обычно он принимается в тот же день, однако носит самостоятельный характер и подлежит отдельной ратификации. Такие протоколы позволяют некоторым сторонам договора устанавливать в отношениях между собой такой обязательственный механизм, который выходит за рамки общего договора и принимается не всеми его участниками, в результате чего создается «двуступенчатая система». Известным примером является Факультативный протокол к Международному пакту о гражданских и политических правах 1966 года;
- протокол, основанный на рамочном договоре, представляет собой содержащий особые материальные обязательства документ, который предназначен для достижения общих целей предшествующей рамочной или зонтичной конвенции. Такие протоколы предусматривают упрощенный и ускоренный процесс заключения договора и особенно популярны в области международного экологического права. В качестве примера можно привести Монреальский протокол по веществам, разрушающим озоновый слой, 1987 года, принятый на основе статей 2 и 8 Венской конвенции об охране озонового слоя 1985 года;
- протокол о внесении изменений представляет собой документ, содержащий положения, которые вносят изменения в один или несколько предшествующих договоров, напримерПротокол 1946 года о внесении изменений в соглашения, конвенции и протоколы о наркотиках;
- протокол в качестве дополнительного договора представляет собой документ, который содержит дополнительные положения к предшествующему договору, например Протокол, касающийся статуса беженцев, 1967 года к Конвенции о статусе беженцев 1951 года;
- протокол (Proces-Verbal) представляет собой документ, который закрепляет определенные договоренности, достигнутые договаривающимися сторонами.
 Ратификация договора, как правило, будет ipso facto включать ратификацию такого протокола;
Ратификация договора, как правило, будет ipso facto включать ратификацию такого протокола; В качестве примера можно привести Монреальский протокол по веществам, разрушающим озоновый слой, 1987 года, принятый на основе статей 2 и 8 Венской конвенции об охране озонового слоя 1985 года;
В качестве примера можно привести Монреальский протокол по веществам, разрушающим озоновый слой, 1987 года, принятый на основе статей 2 и 8 Венской конвенции об охране озонового слоя 1985 года;Ратификация
Ратификация означает международный акт, посредством которого государство выражает свое согласие на обязательность для него договора, если стороны намерены выразить свое согласие посредством такого акта. В случае двусторонних договоров ратификация обычно совершается путем обмена необходимыми документами, а при заключении многосторонних договоров порядок обычно состоит в том, что депозитарий проводит сбор ратификационных грамот всех государств с постоянным информированием о процессе всех сторон. Институт ратификации предоставляет государствам временные рамки для получения требуемого одобрения на внутригосударственном уровне и принятия законодательства, которое является необходимым для вступления данного договора в силу в этом государстве.
В случае двусторонних договоров ратификация обычно совершается путем обмена необходимыми документами, а при заключении многосторонних договоров порядок обычно состоит в том, что депозитарий проводит сбор ратификационных грамот всех государств с постоянным информированием о процессе всех сторон. Институт ратификации предоставляет государствам временные рамки для получения требуемого одобрения на внутригосударственном уровне и принятия законодательства, которое является необходимым для вступления данного договора в силу в этом государстве.
[Статьи 2 (1) (b), 14 (1) и 16 Венской конвенции о праве международных договоров 1969 г.]
Региональные договоры по вопросам ИС
Соглашение между суверенными государствами одного и того же географического региона, касающееся конкретных вопросов из области интеллектуальной собственности.
Регистрация и опубликование договоров
Статья 102 Устава Организации Объединенных Наций предусматривает, что «всякий договор и всякое международное соглашение, заключенные любым Членом Организации после вступления в силу настоящего Устава, должны быть, при первой возможности, зарегистрированы в Секретариате и им опубликованы». В случае нерегистрации стороны не могут ссылаться на такой договор или соглашение ни в одном из органов Организации Объединенных Наций. Регистрация содействует обеспечению прозрачности и доступности текстов договоров для широкой публики. Статья 102 Устава ООН и предшествовавшая ей статья 18 Пакта Лиги Наций берут начало в одном из «Четырнадцати пунктов» Вудро Вильсона, где автор излагает свою идею создания Лиги Наций: «открытые мирные договоры, открыто обсуждённые, после которых не будет никаких тайных международных соглашений какого-либо рода, а дипломатия всегда будет действовать откровенно и на виду у всех».
В случае нерегистрации стороны не могут ссылаться на такой договор или соглашение ни в одном из органов Организации Объединенных Наций. Регистрация содействует обеспечению прозрачности и доступности текстов договоров для широкой публики. Статья 102 Устава ООН и предшествовавшая ей статья 18 Пакта Лиги Наций берут начало в одном из «Четырнадцати пунктов» Вудро Вильсона, где автор излагает свою идею создания Лиги Наций: «открытые мирные договоры, открыто обсуждённые, после которых не будет никаких тайных международных соглашений какого-либо рода, а дипломатия всегда будет действовать откровенно и на виду у всех».
[Статья 80 Венской конвенции о праве международных договоров 1969 г.]
Секретариат, выполняющий функции административного управления
Секретариат, который впоследствии осуществляет административное управление договором.
Соглашения
Термин «соглашение» может иметь может иметь как общее, так и специальное значение. Кроме того, в законодательстве в области региональной экономической интеграции этот термин приобрел специальное значение:
- термин «соглашение» в общем смысле: в самом широком смысле термин «международное соглашение» используется в Венской конвенции о праве международных договоров 1969 г. С одной стороны, в этой конвенции договоры определяются как «международные соглашения», имеющие определенные характеристики. С другой стороны, в этой же конвенции термин «международные соглашения» используется применительно к документам, которые не подпадают под такое определение договора. В статье 3 говорится о «международных соглашениях не в письменной форме». Несмотря на то, что такие устные соглашения встречаются редко, они могут иметь такую же обязательную силу, как и договоры, в зависимости от намерений сторон. Примером устного соглашения может служить обещание, данное министром иностранных дел одного государства своему коллеге из другого государства. Таким образом, термин «международное соглашение» в его общем смысле охватывает самый широкий круг международных документов;
- соглашение как специальный термин: соглашения обычно носят менее официальный характер и касаются более узкого круга вопросов по сравнению с договорами. В соответствии с общей тенденцией термин «соглашение» применяется к двусторонним договорам или многосторонним договорам с ограниченным числом участников. Он специально используется для обозначения документов технического или административного характера, которые подписываются представителями правительственных департаментов, но не подлежат ратификации. Как правило, соглашения касаются вопросов экономического, культурного, научного и технического сотрудничества. Также в соглашениях часто рассматриваются вопросы финансового характера, например, избежание двойного налогообложения, обеспечение гарантий инвестиций и оказание финансовой помощи. ООН и другие международные организации регулярно заключает соглашения с принимающей страной международной конференции или сессии представительного органа Организации. Кроме того, термин «соглашение» используется в названии всеобъемлющих многосторонних соглашений (например, товарных соглашений), особенно в сфере международного экономического права. Использование термина «соглашение» постепенно расширялось в последние несколько десятков лет, и в настоящее время этим словом называется подавляющее большинство международных документов;
- соглашения в программах региональной интеграции: программы региональной интеграции основаны на общих рамочных договорах, в которых устанавливаются основные принципы сотрудничества. Более поздние международные документы, призванные внести изменения в эти рамочные договоры (например, касающиеся присоединения или пересмотра), также именуются договорами. Документы, которые заключаются в рамках конституционного договора или органами региональной организации, обычно называют «соглашениями», чтобы подчеркнуть их отличие от конституционного договора. Например, Римский договор 1957 г. играет роль конституции Европейского сообщества, а договоры, которые ЕС заключает с другими нациями, обычно называются соглашениями. Кроме того, Латиноамериканская ассоциация интеграции (ЛАИ) была создана Договором Монтевидео 1980 г., однако субрегиональные документы, которые были заключены в его рамках, называются соглашениями.
 С одной стороны, в этой конвенции договоры определяются как «международные соглашения», имеющие определенные характеристики. С другой стороны, в этой же конвенции термин «международные соглашения» используется применительно к документам, которые не подпадают под такое определение договора. В статье 3 говорится о «международных соглашениях не в письменной форме». Несмотря на то, что такие устные соглашения встречаются редко, они могут иметь такую же обязательную силу, как и договоры, в зависимости от намерений сторон. Примером устного соглашения может служить обещание, данное министром иностранных дел одного государства своему коллеге из другого государства. Таким образом, термин «международное соглашение» в его общем смысле охватывает самый широкий круг международных документов;
С одной стороны, в этой конвенции договоры определяются как «международные соглашения», имеющие определенные характеристики. С другой стороны, в этой же конвенции термин «международные соглашения» используется применительно к документам, которые не подпадают под такое определение договора. В статье 3 говорится о «международных соглашениях не в письменной форме». Несмотря на то, что такие устные соглашения встречаются редко, они могут иметь такую же обязательную силу, как и договоры, в зависимости от намерений сторон. Примером устного соглашения может служить обещание, данное министром иностранных дел одного государства своему коллеге из другого государства. Таким образом, термин «международное соглашение» в его общем смысле охватывает самый широкий круг международных документов; Он специально используется для обозначения документов технического или административного характера, которые подписываются представителями правительственных департаментов, но не подлежат ратификации. Как правило, соглашения касаются вопросов экономического, культурного, научного и технического сотрудничества. Также в соглашениях часто рассматриваются вопросы финансового характера, например, избежание двойного налогообложения, обеспечение гарантий инвестиций и оказание финансовой помощи. ООН и другие международные организации регулярно заключает соглашения с принимающей страной международной конференции или сессии представительного органа Организации. Кроме того, термин «соглашение» используется в названии всеобъемлющих многосторонних соглашений (например, товарных соглашений), особенно в сфере международного экономического права. Использование термина «соглашение» постепенно расширялось в последние несколько десятков лет, и в настоящее время этим словом называется подавляющее большинство международных документов;
Он специально используется для обозначения документов технического или административного характера, которые подписываются представителями правительственных департаментов, но не подлежат ратификации. Как правило, соглашения касаются вопросов экономического, культурного, научного и технического сотрудничества. Также в соглашениях часто рассматриваются вопросы финансового характера, например, избежание двойного налогообложения, обеспечение гарантий инвестиций и оказание финансовой помощи. ООН и другие международные организации регулярно заключает соглашения с принимающей страной международной конференции или сессии представительного органа Организации. Кроме того, термин «соглашение» используется в названии всеобъемлющих многосторонних соглашений (например, товарных соглашений), особенно в сфере международного экономического права. Использование термина «соглашение» постепенно расширялось в последние несколько десятков лет, и в настоящее время этим словом называется подавляющее большинство международных документов; Более поздние международные документы, призванные внести изменения в эти рамочные договоры (например, касающиеся присоединения или пересмотра), также именуются договорами. Документы, которые заключаются в рамках конституционного договора или органами региональной организации, обычно называют «соглашениями», чтобы подчеркнуть их отличие от конституционного договора. Например, Римский договор 1957 г. играет роль конституции Европейского сообщества, а договоры, которые ЕС заключает с другими нациями, обычно называются соглашениями. Кроме того, Латиноамериканская ассоциация интеграции (ЛАИ) была создана Договором Монтевидео 1980 г., однако субрегиональные документы, которые были заключены в его рамках, называются соглашениями.
Более поздние международные документы, призванные внести изменения в эти рамочные договоры (например, касающиеся присоединения или пересмотра), также именуются договорами. Документы, которые заключаются в рамках конституционного договора или органами региональной организации, обычно называют «соглашениями», чтобы подчеркнуть их отличие от конституционного договора. Например, Римский договор 1957 г. играет роль конституции Европейского сообщества, а договоры, которые ЕС заключает с другими нациями, обычно называются соглашениями. Кроме того, Латиноамериканская ассоциация интеграции (ЛАИ) была создана Договором Монтевидео 1980 г., однако субрегиональные документы, которые были заключены в его рамках, называются соглашениями.Стороны
Согласно публикации «Многосторонние договоры, депонированные Генеральному секретарю», термин «Стороны», который встречается в заглавии каждого договора, включает в себя и «Договаривающиеся государства», и «Стороны». В общем смысле термин «Договаривающиеся государства» означает государства и другие субъекты, способные заключить договор, которые выразили свое согласие на обязательность для них договора, если этот договор еще не вступил в силу или не вступил в силу для данных государств и субъектов; термин «Стороны» означает государства и другие субъекты, способные заключить договор, которые выразили свое согласие на обязательность для них договора, если этот договор вступил в силу для данных государств и субъектов.
Уведомление
Термин «уведомление» означает формальность, посредством которой государство или международная организация сообщает об определенных фактах или событиях, имеющих значение с точки зрения права. К уведомлению все чаще прибегают в качестве средства выражения окончательного согласия. Вместо обмена документами или их депонирования государства могут удовольствоваться уведомлением другой стороны или депозитария о своем согласии. Однако может потребоваться представить уведомление обо всех прочих актах и документах, относящихся к течению срока действия договора.
[Статьи 16 (c), 78 и т.д. Венской конвенции о праве международных договоров 1969 г.]
Установление аутентичности
Термин «установление аутентичности» означает процедуру, посредством которой текст договора становится аутентичным и окончательным. После установления аутентичности текста договора государства не могут в одностороннем порядке менять его положения. Если государства, участвовавшие в переговорах по данному договору, не согласуют конкретные процедуры установления аутентичности, то установление аутентичности договора, как правило, происходит путем подписания, подписания ad referendum или парафирования представителями этих государств.
[Статья 10 Венской конвенции о праве международных договоров 1969 г.]
понятие и перспективы совершенствования законодательного регулирования, Актуальные проблемы социально-экономического развития России
Автор: Спиридонов Андрей Алексеевич, Председатель Генерального Совета Центра развития Государственно-частного партнерства, к.ю.н.
Вопросы взаимоотношения
государства и бизнеса всегда находились в центре пристального внимания любого
современного демократического правового государства. Более того, в последние
годы во всем мире наблюдается тенденция усиления сотрудничества власти и
бизнеса. В первую очередь, это проявляется в таких сферах, как электроэнергетика,
транспорт, здравоохранение, образование. Предприятия в этих сферах, имеют
стратегическое значение и не могут быть приватизированы, но, с другой стороны,
государство не всегда обладает достаточным объемом средств, необходимых для их
поддержки и развития. В этом случае применяется такая форма взаимодействия
власти и бизнеса, как государственно-частное партнерство. Особенную
актуальность тема государственно-частного партнерства приобрела в условиях
мирового финансового кризиса, в связи с которым многие отрасли российской
экономики почувствовали потребность в государственной поддержке. Председатель
Правительства РФ Владимир Путин, открывая совещание в Новомосковске в сентябре
2009 года, назвал «Государственно-частное
партнерство — основой посткризисного развития регионов».
В этом случае применяется такая форма взаимодействия
власти и бизнеса, как государственно-частное партнерство. Особенную
актуальность тема государственно-частного партнерства приобрела в условиях
мирового финансового кризиса, в связи с которым многие отрасли российской
экономики почувствовали потребность в государственной поддержке. Председатель
Правительства РФ Владимир Путин, открывая совещание в Новомосковске в сентябре
2009 года, назвал «Государственно-частное
партнерство — основой посткризисного развития регионов».
Сам термин
«государственно-частное партнерство» является дословным переводом
английского термина «public-private partnership» (PPP) и давно
применяется в зарубежных государствах: Франции, Великобритании, США, Канаде, Австрии,
Бельгии, Дании, Австралии, Израиле, Ирландии, Финляндии, Испании, Португалии,
Греции, Южной Корее, Сингапуре, Чехии. Во Франции такой опыт сотрудничества
государства и бизнеса впервые был реализован еще в 1552 году при постройке
канала по концессионному принципу. В Великобритании стоимость объектов частной
финансовой инициативы, которая является там основной формой ГЧП, в течение
последних 10 лет составляет 24 млрд. фунтов стерлингов, а в течение следующих
трех лет планируется реализовать проекты на общую сумму 12 млрд. фунтов
стерлингов. Ежегодно заключается до 80 новых соглашений. По данным британского
правительства, такие проекты обеспечивают 17% экономии для бюджета страны[1].
В Великобритании стоимость объектов частной
финансовой инициативы, которая является там основной формой ГЧП, в течение
последних 10 лет составляет 24 млрд. фунтов стерлингов, а в течение следующих
трех лет планируется реализовать проекты на общую сумму 12 млрд. фунтов
стерлингов. Ежегодно заключается до 80 новых соглашений. По данным британского
правительства, такие проекты обеспечивают 17% экономии для бюджета страны[1].
По данным Национального совета по частно-государственной кооперации, в США из 65 базовых видов деятельности муниципальных властей (водопровод, канализация, уборка мусора, школьное образование и др.) средний город использует коммерческие фирмы при исполнении 23[2].
В этой связи, представляется целесообразным
изучение нормативного правового регулирования государственно-частного
партнерства в зарубежных государствах, в которых данный вид партнерства уже
давно служит эффективным средством реализации инновационной и инвестиционной
политики, укрепления экономики, расширения инфраструктуры, реализации
масштабных социальных проектов.
В отличие от большинства зарубежных государств, в России государственно-частное партнерство все еще находится на стадии становления и развития соответствующих инструментов. Хотя результаты такого партнерства уже можно наблюдать.[3] Более того, сам термин «государственно-частное партнерство» все чаще стал встречаться в СМИ и сети Интернет. Все чаще с участием экспертов, аналитиков, ученых, представителей органов государственной власти проходят общественные слушания, научные семинары и круглые столы, посвященные выработке предложений по развитию государственно-частного партнерства в России[4]. В органах государственной власти субъектов РФ создаются структурные подразделения по вопросам ГЧП, при министерствах, Государственной Думе созданы экспертные советы по ГЧП, реализуются проекты с применением институтов ГЧП. В настоящее время среди федеральных нормативных правовых актов, тем или иным образом регулирующих отношения, возникающие в сфере ГЧП можно назвать:
— договорные
конструкции, предусмотренные Гражданским кодексом РФ.
— Федеральный закон от 21 июля 2005 года N 115-ФЗ «О концессионных соглашениях»;
— Федеральный закон от 22 июля 2005 года N 116-ФЗ «Об особых экономических зонах в Российской Федерации»;
— Федеральный закон от 21 июля 2005 года N 94-ФЗ «О размещении заказов на поставки товаров, выполнение работ, оказание услуг для государственных и муниципальных нужд»;
— Федеральный закон от 17.05.2007 N 82-ФЗ «О банке развития»;
— Федеральные законы об иных государственных корпорациях: «Роснанотех», ЖКХ, «Олимпстрой»;
— Постановление Правительства РФ от 24.08.2006 N 516 «Об открытом акционерном обществе «Российская венчурная компания».
— Постановление Правительства РФ об Инвестиционном фонде РФ;
— Постановление
Правительства РФ от 4 мая 2005 года N 284 «О государственном учете
результатов научно-исследовательских, опытно-конструкторских и технологических
работ гражданского назначения».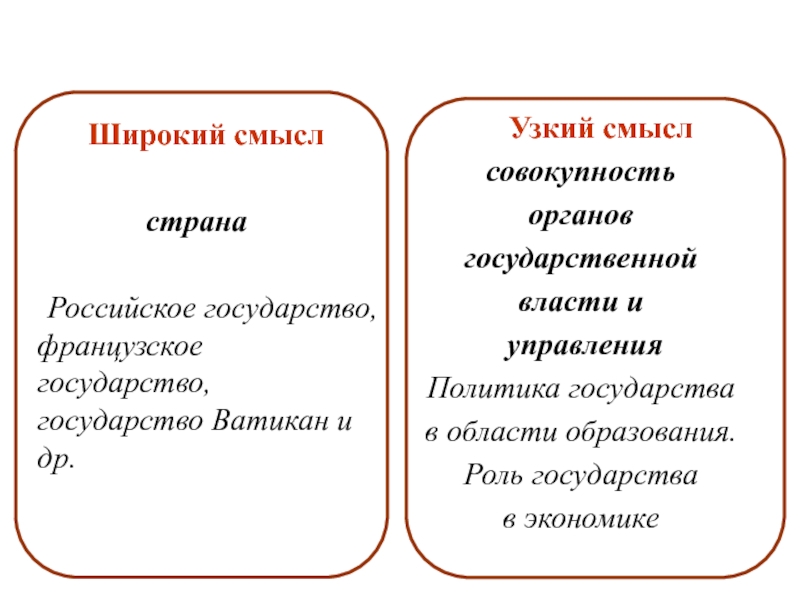
Соответственно действующее российское законодательство предлагает набор инструментов, которые можно применять в проектах, реализуемых на основе ГЧП, это: Инвестиционный фонд РФ, особые экономические зоны, концессии, Российская венчурная компания, ФЦП/ФАИП, региональные фонды развития, государственные корпорации (к примеру, Внешэкономбанк). Широкое применение инструментов ГЧП в России только подтверждает актуальность данного явления для российской экономики в существующих условиях и, следовательно, установления его легального определения.
Однако до сих пор на
федеральном уровне нет общепринятого правового определения понятия ГЧП, хотя оно
и применяется в различных нормативных правовых актах[5].
Региональное законодательство в вопросе о ГЧП продвинулось дальше. В 6
субъектах РФ приняты законы о ГЧП (Санкт-Петербурге, Томской области,
Дагестане, Горном Алтае), в некоторых из них закреплено понятие ГЧП и его
формы. Однако единообразного определения понятия ГЧП также нет. При этом важно
отметить, что и среди специалистов до сих пор нет единого мнения о том, какие
формы взаимодействия власти и бизнеса можно отнести к ГЧП. Так, например, в
литературе под государственно-частным партнерством нередко понимается так
называемая «складчина» государства и бизнеса, некий «институциональный и
организационный альянс» в целях реализации общественно значимых проектов и
программ[6]. Также
пропагандируются под видом ГЧП, такие модели как: социальная ответственность
бизнеса, благотворительность, субсидии частному бизнесу и даже приватизация.
Таким образом, нетрудно отметить, что каждый участник отношений в сфере ГЧП
вкладывает свой смысл в понятие ГЧП, что, прежде всего, связано с отсутствием
общепризнанного определения, что представляет собой ГЧП с точки зрения права.
Однако единообразного определения понятия ГЧП также нет. При этом важно
отметить, что и среди специалистов до сих пор нет единого мнения о том, какие
формы взаимодействия власти и бизнеса можно отнести к ГЧП. Так, например, в
литературе под государственно-частным партнерством нередко понимается так
называемая «складчина» государства и бизнеса, некий «институциональный и
организационный альянс» в целях реализации общественно значимых проектов и
программ[6]. Также
пропагандируются под видом ГЧП, такие модели как: социальная ответственность
бизнеса, благотворительность, субсидии частному бизнесу и даже приватизация.
Таким образом, нетрудно отметить, что каждый участник отношений в сфере ГЧП
вкладывает свой смысл в понятие ГЧП, что, прежде всего, связано с отсутствием
общепризнанного определения, что представляет собой ГЧП с точки зрения права.
Такой разнобой точек зрения на определение
понятия государственно-частного партнерства на наш взгляд, также вызван тем
фактом, что в России только формируется практика и методология ГЧП. Тем не
менее, по нашему убеждению, определение понятия ГЧП является необходимым
условием становления и развития данного института. Важность правильного и
четкого определения понятия ГЧП обусловливается, прежде всего, необходимостью
выработки и законодательного закрепления единой правовой категории, в целях
исключения разногласий, которые могут возникнуть в процессе реализации проектов
на основе ГЧП. Еще Конфуций говорил: «Если понятия не соответствуют
действительности, не соответствуют действительности слова, и если слова не
соответствуют действительности, мы не понимаем друг друга».
Тем не
менее, по нашему убеждению, определение понятия ГЧП является необходимым
условием становления и развития данного института. Важность правильного и
четкого определения понятия ГЧП обусловливается, прежде всего, необходимостью
выработки и законодательного закрепления единой правовой категории, в целях
исключения разногласий, которые могут возникнуть в процессе реализации проектов
на основе ГЧП. Еще Конфуций говорил: «Если понятия не соответствуют
действительности, не соответствуют действительности слова, и если слова не
соответствуют действительности, мы не понимаем друг друга».
В настоящее время, глубоко не вдаваясь в терминологический характер словосочетаний общественно-частное партнерство или частно-государственное партнерство, отметим, что есть несколько подходов к пониманию ГЧП[7].
Условно ГЧП понимают в широком и узком
смыслах. В широком смысле под ГЧП подразумевается любое официальное
конструктивное взаимодействие власти и бизнеса не только в экономике, но и в
политике, культуре, науке.
В узком смысле под государственно-частным партнёрством понимается – взаимодействие бизнеса и власти в процессе реализации социально значимых проектов, имеющих общегосударственное значение.
Рассмотрим основные признаки государственно-частного партнерства.
- сторонами ГЧП являются государство и частный бизнес;
- взаимодействие сторон в ГЧП имеет юридическую основу;
- взаимодействие бизнеса и власти имеет партнерский характер, основанный на учете интересов обоих сторон;
- ГЧП применяется при реализации проектов, имеющих существенную общественно-государственную значимость. ГЧП-проекты — это, прежде всего реализация публичных интересов. Совместные цели и интересы обоих партнеров координируются на основе общегосударственных общественно-полезных целей;
- в процессе реализации проектов на основе ГЧП, объединяются усилия сторон;
- финансовые риски и затраты, а также достигнутые
в ГЧП результаты распределяются между сторонами в пропорциях согласно взаимным
договоренностям, оформленным юридически.

На основе этих признаков можно предложить следующее определение понятия ГЧП, которое представляется возможным рассматривать в качестве основы для законодательного закрепления понятия ГЧП.
ГЧП — это юридически оформленные отношения органов власти и субъектов предпринимательства в отношении объектов, находящихся в юрисдикции государства, основанные на обязательном разделении рисков, учете интересов и координации усилий сторон, осуществляемые в целях наиболее эффективной реализации проектов, имеющих важное общественно-государственное значение.
Не менее важным и дискуссионным вопросом, является вопрос о необходимости принятия комплексного законодательного акта о ГЧП на федеральном уровне. В настоящее время до сих пор среди специалистов нет единства в вопросе о необходимости принятия указанного закона.
По разбросу мнений,
ученых и практиков можно условно разделить на две основные категории:
сторонников принятия закон «Об участии
органов государственной власти и местного самоуправления в ГЧП» (А. В. Баженов, И.Л.
Иванов, И.А. Мартышко и др.) и их оппонентов (О.В. Ревзина, М.Г. Минскова)
некоторые из которых считают, что в принципе вообще можно обойтись без разработки
отраслевого законодательства в сфере ГЧП (Д.Царев, А.Иванюк).
В. Баженов, И.Л.
Иванов, И.А. Мартышко и др.) и их оппонентов (О.В. Ревзина, М.Г. Минскова)
некоторые из которых считают, что в принципе вообще можно обойтись без разработки
отраслевого законодательства в сфере ГЧП (Д.Царев, А.Иванюк).
Основной аргумент оппонентов
принятия закона о ГЧП, заключается в том, что, по мнению указанной группы
авторов, действующее законодательство является достаточным для регулирования
отдельных форм ГЧП и целесообразнее сконцентрировать внимание на внесении
изменений в действующие нормативные правовые акты. «В то же время, — пишут О.В.
Ревзина, М.Г. Минскова, — любые ошибки, допущенные при определении предмета
регулирования федерального закона о ГЧП, могут повлечь за собой ухудшение
правовых условий для реализации ГЧП-проектов в России»[8]. По
мнению Д.Царева, А.Иванюка, «отсутствует необходимость специального
регулирования отношений в сфере ГЧП, в том числе принятия отдельных законов как
по отдельным институтам (формам) ГЧП, так и по ГЧП в целом. К сожалению, именно
по этому направлению пошел российский законодатель. В настоящее время действуют
Федеральные законы N N 94-ФЗ, 115-ФЗ, отдельные федеральные законы по созданию
государственных корпораций в различных областях. Иными словами, наш
законодатель не кодифицирует законодательство, а, наоборот, создает
разрозненные нормативные правовые акты. Такой подход представляется
нецелесообразным и вряд ли внесет ясность в регулирование отношений в сфере
ГЧП. Велика вероятность того, что возникнут коллизия и конкуренция правовых
норм»[9]. По
мнению авторов, положения о концессионных соглашениях могли бы быть включены во
вторую часть ГК РФ, посвященную отдельным видам договоров. «Целесообразно было
бы разработать ряд договоров, отражающих специфику отношений в сфере ГЧП, и
дополнить ими ГК РФ – пишут Д.Царев, А.Иванюк, — В отдельных случаях уже
определенные во второй части ГК РФ договоры могут быть дополнены положениями,
устанавливающими особенности применения таких договоров в рамках ГЧП»[10].
К сожалению, именно
по этому направлению пошел российский законодатель. В настоящее время действуют
Федеральные законы N N 94-ФЗ, 115-ФЗ, отдельные федеральные законы по созданию
государственных корпораций в различных областях. Иными словами, наш
законодатель не кодифицирует законодательство, а, наоборот, создает
разрозненные нормативные правовые акты. Такой подход представляется
нецелесообразным и вряд ли внесет ясность в регулирование отношений в сфере
ГЧП. Велика вероятность того, что возникнут коллизия и конкуренция правовых
норм»[9]. По
мнению авторов, положения о концессионных соглашениях могли бы быть включены во
вторую часть ГК РФ, посвященную отдельным видам договоров. «Целесообразно было
бы разработать ряд договоров, отражающих специфику отношений в сфере ГЧП, и
дополнить ими ГК РФ – пишут Д.Царев, А.Иванюк, — В отдельных случаях уже
определенные во второй части ГК РФ договоры могут быть дополнены положениями,
устанавливающими особенности применения таких договоров в рамках ГЧП»[10].
Рассмотрим аргументы
сторонников закона о ГЧП. «Опираясь на опыт реализации инвестиционных проектов
на принципах государственно-частного партнерства в г. Санкт-Петербурге, — пишет
И.А. Мартышко, — с такой точкой зрения нельзя согласиться. Существующий в
настоящее время в федеральном законодательстве пробел в области ГЧП оставляет
нерешенным множество вопросов, в том числе и такие как статус органов
исполнительной власти, как партнеров в ГЧП, пределы распространения действия
Федерального закона от 21 июля 2005 года N 94-ФЗ «О размещении заказов на
поставки товаров, выполнение работ, оказание услуг для государственных и
муниципальных нужд» на правоотношения участников государственно-частных
партнерств, которые по своей юридической природе, по существу находятся за
пределами действия указанного
федерального закона, а также различные вопросы, связанные с подготовкой
конкурсной документации и проведения конкурсов»[11].
Таким образом, одним из основных аргументов сторонников принятия закона о ГЧП, является несовершенство и недостаточность действующего российского законодательства, в том числе:
«а) недостаточное регулирование в отраслевых законах партнерских отношений органов публичной власти и бизнес-структур, за исключением соглашений о социальном партнерстве, не позволяет четко определять содержание и сферы их применения. Это затрудняет разграничение отдельных видов соглашений как между собой (что не позволяет сформировать их эффективное правовое регулирование), так и с соглашениями, запрещенными антимонопольным законодательством;
б) договоры и соглашения о партнерстве органов публичной власти с бизнес-структурами в публично-правовой сфере (о сотрудничестве в сфере строительства и др., о передаче отдельных государственных (муниципальных) полномочий и т.д.) законодательством не предусмотрены;
в) отсутствие на законодательном уровне четких ориентиров отраслей и сфер, в которых допустимо заключение между органами публичной власти и бизнес-структурами договоров и соглашений публично-правового характера, не позволяет устанавливать взаимоотношения долгосрочного характера;
г) отсутствие в
статусных правовых актах положений о полномочиях органов исполнительной власти
устанавливать с бизнес-структурами партнерские взаимоотношения не позволяет
определить последовательность совершения административных действий при
организации и осуществлении государственно-частных партнерств»[12].
По мнению сторонников закона о ГЧП, предметом регулирования такого закона могут стать следующие вопросы:
- Организация специальной процедуры выбора частного инвестора для заключения контракта ГЧП;
- Обеспечение исполнения обязательств органов власти, принимаемых в рамках контракта ГЧП, в сфере экономического регулирования;
- Приоритет обязательств органов власти, принятых по контракту ГЧП в рамках соответствующей процедуры, для использования в качестве расходных обязательств в бюджете при наступлении случаев, предусмотренных контрактом[13];
По мнению Х.М. Салихова,
«системообразующий нормативный акт мог бы обеспечить максимальную легитимность
действий органов власти в области ГЧП, в частности, по вопросам, не
урегулированным федеральным законодательством и открытым для нормативного
регулирования или практической реализации субъектами Федерации, а также
обеспечил бы судебную защиту соглашениям, заключенным на его основе»[14].
Представляется, что аргументы сторонников закона о ГЧП являются более обоснованными, прежде всего, потому, что развитие ГЧП в России должно опираться, на единую нормативную базу, позволяющую участникам проекта, как властям, так и бизнесу, при реализации совместных проектов взаимодействовать в рамках единого правового поля и «играть» по одним правилам. Кроме того, принятие закона, позволит определить понятийный аппарат ГЧП, а также принципы, механизмы и основные формы ГЧП, сформировать общий подход к реализации проектов на основе ГЧП, а также законодательно регламентировать обязательства сторон и гарантии для частных инвесторов при возникновении различного рода рисков. Принятие закона также могло бы стать положительным сигналом для иностранных инвесторов, которые больше всего опасаются рисков, связанных именно с обязательствами партнера в лице государства.
При этом следует
согласиться с А.А. Зверевым, что помимо наличия четкого законодательства о ГЧП,
необходимо иметь институциональную инфраструктуру для обеспечения
функционирования ГЧП на практике.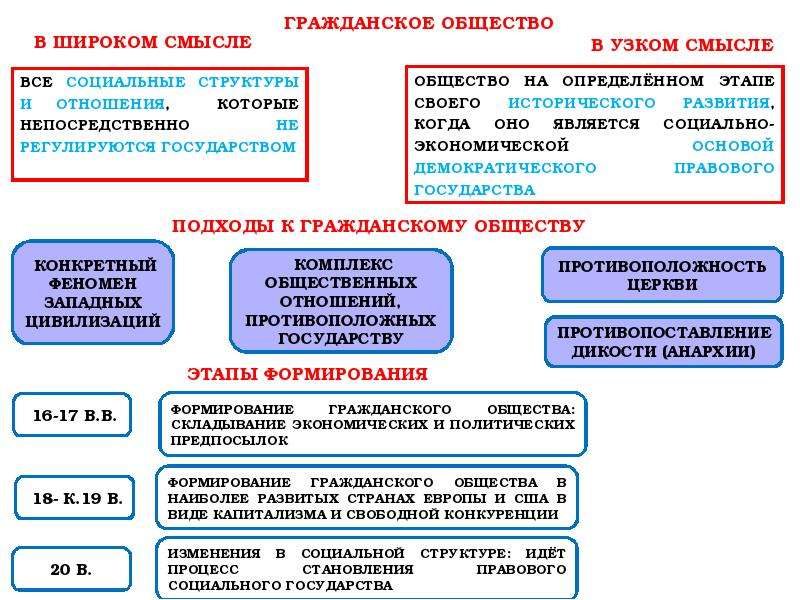 Автор отмечает, что «для использования
различных моделей и обеспечения гибкости, позволяющей изыскивать и оценивать
альтернативные подходы, необходимо создать специализированное подразделение
ГЧП, в целях обобщения опыта с делегированием ему некоторых полномочий –
некоего, «мозгового координирующего центра», который занимался бы подбором
проектов, их оценкой и подготовкой в практической плоскости[15]. Автор
называет это подразделение «национальным Центром ГЧП»[16].
Автор отмечает, что «для использования
различных моделей и обеспечения гибкости, позволяющей изыскивать и оценивать
альтернативные подходы, необходимо создать специализированное подразделение
ГЧП, в целях обобщения опыта с делегированием ему некоторых полномочий –
некоего, «мозгового координирующего центра», который занимался бы подбором
проектов, их оценкой и подготовкой в практической плоскости[15]. Автор
называет это подразделение «национальным Центром ГЧП»[16].
Именно для решения этих задач в рамках Программы Партии «ЕДИНАЯ РОССИЯ» «Диалоги…» и при поддержке Ассоциации молодых предпринимателей России был создан Центр развития ГЧП.
В заключение отметим, что
ГЧП можно рассматривать в качестве одного из основных инструментов достижения
поступательного экономического развития страны. В России государственно-частное
партнерство все еще находится на стадии становления и развития соответствующих
инструментов. Практика с их использованием находится только в начале своего
пути. Вместе с тем, уже сейчас понятно, что качественно реализовать глобальные инфраструктурные
проекты, силами исключительно государства либо бизнеса не представляется
возможным.
Вместе с тем, уже сейчас понятно, что качественно реализовать глобальные инфраструктурные
проекты, силами исключительно государства либо бизнеса не представляется
возможным.
Достичь конкретных результатов высокого качества возможно только при сбалансированной инвестиционной политике в сочетании с сильными сторонами государственного и частного секторов. Поэтому с нашей точки зрения представляется целесообразным, что одним из направлений современной инвестиционной политики должна стать разработка в ближайшем будущем концепции развития в Российской Федерации государственно-частного партнерства, включающей в себя, в том числе единую методологию его организации и осуществления.
[1]
Айрапетян М.С. Зарубежный опыт государственно-частного партнерства. /
Аналитическая записка Правового управления Аппарата Государственной Думы РФ
http://wbase.duma.gov.ru:8080law?d&nd=981605628&mark=r981605004.
[2] См.: Агроскин В. Переходные меры и рецепты реформирования // www.prompolit.ru.
[3] См.: Агроскин В. О кооперации: Три вопроса о частно-государственной кооперации // www.prompolit.ru.
[4] См., «Государственно-частное партнерство: проблемы, тенденции, перспективы» / Материалы Круглого стола Института законодательства и сравнительного правоведения при Правительстве РФ М., 2008 г.
[5] В частности Бюджетный кодекс Российской Федерации и ряд федеральных целевых программ содержат словосочетания, имеющие отношение к государственно-частному партнерству: принципы, проекты, механизмы государственно-частного партнерства. Однако ни в одном из них не раскрывается определение понятия государственно-частного партнерства.
[6] Патоков В.В. Государственно-частное партнерство: Перспективы развития и правовые формы реализации / ИПС «Консультант плюс».
[7] Исходя из содержания понятия
«государственно-частное партнерство» и современной российской
действительности, в России целесообразно применять термин
«государственно-частное партнерство», который четко определяет
ведущую роль государства.
[8]О.В. Ревзина, М.Г. Минскова Государственно-частные партнерства в России – правовое регулирование сегодня перспективы на будущее / Государственно-частное партнерство: пути совершенствования законодательной базы / Сборник статей под общей редакцией Зверева А.А. М. 2009.
[9] Д.Царев, А.Иванюк Государственно-частное партнерство: правовой аспект // «Финансовая газета», 2009, N 17
[10] Д.Царев, А.Иванюк Государственно-частное партнерство: правовой аспект // «Финансовая газета», 2009, N 17
[11]И.А. Мартышко Опыт реализации инвестиционных проектов на принципах государственно-частного партнерства на примере г. Санкт-Петербурга / Государственно-частное партнерство: пути совершенствования законодательной базы / Сборник статей под общей редакцией Зверева А.А. М. 2009.
[12] Хатаева М.А., Цирин А.М. Законодательство о государственно-частном партнерстве: проблемы, тенденции перспективы / Круглого стола на тему «Законодательство о государственно-частном партнерстве в Российской Федерации: проблемы, тенденции, перспективы», проведенного 21 мая 2008 года Институтом законодательства и сравнительного правоведения при Правительстве Российской Федерации
[13] А. В.
Баженов Использование механизмов государственно-частного партнерства как
катализатора антикризисного потенциала развития инфраструктуры /
Государственно-частное партнерство: пути совершенствования законодательной базы
/ Сборник статей под общей редакцией Зверева А.А. М. 2009.
В.
Баженов Использование механизмов государственно-частного партнерства как
катализатора антикризисного потенциала развития инфраструктуры /
Государственно-частное партнерство: пути совершенствования законодательной базы
/ Сборник статей под общей редакцией Зверева А.А. М. 2009.
[14] И.Е.Ильин Государство и бизнес: в русле взаимовыгодного партнерства // «Организация продаж банковских продуктов», 2009, N 1
[15] А.А. Зверев Законодательство о государственно-частном партнерстве: риски и перспективы развития Государственно-частное партнерство: пути совершенствования законодательной базы / Сборник статей под общей редакцией Зверева А.А. М. 2009.
[16] А.А. Зверев, Там же
Влияние архитектуры регулирования на наследуемость в широком и узком смысле
Образец цитирования: Ван Ю, Вик Дж. О., Омхольт С. В., Гьювсланд А. Б. (2013) Влияние архитектуры регулирования на наследуемость в широком и узком смысле. PLoS Comput Biol 9(5):
е1003053.
https://doi.org/10.1371/journal.pcbi.1003053
Б. (2013) Влияние архитектуры регулирования на наследуемость в широком и узком смысле. PLoS Comput Biol 9(5):
е1003053.
https://doi.org/10.1371/journal.pcbi.1003053
Редактор: William Stafford Noble, Вашингтонский университет, Соединенные Штаты Америки
Получено: 10 августа 2012 г.; Принято: 23 марта 2013 г.; Опубликовано: 9 мая 2013 г.
Авторское право: © 2013 Wang et al.Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана Исследовательским советом Норвегии (http://www.rcn.no) в рамках программы eVITA, номер проекта 178901/V30, и проектом Virtual Physiological Rat Project (http:// virtualrat. org) финансируется за счет гранта NIH P50-GM0
org) финансируется за счет гранта NIH P50-GM0
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Наследуемость признака в широком смысле — это доля фенотипической изменчивости, приписываемая генетическим причинам, тогда как наследуемость в узком смысле — это доля, приписываемая аддитивному действию генов.Общая генетическая изменчивость включает дисперсию, объясняемую внутрилокусным доминированием () и межлокусными взаимодействиями (). Причины и важность этой неаддитивной генетической изменчивости, которая отличает две меры наследуемости, были предметом серьезных споров на протяжении более 80 лет (например, [1]–[6]). Недавно с помощью статистических аргументов было показано, что для признаков со многими локусами с экстремальными частотами аллелей большая часть генетической изменчивости становится аддитивной с V G ) обычно >0.5 [3]. В популяциях с промежуточными частотами аллелей, таких как контролируемые скрещивания линий, часто сообщают о более низких соотношениях V A / V G [7], [8]. Это проиллюстрировано в таблице 1, в которой обобщаются расчетные отношения V A / V G из коллекции исследований таких популяций. Этот широкий диапазон соотношений h 2 / H 2 , зарегистрированный для скрещивания линий, не может быть объяснен аргументом частоты аллелей, и предполагаемые объяснения должны быть основаны на том, как регуляторная архитектура лежащих в основе биологических систем формирует карта генотип-фенотип (ГФ).
Недавно с помощью статистических аргументов было показано, что для признаков со многими локусами с экстремальными частотами аллелей большая часть генетической изменчивости становится аддитивной с V G ) обычно >0.5 [3]. В популяциях с промежуточными частотами аллелей, таких как контролируемые скрещивания линий, часто сообщают о более низких соотношениях V A / V G [7], [8]. Это проиллюстрировано в таблице 1, в которой обобщаются расчетные отношения V A / V G из коллекции исследований таких популяций. Этот широкий диапазон соотношений h 2 / H 2 , зарегистрированный для скрещивания линий, не может быть объяснен аргументом частоты аллелей, и предполагаемые объяснения должны быть основаны на том, как регуляторная архитектура лежащих в основе биологических систем формирует карта генотип-фенотип (ГФ).
По нескольким причинам важно понять причинно-следственную подоплеку наблюдаемой изменчивости соотношений h 2 /H 2 внутри биологических систем и между ними.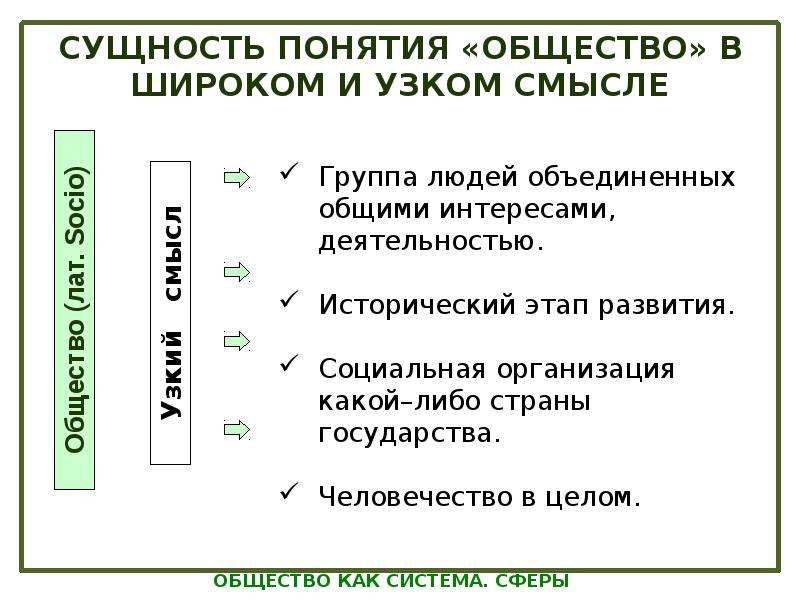 В количественной генетике человека, где обычно используются исследования близнецов, большинство оценок наследуемости относятся к H 2 [9]. В тех случаях, когда h 2 /H 2 является низким, это может привести к нереалистичным ожиданиям относительно того, какая часть основной причинной вариации может быть обнаружена с помощью методов обнаружения линейного QTL [6].С другой стороны, низкая наследуемость данного сложного признака в узком смысле не обязательно означает, что окружающая среда определяет большую часть вариаций. В эволюционной биологии аддитивная изменчивость является основной валютой эволюционной адаптации и эволюционируемости. Важными вопросами в этом контексте являются, например, (i) в какой степени происходит отбор самих регуляторных анатомий для поддержания высокой аддитивной дисперсии, (ii) существуют ли организационные ограничения в построении адаптивных систем, такие, что в некоторых случаях низкий уровень h 2 / H 2 Соотношение обязательно должно возникнуть, пока еще выбрано проксимальное решение? Более того, в продукционной биологии с генетически модифицированными организмами, размножающимися половым путем, хотелось бы гарантировать, что модификации будут передаваться будущим поколениям полностью предсказуемым образом.
В количественной генетике человека, где обычно используются исследования близнецов, большинство оценок наследуемости относятся к H 2 [9]. В тех случаях, когда h 2 /H 2 является низким, это может привести к нереалистичным ожиданиям относительно того, какая часть основной причинной вариации может быть обнаружена с помощью методов обнаружения линейного QTL [6].С другой стороны, низкая наследуемость данного сложного признака в узком смысле не обязательно означает, что окружающая среда определяет большую часть вариаций. В эволюционной биологии аддитивная изменчивость является основной валютой эволюционной адаптации и эволюционируемости. Важными вопросами в этом контексте являются, например, (i) в какой степени происходит отбор самих регуляторных анатомий для поддержания высокой аддитивной дисперсии, (ii) существуют ли организационные ограничения в построении адаптивных систем, такие, что в некоторых случаях низкий уровень h 2 / H 2 Соотношение обязательно должно возникнуть, пока еще выбрано проксимальное решение? Более того, в продукционной биологии с генетически модифицированными организмами, размножающимися половым путем, хотелось бы гарантировать, что модификации будут передаваться будущим поколениям полностью предсказуемым образом. Таким образом, хотелось бы гарантировать, что модификация станет в высокой степени наследуемой в узком смысле.
Таким образом, хотелось бы гарантировать, что модификация станет в высокой степени наследуемой в узком смысле.
В качестве шага к физиологически обоснованному пониманию изменчивости h 2 / H 2 отношений между биологическими системами или процессами мы поставили вопрос: существуют ли регуляторные структуры или определенные классы фенотипов, V A / V G более низкие коэффициенты, чем другие? Решение этого вопроса требует увязки генетической изменчивости с вычислительной биологией в популяционном контексте (т.g., [10]–[19]), так называемое причинно-связное моделирование генотип-фенотип (cGP) [15], [17], [18]. Мы применили этот подход к пяти хорошо проверенным моделям вычислительной биологии, описывающим, соответственно, метаболический путь гликолиза у почкующихся дрожжей [20], сигнальный путь циклического аденозинмонофосфата (цАМФ) у почкующихся дрожжей [21], регуляцию клеточного цикла у почкующихся дрожжей. [22], генная сеть, лежащая в основе циркадных ритмов млекопитающих [23], и ионные каналы, определяющие потенциал действия в сердечных миоцитах мышей [24]. Эти модели различаются по своей регуляторной архитектуре; ниже мы показываем, что они также различаются диапазоном отношений, которые они могут проявлять.В частности, регулирование с положительной обратной связью и колебательное поведение, по-видимому, располагают к низким отношениям V A / V G . Результаты показывают, что наш подход можно использовать в общем виде для изучения того, как соотношение h 2 / H 2 изменяется в зависимости от регуляторной анатомии.
[22], генная сеть, лежащая в основе циркадных ритмов млекопитающих [23], и ионные каналы, определяющие потенциал действия в сердечных миоцитах мышей [24]. Эти модели различаются по своей регуляторной архитектуре; ниже мы показываем, что они также различаются диапазоном отношений, которые они могут проявлять.В частности, регулирование с положительной обратной связью и колебательное поведение, по-видимому, располагают к низким отношениям V A / V G . Результаты показывают, что наш подход можно использовать в общем виде для изучения того, как соотношение h 2 / H 2 изменяется в зависимости от регуляторной анатомии.
Методы
Моделирование моделей cGP
Пять моделей cGP были построены и проанализированы с помощью cgptoolbox (http://github.com/jonovik/cgptoolbox) пакет Python с открытым исходным кодом, разработанный авторами; дополнительный исходный код, относящийся к моделированию в этой статье, доступен по запросу.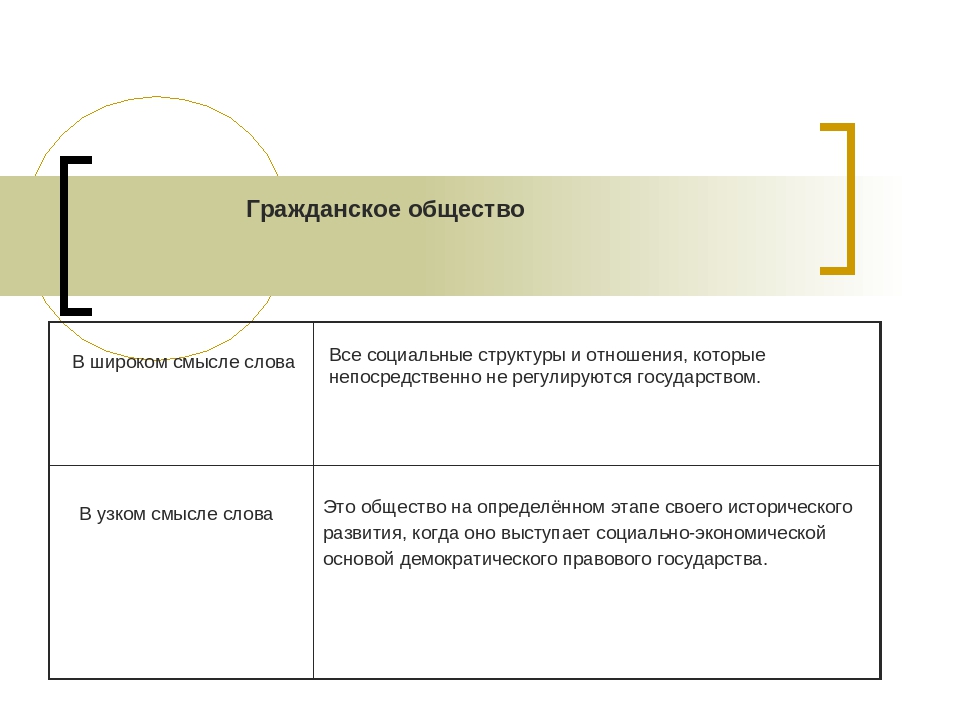 Далее мы описываем три основные части рабочего процесса: (i) сопоставление генотипов с параметрами, (ii) сопоставление параметров с фенотипами, т. е. решение динамических моделей, и (iii) настройка симуляций Монте-Карло, объединяющих два сопоставления (рисунок S1). Для каждой модели мы кратко описываем ее происхождение, программное обеспечение, используемое для ее решения, какие параметры подвергались генетической изменчивости, какие фенотипы были зарегистрированы, а также критерии исключения наборов данных.На рисунках S2, S3, S4, S5, S6 показаны графические представления пяти модельных систем, а текст S1 содержит более подробные описания всех пяти моделей.
Далее мы описываем три основные части рабочего процесса: (i) сопоставление генотипов с параметрами, (ii) сопоставление параметров с фенотипами, т. е. решение динамических моделей, и (iii) настройка симуляций Монте-Карло, объединяющих два сопоставления (рисунок S1). Для каждой модели мы кратко описываем ее происхождение, программное обеспечение, используемое для ее решения, какие параметры подвергались генетической изменчивости, какие фенотипы были зарегистрированы, а также критерии исключения наборов данных.На рисунках S2, S3, S4, S5, S6 показаны графические представления пяти модельных систем, а текст S1 содержит более подробные описания всех пяти моделей.
Преобразование генотипа в параметр.
Для каждой модели следующая процедура была повторена 1000 раз (см. также «Моделирование методом Монте-Карло» ниже) для различных наборов параметров, подлежащих моделируемой генетической изменчивости. Мы начали с выборки трех полиморфных локусов, каждый из которых определял один или два параметра в динамической модели. Таблицы подходящих локусов с соответствующими параметрами и их исходными значениями перечислены в таблицах S1, S2, S3, S4, S5, соответствующих моделям цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно. Наследуемую изменчивость выбранного параметра генерировали для одного биаллельного локуса с аллельными индексами 0 и 1 следующим образом. Сначала равномерно в интервале [0,7, 1,3] были выбраны два числа r 1 и r 2 . Значение параметра для гомозиготы 00 было установлено таким, где b — исходное значение, для гомозиготы 11 значение параметра было равно .Гетерозиготному генотипу 01 было присвоено среднее значение двух гомозигот, что привело к аддитивному сопоставлению генотипов со значениями параметров.
Таблицы подходящих локусов с соответствующими параметрами и их исходными значениями перечислены в таблицах S1, S2, S3, S4, S5, соответствующих моделям цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно. Наследуемую изменчивость выбранного параметра генерировали для одного биаллельного локуса с аллельными индексами 0 и 1 следующим образом. Сначала равномерно в интервале [0,7, 1,3] были выбраны два числа r 1 и r 2 . Значение параметра для гомозиготы 00 было установлено таким, где b — исходное значение, для гомозиготы 11 значение параметра было равно .Гетерозиготному генотипу 01 было присвоено среднее значение двух гомозигот, что привело к аддитивному сопоставлению генотипов со значениями параметров.
цАМФ.
Модель полного сигнального пути цАМФ у S. cerevisiae [21], принимающего в качестве входных данных внешний уровень глюкозы, была загружена в виде кода SBML (http://www. biomedcentral.com/content/supplementary/1752-0509- 3-70-s1.xml) и интегрированы с помощью PySCeS [25]. Генетическая изменчивость была введена в отношении скоростей ассоциации/диссоциации и фосфорилирования/дефосфорилирования сигнальных белков (см. Рисунок S2 и Таблицу S1).Начальные устойчивые концентрации перед добавлением внешней глюкозы, пиковые значения после добавления глюкозы и время, необходимое для достижения пиковых значений клеточных белков, регистрировали как фенотипы (см. Рисунок 1A для иллюстрации фенотипа и Таблицу S6 для описания фенотипа).
biomedcentral.com/content/supplementary/1752-0509- 3-70-s1.xml) и интегрированы с помощью PySCeS [25]. Генетическая изменчивость была введена в отношении скоростей ассоциации/диссоциации и фосфорилирования/дефосфорилирования сигнальных белков (см. Рисунок S2 и Таблицу S1).Начальные устойчивые концентрации перед добавлением внешней глюкозы, пиковые значения после добавления глюкозы и время, необходимое для достижения пиковых значений клеточных белков, регистрировали как фенотипы (см. Рисунок 1A для иллюстрации фенотипа и Таблицу S6 для описания фенотипа).
Рисунок 1. Фенотипы, полученные из моделей cGP.
Графическая иллюстрация фенотипов, зарегистрированных для пяти исследованных моделей cGP. Графики времени (переменная состояния на оси Y, время на оси X) для базового набора параметров отображаются для всех пяти моделей. A. В отсутствие внешней глюкозы все переменные состояния (показана концентрация цАМФ) в модели цАМФ [21] сходятся к стабильному стационарному состоянию (синий кружок на оси ординат). После добавления внешней глюкозы (добавлено 5 мМ в момент времени 50) мы видим быстрое изменение, за которым следует медленный возврат к исходному устойчивому состоянию. В дополнение к исходному стационарному состоянию в качестве фенотипов регистрировали экстремальную концентрацию (верхний синий кружок), а также время достижения экстремума (синяя линия). Б. Концентрации метаболитов (показаны внутренняя глюкоза (GLCi), глюкозо-6-фосфат (G6P) и фруктоза-6-фосфат (F6P)) в модели гликолиза [20] сходятся к стабильному стационарному состоянию, обозначенному незаштрихованными кружками. Стационарные концентрации для 13 метаболитов были зарегистрированы как фенотипы этой модели. C. Для модели клеточного цикла [22] мы записали пиковый уровень и время от основания до пика, как и для циркадной модели (рис. 1D), и, кроме того, мы записали события клеточного цикла, такие как появление почек в то время, когда [BUD] = 1 обозначено черной стрелкой. D. Концентрации мРНК и белка (показаны мРНК для Bmal1 (MB), Cry (MC) и Per (MP)) в циркадной модели [23] сходятся к предельному циклу.
После добавления внешней глюкозы (добавлено 5 мМ в момент времени 50) мы видим быстрое изменение, за которым следует медленный возврат к исходному устойчивому состоянию. В дополнение к исходному стационарному состоянию в качестве фенотипов регистрировали экстремальную концентрацию (верхний синий кружок), а также время достижения экстремума (синяя линия). Б. Концентрации метаболитов (показаны внутренняя глюкоза (GLCi), глюкозо-6-фосфат (G6P) и фруктоза-6-фосфат (F6P)) в модели гликолиза [20] сходятся к стабильному стационарному состоянию, обозначенному незаштрихованными кружками. Стационарные концентрации для 13 метаболитов были зарегистрированы как фенотипы этой модели. C. Для модели клеточного цикла [22] мы записали пиковый уровень и время от основания до пика, как и для циркадной модели (рис. 1D), и, кроме того, мы записали события клеточного цикла, такие как появление почек в то время, когда [BUD] = 1 обозначено черной стрелкой. D. Концентрации мРНК и белка (показаны мРНК для Bmal1 (MB), Cry (MC) и Per (MP)) в циркадной модели [23] сходятся к предельному циклу. В дополнение к периоду колебаний (длинная синяя линия) для каждой из 16 переменных уровень пика (незакрашенный синий кружок), а также время от основания до пика (короткая синяя линия) регистрировались как фенотипы. E. Мы использовали базовый уровень, пиковый уровень, амплитуду, время до пика и время до 25%, 50%, 75% и 90% восстановления потенциала действия и транзиент кальция в качестве фенотипов клеточного уровня модели потенциала действия [24].На рисунке показан потенциал действия.
В дополнение к периоду колебаний (длинная синяя линия) для каждой из 16 переменных уровень пика (незакрашенный синий кружок), а также время от основания до пика (короткая синяя линия) регистрировались как фенотипы. E. Мы использовали базовый уровень, пиковый уровень, амплитуду, время до пика и время до 25%, 50%, 75% и 90% восстановления потенциала действия и транзиент кальция в качестве фенотипов клеточного уровня модели потенциала действия [24].На рисунке показан потенциал действия.
https://doi.org/10.1371/journal.pcbi.1003053.g001
Модель гликолиза.
Модель, опубликованная Teusink et al. [20] описывает гликолиз у S. cerevisiae через кинетику 13 гликолитических ферментов, определяющих потоки переменных состояния метаболитов. Генетическая изменчивость была введена в отношении максимальных скоростей реакции ферментов (см. Рисунок S3 и Таблицу S2). Мы загрузили модель из базы данных BioModels (http://www.ebi.ac.uk/biomodels-main/BIOMD0000000064) в SBML L2 V1 и решили его с помощью PySCeS [25], чтобы найти стабильные устойчивые концентрации метаболитов, которые использовались в качестве фенотипов (см. Рисунок 1B и Таблицу S7). Наборы данных отбрасывались, если один или несколько генотипов не давали стабильного стационарного состояния, что может произойти из-за бифуркации седловидного узла [26].
Рисунок 1B и Таблицу S7). Наборы данных отбрасывались, если один или несколько генотипов не давали стабильного стационарного состояния, что может произойти из-за бифуркации седловидного узла [26].
Модель клеточного цикла.
Модель консенсусных механизмов управления клеточным циклом у S. cerevisae , смоделированная алгебраическими/дифференциальными уравнениями, описывающими непрерывные изменения переменных состояния и разрывы в связи с клеточными событиями [22], получена из репозитория CellML (http //модели.cellml.org/workspace/chen_calzone_csikasznagy_cross_novak_tyson_2004). Генетическая изменчивость была введена в скорость производства и распада различных белков (см. Рисунок S4 и Таблицу S3). Опубликованная модель содержит правила сброса (события) как для параметров, так и для переменных состояния, но реализация CellML включает только правила параметров ( k mad2 , k bub2 и k lte1 ). Правила сброса для переменных состояния [BUD], [SPN] и [ORI], как описано в документе о модели, были реализованы путем решения модели с поиском корня для соответствующих переменных.Модель численно интегрировали с помощью решателя CVODE [27] до сходимости времени клеточного деления, событий клеточного цикла. Пиковые уровни и время до пиковых уровней для концентраций цитозольного белка, а также время событий клеточного деления регистрировали как фенотипы (см. Рисунок 1C для иллюстрации фенотипа и Таблицу S8 для описания фенотипа).
Правила сброса для переменных состояния [BUD], [SPN] и [ORI], как описано в документе о модели, были реализованы путем решения модели с поиском корня для соответствующих переменных.Модель численно интегрировали с помощью решателя CVODE [27] до сходимости времени клеточного деления, событий клеточного цикла. Пиковые уровни и время до пиковых уровней для концентраций цитозольного белка, а также время событий клеточного деления регистрировали как фенотипы (см. Рисунок 1C для иллюстрации фенотипа и Таблицу S8 для описания фенотипа).
Циркадная модель.
Модель циркадных часов млекопитающих, опубликованная Leloup и Goldbeter [23], описывает динамику мРНК и белков трех генов в цитозоле и ядре.Генетическая изменчивость была введена в отношении скорости распада мРНК (см. Рисунок S5 и Таблицу S4). Модель была загружена из репозитория CellML (http://models.cellml.org/workspace/leloup_goldbeter_2004) и интегрирована с помощью CVODE [27] до сходимости к ее предельному циклу. В качестве фенотипов использовали нижние уровни и время от нижнего уровня до пикового значения концентраций мРНК, белков и белковых комплексов. Кроме того, мы записали период колебаний (см. Рисунок 1D для иллюстрации фенотипа и Таблицу S9 для описания фенотипа).
Кроме того, мы записали период колебаний (см. Рисунок 1D для иллюстрации фенотипа и Таблицу S9 для описания фенотипа).
Модель потенциала действия.
Модель [24] является расширением [28] и описывает потенциал действия и кальциевый переходный процесс клетки сердечной мышцы мыши. Мы получили код CellML от авторов, и файл доступен в качестве дополнительного материала в [17]. Численное интегрирование было выполнено с использованием CVODE [27]. Генетическая изменчивость была введена в отношении максимальной проводимости ионных каналов и сродства насоса (см. Рисунок S6 и Таблицу S5). Фенотипы были созданы путем имитации регулярной кардиостимуляции, как это было сделано в [17], [18], при этом ток калия стимула -15 В / с длился 3 мс в начале каждого интервала стимула.Модель моделировалась до сходимости, как описано в [17]; наборы данных, которые не сошлись, отбрасывались. Начальный уровень, пиковый уровень, амплитуда и время до 25, 50, 75 и 90% восстановления потенциала действия и перехода кальция регистрировались как фенотипы клеточного уровня (см. Рисунок 1E для иллюстраций фенотипов и Таблицу S10 для описания фенотипов).
Рисунок 1E для иллюстраций фенотипов и Таблицу S10 для описания фенотипов).
Моделирование методом Монте-Карло.
Для каждой модели мы выполнили 1000 симуляций методом Монте-Карло следующим образом (см. рисунок S1 для иллюстрации).Сначала мы отобрали три полиморфных локуса для введения генетической изменчивости и отобрали карту генотип-параметр, как описано выше. Затем были пронумерованы все 27 возможных трехлокусных генотипов, сопоставлены с 27 наборами параметров, и для каждого набора параметров была решена динамическая модель и получены фенотипы (как описано выше и на рисунке 1). Чтобы избежать артефактов, возникающих из-за численного шума, наборы данных с низкой изменчивостью были исключены из генетического анализа. Абсолютную изменчивость измеряли как разницу между максимальным и минимальным значениями фенотипа в наборе данных, а относительную изменчивость — как отношение абсолютной изменчивости к среднему фенотипу того же набора данных.Пороговые значения для каждого фенотипа и количество наборов данных, превышающих пороги, перечислены в таблицах S6, S7, S8, S9, S10 для моделей цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно.
Результаты/Обсуждение
Системная классификация и размерность фенотипа
Пять моделей cGP, изученных в этой работе, различаются по нескольким параметрам, как по своим функциям, так и по базовой сетевой структуре. Модели гликолиза и цАМФ являются моделями путей с ациклической серией реакций, преобразующих входы в выходы.Модель гликолиза [20] является более продвинутой, чем метаболические модели в более ранних генетически ориентированных исследованиях (например, [3], [31], [32]), поскольку она включает множество различных типов кинетики ферментов, а также регуляцию некоторых ферментов с помощью отрицательной обратной связи. активности ферментов за счет ингибирования продукта. Модель цАМФ [21] содержит ряд петель отрицательной обратной связи, но в целом она также имеет четкую структуру путей, где сигнал глюкозы передается от G-белков к цАМФ и к целевой киназы РКА. Обе эти модели имеют общую относительно простую динамику с решениями, сходящимися к устойчивому устойчивому состоянию (рис. 1А и В).
1А и В).
Напротив, три другие модели демонстрируют циклическое поведение, являющееся результатом взаимодействия между петлями положительной и отрицательной обратной связи (рис. 1 C–E). Однако между этими моделями также есть явные различия. Модель клеток сердца [24] представляет собой возбудимую систему с механизмами обратной связи, включая индуцированное кальцием высвобождение кальция и несколько зависимых от напряжения ионных каналов. В отличие от клеток-водителей ритма, он полагается на внешнюю стимуляцию, чтобы инициировать потенциал действия. Модель циркадного ритма [23], [33] представляет собой сеть экспрессии генов с переплетенными положительными и отрицательными петлями обратной связи транскрипции, дающими осциллятор предельного цикла с устойчивыми колебаниями даже в сплошной темноте.Модель клеточного цикла [22] основывается на петле положительной обратной связи между циклинами В-типа в ассоциации с циклинзависимой киназой и ингибиторами циклинзависимой киназы, которая устанавливает петлю гистерезиса, заставляющую клеточный цикл возникать из переходов между двумя альтернативными стабильными киназами. устойчивые состояния.
устойчивые состояния.
Эта грубая классификация пяти моделей cGP на модели путей и более сложные регуляторные системы ясно отражена в эффективной размерности фенотипов, возникающих в наших симуляциях Монте-Карло.Мы изучили фенотипическую размерность для всех пяти моделей cGP с помощью анализа основных компонентов (PCA) для каждой симуляции Монте-Карло (рис. 2). Во всех смоделированных наборах данных 95% фенотипической изменчивости моделей гликолиза и цАМФ можно объяснить первыми 3 основными компонентами, для моделей клеточного цикла и клеток сердца требуются первые 5 основных компонентов, а для циркадной модели требуется 7 компонентов. Поскольку карты генотип-параметр являются аддитивными для всех пяти моделей, эти различия в эффективной размерности фенотипов высокого уровня указывают на то, что сопоставления параметров с фенотипами проще для моделей путей, чем для трех других моделей.Это вместе с результатами о влиянии положительной обратной связи на статистический эпистаз в сетях регуляции генов [11] позволяет предположить, что модели гликолиза и цАМФ могут приводить к более высоким соотношениям V A / V G , чем другие модели. три модели.
три модели.
Рисунок 2. Размерность фенотипической изменчивости.
Доля общей объясненной фенотипической изменчивости (ось y) по сравнению с количеством основных компонентов (ось x) во всех пяти моделях cGP (обозначены цветом).Для каждого набора данных Монте-Карло матрицы, содержащие полную карту генотип-фенотип для всех M зарегистрированных фенотипов, были стандартизированы до единичной дисперсии перед анализом основных компонентов. Каждая ящичковая диаграмма суммирует объясненную дисперсию для почти 1000 симуляций Монте-Карло.
https://doi.org/10.1371/journal.pcbi.1003053.g002
Отношение аддитивной генетической изменчивости к общей генетической изменчивости
Результаты подтвердили наши ожидания относительно высоких соотношений V A / V G для моделей гликолиза и цАМФ.Кроме того, появилось несколько различных моделей. Модель цАМФ показывает общие самые высокие значения соотношений V A / V G (рис. 3A и таблица S6) со средним и медианным значениями выше 0,99 для всех зарегистрированных фенотипов. Квантиль 0,05 (т. е. только 5 процентов моделирования Монте-Карло показывают более низкие значения, чем это) V A / V G значения были выше 0,97 для всех фенотипов, и не наблюдались значения ниже 0,6.Другими словами, внутри- и межлокусная аддитивная модель действия генов очень хорошо аппроксимирует карты генотип-фенотип, возникающие из этой модели cGP.
3A и таблица S6) со средним и медианным значениями выше 0,99 для всех зарегистрированных фенотипов. Квантиль 0,05 (т. е. только 5 процентов моделирования Монте-Карло показывают более низкие значения, чем это) V A / V G значения были выше 0,97 для всех фенотипов, и не наблюдались значения ниже 0,6.Другими словами, внутри- и межлокусная аддитивная модель действия генов очень хорошо аппроксимирует карты генотип-фенотип, возникающие из этой модели cGP.
Рис. 3. Эмпирическая кумулятивная функция распределения соотношений V A / V G для фенотипов моделей цАМФ (A) и гликолиза (B).
A. Эмпирические кумулятивные функции распределения (ось Y) соотношений V A / V G (ось X) для всех фенотипов, изученных в модели цАМФ: начальные устойчивые концентрации до добавления внешняя глюкоза циклического аденозинмонофосфата (цАМФ), G-белок Ras2a (Ras2a), фактор гуанин-нуклеотидного обмена (Cdc25), протеинкиназа А (PKAi). Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v). Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). См. Таблицу S6 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирическая кумулятивная функция распределения (ось y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза: ацетальдегид (ACE), 1,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).См. Таблицу S7 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v). Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). См. Таблицу S6 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирическая кумулятивная функция распределения (ось y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза: ацетальдегид (ACE), 1,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).См. Таблицу S7 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
https://doi.org/10.1371/journal.pcbi.1003053.g003
Модель гликолиза также имеет средние и медианные значения V A / V G , близкие к 1 для всех фенотипов (рис. 3B). и Таблица S7). Но по сравнению с моделью цАМФ цифры явно ниже; самое низкое зарегистрированное среднее значение (фенотип BPG ) равно 0.9 и 0,05-квантильные значения ниже 0,7 для некоторых фенотипов. Несколько значений ниже 0,5 наблюдаются для всех фенотипов. Распределение соотношений V A / V G для модели клеточного цикла (рисунок S7 и таблица S8) очень похоже на распределение модели гликолиза с самым низким средним значением V A / V G значение 0,93 для время до пика для Sic1 и с 0.05-квантили ниже 0,8 для некоторых фенотипов. V A / V G значения ниже 0,1 наблюдаются для нескольких симуляций методом Монте-Карло в некоторых фенотипах.
Для каждой из моделей цАМФ, гликолиза и клеточного цикла распределения соотношений V A / V G были довольно сходными для всех фенотипов, и подавляющее большинство моделирования методом Монте-Карло показали очень высокие отношения. Модель циркадных часов отличается от этих трех моделей как с точки зрения отображения больших различий между фенотипами, так и с точки зрения наличия гораздо большей доли низких значений (рис. 4А и таблица S9). .Выделяются четыре фенотипа с распределениями V A / V G , которые напоминают равномерное распределение U (0,1). Это время от основания до пика для фосфорилированных и нефосфорилированных белков Per и Cry , и они имеют медианные значения V A / V G в диапазоне от 0,46 до 0,70 и 0,0,036. значения в диапазоне от 0,04 до 0,10. Остальные фенотипы имеют несколько более высокие значения V A / V G , но более половины зарегистрированных фенотипов имеют 0. 05-квантили ниже 0,6. Медианные значения V A / V G ниже 0,9 для большинства фенотипов модели потенциала действия. И все зарегистрированные фенотипы имеют большую долю низких соотношений V A / V G (рис. 4B и таблица S10) с квантилями 0,05 в диапазоне 0,18-0-35. Распределения для потенциалов действия и переходных кальциевых фенотипов весьма сходны, но время до 90% реполяризации для потенциала действия показывает несколько более высокие значения, чем для других.
05-квантили ниже 0,6. Медианные значения V A / V G ниже 0,9 для большинства фенотипов модели потенциала действия. И все зарегистрированные фенотипы имеют большую долю низких соотношений V A / V G (рис. 4B и таблица S10) с квантилями 0,05 в диапазоне 0,18-0-35. Распределения для потенциалов действия и переходных кальциевых фенотипов весьма сходны, но время до 90% реполяризации для потенциала действия показывает несколько более высокие значения, чем для других.
Рисунок 4. Эмпирическая кумулятивная функция распределения соотношений V A / V G для фенотипов циркадной модели (A) и модели потенциала действия (B).
Эмпирические кумулятивные функции распределения (ось Y) отношений V A / V G (ось x) для фенотипов, изученных в циркадной модели и модели клеток сердца. A. На верхней левой панели (Bmal1) показаны фенотипы, связанные с геном bmal1 , включая мРНК (MB), нефосфорилированный/фосфорилированный белок в цитозоле (BC/BCP) и ядре (BN/BNP). Нижняя правая панель (Per) предназначена для на генов, включая мРНК (MP), нефосфорилированный белок (PC) и фосфорилированный белок (PCP). Нижняя концентрация (сплошная линия) и время, необходимое для достижения пика (пунктирная линия) каждого вида, являются зарегистрированными фенотипами. Нижняя левая панель (Cry) связана с геном cry , включая мРНК (MC), нефосфорилированный белок (CC) и фосфорилированный белок (CCP). Верхняя правая панель (комплекс) предназначена для белковых комплексов PCC, PCN, PCCP и PCNP.Также показан период циркадного ритма (Период, пунктирная линия). См. Таблицу S9 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирические кумулятивные функции распределения (ось y) V A / V G отношения (ось x) для фенотипов, изучаемых в модели потенциала действия: время до 25%, 50%, 75 Показаны % и 90% начальных значений, амплитуда, начальные значения (база), пиковые значения, время до достижения пиковых значений потенциала действия (левая панель) и транзиент кальция (правая панель).
Нижняя правая панель (Per) предназначена для на генов, включая мРНК (MP), нефосфорилированный белок (PC) и фосфорилированный белок (PCP). Нижняя концентрация (сплошная линия) и время, необходимое для достижения пика (пунктирная линия) каждого вида, являются зарегистрированными фенотипами. Нижняя левая панель (Cry) связана с геном cry , включая мРНК (MC), нефосфорилированный белок (CC) и фосфорилированный белок (CCP). Верхняя правая панель (комплекс) предназначена для белковых комплексов PCC, PCN, PCCP и PCNP.Также показан период циркадного ритма (Период, пунктирная линия). См. Таблицу S9 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирические кумулятивные функции распределения (ось y) V A / V G отношения (ось x) для фенотипов, изучаемых в модели потенциала действия: время до 25%, 50%, 75 Показаны % и 90% начальных значений, амплитуда, начальные значения (база), пиковые значения, время до достижения пиковых значений потенциала действия (левая панель) и транзиент кальция (правая панель). См. Таблицу S10 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
См. Таблицу S10 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
https://doi.org/10.1371/journal.pcbi.1003053.g004
Все пять моделей cGP способны создавать отношения V A / V G , близкие к 1, и кроме двух фенотипов для циркадной модели все медианные значения V A / V G значительно выше 0.5. Это подтверждает гипотезу [30] о том, что биологические системы имеют тенденцию включать регуляторные механизмы, что в целом приводит к значительной аддитивной генетической изменчивости даже при промежуточных частотах аллелей. Это не означает, что отбор иногда не может привести к регуляторным решениям, стремящимся к низким соотношениям V A / V G ; на самом деле частота низких соотношений V A / V G заметно различалась среди пяти изученных нами моделей. Поскольку карты параметров генотипа были чисто аддитивными, вся неаддитивная генетическая дисперсия является результатом нелинейности моделей ОДУ. Ожидаемым эффектом введения неаддитивности в карты генотип-параметр будет дальнейшее снижение отношений V A / V G .
Поскольку карты параметров генотипа были чисто аддитивными, вся неаддитивная генетическая дисперсия является результатом нелинейности моделей ОДУ. Ожидаемым эффектом введения неаддитивности в карты генотип-параметр будет дальнейшее снижение отношений V A / V G .
Наш вывод о том, что модели со сложной регуляцией, включающей петли положительной обратной связи, имеют тенденцию давать более низкие значения V A / V G , согласуется с предыдущим исследованием сетей регуляции генов [11].Принимая во внимание относительно высокие отношения V A / V G модели клеточного цикла по сравнению с моделями циркадианного цикла и потенциала действия, следующая цитата из обсуждения Тайсона и Новака [34] о том, почему клеточный цикл более информативен как петля гистерезиса, чем как осциллятор предельного цикла (LCO): « Вообще говоря, период LCO является сложной функцией всех кинетических параметров в дифференциальных уравнениях. Однако период цикла клеточного деления зависит только от одного кинетического свойства клетки: времени удвоения ее массы. ”Это, кажется, объясняет, почему карты генотип-фенотип, полученные из моделей клеточного цикла, гораздо более линейны, чем карты из циркадной модели, которая является LCO.
Однако период цикла клеточного деления зависит только от одного кинетического свойства клетки: времени удвоения ее массы. ”Это, кажется, объясняет, почему карты генотип-фенотип, полученные из моделей клеточного цикла, гораздо более линейны, чем карты из циркадной модели, которая является LCO.
Монотонность объясняет большую часть моделей V
A /V G В данной популяции V A / V G является функцией частот аллелей, а также карты GP, и карты GP с сильными взаимодействиями могут по-прежнему давать высокие значения V A / V G значений в популяциях с экстремальными частотами аллелей [3].В популяциях с промежуточными частотами аллелей значения V A / V G определяются в основном формой карты генотип-фенотип и наблюдаемыми различиями между пятью моделями cGP в распределении V Значения / V G побуждают к поиску лежащих в их основе объяснительных принципов.
Недавно предложенная концепция монотонности (или сохранения порядка) отображений GP кажется одним из таких принципов.Короче говоря, карта ГП называется монотонной, если порядок генотипов по содержанию генов (количеству аллелей данного типа) сохраняется в порядке соответствующих фенотипических значений (подробности см. в [30]). На рис. 5 показаны три экстремальных типа карт GP, наблюдаемых в наших симуляциях. Почти аддитивные карты GP, как показано на рисунке 5A, дают значения V A / V G , очень близкие к единице. Карты GP с сильным эпистазом величины, но все еще сохраняющие порядок, обычно дают промежуточные значения V A / V G (рис. 5B), в то время как карты GP сильно немонотонны или нарушают порядок (рис. 5C). ), демонстрирующие сильное сверхдоминирование и/или признаки эпистаза, приводят к значениям V A / V G , близким к нулю.
Рисунок 5. Три различных типа карт генотип-фенотип.
Три различных типа карт генотип-фенотип.
Примеры трех различных типов карт генотип-фенотип, наблюдаемых в наших симуляциях. Для каждой субфигуры значение фенотипа показано на оси ординат, тогда как оси абсцисс, цвета линий и столбцы графика указывают генотип в трех локусах. A. Почти аддитивная карта, иллюстрируемая картой GP времени до пиковой концентрации Cdc25 ( V A / V G = 0.99) в модели цАМФ; B. Полностью монотонная, но неаддитивная карта, представленная на примере карты GP концентрации белка P2G ( V A / V G = 0,41) в модели гликолиза; и, C. Найдена сильно немонотонная карта времени до пиковой концентрации белка PC ( V A / V G = 0,03) из циркадной модели.
https://doi.org/10.1371/журнал.pcbi.1003053.g005
Основываясь на недавних результатах исследований сетей регуляции генов [30], мы ожидали, что три модели cGP со сложной регуляцией, включающей положительную обратную связь, приведут к значительно более немонотонным или нарушающим порядок картам GP, чем две модели пути. Чтобы проверить это, мы измерили количество нарушений порядка во всех смоделированных картах GP (см. Методы) и обнаружили очень четкую закономерность (рис. 6). В то время как наборы данных моделей гликолиза и цАМФ содержали только 1.Карты 1% и 1,3% GP с нарушением порядка для любого локуса, карты клеточного цикла, циркадных моделей и моделей потенциала действия содержали 20,7%, 44,4% и 69,5% соответственно. Более того, монотонные карты GP давали более высокие значения V A / V G , чем немонотонные карты GP для всех пяти моделей cGP (критерий Манна-Уитни; значения p ниже 1e-10 для всех пяти моделей) .
Чтобы проверить это, мы измерили количество нарушений порядка во всех смоделированных картах GP (см. Методы) и обнаружили очень четкую закономерность (рис. 6). В то время как наборы данных моделей гликолиза и цАМФ содержали только 1.Карты 1% и 1,3% GP с нарушением порядка для любого локуса, карты клеточного цикла, циркадных моделей и моделей потенциала действия содержали 20,7%, 44,4% и 69,5% соответственно. Более того, монотонные карты GP давали более высокие значения V A / V G , чем немонотонные карты GP для всех пяти моделей cGP (критерий Манна-Уитни; значения p ниже 1e-10 для всех пяти моделей) .
Рисунок 6. Количество локусов, для которых GP-карта показывает нарушение порядка.
Количество симуляций Монте-Карло, в которых карта GP для данного фенотипа явно нарушает порядок (карты GP с N/A >0.05, см. Методы) показан для модели цАМФ ( A ), модели гликолиза ( B ), модели клеточного цикла ( C ), циркадной модели ( D ) и модели потенциала действия ( Е ).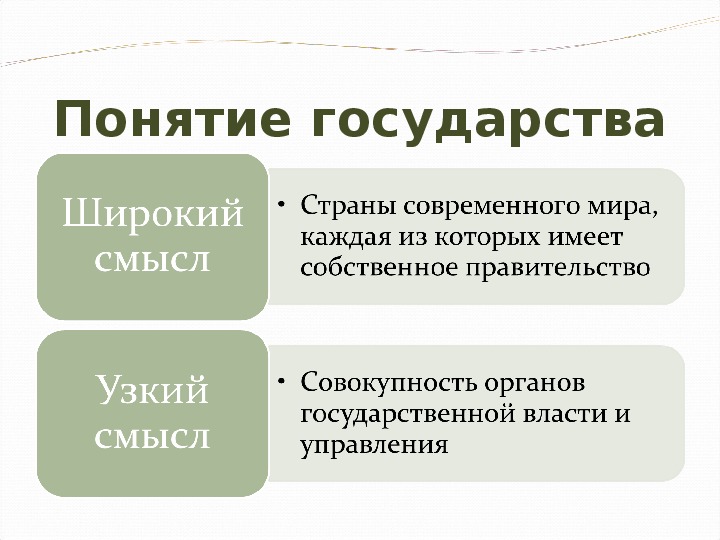 Показаны только фенотипы, в которых хотя бы одно моделирование Монте-Карло привело к нарушению порядка карты GP.
Показаны только фенотипы, в которых хотя бы одно моделирование Монте-Карло привело к нарушению порядка карты GP.
https://doi.org/10.1371/journal.pcbi.1003053.g006
Однако, несмотря на то, что модель гликолиза редко показывает нарушение порядка даже для одного локуса, она обладает значительно меньшими V G значений, чем модель цАМФ.Правдоподобным объяснением является то, что стационарные концентрации метаболитов могут заметно увеличиваться при значениях параметров, близких к точке бифуркации седловидного узла [26]. Результаты моделирования с неустойчивыми устойчивыми состояниями отбрасывались, но в тех случаях, когда один из генотипов (т.е. наборов параметров) приближается к точке бифуркации, не пересекая ее, мы получаем карты генотип-фенотип, как на рис. 5Б, где один генотип (или небольшой set) дает гораздо более высокие фенотипические значения, чем остальные. Такие карты GP, подобные модели дублирующих факторов в Hill et al. [3], как известно, дают низкие отношения V A / V G , несмотря на монотонность. Подобные карты GP, дающие отношения V A / V G , близкие к нулю, были также обнаружены Keightley [32] в его исследовании метаболических моделей, обладающих нулевыми аллелями во всех локусах.
Подобные карты GP, дающие отношения V A / V G , близкие к нулю, были также обнаружены Keightley [32] в его исследовании метаболических моделей, обладающих нулевыми аллелями во всех локусах.
Соображения относительно универсальности результатов
Наша главная причина для ограничения выборочной генетической изменчивости параметров в пределах 30% от опубликованных исходных значений заключалась в том, чтобы избежать качественных (или топологических) изменений в динамике.Такие качественные изменения часто являются биологически реалистичными описаниями нокаутов или других крупных генетических изменений, например потенциалы действия переменной амплитуды (альтернаны) [17]; потеря стабильных циркадных колебаний [23]; и фенотипы нежизнеспособных мутантов клеточного цикла [22]. Однако, поскольку концепции компонентов наследственности и дисперсии определены для фенотипов, демонстрирующих непрерывную, а не дискретную изменчивость, мы стремились избежать здесь таких качественных изменений.
Мы провели моделирование с пятью полиморфными локусами для моделей цАМФ (рис. S8A), гликолиза (рис. S8B), клеточного цикла (рис. S9) и потенциала действия (рис. S10) (циркадная модель явно описывает только три гена).Полученные значения V A / V G были несколько ниже, чем с тремя локусами, но общий вид распределений и четкие различия между моделями не изменились. Это указывает на то, что наши результаты имеют общее значение для олигогенных признаков.
Следует подчеркнуть, что пять исследованных моделей cGP отличаются по ряду других аспектов, чем те, которые выделены здесь, такие как размер системы (количество переменных состояния) и масштабы времени процесса.Эти особенности также могут способствовать наблюдаемой вариации в распределениях отношений V A / V G . Однако такие структурные различия неизбежны, когда целью является сравнение экспериментально подтвержденных моделей, предназначенных для описания конкретных биологических систем. Дополнительный подход заключается в изучении общих моделей, в которых размер системы и структура уравнения фиксированы, а матрица связности может быть изменена для описания семейства систем [35].Это облегчает теоретико-графовое сравнение систем за счет некоторого биологического реализма. Мы ожидаем, что основные выводы из таких исследований будут аналогичны нашим, но вполне может оказаться, что на передний план могут выйти и другие важные общие идеи.
Дополнительный подход заключается в изучении общих моделей, в которых размер системы и структура уравнения фиксированы, а матрица связности может быть изменена для описания семейства систем [35].Это облегчает теоретико-графовое сравнение систем за счет некоторого биологического реализма. Мы ожидаем, что основные выводы из таких исследований будут аналогичны нашим, но вполне может оказаться, что на передний план могут выйти и другие важные общие идеи.
Все модели в нашем исследовании описывают части клеточного механизма, и результирующие фенотипы, таким образом, являются клеточными, а не организменными. Мы не думаем, что это серьезный недостаток с точки зрения основных выводов, которые вытекают из наших результатов.Тем не менее, мы ожидаем, что применение нашего подхода к многомасштабным моделям, включая фенотипы клеток, тканей и целых органов [36], обеспечит значительно улучшенную основу для объяснения того, как свойства карты GP варьируются между биологическими системами и внутри них с точки зрения регуляторной анатомии и ассоциированная генетическая изменчивость [37], [38].
Поскольку наш подход можно использовать вместе с любой моделью вычислительной биологии, он может внести существенный вклад в теоретическую основу, способную предсказать, когда следует ожидать низкого или высокого уровня h 2 / H 2 отношения из принципов регуляторной биологии.Таким образом, причинно-связное моделирование генотип-фенотип считается многообещающим подходом для интеграции причинно-следственных моделей биологических сетей и физиологии с количественной генетикой [39]–[44].
Дополнительная информация
Рисунок S7.
Эмпирическая кумулятивная функция распределения V A / V G 9000s моделей циклов фенотипов. Эмпирические кумулятивные функции распределения (ось Y) отношений V A / V G (ось X) для всех фенотипов, изученных в модели клеточного цикла. Фенотипы делятся на 3 группы. Клеточные события относятся к дискретным событиям, определенным в модельном документе, и включают время почкования (Bud), время репликации ДНК (Rep) и время выравнивания хромосом на метафазных пластинах (Spn). Пиковая концентрация включает концентрацию фосфорилированного комплекса, стимулирующего анафазу (APCP), G1-стабилизирующего белка Cdc6, белка 2 циклина B-типа (Clb2), циклина B-типа, стимулирующего S-фазу (Clb5), стартовой киназы. (Cln2) и белок, стабилизирующий фазу G1 (Sci1).Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1. См. Таблицу S8 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
Фенотипы делятся на 3 группы. Клеточные события относятся к дискретным событиям, определенным в модельном документе, и включают время почкования (Bud), время репликации ДНК (Rep) и время выравнивания хромосом на метафазных пластинах (Spn). Пиковая концентрация включает концентрацию фосфорилированного комплекса, стимулирующего анафазу (APCP), G1-стабилизирующего белка Cdc6, белка 2 циклина B-типа (Clb2), циклина B-типа, стимулирующего S-фазу (Clb5), стартовой киназы. (Cln2) и белок, стабилизирующий фазу G1 (Sci1).Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1. См. Таблицу S8 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
https://doi.org/10.1371/journal.pcbi.1003053.s007
(PDF)
Рисунок S8.
Эмпирическая накопительная функция распределения V A / / / г г г Соотношения для фенотипов лагеря (а) и модели гликолиса (B) с 5 полиморфными локусами . На рис. 3 показаны результаты моделирования с 3 полиморфными локусами. A. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели цАМФ: начальные равновесные концентрации до добавления внешней глюкозы циклического аденозинмонофосфата (цАМФ), G-белка Ras2a (Ras2a), фактора гуанин-нуклеотидного обмена (Cdc25), протеинкиназы А (PKAi). Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v).Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). B. Эмпирическая кумулятивная функция распределения (ось Y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза ацетальдегида (ACE), 1 ,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).
На рис. 3 показаны результаты моделирования с 3 полиморфными локусами. A. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели цАМФ: начальные равновесные концентрации до добавления внешней глюкозы циклического аденозинмонофосфата (цАМФ), G-белка Ras2a (Ras2a), фактора гуанин-нуклеотидного обмена (Cdc25), протеинкиназы А (PKAi). Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v).Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). B. Эмпирическая кумулятивная функция распределения (ось Y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза ацетальдегида (ACE), 1 ,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).
https://doi.org/10.1371/journal.pcbi.1003053.s008
(PDF)
Рисунок S9.
Эмпирическая кумулятивная функция распределения V A / V G соотношения полиморфных клеточных циклов. На рисунке S7 показаны результаты моделирования с 3 полиморфными локусами. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели клеточного цикла.Фенотипы делятся на 3 группы. Клеточные события относятся к дискретным событиям, определенным в модельном документе, и включают время почкования (Bud), время репликации ДНК (Rep) и время выравнивания хромосом на метафазных пластинах (Spn). Пиковая концентрация включает концентрацию фосфорилированного комплекса, стимулирующего анафазу (APCP), G1-стабилизирующего белка Cdc6, белка 2 циклина B-типа (Clb2), циклина B-типа, стимулирующего S-фазу (Clb5), стартовой киназы. (Cln2) и белок, стабилизирующий фазу G1 (Sci1).Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1.
(Cln2) и белок, стабилизирующий фазу G1 (Sci1).Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1.
https://doi.org/10.1371/journal.pcbi.1003053.s009
(PDF)
Рисунок S10.
Эмпирическая кумулятивная функция распределения V A / V G отношений полиморфных фенотипов действия для моделей лоциморфов. На рис. 4В показаны результаты моделирования с 3 полиморфными локусами.Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для фенотипов, изученных в модели потенциала действия: время до 25%, 50%, 75% и 90% показаны начальные значения, амплитуда, начальные значения (база), пиковые значения, время до достижения пиковых значений потенциала действия (левая панель) и транзиент кальция (правая панель).
https://doi.org/10.1371/journal.pcbi.1003053.s010
(PDF)
Таблица S5.
Полиморфные элементы модели потенциала действия. Список элементов и параметров модели потенциала действия, используемых для проявления генетической изменчивости. Имена параметров из таблицы B1 в оригинальной публикации [24], имена, используемые в файле CellML, который доступен в качестве дополнительного материала (имя файла «LNCS model.zip») по адресу doi:10.3389/fphys.2011.00106, и базовые значения в единицах измерения.
https://doi.org/10.1371/journal.pcbi.1003053.s015
(PDF)
Таблица S6.
Краткое описание фенотипов, пороги изменчивости и распределение V A / V G 4
В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
https://doi.org/10.1371/journal.pcbi.1003053.s016
(PDF)
Таблица S7.
Краткое описание фенотипов, пороги изменчивости и распределение V A / V G коэффициенты гликолиза. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
https://doi.org/10.1371/journal.pcbi.1003053.s017
(PDF)
Таблица S8.
Краткое описание фенотипов, пороги изменчивости и распределение V A / V G циклов для модели соотношения клеток.
В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог.Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
https://doi.org/10.1371/journal.pcbi.1003053.s018
(PDF)
Таблица S9.
Краткое описание фенотипов, пороги изменчивости и распределение V A / V G В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.Сокращения: фосфорилированные – phos., цитозольные – cyt., ядерные – nuc., нижняя концентрация – b.c., пиковая концентрация – p.c.
Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.Сокращения: фосфорилированные – phos., цитозольные – cyt., ядерные – nuc., нижняя концентрация – b.c., пиковая концентрация – p.c.
https://doi.org/10.1371/journal.pcbi.1003053.s019
(PDF)
Таблица S10.
Краткое описание фенотипов, пороги изменчивости и распределение V A / V G 4 3s для модели потенциалов действия. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц.Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
https://doi.org/10.1371/journal.pcbi.1003053.s020
(PDF)
Qualia в широком и узком смысле.(A) Определение в широком смысле…
Контекст 1
… В отличие от этого, наше исследование показало, что угловая извилина может вычислять бездействие во время выбора действия, а не только ретроспективно, когда результаты были известны. Это исследование не обнаружило связи между активацией угловой извилины и чувством контроля в пробах с совместимостью, предполагая, что эта область отслеживает сигнал, связанный с трудностью и конфликтом во время выбора действия, а не нормальный плавный поток действия (рис. 1C).Более того, латеральная префронтальная кора, отвечающая за выбор между различными возможными действиями, показала отрицательную корреляцию с активацией угловой извилины только при несовместимо-примированных и отсутствие корреляции при совместимых примированных испытаниях. …
…
Контекст 2
… tem ‘qualia’ используется в двух разных смыслах: широком и узком. Квале в широком смысле относится ко всему сознательному опыту в один момент, который состоит из множества модальностей чувств, зрения, слуха, осязания и т. д., а также множества аспектов восприятия в каждой модальности, например, цвета, формы и движение в поле зрения (рис. 1А).В этом широком смысле весь феноменальный опыт в данный момент времени рассматривается как одно качество. …
Контекст 3
… узкие квалиа кажутся более полезными для исследования сознания. Узкое определение, например, позволяет нам сравнивать квалиа для двух стимулов, представленных в разных пространственных положениях, таких как красный квалиа для стимула А слева и красный квалиа для стимула В справа (рис. 1В). Однако в широком смысле считается, что квалиа для двух объектов в разных местах вызывают разные квалиа, потому что опыт в целом различен из-за их пространственного различия. …
…
Контекст 4
… o C) дают меньше листьев, чем контрольные растения, выращенные при 22 o C (рис. 1А и таблица S1). Эти листья имеют уменьшенный размер и меньшую плотность устьиц, чем листья, развившиеся при 22 o C, даже при хорошем поливе (рис. 1В и таблица S1) [9]. …
Контекст 5
… насыщали водой, герметизировали (для предотвращения потери воды из почвы), взвешивали, рандомизировали и возвращали к обеим температурам. Сходные показатели транспирации наблюдались между растениями, выращенными при 22 o C, и растениями, выращенными при 28 o C при 22 o C (22-22 и 28-22; рисунок 1C и таблица S1).Удивительно, но, учитывая, что у них меньше устьиц, растения, выращенные при 28°C, демонстрировали большую транспирацию при 28°C (28-28), чем растения, выращенные при 22°C ([22][23][24][25][26] [27][28] (Рисунок 1B,C и Таблица S1) …
Контекст 6
… учитывая, что у них меньше устьиц, растения, выращенные при 28 o C, демонстрировали большую транспирацию при 28 o C (28-28) по сравнению с растениями, выращенными при 22°C ([22][23][24][25][26][27][28] (рис. 1B,C и таблица S1). Аналогичное, но преувеличенное, реакция наблюдалась при постоянном освещении, когда устьица не подвергались воздействию сигналов закрытия, вызванных темнотой (рис. S1 и таблица S1) [6]….
1B,C и таблица S1). Аналогичное, но преувеличенное, реакция наблюдалась при постоянном освещении, когда устьица не подвергались воздействию сигналов закрытия, вызванных темнотой (рис. S1 и таблица S1) [6]….
Контекст 7
… 22 o C существенных различий в температуре листьев между 22-22 и 28-22 растениями не наблюдалось (таблица S1). Однако при 28 o C листья 28-28 растений были примерно на 1 o C холоднее, чем 22-28 растений (рис. 1D и таблица S1). Увеличенная разница (>2 o C) наблюдалась между 22-28 и 28-28 растениями при постоянном освещении, что согласуется с повышенной интенсивностью транспирации, наблюдаемой в этих экспериментах (рисунок S1 и таблица S1)….
Контекст 8
… 28 o C, однако листья растений 28-28 были примерно на 1 o C холоднее, чем растения 22-28 (рис. 1D и таблица S1). Увеличенная разница (>2 o C) наблюдалась между 22-28 и 28-28 растениями при постоянном освещении, что согласуется с повышенной интенсивностью транспирации, наблюдаемой в этих экспериментах (рисунок S1 и таблица S1). …
…
Влияние регуляторной архитектуры на наследуемость в широком и узком смысле и продукционно-биологической генетики.Что определяет большую вариацию в зарегистрированных соотношениях
V A / V G из экспериментов с пересечением линий, не совсем понятно. Здесь мы сообщаем, как соотношение V A / V G и, следовательно, соотношение между наследуемостью в узком и широком смысле ( h 2 / H 2 9003) изменяется как функция6 9003 регулирующей архитектуры, лежащей в основе карт генотип-фенотип (GP). Мы изучили пять динамических моделей (пути цАМФ, гликолиза, циркадных ритмов, клеточного цикла и динамики клеток сердца).Мы предположили, что генетическая изменчивость отражается в параметрах модели, и выделили фенотипы, резюмирующие системную динамику. Даже при наложении чисто линейного генотипа на карты параметров и отсутствии вариации окружающей среды мы наблюдали довольно низкие отношения V A / V G . В частности, системы с положительной обратной связью и циклической динамикой давали более немонотонные карты генотип-фенотип и гораздо более низкие отношения V A / V G , чем системы без обратной связи.Результаты показывают, что некоторые регуляторные архитектуры постоянно поддерживают прозрачные отношения между генотипом и фенотипом, тогда как другие архитектуры генерируют более тонкие паттерны. Наш подход можно использовать для систематического выяснения этих взаимосвязей в целом ряде биологических систем.
В частности, системы с положительной обратной связью и циклической динамикой давали более немонотонные карты генотип-фенотип и гораздо более низкие отношения V A / V G , чем системы без обратной связи.Результаты показывают, что некоторые регуляторные архитектуры постоянно поддерживают прозрачные отношения между генотипом и фенотипом, тогда как другие архитектуры генерируют более тонкие паттерны. Наш подход можно использовать для систематического выяснения этих взаимосвязей в целом ряде биологических систем.Резюме автора
Наследуемость в широком смысле признака представляет собой долю фенотипической изменчивости, приписываемую генетическим причинам, в то время как наследуемость в узком смысле представляет собой долю, приписываемую аддитивным генным эффектам.Лучшее понимание того, что лежит в основе изменчивости соотношения двух показателей наследуемости, или эквивалентного отношения аддитивной дисперсии V A к общей генетической изменчивости V G , важно для продукционной биологии, биомедицины и эволюции. Мы обнаруживаем, что сообщаемые значения V A / V G от пересечений линий сильно различаются, и задаемся вопросом, можно ли объяснить биологические механизмы, лежащие в основе таких различий, путем связывания моделей вычислительной биологии с генетикой.С этой целью мы использовали модели пути цАМФ, гликолиза, циркадных ритмов, клеточного цикла и динамики кардиоцитов. Мы предположили аддитивное действие генов от генотипов к параметрам модели и изучили полученные карты GP и соотношения фенотипов на системном уровне. Наши результаты показывают, что некоторые типы регуляторных архитектур неизменно сохраняют прозрачные отношения между генотипом и фенотипом, тогда как другие генерируют более тонкие паттерны.В частности, системы с положительной обратной связью и циклической динамикой приводили к большей немонотонности карты ГП, что приводило к более низким отношениям V A / V G . Наш подход может быть использован для систематического выяснения взаимосвязей между целым рядом биологических систем.
Мы обнаруживаем, что сообщаемые значения V A / V G от пересечений линий сильно различаются, и задаемся вопросом, можно ли объяснить биологические механизмы, лежащие в основе таких различий, путем связывания моделей вычислительной биологии с генетикой.С этой целью мы использовали модели пути цАМФ, гликолиза, циркадных ритмов, клеточного цикла и динамики кардиоцитов. Мы предположили аддитивное действие генов от генотипов к параметрам модели и изучили полученные карты GP и соотношения фенотипов на системном уровне. Наши результаты показывают, что некоторые типы регуляторных архитектур неизменно сохраняют прозрачные отношения между генотипом и фенотипом, тогда как другие генерируют более тонкие паттерны.В частности, системы с положительной обратной связью и циклической динамикой приводили к большей немонотонности карты ГП, что приводило к более низким отношениям V A / V G . Наш подход может быть использован для систематического выяснения взаимосвязей между целым рядом биологических систем.
Введение
Наследуемость признака в широком смысле — это доля фенотипической изменчивости, приписываемая генетическим причинам, тогда как наследуемость в узком смысле — это доля, приписываемая аддитивному действию генов.Общая генетическая изменчивость включает дисперсию, объясняемую внутрилокусным доминированием () и межлокусными взаимодействиями (). Причины и важность этой неаддитивной генетической изменчивости, которая отличает две меры наследуемости, были предметом серьезных споров на протяжении более 80 лет (например, [1]–[6]). Недавно с помощью статистических аргументов было показано, что для признаков со многими локусами с экстремальными частотами аллелей большая часть генетической изменчивости становится аддитивной с V G
) обычно >0.5 [3]. В популяциях с промежуточными частотами аллелей, таких как контролируемые скрещивания линий, часто сообщают о более низких соотношениях V A / V G [7], [8]. Это проиллюстрировано на графике, который обобщает расчетные отношения V A / V G из коллекции исследований таких популяций. Этот широкий диапазон соотношений h 2 / H 2 , зарегистрированный для скрещивания линий, не может быть объяснен аргументом частоты аллелей, и предполагаемые объяснения должны быть основаны на том, как регуляторная архитектура лежащих в основе биологических систем формирует карта генотип-фенотип (ГФ).
Этот широкий диапазон соотношений h 2 / H 2 , зарегистрированный для скрещивания линий, не может быть объяснен аргументом частоты аллелей, и предполагаемые объяснения должны быть основаны на том, как регуляторная архитектура лежащих в основе биологических систем формирует карта генотип-фенотип (ГФ).Таблица 1
Примеры зарегистрированных соотношений V A / V G из экспериментов по пересечению линий.
| Вид | Тип признаков | Количество признаков | Ref. | Мин. В А / В Г | Макс. В А / В Г | Среднее В А / В Г |
| Цыпленок | Рост, вес | 17 | [45] | 0. 03 03 | 0,71 | 0,29 |
| Мышь | Гипероксическая выживаемость | 1 | [46] | — | — | 0,46 |
| Дрозофила меланогастер | Локомоторное поведение | 1 | [47] | — | — | 0,31 |
| Дрозофила меланогастер | Обонятельное поведение | 1 | [48] | — | — | 0. 64 64 |
| Хлопок нагорный | Устойчивость к нематодам | 1 | [49] | — | — | 0,79 |
| Дыня | Цвет и зрелость плода | 2 | [50] a | 0,55 | 0,58 | 0,57 |
| Кукуруза | Морфологические признаки | 17 | [51] б | 0,13 | 1,1 | 0. 59 59 |
| Кукуруза | Устойчивость к пятнистости листьев | 5 | [52] | 0,51 | 0,95 | 0,76 |
| Arabidopsis thaliana | Цветение и морфология и | 22 | [53] c | 0,58 | 1,05 | 0,76 |
| Баклажаны | Признаки, связанные с каллюсом | 4 | [54] | 0,42 | 0. 93 93 | 0,73 |
По нескольким причинам важно понять причинно-следственные связи наблюдаемых вариаций в соотношениях h 2 /H 2 /H 2 внутри и между биологическими системами. В количественной генетике человека, где обычно используются исследования близнецов, большинство оценок наследуемости относятся к H 2 [9]. В тех случаях, когда h 2 /H 2 является низким, это может привести к нереалистичным ожиданиям относительно того, какая часть основной причинной вариации может быть обнаружена с помощью методов обнаружения линейного QTL [6].С другой стороны, низкая наследуемость данного сложного признака в узком смысле не обязательно означает, что окружающая среда определяет большую часть вариаций. В эволюционной биологии аддитивная изменчивость является основной валютой эволюционной адаптации и эволюционируемости. Важными вопросами в этом контексте являются, например, (i) в какой степени происходит отбор самих регуляторных анатомий для поддержания высокой аддитивной дисперсии, (ii) существуют ли организационные ограничения в построении адаптивных систем, такие, что в некоторых случаях низкий уровень h 2 / H 2 Соотношение обязательно должно возникнуть, пока еще выбрано проксимальное решение? Более того, в продукционной биологии с генетически модифицированными организмами, размножающимися половым путем, хотелось бы гарантировать, что модификации будут передаваться будущим поколениям полностью предсказуемым образом. Таким образом, хотелось бы гарантировать, что модификация станет в высокой степени наследуемой в узком смысле.
Таким образом, хотелось бы гарантировать, что модификация станет в высокой степени наследуемой в узком смысле.
В качестве шага к физиологически обоснованному пониманию изменчивости h 2 / H 2 взаимосвязей между биологическими системами или процессами мы поставили вопрос: существуют ли регуляторные структуры или определенные классы фенотипов? V A / V G более низкие отношения, чем другие? Решение этого вопроса требует увязки генетической изменчивости с вычислительной биологией в популяционном контексте (т.g., [10]–[19]), так называемое причинно-связное моделирование генотип-фенотип (cGP) [15], [17], [18]. Мы применили этот подход к пяти хорошо проверенным моделям вычислительной биологии, описывающим, соответственно, метаболический путь гликолиза у почкующихся дрожжей [20], сигнальный путь циклического аденозинмонофосфата (цАМФ) у почкующихся дрожжей [21], регуляцию клеточного цикла у почкующихся дрожжей. [22], генная сеть, лежащая в основе циркадных ритмов млекопитающих [23], и ионные каналы, определяющие потенциал действия в сердечных миоцитах мышей [24]. Эти модели различаются по своей регуляторной архитектуре; ниже мы показываем, что они также различаются диапазоном отношений, которые они могут проявлять.В частности, регулирование с положительной обратной связью и колебательное поведение, по-видимому, располагают к низким отношениям V A / V G . Результаты показывают, что наш подход можно использовать в общем виде для изучения того, как соотношение h 2 / H 2 изменяется в зависимости от регуляторной анатомии.
[22], генная сеть, лежащая в основе циркадных ритмов млекопитающих [23], и ионные каналы, определяющие потенциал действия в сердечных миоцитах мышей [24]. Эти модели различаются по своей регуляторной архитектуре; ниже мы показываем, что они также различаются диапазоном отношений, которые они могут проявлять.В частности, регулирование с положительной обратной связью и колебательное поведение, по-видимому, располагают к низким отношениям V A / V G . Результаты показывают, что наш подход можно использовать в общем виде для изучения того, как соотношение h 2 / H 2 изменяется в зависимости от регуляторной анатомии.
Методы
Моделирование моделей cGP
Пять моделей cGP были построены и проанализированы с помощью cgptoolbox (http://github.com/jonovik/cgptoolbox) пакет Python с открытым исходным кодом, разработанный авторами; дополнительный исходный код, относящийся к моделированию в этой статье, доступен по запросу. Далее мы описываем три основные части рабочего процесса: (i) сопоставление генотипов с параметрами, (ii) сопоставление параметров с фенотипами, т. е. решение динамических моделей, и (iii) настройка симуляций Монте-Карло, объединяющих два сопоставления (рисунок S1). Для каждой модели мы кратко описываем ее происхождение, программное обеспечение, используемое для ее решения, какие параметры подвергались генетической изменчивости, какие фенотипы были зарегистрированы, а также критерии исключения наборов данных.На рисунках S2, S3, S4, S5, S6 показаны графические представления пяти модельных систем, а текст S1 содержит более подробные описания всех пяти моделей.
Далее мы описываем три основные части рабочего процесса: (i) сопоставление генотипов с параметрами, (ii) сопоставление параметров с фенотипами, т. е. решение динамических моделей, и (iii) настройка симуляций Монте-Карло, объединяющих два сопоставления (рисунок S1). Для каждой модели мы кратко описываем ее происхождение, программное обеспечение, используемое для ее решения, какие параметры подвергались генетической изменчивости, какие фенотипы были зарегистрированы, а также критерии исключения наборов данных.На рисунках S2, S3, S4, S5, S6 показаны графические представления пяти модельных систем, а текст S1 содержит более подробные описания всех пяти моделей.
Сопоставление генотипа с параметрами
Для каждой модели следующая процедура повторялась 1000 раз (см. также «Моделирование методом Монте-Карло» ниже) для различных наборов параметров, подлежащих моделируемой генетической изменчивости. Мы начали с выборки трех полиморфных локусов, каждый из которых определял один или два параметра в динамической модели. Таблицы подходящих локусов с соответствующими параметрами и их исходными значениями перечислены в таблицах S1, S2, S3, S4, S5, соответствующих моделям цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно. Наследуемую изменчивость выбранного параметра генерировали для одного биаллельного локуса с аллельными индексами 0 и 1 следующим образом. Сначала равномерно в интервале [0,7, 1,3] были выбраны два числа r 1 и r 2 . Значение параметра для гомозиготы 00 было установлено таким, где b — исходное значение, для гомозиготы 11 значение параметра было равно .Гетерозиготному генотипу 01 было присвоено среднее значение двух гомозигот, что привело к аддитивному сопоставлению генотипов со значениями параметров.
Таблицы подходящих локусов с соответствующими параметрами и их исходными значениями перечислены в таблицах S1, S2, S3, S4, S5, соответствующих моделям цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно. Наследуемую изменчивость выбранного параметра генерировали для одного биаллельного локуса с аллельными индексами 0 и 1 следующим образом. Сначала равномерно в интервале [0,7, 1,3] были выбраны два числа r 1 и r 2 . Значение параметра для гомозиготы 00 было установлено таким, где b — исходное значение, для гомозиготы 11 значение параметра было равно .Гетерозиготному генотипу 01 было присвоено среднее значение двух гомозигот, что привело к аддитивному сопоставлению генотипов со значениями параметров.
Модель цАМФ
Модель полного сигнального пути цАМФ у S. cerevisiae [21], взяв внешний уровень глюкозы в качестве входных данных, был загружен в виде кода SBML (http://www. biomedcentral.com/content/supplementary/1752-0509-3-70-s1.xml) и интегрирован с использованием PySCeS [25]. Генетическая изменчивость была введена в отношении скоростей ассоциации/диссоциации и фосфорилирования/дефосфорилирования сигнальных белков (см. Рисунок S2 и Таблицу S1).Начальные равновесные концентрации перед добавлением внешней глюкозы, пиковые значения после добавления глюкозы и время, необходимое для достижения пиковых значений клеточных белков, регистрировали как фенотипы (см. иллюстрацию фенотипа и Таблицу S6 для описания фенотипа).
biomedcentral.com/content/supplementary/1752-0509-3-70-s1.xml) и интегрирован с использованием PySCeS [25]. Генетическая изменчивость была введена в отношении скоростей ассоциации/диссоциации и фосфорилирования/дефосфорилирования сигнальных белков (см. Рисунок S2 и Таблицу S1).Начальные равновесные концентрации перед добавлением внешней глюкозы, пиковые значения после добавления глюкозы и время, необходимое для достижения пиковых значений клеточных белков, регистрировали как фенотипы (см. иллюстрацию фенотипа и Таблицу S6 для описания фенотипа).
Графическая иллюстрация фенотипов, зарегистрированных для пяти исследованных моделей cGP. Графики времени (переменная состояния на оси Y, время на оси X) для базового набора параметров отображаются для всех пяти моделей. A. В отсутствие внешней глюкозы все переменные состояния (показана концентрация цАМФ) в модели цАМФ [21] сходятся к стабильному стационарному состоянию (синий кружок на оси ординат). После добавления внешней глюкозы (добавлено 5 мМ в момент времени 50) мы видим быстрое изменение, за которым следует медленный возврат к исходному устойчивому состоянию. В дополнение к исходному стационарному состоянию в качестве фенотипов регистрировали экстремальную концентрацию (верхний синий кружок), а также время достижения экстремума (синяя линия). Б. Концентрации метаболитов (показаны внутренняя глюкоза (GLCi), глюкозо-6-фосфат (G6P) и фруктоза-6-фосфат (F6P)) в модели гликолиза [20] сходятся к стабильному стационарному состоянию, обозначенному незаштрихованными кружками. Стационарные концентрации для 13 метаболитов были зарегистрированы как фенотипы этой модели. C. Для модели клеточного цикла [22] мы записали пиковый уровень и время от основания до пика, как и для циркадной модели (), и, кроме того, мы записали события клеточного цикла, такие как появление почек в то время, когда [BUD ] = 1 обозначено черной стрелкой. D. Концентрации мРНК и белка (показаны мРНК для Bmal1 (MB), Cry (MC) и Per (MP)) в циркадной модели [23] сходятся к предельному циклу.
После добавления внешней глюкозы (добавлено 5 мМ в момент времени 50) мы видим быстрое изменение, за которым следует медленный возврат к исходному устойчивому состоянию. В дополнение к исходному стационарному состоянию в качестве фенотипов регистрировали экстремальную концентрацию (верхний синий кружок), а также время достижения экстремума (синяя линия). Б. Концентрации метаболитов (показаны внутренняя глюкоза (GLCi), глюкозо-6-фосфат (G6P) и фруктоза-6-фосфат (F6P)) в модели гликолиза [20] сходятся к стабильному стационарному состоянию, обозначенному незаштрихованными кружками. Стационарные концентрации для 13 метаболитов были зарегистрированы как фенотипы этой модели. C. Для модели клеточного цикла [22] мы записали пиковый уровень и время от основания до пика, как и для циркадной модели (), и, кроме того, мы записали события клеточного цикла, такие как появление почек в то время, когда [BUD ] = 1 обозначено черной стрелкой. D. Концентрации мРНК и белка (показаны мРНК для Bmal1 (MB), Cry (MC) и Per (MP)) в циркадной модели [23] сходятся к предельному циклу. В дополнение к периоду колебаний (длинная синяя линия) для каждой из 16 переменных уровень пика (незакрашенный синий кружок), а также время от основания до пика (короткая синяя линия) регистрировались как фенотипы. E. Мы использовали базовый уровень, пиковый уровень, амплитуду, время до пика и время до 25%, 50%, 75% и 90% восстановления потенциала действия и транзиент кальция в качестве фенотипов клеточного уровня модели потенциала действия [24].На рисунке показан потенциал действия.
В дополнение к периоду колебаний (длинная синяя линия) для каждой из 16 переменных уровень пика (незакрашенный синий кружок), а также время от основания до пика (короткая синяя линия) регистрировались как фенотипы. E. Мы использовали базовый уровень, пиковый уровень, амплитуду, время до пика и время до 25%, 50%, 75% и 90% восстановления потенциала действия и транзиент кальция в качестве фенотипов клеточного уровня модели потенциала действия [24].На рисунке показан потенциал действия.
Модель гликолиза
Модель, опубликованная Teusink et al. [20] описывает гликолиз у S. cerevisiae через кинетику 13 гликолитических ферментов, определяющих потоки переменных состояния метаболитов. Генетическая изменчивость была введена в отношении максимальных скоростей реакции ферментов (см. Рисунок S3 и Таблицу S2). Мы загрузили модель из базы данных BioModels (http://www.ebi.ac.uk/biomodels-main/BIOMD0000000064) в SBML L2 V1 и решили ее с помощью PySCeS [25], чтобы найти стабильные равновесные концентрации метаболитов, которые использовали в качестве фенотипов (см. и Таблицу S7).Наборы данных отбрасывались, если один или несколько генотипов не давали стабильного стационарного состояния, что может произойти из-за бифуркации седловидного узла [26].
и Таблицу S7).Наборы данных отбрасывались, если один или несколько генотипов не давали стабильного стационарного состояния, что может произойти из-за бифуркации седловидного узла [26].
Модель клеточного цикла
Модель консенсусных механизмов контроля клеточного цикла у S. cerevisae , смоделированная алгебраическими/дифференциальными уравнениями, описывающими непрерывные изменения переменных состояния и разрывы из-за клеточных событий [22], была получена из репозиторий CellML (http://models.cellml.org/workspace/chen_calzone_csikasznagy_cross_novak_tyson_2004).Генетическая изменчивость была введена в скорость производства и распада различных белков (см. Рисунок S4 и Таблицу S3). Опубликованная модель содержит правила сброса (события) как для параметров, так и для переменных состояния, но реализация CellML включает только параметр ( k безумный2 , к буб2 и к lte1 ) правил. Правила сброса для переменных состояния [BUD], [SPN] и [ORI], как описано в документе о модели, были реализованы путем решения модели с поиском корня для соответствующих переменных.Модель численно интегрировали с помощью решателя CVODE [27] до сходимости времени клеточного деления, событий клеточного цикла. Пиковые уровни и время до пиковых уровней для концентраций цитозольного белка вместе с синхронизацией событий клеточного деления регистрировали как фенотипы (см. иллюстрации фенотипов и Таблицу S8 для описания фенотипов).
Циркадианная модель
Модель циркадианных часов млекопитающих, опубликованная Leloup и Goldbeter [23], описывает динамику мРНК и белков трех генов в цитозоле и ядре.Генетическая изменчивость была введена в отношении скорости распада мРНК (см. Рисунок S5 и Таблицу S4). Модель была загружена из репозитория CellML (http://models.cellml.org/workspace/leloup_goldbeter_2004) и интегрирована с помощью CVODE [27] до сходимости к ее предельному циклу. В качестве фенотипов использовали нижние уровни и время от нижнего уровня до пикового значения концентраций мРНК, белков и белковых комплексов. Кроме того, мы записали период колебаний (см. иллюстрации фенотипов и Таблицу S9 для описания фенотипов).
Модель потенциала действия
Модель [24] является расширением [28] и описывает потенциал действия и кальциевый переходный процесс в клетке сердечной мышцы мыши. Мы получили код CellML от авторов, и файл доступен в качестве дополнительного материала в [17]. Численное интегрирование было выполнено с использованием CVODE [27]. Генетическая изменчивость была введена в отношении максимальной проводимости ионных каналов и сродства насоса (см. Рисунок S6 и Таблицу S5). Фенотипы были созданы путем имитации регулярной кардиостимуляции, как это было сделано в [17], [18], при этом ток калия стимула -15 В / с длился 3 мс в начале каждого интервала стимула.Модель моделировалась до сходимости, как описано в [17]; наборы данных, которые не сошлись, отбрасывались. Начальный уровень, пиковый уровень, амплитуда и время до 25, 50, 75 и 90% восстановления потенциала действия и кальциевого переходного процесса регистрировались как фенотипы клеточного уровня (см. иллюстрации фенотипов и Таблицу S10 для описания фенотипов).
Моделирование Монте-Карло
Для каждой модели мы выполнили 1000 симуляций Монте-Карло следующим образом (см. рисунок S1 для иллюстрации).Сначала мы отобрали три полиморфных локуса для введения генетической изменчивости и отобрали карту генотип-параметр, как описано выше. Затем были пронумерованы все 27 возможных трехлокусных генотипов, картированы в 27 наборов параметров, и для каждого набора параметров была решена динамическая модель и получены фенотипы (как описано выше и в ). Чтобы избежать артефактов, возникающих из-за численного шума, наборы данных с низкой изменчивостью были исключены из генетического анализа. Абсолютную изменчивость измеряли как разницу между максимальным и минимальным значениями фенотипа в наборе данных, а относительную изменчивость — как отношение абсолютной изменчивости к среднему фенотипу того же набора данных.Пороговые значения для каждого фенотипа и количество наборов данных, превышающих пороги, перечислены в таблицах S6, S7, S8, S9, S10 для моделей цАМФ, гликолиза, клеточного цикла, циркадного ритма и потенциала действия соответственно.
Результаты/Обсуждение
Классификация систем и размерность фенотипа
Пять моделей cGP, изученных в этой работе, различаются по нескольким параметрам, как по своим функциям, так и по базовой сетевой структуре. Модели гликолиза и цАМФ являются моделями путей с ациклической серией реакций, преобразующих входы в выходы.Модель гликолиза [20] является более продвинутой, чем метаболические модели в более ранних генетически ориентированных исследованиях (например, [3], [31], [32]), поскольку она включает множество различных типов кинетики ферментов, а также регуляцию некоторых ферментов с помощью отрицательной обратной связи. активности ферментов за счет ингибирования продукта. Модель цАМФ [21] содержит ряд петель отрицательной обратной связи, но в целом она также имеет четкую структуру путей, где сигнал глюкозы передается от G-белков к цАМФ и к целевой киназы РКА. Обе эти модели имеют общую относительно простую динамику с решениями, сходящимися к устойчивому стационарному состоянию ().
Напротив, три другие модели демонстрируют циклическое поведение в результате взаимодействия между положительными и отрицательными обратными связями (). Однако между этими моделями также есть явные различия. Модель клеток сердца [24] представляет собой возбудимую систему с механизмами обратной связи, включая индуцированное кальцием высвобождение кальция и несколько зависимых от напряжения ионных каналов. В отличие от клеток-водителей ритма, он полагается на внешнюю стимуляцию, чтобы инициировать потенциал действия. Модель циркадного ритма [23], [33] представляет собой сеть экспрессии генов с переплетенными положительными и отрицательными петлями обратной связи транскрипции, дающими осциллятор предельного цикла с устойчивыми колебаниями даже в сплошной темноте.Модель клеточного цикла [22] основывается на петле положительной обратной связи между циклинами В-типа в ассоциации с циклинзависимой киназой и ингибиторами циклинзависимой киназы, которая устанавливает петлю гистерезиса, заставляющую клеточный цикл возникать из переходов между двумя альтернативными стабильными киназами. устойчивые состояния.
Эта грубая классификация пяти моделей cGP на модели путей и более сложные регуляторные системы ясно отражена в эффективной размерности фенотипов, возникающих в наших симуляциях Монте-Карло.Мы изучили фенотипическую размерность для всех пяти моделей cGP с помощью анализа основных компонентов (PCA) для каждой симуляции Монте-Карло (). Во всех смоделированных наборах данных 95% фенотипической изменчивости моделей гликолиза и цАМФ можно объяснить первыми 3 основными компонентами, для моделей клеточного цикла и клеток сердца требуются первые 5 основных компонентов, а для циркадной модели требуется 7 компонентов. Поскольку карты генотип-параметр являются аддитивными для всех пяти моделей, эти различия в эффективной размерности фенотипов высокого уровня указывают на то, что сопоставления параметров с фенотипами проще для моделей путей, чем для трех других моделей.Это вместе с результатами о влиянии положительной обратной связи на статистический эпистаз в сетях регуляции генов [11] позволяет предположить, что модели гликолиза и цАМФ могут приводить к более высоким соотношениям V A / V G , чем другие модели. три модели.
Размерность фенотипической изменчивости.Доля общей объясненной фенотипической изменчивости (ось y) по сравнению с количеством основных компонентов (ось x) во всех пяти моделях cGP (обозначены цветом).Для каждого набора данных Монте-Карло матрицы, содержащие полную карту генотип-фенотип для всех M зарегистрированных фенотипов, были стандартизированы до единичной дисперсии перед анализом основных компонентов. Каждая ящичковая диаграмма суммирует объясненную дисперсию для почти 1000 симуляций Монте-Карло.
Отношение аддитивной генетической дисперсии к общей генетической дисперсии
Результаты подтвердили наши ожидания в отношении высоких соотношений V A / V G для моделей гликолиза и цАМФ.Кроме того, появилось несколько различных моделей. Модель цАМФ показывает общие самые высокие значения соотношений V A / V G (и Таблица S6) со средними и медианными значениями выше 0,99 для всех зарегистрированных фенотипов. Квантиль 0,05 (т. е. только 5 процентов моделирования Монте-Карло показывают более низкие значения, чем это) V A / V G значения были выше 0,97 для всех фенотипов, и не наблюдались значения ниже 0,6.Другими словами, внутри- и межлокусная аддитивная модель действия генов очень хорошо аппроксимирует карты генотип-фенотип, возникающие из этой модели cGP.
Эмпирическая кумулятивная функция распределения соотношений V A / V G для фенотипов моделей цАМФ (A) и гликолиза (B).A. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели цАМФ: начальные равновесные концентрации до добавления внешней глюкозы циклического аденозинмонофосфата (цАМФ), G-белка Ras2a (Ras2a), фактора гуанин-нуклеотидного обмена (Cdc25), протеинкиназы А (PKAi).Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v). Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). См. Таблицу S6 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирическая кумулятивная функция распределения (ось y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза: ацетальдегид (ACE), 1,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).См. Таблицу S7 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
Модель гликолиза также имеет средние и медианные значения, близкие к 1 для всех фенотипов (и Таблица S7). Но по сравнению с моделью цАМФ цифры явно ниже; наименьшее зарегистрированное среднее значение (фенотип BPG ) составляет 0,9, а 0,05-квантильные значения ниже 0.7 для некоторых фенотипов. Несколько значений ниже 0,5 наблюдаются для всех фенотипов. Распределение соотношений V A / V G для модели клеточного цикла (рисунок S7 и таблица S8) очень похоже на распределение модели гликолиза с самым низким средним значением V A / V G значение 0,93 для время до пика для Sic1 и с 0,05-квантилями ниже 0.8 для некоторых фенотипов. V A / V G значения ниже 0,1 наблюдаются для нескольких симуляций методом Монте-Карло в некоторых фенотипах.
Для каждой из моделей цАМФ, гликолиза и клеточного цикла распределения соотношений V A / V G были довольно сходными для всех фенотипов, и подавляющее большинство симуляций методом Монте-Карло показали очень высокие отношения . Модель циркадных часов отличается от этих трех моделей как с точки зрения отображения больших различий между фенотипами, так и с точки зрения наличия гораздо большей доли низких значений (и Таблица S9).Выделяются четыре фенотипа с распределениями V A / V G , которые напоминают равномерное распределение U (0,1). Это время от основания до пика для фосфорилированных и нефосфорилированных белков Per и Cry , и они имеют медианные значения V A / V G в диапазоне от 0,46 до 0,70 и 0,0,036. значения в диапазоне от 0,04 до 0,10. Остальные фенотипы имеют несколько более высокие значения V A / V G , но более половины зарегистрированных фенотипов имеют 0.05-квантили ниже 0,6. Медианные значения V A / V G ниже 0,9 для большинства фенотипов модели потенциала действия. И все зарегистрированные фенотипы имеют большую долю низких соотношений V A / V G (и Таблица S10) с 0,05-квантилями в диапазоне 0,18-0-35. Распределения для потенциалов действия и переходных кальциевых фенотипов весьма сходны, но время до 90% реполяризации для потенциала действия показывает несколько более высокие значения, чем для других.
Эмпирическая кумулятивная функция распределения соотношений V A / V G для фенотипов циркадной модели (A) и модели потенциала действия (B).Эмпирические кумулятивные функции распределения (ось Y) отношений V A / V G (ось x) для фенотипов, изученных в циркадной модели и модели клеток сердца. A. На верхней левой панели (Bmal1) показаны фенотипы, связанные с геном bmal1 , включая мРНК (MB), нефосфорилированный/фосфорилированный белок в цитозоле (BC/BCP) и ядре (BN/BNP).Нижняя правая панель (Per) предназначена для на генов, включая мРНК (MP), нефосфорилированный белок (PC) и фосфорилированный белок (PCP). Нижняя концентрация (сплошная линия) и время, необходимое для достижения пика (пунктирная линия) каждого вида, являются зарегистрированными фенотипами. Нижняя левая панель (Cry) связана с геном cry , включая мРНК (MC), нефосфорилированный белок (CC) и фосфорилированный белок (CCP). Верхняя правая панель (комплекс) предназначена для белковых комплексов PCC, PCN, PCCP и PCNP.Также показан период циркадного ритма (Период, пунктирная линия). См. Таблицу S9 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G . B. Эмпирические кумулятивные функции распределения (ось y) V A / V G отношения (ось x) для фенотипов, изучаемых в модели потенциала действия: время до 25%, 50%, 75 Показаны % и 90% начальных значений, амплитуда, начальные значения (база), пиковые значения, время до достижения пиковых значений потенциала действия (левая панель) и транзиент кальция (правая панель).См. Таблицу S10 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
Все пять моделей cGP способны создавать V A / V G соотношения, близкие к 1, и, кроме двух фенотипов для циркадной модели, все медианные значения V A / / 3 G значительно выше 0,5. Это подтверждает гипотезу [30] о том, что биологические системы склонны включать регуляторные механизмы, что в целом приводит к значительной аддитивной генетической изменчивости даже при промежуточных частотах аллелей.Это не означает, что отбор иногда не может привести к регуляторным решениям, стремящимся к низким соотношениям V A / V G ; на самом деле частота низких соотношений V A / V G заметно различалась среди пяти изученных нами моделей. Поскольку карты параметров генотипа были чисто аддитивными, вся неаддитивная генетическая дисперсия является результатом нелинейности моделей ОДУ. Ожидаемым эффектом введения неаддитивности в карты генотип-параметр будет дальнейшее снижение отношений V A / V G .
Наш вывод о том, что модели со сложной регуляцией, включающей петли положительной обратной связи, имеют тенденцию давать более низкие значения V A / V G , согласуется с предыдущим исследованием сетей регуляции генов [11]. Принимая во внимание относительно высокие отношения V A / V G модели клеточного цикла по сравнению с моделями циркадианного цикла и потенциала действия, следующая цитата из обсуждения Тайсона и Новака [34] о том, почему клеточный цикл более информативен как петля гистерезиса, чем как осциллятор предельного цикла (LCO): « Вообще говоря, период LCO является сложной функцией всех кинетических параметров в дифференциальных уравнениях.Однако период цикла клеточного деления зависит только от одного кинетического свойства клетки: времени удвоения ее массы. ”Это, кажется, объясняет, почему карты генотип-фенотип, полученные из моделей клеточного цикла, гораздо более линейны, чем карты из циркадной модели, которая является LCO.
Монотонность объясняет большую часть паттернов V
A /V GВ данной популяции V A / V G является функцией частот всех элементов, как Карты GP с сильными взаимодействиями все же могут давать высокие значения V A / V G в популяциях с экстремальными частотами аллелей [3].В популяциях с промежуточными частотами аллелей значения V A / V G определяются в основном формой карты генотип-фенотип и наблюдаемыми различиями между пятью моделями cGP в распределении V Значения / V G побуждают к поиску лежащих в их основе объяснительных принципов.
Недавно предложенная концепция монотонности (или сохранения порядка) карт GP кажется одним из таких принципов.Короче говоря, карта ГП называется монотонной, если порядок генотипов по содержанию генов (количеству аллелей данного типа) сохраняется в порядке соответствующих фенотипических значений (подробности см. в [30]). изображает три экстремальных типа карт GP, наблюдаемых в наших симуляциях. Почти аддитивные карты GP, как показано на рисунке, дают значения V A / V G , очень близкие к единице. Карты GP с сильным эпистазом величины, но все еще сохраняющим порядок, обычно приводят к промежуточным значениям V A / V G (), в то время как очень немонотонные или нарушающие порядок карты GP () демонстрируют сильное сверхдоминирование и/или знак эпистаза приводят к значениям V A / V G , близким к нулю.
Три различных типа карт генотип-фенотип.Примеры трех различных типов карт генотип-фенотип, наблюдаемых в наших симуляциях. Для каждой субфигуры значение фенотипа показано на оси ординат, тогда как оси абсцисс, цвета линий и столбцы графика указывают генотип в трех локусах. A. Почти аддитивная карта, иллюстрируемая картой GP времени до пиковой концентрации Cdc25 ( V A / V G = 0,99) в модели цАМФ; Б. Полностью монотонная, но неаддитивная карта, представленная на примере карты GP концентрации белка P2G ( V A / V G = 0,41) в модели гликолиза; и, C. Найдена сильно немонотонная карта времени до пиковой концентрации белка PC ( V A / V G = 0,03) из циркадной модели.
Основываясь на недавних результатах исследований сетей регуляции генов [30], мы ожидали, что три модели cGP со сложной регуляцией, включающей положительную обратную связь, приведут к значительно большему количеству немонотонных или нарушающих порядок карт GP, чем модели с двумя путями.Чтобы проверить это, мы измерили количество нарушений порядка во всех смоделированных картах GP (см. Методы) и обнаружили очень четкую закономерность (). В то время как наборы данных из моделей гликолиза и цАМФ содержали только 1,1% и 1,3% карт GP с нарушением порядка для любого локуса, данные из моделей клеточного цикла, циркадного ритма и потенциала действия содержали 20,7%, 44,4% и 69,5% соответственно. Более того, монотонные карты GP давали более высокие значения V A / V G , чем немонотонные карты GP для всех пяти моделей cGP (критерий Манна-Уитни; значения p ниже 1e-10 для всех пяти моделей) .
Количество локусов, для которых карта GP показывает нарушение порядка.Количество симуляций Монте-Карло, в которых карта GP для данного фенотипа явно нарушает порядок (карты GP с N/A >0,05, см. Методы), показано для модели цАМФ ( A ), модель гликолиза ( B ), модель клеточного цикла ( C ), циркадианную модель ( D ) и модель потенциала действия ( E ). Показаны только фенотипы, в которых хотя бы одно моделирование Монте-Карло привело к нарушению порядка карты GP.
Однако, несмотря на то, что модель гликолиза редко показывает нарушение порядка даже для одного локуса, она обладает гораздо более низкими значениями V A / V G , чем модель цАМФ. Правдоподобным объяснением является то, что стационарные концентрации метаболитов могут заметно увеличиваться при значениях параметров, близких к точке бифуркации седловидного узла [26]. Результаты моделирования с неустойчивыми устойчивыми состояниями отбрасывались, но в тех случаях, когда один из генотипов (т.е. наборы параметров) подходят близко к точке бифуркации, не пересекая ее, мы получаем карты генотип-фенотип, как в случае , где один генотип (или небольшой набор) дает гораздо более высокие фенотипические значения, чем другие. Такие карты GP, подобные модели дублирующих факторов в Hill et al. [3], как известно, дают низкие отношения V A / V G , несмотря на монотонность. Подобные карты GP, дающие отношения V A / V G , близкие к нулю, были также обнаружены Keightley [32] в его исследовании метаболических моделей, обладающих нулевыми аллелями во всех локусах.
Соображения по поводу общности результатов
Наша главная причина для ограничения выборочной генетической изменчивости параметров в пределах 30% от опубликованных исходных значений заключалась в том, чтобы избежать качественных (или топологических) изменений в динамике. Такие качественные изменения часто являются биологически реалистичными описаниями нокаутов или других крупных генетических изменений, например потенциалы действия переменной амплитуды (альтернаны) [17]; потеря стабильных циркадных колебаний [23]; и фенотипы нежизнеспособных мутантов клеточного цикла [22].Однако, поскольку концепции компонентов наследственности и дисперсии определены для фенотипов, демонстрирующих непрерывную, а не дискретную изменчивость, мы стремились избежать здесь таких качественных изменений.
Мы провели моделирование с пятью полиморфными локусами для моделей цАМФ (рис. S8A), гликолиза (рис. S8B), клеточного цикла (рис. S9) и потенциала действия (рис. S10) (циркадная модель явно описывает только три гена). Полученные значения V A / V G были несколько ниже, чем с тремя локусами, но общий вид распределений и четкие различия между моделями не изменились.Это указывает на то, что наши результаты имеют общее значение для олигогенных признаков.
Следует подчеркнуть, что пять исследованных моделей cGP отличаются по ряду других аспектов, чем те, которые выделены здесь, такие как размер системы (количество переменных состояния) и масштабы времени процесса. Эти особенности также могут способствовать наблюдаемой вариации в распределениях отношений V A / V G . Однако такие структурные различия неизбежны, когда целью является сравнение экспериментально подтвержденных моделей, предназначенных для описания конкретных биологических систем.Дополнительный подход заключается в изучении общих моделей, в которых размер системы и структура уравнения фиксированы, а матрица связности может быть изменена для описания семейства систем [35]. Это облегчает теоретико-графовое сравнение систем за счет некоторого биологического реализма. Мы ожидаем, что основные выводы из таких исследований будут аналогичны нашим, но вполне может оказаться, что на передний план могут выйти и другие важные общие идеи.
Все модели в нашем исследовании описывают части клеточного механизма, и результирующие фенотипы, таким образом, являются клеточными, а не организменными.Мы не думаем, что это серьезный недостаток с точки зрения основных выводов, которые вытекают из наших результатов. Тем не менее, мы ожидаем, что применение нашего подхода к многомасштабным моделям, включая фенотипы клеток, тканей и целых органов [36], обеспечит значительно улучшенную основу для объяснения того, как свойства карты GP варьируются между биологическими системами и внутри них с точки зрения регуляторной анатомии и ассоциированная генетическая изменчивость [37], [38].
Поскольку наш подход можно использовать вместе с любой моделью вычислительной биологии, он может внести существенный вклад в теоретическую основу, способную предсказать, когда следует ожидать низкого или высокого уровня V A / V G или h 2 / H 2 соотношения из принципов регуляторной биологии.Таким образом, причинно-связное моделирование генотип-фенотип считается многообещающим подходом для интеграции причинно-следственных моделей биологических сетей и физиологии с количественной генетикой [39]–[44].
Дополнительная информация
Рисунок S1
Блок-схема моделирования и анализа методом Монте-Карло. Блок-схема моделирования методом Монте-Карло, описанного в разделе «Методы» «Моделирование методом Монте-Карло», и последующего анализа, описанного в разделе «Методы» «Статистический анализ».
(PDF)
Рисунок S7
Эмпирическая кумулятивная функция распределения В А / В Г соотношения для фенотипов модели клеточного цикла. Эмпирические кумулятивные функции распределения (ось Y) отношений V A / V G (ось X) для всех фенотипов, изученных в модели клеточного цикла. Фенотипы делятся на 3 группы.Клеточные события относятся к дискретным событиям, определенным в модельном документе, и включают время почкования (Bud), время репликации ДНК (Rep) и время выравнивания хромосом на метафазных пластинах (Spn). Пиковая концентрация включает концентрацию фосфорилированного комплекса, стимулирующего анафазу (APCP), G1-стабилизирующего белка Cdc6, белка 2 циклина B-типа (Clb2), циклина B-типа, стимулирующего S-фазу (Clb5), стартовой киназы. (Cln2) и белок, стабилизирующий фазу G1 (Sci1). Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1.См. Таблицу S8 для дальнейшего описания фенотипов и численных сводок распределения соотношений V A / V G .
(PDF)
Рисунок S8
Эмпирическая кумулятивная функция распределения В А / В Г Соотношения для фенотипов моделей цАМФ (А) и гликолиза (В) с 5 полиморфными локусами. показаны результаты моделирования с 3 полиморфными локусами. A. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели цАМФ: начальные равновесные концентрации до добавления внешней глюкозы циклического аденозинмонофосфата (цАМФ), G-белка Ras2a (Ras2a), фактора гуанин-нуклеотидного обмена (Cdc25), протеинкиназы А (PKAi). Пиковые значения после добавления глюкозы этих белков (cAMPv, Ras2av, Cdc25v и PKAiv), белка-гомолога повторов Келха (Krhv), G-белка Gpa2a (Gpa2av) и фосфодиэстеразы (Pde1v).Время, необходимое для достижения пиковых значений (cAMPt, Ras2at, Cdc25t, PKAit, Krht, Gpa2at, Ped1t). B. Эмпирическая кумулятивная функция распределения (ось Y) отношений V A / V G (ось x) для равновесных концентраций 13 метаболитов в модели гликолиза ацетальдегида (ACE), 1 ,3-бисфосфоглицерат (BPG), фруктозо-1,6-бисфосфат (F16P), фруктозо-6-фосфат (F6P), глюкозо-6-фосфат (G6P), глюкоза в клетке (GLCi), никотинамидадениндинуклеотид (NADH), фосфаты в адениновых нуклеотидах (P), 2-фосфоглиерате (P2G), 3-фосфоглицерате (P3G), фосфоенолпирувате (PEP), пирувате (PYP) и триофосфате (TRIO).
(PDF)
Рисунок S9
Эмпирическая кумулятивная функция распределения В А / В Г соотношения для фенотипов модели клеточного цикла с 5 полиморфными локусами. На рисунке S7 показаны результаты моделирования с 3 полиморфными локусами. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для всех фенотипов, изученных в модели клеточного цикла.Фенотипы делятся на 3 группы. Клеточные события относятся к дискретным событиям, определенным в модельном документе, и включают время почкования (Bud), время репликации ДНК (Rep) и время выравнивания хромосом на метафазных пластинах (Spn). Пиковая концентрация включает концентрацию фосфорилированного комплекса, стимулирующего анафазу (APCP), G1-стабилизирующего белка Cdc6, белка 2 циклина B-типа (Clb2), циклина B-типа, стимулирующего S-фазу (Clb5), стартовой киназы. (Cln2) и белок, стабилизирующий фазу G1 (Sci1).Время достижения пиковых фенотипов включает время достижения пиковых концентраций APCP, Cdc6, Clb2, Clb5, Cln2 и Sci1.
(PDF)
Рисунок S10
Эмпирическая кумулятивная функция распределения В А / В Г соотношения для фенотипов модели потенциала действия с 5 полиморфными локусами. показаны результаты моделирования с 3 полиморфными локусами. Эмпирические кумулятивные функции распределения (ось y) отношений V A / V G (ось x) для фенотипов, изученных в модели потенциала действия: время до 25%, 50%, 75% и 90% показаны начальные значения, амплитуда, начальные значения (база), пиковые значения, время до достижения пиковых значений потенциала действия (левая панель) и транзиент кальция (правая панель).
(PDF)
Таблица S2
Полиморфные элементы модели гликолиза. Список элементов и параметров модели гликолиза, используемых для выявления генетической изменчивости. Названия параметров из оригинальной публикации [20], имена, использованные в файле SBML, полученном с http://www.ebi.ac.uk/biomodels-main/BIOMD0000000064, и базовые значения в единицах измерения.
(PDF)
Таблица S4
Полиморфные элементы модели циркадной модели. Список элементов и параметров циркадной модели, используемых для проявления генетической изменчивости.Имена параметров из (набор параметров 4) в оригинальной публикации [23], имена, используемые в файле CellML «leloup_goldbeter_2004.cellml», полученные с http://models.cellml.org/workspace/leloup_goldbeter_2004/, и базовые значения с единицами измерения.
(PDF)
Таблица S5
Полиморфные элементы модели потенциала действия. Список элементов и параметров модели потенциала действия, используемых для проявления генетической изменчивости. Имена параметров из таблицы B1 в оригинальной публикации [24], имена, используемые в файле CellML, который доступен в качестве дополнительного материала (имя файла «Модель LNCS.zip») по адресу doi:10.3389/fphys.2011.00106 и базовые значения в единицах.
(PDF)
Таблица S6
Краткое описание фенотипов, пороги изменчивости и распределение В А / В Г Отношения для модели цАМФ. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог.Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
(PDF)
Таблица S7
Краткое описание фенотипов, пороги изменчивости и распределение В А / В Г соотношения для модели гликолиза. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц.Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
(PDF)
Таблица S8
Краткое описание фенотипов, пороги изменчивости и распределение В А / В Г соотношения для модели клеточного цикла. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
(PDF)
Таблица S9
Краткое описание фенотипов, пороги изменчивости и распределение В А / В Г соотношения для циркадной модели. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог.Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости. Сокращения: фосфорилированные – phos., цитозольные – cyt., ядерные – nuc., нижняя концентрация – b.c., пиковая концентрация – p.c.
(PDF)
Таблица S10
Краткое описание фенотипов, пороги изменчивости и распределение В А / В Г отношения для модели потенциала действия. В первых трех столбцах перечислены сокращения фенотипов, используемые в этом исследовании, текстовое описание фенотипов и их единиц. Пороги, используемые для фильтрации набора данных с очень низкой относительной и/или абсолютной изменчивостью, перечислены в следующих двух столбцах, за которыми следует количество симуляций Монте-Карло (из 1000), прошедших порог. Последние 7 столбцов содержат квантили и средние значения для наборов данных, прошедших порог изменчивости.
(PDF)
Текст S1
Более подробные описания пяти моделей cGP.
(PDF)
Валидность в широком и узком смысле трех полигенных методов оценки риска рака предстательной железы
Задний план: Существует несколько методов оценки полигенного риска (PRS) для измерения кумулятивного эффекта множественных однонуклеотидных полиморфизмов (SNP), связанных с риском.Их эффективность в прогнозировании риска на индивидуальном уровне изучена недостаточно.
Методы: Мы сравнили эффективность трех методов PRS для оценки риска рака предстательной железы в когорте клинических испытаний, включая оценку генетического риска (GRS), обрезку и определение порога (P + T) и прогнозирование неравновесия по сцеплению (LDpred). Эффективность оценивалась по децилям баллов (валидность в широком смысле) и значениям баллов (валидность в узком смысле).
Результаты: Потребовался процесс обучения для выявления лучшей модели P + T (397 SNP) и модели LDpred (3 011 362 SNP). Напротив, GRS рассчитывался непосредственно на основе 110 установленных SNP, связанных с риском. Для валидности в широком смысле в тестируемой популяции более высокие децили были значительно связаны с более высоким наблюдаемым риском; P тренд был 7,40 × 10 -11 , 7.64 × 10 -13 и 7,51 × 10 -10 для GRS, P + T и LDпред соответственно. Для достоверности с узким смыслом наклон калибровки (1 — лучший) составлял 1,03, 0,77 и 0,87, а средний балл систематической ошибки (0 — лучший) составлял 0,09, 0,21 и 0,10 для GRS, P + T и LDpred соответственно.
Выводы: Показатели GRS были лучше, чем P + T и LDpred. Меньшее количество и хорошо зарекомендовавшие себя SNP GRS также делают его более выполнимым и интерпретируемым для генетического тестирования на индивидуальном уровне.
Ключевые слова: клиническая достоверность; оценка генетического риска; рак простаты.
Происхождение аддитивной генетической изменчивости, обусловленной положительным отбором | Молекулярная биология и эволюция
Аннотация
Фундаментальная теорема Фишера о естественном отборе не предсказывает аддитивной дисперсии приспособленности в естественной популяции. Соответственно, исследования в различных диких популяциях практически не выявили наследуемости в узком смысле ( ч 2 ) признаков, важных для приспособленности.Однако время от времени сообщаются контрпримеры, требующие более глубокого понимания эволюции аддитивной дисперсии. В этом исследовании мы предлагаем адаптивную дивергенцию, за которой следует смешение популяций, как источник аддитивной генетической изменчивости эволюционно важных признаков. Мы экспериментально проверили эту гипотезу, изучив панель из примерно 1000 сегрегантов дрожжей, полученных гибридом двух штаммов дрожжей, испытавших адаптивную дивергенцию. Мы измерили более 400 морфологических признаков дрожжевых клеток и обнаружили сильную положительную корреляцию между ч 2 и их эволюционным значением.Поскольку адаптивная дивергенция, за которой следует смешение популяций, может происходить постоянно, особенно у видов с широким географическим распространением и высокой способностью к миграции (например, у людей), это открытие согласовывает наблюдения обильных аддитивных вариаций в эволюционно важных признаках с фундаментальной теоремой Фишера о естественном отборе. Важно отметить, что обнаруженная роль положительного отбора в продвижении, а не в истощении аддитивной изменчивости предлагает простое объяснение того, почему аддитивная генетическая изменчивость может быть доминирующей в популяции, несмотря на вездесущий эпистаз между генами, наблюдаемый в функциональных тестах.
Введение
Интересным вопросом в генетике является то, как аддитивная генетическая изменчивость сложного признака развилась в популяции, учитывая, что эпистаз между генами, согласно функциональным исследованиям, кажется повсеместным (Costanzo et al. 2010; Sackton and Hartl 2016). Фундаментальная теорема Фишера о естественном отборе предсказывает небольшую аддитивную дисперсию (или наследуемость в узком смысле, ч 2 ) приспособленности, потому что естественный отбор быстро закрепляет аллели с наивысшей приспособленностью (Mousseau and Roff, 1987; Merila and Sheldon, 1999b; Кроу 2002).Расширенное предсказание теоремы состоит в том, что признаки, тесно связанные с приспособленностью (т. е. эволюционно важные признаки), должны иметь меньшее значение h 2 , чем менее связанные с приспособленностью (Круук и др., 2000), потому что реакция на естественный отбор на приспособленности будет определять эволюцию связанных черт (Orr 2009). Отрицательная корреляция между ч 2 и важностью признака была обнаружена во множестве исследований различных видов или популяций (Merila and Sheldon 1999a, 2000; Kruuk et al.2000 г.; Стирлинг и др. 2002 г.; Теплицкий и соавт. 2009 г.; Уилрайт и др. 2014; Степанач и соавт. 2017). Например, для дикой самки благородного оленя ( Cervus elaphus ) h 2 нескольких признаков жизненного цикла, включая общее число потомков, успешность размножения взрослых особей и продолжительность жизни, равны нулю ( Круук и др., 2000). Между тем, морфологические признаки, такие как длина ног и длина челюсти, которые, как считается, менее связаны с приспособленностью, имеют гораздо более высокие значения, чем признаки жизненного цикла.Эта закономерность также верна для ошейниковой мухоловки ( Ficedula albicollis ), воробьев саванны ( Passerculus sandensis ), красноклювой чайки ( Larus novaehollandiae ) и т. д. (Merila and Sheldon 2000; Stirling et al. 2002; Теплицкий и др., 2009 г.; Уилрайт и др., 2014 г.).
Однако есть также сообщения об обильной аддитивной дисперсии важных признаков (Петтай и др., 2005; Теплицкий и др., 2009; Косова и др., 2010; Милот и др., 2011; Чжан, 2012).В частности, иногда наблюдается даже положительная корреляция между ч 2 и важностью признака. Например, в популяции снежных баранов с горы Рэм самая низкая ч 2 была для массы тела при первородстве (0,02), тогда как ч 2 пожизненной плодовитости достигала 0,66 (Reale and Festa). -Бьянше 2000). Было предложено множество объяснений наблюдений. В дополнение к рассмотрению различных компонентов дисперсии ч 2 (Visscher et al.2008) или уравновешивающий отбор (Barton and Keightley 2002; Grieshop and Arnqvist 2018), преобладает точка зрения, согласно которой колебания окружающей среды в сочетании с мутациями могут способствовать поддержанию высокой аддитивной дисперсии приспособленности (Burger and Gimelfarb 2002; Crow 2008; Zhang 2012). Все эти объяснения являются теоретическими, и им не хватает эмпирических доказательств. Что еще более важно, они не предсказывают положительной корреляции между ч 2 и важностью признака.
Мы полагаем, что здесь существенную роль может играть экологический фактор эволюции, а именно миграция.Для данного вида часто существует множество расхождений между популяциями (Пиццо и др., 2008; Свед и др., 2008; Рой и др., 2014). Когда расхождения сочетаются с локальной адаптацией (т. е. адаптивными расхождениями), что довольно часто происходит в природе (Pulido 2007; Liedvogel et al. 2011), аллели с полезными аддитивными эффектами на важные признаки будут предпочтительно закрепляться в популяции. Поскольку в разных популяциях будут отобраны разные гены, последующее смешение популяций в результате миграции приведет к новым популяциям с обильными аддитивными генетическими вариациями по важным признакам.В этом исследовании мы разработали экспериментальную проверку этой разумной гипотезы, выявив цикл рождения и смерти аддитивной дисперсии, обусловленный положительным отбором.
Результаты
Мы исследовали панель, состоящую примерно из 1000 сегрегантов прототрофных гаплоидных дрожжей, полученных в результате скрещивания двух штаммов Saccharomyces cerevisiae (родительский BY и родительский RM). Две родительские линии различаются примерно на 0,5% на уровне геномной последовательности и испытали адаптивную дивергенцию в соответствии с анализом набора признаков основных компонентов (Ho et al.2017). Все около 1000 сегрегантов были генотипированы в предыдущем исследовании (Bloom et al. 2013). Сначала мы проверили сегрегантную панель и удалили сегреганты, которые оказались несовместимыми с указанными генотипами (рис. 1А; см. Материалы и методы).
Рис. 1.
Измерение 405 клеточных морфологических признаков и их наследуемость в сегрегантной популяции дрожжей. ( A ) Экспериментальный процесс в этом исследовании. Как морфологические признаки, так и скорость роста каждого сегреганта измеряли в повторах.( B ) Схематическая диаграмма дрожжевой клетки с основными координатами для характеристики морфологических признаков клетки. ( C ) Распределение CV 405 признаков, рассчитанное для технических повторностей сегрегантов A11_01 (слева) и A11_96 (справа) соответственно. ( D ) CV, полученные для сегрегантов A11_01 и A11_96, согласуются (Pearson’s R = 0,76, P < 2,2 × 10 −16 , N 9003 = 0,76) ( E ) Наследуемость в широком смысле ( H 2 ) и наследуемость в узком смысле ( h 2 ) каждого признака.Столбики погрешностей представляют SE.
Рис. 1.
Измерение 405 клеточных морфологических признаков и их наследуемость в сегрегантной популяции дрожжей. ( A ) Экспериментальный процесс в этом исследовании. Как морфологические признаки, так и скорость роста каждого сегреганта измеряли в повторах. ( B ) Схематическая диаграмма дрожжевой клетки с основными координатами для характеристики морфологических признаков клетки. ( C ) Распределение CV 405 признаков, рассчитанное для технических повторностей сегрегантов A11_01 (слева) и A11_96 (справа) соответственно.( D ) CV, полученные для сегрегантов A11_01 и A11_96, согласуются (Pearson’s R = 0,76, P < 2,2 × 10 −16 , N 9003 = 0,76) ( E ) Наследуемость в широком смысле ( H 2 ) и наследуемость в узком смысле ( h 2 ) каждого признака. Столбики погрешностей представляют SE.
Мы измерили 405 клеточных морфологических признаков для каждого сегреганта с двумя техническими повторениями, следуя предыдущему протоколу с некоторыми изменениями (см. Материалы и методы) (Ohya et al.2005). Эти признаки связаны с признаками материнской клетки и/или почки на разных стадиях, такими как площадь, расстояние, локализация, угол, соотношение и т. д. (рис. 1B). После исключения измерений с недостаточным количеством клеток для расчета признаков мы получили информацию о морфологических признаках для 734 сегрегантов, 73,3% (538/734) из которых имели данные как минимум двух повторностей (дополнительная таблица S1, Дополнительный материал онлайн). Приблизительно 99,5% значений признаков были получены из> 100 клеток данного сегреганта (дополнительный рис.S1, Дополнительный материал онлайн). Сегреганты A11_01 и A11_96 измеряли в каждом эксперименте в качестве технического контроля потенциальной операционной погрешности при культивировании, окрашивании и визуализации. Мы рассчитали измерительный коэффициент дисперсии (CV) каждого морфологического признака, используя 26 технических повторов сегреганта A11_01 или 28 технических повторов сегреганта A11_96 соответственно (см. Материалы и методы). Полученные CV, как правило, были небольшими, более 80% из них были <0,2 (347/405 в A11_01 и 326/405 в A11_96; рис.1С). Кроме того, они были согласованными между измерениями в сегреганте A11_01 и A11_96 (Pearson's R = 0,76, P < 2,2 × 10 −1) Эти данные вместе свидетельствуют об отсутствии сильного группового эффекта в измерениях признаков. Мы также проверили попарно-ранговую корреляцию 405 признаков между 28 техническими повторениями сегреганта A11_96, или 26 повторениями сегреганта A11_01, или двумя повторениями 536 сегрегантов соответственно.Мы наблюдали неизменно сильные корреляции (дополнительный рисунок S2, дополнительный материал онлайн). Большое количество высококачественных количественных признаков одного и того же свойства (т. е. морфологии), измеренных в одних и тех же экспериментальных условиях, предоставило уникальную возможность изучить эволюцию аддитивной генетической изменчивости.
Квантильная нормализация необработанных значений признака была выполнена для обеспечения сопоставимости различных признаков (см. Материалы и методы). Наследуемость в широком смысле H 2 и наследуемость в узком смысле h 2 были оценены для каждого из 405 признаков в соответствии с предыдущим исследованием (Bloom et al.2013) (дополнительная таблица S2, дополнительный материал онлайн). H 2 из 405 признаков варьировались от 0,021 до 0,913 при медиане 0,478; ч 2 варьировались от 0,000 до 0,619 со средним значением 0,240 (рис. 1E). В этом исследовании не было эффектов доминирования, поскольку сегреганты гаплоидны; взаимодействия ген-среда должны быть слабыми, поскольку использовались одни и те же условия культивирования. Таким образом, здесь H 2 — доля фенотипической дисперсии ( V P ), объясняемая аддитивным ( V A ) и неаддитивным (или эпистатическим) эффектами ( V ) ), и ч 2 = В А / В Р .Поскольку учитывались нормализованные значения признаков, V P различных признаков находились в пределах примерно 1,5-кратного диапазона, тогда как h 2 охватывали более 100-кратный диапазон (дополнительный рисунок S3, дополнительный онлайн-материал) . В результате в этом исследовании ч 2 эффективно служили прямой мерой V A , поскольку они сильно коррелировали с пирсоновским R = 0,99 среди 405 признаков (S3, Дополнительный материал онлайн).
Чтобы оценить эволюционное значение морфологических признаков, мы рассчитали их отношение к скорости роста (RTGR). Мы измерили скорость роста каждого сегреганта в тех же условиях, что и для измерения признаков (рис. 1A и дополнительная таблица S3, дополнительный онлайн-материал). Для каждого из 405 признаков мы вычислили коэффициент корреляции Пирсона (Pearson’s R ) между значением признака и скоростью роста клеток среди 734 сегрегантов.После предыдущего исследования (Chen et al. 2018) абсолютное значение Pearson R затем использовалось в качестве RTGR морфологического признака; черты с большим RTGR считаются эволюционно более важными. Значение RTGR варьировалось от 0 до 0,308 со средним значением 0,065, что подчеркивает широкий диапазон эволюционного значения 405 морфологических признаков (рис. 2A).
Рис. 2.
Эволюционно важные признаки, как правило, имеют большую h 2 сегрегантную популяцию.( A ) Сильная положительная корреляция между h 2 и важностью признака, измеренной RTGR (Pearson’s R = 0,55, P < 2,2 × 10 −16 , Столбики погрешностей представляют SE. Серая зона показывает 95% доверительный интервал линии регрессии. ( B ) Определите 59 образцовых признаков из 405 признаков. Тепловая карта показывает сходство признаков в пяти случайно выбранных кластерах, полученных с помощью апкластера, с одним типичным признаком, выделенным оранжевым цветом в каждом кластере.( C ) Корреляция между h 2 и RTGR сохраняется для образцовых признаков (Pearson’s R = 0,52, P = 2,5 × 10 −5 , , , , , , , , ( D ) Корреляция между h 2 и RTGR в основном сохраняется для признаков, характеризующихся на разных стадиях. На верхних диаграммах показаны состояния дрожжевой клетки, которые разделяют клеточные стадии. ( E ) Корреляция между h 2 и RTGR сохраняется для признаков, связанных с клеточной стенкой (окраска FITC-ConA) или ядром (окраска Hoechst).( F ) Положительная корреляция между h 2 и важностью признака, измеренной 1-CV (Pearson’s R = 0,34, P = 1,4 × 10 −9 ,
Рис. 2.
Эволюционно важные признаки, как правило, имеют большую h 2 в сегрегантной популяции. ( A ) Сильная положительная корреляция между h 2 и важностью признака, измеренной с помощью RTGR (Pearson’s R = 0.55, P < 2,2 × 10 −16 , N = 405). Столбики погрешностей представляют SE. Серая зона показывает 95% доверительный интервал линии регрессии. ( B ) Определите 59 образцовых признаков из 405 признаков. Тепловая карта показывает сходство признаков в пяти случайно выбранных кластерах, полученных с помощью апкластера, с одним типичным признаком, выделенным оранжевым цветом в каждом кластере. ( C ) Корреляция между h 2 и RTGR сохраняется для образцовых признаков ( R Пирсона = 0.52, P = 2,5 × 10 −5 , N = 59). ( D ) Корреляция между h 2 и RTGR в основном сохраняется для признаков, характеризующихся на разных стадиях. На верхних диаграммах показаны состояния дрожжевой клетки, которые разделяют клеточные стадии. ( E ) Корреляция между h 2 и RTGR сохраняется для признаков, связанных с клеточной стенкой (окраска FITC-ConA) или ядром (окраска Hoechst). ( F ) Положительная корреляция между h 2 и важностью признака, измеренной 1-CV ( R Пирсона = 0.34, P = 1,4 × 10 −9 , N = 293).
Согласно нашей гипотезе, смешение двух популяций с адаптивной дивергенцией привело бы к образованию новой популяции с более аддитивными вариациями эволюционно более важных признаков. Доступность как h 2 , так и эволюционная важность большого числа признаков позволили провести прямую проверку гипотезы. В соответствии с гипотезой мы обнаружили сильную положительную корреляцию между ч 2 и важностью признака, оцененной с помощью RTGR среди 405 признаков дрожжей ( R Пирсона = 0.55, P < 2,2 × 10 −16 ; инжир. 2А). Поскольку многие признаки коррелируют друг с другом, мы провели кластеризацию распространения сходства и получили 59 кластеров признаков, каждый из которых имеет образцовый признак (рис. 2B и дополнительный рис. S4, дополнительный материал онлайн). Количество признаков, представленных образцовым признаком, варьировалось от 2 до 16 со средним значением 6, и между 59 образцовыми признаками была только слабая корреляция (дополнительный рис. S4, Дополнительный материал онлайн).Сильная положительная корреляция между H 2 и RTGR осталась, когда были рассмотрены только примерные черты (Pearson’s R = 0,52, P = 2,5 × 10 -5 , рис. 2с).
405 признаков представляют морфологию клеток на разных стадиях клеточного цикла. Мы разделили эти признаки на четыре категории в зависимости от состояния почки и ядра (см. Материалы и методы). Признаки стадии A1B и стадии A, как правило, имеют небольшой RTGR, что предполагает менее избирательные ограничения на морфологию двух стадий.Важно отметить, что положительная корреляция между ч 2 и RTGR сохранилась, за исключением признаков стадии А (рис. 2D). Кроме того, поскольку 405 признаков представляют особенности клеточной стенки и ядра, которые окрашиваются двумя разными красителями FITC-ConA и Hoechst соответственно (см. Материалы и методы), мы исследовали 133 признака, связанных с клеточной стенкой, и 272 признака, связанных с ядром. по отдельности. Положительная корреляция между ч 2 и RTGR сохраняется в обеих категориях (рис.2Е).
Предыдущее исследование предполагает, что CV в измерении признаков может служить косвенным показателем важности признаков, с меньшим CV для более важных признаков (Ho and Zhang 2014). Чтобы быть консервативными, мы рассмотрели только 293 признака, которые имеют постоянный CV между A11_01 и A11_96, и использовали среднее значение для представления важности признака (дополнительный рисунок S5, дополнительный материал онлайн). Чтобы соответствовать RTGR в отношении важности признака, мы рассмотрели 1-CV, а не CV. Мы наблюдали положительную корреляцию между ч 2 и 1-CV ( R Пирсона = 0.34, P = 1,4 × 10 −9 ; инжир. 2F), что подтверждает нашу гипотезу. Образец обычно сохранялся за счет рассмотрения образцовых признаков или путем разделения признаков на разные категории (дополнительный рис. S5, дополнительный онлайн-материал).
Затем мы нанесли на карту локусы количественных признаков (QTL) для каждого из признаков. Всего было обнаружено 2505 QTL для 393 признаков (дополнительная таблица S4, дополнительный онлайн-материал), а количество QTL варьировалось от 1 до 19 со средним значением 6 для каждого признака (дополнительный рис.S6, Дополнительный материал онлайн). Было 12 признаков без обнаруживаемых QTL, что соответствует их чрезвычайно низкому уровню ч 2 (медиана ч 2 ~ 0,016). Почти во всех случаях разъемы черты, объясненные обнаруженными QTLS, было близко к H 2 (Pearson’s R = 0,96, P < 2,2 × 10 -16 , рис. 3а), предполагая почти насыщение обнаружения QTL. Это согласуется с предыдущим наблюдением на панели сегрегантов дрожжей (Bloom et al.2015). Каждый из QTL (~90,6%) объяснял небольшую долю (<5%) дисперсии признаков (дополнительный рисунок S6, дополнительный материал онлайн).
Рис. 3.
Анализ QTL 405 признаков. ( A ) Доля фенотипической изменчивости, объясняемая QTL, близка к ч 2 , что предполагает почти насыщение обнаружения QTL. Столбики погрешностей представляют SE. ( B ) Сильная положительная корреляция между номером QTL и h 2 (Pearson’s R = 0.83, P < 2,2 × 10 −16 , N = 405). Серая зона показывает 95% доверительный интервал линии регрессии. ( C ) Умеренно положительная корреляция между числом QTL и важностью признака, измеренной с помощью RTGR (слева) или 1-CV (справа). ( D ) Нет очевидной корреляции между долей генов, влияющих на признак ( f ген ) и важностью признака. Включены только те признаки, исследованные на панели C, которые имеют оценочные значения f гена .
Рис. 3.
Анализ QTL 405 признаков. ( A ) Доля фенотипической изменчивости, объясняемая QTL, близка к ч 2 , что предполагает почти насыщение обнаружения QTL. Столбики погрешностей представляют SE. ( B ) Сильная положительная корреляция между номером QTL и h 2 (Pearson’s R = 0,83, P < 2,2 × 10 −16 , , , Серая зона показывает 95% доверительный интервал линии регрессии.( C ) Умеренно положительная корреляция между числом QTL и важностью признака, измеренной с помощью RTGR (слева) или 1-CV (справа). ( D ) Нет очевидной корреляции между долей генов, влияющих на признак ( f ген ) и важностью признака. Включены только те признаки, исследованные на панели C, которые имеют оценочные значения f гена .
Мы обнаружили, что ч 2 признака сильно коррелируют с количеством QTL (Pearson R = 0.83, P < 2,2 × 10 −16 ; инжир. 3Б). Соответственно, в целом было обнаружено больше QTL для более важных признаков (рис. 3C и дополнительный рисунок S7, дополнительный материал онлайн). Этот результат показал, что после разделения двух родительских дрожжей сегрегантной популяции, исследованной здесь, существует больше дивергентных локусов для важных признаков. Есть два возможных объяснения: во-первых, существует больше генов и, следовательно, больше мутаций, влияющих на важные признаки; во-вторых, более высокая скорость фиксации мутаций, влияющих на важные признаки.Чтобы различить их, мы исследовали данные морфологии клеток, полученные для большого набора мутантов с делецией одного гена дрожжей. Для каждого из признаков мы получили долю генов, влияющих на признак ( f ген ), следуя предыдущему исследованию (Ho and Zhang 2014). Нам не удалось наблюдать более крупный f ген для более важных признаков (рис. 3D), что позволяет предположить, что второе объяснение правдоподобно, хотя количество полных генов, влияющих на признак, не обязательно говорит о количестве естественных вариантов, влияющих на признак.Более высокая скорость фиксации мутаций, влияющих на более важные признаки, указывает на то, что положительный отбор лежит в основе генетической дивергенции родительских дрожжей. Это перекликается с адаптивной фенотипической эволюцией дрожжей S. cerevisiae , предложенной ранее на основании более быстрой фенотипической эволюции более важных признаков (Ho et al. 2017). Следует отметить, что многие QTL, обнаруженные для важного признака, часто демонстрировали противоположные эффекты у родителя (дополнительный рисунок S8, дополнительный онлайн-материал), что указывает на то, что фенотипическая дивергенция между двумя родителями плохо отражает лежащую в основе генетическую дивергенцию.Это может объяснить относительно слабый сигнал адаптивной фенотипической дивергенции между двумя родителями (Ho et al. 2017).
Обсуждение
Фундаментальная теорема Фишера о естественном отборе обеспечивает общую основу для размышлений об эволюции аддитивной генетической изменчивости. Все предыдущие эмпирические исследования по этому вопросу основаны на диких популяциях, и полученные закономерности противоречивы, что часто приписывается смешанным экологическим факторам. Это исследование, насколько нам известно, является первым контролируемым экспериментом по изучению взаимосвязи между аддитивной изменчивостью и эволюционной значимостью большого набора количественных признаков.Преимущество контролируемых экспериментов заключается в том, что все экологические переменные в диких популяциях, такие как питание, паразиты, хищники и т. д., фиксированы. Однако в нашем эксперименте есть одна оговорка. В частности, поскольку предполагаемая адаптивная дивергенция двух родительских дрожжей должна происходить в естественной среде, важность признака, полученная в лабораторных условиях, может не обязательно соответствовать важности естественной среды. Тем не менее, эта проблема, скорее всего, уменьшит корреляцию между ч 2 и важностью признака, недооценивая вклад положительного отбора в происхождение аддитивной дисперсии.
Неожиданная роль положительного отбора, которую может сыграть положительный отбор в развитии аддитивной изменчивости, дает решение давней загадки, а именно, что аддитивные вариации часто широко распространены в популяции, несмотря на повсеместные неаддитивные (или эпистатические) взаимодействия, наблюдаемые между генами в функциональных исследованиях. (Костанцо и др., 2010; Сактон и Хартл, 2016). Предыдущее объяснение загадки рассматривает частоты вариабельных аллелей, которые часто имеют J- или U-образную форму распределения в естественных популяциях, что сводит к минимуму эпистаз, исключая многолокусные генотипы (Hill et al.2008). Это, однако, не может объяснить, почему h 2 сравнительно велико в экспериментальных популяциях с одинаковыми частотами аллелей ~50% (Bloom et al. 2013). В модели адаптивной дивергенции, за которой следует смешение популяций, процесс адаптивной дивергенции эффективно служит фильтром для удаления неаддитивных аллелей, которые могут представлять собой большинство необработанных мутаций. Это связано с тем, что положительный отбор способствует фиксации аддитивных аллелей, но не неаддитивных аллелей. Последующее смешение популяции привело бы к популяции, полной аддитивных дисперсий (рис.4).
Рис. 4.
Цикл рождения-смерти аддитивной изменчивости, управляемой положительным отбором. Процесс рождения объясняется адаптивной дивергенцией, за которой следует смешение популяций. Во время адаптивной дивергенции разные генетические локусы будут положительно отобраны в дивергирующихся популяциях. Хотя неаддитивные вариации ( V Non-A ) могут составлять большинство в исходной популяции, аддитивные аллели, а не неаддитивные аллели, будут предпочтительно фиксированными.Последующее смешение популяции привело бы к новой популяции, полной аддитивных дисперсий ( V A ). Гибель аддитивных дисперсий — типичный процесс, описываемый фундаментальной теоремой Фишера о естественном отборе. Предполагая частые адаптивные дивергенции, за которыми следует смешение популяций, мог бы существовать постоянно активный цикл рождения и смерти, лежащий в основе структуры аддитивных вариаций вида.
Рис. 4.
Цикл рождения-смерти аддитивной изменчивости, управляемой положительным отбором.Процесс рождения объясняется адаптивной дивергенцией, за которой следует смешение популяций. Во время адаптивной дивергенции разные генетические локусы будут положительно отобраны в дивергирующихся популяциях. Хотя неаддитивные вариации ( V Non-A ) могут составлять большинство в исходной популяции, аддитивные аллели, а не неаддитивные аллели, будут предпочтительно фиксированными. Последующее смешение популяции привело бы к новой популяции, полной аддитивных дисперсий ( V A ).Гибель аддитивных дисперсий — типичный процесс, описываемый фундаментальной теоремой Фишера о естественном отборе. Предполагая частые адаптивные дивергенции, за которыми следует смешение популяций, мог бы существовать постоянно активный цикл рождения и смерти, лежащий в основе структуры аддитивных вариаций вида.
Примечательно, что приобретенные аддитивные вариации также истощались бы посредством отбора способом, четко описанным фундаментальной теоремой Фишера о естественном отборе. Мы провели моделирование, чтобы отследить процесс истощения репрезентативного признака с ч 2 ∼ 0.6 в сегрегантной популяции дрожжей и наблюдали быстрое сокращение ч 2 (дополнительный рисунок S9, дополнительный онлайн-материал). Следовательно, существует цикл аддитивной изменчивости рождения и смерти, обусловленный положительным отбором, который может повторяться снова и снова, если предположить повсеместное адаптивное расхождение, за которым следуют популяционные примеси у некоторых видов (рис. 4). Популяции на разных стадиях цикла будут иметь разные структуры аддитивной дисперсии. В соответствии с моделью Райта-Фишера с естественным отбором для новой полезной мутации потребуется ~1000 (или ~10000) поколений с с = 0.01 (или = 0,001) следует зафиксировать в диплоидной популяции N = 100 000, где N — размер популяции, а s — коэффициент отбора (Otto and Whitlock 2013). Время будет короче для аллелей с более чем 1/2 N исходной частотой, как в случае с популяциями, возникшими в результате примеси (Kimura 1983). Эти числа полезны для размышлений о временной шкале рождения и смерти аддитивной дисперсии в цикле.
Предлагаемое происхождение аддитивной дисперсии в этом исследовании имеет особое значение для следующих сценариев.Во-первых, наши знания о количественной генетике часто основаны на исследованиях лабораторных популяций, полученных путем скрещивания двух или нескольких штаммов/линий модельного организма, такого как дрожжи или плодовая муха (Блум и др., 2013; Лонг и др., 2014). Как и в текущем исследовании, все аддитивные дисперсии этих популяций объясняются процессом рождения дисперсии. Во-вторых, при разведении сельскохозяйственных культур или домашнего скота часто используется от нескольких до нескольких десятков поколений (Wiener et al. 1992; Hinze and Lamkey 2003). В результате в структуре аддитивной дисперсии размножающейся популяции также должен доминировать процесс рождения.В-третьих, наши люди имеют как широкое географическое распространение, так и сильную миграционную способность, первая из которых предсказывает частую локальную адаптацию (т. е. адаптивную дивергенцию), а вторая делает возможным повторное смешение популяций (Hellenthal et al., 2014; Fu et al., 2016). Следовательно, управляемый отбором цикл рождения и смерти аддитивной изменчивости мог быть постоянно активен в ходе эволюции человека. Результирующая структура аддитивной дисперсии в современных человеческих популяциях определяет, как можно изучать и понимать сложные человеческие черты.
Материалы и методы
Проверка панели сегреганта
Сегрегантная панель любезно предоставлена д-ром Л. Кругляк. Всего было 1056 сегрегантов в одиннадцати 96-луночных планшетах. Для проверки генотипов случайным образом отбирали 12 сегрегантов в каждом планшете и четыре локуса (MATa, MATα, hphMX4, natMX4) амплифицировали с помощью полимеразной цепной реакции для этих сегрегантов. Сравнивая результаты с генотипами, предоставленными доктором Л. Кругляк, мы обнаружили, что некоторый процент сегрегантов на чашках 8 и 9 не соответствовал друг другу, и не было шаблона для спасения сегрегантов в ряду или в строке, что может быть результатом загрязнений.Затем мы сфокусировали сегреганты в левых девяти чашках с правыми генотипами в следующих экспериментах.
Измерение морфологических признаков клеток
Морфологические признаки каждого сегреганта были измерены в соответствии с протоколом Ohya с некоторыми изменениями (Ohya et al. 2005). Вкратце, сегреганты выращивали в среде YPD (среда дрожжевого экстракта/пептона/декстрозы) до фазы насыщения при 25°C в течение 2 или 3 дней, а затем переносили в новые культуры в экспоненциальную фазу при 25°C в течение 3 или 4 часов.Каждый сегрегант имел две повторности. Клетки фиксировали 3,7% раствором формальдегида. Клеточные стенки окрашивали FITC-ConA (конъюгированный с изотиоцианатом флуоресцеина, конканавалин А, Sigma-Aldrich C7642). Ядра клеток окрашивали смесью Hoechst (Thermo Fisher, раствор Hoechst 33342) вместо DAPI для повышения специфичности. Мы исключили процесс окрашивания актина, потому что краситель актина (родамин фаллоидин) не был стабилен и не мог поддерживать изображение в течение длительного времени в высокопроизводительной автоматизированной обработке изображений.Окрашенные клетки высевали на микропланшеты (Greiner 781091) с ∼5,0 × 10 4 клеток на лунку и получали изображения с помощью IN Cell Analyzer 2200 (GE Healthcare) со 100-кратным объективом. Для каждого сегреганта было две технические повторности, и сегреганты A11_01 и A11_96 культивировались, окрашивались и визуализировались в каждом эксперименте в качестве технического контроля.
Программное обеспечение CalMorph использовалось для анализа изображений для количественной оценки морфологии дрожжей, и было получено 405 количественных признаков (Ohya et al.2005). Сегреганты, число клеток которых для подсчета признаков <80 в обеих двух повторностях, исключали. Значения всех признаков были перечислены в дополнительной таблице S1, дополнительный онлайн-материал. Было получено 734 сегреганта с 405 полученными морфологическими признаками, из которых 73,3% (538/734) имели по крайней мере две повторности. Квантильная нормализация была выполнена для необработанных значений признаков с помощью пакета R preprocessCore для дальнейших расчетов (https://github.com/bmbolstad/preprocessCore).
Признаки, полученные из клеточной стенки или ядра, можно отличить по начальной букве признака, где «С» относится к клеточной стенке, а «D» относится к ядру.Признаки на разных стадиях можно отличить по буквам после соединительной линии. «А» представляет признаки, рассчитанные по клеткам с одним ядром и без почки, «А1В» — признаки, рассчитанные по клеткам с одним ядром в материнской клетке с зачатком или ядром, делящимся на шейке, и «С» — признаки, полученные клетками с одним ядром в материнской клетке и почке. 405 признаков не являются независимыми, и 59 характерных признаков были получены с помощью пакета R «apcluster» (negDistMat, r = 2) с использованием средних нормализованных значений 734 сегрегантов (Frey and Dueck 2007).
Измерение скорости роста клеток
Штаммывыращивали в среде YPD до фазы насыщения при 25 °C в течение 2 или 3 дней, затем разбавляли 1:100 до 100 мкл свежей среды YPD в 96-луночном планшете. Два повтора каждого сегреганта помещали в один и тот же 96-луночный планшет. 96-луночные планшеты помещали на спектрофотометр для микропланшетов Epoch3 (BioTek) и инкубировали при 25°С при встряхивании. Поглощение при 600 нм каждой лунки определяли в час. Измерения длились 24 часа, и все штаммы достигли фазы насыщения.Скорость роста V max , то есть максимальный наклон кривой роста каждой лунки, использовали для оценки приспособленности каждого штамма. Чтобы контролировать позиционное смещение, исходные значения скорости роста в каждой пластине были подобраны с помощью надежной локально взвешенной регрессии с помощью R-пакета «locfit» в соответствии с исследованием Bloom et al. (2013). Средние нормализованные значения скорости роста были взяты за приспособленность каждого сегреганта и перечислены в дополнительной таблице S3, Дополнительный онлайн-материал.
Расчет наследуемости
Поскольку сегрегантная панель была создана Bloom et al., наследуемость в широком смысле ( H 2 ), наследуемость в узком смысле ( h 2 ), аддитивный QTL и дисперсия, объясняемая QTL каждого морфологический признак рассчитывали методами, основанными на исследовании Bloom et al. (2013). Кратко, H 2 рассчитывали по нормализованным значениям признаков сегрегантов с двумя повторностями. H 2 был оценен как σG2/(σG2+σE2), где σG2 — генетическая дисперсия, а σE2 — дисперсия ошибки. Это было выполнено функцией «lmer» в пакете lme4 R (Бейтс и др., 2015). При сравнении с H 2 , h 2 рассчитывали средние нормированные значения признаков сегрегантов с двух повторностей. И сегреганты только с одной репликацией также были включены в другие ситуации. Наследуемость в узком смысле оценивалась как σA2/(σA2+σEV2), где σA2 — аддитивная генетическая дисперсия, а σEV2 — дисперсия ошибки.Пакет R rrBLUP использовался для расчета ч 2 (Endelman 2011). SE H 2 и h 2 были рассчитаны путем удаления-одного складного ножа.
Аддитивный QTL каждого признака был обнаружен с использованием подхода пошагового прямого поиска, разработанного Bloom et al. (2013). Баллы LOD для каждого генотипического маркера и каждого признака рассчитывали как -nln1-r22ln10, где r — коэффициент корреляции Пирсона между генотипами и значениями признаков.Значимые генетические маркеры были обнаружены в четырех раундах с использованием различных порогов LOD, соответствующих 5% FDR, которые составляли 2,68, 2,92, 3,72 и 4,9 соответственно. Линейную модель множественной регрессии оценивали, взяв каждый QTL в качестве независимых переменных каждого признака, а общая фенотипическая дисперсия, объясняемая аддитивным QTL, представляла собой квадрат коэффициента множественной регрессии. Результаты были перечислены в дополнительной таблице S2, дополнительный онлайн-материал.
Вычислить признак RTGR
Для каждого признака рассчитывали средние нормированные значения повторности каждого сегреганта.Для сегрегантов только с одной репликацией значения признака были нормализованными значениями из единственного измерения. Абсолютное отношение Пирсона R между значениями признака и скоростью роста в среде YPD для 734 сегрегантов использовалось в качестве показателя RTGR для каждого признака. Результаты были перечислены в дополнительной таблице S2, дополнительный онлайн-материал.
Вычислить CV признака среди репликаций
Коэффициент вариации для каждого признака был рассчитан с использованием необработанных данных репликаций A11_01 и A11_96 соответственно.Признаки с большим CV могут указывать на их неопределенность, поэтому мы исключили семь признаков с CV >0,4 при использовании CV в качестве показателя важности признака. Чтобы оценить повторяемость двух групп, мы использовали индекс расстояния между двумя группами CV как CV01i-CV96i/(CV01i+CV96i), где CV01i и CV96i были значениями CV для признака i в A11_01 и A11_96, соответственно. . Большое расстояние CV означает, что устойчивость признака к окружающей среде будет разной для разных сегрегантов, поэтому мы исключили 105 признаков с индексом расстояния CV >0.2, чтобы получить набор признаков с последовательными измерениями. Осталось 293 черты. Результаты были перечислены в дополнительной таблице S2, дополнительный онлайн-материал.
Благодарности
Панель для сегрегации дрожжей – подарок доктора Кругляка из Калифорнийского университета в Лос-Анджелесе. Мы благодарны Jianguo Wang за раннюю проверку качества экспериментальных данных и сотрудников лаборатории He за создание платформы для измерения морфологии дрожжевых клеток. Эта работа была поддержана исследовательскими грантами Национального фонда естественных наук Китая (31630042 и 302), Национальной ключевой программы Китая (2017YFA0103504), Научно-технической программы Гуанчжоу (201707020035) и Ведущей программы талантов в науке и технике провинции Гуандун. Провинция.
Вклад авторов
Х.Х. и Л.Л. разработали исследование. З.С., Ю.В., Д.З., З.К., Л.Л. и Х.К. проводил опыты. Л.Л. и Х.Х. проанализировали данные и написали статью с участием всех авторов.
Каталожные номера
Бартон
NH
,Кейтли
ПД.
2002
.Понимание количественной генетической изменчивости
.Nat Rev Genet
.3
(1
):11
–21
.Бейтс
Д
,Махлер
М
,Болкер
БМ
,Уокер
СК.
2015
.Подгонка линейных моделей смешанных эффектов с использованием lme4
.J Stat Soft
.67
(1
):1
–48
.Bloom
JS
,Ehrenreich
,IM
,LOO
WT
,Lite
TL
,Kruglyak
L.
2013
.Поиск источников отсутствия наследуемости в дрожжевом скрещивании
.Природа
494
(7436
):234
–237
.Bloom
JS
,KOTENKO
I
,I
,Sadhu
MJ
,Treusch
S
,
Albert
FW
,Kruglyak
L.
2015
.Генетические взаимодействия вносят меньший вклад в изменение количественных признаков дрожжей, чем аддитивные эффекты
.Нац Коммуна
.6
(8712
):1
–6
.Бургер
Р
,Гимельфарб
А.
2002
.Колебания окружающей среды и роль мутаций в поддержании количественной генетической изменчивости
.Жене Рез
.80
(1
):31
–46
.Чен
H
,Ву
КИ
,Он
Х.
2018
.Генотип-фенотипические отношения в свете естественного отбора
.Мол Биол Эвол
.35
(3
):525
–542
.Costanzo
M
,M
,Baryshnikova
A
,Bellay
J
,KIM
Y
,SPEAR
ED
,SEVER
CS
,DING
H
,Ко
ДЖЛ
,Туфиги
К
,Мостафави
С
, и другие..2010
.Генетический ландшафт клетки
.Наука
327
(5964
):425
–431
.Ворона
Дж.Ф.
2002
.Перспектива: за Фишера, аддитивную генетическую изменчивость и фундаментальную теорему естественного отбора
.Эволюция
56
:1313
–1316
.Ворона
Дж.Ф.
2008
.Поддержание эволюционируемости
.Дж Жене
.87
(4
):349
–353
.Эндельман
JB.
2011
.Ридж-регрессия и другие ядра для геномной селекции с пакетом R rrBLUP
.Геном растения
4
(3
):250
–255
.Фрей
БДЖ
,Дуек
Д.
2007
.Кластеризация путем передачи сообщений между точками данных
.Наука
315
(5814
):972
–976
.фу
Q
,PostH
C
,C
,C
,,
M
,Petr
M
,Mallick
S
,Fernandes
D
,Ungwangler
A
,Хаак
Вт
,Мейер
М
,Митник
А
, и другие..2016
.Генетическая история Европы ледникового периода
.Природа
534
(7606
):200
–205
.Гришоп
К
,Арнквист
Г.
2018
.Инверсия генетической изменчивости в зависимости от пола для приспособленности
.ПЛОС Биол
.16
(12
):e2006810
.Hellentental
G
,Busby
GBJ
,,
G
,
G
,Wilson
JF
,Capelli
C
,Malush
D
,Myers
S.
2014
.Генетический атлас истории примесей человека
.Наука
343
(6172
):747
–751
.Hill
WG
,Goddard
ME
,Visscher
PM.
2008
.Данные и теория указывают в основном на аддитивную генетическую изменчивость сложных признаков
.ПЛОС Жене
.4
(2
):e1000008
.Хинце
Л
,Ламки
К.
2003
.Отсутствие эпистаза по урожайности зерна у элитных гибридов кукурузы
.Растениеводство
.43
(1
):46
–56
.Ho
WC
,Ohya
Y
,Zhang
J.
2017
.Проверка нейтральной гипотезы фенотипической эволюции
.Proc Natl Acad Sci U S A
.114
(46
):12219
–12224
.Хо
WC
,Чжан
Дж.
2014
.Генотип-фенотипическая карта комплексных признаков дрожжей: основные параметры и роль естественного отбора
.Мол Биол Эвол
.31
(6
):1568
–1580
.Кимура
М.
1983
.Нейтральная теория молекулярной эволюции
.Кембридж/Нью-Йорк
:Издательство Кембриджского университета
.Косова
G
,Abney
M
,Ober
C.
2010
.Материалы коллоквиума: наследуемость признаков репродуктивной приспособленности в человеческой популяции
.Proc Natl Acad Sci U S A
.107
(Suppl_1
):1772
–1778
.Kruuk
Le
,LE
,CLUTTON-BROCK
TH
,SLITE
J
,J
,pemberton
JM
,Broterhertone
S
,GUINNES
FE.
2000
.Наследуемость приспособленности в популяции диких млекопитающих
.Proc Natl Acad Sci U S A
.97
(2
):698
–703
.Liedvogel
M
,Akesson
S
,Bensch
S.
2011
.Генетика миграции в движении
.Тренды Ecol Evol
.26
(11
):561
–569
.Лонг
AD
,Макдональд
SJ
,Кинг
EG.
2014
.Препарирование сложных признаков с использованием ресурса синтетической популяции дрозофилы
.Тренды Genet
.30
(11
):488
–495
.Мерила
Дж
,Шелдон
БК.
1999
а.Генетическая архитектура приспособленных и непригодных черт: эмпирические закономерности и развитие идей
.Наследственность
83
(2
):103
–109
.Мерила
Дж
,Шелдон
БК.
1999
б.Генетическая архитектура приспособленных и непригодных черт: эмпирические закономерности и развитие идей
.Наследственность
83
(2
):103
–109
.Мерила
Дж
,Шелдон
БК.
2000
.Пожизненный репродуктивный успех и наследуемость в природе
.Am Nat
.155
(3
):301
–310
.MILOT
E
,Mayer
FM
,FM
,Nussey
DH
,Boisvert
M
,Pelletier
F
,Reale
D.
2011
.Доказательства эволюции в ответ на естественный отбор в современной человеческой популяции
.Proc Natl Acad Sci U S A
.108
(41
):17040
–17045
.Муссо
ТА
,Рофф
ДА.
1987
.Естественный отбор и наследуемость компонентов приспособленности
.Наследственность
59
(2
):181
–197
.Ohya
y
,SESE
J
,J
,Yukawa
M
,Sano
F
,Nakatani
Y
,Saito
TL
,Saka
A
,Фукуда
T
,Исихара
S
,Ока
S
, и другие..2005
.Многомерное и крупномасштабное фенотипирование дрожжевых мутантов
.Proc Natl Acad Sci U S A
.102
(52
):19015
–19020
.Орр
HA.
2009
.Фитнес и его роль в эволюционной генетике
.Nat Rev Genet
.10
(8
):531
–539
.Отто
SP
,Уитлок
МС.
2013
. Вероятность и время фиксации. В: Энциклопедия наук о жизни. Чичестер, Великобритания: John Wiley & Sons, Ltd.Pettay
JE
,JE
,KRUUK
LE
,Jokela
J
,Lummaa
V.
2005
.Наследуемость и генетические ограничения эволюции признаков в жизненном цикле у доиндустриальных людей
.Proc Natl Acad Sci U S A
.102
(8
):2838
–2843
.Pizzo
A
,A
,Roggero
A
,A
,Palestrini
C
,Moczek
AP
,Rolando
A.
2008
.Быстрые различия в форме между естественными и интродуцированными популяциями рогатого жука частично отражают различия между видами
.Эвол Дев
.10
(2
):166
–175
.Пулидо
Ф.
2007
.Генетика и эволюция миграции птиц
.BioScience
57
(2
):165
–174
.Реале
D
,Феста-Бьянше
М.
2000
.Количественная генетика признаков жизненного цикла долгоживущих диких млекопитающих
.Наследственность
85
(6
):593
–603
.ROY
J
,J
,M
,M
,Bradley
BJ
,Guschanski
K
,stephens
CR
,Bucknell
D
,Cirhuza
H
,Кусамба
К
,Кьюнгу
ДЖК
,Смит
В
, и другие..2014
.Недавние расхождения и сокращение численности популяций восточных горилл
.Биол Летт
.10
(11
):20140811
.Sackton
TB
,Hartl
DL.
2016
.Генотипический контекст и эпистаз у особей и популяций
.Сотовый
166
(2
):279
–287
.Стирлинг
DG
,Reale
D
,Roff
DA.
2002
.Отбор, структура и наследуемость поведения
.Дж Эвол Биол
.15
(2
):277
–289
.Свед
JA
,McRae
AF
,Visscher
PM.
2008
.Дивергенция между человеческими популяциями, оцененная по неравновесному сцеплению
.Am J Hum Genet
.83
(6
):737
–743
.Степанач
JL
,McGuigan
K
,Удары
МВт.
2017
.Наследственная микросредовая изменчивость ковариирует с приспособленностью в беспородной популяции Drosophila serrata
.Генетика
206
(4
):2185
–2198
.Теплицкий
C
,Миллс
JA
,Yarrall
JW
,Merila
J.
2009
.Наследуемость компонентов приспособленности в популяции диких птиц
.Эволюция
63
(3
):716
–726
.Visscher
PM
,Hill
WG
,Wray
NR.
2008
.Наследуемость в эпоху геномики – концепции и заблуждения
.Nat Rev Genet
.9
(4
):255
–266
.Wheelwright
NT
,Keller
LF
,Postma
E.
2014
.Влияние типа признака и силы отбора на наследуемость и эволюционируемость в популяции островных птиц
.Эволюция
68
(11
):3325
–3336
.Винер
Г
,Ли
ГДж
,Вуллиамс
Дж.
1992
.Влияние быстрого инбридинга и скрещивания инбредных линий на прирост живой массы овец
.Аним Науки
.55
(1
):89
–99
.Чжан
XS.
2012
.Геометрическая модель фитнес-ландшафта Фишера и вариации фитнеса в меняющейся среде
.Эволюция
66
(8
):2350
–2368
.Примечания автора
© Автор(ы) 2020.Опубликовано Oxford University Press от имени Общества молекулярной биологии и эволюции.
Это статья в открытом доступе, распространяемая в соответствии с лицензией Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии, что оригинал работа цитируется правильно.Рисунок 3. (a) Кровотечение из ногтевого ложа. (b) Аномальные капилляры в ногтевом валике. | Рисунок 5. (a) Телеангиэктазии макулярного типа. (б) Телеангиэктазии сосудистых звездочек. (c) Телеангиэктазии типа болезни Ранду-Ослера-Вебера. г) телеангиэктазии слизистой оболочки губ. |
Уравнение заводчика | Изучайте науку в Scitable
Большинство признаков, интересующих биологов, таких как поведение, морфология, физиология и восприимчивость к болезням, являются количественными. Это означает, что вариации между особями возникают из-за комбинированного эффекта различий в генотипе и в опыте окружающей среды. Генетический компонент этой вариации обычно включает множество генов. Поскольку фенотип возникает в результате сложного взаимодействия генов и окружающей среды, биологи вынуждены применять статистический подход к изучению эволюции количественных признаков.Возможно, самой основной формой этого подхода является уравнение заводчика (Lush, 1937). Уравнение заводчика предсказывает эволюционное изменение признака:
Здесь Z — среднее значение этого признака в популяции, а Δ Ζ — изменение среднего за одно полное поколение. Процесс отбора, будь то естественный или искусственный, входит в предсказание через S, дифференциал отбора, который является мерой связи между значениями признаков и приспособленностью (Falconer & Mackay, 1996).S будет отрицательным, если более низкие значения признака обеспечивают более высокую выживаемость или более высокий репродуктивный успех; положительный, если отбор благоприятствует более высоким значениям признаков. При усеченном отборе, когда для воспроизводства и создания следующего поколения выбирается фиксированная часть популяции, S равна разнице средних значений признаков между отобранными особями и всей популяцией.
Элегантность уравнения селекционера заключается в том, что все сложности многолокусного наследования сведены к h 2. . Единственная статистика, известная как наследуемость в узком смысле, h 2 , представляет собой долю изменчивости признаков, статистически приписываемую аддитивным генетическим эффектам. Если бы мы знали все гены, влияющие на признак, и то, как эти гены влияют на фенотип и частоты альтернативных аллелей в популяции, то мы могли бы вычислить h 2 напрямую. Однако у нас никогда нет всей этой информации. Широкая значимость уравнения селекционера обусловлена тем фактом, что h 2 можно оценить по сходству родственников.Другими словами, h 2 можно вывести, измерив фенотипическое сходство родителей с их потомками или братьев и сестер друг с другом, не зная ни количества генов, влияющих на признак, ни их расположения в геноме, ни того, как они взаимодействуют с окружающая среда. Здесь стоит подчеркнуть, что наследуемость в узком смысле является популяционным параметром, а не свойством отдельных особей. Неспособность полностью оценить это вызвало существенную путаницу в публичной интерпретации оценок человеческих черт h 2 (см. Freeman & Herron 1998 и Visscher et al. 2008 г. для последних комментариев).
Уравнение селекционера используется для различных целей в эволюционной биологии. Наиболее прямое применение — предсказание эволюционных изменений. На рис. 1 представлен пример эволюции размера цветка желтой мартышки ( Mimulus guttatus ). Этот эксперимент начался с двух популяций, которые были генетически эквивалентны и имели средний размер цветка. В каждом случае размер цветка измеряли на 1000 растений. 200 растений с самыми маленькими цветками были отобраны в «низкой» популяции и 200 самых крупных в «высокой» популяции.Выбранные растения случайным образом скрещивали внутри популяций для получения семян для следующего поколения. Этот процесс отчетливо повторялся в каждой популяции на протяжении шести поколений. Как это типично для экспериментов по искусственному отбору, популяции расходятся во времени.Рисунок 1
Разница в среднем размере цветка между популяциями, отобранными по более крупным и более мелким цветкам, соответственно, представлена как функция количества поколений отбора.
Уравнение селекционера можно применить здесь, чтобы предсказать расхождение в среднем размере цветка.Всего было проведено двенадцать поколений селекции (по шесть в каждом направлении). Наследуемость размера цветка оценивается примерно в 0,3 при сравнении размера цветка у других растений этой популяции. Наконец, выбор верхних (или нижних) 20% дает дифференциал отбора S = 1,4 σ, где σ — фенотипическое стандартное отклонение размера цветка в исходной популяции. Расхождение между наблюдаемым и прогнозируемым ответом составляет всего около 4% (рис. 1), что мало, учитывая, что h 2 оценивается с ошибкой.Большие отклонения наблюдаются во многих экспериментах по отбору (Hill & Caballero, 1992), результат, который, вероятно, отражает небольшой размер популяции. Уравнение селекционера является детерминированным и не учитывает напрямую генетический дрейф, случайные изменения частоты аллелей из-за выборки в конечной популяции.
Важным следствием направленного отбора по количественным признакам является то, что он может быстро производить новые фенотипы. Всего после шести поколений селекции мы видим цветы, которые больше и меньше, чем те, которые наблюдались в исходной популяции (рис. 2).Здесь производство новых фенотипов не вызвано новыми мутациями. Вместо этого это вызвано сортировкой многолокусной изменчивости на ранее нереализованные генотипы. В любой момент времени популяция обычно демонстрирует лишь небольшую долю того, что возможно для количественного признака. Например, если всего десять генов вносят вклад в изменчивость признаков (скромное число), существует более 59 000 возможных многолокусных генотипов. Существуют ли определенные генотипы, скажем, производящие экстремальные фенотипы, зависит от размера популяции и частот аллелей.Отбор изменяет частоты аллелей и, таким образом, увеличивает вероятность объединения многих аллелей со схожими фенотипическими эффектами в один и тот же многолокусный генотип. Рисунок 2Биологи-эволюционисты обычно используют уравнение селекционера только с частичной информацией. Если дивергенция (Δ Ζ ) оценена и известны накопленные эффекты отбора, оценка наследуемости признака может быть рассчитана как Δ Ζ /с.Это известно как реализованная наследственность. Напротив, история отбора часто неизвестна, когда дивергенция оценивается путем сравнения средних значений признаков различных природных популяций. Здесь S можно оценить как Δ Z /ч 2 , учитывая оценку наследуемости. Это известно как ретроспективный анализ выбора. Как осознанная наследуемость, так и анализ ретроспективной селекции обременены важными допущениями. В частности, каждый требует, чтобы Δ Z определялся отбором по признаку и чтобы отбор точно характеризовался уравнением селекционера.
Рисунки 1-2 получены в результате эксперимента по искусственному отбору. Уравнение селекционера было успешно применено для предсказания Δ Ζ в естественных популяциях (например, (Grant & Grant 1995)), но есть несколько убедительных примеров. Отчасти это отражает практические трудности точной оценки h 2 и S в природных популяциях. Однако применение также затруднено сильным предположением о том, что измеряемый признак непосредственно порождает различия в приспособленности, характеризуемые S (Morrissey et al. 2010). В природе выживание и размножение будут зависеть от многих отличительных особенностей организма. По этой причине биологи-эволюционисты обычно применяют многомерную форму уравнения селекционера (Lande, 1979; Lande & Arnold, 1983).
Следует отметить, что хотя уравнение селекционера названо в честь предсказания в сельскохозяйственной генетике (Lush, 1937), в эволюционной биологии оно чаще всего используется в качестве концептуального устройства. Сложный процесс, посредством которого отбор вызывает генетические изменения в популяции, распадается на два компонента: фенотипический отбор (S) и реакцию на отбор (h 2 ).Первый определяет взаимосвязь между фенотипом (значениями признаков) и приспособленностью (выживанием и репродуктивным успехом). Последнее определяется отображением генотипа в фенотип. Фенотипический отбор можно измерить в природе для многих организмов, для которых оценка наследственности затруднена или невозможна. Для организмов, за выживанием и размножением которых невозможно следить в дикой природе, исследования наследуемости дают информацию об эволюционном потенциале популяций. Следует признать, что разделение эволюции на фенотипический отбор и реакцию на отбор формально зависит от допущений, которые часто нарушаются в природе (Hadfield 2008; Rauscher 1992).Например, воздействие окружающей среды может одновременно влиять на значения признаков и измерения приспособленности, приводя к ложным оценкам S. Также может быть очень сложно получить точную оценку для отбора, когда многие особи умирают до проявления признака. Если эта «невидимая фракция» (Graffen, 1988) генетически отличается от особей, включенных в расчет S (с точки зрения локусов, влияющих на интересующий признак), то дифференциал отбора снова будет необъективным. Однако, несмотря на эти трудности, перспектива оценки статистических данных, непосредственно связанных с генетическими изменениями, послужила мотивом для большого количества эмпирических исследований в области эволюционной биологии (Kingsolver et al. 2001; Муссо и Рофф, 1987).
.